

Abstract
Exosomes are lipid bilayer vesicles released by multiple cell types. These bioactive vesicles are gradually becoming a leading star in intercellular communication involving in various pathological and physiological process. Exosomes convey specific and bioactive transporting cargos, including lipids, nucleic acids and proteins which can be reflective of their parent cells, rendering them attractive in cell-free therapeutics. Numerous findings have confirmed the crucial role of exosomes in restraining scars, burning, senescence and wound recovery. Moreover, the biology research of exosomes in cutting-edge studies are emerging, allowing for the development of particular guidelines and quality control methodology, which favor their possible application in the future. In this review, we discussed therapeutic potential of exosomes in different relevant mode of dermatologic diseases, as well as the various molecular mechanisms. Furthermore, given the advantages of favorable biocompatibility and transporting capacity, the bioengineering modification of exosomes is also involved.
Keywords: exosomes, cell-free therapy, cell communication, bioengineering, dermatologic diseases
Introduction
The skin is composed of hair, sweat glands, sebaceous glands and other accessory structures, as well as enriched blood vessels, nerve muscles and lymphatics (Wong et al., 2016). The direct contact with the environment leads to various external and internal damage to it. External factors including sunlight, infection, trauma and chemical media invasion caused by antigen or irritant, and internal factors including genetic factors, psychological factors, medical diseases, drug reactions and so on, which jointly bring significant discomfort to patients in appearance, functionality and even psychology. Some intractable skin diseases and severe burns still lack of effective treatments or control measures. For decades, “cell-based” therapy, especially mesenchymal stem cell (MSC) is holding tremendous promise for tissue regeneration. Emerging evidence shows that the cells like MSCs restore the tissue function mainly by paracrine manner, in which exosomes stand out (Phinney and Pittenger, 2017; Phan et al., 2018). Exosomes shed by MSC are safer and more stable than their donor cells concerning the management of live cells (Nikfarjam et al., 2020). These bioactive vesicles are becoming a leading star in cell-cell communication for delivering message from their cells of origin to the recipients (Wortzel et al., 2019; Li C. J. et al., 2020). Study have indicated that MSC-Exo may emerge as a promising therapeutic approach of SARS-CoV-2 pneumonia and in ischemic diseases (Akbari and Rezaie, 2020; Babaei and Rezaie, 2021). The basis, transformation and application of exosomes derived from stem cells and cells of various skin tissues are springing up and obvious progress has been made (Kalluri and Lebleu, 2020). In this review, the regulatory roles of exosomes in skin differentiation, development and the repair of injury will be further elaborated.
Brief Description of Exosomes
Biogenesis and Characterization
Numerous cells types release exosomes into the extracellular environment after fusing with the cytomembrane. Exosomes are membrane vesicle with a size of 30-150 nm. (Kalluri and Lebleu, 2020). Generally, exosomes can be classified as natural and engineered exosomes. The engineered exosomes are artificially designed and modified while natural exosomes can be natively derived from animals and plants. Ulteriorly, animal-derived exosomes, depending on the normal and tumor culture conditions, consist of two parts, normal exosomes and tumor exosomes (Zhang Y. et al., 2020). The heterogeneity of vesicles which originate from various cells are reflective in density, lipid, protein, density and subcellular origin (van der Pol et al., 2012). Multiple signaling pathways are involved in regulating the exosome biogenesis. Exosomes are acknowledgedly released by microvesicle body (MVB) signaling. At the very beginning, the endosome is formed as a small intracellular body by wrapping little intracellular fluid. The endosome is distinguished as early and late endosomes. After the intraluminal vesicles (ILV) formed, the early endosome develops into the late endosome, namely MVB. The MVB can degrade the contents by fusing with the lysosome or release its ILVs by fusing with the plasma membrane in an exocytotic manner, which are exosomes (Théry et al., 2002; Trajkovic et al., 2008; McAndrews and Kalluri, 2019). During this progress, endosomal-sorting complexes required for transport (ESCRT)-dependent and independent pathways are both involved (Figure 1).
FIGURE 1.

Exosomes biogenesis and release. Cytomembrane lipids and membrane-associated proteins are clustered in micro-domains for MVB formation. Soluble components, such as cytosolic proteins and RNAs are recruited for exosome cargo sorting. Other cargos can be specifically sorted via the trans-Golgi network into MVBs. Mechanisms of exosome biogenesis are well-accepted by different pathways, basically distinguished as ESCRT-dependent and ESCRT-independent manner. Notably, ESCRT-III is required for the final formation of the ILVs, while cargo sorting and membrane budding can be regulated by either pathways. MVBs will either undergo degradation by lysosomes and autophagosome (finally lysosomes) or to dock or fuse with the cytomembrane. Docking and fusion are the two final processes required for exosome release whereby Rab family members, SNARE, as well as actin are involved. Treatments such as haparanase, adiponection, transferrin, VPS33B, Ral GTPase, Ca2+, IgM, pH, p53 will help to increase exosome production while ISG15, IFN1, Hrs, GW4869 can block exosome secretion by different mechanisms.
However, after exosomes released, the nomenclature and characterization still not come into a consensus. According to the Minimal Information for studies of Extracellular Vesicles (MISEV), three key areas were addressed: EV isolation/purification, EV characterization and EV functional studies (Lotvall et al., 2014). Regarding the characterization of exosomes, at least three positive protein markers (such as CD9, CD63, CD81, Alix, Hsp70), meanwhile no less than one negative protein marker (such as albumin, calnexin) should be testified. Two different but complementary techniques should be used to characterize single vesicles, such as an electronic or atomic force microscope (which can display near and wide fields) and single particle analyzers (Théry et al., 2018).
Secretion
The secretion of exosomes is a process in which MVB fusing with the plasma membrane and different mechanisms are involved (Record et al., 2011; Azmi et al., 2013; Beach et al., 2014; He et al., 2018; Yang et al., 2019). Several ESCRT and relevant proteins including Rab, Sytenin-1, TSG101, ALIX, VPS4, actin and SNARE are participants in exosome release (Baietti et al., 2012; Colombo et al., 2013; Friand et al., 2015; Ciardiello et al., 2016; Jackson et al., 2017; Guo et al., 2020). The Rab family proteins regulate the exosome secretion in a large extent, in which Rab27a and Rab27b are widely reported to regulate exosome secretion (Ostrowski et al., 2010). Mutant Rab11S25N inhibiting exosome release confirms that Rab11 is involved in their release (Savina et al., 2002). Rab35 localized on the cell surface of oligodendroglia can regulate the density of exosomes (Hsu et al., 2010). SNARE proteins and synaptotagmin family members also help the docking and fusing of vesicles with the cytomembrane and to release the contained exosomes into the extracellular environment (Beach et al., 2014).
Other alternative ways to affect exosome secretion is certain kind of inducement or stimulation of the origin cell. An increase of intracellular Ca2+ and activation of protein kinase C promotes both the generation and delivery of exosomes (Nguyen et al., 2016). In cancer cells, the acute elevation of calcium also causes a quintuple increase in the amount of exosomeALIX+CD63+CD9+ (Messenger et al., 2018). Moreover, Transferrin (Tf)-stimulated exosome release is in a calcium ion -dependent manner (Savina et al., 2003). Heparanase involves in almost every related step from biogenesis to release of the exosomes as well as regulating the exosomal protein cargo (David and Zimmermann, 2020; Purushothaman and Sanderson, 2020). Diabetic sera are also found to affect exosome release by disrupting the normal MSC-derived exosome signaling pathway in vitro (Rezaie et al., 2018b) (see more stimulation methods in Table 1). Though some different mechanisms or ways of exosome biogenesis and release are discussed currently, they are not all-inclusive and there still remains plenty room for fully uncovering exosome pathways in all situations.
TABLE 1.
Approaches to affect exosome release.
| Agent | Mechanisms | Models | Production | References |
| Rab27a/Rab27b | Multivesicular endosomes docking | increase | Ostrowski et al., 2010 | |
| Rab11 | Rab11S25N mediated exosome release | K562 | increase | Savina et al., 2002 |
| Rab35 | GTP-dependent localization | Oligodendroglia cells | increase | Hsu et al., 2010 |
| Ca2+ | a. Activation of protein kinase C b. .Munc13-4/Rab11 dependent pathway | Human RBCs MDA-MB-231 | increase | Nguyen et al., 2016; Messenger et al., 2018 |
| PH | a. Low PH induced high rigidity and sphingomyelin content b. .MVB acidification | Melanoma cells Human cells | increase | Parolini et al., 2009; Sung et al., 2020 |
| Transferrin | Ca2+ dependent manner | K562 | increase | Savina et al., 2003 |
| P53 | a. Regulation of TSAP6 b. .IGF-1/AKT/mTOR signaling | BMDCs cells respond to stress | increase | Lespagnol et al., 2008; Feng, 2010 |
| IgM | BCR activation | CLL cells | increase | Yeh et al., 2015 |
| Haparanase | Accumulation of syndecan-1 | MCF-7 | increase | David and Zimmermann, 2020 |
| Adiponectin | Binding to T-cadherin | MSCs | increase | Nakamura et al., 2020 |
| VPS33B | GD12/Rab11a/Rab27a pathway | HSCs, LICs | increase | Gu et al., 2016 |
| RaI GTPase | colocalization with SYX-5 | nematodes, mammary tumor cells | increase | Hyenne et al., 2015 |
| ISG15 | Targeting TSG101 mediated plasma membrane formation | HEK293 | decrease | Nabhan et al., 2012; Choudhuri et al., 2014 |
| IFN1 | ISG15 triggered MVB-lysosome fusion | HEK293T | decrease | Villarroya-Beltri et al., 2016 |
| GW4869 | nSMAse-independent mechanism | myeloma cells SKBR3 cells | decrease | Stoffel et al., 2016; Menck et al., 2017; Vuckovic et al., 2017 |
Transporting Cargos
Transporting cargos in their lipid bilayer membrane, exosomes successfully pass on the message from parent cells to the recipient cells, which includes proteins, nucleic acids and lipids (Tkach and Théry, 2016; van Niel et al., 2018). The mechanisms of how exosome uptake by target cells remains unclear (Villarroya-Beltri et al., 2014), it is more likely to be a dynamic process. Exosomes may enter cells directly through different mechanisms like the participation of calveolae, clathrin, lipid rafts, direct fusion with cell membrane, receptor-ligand interaction, endocytosis by phagocytosis and also micropinocytosis (Kalluri and Lebleu, 2020). However, at the meantime, the endocytosis process also gives rise to the de novo formation of exosomes by cells. Whether the release of exosomes which generate endogenously or take up occurs separately or together is still hard to tell.
Previously, exosomes are considered as vehicles transporting cargos to discard and unnecessary for their cells of origin, but now accumulating evidence confirms that these vesicles as well as their contents are playing full roles in the physiological and pathological process (Phinney and Pittenger, 2017; Bebelman et al., 2018; van Niel et al., 2018; Kalluri and Lebleu, 2020). The comprehensive exosome-mediated crosstalk between different cell types are now attracting increasing attention, exosomes are recognized as a new “cell-free” therapeutic strategy in regenerative medicine and the roles of exosomes in skin development and cutaneous wound healing will be discussed in detail below.
Exosomes and Dermatologic Disease
Under pathological conditions like diabetes or severe burns, skin function is not fully restored via the normal wound healing process, leading to potentially serious complications such as ulcers or infections (Nuschke, 2014). Therefore, it is crucial to complete wound healing with the least delay after injury. Stem cells, particularly MSCs, present great clinical application prospects in skin tissue regeneration for their advantages of extensive sources, easy expansion in vitro and low rejection after transplantation in vivo. It is well accepted now cells like MSCs releasing repair promoting factors in a paracrine manner. Paracrine is the dominating way to facilitate damage repair, whereby exosome outstands (Phinney and Pittenger, 2017; Mehryab et al., 2020; Rahmani et al., 2020). The findings open up the alternative avenue of cells such as MSCs to exosomes for safer, potent and “cell-free” based therapeutic application. A good deal of preclinical experiments has confirmed the multiple roles of exosomes in various skin injury repair (Figure 2 and Table 2).
FIGURE 2.

Main composition of exosomes and their functions on skin injuries. Despite the different origin and mode of biogenesis, exosomes display a similar appearance, and common composition. Exosomes are featured by tetraspanins, receptors or cell-type specific proteins, lipids, adherin proteins, intracellular trafficking proteins. Deep analysis of extracellular vesicle composition reveal that they convey various cargoes, including nucleic acids, proteins (biogenesis elements, chaperones and signaling pathway molecules) and lipids, which all vary widely between cells and conditions. The vast information will directly affect the fate and function of exosomes in skin injuries. Exosomes participate in multiple cutaneous diseases including laceration, burn, aging, diabetic wound healing, scar formation, skin autoimmune disease and skin cancers, through different biological pathways.
TABLE 2.
Functions of exosomes derived from different origin in cutaneous disease.
| Origin | Recipient | Mechanisms | Functions | References |
| hucMSC | fibroblasts and endothelial cells | miR-21-3p/PTEN/SPRY1 | promote proliferation and migration | Hu et al., 2018 |
| hucMSC | fibroblasts | enriched of alpha-2-macroglobulin | improve the activity and migration ability of skin fibroblasts | Bakhtyar et al., 2018 |
| BM-MSC | dermal vascular endothelium |
|
formation of new blood vessels | El-Tookhy et al., 2017 |
| AdMSC | fibroblasts | bFGF and TGF-β PI3K/Akt signaling pathway | increased proliferation and migration | Zhang W. et al., 2018 |
| autologous human epidermal cells | dermal fibroblasts | activating matrix metalloproteinase-1 | cell proliferation and cause the degradation extracellular matrix | Zhao et al., 2017 |
| hucMSC | dermal fibroblasts epidermis cells |
|
promote the proliferation of skin cells re-epithelialization of epidermis orchestrate the distribution collagen Enhance angiogenesis | Zhang et al., 2015a, 2016 |
| hucMSC | dermal fibroblasts epidermis cells | restrict the scar formation | Zhang et al., 2016 | |
| hucMSC | fibroblasts | miR-21, miR-23a, miR-125b and miR-145 targeted TGF-β/SMAD2 | reduce the expression of α-SMA and collagen deposition control scar formation | Fang et al., 2016 |
| hiPSC | human dermal fibroblasts |

|
protective and anti-aging effect | Oh et al., 2018 |
| human umbilical cord blood | epidermis cells |
|
a certain degree of anti-aging effect | Kim et al., 2017 |
| iPSC | human dermal fibroblasts |

|
protect skin from ultraviolet damage | Oh et al., 2018 |
| 3D fibroblasts spheroids | not mentioned | inducing collagen synthesis | Hu et al., 2019 | |
| Platelet-rich plasma | endothelial cells and fibroblasts | YAP activation | regeneration of chronic skin wounds in diabetic rats | Guo et al., 2017 |
| iPSC | fibroblasts | not clear yet 
|
accelerates diabetic wound healing | Kobayashi et al., 2018 |
| gingival MSC | not mentioned | not clear yet | promote the healing and nerve repair | Shi Q. et al., 2017 |
| melanoma cell | MDSC lymphatic endothelial cells | enhance the tolerance capacity of lymph gland to tumors | immune evasion | Hood, 2016 |
| melanoma cell | skin cells |
|
tumor migration and progression | Alegre et al., 2014; Felicetti et al., 2016 |
| SCC cell | microenvironment cells | TGF-β activation | tumor growth and metastasis | McAndrews and Kalluri, 2019 |
| donor MSC | recipient BM-MSC |
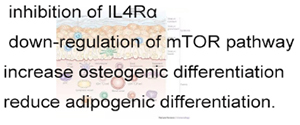
|
caused the rescue of osteopenia impaired BM-MSCs, tight skin, and immune disorders in scleroderma mice | Chen et al., 2017 |
Exosomes and Laceration
Exosomes present favorable repair activity in skin lacerations and full-layer resection. After partial resection of back skin in 12-week-old C57B/6 mice, human umbilical cord MSC derived exosomes (hucMSC-Ex) effectively promote wound angiogenesis and epidermal regeneration in vivo. Further exploration shows that the mechanism is related to miR-21-3p conveyed by exosomes, which regulates cell function by inhibiting its downstream PTEN and SPRY1 target genes (Hu et al., 2018). Protein spectrum reveals that hucMSC-Ex is enriched in a large amount of alpha-2-macroglobulin improve the activity and migration ability of skin fibroblasts (Bakhtyar et al., 2018). Bone marrow MSC (BM-MSC) derived exosomes can accelerate the healing of wound by promoting the neovascularization in the dermal laceration model of large animal dogs, and the mechanism of this repair effect is also related to the fact that BM-MSC exosomes contain various active factors related to angiogenesis (El-Tookhy et al., 2017). Adipose-derived MSC (AdMSC) exosomes are internalized and ingested by skin fibroblasts to accelerate the proliferation and migration of target cells. Expression levels of repair related factors such as collagen, bFGF and TGF- beta are increased, and the PI3K/Akt signaling pathway is activated by AdMSC exosomes in damaged skin cells to accelerate the repair process (Zhang W. et al., 2018). Additionally, dermal fibroblasts can also internalize autologous human epidermal cells derived exosomes in order to promote the proliferation of cell and cause the degradation of some extracellular matrix by activating matrix metalloproteinase-1(MMP-1). Exosomes derived from human epidermal cells are applied in the full-layer skin excision model of rats, whereby the epidermal regeneration is basically completed within two weeks, which also have significant healing effect (Zhao et al., 2017).
Exosomes and Burn
Burns is a common injury in daily life, which occurs in 50 to 100 cases per 10,000 people annually (Lewis, 2013). At present, the treatment of large-scaled scald is mainly through systemic support, wound cleaning, scab excision and surgical skin grafting (Calota et al., 2012), which are difficult to grasp the scab depth and easy to be infected, and in source tension of skin grafts (Eisenbeiß et al., 2012; Lootens et al., 2013; Nacer Khodja et al., 2013). The skin deep burn and scald within 3 weeks is difficult to repair completely by itself, and is easy to form scar tissue (Cuttle et al., 2006; Ribeiro et al., 2009). Thus, shortening the wound healing time is particularly important. In our previous study, we successfully established the deep secondary burn injury model in SD rats, and found that hucMSC-Ex accelerated the cutaneous wound recovery by facilitating the proliferation of skin tissue cells and the re-epithelialization of epidermis, as well as orchestrating the rational distribution of collagen in skin tissue and enhancing angiogenesis (Zhang et al., 2015a,b). Interestingly, long-term (more than 4 weeks) observation showed that hucMSC-Ex conversely inhibited the proliferation of skin cells and the activation of β-catenin, meanwhile, hucMSC-Ex restricted the amplification of skin stem cells, the expression of α-SMA in dermal cells and the deposition of collagen III. Along with the healing process, the cell density varies from low to high, which triggers the Hippo signaling activation. Mechanically, hucMSC-Ex could deliver 14-3-3 zeta protein to combine YAP and LATs, thus speeding up the phosphorylation of YAP to limit the continuous activation β-catenin (Zhang et al., 2016). The dynamic regulation pattern prevents excessive hyperplasia of skin tissue and scar formation after injury (Figure 3).
FIGURE 3.

The dynamically regulatory role of exosomes in burn injury. In the early stage of repairing, cells are in a low density, exosomal Wnt4 activates β-catenin in skin cells to promote proliferation and migration, meanwhile stimulates vascular endothelial cell to enhance angiogenesis. With the progression of wound healing, cell density becomes from low to high, the cell-cell contact triggers the activation of Hippo signaling, leading to p-LATS accumulation. 14-3-4ζ released by exosomes works like a “table” to facilitate the combination of YAP and LATS, so as to accelerate the phosphorylation of YAP, the phosphorylated YAP shows a intracellular retention rather than nuclear translocation to suppress the down-stream gene expression and finally inhibit the cell proliferation and migration. The exosomes mediated dynamic regulation in different stage of wound healing, works as an “accelerate” to promote wound healing in early stage, while as a “brake” to prevent excessive proliferation and scar formation.
Exosomes and Scars
During the proliferation period of skin injury repair, fibroblasts, epidermal cells and vascular endothelial cells jointly promote the formation of vascular network, and in the meantime, cells migrate from the edge to the surface of the wound, reducing the area of the wound surface to promote the smooth progress of epidermal regeneration. In this process, excessive activation and recruitment of fibroblasts under pathological conditions often lead to scar formation and fibrosis (Wynn and Ramalingam, 2012; Van De Water et al., 2013). In our studies, hucMSC-Ex can inhibit tissue over-proliferation and scar formation by activating Hippo signaling pathway in the later stage of burn injury repair, providing a guarantee for the security of exosome in vivo (Zhang et al., 2016). In addition, it is worth noting that the TGF-β family plays a positive regulatory role in cell differentiation, proliferation and metabolism during tissue injury and repair, while the pathological continuous activation of TGF-β activates the SMAD2 signaling pathway, which can easily lead to fiber deposition and scar formation (Frei et al., 2015). HucMSC-Ex could effectively inhibit the activation of fibroblasts in mice after full-layer resection of dorsal skin. A key group of miRNAs, miR-21, miR-23a, miR-125b and miR-145, could inhibit the activation of fibroblasts by targeting the TGF-β/SMAD2 signaling pathway, reduce the expression of α-SMA and collagen deposition, so as to control scar formation (Fang et al., 2016). At the same time, researchers also propose to conduct gene transfection modification on hucMSC-Ex based on this group of miRNA molecules, so as to acquiring a better and safer therapeutic strategy.
Exosomes and Aging
Skin aging is mainly for the genetic factors and easily found in exposure and the exposed parts, with wrinkles and sagging as the main characteristics. Pathologically, the skin epidermis and the dermis thickness become thinner, dermal collagenous fibers and collagen content declines with neatly arranged collagen bundles, clinically characterized by skin with fine wrinkles. hiPSC exosomes are used to confront aging skin, the expression of senescence-associated-β-galactosidase, a natural aging marker, is significantly down-regulated, and the expression of type I collagen in aging fibroblasts increased, thus playing a good protective and anti-aging effect on human dermal fibroblasts (Oh et al., 2018). Human umbilical cord blood derived exosomes penetrate into the epidermal layer of skin within 15 h, and reach a certain degree of anti-aging effect by regulating the expression and distribution of collagen and elastin in skin cells (Kim et al., 2017) and MSCs derived exosomes can also ameliorate aging and age-related diseases (Ahmadi and Rezaie, 2021).
Exosomes and Ultraviolet Damage
Ultraviolet damage of skin is the accumulation of ultraviolet damage in the process of skin aging, which is the result of the interaction of endogenous aging and ultraviolet radiation, featured tissue disintegration, fragmentation and collagen bundle dispersion (Bellei et al., 2018). Matrix metalloproteinases (MMPs) specifically degrade almost all extracellular matrix (ECM) components and play a vital role in photoaging of skin. Photoaging is found to gradually impair the function of human dermal fibroblasts (HDFs), which generate collagen and matrix metalloproteinase in dermis (Jeong et al., 2015; Zorin et al., 2017; Hu et al., 2019). The iPSCs-Ex can increase the expression level of collagen type I in the photo-aged HDFs and reduce the expression level of SA-β-Gal, MMP-1/3 and restore the collagen type I expression in senescent HDFs (Oh et al., 2018). Another interesting research compared exosomes derived from monolayer culture of HDFs (2D HDF-XOs) with that from the three-dimensional spheroids (3D HDF-XOs). The result showed that there was a notably higher level of TIMP-1 and differentially express miRNAs in 3D HDF-XOs (Hu et al., 2019). 3D HDF-XOs mainly work through the downregulation of TNF-α and the upregulation of TGF-β to induce collagenation and antiaging. Additionally, an interdisciplinary study showed that marine sponge spicules (SHSs) significantly enhanced the skin targeted delivery of hucMSC-Ex, which also revealed anti-photoaging effects in mice, including reducing micro-wrinkles and enhancing the expression level of extracellular matrix, and facilitating the skin to recover shortly (Zhang K. et al., 2020).
Exosomes and Diabetic Wound Healing
Diabetic foot ulcer (DFU) is one of the most common and major complications of diabetes. At present, the pathogenesis of DFU is considered to be the result of the interaction between peripheral blood vessels and peripheral neuropathy. The complexity of the pathogenesis and the difficulty of wound healing make it one of the problems that threaten the life and health of diabetic patients (Iversen et al., 2009; Goodall et al., 2020). Our previous research has confirmed that hucMSC-Ex can alleviate insulin resistance in type 2 diabetic individuals, increase the distribution of GLUT on the surface of liver, muscle, fat and other tissue membranes, and improve the amount and activity of glucose metabolism enzymes in tissues such as liver, muscle and fat, promote the body’s intake and utilization of glucose in the blood to reduce blood sugar in individuals with type 2 diabetes. Specific mechanisms such as exosome directly carrying GLUT molecules to receptor cell (liver, muscle, and fat) membranes, which facilitate extracellular glucose transportation, provide a basic mechanism for exosomes to treat diabetes-related complications (Sun et al., 2018). Conversely, study indicated that diabetes could decrease angiogenic and restorative effect of stem cells in vitro (Rezaie et al., 2018a). In diabetic mice, human fibroblast-derived exosomes can promote the acceleration of wound repair (Geiger et al., 2015). The platelet-rich plasma-derived exosomes can activate YAP and promote the regeneration of chronic skin wounds in diabetic rats (Guo et al., 2017). iPSC exosomes enhance fibroblast migration and proliferation, accelerates wound healing and local nerve regeneration in diabetic foot (Kobayashi et al., 2018). The gingival MSC exosome and the synovial MSC exosome have also been reported to promote the healing and nerve repair of diabetic ulcer wounds, however, the specific mechanism requires to be refined (Shi Q. et al., 2017).
Exosomes and Skin Cancer
Exosomes are closely related to tumorigenesis, such as U87 MG human glioblastoma, colorectal cancer, skin cancer (Ahmadi et al., 2021; Soraya et al., 2021). Study showed that exosomal cargos modulate autophagy and play pivotal roles in neoplasis, tumor heomostasis and metastasis (Hassanpour et al., 2020; Jafari et al., 2020; Salimi et al., 2020). Exosomes in skin cancer microenvironment are secreted and enriched, provide cancerous cell the platform to metastasize and attacking the host’s immune system (Gowda et al., 2020). Melanoma is the most malignant of skin neoplasms, and melanoma cell-derived exosomes affect the sentinel lymph node genes in a variety of ways to promote tumor progression. In melanoma cells, Wnt5a promotes the release of immunoregulatory and proangiogenic factors from exosomes (Ekström et al., 2014), while melanoma cell-derived exosomes can regulate lymphatic endothelial cells and myeloid-derived suppressor cells (MDSCs) to enhance the tolerance capacity of lymph gland to tumors, finally achieving immune evasion (Hood, 2016). Metastatic melanomas release exosomes carrying PD-L1, which also be able to suppress CD8+T cells function and facilitates tumor growth (Chen et al., 2018). Besides, exosomes can also contain miRNAs, such as miR-222, activates PI3K/AKT signaling pathways in skin cells to promote tumorigenesis (Felicetti et al., 2016). Human metastatic melanoma serum exosomes show a high expression level of miRNA-17, miRNA-19a, miRNA-126 (Pfeffer et al., 2015). MiRNA-125b is selectively expressed by tumor cells due to its ability to inhibit tumor-associated c-Jun protein expression (Alegre et al., 2014). Due to tissue-specific miRNAs can be encapsulated in EVs and distributed in multiple biological fluids, they may serve as a tumor biomarker in neoplasia early progression (Jafari et al., 2019).
In squamous cell carcinoma (SCC), exosomes also serve as an important vehicle for tumor and microenvironment communication, and reactivate tumor-associated signals such as TGF-β in order to promote metabolism and growth of cancer cells (McAndrews and Kalluri, 2019).
Exosomes and Other Skin Lesions
Scleroderma is characterized by inflammatory, degeneration, thickening, and fibrosis, which in turn is hardened and atrophied, thus causing multiple system damage. Studies have reported a significant reduction in exosome levels in peripheral blood serum from patients with scleroderma, which is highly likely to be associated with impaired exosome transport system caused by vascular abnormalities (Nakamura et al., 2016a). This decrement of exosome production in scleroderma patients’ serum may be attributed to the disordered transport of exosomes which is from the skin epithelium to the blood flow, resulting in the down-regulation of collagen I that delays wound repair, leading to a higher susceptibility to cutaneous ulcer (Wermuth et al., 2017). Recently, it has been reported that miR-151-5p is a diagnostic and therapeutic potential biomarker of scleroderma. Chen et al. (2017) found that exosomal miR-151-5p released from donor MSCs could inhibit the expression level of IL4Rα and suppress the activation of mTOR pathway, which leads to an increase in osteogenic differentiation and decrease in adipogenic differentiation. Further, miR-151-3p delivered systemically protected the scleroderma mice from immunologic derangement, tight skin, osteopenia and impaired bone marrow MSCs (Chen et al., 2017). Wermuth et al. (2017) found that exosomes derived from scleroderma patients cell culture supernatant were associated with the stimulation of genes expression, including collagen type III alpha 1 chain, COL1A1 and fibronectin-1 encoding ECM components (Chen et al., 2017). In profiles of neutrophils exosomes derived from scleroderma patients, 281 dysregulated lncRNAs and 22 dysregulated miRNAs are identified as well as further analysis were conducted of their target genes and downstream signaling pathways, which could be potential therapeutic targets and biomarkers in scleroderma (Li L. et al., 2020).
Psoriasis is an autoimmune disease of the skin, often accompanied by excessive proliferation of keratinocytes, while iron reserves often affect the working state of the patient’s immune system. Some researchers have found that important proteins regulating iron reserve such as hepcidin and soluble transferrin receptor are at low expression levels in serum exosome of patients with psoriasis, presumably affects immune function and thus affects the course of the disease (El-Rifaie et al., 2018).
Exosomes Participate in Various Skin Damage Repair Mechanisms
Coagulation
Exosomes can be extracted from saliva, mononuclear macrophages, and platelet secretions. After skin trauma, obstruction of coagulation function affects the repair time of wounds, and the blocked coagulation function is relevant to the change of tissue factor (TF) conformation or the presence of TF inhibitory molecules. Saliva-derived exosomes can shorten wound clotting time by transporting active TF in combination with factor VII. At the same time, these different sources of exosomes secreted into the intracellular space can also activate coagulation by transmitting TF, for example, exosome secreted by monocytes can bind to platelets and release their own TF into platelets to activate downstream cascades to promote thrombin and fibrin clot formation (Biró et al., 2003; Del Conde et al., 2005).
Proliferation
Proliferation of skin cells plays a vital role in skin damage repair. At present, exosomes from various tissue and cell regulate the proliferation of skin cells, such as MSC exosomes (Du et al., 2014; Shabbir et al., 2015; Jiang et al., 2020), mouse ESC exosomes (Jeong et al., 2014), endothelial progenitor cell-derived exosomes (Li et al., 2016) and fibroblast-derived exosomes. These exosomes functioning in regulating cell proliferation in two ways, activate cell cycle related genes or increase growth factor expression levels. The exosomal RNAs or proteins are delivered to the target cell, such as Wnt4 transportation to motivate the β-catenin signaling pathway in the epidermal cell (Zhang et al., 2015a), or delivering the small RNAs to target the downstream Akt, Erk, MAPK signaling pathway, etc. (Zhang et al., 2016; Guo et al., 2017), ultimately attributed to the regulation of cell cycle-related genes such as cyclinA1, cyclinD1, cyclinD3 to promote cell proliferation. Additionally, exosomes can also activate interleukin-6 (IL-6), insulin-like growth factor-1 (IGF-1), vascular endothelial growth factor (VEGF) and other growth factors, which stimulate damaged skin cells proliferation. The proliferative effect of exosomes can be caused both by paracrine affecting surrounding cells and autocrine to de novo affect itself.
Migration
Proliferation and migration of cells are pivotal for wound healing. With the onset of the clotting period, immunoinflammatory cells such as macrophages and neutrophils are first recruited to the damaged site to get rid of necrotic tissue fragments as well as phagocytose infected microbe. Subsequently, epidermal cells and dermal fibroblasts begin to migrate to the injured sites to fill the vacancies and promote wound healing (Singer and Clark, 1999). In this process, exosomes from various cells are involved in the regulation of cell migration, such as MSCs, keratinocytes, endothelial cells and fibroblast-derived exosomes (Cheng et al., 2008). Keratinocyte-derived exosomes can promote their own migration by transporting heat shock protein 90α (HSP90α) in an autocrine manner. In addition, keratinocyte-derived exosomes can also promote the migration of vascular endothelial cells, thereby promoting the regeneration and repair of blood vessels (Leoni et al., 2015). Moreover, keratinocyte-derived exosomes enhance the migration of dermal fibroblasts by transporting HSP 90α, which mechanically relate to its interaction with cell surface receptor LRP-1/CD91 (McCready et al., 2010). MSC exosomes regulate fibroblast migration by activation of Erk1/2 signaling pathway, resulting in up-regulating the expression of IL-6, cyclin1, MMP-1, N-cadherin and other factors in target cells (Huang et al., 2015; Hu et al., 2016).
Vascularization
Angiogenesis provides oxygen for repairing tissues and supply nutrients. The proliferation of vascular endothelial cells is an essential constituent of it. Moreover, it also requires interaction with angiogenic, endothelial cells and the surrounding extracellular matrix environment (Liekens et al., 2001). The wound area exhibits chemotactic effect on vascular endothelial cells, enabling endothelial cells to penetrate the vascular basement membrane, enter the extracellular matrix, form a lumen-like structure and continuously extend and branch, and finally generate a new vascular network. In this process, human MSC and endothelial cell-derived exosome can increase the density of neovascularization in the wound area, which is related to the promotion of endothelial cell proliferation and migration. In addition, endothelial progenitor cells and platelet-rich plasma-derived exosomes can activate the expression of pro-angiogenic genes in target cells, such as IL-8, IL-6, fiber growth factor 2, E-selectin, etc. The expression of these factors will further activate Erk1/2 and other related signaling pathways, activate downstream VEGFA, VEGFR-2, c-Myc, cyclinD1 and other target genes, thereby further promoting neovascularization (Hromada et al., 2017). In our previous studies, we found that hucMSC-Ex repaired wounds by promoting Wnt4/β-catenin signaling pathway in vascular endothelial cells (Zhang et al., 2015a). Conversely, some researchers find exosomes to inhibit the formation of blood vessels by binding to CD36 on the surface of target cells (Ramakrishnan et al., 2016). Therefore, despite of a great potential in regulating angiogenesis, it is still necessary to specifically analyze the source, type and specific pathological conditions when exosomes are used.
Skin Stem Cell Mobilization and Participation
Stem cells are the important reason to explain the super self-repair ability of skin. Epidermal stem/progenitor cells generally locate in the basal layer of the epidermis to meet the daily keratin renewal metabolism or the need to fulfill new cells after skin damage. The proliferation and differentiation of skin stem cells are affected by multiple factors, and finally achieve the balance between proliferation and differentiation (Li and Sen, 2016; Yang et al., 2020). In the model of deep second-degree scald repair in rats, we confirm that hucMSC-Ex can activate CK19 and α6 integrin-positive epidermal stem cells in the early stage of repair, and make the epidermal stem cells return to resting state after the repair is completed. The mechanism may be related to the dynamic regulation between Wnt/β-catenin signaling pathway and Hippo/YAP signaling pathway (Zhang et al., 2016). In addition, epidermal stem cells are involved in epidermal regeneration in the early post-traumatic period, but do not play a role in late and normal epidermal renewal. Exosomes from autologous stem cells are also effective in promoting proliferation, angiogenesis and wound healing of damaged skin cells (Lu et al., 2019).
Immunoregulation
After skin trauma and infection, a great quantity of immune cells are recruited to the lesion region, Exosomes transport pathological nucleic acids, proteins and lipids, and take part in antigen-promoting, immune response-related receptors activation, and defense induction, which therefore arouse interests to consider exosomes as good immune-based therapeutics and vaccine candidates (Schorey and Harding, 2016).
Skin and immune cells mediate the corresponding immunological effects by secreting exosomes. Keratinocytes expedite the stimulation and maturation of dendritic cells by secreting exosomes, enhance the expression of CD40, and produce a large number of cytokines such as IL-6, IL-10, IL-12 (Kotzerke et al., 2013). Exosomes from circulating immunological cells such as monocytes can also promote the expression of MMP-1 in dermal fibroblasts by transporting 14-3-3 protein, thereby promoting cell migration to the wound area (Medina and Ghahary, 2010). Moreover, MSC exosomes are involved in autologous skin transplantation by regulating the inhibitory immune cell Treg (Zhang B. et al., 2018; Showalter et al., 2019), and exosomes from sweat can also regulate skin immunity, which is the donor exosomes trigger alloreactive T cell responses after transplantation (Marino et al., 2016). Furthermore, the M2 Macrophage-derived exosomes (M2-Exo) induce M1-M2 conversion in vitro to promote wound healing by facilitating neovascularization, collagen deposition as well as re-epithelialization (Kim et al., 2019). In recent years, immune cell such as dendritic cell-derived exosomes are used to stimulate T cells to activate host anti-tumor immunity, and have entered Phase I and Phase II clinical trials (Besse et al., 2016; Figure 4). Conversely, some studies have found that after skin transplantation, some donor-derived exosomes unexpectedly trigger an immune response and attack receptor tissue cells by activating autologous T-cell immunity, which reminds us the immune system in skin is quite a complex network that every type of cells inside is possibly to communicate with other cell types nearby or in a distance, some of the “cross-talk” mechanisms are unlocked while some are not, the function and application of exosomes in immune regulation remains to be more fully lucubrated (Morelli et al., 2017; Benichou et al., 2020).
FIGURE 4.

The immunoregulatory roles of exosomes in skin diseases. The skin relies on the immune system for essential functions, such as thermoregulation and infection control. The wound healing process is a complex one with at least three stages, while immune cells participate in them all. Neutrophils and macrophages infiltrate into the wound healing as soon as the inflammatory stage begins. The wound area is a complicated microenvironment that cells inside of it like epidermic cells, fibroblasts, stroma cells can interact with immune cells including T-cells, Tregs, dendritic cells and macrophages by releasing specific bio-molecules, jointly affect the fate of disease.
Extracellular Matrix Remodeling
Exosomes are involved in the hemorrhagic, inflammatory and proliferative phase of skin damage repairing as well as the tissue remodeling. Exosomes from various cells have shown an effective regulatory function on extracellular matrix. Elastin is an important component of the extracellular matrix maintenance structure, and exosomes can promote elastin expression during tissue remodeling (Zhang et al., 2015c). Collagen mainly secreted by fibroblasts also takes part in tissue remodeling. Adipose-derived MSC exosomes can effectively promote the proliferation and secretion capacity of dermal fibroblasts (Fang et al., 2019). Exosomes are also able to promote collagen formation in the extracellular matrix through its membrane surface LOXL2 protein (de Jong et al., 2016). Other identical studies revealed that exosomes could be isolated in fiber networks, demonstrating their involvement in the formation and functional regulation of extracellular matrices (Huleihel et al., 2016). In addition, exosomes can accelerate the repairing process and restrict scab size by interacting with the Annexin A1 and formyl peptide receptors to complete tissue remodeling (Nakamura et al., 2016b).
Application Prospect and Open Question
Exosomes and Scar Free Treatment
Skin wound healing is often accompanied by scab and scar formation, when the skin tissue is seriously damaged, exceeding its self-healing capacity, the wound area will be replaced by fibrous scars. The atrophy period of scar formation will bring physiological discomfort to the patient, and after the scar formation will certainly affect the local appearance and function. Our previous findings indicate that hucMSC-Ex retain YAP in the cytoplasm by transporting the active 14-3-3 molecule at the later stage of repairing, thus inhibiting the excessive proliferation of skin cells and the formation of fibrosis (Zhang et al., 2016). HucMSC-Ex also inhibit scar formation by inhibiting TGF- beta/SMAD2 pathway activation in fibroblasts by transshipping mir-23a, mir-125b and mir-145b, etc., thereby inhibiting myofibroblast activation (Chen J. et al., 2019). Meanwhile, AdMSC exosomes can reduce scar area and inhibit the differentiation of fibroblasts into myofibroblasts, mechanically related to the inhibition of TGF- beta 1 signaling and as well as ERK/MAPK signaling activation in fibroblasts (Wang et al., 2017). However, scars such as hypertrophic scars, resulting from alterations in the normal processes of cutaneous wound healing are finally featured by excessive deposition of collagen, persistent inflammation and fibrosis. New therapies including exosomes are designed to no more than minimizing scarring and accelerating wound healing immediately to avoid scar formation at an early stage. The hypertrophic scar treatments and even the keloid treatments still call for intensive studies.
Exosomes as Bio-Markers for Cutaneous Diseases
Exosomes are carrying a variety of bioactive nucleic acids or proteins, which have been widely studied and reported as diagnostic markers of various diseases. Both the quantity and contents may alter when pathological conditions occur. In comparison with healthy controls, serum derived exosomes are more secretion in cutaneous diseases, especially tumors such as melanoma. Moreover, exosomes derived from melanoma patients are proved to convey an increased level of S100B (Alegre et al., 2016; Isola et al., 2016). Due to EV-related miRNAs are tissue-specific and exist in most biological fluids, they may serve as a tumor biomarker in early diagnosis, treatment response and prognosis of cancer (Mirzaei et al., 2016; Chen L. et al., 2019; Rahbarghazi et al., 2019). Exosomal miR-17, miR-19a, miR-21, miR-126, and miR-149 were increased in serum of patients diagnosed with melanoma (Chen L. et al., 2019). The expression level of exosomal miR-222 derived from melanoma samples was consistent with the metastatic capacity of melanoma cells (Felicetti et al., 2016). With the exception of skin tumors, the numbers of exosomes in the serum of patients with scleroderma is also remarkably higher than healthy group, which was of certain diagnostic value.
Recently, by using mass spectrometry and RNA sequencing, vast information has been acquired, which can be reference to the disease of interest. For example, profiles of neutrophils derived exosomes revealed 281 dysregulated lncRNAs and 22 dysregulated miRNAs as well as the predicted target genes and reactome pathways such as Wnt, IL-23, AMPK, and NOTCH signaling in systemic sclerosis, which could be high diagnostic value in the disease (Li L. et al., 2020). A mass spectrometry-based proteomic characterization of melanoma exosomes identified 1507 tumor-related proteins, most of which participated in cell proliferation, migration, EMT and neovascularization, which indicated a potential for further research identifying new biomarkers (Surman et al., 2020). Nevertheless, to be a competent diagnostic marker, more comprehensive case data and criteria are needed to promote its transformation from basic science to clinic. In order to realize the diagnostic and prognostic potential of exosomes containing distinct biological markers, it is essential to evolve the proper exosome isolation methods. Effective and clean without contamination separation of exosomes in circulation should be conducted to gain valid information for clinical application, the selected cargos should be specific and sensitive.
Tailored Exosome as New Therapeutics for Cutaneous Disease
The positive role of exosomes in cutaneous wounds has been continuously recognized. At present, a craze of engineering modification of exosomes has been set off in the research field, which maximizes the advantages of exosome by gene manipulation of origin cells, drug or therapeutic molecular loading by physical incubation, the freeze-thaw cycling, electroporation, sonication and other techniques (Figure 5). The well-designed exosomes could erase the limitations of natural exosomes and increase the targeting capability, half-life in circulation as well as the cargo concentrations.
FIGURE 5.

The bioengineering modification of exosomes. This is a simplified illustration of methods for modification of therapeutics into exosomes. The methods are basically divided into two parts, the gene manipulation modification and the post-translational modification. The first one is to use virus mediated system or gene editing techniques to modify the parental cells before exosome extraction. The other way is to directly modify exosomes after the isolation process by various method including incubation, electroporation, sonication, extrusion, hypotonic dialysis, freeze-thaw cycles, saponin, CaCl2 and lipofectamine reagent.
The endogenous way to get proteins encapsulated into exosomes is mainly by protein fusion which depends on the available cellular machinery for classifying therapeutic proteins into exosomes (Nasiri Kenari et al., 2020). Proteins such as lactadherin, lamp2b, which exist on the membrane of exosomes are used to deliver therapeutic siRNA, miRNA to the site of interest, tetraspanin family members CD9, CD63, CD81 are mostly used as a candidate for fusion of fluorescent proteins to trace exosomes in vitro and in vivo. In addition, tailored exosomes by transferring certain specific miRNA or siRNA has also been explored for disease treatment. By gene manipulation of synovial MSCs, exosomes express a high level of miR-126 to promote the angiogenesis in skin wound (Tao et al., 2017). Other pre-treatment of the origin cells like chemical agents, hypoxia is also effective for exosome modification. Melatonin-pretreated MSC-derived exosomes can enhance diabetic wound healing by regulating macrophage M1-M2 conversion (Liu et al., 2020). In our previous study, we found that small molecule drugs 3,3′-diindolymethane (DIM), as a natural small-molecule drug extracted from cruciferous plants, can activate hucMSC-Ex and enhance the dry and paracrine ability of MSC by carrying active Wnt11 molecules, so as to enhancing the repairing effect of deep second-degree scald wounds (Shi H. et al., 2017).
Exosomes also can be modified exogenously after their isolation process. Approaches including electroporation, incubation, sonication, extrusion, hypotonic dialysis, freeze-thaw cycles, saponin, CaCl2 and lipofectamine reagent are used for exosomes loading. Sources such as platelet derived exosomes loaded with curcumin on polysaccharide, can more effectively promote diabetic foot ulcer wound repair (Brennan et al., 2020). MSC derived exosomes assembled in a chitosan/silk hydro gel (Shiekh et al., 2020) and in a self-healing, anti-bacterial polypeptide hydro gel (Wang et al., 2019) promoted diabetic wound healing. It can be predicted that exosome modification will make it play its maximum advantages in cutaneous wound healing. Meanwhile, it remains to be determined the specific mechanism of cargo loading, release, uptake, and destiny of vesicle in recipient tissue.
Future Application Prospects and Problems of Exosomes
Exosomes can significantly repair cutaneous damage, provide new therapeutic strategies for the treatment of various diseases, and lay an important foundation for biopharmaceuticals based on exosome modification. However, there still exists many unanswered questions for exosome utilization from basic science to clinic: (1) The scalability of exosomes is a major challenge currently. The future clinical use for exosomes depends on a mass-produced and GMP-compliant production process. New approaches like tangential flow filtration can concentrate large quantities of cell-conditioned media for exosome enrichment (Heinemann et al., 2014). (2) There is an urgent need for precise and accurate characterization of exosomes to better evaluate their heterogeneity, their cargo, and functions. A new criterial for exosome identification is put forward by MISEV (Théry et al., 2018). (3) The use of supraphysiological numbers of exosomes still needs to be re-considered, for example, the dose, time duration as well as the route of administration. (4) It is still unclear that how the multiple information conveyed by exosomes can be fully used to develop more precise or early-phase biomarker to help clinical decision making. (5) The application of exosomes as therapeutics relies on producing homogeneous exosomes on a large scale by finding optimal producer cells, culture conditions as well as stimulation conditions, subsequently, getting the approval by the FDA for clinical applications. A large amount of studies has demonstrated the role of exosomes, especially MSC-exosomes in tissue regeneration, immunomodulation and tumor. However, exosomes under different conditions may have opposite effect, for example, exosomes from aged MSC shows no effect on skin regeneration. Further investigation is still required to uncover the multifaceted function of exosomes.
Ultimately, although we have already acquired certain stock of knowledge on exosomes and their functions on skin biology, there still much room for exploration on the potential of exosomes in dermatologic disease is diagnose and treatment.
Author Contributions
HS did the conceptualization. HS and YS wrote. MW and DY collected the data. HQ and WX reviewed and edited. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
- MVB
multivesicular body
- MSC
mesenchymal stem cells
- ILV
intraluminal vesicles
- EV
extracellular vesicle
- ESCRT
endosomal sorting complex required for transport
- TSG101
tumor susceptibility gene 101 protein
- ALIX
ALG-2 interacting protein X
- VPS
vacuolar protein sorting-associated protein
- SNARE
soluble NSF Attachment Protein Receptor
- DPC-Ex
dermal papilla cells derived exosomes
- MAPK
mitogen-activated protein kinase
- EMT
epithelial-to-mesenchymal transition
- MSC
mesenchymal stem cell
- hucMSC
human umbilical cord derived MSC
- BM-MSC
bone marrow derived MSC
- hiPSCs
human induced pluripotent stem cells
- MMPs
matrix metalloproteinases
- 3D
three-dimensional
- TIMP-1
tissue inhibitor of metalloproteinases-1
- SHSs
sponge sp. spicules
- DFU
diabetic foot ulcer
- GLUT
glucose transporter
- MDSC
myeloid-derived suppressor cell
- SCC
squamous cell carcinoma
- TF
tissue factor
- IGF-1
insulin-like growth factor-1
- IL-6
interleukin-6
- VEGF
vascular endothelial growth factor
- HSP90α
heat shock protein 90α
- AdMSC
adipose derived MSC
- DIM
3,3 ′-diindolymethane
- MISEV
minimal information for studies of extracellular vesicles
- FDA
Food and Drug Administration.
Funding
This work was supported by the National Natural Science Foundation of China (Grant 81971757), National Natural Science Youth Foundation of China (Grant 82001975), the Natural Science Youth Foundation of the Jiangsu Province (Grant BK20190841), the National Post-doctoral Science Foundation of China (Grant 2020M671371), and Zhenjiang Key Laboratory of High Technology Research on Exosomes Foundation and Transformation Application (Grant ss2018003). Introduction program of high-level innovative and entrepreneurial talents in Jiangsu province.
References
- Ahmadi M., Rezaie J. (2021). Ageing and mesenchymal stem cells derived exosomes: molecular insight and challenges. Cell Biochem. Funct. 39 60–66. 10.1002/cbf.3602 [DOI] [PubMed] [Google Scholar]
- Ahmadi M., Jafari R., Mahmoodi M., Rezaie J. (2021). The tumorigenic and therapeutic functions of exosomes in colorectal cancer: opportunity and challenges. Cell Biochem. Funct. 39 468–477. 10.1002/cbf.3622 [DOI] [PubMed] [Google Scholar]
- Akbari A., Rezaie J. (2020). Potential therapeutic application of mesenchymal stem cell-derived exosomes in SARS-CoV-2 pneumonia. Stem Cell Res. Ther. 11:356. 10.1186/s13287-020-01866-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alegre E., Sanmamed M. F., Rodriguez C., Carranza O., Martín-Algarra S., González A. (2014). Study of circulating microRNA-125b levels in serum exosomes in advanced melanoma. Arch. Pathol. Lab. Med. 138 828–832. 10.5858/arpa.2013-0134-OA [DOI] [PubMed] [Google Scholar]
- Alegre E., Zubiri L., Perez-Gracia J. L., González-Cao M., Soria L., Martín-Algarra S., et al. (2016). Circulating melanoma exosomes as diagnostic and prognosis biomarkers. Clin. Chim. Acta 454 28–32. 10.1016/j.cca.2015.12.031 [DOI] [PubMed] [Google Scholar]
- Azmi A. S., Bao B., Sarkar F. H. (2013). Exosomes in cancer development, metastasis, and drug resistance: a comprehensive review. Cancer Metastasis Rev. 32 623–642. 10.1007/s10555-013-9441-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babaei M., Rezaie J. (2021). Application of stem cell-derived exosomes in ischemic diseases: opportunity and limitations. J. Transl. Med. 19:196. 10.1186/s12967-021-02863-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baietti M. F., Zhang Z., Mortier E., Melchior A., Degeest G., Geeraerts A., et al. (2012). Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat. Cell Biol. 14 677–685. 10.1038/ncb2502 [DOI] [PubMed] [Google Scholar]
- Bakhtyar N., Jeschke M. G., Herer E., Sheikholeslam M., Amini-Nik S. (2018). Exosomes from acellular Wharton’s jelly of the human umbilical cord promotes skin wound healing. Stem Cell Res. Ther. 9:193. 10.1186/s13287-018-0921-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beach A., Zhang H. G., Ratajczak M. Z., Kakar S. S. (2014). Exosomes: an overview of biogenesis, composition and role in ovarian cancer. J. Ovarian Res. 7:14. 10.1186/1757-2215-7-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bebelman M. P., Smit M. J., Pegtel D. M., Baglio S. R. (2018). Biogenesis and function of extracellular vesicles in cancer. Pharmacol. Ther. 188 1–11. 10.1016/j.pharmthera.2018.02.013 [DOI] [PubMed] [Google Scholar]
- Bellei B., Migliano E., Tedesco M., Caputo S., Papaccio F., Lopez G., et al. (2018). Adipose tissue-derived extracellular fraction characterization: biological and clinical considerations in regenerative medicine. Stem Cell Res. Ther. 9:207. 10.1186/s13287-018-0956-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benichou G., Wang M., Ahrens K., Madsen J. C. (2020). Extracellular vesicles in allograft rejection and tolerance. Cell. Immunol. 349:104063. 10.1016/j.cellimm.2020.104063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Besse B., Charrier M., Lapierre V., Dansin E., Lantz O., Planchard D., et al. (2016). Dendritic cell-derived exosomes as maintenance immunotherapy after first line chemotherapy in NSCLC. Oncoimmunology 5:e1071008. 10.1080/2162402x.2015.1071008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biró E., Sturk-Maquelin K. N., Vogel G. M., Meuleman D. G., Smit M. J., Hack C. E., et al. (2003). Human cell-derived microparticles promote thrombus formation in vivo in a tissue factor-dependent manner. J. Thromb. Haemost. 1 2561–2568. 10.1046/j.1538-7836.2003.00456.x [DOI] [PubMed] [Google Scholar]
- Brennan M., Layrolle P., Mooney D. J. (2020). Biomaterials functionalized with MSC secreted extracellular vesicles and soluble factors for tissue regeneration. Adv. Funct. Mater. 30:1909125. 10.1002/adfm.201909125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calota D. R., Nitescu C., Florescu I. P., Lascar I. (2012). Surgical management of extensive burns treatment using allografts. J. Med. Life 5 486–490. [PMC free article] [PubMed] [Google Scholar]
- Chen C., Wang D., Moshaverinia A., Liu D., Kou X., Yu W., et al. (2017). Mesenchymal stem cell transplantation in tight-skin mice identifies miR-151-5p as a therapeutic target for systemic sclerosis. Cell Res. 27 559–577. 10.1038/cr.2017.11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen G., Huang A. C., Zhang W., Zhang G., Wu M., Xu W., et al. (2018). Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 560 382–386. 10.1038/s41586-018-0392-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J., Zhou R., Liang Y., Fu X., Wang D., Wang C. (2019). Blockade of lncRNA-ASLNCS5088-enriched exosome generation in M2 macrophages by GW4869 dampens the effect of M2 macrophages on orchestrating fibroblast activation. FASEB J. 33 12200–12212. 10.1096/fj.201901610 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen L., Karisma V. W., Liu H., Zhong L. (2019). MicroRNA-300: a transcellular mediator in exosome regulates melanoma progression. Front. Oncol. 9:1005. 10.3389/fonc.2019.01005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng C. F., Fan J., Fedesco M., Guan S., Li Y., Bandyopadhyay B., et al. (2008). Transforming growth factor alpha (TGFalpha)-stimulated secretion of HSP90alpha: using the receptor LRP-1/CD91 to promote human skin cell migration against a TGFbeta-rich environment during wound healing. Mol. Cell. Biol. 28 3344–3358. 10.1128/mcb.01287-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choudhuri K., Llodrá J., Roth E. W., Tsai J., Gordo S., Wucherpfennig K. W., et al. (2014). Polarized release of T-cell-receptor-enriched microvesicles at the immunological synapse. Nature 507 118–123. 10.1038/nature12951 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ciardiello C., Cavallini L., Spinelli C., Yang J., Reis-Sobreiro M., de Candia P., et al. (2016). Focus on extracellular vesicles: new frontiers of cell-to-cell communication in cancer. Int. J. Mol. Sci. 17:175. 10.3390/ijms17020175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colombo M., Moita C., van Niel G., Kowal J., Vigneron J., Benaroch P., et al. (2013). Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J. Cell Sci. 126 5553–5565. 10.1242/jcs.128868 [DOI] [PubMed] [Google Scholar]
- Cuttle L., Kempf M., Phillips G. E., Mill J., Hayes M. T., Fraser J. F., et al. (2006). A porcine deep dermal partial thickness burn model with hypertrophic scarring. Burns 32 806–820. 10.1016/j.burns.2006.02.023 [DOI] [PubMed] [Google Scholar]
- David G., Zimmermann P. (2020). Heparanase involvement in exosome formation. Adv. Exp. Med. Biol. 1221 285–307. 10.1007/978-3-030-34521-1_10 [DOI] [PubMed] [Google Scholar]
- de Jong O. G., van Balkom B. W., Gremmels H., Verhaar M. C. (2016). Exosomes from hypoxic endothelial cells have increased collagen crosslinking activity through up-regulation of lysyl oxidase-like 2. J. Cell. Mol. Med. 20 342–350. 10.1111/jcmm.12730 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Conde I., Shrimpton C. N., Thiagarajan P., López J. A. (2005). Tissue-factor-bearing microvesicles arise from lipid rafts and fuse with activated platelets to initiate coagulation. Blood 106 1604–1611. 10.1182/blood-2004-03-1095 [DOI] [PubMed] [Google Scholar]
- Du T., Ju G., Wu S., Cheng Z., Cheng J., Zou X., et al. (2014). Microvesicles derived from human Wharton’s jelly mesenchymal stem cells promote human renal cancer cell growth and aggressiveness through induction of hepatocyte growth factor. PLoS One 9:e96836. 10.1371/journal.pone.0096836 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenbeiß W., Siemers F., Amtsberg G., Hinz P., Hartmann B., Kohlmann T., et al. (2012). Prospective, double-blinded, randomised controlled trial assessing the effect of an octenidine-based hydrogel on bacterial colonisation and epithelialization of skin graft wounds in burn patients. Int. J. Burns Trauma 2 71–79. [PMC free article] [PubMed] [Google Scholar]
- Ekström E. J., Bergenfelz C., von Bülow V., Serifler F., Carlemalm E., Jönsson G., et al. (2014). WNT5A induces release of exosomes containing pro-angiogenic and immunosuppressive factors from malignant melanoma cells. Mol. Cancer 13:88. 10.1186/1476-4598-13-88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Rifaie A. A., Sabry D., Doss R. W., Kamal M. A., Abd El Hassib D. M. (2018). Heme oxygenase and iron status in exosomes of psoriasis patients. Arch. Dermatol. Res. 310 651–656. 10.1007/s00403-018-1852-6 [DOI] [PubMed] [Google Scholar]
- El-Tookhy O. S., Shamaa A. A., Shehab G. G., Abdallah A. N., Azzam O. M. (2017). Histological evaluation of experimentally induced critical size defect skin wounds using exosomal solution of mesenchymal stem cells derived microvesicles. Int. J. Stem Cells 10 144–153. 10.15283/ijsc17043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang S., Xu C., Zhang Y., Xue C., Yang C., Bi H., et al. (2016). Umbilical cord-derived mesenchymal stem cell-derived exosomal microRNAs suppress myofibroblast differentiation by inhibiting the transforming growth factor-β/SMAD2 pathway during wound healing. Stem Cells Transl. Med. 5 1425–1439. 10.5966/sctm.2015-0367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang Y., Zhang Y., Zhou J., Cao K. (2019). Adipose-derived mesenchymal stem cell exosomes: a novel pathway for tissues repair. Cell Tissue Bank. 20 153–161. 10.1007/s10561-019-09761-y [DOI] [PubMed] [Google Scholar]
- Felicetti F., De Feo A., Coscia C., Puglisi R., Pedini F., Pasquini L., et al. (2016). Exosome-mediated transfer of miR-222 is sufficient to increase tumor malignancy in melanoma. J. Transl. Med. 14:56. 10.1186/s12967-016-0811-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng Z. (2010). p53 regulation of the IGF-1/AKT/mTOR pathways and the endosomal compartment. Cold Spring Harb. Perspect. Biol. 2:a001057. 10.1101/cshperspect.a001057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frei K., Gramatzki D., Tritschler I., Schroeder J. J., Espinoza L., Rushing E. J., et al. (2015). Transforming growth factor-β pathway activity in glioblastoma. Oncotarget 6 5963–5977. 10.18632/oncotarget.3467 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friand V., David G., Zimmermann P. (2015). Syntenin and syndecan in the biogenesis of exosomes. Biol. Cell 107 331–341. 10.1111/boc.201500010 [DOI] [PubMed] [Google Scholar]
- Geiger A., Walker A., Nissen E. (2015). Human fibrocyte-derived exosomes accelerate wound healing in genetically diabetic mice. Biochem. Biophys. Res. Commun. 467 303–309. 10.1016/j.bbrc.2015.09.166 [DOI] [PubMed] [Google Scholar]
- Goodall R. J., Ellauzi J., Tan M. K. H., Onida S., Davies A. H., Shalhoub J. (2020). A systematic review of the impact of foot care education on self efficacy and self care in patients with diabetes. Eur. J. Vasc. Endovasc. Surg. 60 282–292. 10.1016/j.ejvs.2020.03.053 [DOI] [PubMed] [Google Scholar]
- Gowda R., Robertson B. M., Iyer S., Barry J., Dinavahi S. S., Robertson G. P. (2020). The role of exosomes in metastasis and progression of melanoma. Cancer Treat. Rev. 85:101975. 10.1016/j.ctrv.2020.101975 [DOI] [PubMed] [Google Scholar]
- Gu H., Chen C., Hao X., Wang C., Zhang X., Li Z., et al. (2016). Sorting protein VPS33B regulates exosomal autocrine signaling to mediate hematopoiesis and leukemogenesis. J. Clin. Invest. 126 4537–4553. 10.1172/JCI87105 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Guo L., Zhang Y., Wei R., Zhang X., Wang C., Feng M. (2020). Proinflammatory macrophage-derived microvesicles exhibit tumor tropism dependent on CCL2/CCR2 signaling axis and promote drug delivery via SNARE-mediated membrane fusion. Theranostics 10 6581–6598. 10.7150/thno.45528 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo S. C., Tao S. C., Yin W. J., Qi X., Yuan T., Zhang C. Q. (2017). Exosomes derived from platelet-rich plasma promote the re-epithelization of chronic cutaneous wounds via activation of YAP in a diabetic rat model. Theranostics 7 81–96. 10.7150/thno.16803 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hassanpour M., Rezabakhsh A., Rezaie J., Nouri M., Rahbarghazi R. (2020). Exosomal cargos modulate autophagy in recipient cells via different signaling pathways. Cell Biosci. 10:92. 10.1186/s13578-020-00455-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He C., Zheng S., Luo Y., Wang B. (2018). Exosome theranostics: biology and translational medicine. Theranostics 8 237–255. 10.7150/thno.21945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heinemann M. L., Ilmer M., Silva L. P., Hawke D. H., Recio A., Vorontsova M. A., et al. (2014). Benchtop isolation and characterization of functional exosomes by sequential filtration. J. Chromatogr. A 1371 125–135. 10.1016/j.chroma.2014.10.026 [DOI] [PubMed] [Google Scholar]
- Hood J. L. (2016). Melanoma exosomes enable tumor tolerance in lymph nodes. Med. Hypotheses 90 11–13. 10.1016/j.mehy.2016.02.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hromada C., Mühleder S., Grillari J., Redl H., Holnthoner W. (2017). Endothelial extracellular vesicles-promises and challenges. Front. Physiol. 8:275. 10.3389/fphys.2017.00275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu C., Morohashi Y., Yoshimura S., Manrique-Hoyos N., Jung S., Lauterbach M. A., et al. (2010). Regulation of exosome secretion by Rab35 and its GTPase-activating proteins TBC1D10A-C. J. Cell Biol. 189 223–232. 10.1083/jcb.200911018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu L., Wang J., Zhou X., Xiong Z., Zhao J., Yu R., et al. (2016). Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Sci. Rep. 6:32993. 10.1038/srep32993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu S., Li Z., Cores J., Huang K., Su T., Dinh P. U., et al. (2019). Needle-free injection of exosomes derived from human dermal fibroblast spheroids ameliorates skin photoaging. ACS Nano 13 11273–11282. 10.1021/acsnano.9b04384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu Y., Rao S. S., Wang Z. X., Cao J., Tan Y. J., Luo J., et al. (2018). Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function. Theranostics 8 169–184. 10.7150/thno.21234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang P., Bi J., Owen G. R., Chen W., Rokka A., Koivisto L., et al. (2015). Keratinocyte microvesicles regulate the expression of multiple genes in dermal fibroblasts. J. Invest. Dermatol. 135 3051–3059. 10.1038/jid.2015.320 [DOI] [PubMed] [Google Scholar]
- Huleihel L., Hussey G. S., Naranjo J. D., Zhang L., Dziki J. L., Turner N. J., et al. (2016). Matrix-bound nanovesicles within ECM bioscaffolds. Sci. Adv. 2:e1600502. 10.1126/sciadv.1600502 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyenne V., Apaydin A., Rodriguez D., Spiegelhalter C., Hoff-Yoessle S., Diem M., et al. (2015). RAL-1 controls multivesicular body biogenesis and exosome secretion. J. Cell Biol. 211 27–37. 10.1083/jcb.201504136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isola A. L., Eddy K., Chen S. (2016). Biology, therapy and implications of tumor exosomes in the progression of melanoma. Cancers 8:110. 10.3390/cancers8120110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iversen M. M., Midthjell K., Tell G. S., Moum T., Ostbye T., Nortvedt M. W., et al. (2009). The association between history of diabetic foot ulcer, perceived health and psychological distress: the Nord-Trøndelag health study. BMC Endocr. Disord. 9:18. 10.1186/1472-6823-9-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson C. E., Scruggs B. S., Schaffer J. E., Hanson P. I. (2017). Effects of inhibiting VPS4 support a general role for ESCRTs in extracellular vesicle biogenesis. Biophys. J. 113 1342–1352. 10.1016/j.bpj.2017.05.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jafari D., Malih S., Eslami S. S., Jafari R., Darzi L., Tarighi P., et al. (2019). The relationship between molecular content of mesenchymal stem cells derived exosomes and their potentials: opening the way for exosomes based therapeutics. Biochimie 165 76–89. 10.1016/j.biochi.2019.07.009 [DOI] [PubMed] [Google Scholar]
- Jafari R., Rahbarghazi R., Ahmadi M., Hassanpour M., Rezaie J. (2020). Hypoxic exosomes orchestrate tumorigenesis: molecular mechanisms and therapeutic implications. J. Transl. Med. 18:474. 10.1186/s12967-020-02662-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeong D., Jo W., Yoon J., Kim J., Gianchandani S., Gho Y. S., et al. (2014). Nanovesicles engineered from ES cells for enhanced cell proliferation. Biomaterials 35 9302–9310. 10.1016/j.biomaterials.2014.07.047 [DOI] [PubMed] [Google Scholar]
- Jeong J. H., Fan Y., You G. Y., Choi T. H., Kim S. (2015). Improvement of photoaged skin wrinkles with cultured human fibroblasts and adipose-derived stem cells: a comparative study. J. Plast. Reconstr. Aesthet. Surg. 68 372–381. 10.1016/j.bjps.2014.10.045 [DOI] [PubMed] [Google Scholar]
- Jiang T., Wang Z., Sun J. (2020). Human bone marrow mesenchymal stem cell-derived exosomes stimulate cutaneous wound healing mediates through TGF-β/Smad signaling pathway. Stem Cell Res. Ther. 11:198. 10.1186/s13287-020-01723-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalluri R., Lebleu V. S. (2020). The biology, function, and biomedical applications of exosomes. Science 367:eaau6977. 10.1126/science.aau6977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H., Wang S. Y., Kwak G., Yang Y., Kwon I. C., Kim S. H. (2019). Exosome-guided phenotypic switch of M1 to M2 macrophages for cutaneous wound healing. Adv. Sci. 6:1900513. 10.1002/advs.201900513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim Y. J., Yoo S. M., Park H. H., Lim H. J., Kim Y. L., Lee S., et al. (2017). Exosomes derived from human umbilical cord blood mesenchymal stem cells stimulates rejuvenation of human skin. Biochem. Biophys. Res. Commun. 493 1102–1108. 10.1016/j.bbrc.2017.09.056 [DOI] [PubMed] [Google Scholar]
- Kobayashi H., Ebisawa K., Kambe M., Kasai T., Suga H., Nakamura K., et al. (2018). Effects of exosomes derived from the induced pluripotent stem cells on skin wound healing. Nagoya J. Med. Sci. 80 141–153. 10.18999/nagjms.80.2.141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kotzerke K., Mempel M., Aung T., Wulf G. G., Urlaub H., Wenzel D., et al. (2013). Immunostimulatory activity of murine keratinocyte-derived exosomes. Exp. Dermatol. 22 650–655. 10.1111/exd.12230 [DOI] [PubMed] [Google Scholar]
- Leoni G., Neumann P. A., Kamaly N., Quiros M., Nishio H., Jones H. R., et al. (2015). Annexin A1-containing extracellular vesicles and polymeric nanoparticles promote epithelial wound repair. J. Clin. Invest. 125 1215–1227. 10.1172/jci76693 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lespagnol A., Duflaut D., Beekman C., Blanc L., Fiucci G., Marine J., et al. (2008). Exosome secretion, including the DNA damage-induced p53-dependent secretory pathway, is severely compromised in TSAP6/Steap3-null mice. Cell Death Differ. 15 1723–1733. 10.1038/cdd.2008.104 [DOI] [PubMed] [Google Scholar]
- Lewis C. J. (2013). Stem cell application in acute burn care and reconstruction. J. Wound Care 22 7–8, 10, 12–16. 10.12968/jowc.2013.22.1.7 [DOI] [PubMed] [Google Scholar]
- Li C. J., Fang Q. H., Liu M. L., Lin J. N. (2020). Current understanding of the role of adipose-derived extracellular vesicles in metabolic homeostasis and diseases: communication from the distance between cells/tissues. Theranostics 10 7422–7435. 10.7150/thno.42167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J., Sen G. L. (2016). Post-transcriptional mechanisms regulating epidermal stem and progenitor cell self-renewal and differentiation. J. Invest. Dermatol. 136 746–752. 10.1016/j.jid.2015.12.030 [DOI] [PubMed] [Google Scholar]
- Li L., Zuo X., Liu D., Luo H., Zhu H. (2020). The profiles of miRNAs and lncRNAs in peripheral blood neutrophils exosomes of diffuse cutaneous systemic sclerosis. J. Dermatol. Sci. 98 88–97. 10.1016/j.jdermsci.2020.02.009 [DOI] [PubMed] [Google Scholar]
- Li X., Jiang C., Zhao J. (2016). Human endothelial progenitor cells-derived exosomes accelerate cutaneous wound healing in diabetic rats by promoting endothelial function. J. Diabetes Complications 30 986–992. 10.1016/j.jdiacomp.2016.05.009 [DOI] [PubMed] [Google Scholar]
- Liekens S., De Clercq E., Neyts J. (2001). Angiogenesis: regulators and clinical applications. Biochem. Pharmacol. 61 253–270. 10.1016/s0006-2952(00)00529-3 [DOI] [PubMed] [Google Scholar]
- Liu W., Yu M., Xie D., Wang L., Ye C., Zhu Q., et al. (2020). Melatonin-stimulated MSC-derived exosomes improve diabetic wound healing through regulating macrophage M1 and M2 polarization by targeting the PTEN/AKT pathway. Stem Cell Res. Ther. 11:259. 10.1186/s13287-020-01756-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lootens L., Brusselaers N., Beele H., Monstrey S. (2013). Keratinocytes in the treatment of severe burn injury: an update. Int. Wound J. 10 6–12. 10.1111/j.1742-481X.2012.01083.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lotvall J., Hill A. F., Hochberg F., Buzas E. I., Di Vizio D., Gardiner C., et al. (2014). Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the International society for extracellular vesicles. J. Extracell. Vesicles 3:26913. 10.3402/jev.v3.26913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu M., Peng L., Ming X., Wang X., Cui A., Li Y., et al. (2019). Enhanced wound healing promotion by immune response-free monkey autologous iPSCs and exosomes vs. their allogeneic counterparts. EBioMedicine 42 443–457. 10.1016/j.ebiom.2019.03.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marino J., Babiker-Mohamed M. H., Crosby-Bertorini P., Paster J. T., LeGuern C., Germana S., et al. (2016). Donor exosomes rather than passenger leukocytes initiate alloreactive T cell responses after transplantation. Sci. Immunol. 1:aaf8759. 10.1126/sciimmunol.aaf8759 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McAndrews K. M., Kalluri R. (2019). Mechanisms associated with biogenesis of exosomes in cancer. Mol. Cancer 18:52. 10.1186/s12943-019-0963-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCready J., Sims J. D., Chan D., Jay D. G. (2010). Secretion of extracellular hsp90alpha via exosomes increases cancer cell motility: a role for plasminogen activation. BMC Cancer 10:294. 10.1186/1471-2407-10-294 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medina A., Ghahary A. (2010). Transdifferentiated circulating monocytes release exosomes containing 14-3-3 proteins with matrix metalloproteinase-1 stimulating effect for dermal fibroblasts. Wound Repair Regen. 18 245–253. 10.1111/j.1524-475X.2010.00580.x [DOI] [PubMed] [Google Scholar]
- Mehryab F., Rabbani S., Shahhosseini S., Shekari F., Fatahi Y., Baharvand H., et al. (2020). Exosomes as a next-generation drug delivery system: an update on drug loading approaches, characterization, and clinical application challenges. Acta Biomater. 113 42–62. 10.1016/j.actbio.2020.06.036 [DOI] [PubMed] [Google Scholar]
- Menck K., Sönmezer C., Worst T. S., Schulz M., Dihazi G. H., Streit F., et al. (2017). Neutral sphingomyelinases control extracellular vesicles budding from the plasma membrane. J. Extracell. Vesicles 6:1378056. 10.1080/20013078.2017.1378056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messenger S. W., Woo S. S., Sun Z., Martin T. F. J. (2018). A Ca(2+)-stimulated exosome release pathway in cancer cells is regulated by Munc13-4. J. Cell Biol. 217 2877–2890. 10.1083/jcb.201710132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirzaei H., Gholamin S., Shahidsales S., Sahebkar A., Jaafari M. R., Mirzaei H. R., et al. (2016). MicroRNAs as potential diagnostic and prognostic biomarkers in melanoma. Eur. J. Cancer 53 25–32. 10.1016/j.ejca.2015.10.009 [DOI] [PubMed] [Google Scholar]
- Morelli A. E., Bracamonte-Baran W., Burlingham W. J. (2017). Donor-derived exosomes: the trick behind the semidirect pathway of allorecognition. Curr. Opin. Organ Transpl. 22 46–54. 10.1097/mot.0000000000000372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nabhan J. F., Hu R., Oh R. S., Cohen S. N., Lu Q. (2012). Formation and release of arrestin domain-containing protein 1-mediated microvesicles (ARMMs) at plasma membrane by recruitment of TSG101 protein. Proc. Natl. Acad. Sci. U.S.A. 109 4146–4151. 10.1073/pnas.1200448109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nacer Khodja A., Mahlous M., Tahtat D., Benamer S., Larbi Youcef S., Chader H., et al. (2013). Evaluation of healing activity of PVA/chitosan hydrogels on deep second degree burn: pharmacological and toxicological tests. Burns 39 98–104. 10.1016/j.burns.2012.05.021 [DOI] [PubMed] [Google Scholar]
- Nakamura K., Jinnin M., Fukushima S., Ihn H. (2016a). Exosome expression in the skin and sera of systemic sclerosis patients, and its possible therapeutic application against skin ulcer. J. Dermatol. Sci. 84 e97–e98. [Google Scholar]
- Nakamura K., Jinnin M., Harada M., Kudo H., Nakayama W., Inoue K., et al. (2016b). Altered expression of CD63 and exosomes in scleroderma dermal fibroblasts. J. Dermatol. Sci. 84 30–39. 10.1016/j.jdermsci.2016.06.013 [DOI] [PubMed] [Google Scholar]
- Nakamura Y., Kita S., Tanaka Y., Fukuda S., Obata Y., Okita T., et al. (2020). Adiponectin stimulates exosome release to enhance mesenchymal stem-cell-driven therapy of heart failure in mice. Mol. Ther. 28 2203–2219. 10.1016/j.ymthe.2020.06.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nasiri Kenari A., Cheng L., Hill A. F. (2020). Methods for loading therapeutics into extracellular vesicles and generating extracellular vesicles mimetic-nanovesicles. Methods 177 103–113. 10.1016/j.ymeth.2020.01.001 [DOI] [PubMed] [Google Scholar]
- Nguyen D. B., Ly T. B., Wesseling M. C., Hittinger M., Torge A., Devitt A., et al. (2016). Characterization of microvesicles released from human red blood cells. Cell. Physiol. Biochem. 38 1085–1099. 10.1159/000443059 [DOI] [PubMed] [Google Scholar]
- Nikfarjam S., Rezaie J., Zolbanin N. M., Jafari R. (2020). Mesenchymal stem cell derived-exosomes: a modern approach in translational medicine. J. Transl. Med. 18:449. 10.1186/s12967-020-02622-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuschke A. (2014). Activity of mesenchymal stem cells in therapies for chronic skin wound healing. Organogenesis 10 29–37. 10.4161/org.27405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh M., Lee J., Kim Y. J., Rhee W. J., Park J. H. (2018). Exosomes derived from human induced pluripotent stem cells ameliorate the aging of skin fibroblasts. Int. J. Mol. Sci. 19:1715. 10.3390/ijms19061715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ostrowski M., Carmo N. B., Krumeich S., Fanget I., Raposo G., Savina A., et al. (2010). Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat. Cell Biol. 12 19–30. 10.1038/ncb2000 [DOI] [PubMed] [Google Scholar]
- Parolini I., Federici C., Raggi C., Lugini L., Palleschi S., De Milito A., et al. (2009). Microenvironmental pH is a key factor for exosome traffic in tumor cells. J. Biol. Chem. 284 34211–34222. 10.1074/jbc.M109.041152 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeffer S. R., Grossmann K. F., Cassidy P. B., Yang C. H., Fan M., Kopelovich L., et al. (2015). Detection of exosomal miRNAs in the plasma of melanoma patients. J. Clin. Med. 4 2012–2027. 10.3390/jcm4121957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan J., Kumar P., Hao D., Gao K., Farmer D., Wang A. (2018). Engineering mesenchymal stem cells to improve their exosome efficacy and yield for cell-free therapy. J. Extracell. Vesicles 7:1522236. 10.1080/20013078.2018.1522236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phinney D. G., Pittenger M. F. (2017). Concise review: MSC-derived exosomes for cell-free therapy. Stem Cells 35 851–858. 10.1002/stem.2575 [DOI] [PubMed] [Google Scholar]
- Purushothaman A., Sanderson R. D. (2020). Heparanase: a dynamic promoter of myeloma progression. Adv. Exp. Med. Biol. 1221 331–349. 10.1007/978-3-030-34521-1_12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahbarghazi R., Jabbari N., Sani N. A., Asghari R., Salimi L., Kalashani S. A., et al. (2019). Tumor-derived extracellular vesicles: reliable tools for cancer diagnosis and clinical applications. Cell Commun. Signal. 17:73. 10.1186/s12964-019-0390-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahmani A., Saleki K., Javanmehr N., Khodaparast J., Saadat P., Nouri H. R. (2020). Mesenchymal stem cell-derived extracellular vesicle-based therapies protect against coupled degeneration of the central nervous and vascular systems in stroke. Ageing Res. Rev. 62:101106. 10.1016/j.arr.2020.101106 [DOI] [PubMed] [Google Scholar]
- Ramakrishnan D. P., Hajj-Ali R. A., Chen Y., Silverstein R. L. (2016). Extracellular vesicles activate a CD36-dependent signaling pathway to inhibit microvascular endothelial cell migration and tube formation. Arterioscler. Thromb. Vasc. Biol. 36 534–544. 10.1161/atvbaha.115.307085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Record M., Subra C., Silvente-Poirot S., Poirot M. (2011). Exosomes as intercellular signalosomes and pharmacological effectors. Biochem. Pharmacol. 81 1171–1182. 10.1016/j.bcp.2011.02.011 [DOI] [PubMed] [Google Scholar]
- Rezaie J., Nejati V., Khaksar M., Oryan A., Aghamohamadzadeh N., Shariatzadeh M. A., et al. (2018b). Diabetic sera disrupted the normal exosome signaling pathway in human mesenchymal stem cells in vitro. Cell Tissue Res. 374 555–565. 10.1007/s00441-018-2895-x [DOI] [PubMed] [Google Scholar]
- Rezaie J., Mehranjani M. S., Rahbarghazi R., Shariatzadeh M. A. (2018a). Angiogenic and restorative abilities of human mesenchymal stem cells were reduced following treatment with serum from diabetes mellitus type 2 patients. J. Cell. Biochem. 119 524–535. 10.1002/jcb.26211 [DOI] [PubMed] [Google Scholar]
- Ribeiro M. P., Espiga A., Silva D., Baptista P., Henriques J., Ferreira C., et al. (2009). Development of a new chitosan hydrogel for wound dressing. Wound Repair Regen. 17 817–824. 10.1111/j.1524-475X.2009.00538.x [DOI] [PubMed] [Google Scholar]
- Salimi L., Akbari A., Jabbari N., Mojarad B., Vahhabi A., Szafert S., et al. (2020). Synergies in exosomes and autophagy pathways for cellular homeostasis and metastasis of tumor cells. Cell Biosci. 10:64. 10.1186/s13578-020-00426-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savina A., Furlán M., Vidal M., Colombo M. I. (2003). Exosome release is regulated by a calcium-dependent mechanism in K562 cells. J. Biol. Chem. 278 20083–20090. 10.1074/jbc.M301642200 [DOI] [PubMed] [Google Scholar]
- Savina A., Vidal M., Colombo M. I. (2002). The exosome pathway in K562 cells is regulated by Rab11. J. Cell Sci. 115 2505–2515. [DOI] [PubMed] [Google Scholar]
- Schorey J. S., Harding C. V. (2016). Extracellular vesicles and infectious diseases: new complexity to an old story. J. Clin. Invest. 126 1181–1189. 10.1172/jci81132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shabbir A., Cox A., Rodriguez-Menocal L., Salgado M., Van Badiavas E. (2015). Mesenchymal stem cell exosomes induce proliferation and migration of normal and chronic wound fibroblasts, and enhance angiogenesis in vitro. Stem Cells Dev. 24 1635–1647. 10.1089/scd.2014.0316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi H., Xu X., Zhang B., Xu J., Pan Z., Gong A., et al. (2017). 3,3’-diindolylmethane stimulates exosomal Wnt11 autocrine signaling in human umbilical cord mesenchymal stem cells to enhance wound healing. Theranostics 7 1674–1688. 10.7150/thno.18082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi Q., Qian Z., Liu D., Sun J., Wang X., Liu H., et al. (2017). GMSC-derived exosomes combined with a chitosan/silk hydrogel sponge accelerates wound healing in a diabetic rat skin defect model. Front. Physiol. 8:904. 10.3389/fphys.2017.00904 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiekh P. A., Singh A., Kumar A. (2020). Exosome laden oxygen releasing antioxidant and antibacterial cryogel wound dressing OxOBand alleviate diabetic and infectious wound healing. Biomaterials 249:120020. 10.1016/j.biomaterials.2020.120020 [DOI] [PubMed] [Google Scholar]
- Showalter M. R., Wancewicz B., Fiehn O., Archard J. A., Clayton S., Wagner J., et al. (2019). Primed mesenchymal stem cells package exosomes with metabolites associated with immunomodulation. Biochem. Biophys. Res. Commun. 512 729–735. 10.1016/j.bbrc.2019.03.119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singer A. J., Clark R. A. (1999). Cutaneous wound healing. N. Engl. J. Med. 341 738–746. 10.1056/nejm199909023411006 [DOI] [PubMed] [Google Scholar]
- Soraya H., Sani N. A., Jabbari N., Rezaie J. (2021). Metformin increases exosome biogenesis and secretion in U87 MG human glioblastoma cells: a possible mechanism of therapeutic resistance. Arch. Med. Res. 52 151–162. 10.1016/j.arcmed.2020.10.007 [DOI] [PubMed] [Google Scholar]
- Stoffel W., Hammels I., Jenke B., Binczek E., Schmidt-Soltau I., Brodesser S., et al. (2016). Neutral sphingomyelinase (SMPD3) deficiency disrupts the Golgi secretory pathway and causes growth inhibition. Cell Death Dis. 7:e2488. 10.1038/cddis.2016.385 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun Y., Shi H., Yin S., Ji C., Zhang X., Zhang B., et al. (2018). Human mesenchymal stem cell derived exosomes alleviate type 2 diabetes mellitus by reversing peripheral insulin resistance and relieving β-cell destruction. ACS Nano 12 7613–7628. 10.1021/acsnano.7b07643 [DOI] [PubMed] [Google Scholar]
- Sung B., von Lersner A., Guerrero J., Krystofiak E., Inman D., Pelletier R., et al. (2020). A live cell reporter of exosome secretion and uptake reveals pathfinding behavior of migrating cells. Nat. Commun. 11:2092. 10.1038/s41467-020-15747-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surman M., Kȩdracka-Krok S., Hoja-Łukowicz D., Jankowska U., Drożdż A., Stȩpień E., et al. (2020). Mass spectrometry-based proteomic characterization of cutaneous melanoma ectosomes reveals the presence of cancer-related molecules. Int. J. Mol. Sci. 21:2934. 10.3390/ijms21082934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tao S. C., Guo S. C., Li M., Ke Q. F., Guo Y. P., Zhang C. Q. (2017). Chitosan wound dressings incorporating exosomes derived from microRNA-126-overexpressing synovium mesenchymal stem cells provide sustained release of exosomes and heal full-thickness skin defects in a diabetic rat model. Stem Cells Transl. Med. 6 736–747. 10.5966/sctm.2016-0275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Théry C., Witwer K. W., Aikawa E., Alcaraz M. J., Anderson J. D., Andriantsitohaina R., et al. (2018). Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International society for extracellular vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 7:1535750. 10.1080/20013078.2018.1535750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Théry C., Zitvogel L., Amigorena S. (2002). Exosomes: composition, biogenesis and function. Nat. Rev. Immunol. 2 569–579. 10.1038/nri855 [DOI] [PubMed] [Google Scholar]
- Tkach M., Théry C. (2016). Communication by extracellular vesicles: where we are and where we need to go. Cell 164 1226–1232. 10.1016/j.cell.2016.01.043 [DOI] [PubMed] [Google Scholar]
- Trajkovic K., Hsu C., Chiantia S., Rajendran L., Wenzel D., Wieland F., et al. (2008). Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 319 1244–1247. 10.1126/science.1153124 [DOI] [PubMed] [Google Scholar]
- Van De Water L., Varney S., Tomasek J. J. (2013). Mechanoregulation of the myofibroblast in wound contraction, scarring, and fibrosis: opportunities for new therapeutic intervention. Adv. Wound Care 2 122–141. 10.1089/wound.2012.0393 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Pol E., Boing A. N., Harrison P., Sturk A., Nieuwland R. (2012). Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol. Rev. 64 676–705. 10.1124/pr.112.005983 [DOI] [PubMed] [Google Scholar]
- van Niel G., D’Angelo G., Raposo G. (2018). Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 19 213–228. 10.1038/nrm.2017.125 [DOI] [PubMed] [Google Scholar]
- Villarroya-Beltri C., Baixauli F., Gutiérrez-Vázquez C., Sánchez-Madrid F., Mittelbrunn M. (2014). Sorting it out: regulation of exosome loading. Semin. Cancer Biol. 28 3–13. 10.1016/j.semcancer.2014.04.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villarroya-Beltri C., Baixauli F., Mittelbrunn M., Fernández-Delgado I., Torralba D., Moreno-Gonzalo O., et al. (2016). ISGylation controls exosome secretion by promoting lysosomal degradation of MVB proteins. Nat. Commun. 7:13588. 10.1038/ncomms13588 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vuckovic S., Vandyke K., Rickards D. A., McCauley Winter P., Brown S. H. J., Mitchell T. W., et al. (2017). The cationic small molecule GW4869 is cytotoxic to high phosphatidylserine-expressing myeloma cells. Br. J. Haematol. 177 423–440. 10.1111/bjh.14561 [DOI] [PubMed] [Google Scholar]
- Wang C., Wang M., Xu T., Zhang X., Lin C., Gao W., et al. (2019). Engineering bioactive self-healing antibacterial exosomes hydrogel for promoting chronic diabetic wound healing and complete skin regeneration. Theranostics 9 65–76. 10.7150/thno.29766 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L., Hu L., Zhou X., Xiong Z., Zhang C., Shehada H. M. A., et al. (2017). Exosomes secreted by human adipose mesenchymal stem cells promote scarless cutaneous repair by regulating extracellular matrix remodelling. Sci. Rep. 7:13321. 10.1038/s41598-017-12919-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wermuth P. J., Piera-Velazquez S., Jimenez S. A. (2017). Exosomes isolated from serum of systemic sclerosis patients display alterations in their content of profibrotic and antifibrotic microRNA and induce a profibrotic phenotype in cultured normal dermal fibroblasts. Clin. Exp. Rheumatol. 35 Suppl. 106 21–30. [PMC free article] [PubMed] [Google Scholar]
- Wong R., Geyer S., Weninger W., Guimberteau J. C., Wong J. K. (2016). The dynamic anatomy and patterning of skin. Exp. Dermatol. 25 92–98. 10.1111/exd.12832 [DOI] [PubMed] [Google Scholar]
- Wortzel I., Dror S., Kenific C. M., Lyden D. (2019). Exosome-mediated metastasis: communication from a distance. Dev. Cell 49 347–360. 10.1016/j.devcel.2019.04.011 [DOI] [PubMed] [Google Scholar]
- Wynn T. A., Ramalingam T. R. (2012). Mechanisms of fibrosis: therapeutic translation for fibrotic disease. Nat. Med. 18 1028–1040. 10.1038/nm.2807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang B., Chen Y., Shi J. (2019). Exosome biochemistry and advanced nanotechnology for next-generation theranostic platforms. Adv. Mater. 31:e1802896. 10.1002/adma.201802896 [DOI] [PubMed] [Google Scholar]
- Yang R., Yang S., Zhao J., Hu X., Chen X., Wang J., et al. (2020). Progress in studies of epidermal stem cells and their application in skin tissue engineering. Stem Cell Res. Ther. 11:303. 10.1186/s13287-020-01796-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeh Y., Ozer H., Lehman A., Maddocks K., Yu L., Johnson A., et al. (2015). Characterization of CLL exosomes reveals a distinct microRNA signature and enhanced secretion by activation of BCR signaling. Blood 125 3297–3305. 10.1182/blood-2014-12-618470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang B., Shi Y., Gong A., Pan Z., Shi H., Yang H., et al. (2016). HucMSC exosome-delivered 14-3-3ζ orchestrates self-control of the Wnt response via modulation of YAP during cutaneous regeneration. Stem Cells 34 2485–2500. 10.1002/stem.2432 [DOI] [PubMed] [Google Scholar]
- Zhang B., Wang M., Gong A., Zhang X., Wu X., Zhu Y., et al. (2015a). HucMSC-exosome mediated-Wnt4 signaling is required for cutaneous wound healing. Stem Cells 33 2158–2168. 10.1002/stem.1771 [DOI] [PubMed] [Google Scholar]
- Zhang B., Wu X., Zhang X., Sun Y., Yan Y., Shi H., et al. (2015b). Human umbilical cord mesenchymal stem cell exosomes enhance angiogenesis through the Wnt4/β-catenin pathway. Stem Cells Transl. Med. 4 513–522. 10.5966/sctm.2014-0267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J., Guan J., Niu X., Hu G., Guo S., Li Q., et al. (2015c). Exosomes released from human induced pluripotent stem cells-derived MSCs facilitate cutaneous wound healing by promoting collagen synthesis and angiogenesis. J. Transl. Med. 13:49. 10.1186/s12967-015-0417-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang B., Yeo R. W. Y., Lai R. C., Sim E. W. K., Chin K. C., Lim S. K. (2018). Mesenchymal stromal cell exosome-enhanced regulatory T-cell production through an antigen-presenting cell-mediated pathway. Cytotherapy 20 687–696. 10.1016/j.jcyt.2018.02.372 [DOI] [PubMed] [Google Scholar]
- Zhang K., Yu L., Li F. R., Li X., Wang Z., Zou X., et al. (2020). Topical application of exosomes derived from human umbilical cord mesenchymal stem cells in combination with sponge spicules for treatment of photoaging. Int. J. Nanomed. 15 2859–2872. 10.2147/ijn.S249751 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W., Bai X., Zhao B., Li Y., Zhang Y., Li Z., et al. (2018). Cell-free therapy based on adipose tissue stem cell-derived exosomes promotes wound healing via the PI3K/Akt signaling pathway. Exp. Cell Res. 370 333–342. [DOI] [PubMed] [Google Scholar]
- Zhang Y., Bi J., Huang J., Tang Y., Du S., Li P. (2020). Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int. J. Nanomed. 15 6917–6934. 10.2147/IJN.S264498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao B., Zhang Y., Han S., Zhang W., Zhou Q., Guan H., et al. (2017). Exosomes derived from human amniotic epithelial cells accelerate wound healing and inhibit scar formation. J. Mol. Histol. 48 121–132. 10.1007/s10735-017-9711-x [DOI] [PubMed] [Google Scholar]
- Zorin V., Zorina A., Cherkasov V., Deev R., Kopnin P., Isaev A. (2017). Clinical-instrumental and morphological evaluation of the effect of autologous dermal fibroblasts administration. J. Tissue Eng. Regen. Med. 11 778–786. 10.1002/term.1976 [DOI] [PubMed] [Google Scholar]