

Abstract
When people are stuck on a problem, they sometimes benefit from an incubation period –a break from working on the problem. Anecdotes and empirical evidence suggest that sleeping during incubation is useful, but the mechanisms remain poorly understood. We examined how targeted memory reactivation during sleep, which boosts next-day solving, relates to forgetting fixation, a well-supported explanation of awake incubation. In evening sessions, participants attempted puzzles, while a unique sound cue played during each puzzle. Half the time, puzzles included fixating information reinforcing an incorrect representation. Later, during deep sleep, sounds associated with half of participants’ previously unsolved puzzles were presented. The sounds should strengthen puzzle memories and reduce forgetting of the fixating information. In morning solving, overnight cueing reliably interacted with fixating information: participants solved numerically more cued than uncued puzzles, but only when puzzles included fixating information. These results suggest that additional processing occurred beyond simple fixation forgetting.
Keywords: problem solving, incubation, sleep, targeted memory reactivation, restructuring
Sleep has been associated with problem solving in anecdotes from famous scientists and artists (Barrett, 2001; Mazzarello, 2000) as well as several research experiments (Beijamini et al., 2014; Sio et al., 2013; Wagner et al., 2004). Sleep can be thought of as a special kind of incubation period – a time away from the problem during which the problem-solver is not intentionally, consciously thinking about or actively working to solve the problem. Previous research demonstrates that under certain waking conditions, people solve problems better after a period of incubation, compared to a period of continuous work, and several theories of how awake incubation periods improve problem solving have been proposed (for review, Dodds et al., 2012; Gilhooly, 2016; Sio & Ormerod, 2009). However, while there is some evidence that sleep facilitates creative problem solving, the mechanisms underlying this effect are not yet clear.
Incubation has long been proposed to be a critical component of the problem solving process, particularly after a problem solver reaches an impasse (Wallas, 1926). Many researchers agree that overcoming this impasse benefits from restructuring – that is, recognizing or using different relations between problem elements to reorganize their conception of the problem or approach it in a new way (for review, Schooler & Melcher, 1995; Sio & Ormerod, 2009). After restructuring, participants often experience a feeling of ‘insight’ whereby the solution is revealed suddenly and holistically (Kounios & Beeman, 2014).
Research on memory processing during sleep provides evidence that could be consistent with the idea that unconscious processing during sleep permits restructuring in ways that benefit post-sleep solving. During sleep, external input is considerably reduced and information processing is generally considered unconscious; yet, meaningful cognitive processing occurs during sleep. Sleep strengthens memory such that information is better retained after a period of sleep than an equivalent period awake (for review see Diekelmann & Born, 2010; Rasch & Born, 2013a; Schönauer & Gais, 2017). In addition, sleep also facilitates memory reorganization (Lewis et al., 2018; Payne, 2011) such that people are better able to recognize and extract underlying patterns and regularities (Durrant et al., 2015; Ellenbogen et al., 2007; Lewis & Durrant, 2011; Wagner et al., 2004), become consciously aware of previously unconscious information (Cousins et al., 2014; Fischer et al., 2006), and integrate new information with previously learned material (Dumay & Gaskell, 2007; Sio et al., 2013). All of these examples of memory reorganization could be considered forms of restructuring resulting in new information patterns.
Memory processing during sleep can be researched by modulating the strength or target of this processing using Targeted Memory Reactivation (TMR, Oudiette & Paller, 2013). As people learn new information, to-be-learned stimuli are paired with sensory cues such as sounds (Rudoy et al., 2009) or smells (Rasch et al., 2007). The associated cues are then presented during sleep without waking the sleeper. The cues are thought to bias naturally-occurring memory reactivation processes, cueing people to specifically reactivate the sound or smell’s associated, learned information. TMR is associated with memory strengthening (Hu et al., 2020; Oudiette & Paller, 2013) and memory reorganization (e.g. Cousins et al., 2014) of the targeted stimuli.
In a recent study, we used TMR to selectively target puzzles for processing during sleep, enhancing solving the following morning (Sanders et al., 2019). During evening sessions, participants attempted to solve puzzles one at a time, each puzzle paired with a distinct, unrelated sound cue. Then, for half of the puzzles that people had failed to solve in previous evenings, we presented the associated sound cues during sleep. Upon reattempting their previously unsolved puzzles the following morning, participants solved more of the cued puzzles than the uncued puzzles, and solving of the cued puzzles was tentatively associated with better memory for the cued puzzle details. To understand what processes underlie this TMR solving benefit, and how successful problem incubation may occur during sleep, we consider general incubation theories proffered to explain awake incubation effects and how they may apply to sleep.
Incubation Theories
Three alternative, but not mutually exclusive, theories suggest different ways that awake incubation periods help people subsequently solve problems. 1) Unconscious processing: While not actively engaged with the problem, some background process(es) allows access to new critical information or relation between problem elements that improves a person’s chance of solving the problem (Gilhooly, 2019; Ritter & Dijksterhuis, 2014). 2) Opportunistic assimilation: While not actively working on the problem, people serendipitously encounter hints in the environment that lead to useful approaches or solutions (Seifert et al., 1995). 3) Forgetting fixation: On first solving attempts, people are initially fixated on incorrect strategies or associations that interfere with solving; but, after time away from the problem, they forget these incorrect ideas, allowing them to more quickly identify better solving strategies, associations, or ideas when they later return to the problem (e.g., Smith, 1995a, 1995b).
Cueing-facilitated problem-solving during sleep is consistent with unconscious processing theory. As mentioned above, processing during sleep is generally considered outside of people’s awareness yet memory transformation occurs (e.g. Ellenbogen et al., 2007). In addition, the only difference between cued and uncued puzzles in our prior study was the presentation of related sounds during sleep that purportedly reactivate the memory, suggesting additional processing took place. Spreading activation from the problem information to related ideas and concepts in memory is often thought to constitute unconscious processing during awake incubation (Gilhooly, 2019; Hélie & Sun, 2010; Yaniv & Meyer, 1987). Indeed, people attempting to solve problems demonstrate solution-related priming (Bowden & Beeman, 1998) and solution sensitivity (Bowers et al., 1990) without solving. People also demonstrate improved solving after an incubation that occurs immediately after seeing the problem but before actively working on it and becoming fixated, suggesting that continued processing occurs during the break (Gilhooly et al., 2012). Moreover, after initially seeing a problem, task difficulty and content during the following awake incubation period – putatively related to the amount of cognitive resources available to engage in background processing – affects solving (Baird et al., 2012; Gilhooly et al., 2013) and the accessibility of information that could lead to solving (Ritter & Dijksterhuis, 2014) suggesting that processing continues outside of participants’ awareness and immediate cognitive focus.
Opportunistic assimilation, while well-supported as a theory of awake incubation, both anecdotally (e.g. Swearingen, 2016) and experimentally (e.g., Seifert, et al., 1995), is unlikely to explain sleep’s facilitation of problem solving as usually no new information is externally presented to participants during sleep. One study presented solution hints to participants prior to the problems, rather than during the incubation period, and found that REM (Rapid Eye Movement) sleep helped people utilize the hints (Cai et al., 2009). However, as opposed to encountering and recognizing the hints during the incubation period, this study suggests that sleep facilitated the integration of information previously encountered while awake, so sleep more likely involved unconscious processing of that previously encountered hint. In addition, dreams could be considered a type of internal environment from which hints or the solution itself could be drawn. Indeed, anecdotal evidence of sleep-facilitated problem solving often highlights a dream that illuminated the solution (Barrett, 2001); however, because dreams are part of an internal mental environment, it is hard to distinguish this from the unconscious processing theory. Most importantly, the sound cues presented during TMR do not provide any additional information related to the puzzles, so there is no greater chance for opportunistic assimilation of hints when puzzles are cued overnight. Therefore, opportunistic assimilation cannot explain the previously found cueing effect, so we will not consider it as an explanation for TMR-facilitated incubation during sleep.
Finally, the forgetting fixation theory is supported by numerous laboratory studies on awake incubation (e.g., Smith, 1995a, 1995b); however, how it might apply to sleep incubation is less clear. In most studies investigating fixation forgetting, participants initially see problems accompanied by red herrings (false solutions). Then, after a short break, they return to reattempt their unsolved puzzles, and solve more puzzles after the incubation period than an equal period of time continuously working on the puzzles (Kohn & Smith, 2009; Smith & Blankenship, 1989, 1991). These studies suggest that changes in environmental (Beda & Smith, 2018) and temporal (Smith & Beda, 2019) contexts improve problem solving through the forgetting or weakening of incorrect solution paths.
However, current evidence for processing during sleep, and especially for improved solving following TMR, seems less consistent with a simple forgetting fixation process for asleep incubation periods. Based on decades of research, sleep is primarily associated with memory strengthening compared to similar periods awake rather than weakening or forgetting (Rasch & Born, 2013), although differential memory processing during sleep is possible under certain circumstances. For example, sleep enhances memory for emotional objects but not the neutral backgrounds they were studied with (Payne et al., 2008). Also, directing participants to remember some items and forget others leads to worse memory for the ‘to forget’ items (Saletin et al., 2011) and reactivating the act of forgetting with learned stimuli during sleep, reduces the targeted item memory compared to unreactived items (Simon et al., 2018). Finally, in our TMR study, cueing the puzzles by playing associated sounds during sleep should strengthen puzzle memory, including whatever information was inducing fixation. Thus, cued puzzles should have decreased fixation forgetting – yet people solved them better than uncued puzzles. Taken as a whole, sleep incubation studies suggest, at a minimum, an additional prioritization process to preferentially maintain some memories over others, rather than a simple, passive, forgetting mechanism.
The Current Research
The current research investigates the roles of unconscious processing and forgetting fixation in sleep-cueing-facilitated problem solving. Therefore, similar to previous studies of forgetting fixation, we adapted puzzles from Sanders et al., (2019) into two versions of each puzzle, a More-Fixating and a Less-Fixating version, by selectively adding (or removing) misleading information to each puzzle. The two versions of each puzzle are identical besides the small alteration, and require the same solution.
For all puzzles, cues should trigger reactivation of puzzle information. For the More-Fixating puzzles, cues will also reactivate fixating information. According to the standard passive forgetting fixation hypothesis, this should impede forgetting of the fixating information, and reduce any benefits of cueing. In this case, we would expect the effect of cueing on problem solving to be smaller for the More-Fixating versions of the puzzles than for the Less-Fixating versions of the puzzles. However, if unconscious processing occurs, co-activating both the puzzle information and the fixating information at the same time could facilitate a memory reorganization process. In this case, we would expect the effect of cueing on problem solving to be larger for the More-Fixating versions of the puzzles. Ideally, these results would occur with concomitant effects on memory for the puzzle details and their associated fixating information (in the More-Fixating versions), but the materials were optimized to measure solving, and we can only roughly measure memory for the puzzles and fixating information.
Method
Participants
Sixty-five participants (51 Female) from the Northwestern University community enrolled in the experiment (age range: 18–26, M = 19.92, SD = 1.66). Participants were fluent speakers of English who reported no history of neurological or sleep disorders. As the data were collected, 6 participants failed to complete all four sessions (two evening and two morning) of the experiment; therefore, we continued to collect data, without looking at the results, until we acquired approximately 60 complete datasets (59 due to a scheduling error). We excluded one additional participant who failed to comply with study procedures. After exclusions, 58 participants (46 Female, age range: 18–26, M = 19.82, SD = 1.62) remained in the analysis. Participants provided informed consent and were monetarily compensated. Procedures were approved by the Northwestern University Institutional Review Board.
Materials
To test the forgetting fixation and unconscious processing hypotheses, we adapted 40 puzzles (32 from Sanders, Osburn, Paller, & Beeman (2019), and 8 new). The puzzles were initially gathered from a variety of sources including previous research on insight problem solving. They were selected to be difficult and require thinking about the problem in a non-dominant way to reach the solution. Puzzles came from four different general categories (matchstick, rebus, spatial, and verbal) in order to enhance each puzzle’s distinctiveness. We then created two versions of each of the 40 puzzles, a More-Fixating version and a Less-Fixating version (see Appendix A for several examples). The two versions of each puzzle were identical and required the same overall solution, except that the More-Fixating version of each puzzle was designed to reinforce a dominant but incorrect representation or approach to the puzzle, making it more salient to the problem solver. For most of the puzzles (n = 28 puzzles), the More-Fixating version contained a distinctly separate piece of information such as a hint (explaining an attempt that does not work, e.g. Appendix A rebus puzzle) or example (reinforcing a rule mentioned in the puzzle, e.g. Appendix A spatial puzzle) that provided relevant information but also made an incorrect approach to the puzzle more salient. For a smaller subset of the puzzles (n = 12 puzzles), the fixating information was integrated into the puzzle, for example as an added rule or detail in the written description, or a different way of presenting the initial setup of a puzzle (e.g. Appendix A matchstick puzzle). Finally, of the 40 original versions of the puzzles taken from the previous study, 31 were used as the Less-Fixating version (and information was added to make the More-Fixating version), 7 were used as the More-Fixating version (and information was removed to make the Less-Fixating version) and 2 were adjusted by both removing fixating information in the Less-Fixating version and adding fixating information in the More-Fixating version. Participants experienced both More-Fixating and Less-Fixating puzzles each night but never both versions of the same puzzle; across participants, the two versions were seen about equally often.
Forty sound clips were selected from Sanders et al. (2019). The sounds were segments of instrumental music and sound effects of differing genres and instrumentation to maintain each sound’s distinctiveness. Sound cues were then generated that were 15 seconds in length and consist of 8–12 seconds of sound followed by 3–7 seconds of silence to reduce habituation to the sound. Sound cues and puzzles were randomly paired for each participant and were therefore not directly meaningful to the solving attempt.
Procedure
Participants completed 2 sets of evening-overnight-morning session pairs (see Figure 1), with 6 unsolved puzzles attained in each evening session: 3 More-Fixating and 3 Less-Fixating puzzles; 3 of which were Cued and 3 of which were Uncued. On a given night, at least one puzzle fell into each of the 4 experimental conditions. Across the two nights, this resulted in a total of 12 puzzles (3 each of Matchstick, Rebus, Spatial, and Verbal, 3 More-Fixating Cued, 3 Less-Fixating Cued, 3 More-Fixating Uncued, and 3 Less-Fixating Uncued) participants failed to solve before incubating during overnight sleep.
Figure 1.

Experimental procedure that was repeated twice. Each evening, participants attempted puzzles while a distinct, 15-s sound clip looped for each one. Half of the puzzles contained additional, misleading information while the other half did not. New puzzles were presented until 6 puzzles remained unsolved. Then participants were tested on recalling which puzzle went with each sound, with feedback. Participants took home a portable EEG sleep monitoring and cueing system. When it detected SWS, it presented sound cues. The following morning, participants returned to the laboratory to complete sound-puzzle and puzzle-details recall tests, and then attempted to solve the puzzles that were unsolved the previous night.
Evening.
Participants generally completed evening sessions between 5 and 8 PM. In the first evening session, participants were instructed how to use the Sleep Monitoring and Cueing System (SMCS, see below) that they later took home to monitor their sleep. Participants were then introduced to the procedure and the puzzle types. Then, participants attempted puzzles one at a time for 2 minutes each. Before each puzzle, participants were told the type of puzzle (Rebus, Spatial, Matchstick, or Verbal). The puzzle’s sound cue then played once with a blank screen to familiarize the participants with the sound. Participants then read the puzzle and indicated if they were familiar with the puzzle, in which case it was skipped and the next puzzle was presented. If the puzzle was unfamiliar, participants attempted to solve the puzzle for 2 minutes while the sound cue played and continuously looped.
During the solving period, if participants had a solution idea, they pressed the spacebar and said their solution aloud. If the offered solution was incorrect, the experimenter explained why the solution would not work and the participant had the remaining time to continue working on the puzzle. If participants correctly solved the puzzle, it was excluded from the remainder of the experiment and replaced with a new puzzle. If participants did not solve the puzzle, they stopped working on the puzzle, a blank screen appeared, and the sound cue played one more time through (15 seconds). During the final 15 seconds of the sound cue, participants were instructed to try to memorize the sound-puzzle pairing.
Participants continued to attempt puzzles until six puzzles remained unsolved. To maximize the distinctiveness of the puzzles, we presented puzzles such that, across all nights of the study, an approximately equal number of puzzles from each category remained unsolved and incubation-ready at the end of each evening session. For instance, if participants solved a Spatial puzzle, an additional Spatial puzzle was presented later in the session.
At the end of each evening session, we tested participants’ memory for the sound-puzzle pairings to further reinforce the association of each unique puzzle with its unique sound. Participants heard a sound clip, reported the puzzle that was paired with the sound, and then, regardless of accuracy, they saw the correct puzzle presented again with the sound for a few seconds and were told to remind themselves of the sound-puzzle pairing. Participants completed 3 rounds of the sound-puzzle recall test. By the third round, participants correctly recalled 81.8% (SD = 17.0%) of puzzles in response to the sound cues. Participants’ accuracy was comparable for puzzles that would later be Cued (M = 80.2%, SD = 23.2%) and those that would later not be cued (M = 83.2%, SD = 16.5%, t(56) = −1.12, p = .268, dz = 0.15). Participants’ accuracy was also comparable for puzzles in the Less-Fixating (M = 80.8%, SD = 19.9%) and the More-Fixating (M = 82.8%, SD = 19.0%, t(56) = −0.79, p = .432, dz = 0.10) conditions. An algorithm determined which puzzles were cued overnight to keep the experimenter blind to the puzzle condition. It attempted to select sounds for cueing such that, across participants, each puzzle was equally often Cued versus Uncued. Due to evening solving difficulty varying by puzzle and fixating condition and prioritizing equal numbers of Cued/Uncued and More-Fixating/Less-Fixating puzzles each night, it was challenging to balance individual puzzle presentation across all conditions. Individual puzzles ranged from 27.3% to 66.7% of the time in the Cued condition, and 20.0% to 81.8% of the time in the More-Fixating condition.
Before leaving the lab, participants were asked to avoid thinking about or attempting to solve the unsolved puzzles between the evening and morning sessions. Puzzles that participants reported solving after the evening session but before sleep were removed from the analyses.
Overnight.
Upon leaving the lab, participants took home a Sleep Monitoring and Cueing System (SMCS, the Asimov by Sheepdog Sciences, Inc.). The SMCS includes a modified transmitter (from Zeo Inc.) and a laptop computer with adapted software and hardware to receive wirelessly transmitted signals from the transmitter. This device reliably classifies sleep into component stages similar to standard polysomnographic staging (Cellini et al., 2015; Griessenberger et al., 2013; Shambroom et al., 2012; Tonetti et al., 2013) and has been successfully used in prior studies to reactivate memories during slow wave sleep (Honma et al., 2016), as well as to reactivate puzzle memories during sleep-incubation (Sanders et al., 2019). Participants placed the SMCS near their bed on a stable, elevated surface. Prior to sleeping, participants snapped three sensors (single-use silver/silver-chloride electrodes) into the Zeo wireless transmitter. Participants filled each sensor with an electrolyte (signa gel), exposed the adhesive backing on the sensors, and affixed the transmitter to their forehead. Once the SMCS was started, it employed an algorithm to determine a participant’s stage of sleep every 30 seconds.
When the SMCS detected participants were asleep, it began quietly playing pink noise. The pink noise continued throughout the entire night to reduce the likelihood that the puzzle-associated sound cues would wake up participants. After the SMCS detected more than 1.5 minutes of slow wave sleep (SWS), it presented one of the sounds associated with an unsolved puzzle from the evening session. If the sleep staging algorithm detected a participant was no longer in SWS, the sound cues immediately stopped and only resumed when the algorithm again detected SWS. Over the course of each night, half of the unsolved-puzzle-associated sounds were presented. The three chosen sounds were presented in a random order that then repeated throughout the night. Each sound played for 3 minutes (12 repetitions of the 15-second sound cue) before switching to the next sound. On average, each intended cue was presented 72.33 times (18.08 minutes) total across the night of sleep (SD = 43.72, range = 0 to 192) and cueing was similar for Less-Fixating (M = 73.29, SD = 44.16) and More-Fixating puzzle versions (M = 71.48, SD = 43.45, t(288) = 0.35, p = .725, dz = 0.02). If intended cues were not presented at all due to the SMCS detecting little to no SWS, the associated puzzles were later recategorized as Uncued (see Data Exclusions below).
Morning.
When participants woke up in the morning, they removed the transmitter from their forehead, stopped the recording, and returned to the lab with the SMCS. Participants generally completed morning sessions between 8 AM and 11 AM. During the morning sessions, participants first answered a questionnaire about the quality of their sleep, whether they heard any sounds during the night and, if so, which ones; and whether they solved any of the unsolved puzzles between the evening and morning sessions. Participants then completed a quick sound-puzzle recall test at the beginning of the morning session. Each sound was presented once, and then participants reported aloud what puzzle went with the sound. Overall participants remembered 69.5% (SD = 22.4%) of the sound-puzzle pairings. Participants’ accuracy was comparable for puzzles that were Cued (M = 68.6%, SD = 26.9%) and those that were not cued (M = 70.4%, SD = 26.7%, t(57) = −0.41, p = .680, dz = 0.05) and for the Less-Fixating (M = 69.7%, SD = 25.5%) and More-Fixating (M = 69.7%, SD = 25.8%, t(57) = 0.02, p = .986, dz = 0.003) puzzles.
After this quick sound-puzzle memory test, participants saw the full version of each of the unsolved puzzles from the previous evening, one at a time, and attempted to solve them. Before each puzzle was presented, participants were told what type of puzzle would be next (Matchstick, Rebus, Spatial, or Verbal). Participants then saw the puzzle title and recalled aloud as many details about the puzzle as they could remember. To prevent participants from considering the fixating information less important and therefore not worth reporting even if they remembered it, the instructions emphasized that participants should report all aspects of the puzzle including any details, examples, or hints. They then saw the full puzzle for four minutes and attempted to solve it. As in the evening session, when participants had an idea, they pressed the spacebar and reported their solution. If the solution was incorrect, the experimenter explained why the solution would not work and then participants had the remaining time to work on the puzzle. If participants did not solve the puzzle in the allotted time, they reported any final thoughts or guesses and, if still unsolved, they were told the solution to the puzzle before the next puzzle was presented. Experimenters in the morning sessions were blind to whether a puzzle was cued or not during the previous overnight session.
Memory Scoring.
Before attempting to solve a puzzle in the morning, participants were presented with the puzzle’s title and asked to verbally report as many details about the puzzle as they can remember. Participants were given as much time as they needed, and the experimenter advanced the screen when the participant said he or she was done. All responses to the memory prompt were recorded and later transcribed for all participants who consented to be audio recorded. For the six participants who did not consent to audio recording, memory scores were calculated based on the experimenter’s notes. To quantify participants’ memory for the puzzle details (the number of which varied by puzzle types and by specific puzzles), we created a scoring guide for each puzzle that consisted of a list of all the elements in the puzzle that a participant could potentially remember including actors, objects, goals, rules, and minor details. Using the transcription of participants’ responses, two independent raters coded each response for the presence or absence of each of the elements from the scoring guide. Initially, approximately one third of the data, equally spread across the forty puzzles and two fixation conditions, was coded by both raters (total agreement = 91.8%, Cohen’s kappa = 0.84, p < .001). Disagreements were reconciled through discussion among the raters. After the high level of agreement and discussion of discrepancies, each rater then scored half of the remaining two thirds of the data individually. Raters were blind to each puzzle’s cueing and fixating conditions and whether the puzzle was solved. We restricted our analysis to puzzle elements that were relevant to the solution of the puzzle and were not provided by the puzzle’s title. These relevant puzzle elements were identical across the More-Fixating and Less-Fixating versions of the puzzle. We then calculated the percent of relevant puzzle elements that were remembered out of the total number of relevant puzzle elements that could be remembered for each puzzle. Several puzzles’ relevant elements were completely contained in the puzzle’s title (for example the rebus puzzle, GESG, is just those four letters and therefore “GESG” is also the puzzle’s title). These puzzles were excluded from the memory analysis. In addition to the puzzle’s relevant details, for the More-Fixating versions of the puzzles only, we separately calculated the proportion of the fixating information that participants remembered.
Data Exclusions.
As in Sanders et al. (2019), several pre-determined data exclusion criteria were employed. From the 58 participants included in the analysis, 5 nights (4.3% of the data) were excluded from analysis due to the SMCS not picking up enough SWS to cue any sounds while the participant slept, likely due to bad adhesion of the sensor to the participant’s forehead. If at least some sound cues were presented during a night of sleep, but a specific sound was not, the unplayed sound was included in the analysis but marked as Uncued (2.2% of the data). Individual puzzles were excluded if the puzzle was solved between the evening session and the participant falling asleep the night before (1.7% of the data), the participant reported hearing the sound associated with the puzzle during their night of sleep (4.5% of the data), or the experimenter incorrectly classified a puzzle as solved/unsolved (2.0% of the data). Finally, the first 16 participants in this experiment attempted Compound Remote Associate (CRA) items in addition to the matchstick, rebus, spatial and verbal puzzles until 8 puzzles remained unsolved (the 6 described + 2 CRAs). We removed the CRA puzzles from the experiment for the remaining 42 participants due to low puzzle recall and we excluded the first 16 participants’ CRA puzzles from the analysis here.
Results
The effects of fixating information and overnight cueing on participants’ solving and puzzle memory the following morning were primarily analyzed using repeated measures analysis of variance (ANOVA) and correlations. P-values are reported for 2-tailed comparisons, except where noted for specific directional predictions. We additionally report results from a logistic mixed-effects regression model fitted in R (R Core Team, 2019) using the glmer() and lmer() functions of the lme4 package (Bates et al., 2015) that included random intercepts for both subject and item as random effects (see Supplemental Material).
Fixation Manipulation Check
In this study we manipulated the stimuli to create one version that was more likely to induce fixation and a second version that was less likely to induce fixation. If the manipulation was successful, participants should solve puzzles more often in the Less-Fixating condition than in the More-Fixating condition, despite the puzzles being nearly identical and requiring the same solution. Indeed, in the evening participants solved more puzzles when the puzzles were presented in their Less-Fixating version (M = 18.3%, SD = 17.3%) than in their More-Fixating version (M = 13.5%, SD = 12.9%, t(57) = 1.89, p = .032, one-tailed, dz = 0.25, Figure 2a). Similar differences remained in the morning session with participants solving more puzzles when the puzzles were presented in their Less-Fixating version (M = 26.4%, SD = 23.8%) than in their More-Fixating version (M = 20.3%, SD = 19.8%) puzzles (t(57) = 1.61, p = .056, one-tailed, dz = 0.21, Figure 2b).
Figure 2.

Solving rates for Less-Fixating (LF) and More-Fixating (MF) puzzles. (a) Evening solving rates. (b) Morning solving rates. Error bars represent within-subject standard error.
Morning Solving
As previously stated, presenting associated sound cues during sleep should trigger the reactivation of puzzle information. For the More-Fixating puzzles, the cues will also reactivate the fixating information. If passive forgetting fixation occurs, cueing should impede forgetting of the fixating information in the More-Fixating version of the puzzles, reducing any benefits of cueing on solving. In contrast, if unconscious processing occurs, reactivating both the puzzle information and the fixating information at the same time could facilitate memory reorganization enhancing the benefits of cueing. Therefore, passive forgetting fixation predicts a smaller effect of cueing on problem solving for the More-Fixating than Less-Fixating puzzles whereas unconscious processing predicts a larger cueing effect for the More-Fixating versions of the puzzles.
To test this hypothesis, we conducted a 2 (Cue) x 2 (Fixation) repeated measures ANOVA. We did not find a main effect of Cue (F(1,54) = 0.18, p = .677, ηp2 = .003) or Fixation (F(1,54) = 2.54, p = .117, ηp2 = .05). However, most importantly, we did find a significant crossover interaction (F(1,54) = 4.56, p = .037, ηp2 = .08). Descriptively, the cueing effect was larger in the More-Fixating than the Less-Fixating puzzles, consistent with the unconscious processing hypothesis. Planned contrasts did not find statistical differences between solving of the Cued and Uncued puzzles for either the More-Fixating puzzle versions (Mcued = 24.7%, SDcued = 30.8%, Muncued = 15.6%, SDuncued = 19.9%, t(56) = 1.70, p = .094, dz = 0.23 or Less-Fixating puzzle versions (Mcued = 22.7%, SDcued = 31.7%, Muncued = 27.6%, SDuncued = 28.7%, t(54) = −1.00, p = .321, dz = 0.13, Figure 3). Phrased another way, when puzzles were not cued, participants solved more of the puzzles presented in their Less-Fixating version than their More-Fixating version (t(56) = 3.02, p = .004, dz = 0.40). However, cueing erased the Fixation effect such that participants solved puzzles equally well in their Less-Fixating and More-Fixating versions (t(54) = −.30, p = .766, dz = 0.04).
Figure 3.

Solving rates for Uncued and Cued puzzles presented in their Less-Fixating (left) and More-Fixating (right) puzzle versions. Error bars represent within-subject standard error.
Morning Shared Memory
Although puzzle materials were designed to test solving rates, we also examined participants’ memory for the relevant details of the puzzles prior to seeing and reattempting the puzzles again in the morning. Based on the prior literature, if we had a more sensitive memory measure we would expect to find better puzzle memory for cued compared to uncued puzzles and for memory to predict solving more in the cued than the uncued condition. We did not expect an interaction with the fixation condition because the same relevant puzzle details were present in both the More-Fixating and Less-Fixating versions of the puzzles.
To test these hypotheses, we conducted a 2 (Cue) x 2 (Fixation) repeated measures ANOVA. We did not find a main effect of Cue. Participants recalled similar amounts of solution- (not fixation-) relevant details from Cued (M = 77.7%, SD = 18.8%) and Uncued (M = 81.2%, SD = 11.6%) puzzles (F(1,52) = 0.55, p = .462, ηp2 = .01). We also did not find a main effect of Fixation. Participants remembered similar amounts of relevant puzzle details for the Less-Fixating (M = 81.5%, SD = 13.2%) and More-Fixating versions of the puzzles (M = 78.2%, SD = 14.15%, F(52) = 0.39, p = .534, ηp2 = .01). Finally, Fixation did not interact with cueing for memory scores (F(1,52) = 0.61, p = .438, ηp2 = .01).
As in Sanders et al. (2019), we also examined correlations between solving and memory for relevant puzzle details (uncorrected for multiple comparisons), which were the same for both the Less-Fixating and More-Fixating puzzle versions. Not surprisingly, given that puzzles were not designed to measure memory, most of the correlations between memory and solving were not reliable on their own. However, for sake of completeness these correlations are listed in Tables S1 and S2. Although we cannot draw strong conclusions, we note that More-Fixating and Less-Fixating puzzles showed opposite patterns in terms of how cueing interacted with the relation between solving and memory. For More-Fixating puzzles, the better participants remembered Cued puzzles, the more they solved Cued puzzles (r(55) = .20, p = .143) while the better participants remembered Uncued puzzles, the less they solved Uncued puzzles (r(55) = - .20, p = .137, see Table S2), and these correlations significantly differed from each other (z = 2.11, p = .036) although neither was individually different from zero. In contrast, for the Less-Fixating puzzles, the opposite pattern of correlations between solving and memory was found. For Cued puzzles the better participants remembered the puzzles, the fewer puzzles they solved (r(53) = −.19, p = .161) while for Uncued puzzles the better participants remembered the puzzles, the more puzzles they solved (r(53) = .17, p = .210, see Table S2); although the difference between these correlations was not reliable (z = - 1.80, p = .072), but was in the opposite direction from that of the More-Fixating puzzles.
Morning Fixation Memory
In addition to the details that were shared across the Less-Fixating and More-Fixating versions of the puzzles, More-Fixating puzzle versions contained additional fixating information meant to reinforce incorrect approaches to the puzzle (see Materials). We therefore tested whether, within the More-Fixating puzzles, cueing affected memory for this additional information. As for the solution-relevant analyses we do not find statistically reliable results; however, we again note an interesting pattern. Participants remembered the fixating information similarly well for Cued (M = 35.5%, SD = 33.2%) and Uncued puzzles (M = 38.3%, SD = 27.4%, t(56) = −0.39, p = .699, dz = 0.05). If anything, participants remembered the fixating information worse when puzzles were cued than when puzzles were not cued. In addition, memory for fixating information did not reliably correlate with solving within the More-Fixating puzzles, although all correlations were descriptively negative (Overall: r(56) = - .20, p = .131; Cued: r(56) = - .14, p = .311; Uncued: r(56) = - .03, p = .847) suggesting that, if anything, the more likely participants forgot the fixating information for cued puzzles, the better they solved them.
Finally, we tested whether differential forgetting of the fixating compared to the relevant information predicted solving. We calculated a relevant minus fixation difference score for each participant’s More-Fixating puzzle memory: percent of relevant information recalled minus percent of fixating information recalled (only occurring the More-Fixating condition). This relevant-fixation differential positively correlated with solving the More-Fixating puzzles (r(56) = .30, p = .022) such that the better participants remembered relevant information compared to fixating information, the better they solved the More-Fixating puzzles. In addition, although not statistically significant, again we find a differential pattern such that the correlation was positive for the Cued puzzles (r(56) = .22, p = .095) and negative for the Uncued puzzles ( r(55) = −.10, p = .481) although these correlations did not differ from each other (z = 1.69, p = .091). Together these results provide a preliminary hint that differential memory for relevant compared to irrelevant information may relate to better solving particularly when puzzles are cued overnight.
Comparison to Prior Study
Contrary to our expectations, we did not find an overall cueing effect across the two fixation versions of the puzzles in this study. One possibility is that adjusting the puzzles to make them More-/Less- fixating created variation in the cueing effect. Therefore, we examined the effect of cueing on the unaltered puzzles that are identical to the versions Sanders et al. (2019) used in their study. Of the 40 puzzles, 32 were taken from the previous study. Then, of those 32, only one of the versions (either the More-Fixating or Less-Fixating version) was identical to the puzzle from the prior study. Therefore, this replication analysis is underpowered to detect statistically reliable results due to substantial reduction of the data that could be included leading to less stable measurement of solution rates. Specifically, only 232 total data points are included in this analysis and of these puzzles, on average, each participant experienced only 1.76 Cued and 2.24 Uncued puzzles substantially reducing the reliability of the solving measure. To slightly mitigate this, instead of calculating solving across the 58 participants, we calculated the Cued and Uncued solving rates across the 32 unaltered puzzles (on average, 3.19 Cued and 4.06 Uncued observations per puzzle) and compare it to the solving rates of the same 32 puzzles from Sanders et al. (2019).
Evening solving rates of the unaltered puzzles were similar for this study (M = 14.6%, SD = 13.7%) and the previous study (M = 16.5%, SD = 16.41%, t(31) = −0.68, p = .501, dz = 0.12) and solving rates were positively correlated (r(30) = .44, p = .012). Morning solving rates were also similar for this study (M = 28.3%, SD = 20.5%) and the previous study (M = 27.1%, SD = 18.3%, t(31) = 0.31, p = .761, dz = 0.05) suggesting that, on average, solving ability was similar across the two studies. In addition, in this study the puzzles were, descriptively but not reliably, solved more often in the Cued (M = 33.3%, SD = 34.9%) than the Uncued condition (M = 27.6%, SD = 26.3%, t(30) = 0.86, p = .199, one-tailed, dz = 0.15, see Figure 4a). Although the difference in this study was not reliable, the Cued and Uncued solving rates are similar to those found in the original study for the same subset of puzzles (Mcued = 33.2%, SDcued = 21.9%, Muncued = 23.8%, SDuncued = 21.6%) and, in a 2 (Cueing) x 2 (Study) repeated measures ANOVA we find a main effect of Cueing (F(1,30) = 4.38, p = .045, ηp2 = .13, but no main effect of Study (F(1,30) = .36, p = .554, ηp2 = .01) nor an interaction (F(1,30) = .13, p = .720, ηp2 = .004). To investigate whether these results provide evidence for a replication of the original study, we calculated a replication Bayes Factor (Verhagen & Wagenmakers, 2014)2. Our analysis yielded ambiguous evidence for replication (BF = 0.60) suggesting that the data are 1.66 times more likely to have occurred under the null hypothesis than under the replication hypothesis. We additionally found an equality-of-effect-size BF of 3.98 suggesting modest evidence that the cueing effects were of similar size in the two studies using the identical puzzles. Taken together, these results provide an underpowered, descriptive replication of an overall cueing effect on problem solving.
Figure 4.

Solving rates for only puzzle versions that are identical between the current study and Sanders et al., 2019. (a) Uncued vs. Cued in the current study for the shared puzzles. (b) Uncued vs. Cued puzzle solutions in the Sanders et al. 2019 study for the shared puzzles. Error bars represent within-puzzle standard error.
Discussion
In this study we examined how reactivating puzzles for processing during sleep facilitates problem solving in the context of two theories of awake incubation facilitation of problem solving, Forgetting Fixation and Unconscious Processing. Participants attempted More-Fixating and Less-Fixating versions of puzzles in an evening session. Then, overnight half of each of the types of puzzles were cued during overnight sleep using paired sound cues. Cueing (sound cue presented versus not presented) interacted with puzzle version (More-Fixating vs. Less-Fixating) to affect the number of puzzles participants solved in the morning. Although the cueing effects (Cued minus Uncued solving) were not statistically reliable within each puzzle version, the interaction was reliable because cueing affected participants’ solving in opposite directions for the two puzzle types: participants solved slightly more Cued than Uncued of the More-Fixating versions of the puzzles, while solving slightly fewer Cued than Uncued of the Less-Fixating versions of the puzzles.
Overall, these results argue against a simple, passive forgetting fixation theory, which predicts that cueing should lead to poorer solving when more fixating information was included. Given the previous literature demonstrating targeted memory reactivation during sleep strengthens memory (Hu et al., 2020; Oudiette & Paller, 2013), the sound cues likely reactivated all the puzzle information, including both the relevant puzzle details and the additional fixating information included in the More-Fixating puzzles. Yet, we found that cueing erased the effect of adding fixating information to the puzzles. These results suggest that instead of impeding problem solving, reactivating fixating information alongside the puzzle memory enabled unconscious processing to occur such that the fixating information could be overcome.
Although the puzzles were designed for solving, not memory, we examined the relations between solution-relevant details recall, and fixating-information details recall and solving. No strong conclusions can be drawn because most of these under-powered comparisons were not statistically reliable, but the overall pattern suggests tentative, supplemental support that unconscious processing is involved when sleep-cueing improves next-day solving. Rather than uniform memory strengthening or forgetting, the pattern looks like differential forgetting. Specifically, participants who remember solution-relevant details better than fixating information details solved reliably more of the More-Fixating puzzles. Although inconclusive, these results are consistent with the interpretation that rather than universal memory strengthening or weakening, cueing facilitated better memory for relevant parts of the puzzle over others which in turn facilitated solving. The individual patterns of results for solution-relevant puzzle details and fixating-information details are not statistically reliable but tentatively support this position. Within the More-Fixating puzzles, Cued memory for relevant puzzle was positively associated with Cued solving while Uncued memory for relevant puzzle details was negatively associated with worse Uncued solving, and these correlations statistically differed from each other although neither was reliable on its own. The opposing correlations suggest that cueing did impact solving although, as expected, our measures of memory may not be sensitive enough to detect an overall cueing effect on memory for the puzzle details. In contrast, memory for fixating information in More-Fixating puzzles generally predicted worse solving, although not statistically reliable.
Although preliminary and inconclusive, this overall pattern tentatively suggests that when cueing differentially affected memory such that relevant information was strengthened and fixating information was more likely to be forgotten, solving was also facilitated. This pattern is in line with research that suggests sleep may enhance differential memory strengthening, particularly if it is directed to be forgotten (Saletin et al., 2011; Simon et al., 2018) or of a lower value or salience than other information (Cunningham & Payne, 2017; Oudiette et al., 2013; Payne et al., 2008). Rather than directly opposing forgetting fixation and unconscious processing theories, this would imply a hybrid theory whereby unconscious processing occurs to identify, prioritize, and reorganize memory but that the outcome results in forgetting fixating information.
The extent and type of unconscious processing that may occur during asleep incubation remains to be explored. A simple, passive fixation forgetting caused by the passage of time cannot explain why cueing would differentially impact More-Fixating and Less-Fixating puzzles such that cueing would benefit More-Fixating puzzles more than Less-Fixating puzzles. This is especially unlikely since participants saw the puzzles with the accompanying fixating information again in the morning preventing the fixating information from being fully forgotten. However, an adaptation of the forgetting fixation hypothesis to allow for differential forgetting could suggest that reactivation of the puzzle memory during sleep temporarily destabilizes the memory (Nadel et al., 2012), presenting another opportunity for forgetting to occur. In this account, an additional process would have to determine what information is worth consolidating versus forgetting, perhaps through tagging of the fixating information as less relevant (Cunningham & Payne, 2017; Stickgold & Walker, 2013) or through coherence of the puzzle information (Bowers et al., 1990; Yaniv & Meyer, 1987). In these cases, differential forgetting is responsible for the cueing effect on solving.
Alternatively, differential forgetting of fixating versus solution-relevant information could occur as a side effect of problem restructuring during sleep. What form this restructuring might take is unknown and could potentially differ for different puzzles. Some possibilities include spreading activation connecting puzzle information to relevant prior knowledge, re-prioritizing the critical puzzle information, or discovering links between parts of the puzzle that were previously unconnected. Although inspired by research on sleep and memory reorganization, these ideas closely relate to theories of restructuring during awake problem solving. For example, Representational Change Theory (RCT, Knoblich et al., 1999; Ohlsson, 1992; Öllinger et al., 2013) suggests that people solve these types of problems through relaxing prepotent constraints or decomposing the elements of the problem and recombining them in new ways. These processes could also occur during sleep. Constraint relaxation is similar to the reprioritization and differential forgetting we propose here whereby incorrect assumptions that are reinforced with cueing may be strategically deprioritized during sleep such that when returning to the puzzle, they are no longer as salient. Similarly, chunk decomposition may occur during sleep through reactivation of the puzzle elements leading to individual components being reorganized and connected to participants’ prior knowledge. Then, as a result of restructuring, the fixating information could be forgotten.
In contrast, a different interpretation of these results suggests that adding the fixating information may have acted as a hint by directing participants’ attention to the challenging part of the puzzle or otherwise making the problem space more clearly understood. However, this explanation is unlikely because the More-Fixating versions of the puzzles were less likely to be solved as a whole, suggesting that the additional information made the puzzle more challenging, not less so.
The lack of an overall cueing effect in the study is unexpected because we previously found a cueing effect in a prior study (Sanders et al., 2019). However, when we restrict our analysis to only versions of the puzzles that were unaltered from those in the prior study, we find a similarly sized cueing effect (although not reliably different from zero). Although inconclusive, this suggests that variations in the puzzles caused by manipulating the fixating information may have reduced the cueing effectiveness. When creating the More-/Less-Fixating versions of the puzzles, we endeavored to identify and reinforce the most salient piece of fixating information in each puzzle. However, due to the variation in types of puzzles and types of fixating information, this led to a heterogeneous set of puzzles. Future studies could focus on a smaller set of puzzles and types of fixating information to reduce this variability.
We found an overall effect of fixation such that participants solved the same puzzles less often when the puzzles were presented with an additional piece of misleading information; however, the size of this effect was smaller than many other forgetting fixation studies. One possibility is that the complexity and distinctiveness of the puzzles - a strength in the TMR paradigm so puzzles are not easily confused - may conversely result in large variability in the effectiveness of the added fixating information. Even within a single complex puzzle, there are often multiple ways participants can become stuck (e.g. Kershaw & Ohlsson, 2004) and it is not possible here to determine whether adding the fixating information truly increased the salience of the intended misconception or in a different way. To begin to address this concern, some future research could use a think-aloud protocol (e.g. Cranford & Moss, 2012) to more precisely assess whether participants become stuck in the way the misleading information intended, and more so in the More-Fixating than Less-Fixating versions of the puzzles.
Our design differed from prior forgetting fixation studies in several ways. First, the puzzles we used are more complex than the simple word puzzles often used in the prior literature (e.g. Smith & Blankenship, 1989, 1991). This complexity makes it challenging to precisely determine how participants get fixated, and the levels and types of fixations may vary across participants. Indeed, participants’ own self-generated fixations may be more effective than experimenter designed fixations that externally reinforce one of the many ways participants could potentially get stuck. Future experiments could consider determining participants’ own individual fixations for each puzzle (e.g., through think-aloud protocols) and cueing their individual fixating information specifically. Second, to prevent participants’ awareness of the fixating information, the same version of the puzzle was presented again in the morning session rather than removing the fixating information upon retest as in many other studies. This enhances the likelihood that any advantageous processing resulting in increased solving occurred during the incubation period. In fact, it is possible that if the fixating information is removed at retest, the cueing effect for More-Fixating puzzles may be even larger because participants are not reminded of the fixating information again. Finally, the incubation period we employed was much longer (hours compared to minutes) than standard forgetting fixation studies. It is possible that at different incubation durations and under different conditions, some processes and incubation theories may be more linked to incubation effects than others.
In conclusion, we found a stronger cueing effect for More-Fixating compared to Less-Fixating puzzles, arguing against a simple passive forgetting fixation hypothesis. Furthermore the pattern of correlations between memory and problem solving tentatively suggest a hybrid theory, in which restructuring during sleep may selectively enhance relevant information while also devaluing less relevant, fixating information. Future research will continue to further elucidate the cognitive mechanisms that enhance problem solving during sleep; and sleep, as a unique kind of incubation period, may reciprocally enhance our understanding of incubation more generally.
Supplementary Material
Acknowledgements
We thank Ken A. Paller, Susan Florczak, Todd Anderson, Daniel Wetmore, and Sheepdog Sciences for the use of, and help with, the sleep monitoring and cueing system devices. We additionally thank Christopher Krause, Lane Patterson, Sophia McCullough, Alec Friswold, and James Crisafulli for data-collection and memory-coding assistance.
Funding Details
This work was supported by the National Institutes of Health’s National Institute of Child Health and Human Development awarded to M. Beeman under Grant R03HD087111.
Appendix A
| Puzzle Type | Less-Fixating Version | More-Fixating Version | Dominant Representation Reinforced | Solution |
|---|---|---|---|---|
|
| ||||
| Matchstick | Arrange these 7 matches to form 4 equal-sized triangles. All matches must be flat on the surface.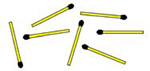
|
Arrange these 7 matches to form 4 equal-sized triangles. All matches must be flat on the surface.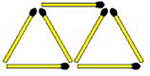
|
When participants hear equal-sized they often start by forming equilateral triangles. Starting with an equilateral triangle structure reinforces this approach. |
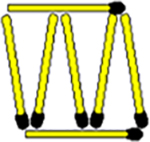
|
|
| ||||
| Rebus |

|

|
Participants often focus on the smaller words present in the puzzle (“no”, “hi”), the hint focuses the problem-solver on these smaller words. | H2O (water) |
| Hint: the word “no” is not in the answer. | ||||
|
| ||||
| Spatial | How can a round birthday cake be cut into eight pieces of equal size with only three straight cuts? | How can a round birthday cake be cut into eight pieces of equal size with only three straight cuts? For example: This cake is cut into eight pieces of equal size but with 4 straight cuts |
The two-dimensional picture and the depiction of standard cutting of eighths reinforces both a flat, 2D representation of the problem and the dominant way a cake is usually cut (from above). | Two vertical cuts to cut the cake into quarters and one horizontal cut parallel to the plate to divide the quarters into eighths. |
| ||||
|
| ||||
| Verbal | Two blind men walk into the same store at the same time on a cool summer night and with some help from a salesperson each one gets two pairs of white socks and two pairs of black socks. The two men arrive at the checkout counter at the same time and the cashier, so surprised by this event, puts all the socks into one bag, giving it to one man while giving the other man an empty bag. Outside the two blind men discover what happened and all by themselves with no help from anyone else they sort the socks so that each man now has two pairs of white socks and two pairs of black socks. How did they accomplish this? | Two blind men walk into the same store at the same time on a cool summer night and with some help from a salesperson each one gets two pairs of white socks and two pairs of black socks. The two men arrive at the checkout counter at the same time and the cashier, so surprised by this event, puts all the socks into one bag, giving it to one man while giving the other man an empty bag. Outside the two blind men discover what happened and all by themselves with no help from anyone else they sort the socks so that each man now has two pairs of white socks and two pairs of black socks. How did they accomplish this? Hint: The socks are the same size and shape and were not put in the bag in any particular order. |
The hint reinforces two approaches participants often take – identifying physical characteristics of the socks that might distinguish them from each other, or focusing on procedural parts of the puzzle. | Pairs of socks consist of two socks connected to each other. Separate each pair of socks so that each man gets one of the two socks in the pair. At the end, each man will have 4 single black socks and 4 single white socks (a total of 2 pairs of each). |
Appendix B
Morning solving rates for each puzzle overall and by Fixation and Cueing conditions
| Puzzle | Puzzle Type | Solved (%) | Solved Less-Fixating Uncued (%) | Solved Less-Fixating Cued (%) | Solved More-Fixating Uncued (%) | Solved More-Fixating Cued (%) |
|---|---|---|---|---|---|---|
| 3 Triangles to 4 Triangles | matchstick | 21.7 | 0.0 | 50.0 | 0.0 | 40.0 |
| 4 Complete Squares | matchstick | 10.0 | 25.0 | 0.0 | 14.3 | 0.0 |
| 6 to 9 | matchstick | 16.7 | 25.0 | 0.0 | 0.0 | 33.3 |
| 7 Matches 4 Triangles | matchstick | 21.7 | 33.3 | 14.3 | 20.0 | 0.0 |
| Divide in Two | matchstick | 18.2 | 14.3 | 40.0 | 16.7 | 0.0 |
| Equation | matchstick | 30.8 | 0.0 | 50.0 | 20.0 | 50.0 |
| Fish | matchstick | 80.0 | 75.0 | 50.0 | 100.0 | 100.0 |
| Matchstick Conversion | matchstick | 36.4 | 0.0 | 100.0 | 50.0 | 50.0 |
| Pig | matchstick | 37.5 | 0.0 | 100.0 | 50.0 | 25.0 |
| 0BScMAPhD | rebus | 35.7 | 33.3 | 50.0 | 25.0 | 33.3 |
| ALL | rebus | 9.1 | 0.0 | 0.0 | 0.0 | 33.3 |
| BEND DRAW DRAW | rebus | 63.6 | 100.0 | 100.0 | 50.0 | 33.3 |
| GESG | rebus | 40.0 | 100.0 | 0.0 | 0.0 | 50.0 |
| Hearts | rebus | 17.4 | 10.0 | 0.0 | 33.3 | 100.0 |
| HIJKLMNO | rebus | 5.3 | 0.0 | 16.7 | 0.0 | 0.0 |
| HOROBOD | rebus | 20.0 | 66.7 | 0.0 | 0.0 | 0.0 |
| JUSTTTTING | rebus | 17.6 | 0.0 | 16.7 | 25.0 | 100.0 |
| RPI | rebus | 6.7 | 16.7 | 0.0 | 0.0 | 0.0 |
| SMOKEG | rebus | 30.8 | 66.7 | 33.3 | 25.0 | 0.0 |
| 12 Coin Square | spatial | 22.2 | 28.6 | 0.0 | 100.0 | 20.0 |
| 8 Coins | spatial | 16.7 | 50.0 | 0.0 | 0.0 | 0.0 |
| 9 Dot | spatial | 8.3 | 0.0 | 0.0 | 0.0 | 50.0 |
| Birthday Cake | spatial | 21.4 | 50.0 | 50.0 | 0.0 | 0.0 |
| Chain | spatial | 33.3 | 75.0 | 100.0 | 0.0 | 27.3 |
| Figure | spatial | 21.4 | 0.0 | 33.3 | 16.7 | 25.0 |
| Flipped Triangle | spatial | 44.4 | 40.0 | 100.0 | 33.3 | |
| Lines of Pennies | spatial | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Rabbit Pens | spatial | 54.5 | 25.0 | 66.7 | 75.0 | |
| Trees | spatial | 16.7 | 20.0 | 50.0 | 0.0 | 0.0 |
| Two Rings | spatial | 40.0 | 50.0 | 100.0 | 0.0 | 33.3 |
| 10 Bags of 10 Coins | verbal | 15.0 | 0.0 | 20.0 | 0.0 | 50.0 |
| Cleopatra’s Diamonds | verbal | 16.7 | 0.0 | 25.0 | 50.0 | 0.0 |
| Lightbulb | verbal | 14.3 | 33.3 | 33.3 | 0.0 | 0.0 |
| Murder Trial | verbal | 13.3 | 0.0 | 0.0 | 16.7 | 16.7 |
| Power Cords | verbal | 33.3 | 0.0 | 0.0 | 100.0 | |
| Socks | verbal | 14.3 | 25.0 | 0.0 | 0.0 | 25.0 |
| The Balloon Game | verbal | 23.5 | 50.0 | 0.0 | 20.0 | 0.0 |
| Three Fuses | verbal | 4.3 | 14.3 | 0.0 | 0.0 | 0.0 |
| Three Suitors | verbal | 23.5 | 40.0 | 25.0 | 20.0 | 0.0 |
| Truth Tellers and Liars | verbal | 17.6 | 20.0 | 28.6 | 0.0 | 0.0 |
Footnotes
Disclosure of Interest
No potential conflict of interest was reported by the authors.
This analysis incorporates the t test statistic and degrees of freedom from the original study into the analysis to derive a Bayes Factor (BF) that indicates the likelihood that the replication data more closely reflect the hypothesis that Cued puzzles are solved more than Uncued puzzles, or the null hypothesis. BFs greater than 1 indicate evidence for the experimental hypothesis, BFs less than 1 indicate evidence for the null hypothesis. Generally BFs between 0.33 and 3 are considered inconclusive while BFs less than 0.1 or greater than 10 are considered strong evidence (Wetzels et al., 2011).
Data Availability Statement
The data that support the findings of this study are available at https://osf.io/bucpe/.
References
- Baird B, Smallwood J, Mrazek MD, Kam JWY, Franklin MS, & Schooler JW (2012). Inspired by distraction mind wandering facilitates creative incubation. Psychological Science, 23(10), 1117–1122. 10.1177/0956797612446024 [DOI] [PubMed] [Google Scholar]
- Barrett Deirdre. (2001). The committee of sleep: How artists, scientists, and athletes use dreams for creative problem-solving—And how you can, too. Crown Publishers. [Google Scholar]
- Bates D, Machler M, Bolker B, & Walker S (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software, 67(1), 1–48. 10.18637/jss.v067.i01 [DOI] [Google Scholar]
- Beda Z, & Smith SM (2018). Chasing red herrings: Memory of distractors causes fixation in creative problem solving. Memory & Cognition, 46(5), 671–684. 10.3758/s13421-018-0799-3 [DOI] [PubMed] [Google Scholar]
- Beijamini F, Pereira SIR, Cini FA, & Louzada FM (2014). After being challenged by a video game problem, sleep increases the chance to solve it. PLOS ONE, 9(1), e84342. 10.1371/journal.pone.0084342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowden EM, & Beeman MJ (1998). Getting the right idea: Semantic activation in the right hemisphere may help solve insight problems. Psychological Science, 9(6), 435–440. 10.1111/1467-9280.00082 [DOI] [Google Scholar]
- Bowers KS, Regehr G, Balthazard C, & Parker K (1990). Intuition in the context of discovery. Cognitive Psychology, 22(1), 72–110. 10.1016/0010-0285(90)90004-N [DOI] [Google Scholar]
- Cai DJ, Mednick SA, Harrison EM, Kanady JC, & Mednick SC (2009). REM, not incubation, improves creativity by priming associative networks. Proceedings of the National Academy of Sciences, 106(25), 10130–10134. 10.1073/pnas.0900271106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cellini N, McDevitt EA, Ricker AA, Rowe KM, & Mednick SC (2015). Validation of an automated wireless system for sleep monitoring during daytime naps. Behavioral Sleep Medicine, 13(2), 157–168. 10.1080/15402002.2013.845782 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cousins JN, El-Deredy W, Parkes LM, Hennies N, & Lewis PA (2014). Cued memory reactivation during slow-wave sleep promotes explicit knowledge of a motor sequence. The Journal of Neuroscience, 34(48), 15870–15876. 10.1523/JNEUROSCI.1011-14.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cranford E, & Moss J (2012). Is insight always the same? A protocol analysis of insight in compound remote associate problems. The Journal of Problem Solving, 4(2). 10.7771/1932-6246.1129 [DOI] [Google Scholar]
- Cunningham TJ, & Payne JD (2017). Emotional memory consolidation during sleep. In Cognitive Neuroscience of Memory Consolidation (pp. 133–159). Springer, Cham. 10.1007/978-3-319-45066-7_9 [DOI] [Google Scholar]
- Diekelmann S, & Born J (2010). The memory function of sleep. Nature Reviews Neuroscience, 11(2), 114–126. [DOI] [PubMed] [Google Scholar]
- Dodds RA, Ward TB, & Smith SM (2012). A review of experimental research on incubation in problem solving and creativity. In Runco MA (Ed.), Creativity research handbook (Vol. 3, pp. 251–284). Hampton Press. [Google Scholar]
- Dumay N, & Gaskell MG (2007). Sleep-associated changes in the mental representation of spoken words. Psychological Science, 18(1), 35–39. 10.1111/j.1467-9280.2007.01845.x [DOI] [PubMed] [Google Scholar]
- Durrant SJ, Cairney SA, McDermott C, & Lewis PA (2015). Schema-conformant memories are preferentially consolidated during REM sleep. Neurobiology of Learning and Memory, 122, 41–50. 10.1016/j.nlm.2015.02.011 [DOI] [PubMed] [Google Scholar]
- Ellenbogen JM, Hu PT, Payne JD, Titone D, & Walker MP (2007). Human relational memory requires time and sleep. Proceedings of the National Academy of Sciences, 104(18), 7723–7728. 10.1073/pnas.0700094104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer S, Drosopoulos S, Tsen J, & Born J (2006). Implicit learning–explicit knowing: A role for sleep in memory system interaction. Journal of Cognitive Neuroscience, 18(3), 311–319. 10.1162/jocn.2006.18.3.311 [DOI] [PubMed] [Google Scholar]
- Gilhooly KJ (2016). Incubation and intuition in creative problem solving. Frontiers in Psychology, 7. 10.3389/fpsyg.2016.01076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilhooly KJ (2019). Incubation in problem solving and creativity: Unconscious processes. Routledge. 10.4324/9781315147611 [DOI] [Google Scholar]
- Gilhooly KJ, Georgiou G, & Devery U (2013). Incubation and creativity: Do something different. Thinking & Reasoning, 19(2), 137–149. 10.1080/13546783.2012.749812 [DOI] [Google Scholar]
- Gilhooly KJ, Georgiou GJ, Garrison J, Reston JD, & Sirota M (2012). Don’t wait to incubate: Immediate versus delayed incubation in divergent thinking. Memory & Cognition, 40(6), 966–975. 10.3758/s13421-012-0199-z [DOI] [PubMed] [Google Scholar]
- Griessenberger H, Heib DPJ, Kunz AB, Hoedlmoser K, & Schabus M (2013). Assessment of a wireless headband for automatic sleep scoring. Sleep and Breathing, 17(2), 747–752. 10.1007/s11325-012-0757-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hélie S, & Sun R (2010). Incubation, insight, and creative problem solving: A unified theory and a connectionist model. Psychological Review, 117(3), 994–1024. 10.1037/a0019532 [DOI] [PubMed] [Google Scholar]
- Honma M, Plass J, Brang D, Florczak SM, Grabowecky M, & Paller KA (2016). Sleeping on the rubber-hand illusion: Memory reactivation during sleep facilitates multisensory recalibration. Neuroscience of Consciousness, 2016(1), niw020. 10.1093/nc/niw020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu X, Cheng LY, Chiu MH, & Paller KA (2020). Promoting memory consolidation during sleep: A meta-analysis of targeted memory reactivation. Psychological Bulletin, 146(3), 218–244. 10.1037/bul0000223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kershaw TC, & Ohlsson S (2004). Multiple causes of difficulty in insight: The case of the nine-dot problem. Journal of Experimental Psychology: Learning, Memory, and Cognition, 30(1), 3–13. 10.1037/0278-7393.30.1.3 [DOI] [PubMed] [Google Scholar]
- Knoblich G, Ohlsson S, Haider H, & Rhenius D (1999). Constraint relaxation and chunk decomposition in insight problem solving. Journal of Experimental Psychology: Learning, Memory, and Cognition, 25(6), 1534–1555. 10.1037/0278-7393.25.6.1534 [DOI] [Google Scholar]
- Kohn N, & Smith SM (2009). Partly versus completely out of your mind: Effects of incubation and distraction on resolving fixation. The Journal of Creative Behavior, 43(2), 102–118. 10.1002/j.2162-6057.2009.tb01309.x [DOI] [Google Scholar]
- Kounios J, & Beeman M (2014). The cognitive neuroscience of insight. Annual Review of Psychology, 65(1), 71–93. 10.1146/annurev-psych-010213-115154 [DOI] [PubMed] [Google Scholar]
- Lewis PA, & Durrant SJ (2011). Overlapping memory replay during sleep builds cognitive schemata. Trends in Cognitive Sciences, 15(8), 343–351. 10.1016/j.tics.2011.06.004 [DOI] [PubMed] [Google Scholar]
- Lewis PA, Knoblich G, & Poe G (2018). How memory replay in sleep boosts creative problem-solving. Trends in Cognitive Sciences, 22(6), 491–503. 10.1016/j.tics.2018.03.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazzarello P (2000). What dreams may come? Nature, 408(6812), 523–523. 10.1038/35046170 [DOI] [PubMed] [Google Scholar]
- Nadel L, Hupbach A, Gomez R, & Newman-Smith K (2012). Memory formation, consolidation and transformation. Neuroscience & Biobehavioral Reviews, 36(7), 1640–1645. 10.1016/j.neubiorev.2012.03.001 [DOI] [PubMed] [Google Scholar]
- Ohlsson S (1992). Information-processing explanations of insight and related phenomena. In Keane MT & Gilhooly KJ (Eds.), Advances in the psychology of thinking (pp. 1–43). Harvester Wheatsheaf. [Google Scholar]
- Öllinger M, Jones G, Faber AH, & Knoblich G (2013). Cognitive mechanisms of insight: The role of heuristics and representational change in solving the eight-coin problem. Journal of Experimental Psychology: Learning, Memory, and Cognition, 39(3), 931–939. 10.1037/a0029194 [DOI] [PubMed] [Google Scholar]
- Oudiette D, Antony JW, Creery JD, & Paller KA (2013). The role of memory reactivation during wakefulness and sleep in determining which memories endure. Journal of Neuroscience, 33(15), 6672–6678. 10.1523/JNEUROSCI.5497-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oudiette D, & Paller KA (2013). Upgrading the sleeping brain with targeted memory reactivation. Trends in Cognitive Sciences, 17(3), 142–149. 10.1016/j.tics.2013.01.006 [DOI] [PubMed] [Google Scholar]
- Payne JD (2011). Learning, memory, and sleep in humans. Sleep Medicine Clinics, 6(1), 15–30. 10.1016/j.jsmc.2010.12.005 [DOI] [Google Scholar]
- Payne JD, Stickgold R, Swanberg K, & Kensinger EA (2008). Sleep preferentially enhances memory for emotional components of scenes. Psychological Science, 19(8), 781–788. 10.1111/j.1467-9280.2008.02157.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team. (2019). R: A language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/ [Google Scholar]
- Rasch B, & Born J (2013). About sleep’s role in memory. Physiological Reviews, 93(2), 681–766. 10.1152/physrev.00032.2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasch B, Büchel C, Gais S, & Born J (2007). Odor cues during slow-wave sleep prompt declarative memory consolidation. Science, 315(5817), 1426–1429. 10.1126/science.1138581 [DOI] [PubMed] [Google Scholar]
- Ritter SM, & Dijksterhuis A (2014). Creativity—The unconscious foundations of the incubation period. Frontiers in Human Neuroscience, 8, 215. 10.3389/fnhum.2014.00215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudoy JD, Voss JL, Westerberg CE, & Paller KA (2009). Strengthening individual memories by reactivating them during sleep. Science, 326(5956), 1079–1079. 10.1126/science.1179013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saletin JM, Goldstein AN, & Walker MP (2011). The role of sleep in directed forgetting and remembering of human memories. Cerebral Cortex (New York, NY), 21(11), 2534–2541. 10.1093/cercor/bhr034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders KEG, Osburn S, Paller KA, & Beeman M (2019). Targeted memory reactivation during sleep improves next-day problem solving. Psychological Science, 30(11), 1616–1624. 10.1177/0956797619873344 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schönauer M, & Gais S (2017). The effect of sleep on multiple memory systems. In Cognitive Neuroscience of Memory Consolidation (pp. 105–115). Springer, Cham. 10.1007/978-3-319-45066-7_7 [DOI] [Google Scholar]
- Seifert CM, Meyer DE, Davidson N, Patalano AL, & Yaniv I (1995). Demystification of cognitive insight: Opportunistic assimilation and the prepared-mind perspective. In Sternberg RJ, Davidson JE, Sternberg RJ (Ed), & Davidson JE (Ed) (Eds.), The nature of insight. (1994–98855-003; pp. 65–124). The MIT Press. [Google Scholar]
- Shambroom JR, Fábregas SE, & Johnstone J (2012). Validation of an automated wireless system to monitor sleep in healthy adults. Journal of Sleep Research, 21(2), 221–230. 10.1111/j.1365-2869.2011.00944.x [DOI] [PubMed] [Google Scholar]
- Simon KCNS, Gómez RL, & Nadel L (2018). Losing memories during sleep after targeted memory reactivation. Neurobiology of Learning and Memory, 151, 10–17. 10.1016/j.nlm.2018.03.003 [DOI] [PubMed] [Google Scholar]
- Sio UN, Monaghan P, & Ormerod T (2013). Sleep on it, but only if it is difficult: Effects of sleep on problem solving. Memory & Cognition, 41(2), 159–166. [DOI] [PubMed] [Google Scholar]
- Sio UN, & Ormerod TC (2009). Does incubation enhance problem solving? A meta-analytic review. Psychological Bulletin, 135(1), 94–120. 10.1037/a0014212 [DOI] [PubMed] [Google Scholar]
- Smith SM (1995a). Fixation, incubation, and insight in memory and creative thinking. In Smith SM, Ward TB, Finke RA, Smith SM (Ed), Ward TB (Ed), & Finke RA (Ed) (Eds.), The creative cognition approach. (1995–97533-006; pp. 135–156). The MIT Press. [Google Scholar]
- Smith SM (1995b). Getting into and out of mental ruts: A theory of fixation, incubation, and insight. In Sternberg RJ, Davidson JE, Sternberg RJ (Ed), & Davidson JE (Ed) (Eds.), The nature of insight. (1994–98855-007; pp. 229–251). The MIT Press. [Google Scholar]
- Smith SM, & Beda Z (2019). Old problems in new contexts: The context-dependent fixation hypothesis. Journal of Experimental Psychology: General. 10.1037/xge0000615 [DOI] [PubMed] [Google Scholar]
- Smith SM, & Blankenship SE (1989). Incubation effects. Bulletin of the Psychonomic Society, 27(4), 311–314. 10.3758/BF03334612 [DOI] [Google Scholar]
- Smith SM, & Blankenship SE (1991). Incubation and the persistence of fixation in problem solving. The American Journal of Psychology, 104(1), 61–87. 10.2307/1422851 [DOI] [PubMed] [Google Scholar]
- Stickgold R, & Walker MP (2013). Sleep-dependent memory triage: Evolving generalization through selective processing. Nature Neuroscience, 16(2), 139–145. 10.1038/nn.3303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swearingen J (2016, November 1). An idea that stuck: How George de Mestral invented Velcro. The Vindicated. http://nymag.com/vindicated/2016/11/an-idea-that-stuck-how-george-de-mestral-invented-velcro.html [Google Scholar]
- Tonetti L, Cellini N, de Zambotti M, Fabbri M, Martoni M, Fábregas SE, Stegagno L, & Natale V (2013). Polysomnographic validation of a wireless dry headband technology for sleep monitoring in healthy young adults. Physiology & Behavior, 118, 185–188. 10.1016/j.physbeh.2013.05.036 [DOI] [PubMed] [Google Scholar]
- Verhagen J, & Wagenmakers E-J (2014). Bayesian tests to quantify the result of a replication attempt. Journal of Experimental Psychology: General, 143(4), 1457–1475. 10.1037/a0036731 [DOI] [PubMed] [Google Scholar]
- Wagner U, Gais S, Haider H, Verleger R, & Born J (2004). Sleep inspires insight. Nature, 427(6972), 352–355. 10.1038/nature02223 [DOI] [PubMed] [Google Scholar]
- Wallas G (1926). The art of thought (1926–10372-000). J. Cape: London. [Google Scholar]
- Wetzels R, Matzke D, Lee MD, Rouder JN, Iverson GJ, & Wagenmakers E-J (2011). Statistical Evidence in Experimental Psychology: An Empirical Comparison Using 855 t Tests. Perspectives on Psychological Science, 6(3), 291–298. 10.1177/1745691611406923 [DOI] [PubMed] [Google Scholar]
- Yaniv I, & Meyer DE (1987). Activation and metacognition of inaccessible stored information: Potential bases for incubation effects in problem solving. Journal of Experimental Psychology: Learning, Memory, and Cognition, 13(2), 187–205. 10.1037/0278-7393.13.2.187 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data that support the findings of this study are available at https://osf.io/bucpe/.