

Abstract
The dynamic regulation of blood flow is essential for meeting the high metabolic demands of the brain and maintaining brain function. Cerebral blood flow is regulated primarily by 1) the intrinsic mechanisms that determine vascular contractility and 2) signals from neurons and astrocytes that alter vascular contractility. Stimuli from neurons and astrocytes can also initiate a signaling cascade in the brain capillary endothelium to increase regional blood flow. Recent studies provide evidence that TRP channels in endothelial cells, smooth muscle cells, neurons, astrocytes, and perivascular nerves control cerebrovascular contractility and cerebral blood flow. TRP channels exert their functional effects either through cell membrane depolarization or by serving as a Ca2+ influx pathway. Endothelial cells and astrocytes also maintain the integrity of the blood-brain barrier. Both endothelial cells and astrocytes express TRP channels, and an increase in endothelial TRP channel activity has been linked with a disrupted endothelial barrier function. Therefore, TRP channels can play a potentially important role in regulating blood-brain barrier integrity. Here, we review the regulation of cerebrovascular contractility by TRP channels under healthy and disease conditions and their potential roles in maintaining blood-brain barrier function.
Keywords: TRP channels, Cerebral Microcirculation, Neurovascular Coupling, Blood-brain barrier, Cerebral Blood Flow
Introduction
The brain has high metabolic and energy needs. Since neurons cannot store oxygen and glucose, they rely heavily on a continuous blood supply to fulfill their metabolic requirements. The metabolic requirements are not uniform throughout the brain but instead increased in the regions of high neuronal activity. Therefore, a cellular cross-talk between neurons and the vasculature is of utmost importance in supporting the areas with high neuronal activity. Interruption of this “neurovascular” communication can impair regional blood flow within the brain and have detrimental effects on neuronal structure and function [59]. Contractility of the cerebral vasculature is a crucial determinant of cerebral blood flow. Vascular contractility, in turn, is determined by mechanisms intrinsic to blood vessels and mediators released by neighboring neurons and astrocytes [4, 13]. Intraluminal pressure is one of the most critical regulators of cerebrovascular contractility. Cerebral arteries and arterioles constrict in response to increased intraluminal pressure. This phenomenon, called myogenic constriction, regulates capillary hydrostatic pressure and cerebral blood flow. Myogenic constriction involves depolarization of smooth muscle cell (SMC) membrane and subsequent Ca2+ influx that results in SMC contraction [9, 59]. Localized changes in the levels of O2, H+, K+, Ca2+, glucose, adenosine, and adenosine triphosphate (ATP) can also alter vascular contractility and cerebral blood flow. Additionally, mediators released by nearby astrocytes and neurons can act on endothelial cells (ECs) and SMCs to alter vascular contractility and regional blood flow in the brain.
The cerebral circulation exhibits unique characteristics that minimize injury by maintaining optimal perfusion in response to physiological and pathological stimuli. The brain represents only 2% of the bodyweight but receives approximately 15% of the cardiac output. Blood supply to the brain occurs via two pairs of arteries: right and left common carotid arteries and right and left vertebral arteries. Each common carotid artery divides into the external carotid artery and internal carotid artery, the cerebrum’s principal blood supplier. The basilar artery and two internal carotid arteries form an anastomotic ring at the brain’s base (circle of Willis). The anterior, middle, and posterior cerebral arteries that branch to smaller arteries and arterioles run along the surface and then penetrate the brain parenchyma. The vascular tree branches further into arterioles and capillaries, dramatically increasing the overall vascular area, slowing the blood flow, and allowing diffusion of nutrients and metabolites [22, 23, 105].
The cerebrovascular wall consists of multiple cell types, including ECs, SMCs, pericytes, astrocytes, microglia, perivascular macrophages (PVMs), and perivascular fibroblast-like cells [121]. Vascular contraction and dilation are differentially regulated in the different segments of the cerebrovascular tree. Large arteries (carotid artery and middle cerebral artery) have multiple layers of contractile SMCs, which encircle the endothelial basement membrane. The number of SMCs decreases as the arteries become smaller (pial artery), eventually diminishing to a single cell layer in arterioles (penetrating parenchymal arterioles). The perivascular sensory nerves are limited to the pial arteries. A continuous endothelial monolayer forms the inner layer of the vascular walls. Adjacent ECs are securely connected to one another by tight junctions. On the luminal side, ECs are covered by glycocalyx glycoproteins, proteoglycans, and glycosaminoglycans [99]. Pericytes are embedded into the basement membrane of capillaries and pre-capillary arterioles and play a crucial role in regulating CBF and blood-brain barrier integrity (Fig. 1–3).
Figure 1: TRP channels regulate pial arterial contractility.
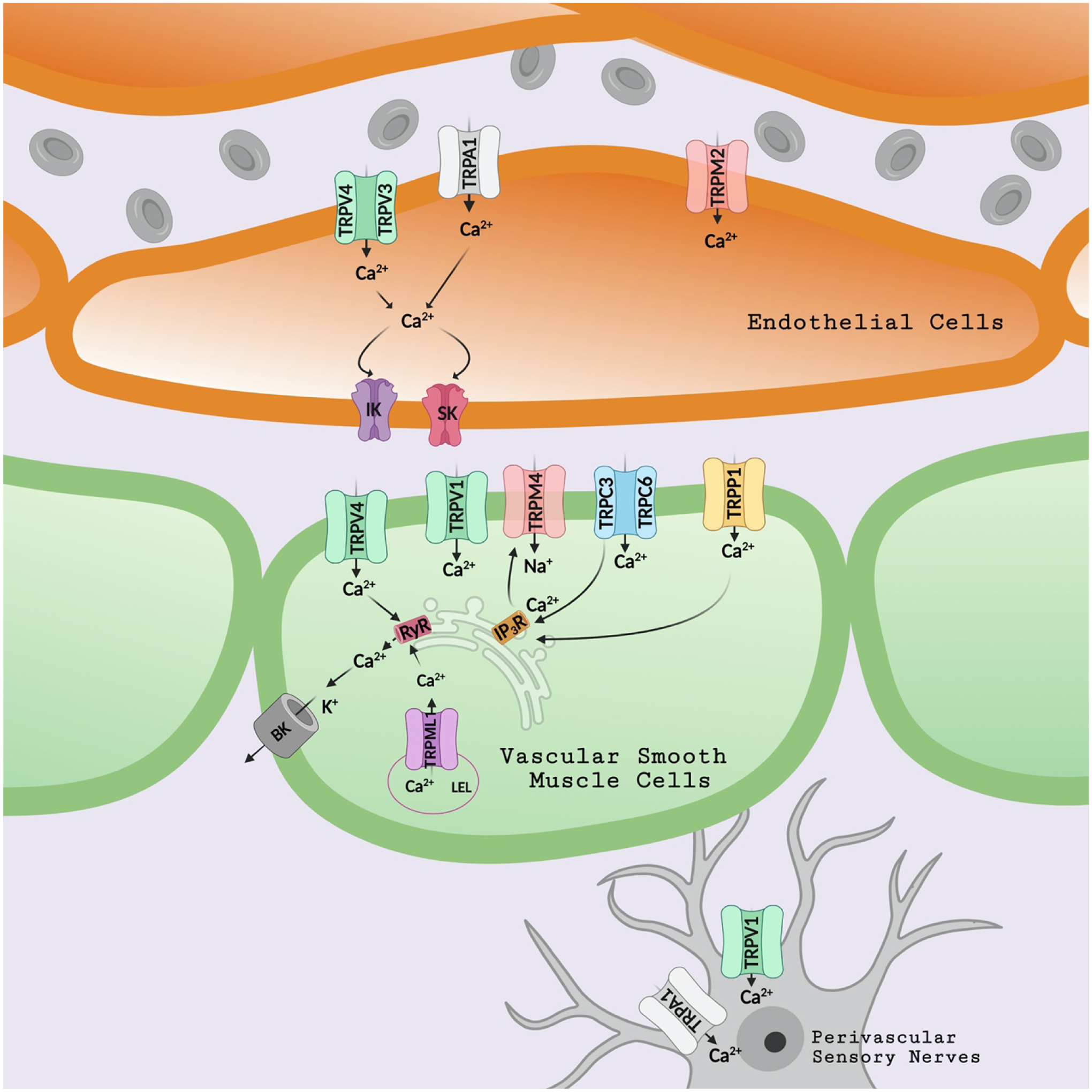
Endothelial Cells: Ca2+ influx through TRPV3, TRPV4, and TRPA1 channels activates nearby intermediate (IK) and small (SK) conductance Ca2+-activated potassium channels, causing vasodilation. TRPM2 channels are another Ca2+ entry pathway in cerebral endothelial cells. Smooth muscle cells: Ca2+ influx through plasma membrane TRPV4 channels or Ca2+ release through late endo-lysosome (LEL) TRPML1 channel activates ryanodine receptors (RyR), resulting in Ca2+ release (Ca2+ Sparks) from the sarcoplasmic reticulum (SR). Ca2+ sparks, in turn, stimulate large-conductance Ca2+-activated potassium (BK) channels, leading to SMC relaxation and vasodilation. Ca2+ entry through TRPC6, TRPC3, and TRPP1 channels triggers Ca2+ release from the SR via IP3 receptors (IP3R). IP3R Ca2+ signals activate sodium influx through TRPM4 channels, leading to SMC depolarization and vasoconstriction. Perivascular Sensory Nerves: Ca2+ entry via TRPA1 and TRPV1 channels induces the release of CGRP (calcitonin gene-ralated peptide) and vasodilation.
Figure 3: TRP channel regulation of neurovascular communication at the level of capillaries.
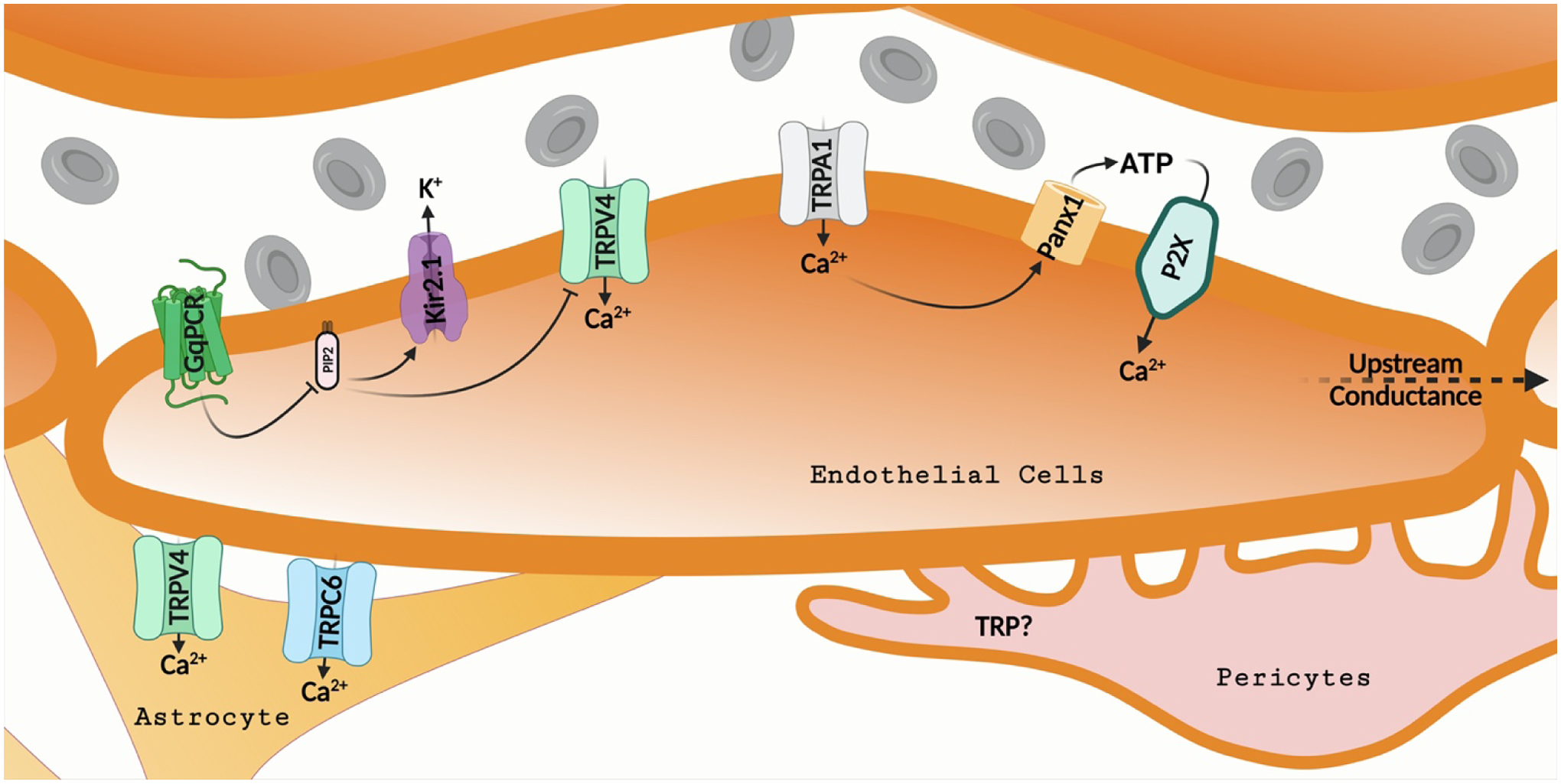
Ca2+ influx through TRPV4 channels is promoted by Gq protein-coupled receptor (GqPCR) signaling in the capillary endothelium. Activation of phospholipase C by Gq proteins lowers phosphatidylinositol 4,5-bisphosphate (PIP2) levels. PIP2 inhibits TRPV4 channels and activates inwardly rectifying potassium (Kir) channels. Thus, GqPCR signaling disinhibits TRPV4 channels and lowers the activity of Kir channels. Ca2+ influx signals through TRPA1 channels stimulate nearby Pannexin 1 channels, promoting ATP release. ATP activates Ca2+-permeable purinergic P2X receptor ion channels. The capillary Ca2+ events propagate upstream to the parenchymal arteriolar endothelium resulting in vasodilation. The role of TRP channels in pericytes is largely unknown.
The term “neurovascular communication (NVC)” represents the interactions of neurons or astrocytes with cerebrovascular ECs and SMCs [23, 47]. Vascular cells (ECs, SMCs, pericytes), astrocytes, and neurons work in tandem to regulate blood flow in the brain. Astrocytes are the regulatory hubs that receive signals from the active neurons and relay them to blood vessels, facilitating increased blood flow to the active neurons [5]. The network of astrocytes projects from neurons to parenchymal arterioles/capillaries, where astrocytic “endfeet” encase the parenchymal arterioles/capillaries. ECs and astrocytes also maintain the integrity of the blood-brain barrier [14]. The blood-brain barrier is absolutely essential for excluding neurotoxic plasma-derived components, red blood cells, leukocytes, and pathogens from the central nervous system (CNS) [4]. Blood-brain barrier integrity is maintained by specialized tight junctions that limit the hydraulic conductivity and transcellular movement. Cellular signaling mechanisms in the neurovascular unit control cerebral blood flow and blood-brain barrier integrity. While NVC implies passage of information from neurons to the vasculature, communication in the opposite direction (vasculo-neuronal coupling or VNC) has also been proposed. Together, NVC and VNC balance the energy supply with demands while preventing brain hyperperfusion or hypoperfusion. It is thought that the NVC overrides VNC during high neuronal activity [93].
Transient receptor potential (TRP) ion channels have emerged as essential regulators of cerebrovascular function and neurovascular communication. TRP channels are members of the superfamily of cation-conducting membrane proteins widely expressed in both excitable and non-excitable cells. TRP channels are activated by diverse stimuli, including temperature, light, pressure, changes in osmolarity, and G protein-coupled receptor (GPCR) signaling. Several TRP channels are expressed in the cerebral vasculature and neurovascular unit. TRP channels, based on their sequence homology, are classified into six subfamilies: TRPC (Canonical), TRPV (Vanilloid), TRPA (Ankyrin), TRPM (Melastatin), TRPP (Polycystic), and TRPML (Mucolipin). Broadly speaking, TRP channels act as cellular sensors that alter cellular function primarily by changing membrane potential and/or increasing cellular Ca2+. The molecular structure of TRP channels is characterized by four subunits, each presenting six hydrophobic transmembrane segments (S1-S6) and cytosolic N-and C- terminal tails [2, 8]. A short hydrophobic conserved domain among TRPV, TRPM, and TRPC subfamilies, referred to as the “TRP box,” is located after S6. TRP box is implicated in fine-tuning TRPV, TRPM, and TRPC channel activity.
TRP channels play crucial roles in various biological processes in the brain, including initiation of pain, thermoregulation, salivary fluid secretion, inflammation, pressure regulation, and Ca2+ and Mg2+ homeostasis. Particularly in blood vessels, GPCR signaling and other endogenous stimuli can activate TRP channels and enable their physiological roles. Accumulating evidence suggests that sex hormones can influence TRP channels expression and function, supporting potentially important roles for TRP channels in determining the sex differences in NVC and VNC [62]. TRP channels in cerebrovascular endothelium have primarily been associated with vasodilation. In contrast, TRP channels in SMCs can cause vasoconstriction or vasodilation depending on the channel involved and its downstream signaling targets. Here, we provide an overview of the role of cerebrovascular TRP channels in neurovascular communication and cerebral blood flow regulation.
TRPM channels
TRPM channels in both the SMC and EC layers play an essential role in regulating cerebrovascular function. The TRPM subfamily contains eight members: TRPM1-TRPM8. Most TRPM subfamily members are non-selective, Ca2+-permeable cation channels, except for TRPM4 and TRPM5, which are impermeable to Ca2+ [50].
TRPM4 channel has emerged as a critical player in pressure-induced depolarization of cerebrovascular SMCs. The unitary conductance of the TRPM4 channel is approximately 24 pS. TRPM4 channel is present in SMCs from the cerebral pial and parenchymal arterioles [67] (Fig. 1, Fig. 2). It is a Ca2+-activated, Ca2+-impermeable non-selective cation channel that contains binding sites for calmodulin (CaM) and ATP, and protein kinase C (PKC) phosphorylation sites [81]. Earley and colleagues presented the first evidence that SMC TRPM4 channels contribute to myogenic constriction of rat cerebral arteries [32, 96]. Further studies showed that TRPM4 channel-induced SMC depolarization and contraction are enhanced by angiotensin II receptor 1 (AT1R) stimulation [31] and PKC activation [37, 67] in male Sprague-Dawley rats. On the contrary, PKC inhibition impaired myogenic constriction of cerebral arteries [31]. Suppression of TRPM4 channel expression using antisense oligonucleotide caused a 70% to 85% decrease in myogenic constriction, further supporting an essential role of TRPM4 channels in the development of myogenic constriction of cerebral arteries. TRPM4 channels in cerebral arteries were also critical for purinergic P2Y receptor activation-induced vasoconstriction [67] and acted as a crucial link between P2Y receptors and myogenic constriction. Notably, TRPM4 channels were upregulated in a rat model of subarachnoid hemorrhage, increasing myogenic constriction of cerebral arteries and lowering cerebral blood flow [40]. Gonzales and colleagues observed that co-localization of TRPM4 channels with TRPC6 channels in cerebral artery myocytes facilitates stretch-induced membrane depolarization and myogenic constriction [41] in male Sprague-Dawley rats. The researchers explained that stretch activates TRPC6 channels directly or through AT1R-phospholipase Cγ1 (PLCγ1) signaling. The resulting Ca2+ influx activates inositol 1,4,5-trisphosphate (IP3) receptors on the sarcoplasmic reticular (SR) membrane. IP3 receptor-mediated Ca2+ release promotes the activity of TRPM4 channels on the SMC membrane and leads to SMC membrane depolarization.
Figure 2: TRP channel regulation of parenchymal arteriolar contractility.
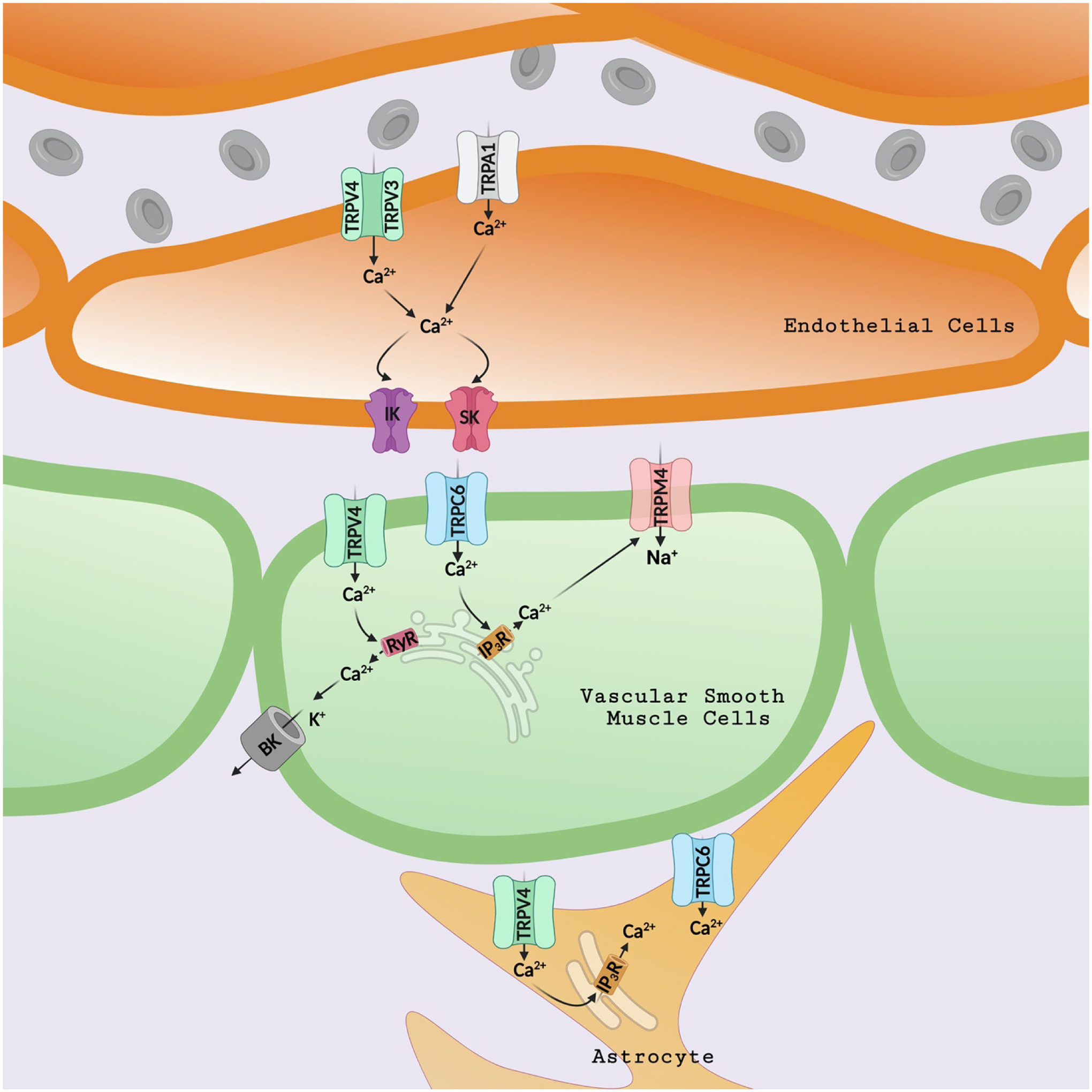
Endothelial Cells: Ca2+ influx through TRPV3, TRPV4, and TRPA1 channels activates nearby intermediate (IK) and small (SK) conductance Ca2+-activated potassium channels, causing vasodilation. Smooth muscle cells: Ca2+ entry through TRPC6 channels stimulates Ca2+ release from the sarcoplasmic reticulum (SR) via inositol triphosphate receptors (IP3R). IP3R-mediated Ca2+ release activates sodium currents through TRPM4 channels, depolarizing the SMC membrane and causing vasoconstriction. Ca2+ influx through TRPV4 channels stimulates Ca2+ release from the SR through ryanodine receptors (RyR) and activates large-conductance Ca2+-activated potassium channels (BK), leading to SMC hyperpolarization and vasodilation. Astrocytic Endfeet: Ca2+ signals through TRPV4 channels are amplified by Ca2+-induced Ca2+ release via IP3Rs. This mechanism increases boosts neurovascular coupling and dilates parenchymal arterioles. TRPC6 channel is another essential regulator of astrocytic Ca2+ homeostasis.
TRPM2 channel is a Ca2+-permeable non-selective cation channel highly expressed in the CNS [124], cerebral ECs [85], microglia [76], and pericytes [54]. TRPM2 channel is equally permeable to divalent and monovalent cations [101]. The channel is localized in the plasma membrane [120] and/or lysosomal compartments [64, 133]. TRPM2 channel embodies both ion channel functionality and enzymatic ADP-ribose (ADPR) hydrolase activity [119]. Several extracellular stimuli, including reactive oxygen species (ROS), H2O2, amyloid β peptide (Aβ), and Tumor necrosis factor- α (TNFα) were shown to induce TRPM2 channel activity via metabolic production of intracellular poly-ADPR [61, 107]. TRPM2 channel has also been proposed as an essential regulator of endothelial Ca2+ homeostasis. In dementia of Alzheimer’s disease (AD), the extracellular accumulation of Aβ triggered oxidative-nitrosative stress that activated the DNA repair enzyme poly-ADPR polymerase. The increase in poly-ADPR potentiated TRPM2 channel activity in ECs, leading to endothelial Ca2+ overload, endothelial dysfunction, and neurovascular impairment [85]. In the transient middle cerebral artery occlusion (tMCAO) model of stroke in male Sprague Dawley rats, microglial TRPM2 channel was upregulated through microglial activation and cellular infiltration [36]. It was proposed that under ischemia-reperfusion (I/R) conditions, persistent activation of TRPM2 channel results in prolonged membrane depolarization and elevation of [Ca2+]i and [Na+]I, resulting in microglial death. Therefore, targeting TRPM2 channels could be a therapeutic avenue to counteract the deleterious cerebrovascular effects of AD and I/R. Jia et al. [53] reported an important role for TRPM2 channels in sex differences during cerebral ischemia-induced oxidative stress. Inhibition or knockdown of TRPM2 channel significantly protected ischemia-induced neuronal death and reduced the infarct volume in male mice but not in female mice. Recent studies also show that TRPM7 channels contribute to ageing-related dementia [134], whereas TRPM8 channels inhibit TRPV1 channel-associated hypothermia [35]. Thus, the roles of different TRPM channels in the neurovascular system and sex-differences in neurovascular function is an emerging area of research.
TRPA1 channels
Recent developments establish that endothelial TRPA1 channel is a crucial Ca2+ entry pathway in neurovascular coupling and regulation of cerebral blood flow. TRPA1, the sole member of the mammalian ankyrin TRP subfamily, is a large-conductance (~100 pS) [79], Ca2+-permeable cation channel. TRPA1 channel is permeable to both monovalent and divalent cations, but its relative permeability is higher for Ca2+ than for K+ or Na+ [11]. Moreover, the activity of TRPA1 channels is regulated by both intracellular and extracellular Ca2+. TRPA1 channels are activated by several pungent natural compounds in food, including allicin (garlic), allyl isothiocyanate (mustard), and cinnamon (cinnamaldehyde) [6]. Importantly, TRPA1 channels are critical sensors of cellular redox signaling, a hallmark of cerebrovascular disease [90].
Endothelial TRPA1 channels in pial arteries (Fig. 1), parenchymal arterioles (Fig. 2), and capillaries (Fig. 3) play crucial roles in regulating cerebral blood flow. Pires et al. found that hypoxia and cerebral ischemia dilate cerebral pial arteries and parenchymal arterioles via activation of endothelial TRPA1 channels [90]. Acute hypoxia increased the frequency of TRPA1 sparklets (optically detected elementary Ca2+ influx signals [108, 113]) and dilated mouse cerebral arteries and penetrating parenchymal arterioles. Further, increased endothelial TRPA1 channel activity reduced cerebral infarct size. Hypoxia-induced dilation of cerebral arteries was disrupted by antioxidants, TRPA1 antagonist A967079, and EC-specific TRPA1 deletion in mice. In a recent study, Thakore et al. showed that capillary endothelial TRPA1 channels initiate a dilatory signal that is transmitted to upstream parenchymal arterioles. Ca2+ signals from capillary TRPA1 channels propagate upstream through the capillary network via endothelial Pannexin1 (Panx1, ATP release channel on the cell membrane), extracellular ATP release, and subsequent activation of Ca2+-permeable purinergic P2X receptor ion channels [116]. ATP derived from astrocytes, neurons, and capillary ECs can activate purinergic P2X receptors on adjacent ECs. Ca2+ signals initiated in the capillary endothelium propagate upstream to the arteriolar endothelium, where they activate Ca2+-sensitive SK and IK (small and intermediate conductance K+ channels, respectively) channels. SK and IK channel activation results in EC membrane hyperpolarization, which is transmitted to arteriolar SMCs via gap junctions, thereby dilating the parenchymal arterioles. Together, these studies demonstrated a central role for endothelial TRPA1 channels in neurovascular coupling [116].
TRPA1 channel is also found in sensory neurons [7, 110]. Neuronal activation leads to the release of calcitonin gene-related peptide (CGRP), a potent vasodilator. TRPA1 channels co-localize with CGRP in primary sensory neurons [110]. Activation of neuronal TRPA1 channels by mustard oil increases CGRP release and dilates cerebral vasculature [63], causing headaches. This effect is blocked by a CGRP antagonist or TRPA1 channel inhibitor. Similarly, in diet-induced obese rats, higher TRPA1 channel activity in trigeminal ganglia increased the CGRP release and elevated meningeal blood flow [71]. Moreover, activation of TRPA1 channels in perivascular sensory neurons triggered the release of CGRP, decreased vascular resistance, and increased cerebral blood flow [87]. A recent study also showed that TRPA1 channels may play a crucial role in ageing-related memory decline [12]. Thus, activation of endothelial TRPA1 channels has neuroprotective effects, whereas activation of neuronal TRPA1 channels can have detrimental consequences in the brain.
TRPP channels
TRPP1 and TRPP2 are Ca2+-permeable, non-selective cation channels with a large unitary conductance (~100 pS) [69]. Recent studies have revealed vital roles for TRPP1 channels in regulating myogenic constriction in different vascular beds [80, 106]. TRPP1 channels are expressed in cerebrovascular SMCs [80] and ECs [10], located on the endoplasmic reticular membrane [104] and plasma membrane [69]. In human and rat cerebral artery SMCs, TRPP1 channels are predominantly expressed at the plasma membrane [80] and contribute to myogenic constriction. RNAi-mediated knockdown of TRPP1 reduced the swelling-activated cation currents in isolated cerebrovascular SMCs and suppressed the development of myogenic constriction in cerebral arteries [80]. Moreover, TRPP1 haploinsufficient mice displayed impaired SMC Ca2+ homeostasis and elevated blood pressure [95]. Abdi and colleagues showed that TRPP2 channels interact with ryanodine receptors (RyR) and IP3 receptors in mouse cerebral artery SMCs [1]. Moreover, TRPP2 haploinsufficiency resulted in arterial remodeling of mouse middle cerebral arteries. Thus, recent evidence supports crucial roles for TRPP channels in regulating SMC Ca2+ homeostasis, although further studies are needed to understand these channels’ regulatory mechanisms and physiological roles in the vasculature. The development of specific pharmacological activators or inhibitors of TRPP channels will significantly improve our understanding of their properties and functional roles.
TRPML channels
The TRPML subfamily consists of Ca2+-permeable, non-selective cation channels (TRPML1, TRPML2, and TRPML3). TRPML1 is ubiquitously expressed, TRPML2 is expressed in lymphoid and myeloid tissues, and TRPML3 is expressed in the cochlea and the melanocytes [19]. Unlike other TRP channels present on the plasma membrane, TRPML1 is predominantly localized at the late endosomal and lysosomal (LELs) membranes. TRPML1 channels are permeable to Ca2+, Fe2+, Zn2+, Na+, and K+. The unitary conductance of the TRPML1 channel is 45 pS [125]. Recent studies indicate that TRPML1 channels are expressed in the SMCs from cerebral arteries [118], where they form a stable complex with RyR2s and are functionally coupled with RyR2s. The TRPML1-RyR2 signaling subsequently activates Ca2+-sensitive, large-conductance K+ (BK) channels in cerebrovascular SMCs, resulting in vasodilation. Global deficiency of the TRPML1 channel results in vascular hypercontractility and hypertension [118]. Therefore, SMC TRPML1 channel appears to be a vital regulator of cerebrovascular resistance and systemic blood pressure.
TRPC channels
TRPC subfamily is grouped into three major categories based on the amino acid sequence homology and functional similarities; TRPC1/4/5, TRPC2, and TRPC3/6/7 [23, 75]. The TRPC channel family consists of Ca2+-permeable, non-selective cation channels (TRPC1–7). TRPC1 functions as a receptor-operated, diacylglycerol (DAG)- and IP3 receptor-activated cation channel. TRPC2 is different from other TRPC subfamily members in that it encodes a non-functional truncated protein (pseudogene) in humans [122]. TRPC4 and TRPC5 function as non-selective cation channels activated by Gq/11 family GPCRs and receptor tyrosine kinases [103]. TRPC3, TRPC6, and TRPC7 show non-selective and double-rectifying cation currents when activated by Gq/11-coupled receptors or DAG as either homomeric or heteromeric channels [48].
TRPC3 channels are expressed in cerebral ECs [100], neurons [17], and SMCs [128]. The unitary conductance for the TRPC3 channel is approximately 68 pS [92]. TRPC3 channel is a key player in the receptor-mediated contraction of cerebral artery SMCs. Reading and colleagues showed that suppression of SMC TRPC3 channel expression with antisense oligonucleotide reduced uridine triphosphate (UTP)-induced constriction of rat cerebral arteries, but not pressure-induced constriction [97]. Further, IP3 receptor activation was shown to induce constriction through IP3 receptor–TRPC3 channel signaling in cerebral arteries [128]. In hyperperfusion disorders such as epilepsy and seizures, TRPC3 and TRPC6 channels are upregulated in hippocampal neurons [89, 131]. The upregulation of neuronal TRPC3 and TRPC6 channels correlates with impaired cerebral blood flow and a high degree of moment-to-moment variability in neuronal activity [24]. Deleting TRPC3 channels from SMCs rescued cerebral blood flow and neuronal activity in mice [24], implying a harmful effect of SMC TRPC3 channel activity on neurovascular communication.
TRPC6 channels are an integral part of the force-sensing complex in cerebral artery SMCs and a key regulator of myogenic constriction [41] (Fig. 1). Gonzales et al. showed that Ca2+ influx through TRPC6 channels in rat cerebral artery SMCs is activated by a direct mechanical stimulation or through the PLCγ–DAG pathway. Ca2+ influx through TRPC6 channels can increase IP3 receptor-mediated Ca2+ release from the SR, which, in turn, stimulates nearby TRPM4 channels [41]. In another study, Welsh and colleagues found that antisense oligonucleotides targeting TRPC6 channel mRNA attenuated pressure-induced membrane depolarization of rat cerebral artery SMCs and reduced myogenic constriction [127]. Notably, global TRPC6−/− mice showed increased myogenic constriction of cerebral arteries and elevated blood pressure. This surprising finding was attributed to a compensatory upregulation of TRPC3 channels in SMCs [28]. Thus, SMC TRPC3 and TRPC6 channels appear to be crucial controllers of receptor-mediated and myogenic constriction of cerebral arteries, respectively.
In forebrain ischemia, Ca2+ influx through SMC TRPC1 and TRPC6 channels and endothelin-1-induced constriction of middle cerebral arteries are increased [55]. Ischemia/reperfusion (I/R) and oxygen/glucose deprivation also upregulated TRPC6 channels in primary cortical neurons [18], contributing to Na+ entry, NMDA receptor activation, and Ca2+ overload that ultimately resulted in neuronal death. Deletion of TRPC6 channel protected the neurons against I/R-induced damage and excitatory neurotoxins such as glutamate and NMDA. On the contrary, the SMC TRPC6 channel was downregulated in the middle cerebral artery occlusion (MCAO) mouse model of I/R [68]. The brain infarct lesion, apoptosis, and interleukin-6 (IL-6) and IL-1β levels were increased, and the astrocyte population was reduced in this mouse model. Overexpression of astrocytic TRPC6 channel or HYP9, a selective TRPC6 agonist, attenuated the damage to astrocytes in this model [68]. Remarkably, TRPC3/6/7 triple knockout mouse ameliorated brain damage through decreased nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) expression and increased AKT phosphorylation, resulting in reduced astrocyte apoptosis [20, 94]. Overall, TRPC channels may serve as a crucial link between the nervous system and the vasculature in health and disease.
TRPC channel activity has also been linked with the maintenance of blood-brain barrier function. The expression of TRPC3 channel was upregulated in neurons and ECs within the brain’s piriform cortex in status epilepticus [100]. Upregulation of endothelial TRPC3 channels lowered the levels of SMI-71, a rat endothelial blood-brain barrier antigen, a finding that correlated with blood-brain barrier disruption and neuronal damage. On the contrary, Chlorpyrifos, an insecticide, decreased the expression of endothelial TRPC4 channels and disrupted the blood-brain barrier in Sprague–Dawley rats [66]. An in-depth assessment of the role of TRPC channels in maintaining blood-brain barrier integrity is awaited.
TRPV channels
TRPV channels are expressed on the cell membranes of SMCs, ECs, and perivascular nerves, where they serve as signal transducers activated by endogenous ligands, heat, mechanical stimuli, and osmotic stress. High unitary conductance and permeability for Ca2+ are characteristic properties of TRPV channels.
The first TRPV subfamily member, TRPV1, has a unitary conductance of 35 to 70 pS [15]. TRPV1 channels are essential for thermal and chemical nociception in sensory neurons and are activated by capsaicin, changes in pH, and temperature in the noxious range (>42 °C) [8]. TRPV1 channel activation in perivascular sensory nerves stimulates the release of CGRP and leads to vasodilation [43, 123]. TRPV1 reporter mice clearly showed the expression of TRPV1 channels in perivascular sensory neurons [16] and SMCs from cerebral arteries [88]. In contrast to the dilatory effect of neuronal TRPV1 channel activity, SMC TRPV1 channels induce vasoconstriction [16, 88]. Therefore, the activation of TRPV1 channels on SMCs could counteract nerve-related changes in vascular reactivity in response to endogenous TRPV1 agonists.
Pegorini et al. demonstrated that in vivo pretreatment with capsaicin, a TRPV1 channel agonist, was neuroprotective in a rodent model of global cerebral ischemia [86]. In a later study, Khatibi and colleagues confirmed these findings in a different model of brain ischemia [57]. Pre-ischemia injection of capsaicin reduced cerebral infarct volume and normalized vasoreactivity [57]. The authors speculated that capsaicin-induced desensitization of TRPV1 channels prevents Ca2+ overload and neurotoxicity following ischemia. Furthermore, genetic deletion of TRPV1 was protective against ischemia-induced neurological and motor deficits [77]. In a more recent study, Hakimizadeh and colleagues showed that inhibiting TRPV1 channels post-ischemia limits neuronal damage by lowering the expression of toll-like receptor 2 (TLR2) and TLR4 [42]. Indeed, TLR2 and TLR4 are upregulated following cerebral ischemia [42, 65] and play an essential role in sustaining neuronal inflammation and death [115]. Based on these reports, TRPV1 channels could be a novel therapeutic target against stroke-induced neuronal damage, and their roles in the pathogenesis of cerebral ischemia need to be further investigated.
Another TRPV subfamily member, TRPV4, is permeable to Ca2+ and Na+ and is widely distributed in the neurovascular system, found in ECs [45], SMCs [30], astrocytes [29], and neurons [73]. The single-channel conductance of TRPV4 channel is in the range of 90–100 pS for outward currents and 50–60 pS for inward currents [33]. In cerebral pial arteries, endogenous agonist 11,12 epoxyeicosatrienoic acid (11,12-EET) activated SMC TRPV4 channel currents. Ca2+ influx through TRPV4 channels signaled through RyR–BK channel to induce SMC hyperpolarization and vasodilation [30]. Elementary Ca2+ release signals (Ca2+ sparks) through RyRs located on the SR membrane activate BK channels [126]. Reducing the expression of TRPV4 channels with antisense oligonucleotides prevented the 11,12-EET-induced SMC hyperpolarization and dilation of cerebral pial arteries from male Sprague–Dawley rats. In SMCs from penetrating parenchymal arterioles, PKC anchoring by A-kinase-anchoring protein 150 (AKAP150) facilitated Ang II-activation of SMC TRPV4 channels. Furthermore, the AKAP150-TRPV4 channel interactions in SMCs were highly dynamic, with activation of Ang II signaling increasing the proximity between AKAP150 and TRPV4 channels [74]. Thus, TRPV4 channels in cerebrovascular SMCs have been associated with SMC relaxation and vasodilation (Fig. 1, Fig. 2). In this regard, important sex differnces were observed in the activity of SMC TRPV4 channels from cerebral arteries of male and female mice, with the overall activity being lower in female mice [114].
Endothelial TRPV4 channel activity promotes endothelium-dependent dilation in the cerebral arteries and is impaired in cerebrovascular disorders. Chang and colleagues showed that endothelial TRPV4 channels dilate pial arteries. This group further demonstrated that TRPV4 channel-dependent dilation of pial arteries is impaired in a mouse model of Alzheimer’s disease [132]. Using TRPV4 knockout rats, a recent study by Dorrance and colleagues reported that endothelial TRPV4 channel signaling is essential for parenchymal arteriolar dilation and cognitive function [26]. Earlier, Dorrance and colleagues showed that mineralocorticoid receptor antagonism increases TRPV4 channel expression in parenchymal arterioles and improves cognitive function [27] in hypertension, providing further evidence that endothelial TRPV4 channels have a protective effect on cognitive function.
Cerebral capillary ECs also show a robust expression and function of TRPV4 channels (Fig. 3). In the arterial ECs, coupling of TRPV4 channels with IK/SK channels or eNOS [72, 82, 108] results in SMC relaxation and vasodilation. Of note, the TRPV4-eNOS or TRPV4-IK/SK channel coupling is known to occur at endothelial projections to SMCs [84]. In the capillaries, however, there are no SMCs surrounding the ECs. Further, IK/SK channel currents were not observed in capillary ECs [70], suggesting a distinct TRPV4 channel-dependent signaling in capillary ECs compared to arterial ECs. Capillary ECs have been shown to express eNOS [75]. Therefore, it is plausible that TRPV4 channels in capillary ECs activate eNOS, although this possibility has not been verified. Harraz et al. showed that TRPV4 channels in capillary ECs are tonically inhibited by basal levels of phosphatidylinositol 4,5-bisphosphate (PIP2) [45]. PIP2 is a plasma membrane lipid hydrolyzed by Gq protein-coupled receptor (GqPCR) signaling into IP3 and DAG. In capillary ECs, activation of GqPCR reduced PIP2 levels and disinhibited TRPV4 channels, simultaneously reducing inwardly rectifying potassium (Kir2.1) channel activity [44]. Thus, PIP2 acts as a functional switch for TRPV4 and Kir2.1 channels (Fig. 3), both significant players in cerebral blood flow regulation.
The expression of TRPV4 channels at the astrocytic endfeet [9, 10] suggests that the channels may play a vital role in neurovascular communication. Astrocytic endfeet are critical microdomains for coupling neuronal activity with changes in cerebral blood flow [32]. In this regard, Straub et al. optically recorded the Ca2+ signals occurring at the astrocytic endfeet [111]. Neuronal activity induced a rapid increase in endfeet Ca2+, which preceded the dilation of adjacent arterioles. The authors concluded that spatially restricted IP3 receptor Ca2+ signals at the endfeet are sufficient to induce vasodilation [111]. Later, Dunn et al. provided the first evidence that Ca2+ signals through TRPV4 channels increased the strength of neurovascular coupling by augmenting neuronal activation-induced endfoot Ca2+ and arteriolar dilation. The authors reported that the amplification of TRPV4 Ca2+ signals by Ca2+-induced Ca2+ release via IP3 receptors [29] further boosts neurovascular coupling. Interestingly, Kim et al. demonstrated that astrocytic TRPV4 channels mediate pressure-induced constriction of parenchymal arterioles, thus modulating vascular contractility to match the neuronal activity [58]. Taken together, TRPV4 channel activity appears to be central to the physiological function of multiple cell types involved in neurovascular communication.
TRPV3 channels are large unitary conductance (~150–200 pS), Ca2+ and Na+ permeable cation channels. They are expressed in the endothelium of cerebral pial arteries and parenchymal arterioles [91]. Activation of TRPV3 channels causes endothelium-dependent hyperpolarization of SMCs in pial arteries and parenchymal arterioles. Ca2+ influx through TRPV3 channels activates SK and IK channel currents, resulting in endothelium-dependent vasodilation of pial arteries and parenchymal arterioles [91]. Moreover, the Ca2+ flux through a single TRPV3 channel was found to be greater than other TRP channels [91]; approximately three times greater than TRPV4 channels [112] and 1.5 times greater than TRPA1 channels [113]. Further studies are needed to understand the physiological activators and regulatory mechanisms for endothelial TRPV3 channels.
Conclusion and Perspectives
TRP channels in vascular ECs and SMCs, astrocytes, perivascular nerves, and neurons play a critical role in ensuring resource delivery for neural processing and neural homeostasis. TRP channels accomplish this by 1) directly regulating cerebrovascular contractility; and 2) indirectly influencing cerebrovascular contractility via signaling from neurons, astrocytes, and perivascular nerves. TRP channels control cerebrovascular contractility by acting as a Ca2+ influx pathway or depolarizing the membrane potential. Current evidence suggests that TRPA1 and TRPV channels are the main Ca2+ influx pathways in cerebrovascular ECs. TRPM4, TRPC3, TRPC6, TRPP, and TRPV are the main TRP channels in cerebrovascular SMCs. The diverse biophysical properties of TRP channels and their distinct downstream signaling targets enable their involvement in multiple signaling pathways that control cerebral blood flow. The involvement of TRP channels in cerebral blood flow autoregulation could also be determined by the local stimuli that activate a specific TRP channel, thereby ensuring functional specificity. Cerebral arteries and arterioles are hypercontractile in vascular disorders [51]. Moreover, neurovascular communication is impaired in vascular disorders such as hypertension and stroke [51, 52]. Further research is needed to determine the contribution of TRP channels in impaired neurovascular communication in vascular disorders.
Pericytes line capillaries in the brain. Recent studies show that brain pericytes have a substantial effect on cerebral blood flow [46]. TRP channel-dependent signaling mechanisms in brain capillary pericytes remain unexplored and may be an exciting direction for future investigations. Further, many TRP channels in ECs and SMCs require scaffolding/anchoring proteins for normal function [74, 83]. AKAP150 is an example of a scaffolding protein that facilitates post-translational modifications of TRP channels by anchoring kinases such as Protein Kinase A (PKA) and PKC [60]. Indeed, AKAP150 is expressed in multiple cell types in the brain. In a recent study, Tajada and collegues demonstrated the importance of AKAP150-PKC complexes in enhancing TRPV4 channel activity in myocytes isolated from mouse pial and parenchymal arteries [114]. Interestignly, Pagán and collegues identified a novel neuroprotective role for AKAP150 in astrocytes. AKAP150 was upregulated in astrocytes under ischemic conditions to facilitate astrocytic K+ clearance [98]. These studies provide initial evidence that AKAP150 may be a significant player in the regulation of neurovascular comunication.
Caveolin-1 is another important structural protein found within caveolae. Chow and colleagues showed an essential role for endothelial caveolae in neurovascular coupling [21]. Endothelial caveolin-1 anchors eNOS [39], and TRP channel activation has also been linked with eNOS activation [25]. Moreover, caveolin-1 has emerged as a vital scaffolding protein for TRP channels [25]. Therefore, it will be interesting to investigate whether caveolin-1–TRP channel interaction determines eNOS activity, neurovascular coupling, and cerebral blood flow. In systemic arteries, endothelial TRP channels play a central role in myoendothelial feedback, which involves SMC to EC signaling that limits SMC contraction. Endothelial TRPV4 channels were essential for negatively regulating SMC α1 adrenergic receptor-mediated constriction of systemic arteries [49]. Myoendothelial feedback mechanisms have not been studied in the cerebral vasculature and may play potentially important roles in regulating cerebral blood flow. IK/SK channels are not present in the brain capillary endothelium. However, Kir2.1 channels are functional and are crucial for conducted hyperpolarization. TRPV4 channel-induced vasodilation occurred through downstream activation of endothelial Kir2.1 channels in the systemic arteries [109]. Kir2.1 channels were also involved in flow-mediated dilation of systemic arteries [3]. However, a functional interaction between TRPV4 and Kir2.1 channels and its physiological roles in the brain capillary endothelium remain unknown.
Perivascular macrophages (PVM) were recently shown to impair neurovascular coupling through NADPH 2 Oxidase (NOX2) activation and production of superoxide radicals. This effect seems to be mediated by angiotensin II, which activates angiotensin receptor 1 expressed on PVM [34, 56, 129]. Other studies suggest that TRP channels can influence ATR1 activity [130]. Considering the expression of TRP channels in macrophages [102], future studies of PVM TRP channels may provide interesting insigths into the impairment of neurovascular communication.
Finally, the role of TRP channels in controlling the integrity of the blood-brain barrier remains largely unexplored. Endothelial TRP channels have been associated with increased capillary permeability [38, 117]. In this regard, caveolin-1 is an essential component of the transcellular barrier. An increased expression of caveolin-1 was linked with blood-brain barrier leakage following ischemic brain insult [78]. Therefore, the interaction of TRP channels with caveolin-1 or other scaffolding proteins in endothelial cells and astrocytes will be an exciting area for future research on the blood-brain barrier.
Highlights.
Overview of neurovascular communication and blood brain barrier function
Biophysical properties and roles of individual TRP channels involved in neurovascular communication
Conclusions and Future Research Directions
Funding.
This work was supported by grants from the NIH to SKS (HL146914 and HL142808).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- [1].Abdi A, Mazzocco C, Légeron FP, Yvert B, Macrez N, Morel JL, TRPP2 modulates ryanodine- and inositol-1,4,5-trisphosphate receptors-dependent Ca2+ signals in opposite ways in cerebral arteries. Cell Calcium, Vol. 58, 2015, pp. 467–475. [DOI] [PubMed] [Google Scholar]
- [2].Adamian L, Liang J, Prediction of transmembrane helix orientation in polytopic membrane proteins. BMC Structural Biology, Vol. 6, 2006, pp. 1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Ahn SJ, Fancher IS, Bian JT, Zhang CX, Schwab S, Gaffin R, Phillips SA, Levitan I, Inwardly rectifying K(+) channels are major contributors to flow-induced vasodilatation in resistance arteries, J Physiol 595 (2017) 2339–2364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Armstead WM, Cerebral Blood Flow Autoregulation and Dysautoregulation. Anesthesiol Clin, Vol. 34, 2016, pp. 465–477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Attwell D, Buchan AM, Charpak S, Lauritzen M, MacVicar BA, Newman EA, Glial and neuronal control of brain blood flow. Nature, Vol. 468, 2010, pp. 232–243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Bandell M, Story GM, Hwang SW, Viswanath V, Eid SR, Petrus MJ, Earley TJ, Patapoutian A, Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron, Vol. 41, 2004, pp. 849–857. [DOI] [PubMed] [Google Scholar]
- [7].Bautista DM, Movahed P, Hinman A, Axelsson HE, Sterner O, Högestätt ED, Julius D, Jordt SE, Zygmunt PM, Pungent products from garlic activate the sensory ion channel TRPA1. Proceedings of the National Academy of Sciences of the United States of America, Vol. 102, 2005, pp. 12248–12252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Baylie RL, Brayden JE, TRPV channels and vascular function. Acta Physiologica, Vol. 203, 2011, pp. 99–116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Bayliss WM, On the local reactions of the arterial wall to changes of internal pressure. The Journal of Physiology, Vol. 28, 1901, p. 1902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Berrout J, Jin M, O’Neil RG, Critical role of TRPP2 and TRPC1 channels in stretch-induced injury of blood-brain barrier endothelial cells. Brain Research, Vol. 1436, Elsevier B.V., 2012, pp. 1–12. [DOI] [PubMed] [Google Scholar]
- [11].Bobkov YV, Corey EA, Ache BW, The pore properties of human nociceptor channel TRPA1 evaluated in single channel recordings. Biochimica et Biophysica Acta - Biomembranes, Vol. 1808, Elsevier B.V., 2011, pp. 1120–1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Borbely E, Payrits M, Hunyady A, Mezo G, Pinter E, Important regulatory function of transient receptor potential ankyrin 1 receptors in age-related learning and memory alterations of mice, Geroscience 41 (2019) 643–654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Brayden JE, Earley S, Nelson MT, Reading S, Transient receptor potential (TRP) channels, vascular tone and autoregulation of cerebral blood flow. Clinical and Experimental Pharmacology and Physiology, Vol. 35, 2008, pp. 1116–1120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Cardoso FL, Brites D, Brito MA, Looking at the blood-brain barrier: molecular anatomy and possible investigation approaches, Brain Res Rev 64 (2010) 328–363. [DOI] [PubMed] [Google Scholar]
- [15].Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D, The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature, Vol. 389, 1997, pp. 816–824. [DOI] [PubMed] [Google Scholar]
- [16].Cavanaugh DJ, Chesler AT, Jackson AC, Sigal YM, Yamanaka H, Grant R, O’Donnell D, Nicoll RA, Shah NM, Julius D, Basbaum AI, Trpv1 reporter mice reveal highly restricted brain distribution and functional expression in arteriolar smooth muscle cells. Journal of Neuroscience, Vol. 31, 2011, pp. 5067–5077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Chen J, Li Z, Hatcher JT, Chen QH, Chen L, Wurster RD, Chan SL, Cheng Z, Deletion of TRPC6 attenuates NMDA receptor-mediated Ca2+ entry and Ca2+-induced neurotoxicity following cerebral ischemia and oxygen-glucose deprivation. Frontiers in Neuroscience, Vol. 11, 2017, pp. 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Chen J, Li Z, Hatcher JT, Chen QH, Chen L, Wurster RD, Chan SL, Cheng Z, Deletion of TRPC6 Attenuates NMDA Receptor-Mediated Ca(2+) Entry and Ca(2+)-Induced Neurotoxicity Following Cerebral Ischemia and Oxygen-Glucose Deprivation, Front Neurosci 11 (2017) 138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Chen Q, She J, Zeng W, Guo J, Xu H, Bai XC, Jiang Y, Structure of mammalian endolysosomal TRPML1 channel in nanodiscs. Nature, Vol. 550, Nature Publishing Group, 2017, pp. 415–418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Chen X, Lu M, He X, Ma L, Birnbaumer L, Liao Y, TRPC3/6/7 Knockdown Protects the Brain from Cerebral Ischemia Injury via Astrocyte Apoptosis Inhibition and Effects on NF-кB Translocation. Molecular Neurobiology, Vol. 54, 2017, pp. 7555–7566. [DOI] [PubMed] [Google Scholar]
- [21].Chow BW, Nunez V, Kaplan L, Granger AJ, Bistrong K, Zucker HL, Kumar P, Sabatini BL, Gu C, Caveolae in CNS arterioles mediate neurovascular coupling, Nature 579 (2020) 106–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Cipolla MJ, The cerebral circulation. INTEGRATED SYSTEMS PHYSIOLOGY: FROM MOLECULE TO FUNCTION, Vol. 2, 2010, p. 117. [Google Scholar]
- [23].Cipolla MJ, The cerebral circulation. Integrated systems physiology: from molecule to function. Morgan & Claypool Life Sciences, 2009, pp. 1170–1172. [PubMed] [Google Scholar]
- [24].Cozart MA, Phelan KD, Wu H, Mu S, Birnbaumer L, Rusch NJ, Zheng F, Vascular smooth muscle TRPC3 channels facilitate the inverse hemodynamic response during status epilepticus. Scientific Reports, Vol. 10, Springer US, 2020, pp. 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Daneva Z, Marziano C, Ottolini M, Chen YL, Baker TM, Kuppusamy M, Zhang A, Ta HQ, Reagan CE, Mihalek AD, Kasetti RB, Shen Y, Isakson BE, Minshall RD, Zode GS, Goncharova EA, Laubach VE, Sonkusare SK, Caveolar peroxynitrite formation impairs endothelial TRPV4 channels and elevates pulmonary arterial pressure in pulmonary hypertension, Proc Natl Acad Sci U S A 118 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Diaz-Otero JM, Yen TC, Ahmad A, Laimon-Thomson E, Abolibdeh B, Kelly K, Lewis MT, Wiseman RW, Jackson WF, Dorrance AM, Transient receptor potential vanilloid 4 channels are important regulators of parenchymal arteriole dilation and cognitive function, Microcirculation 26 (2019) e12535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Diaz-Otero JM, Yen TC, Fisher C, Bota D, Jackson WF, Dorrance AM, Mineralocorticoid receptor antagonism improves parenchymal arteriole dilation via a TRPV4-dependent mechanism and prevents cognitive dysfunction in hypertension, Am J Physiol Heart Circ Physiol 315 (2018) H1304–H1315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Dietrich A, Mederos y Schnitzler M, Gollasch M, Gross V, Storch U, Dubrovska G, Obst M, Yildirim E, Salanova B, Kalwa H, Essin K, Pinkenburg O, Luft FC, Gudermann T, Birnbaumer L, Increased Vascular Smooth Muscle Contractility in TRPC6−/− Mice. Molecular and Cellular Biology, Vol. 25, 2005, pp. 11191–11191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Dunn KM, Hill-Eubanks DC, Liedtke WB, Nelson MT, TRPV4 channels stimulate Ca2+-induced Ca2+ release in astrocytic endfeet and amplify neurovascular coupling responses. Proceedings of the National Academy of Sciences of the United States of America, Vol. 110, 2013, pp. 6157–6162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Earley S, Heppner TJ, Nelson MT, Brayden JE, TRPV4 forms a novel Ca2+ signaling complex with ryanodine receptors and BKCa channels. Circulation Research, Vol. 97, 2005, pp. 1270–1279. [DOI] [PubMed] [Google Scholar]
- [31].Earley S, Straub SV, Brayden JE, Protein kinase C regulates vascular myogenic tone through activation of TRPM4. American Journal of Physiology - Heart and Circulatory Physiology, Vol. 292, 2007, pp. 2613–2622. [DOI] [PubMed] [Google Scholar]
- [32].Earley S, Waldron BJ, Brayden JE, Critical role for transient receptor potential channel TRPM4 in myogenic constriction of cerebral arteries. Circulation Research, Vol. 95, 2004, pp. 922–929. [DOI] [PubMed] [Google Scholar]
- [33].Everaerts W, Nilius B, Owsianik G, The vanilloid transient receptor potential channel TRPV4: from structure to disease, Prog Biophys Mol Biol 103 (2010) 2–17. [DOI] [PubMed] [Google Scholar]
- [34].Faraco G, Sugiyama Y, Lane D, Garcia-Bonilla L, Chang H, Santisteban MM, Racchumi G, Murphy M, Van Rooijen N, Anrather J, Iadecola C, Perivascular macrophages mediate the neurovascular and cognitive dysfunction associated with hypertension, J Clin Invest 126 (2016) 4674–4689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Feketa VV, Zhang Y, Cao Z, Balasubramanian A, Flores CM, Player MR, Marrelli SP, Transient receptor potential melastatin 8 channel inhibition potentiates the hypothermic response to transient receptor potential vanilloid 1 activation in the conscious mouse, Crit Care Med 42 (2014) e355–363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Fonfria E, Mattei C, Hill K, Brown JT, Randall A, Benham CD, Skaper SD, Campbell CA, Crook B, Murdock PR, Wilson JM, Maurio FP, Owen DE, Tilling PL, McNulty S, TRPM2 is elevated in the tMCAO stroke model, transcriptionally regulated, and functionally expressed in C13 microglia. Journal of Receptors and Signal Transduction, Vol. 26, 2006, pp. 179–198. [DOI] [PubMed] [Google Scholar]
- [37].Garcia ZI, Bruhl A, Gonzales AL, Earley S, Basal protein kinase Cδ activity is required for membrane localization and activity of TRPM4 channels in cerebral artery smooth muscle cells. Channels, Vol. 5, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Genova T, Gaglioti D, Munaron L, Regulation of Vessel Permeability by TRP Channels, Front Physiol 11 (2020) 421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Goligorsky MS, Li H, Brodsky S, Chen J, Relationships between caveolae and eNOS: everything in proximity and the proximity of everything, Am J Physiol Renal Physiol 283 (2002) F1–10. [DOI] [PubMed] [Google Scholar]
- [40].Gong Y, Du M.y., Yu H.l., Yang Z.y., Li Y.j., Zhou L, Mei R, Yang L, Wang F, Increased TRPM4 Activity in Cerebral Artery Myocytes Contributes to Cerebral Blood Flow Reduction After Subarachnoid Hemorrhage in Rats. Neurotherapeutics, Vol. 16, Neurotherapeutics, 2019, pp. 901–911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Gonzales AL, Yang Y, Sullivan MN, Sanders L, Dabertrand F, Hill-Eubanks DC, Nelson MT, Earley S, A PLCγ1-dependent, force-sensitive signaling network in the myogenic constriction of cerebral arteries. Science Signaling, Vol. 7, 2014, pp. 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Hakimizadeh E, Shamsizadeh A, Roohbakhsh A, Kazemi Arababadi M, Hajizadeh MR, Shariati M, Fatemi I, Moghadam-Ahmadi A, Bazmandegan G, Rezazadeh H, Allahtavakoli M, TRPV1 receptor-mediated expression of toll-like receptors 2 and 4 following permanent middle cerebral artery occlusion in rats. Iranian Journal of Basic Medical Sciences, Vol. 20, 2017, pp. 863–869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Han S-P, Naes L, Westfall TC, Impaired vasodilation in response to perivascular nerve stimulation in mesenteric arteries of TRPV1-null mutant mice. Biochemical and Biophysical Research Communications, Vol. 168, 1990, pp. 786–791. [DOI] [PubMed] [Google Scholar]
- [44].Harraz OF, Hill-Eubanks D, Nelson MT, PIP2: A critical regulator of vascular ion channels hiding in plain sight. Proceedings of the National Academy of Sciences of the United States of America, Vol. 117, 2020, pp. 20378–20389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Harraz OF, Longden TA, Eubanks DH, Nelson MT, PIP2depletion promotes TRPV4 channel activity in mouse brain capillary endothelial cells. eLife, Vol. 7, 2018, pp. 1–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Hartmann DA, Berthiaume AA, Grant RI, Harrill SA, Koski T, Tieu T, McDowell KP, Faino AV, Kelly AL, Shih AY, Brain capillary pericytes exert a substantial but slow influence on blood flow, Nat Neurosci 24 (2021) 633–645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Hirsch S, Reichold J, Schneider M, Székely G, Weber B, Topology and hemodynamics of the cortical cerebrovascular system. Journal of Cerebral Blood Flow and Metabolism, Vol. 32, 2012, pp. 952–967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Hofmann T, Obukhov AG, Schaefer M, Harteneck C, Gudermann T, Schultz G, Direct activation of human TRPC6 and TRPC3 channels by diacylglycerol. Nature, Vol. 397, 1999, pp. 259–263. [DOI] [PubMed] [Google Scholar]
- [49].Hong K, Cope EL, DeLalio LJ, Marziano C, Isakson BE, Sonkusare SK, TRPV4 (Transient Receptor Potential Vanilloid 4) Channel-Dependent Negative Feedback Mechanism Regulates Gq Protein-Coupled Receptor-Induced Vasoconstriction, Arterioscler Thromb Vasc Biol 38 (2018) 542–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Huang Y, Fliegert R, Guse AH, Lü W, Du J, A structural overview of the ion channels of the TRPM family. Cell Calcium, Vol. 85, Elsevier, 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Iadecola C, Davisson RL, Hypertension and cerebrovascular dysfunction, Cell Metab 7 (2008) 476–484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Iadecola C, Gottesman RF, Neurovascular and Cognitive Dysfunction in Hypertension, Circ Res 124 (2019) 1025–1044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Jia J, Verma S, Nakayama S, Quillinan N, Grafe MR, Hurn PD, Herson PS, Sex differences in neuroprotection provided by inhibition of TRPM2 channels following experimental stroke, J Cereb Blood Flow Metab 31 (2011) 2160–2168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Jiang Q, Gao Y, Wang C, Tao R, Wu Y, Zhan K, Liao M, Lu N, Lu Y, Wilcox CS, Luo J, Jiang LH, Yang W, Han F, Nitration of TRPM2 as a Molecular Switch Induces Autophagy during Brain Pericyte Injury. Antioxidants and Redox Signaling, Vol. 27, 2017, pp. 1297–1316. [DOI] [PubMed] [Google Scholar]
- [55].Johansson SE, Andersen XEDR, Hansen RH, Povlsen GK, Edvinsson L, Cerebrovascular endothelin-1 hyper-reactivity is associated with transient receptor potential canonical channels 1 and 6 activation and delayed cerebral hypoperfusion after forebrain ischaemia in rats. Acta Physiologica, Vol. 214, 2015, pp. 376–389. [DOI] [PubMed] [Google Scholar]
- [56].Kazama K, Anrather J, Zhou P, Girouard H, Frys K, Milner TA, Iadecola C, Angiotensin II impairs neurovascular coupling in neocortex through NADPH oxidase-derived radicals, Circ Res 95 (2004) 1019–1026. [DOI] [PubMed] [Google Scholar]
- [57].Khatibi NH, Lee LK, Zhou Y, Chen W, Rolland W, Fathali N, Martin R, Applegate R, Stier G, Zhang JH, Capsaicin Pre-treatment Provides Neurovascular Protection Against Neonatal Hypoxic-Ischemic Brain Injury in Rats. Acta Neurochirurgica, Vol. 111, 2011, pp. 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Kim KJ, Iddings JA, Stern JE, Blanco VM, Croom D, Kirov SA, Filosa JA, Astrocyte contributions to flow/pressure-evoked parenchymal arteriole vasoconstriction, J Neurosci 35 (2015) 8245–8257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Kisler K, Nelson AR, Montagne A, Zlokovic BV, Cerebral blood flow regulation and neurovascular dysfunction in Alzheimer disease. Nature Reviews Neuroscience, Vol. 18, Nature Publishing Group, 2017, pp. 419–434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Klauck TM, Faux MC, Labudda K, Langeberg LK, Jaken S, Scott JD, Coordination of three signaling enzymes by AKAP79, a mammalian scaffold protein, Science 271 (1996) 1589–1592. [DOI] [PubMed] [Google Scholar]
- [61].Kraft R, Harteneck C, The mammalian melastatin-related transient receptor potential cation channels: An overview. Pflugers Archiv European Journal of Physiology, Vol. 451, 2005, pp. 204–211. [DOI] [PubMed] [Google Scholar]
- [62].Kumar A, Kumari S, Majhi RK, Swain N, Yadav M, Goswami C, Regulation of TRP channels by steroids: Implications in physiology and diseases, Gen Comp Endocrinol 220 (2015) 23–32. [DOI] [PubMed] [Google Scholar]
- [63].Kunkler PE, Ballard CJ, Oxford GS, Hurley JH, TRPA1 receptors mediate environmental irritant-induced meningeal vasodilatation. Pain, Vol. 152, 2011, pp. 38–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Lange I, Yamamoto S, Partida-Sanchez S, Mori Y, Fleig A, Penner R, TRPM2 functions as a lysosomal Ca2+-release channel in beta cells, Sci Signal 2 (2009) ra23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Lehnardt S, Lehmann S, Kaul D, Tschimmel K, Hoffmann O, Cho S, Krueger C, Nitsch R, Meisel A, Weber JR, Toll-like receptor 2 mediates CNS injury in focal cerebral ischemia, J Neuroimmunol 190 (2007) 28–33. [DOI] [PubMed] [Google Scholar]
- [66].Li W, Ehrich M, Transient alterations of the blood-brain barrier tight junction and receptor potential channel gene expression by chlorpyrifos. Journal of Applied Toxicology, Vol. 33, 2013, pp. 1187–1191. [DOI] [PubMed] [Google Scholar]
- [67].Li Y, Baylie RL, Tavares MJ, Brayden JE, TRPM4 channels couple purinergic receptor mechanoactivation and myogenic tone development in cerebral parenchymal arterioles. Journal of Cerebral Blood Flow and Metabolism, Vol. 34, 2014, pp. 1706–1714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Liu L, Chen M, Lin K, Xiang X, Yang J, Zheng Y, Xiong X, Zhu S, TRPC6 Attenuates Cortical Astrocytic Apoptosis and Inflammation in Cerebral Ischemic/Reperfusion Injury, Front Cell Dev Biol 8 (2020) 594283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Liu X, Vien T, Duan J, Sheu SH, DeCaen PG, Clapham DE, Polycystin-2 is an essential ion channel subunit in the primary cilium of the renal collecting duct epithelium, Elife 7 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Longden TA, Dabertrand F, Koide M, Gonzales AL, Tykocki NR, Brayden JE, Hill-Eubanks D, Nelson MT, Capillary K(+)-sensing initiates retrograde hyperpolarization to increase local cerebral blood flow, Nat Neurosci 20 (2017) 717–726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Marics B, Peitl B, Varga A, Pazmandi K, Bacsi A, Nemeth J, Szilvassy Z, Jancso G, Dux M, Diet-induced obesity alters dural CGRP release and potentiates TRPA1-mediated trigeminovascular responses, Cephalalgia 37 (2017) 581–591. [DOI] [PubMed] [Google Scholar]
- [72].Marziano C, Hong K, Cope EL, Kotlikoff MI, Isakson BE, Sonkusare SK, Nitric Oxide-Dependent Feedback Loop Regulates Transient Receptor Potential Vanilloid 4 (TRPV4) Channel Cooperativity and Endothelial Function in Small Pulmonary Arteries, J Am Heart Assoc 6 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].McCray BA, Diehl E, Sullivan JM, Aisenberg WH, Zaccor NW, Lau AR, Rich DJ, Goretzki B, Hellmich UA, Lloyd TE, Sumner CJ, Neuropathy-causing TRPV4 mutations disrupt TRPV4-RhoA interactions and impair neurite extension, Nat Commun 12 (2021) 1444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Mercado J, Baylie R, Navedo MF, Yuan C, Scott JD, Nelson MT, Brayden JE, Santana LF, Local control of TRPV4 channels by AKAP150-targeted PKC in arterial smooth muscle, J Gen Physiol 143 (2014) 559–575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Meyer DJ Jr., Huxley VH, Capillary hydraulic conductivity is elevated by cGMP-dependent vasodilators, Circ Res 70 (1992) 382–391. [DOI] [PubMed] [Google Scholar]
- [76].Miyanohara J, Kakae M, Nagayasu K, Nakagawa T, Mori Y, Arai K, Shirakawa H, Kaneko S, TRPM2 Channel Aggravates CNS Inflammation and Cognitive Impairment via Activation of Microglia in Chronic Cerebral Hypoperfusion, J Neurosci 38 (2018) 3520–3533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Miyanohara J, Shirakawa H, Sanpei K, Nakagawa T, Kaneko S, A pathophysiological role of TRPV1 in ischemic injury after transient focal cerebral ischemia in mice, Biochem Biophys Res Commun 467 (2015) 478–483. [DOI] [PubMed] [Google Scholar]
- [78].Nag S, Venugopalan R, Stewart DJ, Increased caveolin-1 expression precedes decreased expression of occludin and claudin-5 during blood-brain barrier breakdown, Acta Neuropathol 114 (2007) 459–469. [DOI] [PubMed] [Google Scholar]
- [79].Nagata K, Duggan A, Kumar G, García-Añoveros J, Nociceptor and hair cell transducer properties of TRPA1, a channel for pain and hearing. Journal of Neuroscience, Vol. 25, 2005, pp. 4052–4061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [80].Narayanan D, Bulley S, Leo MD, Burris SK, Gabrick KS, Boop FA, Jaggar JH, Smooth muscle cell transient receptor potential polycystin-2 (TRPP2) channels contribute to the myogenic response in cerebral arteries. Journal of Physiology, Vol. 591, 2013, pp. 5031–5046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Nilius B, Prenen J, Tang J, Wang C, Owsianik G, Janssens A, Voets T, Zhu MX, Regulation of the Ca2+ sensitivity of the nonselective cation channel TRPM4. Journal of Biological Chemistry, Vol. 280, © 2005 ASBMB. Currently published by Elsevier Inc; originally published by American Society for Biochemistry and Molecular Biology., 2005, pp. 6423–6433. [DOI] [PubMed] [Google Scholar]
- [82].Ottolini M, Daneva Z, Chen YL, Cope EL, Kasetti RB, Zode GS, Sonkusare SK, Mechanisms underlying selective coupling of endothelial Ca2+ signals with eNOS vs. IK/SK channels in systemic and pulmonary arteries. Journal of Physiology, Vol. 598, 2020, pp. 3577–3596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Ottolini M, Hong K, Cope EL, Daneva Z, DeLalio LJ, Sokolowski JD, Marziano C, Nguyen NY, Altschmied J, Haendeler J, Johnstone SR, Kalani MY, Park MS, Patel RP, Liedtke W, Isakson BE, Sonkusare SK, Local Peroxynitrite Impairs Endothelial Transient Receptor Potential Vanilloid 4 Channels and Elevates Blood Pressure in Obesity, Circulation 141 (2020) 1318–1333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Ottolini M, Sonkusare SK, The Calcium Signaling Mechanisms in Arterial Smooth Muscle and Endothelial Cells. Comprehensive Physiology, Vol. 11, 2021, pp. 1831–1869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Park L, Wang G, Moore J, Girouard H, Zhou P, Anrather J, Iadecola C, The key role of transient receptor potential melastatin-2 channels in amyloid-β-induced neurovascular dysfunction. Nature Communications, Vol. 5, Nature Publishing Group, 2014, pp. 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Pegorini S, Braida D, Verzoni C, Guerini-Rocco C, Consalez GG, Croci L, Sala M, Capsaicin exhibits neuroprotective effects in a model of transient global cerebral ischemia in Mongolian gerbils, Br J Pharmacol 144 (2005) 727–735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Peixoto-Neves D, Soni H, Adebiyi A, CGRPergic Nerve TRPA1 Channels Contribute to Epigallocatechin Gallate-Induced Neurogenic Vasodilation. ACS Chemical Neuroscience, Vol. 10, 2019, pp. 216–220. [DOI] [PubMed] [Google Scholar]
- [88].Phan TX, Ton HT, Gulyás H, Pórszász R, Tóth A, Russo R, Kay MW, Sahibzada N, Ahern GP, TRPV1 expressed throughout the arterial circulation regulates vasoconstriction and blood pressure. Journal of Physiology, Vol. 598, 2020, pp. 5639–5659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Phelan KD, Shwe UT, Cozart MA, Wu H, Mock MM, Abramowitz J, Birnbaumer L, Zheng F, TRPC3 channels play a critical role in the theta component of pilocarpine-induced status epilepticus in mice. Epilepsia, Vol. 58, 2017, pp. 247–254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Pires PW, Earley S, Neuroprotective effects of trpa1 channels in the cerebral endothelium following ischemic stroke. eLife, Vol. 7, 2018, pp. 1–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Pires PW, Sullivan MN, Pritchard HAT, Robinson JJ, Earley S, Unitary TRPV3 channel ca2+ influx events elicit endothelium-dependent dilation of cerebral parenchymal arterioles. American Journal of Physiology - Heart and Circulatory Physiology, Vol. 309, 2015, pp. H2031–H2041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Poteser M, Graziani A, Rosker C, Eder P, Derler I, Kahr H, Zhu MX, Romanin C, Groschner K, TRPC3 and TRPC4 associate to form a redox-sensitive cation channel: Evidence for expression of native TRPC3-TRPC4 heteromeric channels in endothelial cells. Journal of Biological Chemistry, Vol. 281, © 2006 ASBMB. Currently published by Elsevier Inc; originally published by American Society for Biochemistry and Molecular Biology., 2006, pp. 13588–13595. [DOI] [PubMed] [Google Scholar]
- [93].Presa JL, Saravia F, Bagi Z, Filosa JA, Vasculo-Neuronal Coupling and Neurovascular Coupling at the Neurovascular Unit: Impact of Hypertension, Front Physiol 11 (2020) 584135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [94].Prikhodko V, Chernyuk D, Sysoev Y, Zernov N, Okovityi S, Popugaeva E, Potential Drug Candidates to Treat TRPC6 Channel Deficiencies in the Pathophysiology of Alzheimer’s Disease and Brain Ischemia. Cells, Vol. 9, 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Qian Q, Hunter LW, Li M, Marin-Padilla M, Prakash YS, Somlo S, Harris PC, Torres VE, Sieck GC, Pkd2 haploinsufficiency alters intracellular calcium regulation in vascular smooth muscle cells. Human Molecular Genetics, Vol. 12, 2003, pp. 1875–1880. [DOI] [PubMed] [Google Scholar]
- [96].Reading SA, Brayden JE, Central role of TRPM4 channels in cerebral blood flow regulation. Stroke, Vol. 38, 2007, pp. 2322–2328. [DOI] [PubMed] [Google Scholar]
- [97].Reading SA, Earley S, Waldron BJ, Welsh DG, Brayden JE, TRPC3 mediates pyrimidine receptor-induced depolarization of cerebral arteries. American Journal of Physiology - Heart and Circulatory Physiology, Vol. 288, 2005, pp. 2055–2061. [DOI] [PubMed] [Google Scholar]
- [98].Rivera-Pagan AF, Mendez-Gonzalez MP, Rivera-Aponte DE, Malpica-Nieves CJ, Melnik-Martinez KV, Zayas-Santiago A, Maldonado-Martinez G, Shuba YM, Skatchkov SN, Eaton MJ, A-Kinase-Anchoring Protein (AKAP150) is expressed in Astrocytes and Upregulated in Response to Ischemia, Neuroscience 384 (2018) 54–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Ross JM, Kim C, Allen D, Crouch EE, Narsinh K, Cooke DL, Abla AA, Nowakowski TJ, Winkler EA, The Expanding Cell Diversity of the Brain Vasculature, Front Physiol 11 (2020) 600767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Ryu HJ, Kim JE, Kim YJ, Kim JY, Kim WI, Choi SY, Kim MJ, Kang TC, Endothelial transient receptor potential conical channel (TRPC)-3 activation induces vasogenic edema formation in the rat piriform cortex following status epilepticus. Cellular and Molecular Neurobiology, Vol. 33, 2013, pp. 575–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Sano Y, Inamura K, Miyake A, Mochizuki S, Yokoi H, Matsushime H, Furuichi K, Immunocyte Ca2+ influx system mediated by LTRPC2. Science, Vol. 293, 2001, pp. 1327–1330. [DOI] [PubMed] [Google Scholar]
- [102].Santoni G, Morelli MB, Amantini C, Santoni M, Nabissi M, Marinelli O, Santoni A, “Immuno-Transient Receptor Potential Ion Channels”: The Role in Monocyte- and Macrophage-Mediated Inflammatory Responses, Front Immunol 9 (2018) 1273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Schaefer M, Plant TD, Obukhov AG, Hofmann T, Gudermann T, Schultz G, Receptor-mediated regulation of the nonselective cation channels TRPC4 and TRPC5. Journal of Biological Chemistry, Vol. 275, 2000, pp. 17517–17526. [DOI] [PubMed] [Google Scholar]
- [104].Scheffers MS, Le H, Van Der Bent P, Leonhard W, Prins F, Spruit L, Breuning MH, De Heer E, Peters DJM, Distinct subcellular expression of endogenous polycystin-2 in the plasma membrane and Golgi apparatus of MDCK cells. Human Molecular Genetics, Vol. 11, 2002, pp. 59–67. [DOI] [PubMed] [Google Scholar]
- [105].Segarra M, Aburto MR, Hefendehl J, Acker-Palmer A, Neurovascular interactions in the nervous system. Annual Review of Cell and Developmental Biology, Vol. 35, 2019, pp. 615–635. [DOI] [PubMed] [Google Scholar]
- [106].Sharif-Naeini R, Folgering JHA, Bichet D, Duprat F, Lauritzen I, Arhatte M, Jodar M, Dedman A, Chatelain FC, Schulte U, Retailleau K, Loufrani L, Patel A, Sachs F, Delmas P, Peters DJM, Honoré E, Polycystin-1 and −2 Dosage Regulates Pressure Sensing. Cell, Vol. 139, Elsevier Ltd, 2009, pp. 587–596. [DOI] [PubMed] [Google Scholar]
- [107].Sita G, Hrelia P, Graziosi A, Ravegnini G, Morroni F, TRPM2 in the Brain: Role in Health and Disease. Cells, Vol. 7, 2018, p. 82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [108].Sonkusare SK, Bonev AD, Ledoux J, Liedtke W, Kotlikoff MI, Heppner TJ, Hill-eubanks DC, Nelson MT, Elementary Ca2+ Signals Through Endothelial TRPV4 Channels Regulate Vascular Function. Science, Vol. 336, 2012, pp. 597–602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [109].Sonkusare SK, Dalsgaard T, Bonev AD, Nelson MT, Inward rectifier potassium (Kir2.1) channels as end-stage boosters of endothelium-dependent vasodilators, J Physiol 594 (2016) 3271–3285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Story GM, Peier AM, Reeve AJ, Eid SR, Mosbacher J, Hricik TR, Earley TJ, Hergarden AC, Andersson DA, Hwang SW, McIntyre P, Jegla T, Bevan S, Patapoutian A, ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell, Vol. 112, 2003, pp. 819–829. [DOI] [PubMed] [Google Scholar]
- [111].Straub SV, Bonev AD, Wilkerson MK, Nelson MT, Dynamic inositol trisphosphate-mediated calcium signals within astrocytic endfeet underlie vasodilation of cerebral arterioles, J Gen Physiol 128 (2006) 659–669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [112].Sullivan MN, Francis M, Pitts NL, Taylor MS, Earley S, Optical recording reveals novel properties of GSK1016790A-induced vanilloid transient receptor potential channel TRPV4 activity in primary human endothelial cells. Molecular Pharmacology, Vol. 82, 2012, pp. 464–472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [113].Sullivan MN, Gonzales AL, Pires PW, Bruhl A, Leo MD, Li W, Oulidi A, Boop FA, Feng Y, Jaggar JH, Welsh DG, Earley S, Vascular biology: Localized TRPA1 channel Ca2+ signals stimulated by reactive oxygen species promote cerebral artery dilation. Science Signaling, Vol. 8, 2015, pp. 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [114].Tajada S, Moreno CM, O’Dwyer S, Woods S, Sato D, Navedo MF, Santana LF, Distance constraints on activation of TRPV4 channels by AKAP150-bound PKCalpha in arterial myocytes, J Gen Physiol 149 (2017) 639–659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Tang SC, Arumugam TV, Xu X, Cheng A, Mughal MR, Jo DG, Lathia JD, Siler DA, Chigurupati S, Ouyang X, Magnus T, Camandola S, Mattson MP, Pivotal role for neuronal Toll-like receptors in ischemic brain injury and functional deficits, Proc Natl Acad Sci U S A 104 (2007) 13798–13803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [116].Thakore P, Alvarado MG, Ali S, Mughal A, Pires PW, Yamasaki E, Pritchard HAT, Isakson BE, Tran CHT, Earley S, Brain endothelial cell trpa1 channels initiate neurovascular coupling. eLife, Vol. 10, 2021, pp. 1–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [117].Thakore P, Earley S, Transient Receptor Potential Channels and Endothelial Cell Calcium Signaling, Compr Physiol 9 (2019) 1249–1277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [118].Thakore P, Pritchard HAT, Griffin CS, Yamasaki E, Drumm BT, Lane C, Sanders KM, Earley YF, Earley S, TRPML1 channels initiate Ca2+ sparks in vascular smooth muscle cells. Science Signaling, Vol. 13, 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [119].Tóth B, Iordanov I, Csanády L, Putative chanzyme activity of TRPM2 cation channel is unrelated to pore gating. Proceedings of the National Academy of Sciences of the United States of America, Vol. 111, 2014, pp. 16949–16954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [120].Turlova E, Feng ZP, Sun HS, The role of TRPM2 channels in neurons, glial cells and the blood-brain barrier in cerebral ischemia and hypoxia, Acta Pharmacol Sin 39 (2018) 713–721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [121].Vanlandewijck M, He L, Mae MA, Andrae J, Ando K, Del Gaudio F, Nahar K, Lebouvier T, Lavina B, Gouveia L, Sun Y, Raschperger E, Rasanen M, Zarb Y, Mochizuki N, Keller A, Lendahl U, Betsholtz C, A molecular atlas of cell types and zonation in the brain vasculature, Nature 554 (2018) 475–480. [DOI] [PubMed] [Google Scholar]
- [122].Vannier B, Peyton M, Boulay G, Brown D, Qin N, Jiang M, Zhu X, Birnbaumer L, Mouse trp2, the homologue of the human trpc2 pseudogene, encodes mTrp2, a store depletion-activated capacitative Ca2+ entry channel. Proceedings of the National Academy of Sciences of the United States of America, Vol. 96, 1999, pp. 2060–2064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [123].Wang LH, Luo M, Wang Y, Galligan JJ, Wang DH, Impaired vasodilation in response to perivascular nerve stimulation in mesenteric arteries of TRPV1-null mutant mice. Journal of Hypertension, Vol. 24, 2006, pp. 2399–2408. [DOI] [PubMed] [Google Scholar]
- [124].Wang Q, Liu N, Ni YS, Yang JM, Ma L, Lan XB, Wu J, Niu JG, Yu JQ, TRPM2 in ischemic stroke: Structure, molecular mechanisms, and drug intervention. Channels, Vol. 15, Taylor & Francis, 2021, pp. 136–154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [125].Wang W, Zhang X, Gao Q, Xu H, Trpml1: An ion channel in the lysosome. Handbook of Experimental Pharmacology, Vol. 222, 2014, pp. 631–645. [DOI] [PubMed] [Google Scholar]
- [126].Wellman GC, Nathan DJ, Saundry CM, Perez G, Bonev AD, Penar PL, Tranmer BI, Nelson MT, Ca2+ sparks and their function in human cerebral arteries, Stroke 33 (2002) 802–808. [DOI] [PubMed] [Google Scholar]
- [127].Welsh DG, Morielli AD, Nelson MT, Brayden JE, Transient receptor potential channels regulate myogenic tone of resistance arteries. Circulation Research, Vol. 90, 2002, pp. 248–250. [DOI] [PubMed] [Google Scholar]
- [128].Xi Q, Adebiyi A, Zhao G, Chapman KE, Waters CM, Hassid A, Jaggar JH, IP3 constricts cerebral arteries via IP3 receptor-mediated TRPC3 channel activation and independently of sarcoplasmic reticulum Ca2+ release. Circulation Research, Vol. 102, 2008, pp. 1118–1126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [129].Yanagitani Y, Rakugi H, Okamura A, Moriguchi K, Takiuchi S, Ohishi M, Suzuki K, Higaki J, Ogihara T, Angiotensin II type 1 receptor-mediated peroxide production in human macrophages, Hypertension 33 (1999) 335–339. [DOI] [PubMed] [Google Scholar]
- [130].Zaccor NW, Sumner CJ, Snyder SH, The nonselective cation channel TRPV4 inhibits angiotensin II receptors, J Biol Chem 295 (2020) 9986–9997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [131].Zeng C, Zhou P, Jiang T, Yuan C, Ma Y, Feng L, Liu R, Tang W, Long X, Xiao B, Tian F, Upregulation and Diverse Roles of TRPC3 and TRPC6 in Synaptic Reorganization of the Mossy Fiber Pathway in Temporal Lobe Epilepsy. Molecular Neurobiology, Vol. 52, 2015, pp. 562–572. [DOI] [PubMed] [Google Scholar]
- [132].Zhang L, Papadopoulos P, Hamel E, Endothelial TRPV4 channels mediate dilation of cerebral arteries: impairment and recovery in cerebrovascular pathologies related to Alzheimer’s disease, Br J Pharmacol 170 (2013) 661–670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [133].Zhao Q, Li J, Ko WH, Kwan YW, Jiang L, Sun L, Yao X, TRPM2 promotes autophagic degradation in vascular smooth muscle cells, Sci Rep 10 (2020) 20719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [134].Zhao Y, Kiss T, DelFavero J, Li L, Li X, Zheng L, Wang J, Jiang C, Shi J, Ungvari Z, Csiszar A, Zhang XA, CD82-TRPM7-Numb signaling mediates age-related cognitive impairment, Geroscience 42 (2020) 595–611. [DOI] [PMC free article] [PubMed] [Google Scholar]