

ABSTRACT
The orientation of the mitotic spindle determines the direction of cell division, and therefore contributes to tissue shape and cell fate. Interaction between the multifunctional scaffolding protein Discs large (Dlg) and the canonical spindle orienting factor GPSM2 (called Pins in Drosophila and LGN in vertebrates) has been established in bilaterian models, but its function remains unclear. We used a phylogenetic approach to test whether the interaction is obligate in animals, and in particular whether Pins/LGN/GPSM2 evolved in multicellular organisms as a Dlg-binding protein. We show that Dlg diverged in C. elegans and the syncytial sponge Opsacas minuta and propose that this divergence may correspond with differences in spindle orientation requirements between these organisms and the canonical pathways described in bilaterians. We also demonstrate that Pins/LGN/GPSM2 is present in basal animals, but the established Dlg-interaction site cannot be found in either Placozoa or Porifera. Our results suggest that the interaction between Pins/LGN/GPSM2 and Dlg appeared in Cnidaria, and we therefore speculate that it may have evolved to promote accurate division orientation in the nervous system. This work reveals the evolutionary history of the Pins/LGN/GPSM2-Dlg interaction and suggests new possibilities for its importance in spindle orientation during epithelial and neural tissue development.
KEY WORDS: Spindle orientation, Discs large, GPSM2, LGN, Pins
Summary: Discs large and GPSM2 interact to drive cell division orientation. In this article the authors investigate the evolutionary history of this interaction and speculate on its importance to development.
INTRODUCTION
Discs large (Dlg) is a multifunctional scaffolding protein (Noirot-Timothée and Noirot, 1980; Woods and Bryant, 1994; Woods et al., 1996). Like other MAGUK (membrane associated guanylate kinase) proteins, Dlg is comprised of three N-terminal PDZ domains, an SH3 domain, and a C-terminal GUK (Guanylate Kinase) domain that is catalytically inactive and binds phosphorylated proteins (Woods and Bryant, 1994; Woods et al., 1996; Zhu et al., 2011a, 2012). Though well-studied for its roles in epithelial cell polarity and synaptic function, Dlg has also been identified as a factor involved in orientation of the mitotic spindle (Bellaiche et al., 2001; Bergstralh et al., 2013; Chanet et al., 2017; Johnston et al., 2009; Nakajima et al., 2013; Saadaoui et al., 2014; Siegrist and Doe, 2005).
The canonical spindle orientation complex is comprised of at least three evolutionarily conserved proteins: Mud/NuMA (mushroom body defect in Drosophila, nuclear mitotic apparatus in vertebrates), GPSM2 (G-protein signaling modulator, also called Pins in Drosophila and LGN in vertebrates), and the microtubule motor protein dynein (Fig. 1). Mud/NuMA and dynein combine to produce a pulling force that reels the spindle into alignment by walking towards the minus end of astral microtubules. Pins/LGN/GPSM2 is thought to work as an adaptor; it binds to Mud/NuMA through its N-terminal TPRs (tetratricopeptide repeats) and to myristoylated Gαi through its C-terminal GoLoco domains, thereby anchoring the complex to the cell cortex (Fig. 1). This structure is conserved across taxa, though the number of TPR and GoLoco domains varies between and within phyla (Wavreil and Yajima, 2020). A notable exception is the nematode worm, in which two functional homologs to Pins/LGN/GPSM2, called GPR1 and 2, have a unique structure that lack GoLoco domains and may contain one or two TPRs, though this is debated (Nguyen-Ngoc et al., 2007; Siller and Doe, 2009; Willard et al., 2004).
Fig. 1.
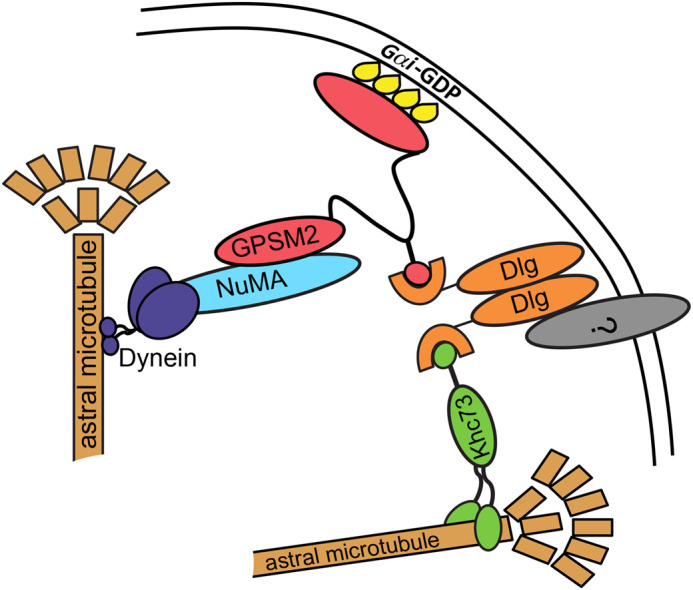
The canonical spindle orientation complex. The spindle orientation complex proteins Mud/NuMA (light blue), Pins/LGN/GPSM2 (fuchsia), and Dlg (orange). Khc73 (green), a potential spindle orientation factor, is also shown. This complex functions to capture microtubules and link the motor protein dynein (dark blue) to the cortex.
Extensive evidence links Dlg to spindle orientation through direct interaction with GPMS2, mediated by the Dlg GUK domain (Bellaiche et al., 2001; Johnston et al., 2012). Crystallization of Pins/LGN/GPSM2 and Dlg from Drosophila melanogaster and Mus musculus reveals that binding relies on phosphorylation of Pins/LGN/GPSM2 at a serine residue (S436 in Drosophila, S401 in vertebrates) in the unstructured linker between the TPR and GoLoco domains (Johnston et al., 2012; Woods et al., 1996; Zhu et al., 2011a). In agreement, expression of the non-phosphorylatable S401A variant in MDCK cell cysts or chick neuroepithelium causes spindle misorientation at metaphase (Hao et al., 2010; Saadaoui et al., 2014). Additionally, Drosophila follicle epithelial cells homozygous for dlg1P20, which encodes a premature truncation that disrupts the GUK domain, orient spindles randomly (Bergstralh et al., 2013).
Two models, which are not exclusive, have been proposed to explain the function of this interaction. The first is that Dlg functions as a positioning cue that recruits and/or restricts Pins/LGN/GPSM2 to distinct regions of the cell cortex. This mechanism has been described in Drosophila follicular and embryonic epithelial cells and in chick neuroepithelial cells (Bergstralh et al., 2013; Chanet et al., 2017; Saadaoui et al., 2014). Alternative positioning cues, including Inscuteable, E-Cadherin, and Afadin, have also been reported to influence Pins/LGN/GPSM2 positioning through direct interaction, and like Dlg, they may function in a cell-specific manner (Carminati et al., 2016; Gloerich et al., 2017; Yu et al., 2000). The second model is that Dlg acts to link the canonical complex to a microtubule motor protein called Khc73, thereby promoting microtubule capture (Siegrist and Doe, 2005; Zhu et al., 2016). This interaction also uses the GUK domain, which can bind the Khc73 MBS (MAGUK binding stalk) domain in a manner that relies on phosphorylation at a region outside the binding site (Zhu et al., 2016). A challenge faced in reconciling these two models is the problem of how Dlg could interact with both proteins in the same spindle orientation complex. However, in vitro studies suggest that Dlg exists as a dimer, in which case one monomer could bind to Pins/LGN/GPSM2 and the other to Khc73 (Fig. 1) (Marfatia et al., 2000).
In mammals, the phenotypic changes associated with spindle misorientation are predominantly observed in the nervous system, where they underlie diseases such as primary microcephaly, Chudley-McCullough syndrome, and Huntington's disease (Barnat et al., 2020; Godin and Humbert, 2011; Godin et al., 2010; Higgins et al., 2010; MacDonald et al., 1993; Noatynska et al., 2012). For example, exome sequencing and homozygosity mapping identified a mutation in human Pins/LGN/GPSM2 associated with the sensorineural hearing loss and brain malformations in Chudley-McCullough syndrome (Doherty et al., 2012). Additional experiments in mice demonstrated that this mutation causes profound deafness in otherwise phenotypically normal mice (Bhonker et al., 2016). Since the truncated Pins/LGN/GPSM2 mutant lacks GoLoco domains, the interaction between Pins/LGN/GPSM2 and Gαi is disrupted, but only appears to affect hearing and brain formation. Indeed, Gαi is essential for central auditory processing, inner hair cell synapse maturation, and hair bundle shape in mice (Beer-Hammer et al., 2018).
Neural defects due to spindle misorientation are also observed outside of vertebrates. In Drosophila, mud (Mud/NuMA) mutants are viable and demonstrate only a few defects: morphological changes in the mushroom bodies of the brain (Bowman et al., 2006; Prokop and Technau, 1994), slightly smaller wings, and female sterility (Zhou et al., 2019).
Variations in the requirements for spindle orientation between different tissues is well established. An important example is the presence of the protein Insc (Inscuteable) in neuroblasts but not in epithelia. In bilaterian models, Insc orients asymmetrical neuroblast division in the apical–basal axis by apically localizing Pins/LGN/GPSM2 (Yu et al., 2000). Unlike Dlg, which interacts with Pins/LGN/GPSM2 at the phosphorylated linker, Insc localizes Pins/LGN/GPSM2 by binding to the TPR domains (Yuzawa et al., 2011). Interestingly, the asymmetric localization of Pins/LGN/GPSM2 in neuroblast spindle orientation can be mediated through either Insc or Dlg, each with a distinct pathway (Siegrist and Doe, 2005).
While the importance of the interaction between Dlg and Pins/LGN/GPSM2 has been demonstrated in bilaterian models, whether these proteins and their interactions are conserved in more ancient animals remains unknown. If this interaction is conserved across all animal phyla, then the interaction between Pins/LGN/GPSM2 and Dlg may be essential to multicellularity. Alternatively, the interaction could have evolved as tissue types and architectures diversified within the animal kingdom. Therefore, determining when the Dlg and Pins/LGN/GPSM2 interaction arose may reveal the possible functions of the interaction in spindle orientation.
RESULTS
The GUK domain is conserved in basal animals
The Dlg GUK domain is found in animals and the single-celled eukaryotes choanoflagellates and Filasterea (Mendoza et al., 2010; te Velthuis et al., 2007). Our analysis indicates that the GUK domain is evolutionarily conserved; global alignment reveals sequence similarity at the Dlg C-terminus, and computationally predicted structures at this region resemble the experimentally determined structure (Fig. 2A,B). Conserved residues at the Dlg GUK domain, specifically at the GMP binding (BM) subdomain, which interact with the phosphorylated Pins/LGN/GPSM2 linker have been identified in Homo sapiens, M. musculus, Danio rerio, and Drosophila (Anderson et al., 2016; Johnston et al., 2011, 2012). Included in these residues is the proline residue responsible for the functional switch from GMP to phosphoprotein binding activity. We extended this analysis to other bilaterians and found that the GB subdomain is highly conserved. Similarity is also evident outside of bilaterian animals, with on average 15% greater identity locally than globally in Cnidaria, Placozoa, and Porifera, though we could not identify a Dlg homolog in Ctenophora (Fig. 2C). We also identified Dlg homologs in the choanoflagellates Salpingoeca rosetta and Monosiga brevicollis and show here that GB domains in these proteins share an average of 54% and 56% identity (respectively) with animals (Fig. 2C). Together, these results agree with previous work suggesting that the ability of Dlg to bind Pins/LGN/GPSM2 has been retained throughout its evolutionary history (Anderson et al., 2016). We also identified two notable exceptions: C. elegans and the poriferan O. minuta. For both organisms, we observed sequence divergence from other animals in the global and GB domain alignments (Fig. 2C, Fig. S1A,B).
Fig. 2.

Evolutionary conservation of Discs large. (A) Dlg sequences are aligned and colored based on conservation. The H. sapiens Dlg domain map is used to illustrate conserved domains. (B) Predicted structure (trRosetta) of Dlg GUK domain. (C) Dlg GB subdomain sequence alignment with organisms included in this study and choanoflagellates (M. brevicollis and S. rosetta). Asterisks indicate residues that are important for the interaction with phosphorylated Pins/LGN/GPSM2.
Syncytial sponges lack conservation at the GUK-GB subdomain
There are four classes of sponge within the Porifera phylum: Calcarea, Demospongia Hexactinellida, and Homoscleromorpha. Hexactinellida cell types are mainly syncytial, while Calcarea, Demospongia, and Homoscleromorpha contain entirely mononuclear cells. We found that the Dlg protein sequence has diverged in O. minuta, a member of Hexactinellida. Of the seven amino acids that interact with the Pins/LGN/GPSM2 linker, only three are conserved in O. minuta while five are conserved in Amphimedon queenslandica (Demospongiae) and all seven are conserved in Oscarella lobularis (Homoscleromorpha). Furthermore, the functional proline residue is conserved in A. queenslandica and O. lobularis but is replaced by a leucine in O. minuta (Fig. 2C) (Johnston et al., 2012). We considered the possibility that these differences could reflect low quality of the O. minuta genome annotation. To test for this, we examined the polarity proteins aPKC and Scribble. These sequences are equally conserved across all three sponges (not shown).
The Khc73 MBS domain is conserved across animals
Before investigating conservation of the Dlg-interacting region in Pins/LGN/GPSM2, we first asked to what extent sequence conservation should be expected. To address this question, we examined the Dlg-interacting region in Khc-73, a well-established binding partner for Dlg, with particular attention paid to sequences in animals outside of the Bilaterian clade (Fig. 3A). Khc73 is globally conserved across H. sapiens, the choanoflagellate S. rosetta, and Filasterea C. owczarzaki (Anderson et al., 2016) and locally conserved at a 14-3-3 binding motif across H. sapiens, X. laevis, D. rerio, and Drosophila (Lu and Prehoda, 2013). However, it is unknown whether the MAGUK-binding stalk is conserved within the animal kingdom. We found that the MBS domain sequence is highly conserved across phyla (Fig. 3B, Fig. S1C).
Fig. 3.

The Khc73 MBS domain is locally conserved in Porifera, Cnidaria, and Bilateria. (A) Khc73 sequences are aligned and colored based on conservation. The H. sapiens Khc domain map is used to illustrate conserved domains. (B) Sequence alignment of identified Khc73 MBS domains.
Identification of a candidate Pins/LGN/GPSM2 in Porifera
We next sought to identify Pins/LGN/GPSM2 orthologs in basal animals. BLAST searching against Ctenophora and Porifera genomes recovered no hits for Pins/LGN/GPSM2 in ctenophores and only one hit, a Pins/LGN/GPSM2-like protein [XP_019856151.1; NW_003546593.1 (14479...19908, complement)] in the sponge A. queenslandica. This sequence lacked an apparent linker region and GoLoco domains. Manual analysis of the Amphimedon queenslandica Annotation Release 102 genomic sequence revealed an additional annotated sequence with homology to the GoLoco domains [XP_003389018.1; NW_003546593.1 (9550…10710, complement)] located 3769 nucleotides past the C-terminus of XP_019856151.1. These sequences were combined to generate a candidate full length Pins/LGN/GPSM2 sequence (Fig. 4A, Fig. S1D).
Fig. 4.

Evolutionary conservation of Pins/LGN/GPSM2. (A) A reconstructed sequence for the Pins/LGN/GPSM2 ortholog in the poriferan A. queenslandica. (B) Identified Pins/LGN/GPSM2 sequences are aligned and colored based on conservation. The H. sapiens Pins/LGN/GPSM2 domain map is used to illustrate conserved domains. (C) Predicted structure (trRosetta) of Pins/LGN/GPSM2. Arrow indicates E. diaphana linker. (D) Sequence alignment at the Pins/LGN/GPSM2 linker region. (+) in consensus sequences indicate positions where two residues share the modal value. Asterisk indicates the phosphorylation site.
We considered the possibility that exons past the N-terminus of XP_019856151.1 or past the C-terminus of XP_003389018.1 were missed in the manual reconstruction. We therefore performed global sequence alignment comparing the Pins/LGN/GPSM2 sequence in A. queenslandica to all other organisms included in this study. Based on this analysis, missing exons are likely to be located past the N-terminus of XP_019856151.1, as all organisms except A. queenslandica contain an additional 20-80 amino acids at the N-terminus, corresponding to the first TPR domain in H. sapiens Pins/LGN/GPSM2. It is also possible that an exon is missing past the C-terminus of XP_003389018.1, although other organisms in this study only contain an additional 5-20 amino acids at the C-terminus, and these do not correspond with any functional domains.
Conservation of the Pins/LGN/GPSM2 linker region
A Pins/LGN/GPSM2 ortholog has been identified in choanoflagellates but lacks the necessary phosphorylation site to interact with Dlg. Because this site has been previously identified in multiple animal phyla, the question arises of whether it arose to facilitate multicellularity. A challenge to addressing this question is that the representative cnidarian sequence (Hydra vulgaris XM_004209432.1) used in the earlier alignment is no longer annotated as part of the H. vulgaris genome and has been removed from the NBCI GenBank (Anderson et al., 2016). We therefore examined Pins/LGN/GPSM2 sequences from the cnidarians A. tenebrosa, E. diaphana, N. vectensis, O. faveolate, P. damicornis, and S. pistillata and identified a highly conserved region with a consensus sequence of RKHSN++ADKRK (+ denotes positions where two residues share the modal value) (Fig. S2). The conserved serine residue is predicted to be a phosphorylation site with >99% confidence (Blom et al., 1999). While E. diaphana and N. vectensis contain a second phosphorylation site at a serine residue two amino acids away from the conserved phosphorylation site, the bilaterian linker contains a glutamic acid residue at this same location and therefore this double phosphorylation is unlikely to affect the interaction between Dlg and Pins/LGN/GPSM2 structurally or electrostatically. Together, these results are consistent with the possibility that interaction between Dlg and phosphorylated Pins/LGN/GPSM2 extends to cnidarians.
Previous work suggests that the mitotic kinase Aurora A phosphorylates Pins/LGN/GPSM2 and is required for spindle orientation in cultured cells with induced cortical polarity (Johnston et al., 2009). Aurora A phosphorylates proteins that contain an R/K/N-R-X-S/T-B motif, where B is any hydrophobic residue except proline (Ferrari et al., 2005). This motif is readily apparent in bilaterian Pins/LGN/GPSM2 proteins; the relevant serine residue is situated within a highly conserved consensus sequence (RRHSMENMELMKLTP) (Anderson et al., 2016; Johnston et al., 2009; Saadaoui et al., 2017). An exception is the nematode worm. An earlier study raised the question of whether an aspartic acid in the C. elegans linker takes the place of the phosphorylated serine, acting as a phosphomimetic that permits constitutive binding to Dlg (Johnston et al., 2009). Given the low sequence conservation of the GB subdomain of C. elegans Dlg, we suggest this possibility is unlikely.
The conserved phosphorylation target sequence in cnidarians does not match the bilaterian target. All identified cnidarian linker sequences contain either a charged or polar residue (R/K/N) at the B position (Fig. S2). The efficiency (Kcat/Km) of human Aurora A phosphorylation of synthetic peptides is reduced by 85% when the B position is occupied by an arginine versus a leucine (Ferrari et al., 2005). This observation suggests the possibility that Aurora A does not phosphorylate the Pins/LGN/GPSM2 linker in cnidarians, though it is also possible that cnidarian Aurora A has a distinct target sequence. We were unable to identify a conserved target sequence in other Aurora-A substrates, namely NEDD1, NDC80, P150-glued, MAP9, Astrin, and Haspin.
The Pins/LGN/GPSM2 linker region is not conserved in Placozoa and Porifera
The Pins/LGN/GPSM2 linker regions in Placozoa and Porifera do not readily align with bilaterian and cnidarian sequences (Fig. 4A-C). We considered whether alternate phosphorylation sites could mediate the interaction with Dlg.
T. adhaerens (Placozoan) Pins/LGN/GPSM2 contains two predicted serine and one predicted threonine phosphorylation site between amino acids 381-420 (Fig. S3A) (Blom et al., 1999). While a previous report suggested that the threonine is the conserved phosphorylation site, we consider this possibility unlikely. Firstly, the residue is a serine in cnidarian and bilaterian animals. Secondly, we do not find sequence similarity in the flanking region. Thirdly, the proximity of this site to other phosphorylated resides is expected to present a steric challenge to Dlg, which interacts with mono-phosphorylated Pins/LGN/GPSM2 in other organisms.
A. queenslandica (Poriferan) GPMS2 contains two predicted serine phosphorylation sites between amino acids 328-359, presenting the same challenge (Fig. S3B). Given similarity in length and sequence to other Pins/LGN/GPSM2 homologs, we expect that the entire length of the disordered linker region is included in our reconstruction (Fig. 4B). Nevertheless, we considered the possibility of potential missing exons between XP_019856151.1 and XP_003389018.1. To test for this, we reverse translated nucleotide sequences of regions with high exon character located between NW_003546593.1 (14479...19908, complement) and NW_003546593.1 (9550…10710, complement) in BLAST (Madeira et al., 2019). We identified and translated eight potential exons and determined whether these amino acid sequences contained phosphorylation sites and were similar to the bilaterian and cnidarian linkers. When determining whether these exons contained phosphorylation sites, we flanked the exon amino acid sequence with the last 15 amino acids from XP_019856151.1 and the first 15 amino acids from XP_003389018.1 to ensure that the properties of the surrounding amino acids were taken into account (Fig. S3C). Only one of these [NW_003546593.1 (12781…12966, complement)] contained phosphorylation sites (Fig. S3D), but the sequence was not similar to bilaterian or cnidarian linker sequences. Our analysis suggests that Dlg and Pins/LGN/GPSM2 do not interact in Placozoa or Porifera.
DISCUSSION
Dlg is necessary for spindle orientation across many bilaterian models. Here we provide evidence that the interaction between Dlg and Khc73 is present throughout the animal kingdom but that the interaction between Dlg and Pins/LGN/GPSM2 evolved in Cnidaria. While it remains unclear whether or not Dlg functions as a positioning cue or to assist in microtubule capture in bilaterian spindle orientation, our results suggest that neither function is required for multicellularity.
Dlg diverged in O. minuta
Hexactinellida is a class of sponges classified by the presence of the trabecular syncytium, a continuous multinucleated tissue that is the main component of the organism (Reiswig and Mehl, 1991). This class of animals is therefore an ideal system to investigate whether the canonical spindle orientation machinery is necessary for nuclear-only divisions. Of the three Poriferan GUK domain sequences identified and analyzed in this study, one species: O. minuta, a hexactinellid, showed an unusually low average percent identity. The same trend was observed in the global Dlg sequence. Given that the GB subdomain participates in many interactions important for spindle orientation, a likely explanation for this divergence is that spindle orientation is not required for the development of the trabecular syncytium. Therefore, it would not appear necessary to orient the plane of division in cells with only nuclear, and not cytoplasmic, divisions. Furthermore, how the spindle orientation complex, consisting of many cortical proteins, could interact with nuclei towards the center of the cytoplasmic mass is unclear.
Dlg and Pins/LGN/GPSM2 may have coevolved in C. elegans
The C. elegans Pins/LGN/GPSM2 orthologs, GPR1/2, demonstrate low sequence conservation with Pins/LGN/GPSM2 proteins despite maintaining its role in spindle orientation (Colombo et al., 2003; Srinivasan et al., 2003; Wavreil and Yajima, 2020). GPR1/2 share only an average of 23% identity with all other animals in our analyses. Similarly, C. elegans Dlg shares only an average of 43% identity with all other organisms. The observation that C. elegans Dlg and Pins/LGN/GPSM2 sequences share lower than expected percent identity with other bilaterians, and animals overall, is consistent with the possibility that Dlg and Pins/LGN/GPSM2 coevolved in C. elegans. The coevolution could be the cause or consequence of differences observed in C. elegans asymmetrical cell divisions (ACD) compared to other bilaterians. GPR1/2 and LIN-5 (Mud/NuMA) act downstream of cell polarity factors in C. elegans embryos (Colombo et al., 2003; Gotta et al., 2003; Srinivasan et al., 2003) whereas Drosophila Pins (Pins/LGN/GPSM2), Mud (Mud/NuMA), and polarity factors are mutually dependent on one another (Chia et al., 2008; Schober et al., 1999; Wavreil and Yajima, 2020; Yu et al., 2000). Furthermore, C. elegans use an evidently unique system that relies on the nematode-specific protein LET-99 to localize GPR1/2 (Krueger et al., 2010; Tsou et al., 2002). Together with previous work, our analysis suggests that spindle orientation in C. elegans may be best viewed as an unusual system for comparison, rather than a broadly applicable model.
Distinction between the Cnidarian and Bilaterian linkers
While site prediction software (NetPhos) predicts phosphorylation in the cnidarian linker with >99% confidence, the consensus sequence that we identified in these animals is markedly distinct from that in bilaterians. First, although human Aurora-A is unable to efficiently phosphorylate synthetic peptides with a charged residue at the B position, it is possible that Aurora-A can phosphorylate Pins/LGN/GPSM2 in vivo in cnidarians. Another possibility is that cnidarians use an alternative kinase to phosphorylate Pins/LGN/GPSM2. A strong candidate is aPKC (atypical Protein Kinase C), which is implicated in cell polarity and spindle orientation (Cui et al., 2007; Hao et al., 2010; Joberty et al., 2000; Zhu et al., 2011b).
Pins/LGN/GPSM2 may have interphase functions in Placozoa and Porifera
If the interaction between Pins/LGN/GPSM2 and Dlg arose in Cnidaria, what is the function of Pins/LGN/GPSM2 in Placozoa and Porifera? The poriferan and placozoan Pins/LGN/GPSM2 may still be able to orient cell divisions; the GoLoco and TPR domains are conserved, indicating that the interactions between Pins/LGN/GPSM2 and Mud/NuMA and Pins/LGN/GPSM2 and Gαi are likely conserved as well. While Pins/LGN/GPSM2 may interact with a more diverse group of proteins in higher ordered organisms, it is possible that localizing Mud/NuMA to the cortex is sufficient for Pins/LGN/GPSM2 to orient spindles in Porifera and Placozoa. In the Drosophila wing disc epithelium, spindle orientation is not disrupted by dlg1P20 mutants, which can still regulate apical-basal polarity factors but cannot interact with Pins due to a truncation in the GUK domain. Wing disc epithelium spindle orientation is also preserved in pins62 mutants, which are null (Bergstralh et al., 2016). Both observed effects are due to the presence of a Pins-independent mechanism of Mud localization (Bergstralh et al., 2016; Bosveld et al., 2016; David et al., 2005).
Another possibility is that poriferan and placozoan Pins/LGN/GPSM2 is involved in interphase cell behaviors. Though interphase Pins/LGN/GPSM2 functions are less understood, Pins/LGN/GPSM2 has been shown to play a role in endothelial interphase cells. While Pins/LGN/GPSM2 is dispensable for spindle orientation in this population, it is required for endothelial cell–cell adhesion, cell–matrix adhesion, and migration (Wright et al., 2015). The disruption of cell contacts and migration in Pins/LGN/GPSM2 knockdown endothelial cells is likely due to Pins/LGN/GPSM2 regulating microtubule dynamics and focal adhesion turnover, though the exact mechanisms have not been elucidated. This finding demonstrates that Pins/LGN/GPSM2 can have functions that are independent of spindle orientation. However, a final possibility is that Pins/LGN/GPSM2 could function to both localize Mud/NuMA in metaphase cells and regulate interphase behaviors in Placozoa and Porifera.
The interaction between Dlg and Pins/LGN/GPSM2 may be required for proper neural, but not epithelial, tissue development
Spindle orientation defects appear to have more severe consequences in nervous tissue versus epithelial tissue, as mutations in spindle orientation factors have been associated with brain abnormalities in otherwise phenotypically normal animals. Our results support the possibility that Dlg and Pins/LGN/GPSM2 interact in Cnidaria and Bilateria, but not it Porifera or Placozoa. Since the latter phyla do not have neural tissue, it is tempting to speculate that the interaction evolved to help drive neurodevelopment. However, we were unable to identify either protein in Ctenophora, which also have neurons.
Recent work in molecular phylogenetics place ctenophores as the sister group to all other animals (Moroz et al., 2014; Ryan et al., 2013). One possibility is that the common ancestor to all animal clades had neurons, but they were lost in Placozoa and Porifera. However, transcriptome analysis of ctenophores revealed that they lack, or express in non-neural tissue, the bilaterian neuronal pathway genes. These data support the idea that ctenophore nervous systems evolved independently from those in cnidarians and bilaterians (Moroz et al., 2014).
The mechanisms through which the nervous system evolved in Cnidaria and Bilateria are unresolved. A prominent hypothesis is that neurons evolved from epithelial tissue that gained the ability to sense external stimuli and transmit electrical and chemical signals to neighboring cells (Mackie, 1970). This theory is supported by the biological function of specialized cnidarian epithelial cells called nematocytes. Even in the absence of nerve cells, nematocytes can cause discharge of the nematocysts in response to chemical and mechanical stimuli (Aerne et al., 1991; Miljkovic-Licina et al., 2004). Additionally, similarities established between the epithelial septate junctions and neural synapse junctions suggest they are evolutionarily related (Banerjee et al., 2006; Harden et al., 2016) and recent work in the Drosophila follicular epithelium demonstrates that the epithelial cell reintegration and IgCAM-mediated axon growth occur through the same mechanism (Cammarota et al., 2020). The identification of IgCAMs in T. adhaerens, which contain epithelial cells but not neural, strongly suggests that the epithelial pathway was co-opted for axon growth and pathfinding when nervous tissue evolved from epithelia (Cammarota et al., 2020).
Our work may also provide insights into how epithelial cell processes may have been adopted in neurons. One possibility is that the tight regulation of ACD in the nervous system evolved from pre-existing ACD mechanisms. While the Insc-Pins/LGN/GPSM2 interaction is the main contributor to ACD in neuroblasts, Dlg has also been shown to regulate ACD in Drosophila sensory organ precursor (SOP) cells, which do not endogenously express Insc (Bellaiche et al., 2001). Additionally, Dlg is required for ACD of Drosophila neuroblasts in Insc null mutants (Siegrist and Doe, 2005). We therefore postulate that the interaction between Dlg and Pins/LGN/GPSM2 arose to regulate ACD as cell type and tissue diversity increased within the animal kingdom. Insc then co-opted this system of ACD regulation in neural tissues, where tight regulation of ACD is a crucial for normal animal development.
Several questions remain unresolved. Firstly, we were unable to identify Insc outside of the bilaterian clade, suggesting that Cnidaria do not express Insc. Does Dlg then regulate all neural ACD in this phylum? Additionally, what mechanisms regulate neural ACD in Ctenophora if their nervous system evolved independent from Cnidaria and Bilateria? The answers to these questions may further elucidate the functional consequences of interactions in the spindle orientation complex and how they relate to neurodevelopment.
The findings presented in this study indicate that the mechanisms of spindle orientation have undergone substantial evolution within the animal kingdom. While canonical spindle orientation proteins Pins/LGN/GPSM2 and Dlg can be identified in basal organisms, sequence alignments reveal that their interaction is likely not required for multicellularity. Rather, the interaction between Pins/LGN/GPSM2 and Dlg appears at the origin of neurons, suggesting it may have served as a template for asymmetrical cell division regulation in organisms with increasingly diverse and complex tissue development.
METHODS
Protein sequence identification and alignment
The non-redundant protein sequences (nr) database and PSI-BLAST Iteration 1 (Altschul et al., 1997) program were used for all NCBI BLAST searches (Sayers et al., 2020a,b). Reciprocal BLAST analysis was conducted to confirm identity. This approach confirmed that the identified sequence for O. minuta is not actually a different, closely related MAGUK family protein using O. minuta Dlg as the query sequence against the H. sapiens genome resulted in only hits against Dlg isoforms with E-values <10−10.
Using H. sapiens Pins/LGN/GPSM2 (LGN; NP_001307967.1) and Dlg (NP_001353143.1) as the query sequence, we performed a BLAST search against common bilaterian model organisms (M. musculus, X. laevis, D. rerio, S. purpuratus, D. melanogaster, C. gigas). Since spindle orientation factors have not been studied in basal animals, the entire Cnidarian (taxid:6073), Placazoan (taxid:10226), Poriferan (taxid:6040) and Ctenophoran (taxid: 10197) phyla were searched for homologous sequences. With the exception of C. elegans, Pins/LGN/GPSM2 sequences were identified in the bilaterian models as well as N. vectensis, A. tenebrosa, E. diaphana, and T. adhaerens each with >30% identity to H. sapiens Pins/LGN/GPSM2 and an E-value <10−10. Similarly, Dlg sequences were identified in the bilaterian models, N. vectensis, A. tenebrosa, E. diaphana, T. adhaerens, O. lobularis, and O. minuta each with >30% identity to H. sapiens Dlg and an E-value of<10−10 (Fig. S1). These results indicate that the identified sequences share significant similarity with human Pins/LGN/GPSM2 (LGN) and therefore are homologs of human Pins/LGN/GPSM2 (Pearson, 2013). While Pins/LGN/GPSM2 was identified in A. queenslandica, only a partial Dlg sequence, containing the GUK domain, was identified in A. queenslandica (XP_011408781.2) and therefore this organism was not included in the Dlg alignment.
Using H. sapiens Khc73 (KIF13B) as the query sequence (Q9NQT8), we identified potential orthologs in all organisms except for S. purpuratus and T. adhaerens. Reciprocal BLAST searching confirmed that identified sequences are the best hit Khc73 homologs in M. musculus, X. laevis, and D. rerio. However, the reciprocal BLAST for all other identified sequences indicated the best hit as KIF13A, a kinesin-motor protein that is closely related to Khc73. To confirm whether or not identified sequences are orthologs to KIF13A or Khc73, we created and analyzed the conservation domain map. While Khc73 contains an N-terminal Cap-Gly domain, KIF13A does not. Therefore, conservation at this region can be used to distinguish between the two proteins. The conservation domain map revealed a highly conserved region at the N-terminus of identified sequences, which we confirmed to be conserved at the H. sapiens Cap-Gly domain by analyzing the corresponding amino acid sequences. However, the identified sequence for C. elegans was truncated at the N-terminus and did not contain the Cap-Gly region, indicating it either is missing an exon at the N-terminus or is not the ortholog of Khc73. Therefore, C. elegans, along with T. adhaerens and S. purpuratus, were excluded from our analysis.
Alignments were created using COBALT (constraint-based alignment tool) (Papadopoulos and Agarwala, 2007) and were confirmed using T-coffee (Notredame et al., 2000) and ClustalX (Jeanmougin et al., 1998). Global conservation maps were created from the Multiple Sequence Alignment Viewer application where conservation was determined by relative entropy threshold of the aligned residues. Alignments from COBALT were downloaded as FASTA files and then were viewed in Jalview 2 (Waterhouse et al., 2009) where local sequence alignment figures were created.
Percent identity for matrices was calculated using EMBL-EBI Simple Phylogeny (Madeira et al., 2019). Nucleotide sequences from A. queenslandica exons were translated into amino acid sequences using EMBOSS EMBL-EBI Transeq (Madeira et al., 2019) with reverse set to true.
Phosphorylation site prediction
Phosphorylation sites at predicted Pins/LGN/GPSM2 linker sequences were determined using NetPhos 3.1 (Blom et al., 1999). Serine, threonine, and tyrosine residues were considered, and graphics were modeled after the NetPhos 3.1 Server output. Only phosphorylation sites predicted with>90% confidence were considered to be likely phosphorylation sites in this study.
Protein structure prediction
We used the trRosetta (transformed-restrained Rosetta) algorithm to predict the structure of identified Pins/LGN/GPSM2 and Dlg sequences (Hiranuma et al., 2021; Yang et al., 2020). Identified amino acids sequences were uploaded to the Robetta server where the trRosetta algorithm was used to create a PDB file with predicted protein structures (Hiranuma et al., 2021; Yang et al., 2020). Angstrom error graphs were also obtained through the Robetta server. The PyMOL Molecular Graphics System, Version 2.4.1 was used to create 3D visualizations of protein structures. Structures were imported to PyMOL and aligned using the alignment plugin.
Predicted Pins/LGN/GPSM2 structures have conserved secondary structures at previously defined domains. Each TPR consisted of a helix-turn-helix motif while each GoLoco domain contains an alpha helix. These results provide further evidence that the sequences presented in this study are orthologous. However, due to high angstrom error estimates within unstructured regions of these proteins, including the Pins/LGN/GPSM2 linker region, these structures could not be used to predict interactions between Pins/LGN/GPSM2 and Dlg.
Supplementary Material
Acknowledgements
We are grateful to J. David Lambert, Michael Welte, and members of the Bergstralh lab for their questions and comments.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
E.A.S. designed and performed the analyses, and E.A.S. wrote the manuscript. D.T.B. conceived and guided the project.
Funding
This work was supported by an NSF CAREER award to D.T.B. and NIH grant R01GM125839.
References
- Aerne, B. L., Stidwill, R. P. and Tardent, P. (1991). Nematocyst discharge in Hydra does not require the presence of nerve cells. J. Exp. Zool. 258, 137-141. 10.1002/jez.1402580115 [DOI] [Google Scholar]
- Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W. and Lipman, D. J. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson, D. P., Whitney, D. S., Hanson-Smith, V., Woznica, A., Campodonico-Burnett, W., Volkman, B. F., King, N., Prehoda, K. E. and Thornton, J. W. (2016). Evolution of an ancient protein function involved in organized multicellularity in animals. Elife 5, e10147. 10.7554/eLife.10147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banerjee, S., Sousa, A. D. and Bhat, M. A. (2006). Organization and function of septate junctions. Cell Biochem. Biophys. 46, 65-77. 10.1385/CBB:46:1:65 [DOI] [PubMed] [Google Scholar]
- Barnat, M., Capizzi, M., Aparicio, E., Boluda, S., Wennagel, D., Kacher, R., Kassem, R., Lenoir, S., Agasse, F., Braz, B. Y.et al. (2020). Huntington's disease alters human neurodevelopment. Science 369, 787-793. 10.1126/science.aax3338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beer-Hammer, S., Lee, S. C., Mauriac, S. A., Leiss, V., Groh, I. A. M., Novakovic, A., Piekorz, R. P., Bucher, K., Chen, C., Ni, K.et al. (2018). Gαi Proteins are Indispensable for Hearing. Cell. Physiol. Biochem. 47, 1509-1532. 10.1159/000490867 [DOI] [PubMed] [Google Scholar]
- Bellaiche, Y., Radovic, A., Woods, D. F., Hough, C. D., Parmentier, M.-L., O'Kane, C. J., Bryant, P. J. and Schweisguth, F. (2001). The partner of inscuteable/discs-large complex is required to establish planar polarity during asymmetric cell division in Drosophila. Cell 106, 355–366. 10.1016/S0092-8674(01)00444-5 [DOI] [PubMed] [Google Scholar]
- Bergstralh, D. T., Lovegrove, H. E. and St Johnston, D. (2013). Discs large links spindle orientation to apical-basal polarity in Drosophila epithelia. Curr. Biol. 23, 1707-1712. 10.1016/j.cub.2013.07.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergstralh, D. T., Lovegrove, H. E., Kujawiak, I., Dawney, N. S., Zhu, J., Cooper, S., Zhang, R. and Johnston, D. S. (2016). Pins is not required for spindle orientation in the Drosophila wing disc. Development 143, 2573-2581. 10.1242/dev.135475 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhonker, Y., Abu-Rayyan, A., Ushakov, K., Amir-Zilberstein, L., Shivatzki, S., Yizhar-Barnea, O., Elkan-Miller, T., Tayeb-Fligelman, E., Kim, S. M., Landau, M.et al. (2016). The GPSM2/LGN GoLoco motifs are essential for hearing. Mamm. Genome 27, 29-46. 10.1007/s00335-015-9614-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blom, N., Gammeltoft, S. and Brunak, S. (1999). Sequence and structure-based prediction of eukaryotic protein phosphorylation sites1 1Edited by F. E. Cohen. J. Mol. Biol. 294, 1351-1362. 10.1006/jmbi.1999.3310 [DOI] [PubMed] [Google Scholar]
- Bosveld, F., Markova, O., Guirao, B., Martin, C., Wang, Z., Pierre, A., Balakireva, M., Gaugue, I., Ainslie, A., Christophorou, N.et al. (2016). Epithelial tricellular junctions act as interphase cell shape sensors to orient mitosis. Nature 530, 495-498. 10.1038/nature16970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowman, S. K., Neumüller, R. A., Novatchkova, M., Du, Q. and Knoblich, J. A. (2006). The Drosophila NuMA homolog mud regulates spindle orientation in asymmetric cell division. Dev. Cell 10, 731-742. 10.1016/j.devcel.2006.05.005 [DOI] [PubMed] [Google Scholar]
- Cammarota, C., Finegan, T. M., Wilson, T. J., Yang, S. and Bergstralh, D. T. (2020). An axon-pathfinding mechanism preserves epithelial tissue integrity. Curr. Biol. 30, 5049-5057.e3. 10.1016/j.cub.2020.09.061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carminati, M., Gallini, S., Pirovano, L., Alfieri, A., Bisi, S. and Mapelli, M. (2016). Concomitant binding of Afadin to LGN and F-actin directs planar spindle orientation. Nat. Struct. Mol. Biol. 23, 155-163. 10.1038/nsmb.3152 [DOI] [PubMed] [Google Scholar]
- Chanet, S., Sharan, R., Khan, Z. and Martin, A. C. (2017). Myosin 2-induced mitotic rounding enables columnar epithelial cells to interpret cortical spindle positioning cues. Curr. Biol. 27, 3350-3358.e3. 10.1016/j.cub.2017.09.039 [DOI] [PubMed] [Google Scholar]
- Chia, W., Somers, W. G. and Wang, H. (2008). Drosophila neuroblast asymmetric divisions: cell cycle regulators, asymmetric protein localization, and tumorigenesis. J. Cell Biol. 180, 267-272. 10.1083/jcb.200708159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colombo, K., Grill, S. W., Kimple, R. J., Willard, F. S., Siderovski, D. P. and Gönczy, P. (2003). Translation of polarity cues into asymmetric spindle positioning in caenorhabditis elegans embryos. Science 300, 1957-1961. 10.1126/science.1084146 [DOI] [PubMed] [Google Scholar]
- Cui, S., Otten, C., Rohr, S., Abdelilah-Seyfried, S. and Link, B. A. (2007). Analysis of aPKCλ and aPKCζ reveals multiple and redundant functions during vertebrate retinogenesis. Mol. Cell. Neurosci. 34, 431-444. 10.1016/j.mcn.2006.11.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- David, N. B., Martin, C. A., Segalen, M., Rosenfeld, F., Schweisguth, F. and Bellaiche, Y. (2005). Drosophila Ric-8 regulates Gαi cortical localization to promote Gαi-dependent planar orientation of the mitotic spindle during asymmetric cell division. Nat. Cell Biol. 7, 1083-1090. 10.1038/ncb1319 [DOI] [PubMed] [Google Scholar]
- Doherty, D., Chudley, A. E., Coghlan, G., Ishak, G. E., Innes, A. M., Lemire, E. G., Rogers, R. C., Mhanni, A. A., Phelps, I. G., Jones, S. J. M.et al. (2012). GPSM2 mutations cause the brain malformations and hearing loss in Chudley-McCullough syndrome. Am J Hum Genetics 91, 209. 10.1016/j.ajhg.2012.06.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrari, S., Marin, O., Pagano, M. A., Meggio, F., Hess, D., El-Shemerly, M., Krystyniak, A. and Pinna, L. A. (2005). Aurora-A site specificity: a study with synthetic peptide substrates. Biochem. J. 390, 293-302. 10.1042/BJ20050343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gloerich, M., Bianchini, J. M., Siemers, K. A., Cohen, D. J. and Nelson, W. J. (2017). Cell division orientation is coupled to cell–cell adhesion by the E-cadherin/LGN complex. Nat. Commun. 8, 13996. 10.1038/ncomms13996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godin, J. D. and Humbert, S. (2011). Mitotic spindle: Focus on the function of huntingtin. Int. J. Biochem. Cell Biol. 43, 852-856. 10.1016/j.biocel.2011.03.009 [DOI] [PubMed] [Google Scholar]
- Godin, J. D., Colombo, K., Molina-Calavita, M., Keryer, G., Zala, D., Charrin, B. C., Dietrich, P., Volvert, M.-L., Guillemot, F., Dragatsis, I.et al. (2010). Huntingtin is required for mitotic spindle orientation and mammalian neurogenesis. Neuron 67, 392-406. 10.1016/j.neuron.2010.06.027 [DOI] [PubMed] [Google Scholar]
- Gotta, M., Dong, Y., Peterson, Y. K., Lanier, S. M. and Ahringer, J. (2003). Asymmetrically distributed C. elegans homologs of AGS3/PINS control spindle position in the early embryo. Curr. Biol. 13, 1029-1037. 10.1016/S0960-9822(03)00371-3 [DOI] [PubMed] [Google Scholar]
- Hao, Y., Du, Q., Chen, X., Zheng, Z., Balsbaugh, J. L., Maitra, S., Shabanowitz, J., Hunt, D. F. and Macara, I. G. (2010). Par3 controls epithelial spindle orientation by aPKC-mediated phosphorylation of apical pins. Curr. Biol. 20, 1809-1818. 10.1016/j.cub.2010.09.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harden, N., Wang, S. J. H. and Krieger, C. (2016). Making the connection – shared molecular machinery and evolutionary links underlie the formation and plasticity of occluding junctions and synapses. J. Cell Sci. 129, 3067-3076. 10.1242/jcs.186627 [DOI] [PubMed] [Google Scholar]
- Higgins, J., Midgley, C., Bergh, A.-M., Bell, S. M., Askham, J. M., Roberts, E., Binns, R. K., Sharif, S. M., Bennett, C., Glover, D. M.et al. (2010). Human ASPM participates in spindle organisation, spindle orientation and cytokinesis. BMC Cell Biol. 11, 85. 10.1186/1471-2121-11-85 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiranuma, N., Park, H., Baek, M., Anishchenko, I., Dauparas, J. and Baker, D. (2021). Improved protein structure refinement guided by deep learning-based accuracy estimation. Nat. Commun. 12, 1340. 10.1038/s41467-021-21511-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeanmougin, F., Thompson, J. D., Gouy, M., Higgins, D. G. and Gibson, T. J. (1998). Multiple sequence alignment with Clustal X. Trends Biochem. Sci. 23, 403-405. 10.1016/S0968-0004(98)01285-7 [DOI] [PubMed] [Google Scholar]
- Joberty, G., Petersen, C., Gao, L. and Macara, I. G. (2000). The cell-polarity protein Par6 links Par3 and atypical protein kinase C to Cdc42. Nat. Cell Biol. 2, 531-539. 10.1038/35019573 [DOI] [PubMed] [Google Scholar]
- Johnston, C. A., Hirono, K., Prehoda, K. E. and Doe, C. Q. (2009). Identification of an aurora-A/PinsLINKER/ Dlg spindle orientation pathway using induced cell polarity in S2 cells. Cell 138, 1150-1163. 10.1016/j.cell.2009.07.041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnston, C. A., Whitney, D. S., Volkman, B. F., Doe, C. Q. and Prehoda, K. E. (2011). Conversion of the enzyme guanylate kinase into a mitotic-spindle orienting protein by a single mutation that inhibits GMP-induced closing. Proc National Acad Sci 108, E973-E978. 10.1073/pnas.1104365108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnston, C. A., Doe, C. Q. and Prehoda, K. E. (2012). Structure of an enzyme-derived phosphoprotein recognition domain. Plos One 7, e36014. 10.1371/journal.pone.0036014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krueger, L. E., Wu, J.-C., Tsou, M.-F. B. and Rose, L. S. (2010). LET-99 inhibits lateral posterior pulling forces during asymmetric spindle elongation in C. elegans embryos. J Cell Biol. 189, 481-495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu, M. S. and Prehoda, K. E. (2013). A NudE/14-3-3 pathway coordinates dynein and the kinesin Khc73 to position the mitotic spindle. Dev. Cell 26, 369-380. 10.1016/j.devcel.2013.07.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacDonald, M. E., Ambrose, C. M., Duyao, M. P., Myers, R. H., Lin, C., Srinidhi, L., Barnes, G., Taylor, S. A., James, M., Groot, N.et al. (1993). A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. Cell 72, 971-983. 10.1016/0092-8674(93)90585-E [DOI] [PubMed] [Google Scholar]
- Mackie, G. O. (1970). Neuroid conduction and the evolution of conducting tissues. Q. Rev. Biol. 45, 319-332. 10.1086/406645 [DOI] [PubMed] [Google Scholar]
- Madeira, F., Park, Y. M., Lee, J., Buso, N., Gur, T., Madhusoodanan, N., Basutkar, P., Tivey, A. R. N., Potter, S. C., Finn, R. D.et al. (2019). The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 47, W636-W641. 10.1093/nar/gkz268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marfatia, S. M., Byron, O., Campbell, G., Liu, S.-C. and Chishti, A. H. (2000). Human homologue of the drosophila discs large tumor suppressor protein forms an oligomer in solution identification of the self-association site*. J. Biol. Chem. 275, 13759-13770. 10.1074/jbc.275.18.13759 [DOI] [PubMed] [Google Scholar]
- de Mendoza, A., Suga, H. and Ruiz-Trillo, I. (2010). Evolution of the MAGUK protein gene family in premetazoan lineages. BMC Evol. Biol. 10, 93. 10.1186/1471-2148-10-93 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miljkovic-Licina, M., Gauchat, D. and Galliot, B. (2004). Neuronal evolution: analysis of regulatory genes in a first-evolved nervous system, the hydra nervous system. Biosystems 76, 75-87. 10.1016/j.biosystems.2004.05.030 [DOI] [PubMed] [Google Scholar]
- Moroz, L. L., Kocot, K. M., Citarella, M. R., Dosung, S., Norekian, T. P., Povolotskaya, I. S., Grigorenko, A. P., Dailey, C., Berezikov, E., Buckley, K. M.et al. (2014). The ctenophore genome and the evolutionary origins of neural systems. Nature 510, 109-114. 10.1038/nature13400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakajima, Y., Meyer, E. J., Kroesen, A., McKinney, S. A. and Gibson, M. C. (2013). Epithelial junctions maintain tissue architecture by directing planar spindle orientation. Nature 500, 359-362. 10.1038/nature12335 [DOI] [PubMed] [Google Scholar]
- Nguyen-Ngoc, T., Afshar, K. and Gönczy, P. (2007). Coupling of cortical dynein and Gα proteins mediates spindle positioning in Caenorhabditis elegans. Nat. Cell Biol. 9, 1294-1302. 10.1038/ncb1649 [DOI] [PubMed] [Google Scholar]
- Noatynska, A., Gotta, M. and Meraldi, P. (2012). Mitotic spindle (DIS)orientation and DISease: Cause or consequence? J. Cell Biol. 199, 1025-1035. 10.1083/jcb.201209015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noirot-Timothée, C. and Noirot, C. (1980). Septate and scalariform junctions in arthropods. Int. Rev. Cytol. 63, 97-140. 10.1016/S0074-7696(08)61758-1 [DOI] [PubMed] [Google Scholar]
- Notredame, C., Higgins, D. G. and Heringa, J. (2000). T-coffee: a novel method for fast and accurate multiple sequence alignment11Edited by J. Thornton. J. Mol. Biol. 302, 205-217. 10.1006/jmbi.2000.4042 [DOI] [PubMed] [Google Scholar]
- Papadopoulos, J. S. and Agarwala, R. (2007). COBALT: constraint-based alignment tool for multiple protein sequences. Bioinformatics 23, 1073-1079. 10.1093/bioinformatics/btm076 [DOI] [PubMed] [Google Scholar]
- Pearson, W. R. (2013). An introduction to sequence similarity (“homology”) searching. Curr. Protoc. Bioinform. 42, 3.1.1- 3.1.8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prokop, A. and Technau, G. M. (1994). Normal function of the mushroom body defect gene of Drosophila is required for the regulation of the number and proliferation of neuroblasts. Dev. Biol. 161, 321-337. 10.1006/dbio.1994.1034 [DOI] [PubMed] [Google Scholar]
- Reiswig, H. M. and Mehl, D. (1991). Tissue organization of Farrea occa (Porifera, Hexactinellida). Zoomorphology 110, 301-311. 10.1007/BF01668020 [DOI] [Google Scholar]
- Ryan, J. F., Pang, K., Schnitzler, C. E., Nguyen, A.-D., Moreland, R. T., Simmons, D. K., Koch, B. J., Francis, W. R., Havlak, P., Program, N. C. S.et al. (2013). The genome of the ctenophore Mnemiopsis leidyi and its implications for cell type evolution. Science 342, 1242592. 10.1126/science.1242592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saadaoui, M., Machicoane, M., di Pietro, F., Etoc, F., Echard, A. and Morin, X. (2014). Dlg1 controls planar spindle orientation in the neuroepithelium through direct interaction with LGN. J. Cell Biol. 206, 707-717. 10.1083/jcb.201405060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saadaoui, M., Konno, D., Loulier, K., Goiame, R., Jadhav, V., Mapelli, M., Matsuzaki, F. and Morin, X. (2017). Loss of the canonical spindle orientation function in the Pins/LGN homolog AGS3. EMBO Rep. 18, 1509-1520. 10.15252/embr.201643048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sayers, E. W., Beck, J., Bolton, E. E., Bourexis, D., Brister, J. R., Canese, K., Comeau, D. C., Funk, K., Kim, S., Klimke, W.et al. (2020a). Database resources of the national center for biotechnology information. Nucleic Acids Res. 49, gkaa892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sayers, E. W., Cavanaugh, M., Clark, K., Pruitt, K. D., Schoch, C. L., Sherry, S. T. and Karsch-Mizrachi, I. (2020b). GenBank. Nucleic Acids Res. 49, gkaa1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schober, M., Schaefer, M. and Knoblich, J. A. (1999). Bazooka recruits Inscuteable to orient asymmetric cell divisions in Drosophila neuroblasts. Nature 402, 548-551. 10.1038/990135 [DOI] [PubMed] [Google Scholar]
- Siegrist, S. E. and Doe, C. Q. (2005). Microtubule-induced pins/gαi cortical polarity in Drosophila neuroblasts. Cell 123, 1323-1335. 10.1016/j.cell.2005.09.043 [DOI] [PubMed] [Google Scholar]
- Siller, K. H. and Doe, C. Q. (2009). Spindle orientation during asymmetric cell division. Nat. Cell Biol. 11, 365-374. 10.1038/ncb0409-365 [DOI] [PubMed] [Google Scholar]
- Srinivasan, D. G., Fisk, R. M., Xu, H. and van den Heuvel, S. (2003). A complex of LIN-5 and GPR proteins regulates G protein signaling and spindle function in C. elegans. Gene Dev 17, 1225-1239. 10.1101/gad.1081203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- te Velthuis, A. J., Admiraal, J. F. and Bagowski, C. P. (2007). Molecular evolution of the MAGUK family in metazoan genomes. BMC Evol. Biol. 7, 129. 10.1186/1471-2148-7-129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsou, M.-F. B., Hayashi, A., DeBella, L. R., McGrath, G. and Rose, L. S. (2002). LET-99 determines spindle position and is asymmetrically enriched in response to PAR polarity cues in C. elegans embryos. Development 129, 4469-4481. [DOI] [PubMed] [Google Scholar]
- Waterhouse, A. M., Procter, J. B., Martin, D. M. A., Clamp, M. and Barton, G. J. (2009). Jalview Version 2—a multiple sequence alignment editor and analysis workbench. Bioinformatics 25, 1189-1191. 10.1093/bioinformatics/btp033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wavreil, F. D. M. and Yajima, M. (2020). Diversity of activator of G-protein signaling (AGS)-family proteins and their impact on asymmetric cell division across taxa. Dev. Biol. 465, 89-99. 10.1016/j.ydbio.2020.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willard, F. S., Kimple, R. J. and Siderovski, D. P. (2004). Return of the GDI: the GoLoco motif in cell division. Annu. Rev. Biochem. 73, 925-951. 10.1146/annurev.biochem.73.011303.073756 [DOI] [PubMed] [Google Scholar]
- Woods, D. F. and Bryant, P. J. (1994). Tumor suppressor genes and signal transduction in Drosophila. Princess Takamatsu S 24, 1-13. [PubMed] [Google Scholar]
- Woods, D. F., Hough, C., Peel, D., Callaini, G. and Bryant, P. J. (1996). Dlg protein is required for junction structure, cell polarity, and proliferation control in Drosophila epithelia. J Cell Biology 134, 1469-1482. 10.1083/jcb.134.6.1469 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright, C. E., Kushner, E. J., Du, Q. and Bautch, V. L. (2015). LGN directs interphase endothelial cell behavior via the microtubule network. PLoS ONE 10, e0138763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, J., Anishchenko, I., Park, H., Peng, Z., Ovchinnikov, S. and Baker, D. (2020). Improved protein structure prediction using predicted interresidue orientations. Proc. Natl. Acad. Sci. USA 117, 1496-1503. 10.1073/pnas.1914677117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, F., Morin, X., Cai, Y., Yang, X. and Chia, W. (2000). Analysis of partner of inscuteable, a novel player of Drosophila asymmetric divisions, reveals two distinct steps in inscuteable apical localization. Cell 100, 399-409. 10.1016/S0092-8674(00)80676-5 [DOI] [PubMed] [Google Scholar]
- Yuzawa, S., Kamakura, S., Iwakiri, Y., Hayase, J. and Sumimoto, H. (2011). Structural basis for interaction between the conserved cell polarity proteins Inscuteable and Leu-Gly-Asn repeat-enriched protein (LGN). Proc. Natl. Acad. Sci. USA 108, 19210-19215. 10.1073/pnas.1110951108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou, Z., Alégot, H. and Irvine, K. D. (2019). Oriented Cell Divisions Are Not Required for Drosophila Wing Shape. Curr. Biol. 29, 856-864.e3. 10.1016/j.cub.2019.01.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu, J., Shang, Y., Xia, C., Wang, W., Wen, W. and Zhang, M. (2011a). Guanylate kinase domains of the MAGUK family scaffold proteins as specific phospho–protein–binding modules. EMBO J. 30, 4986-4997. 10.1038/emboj.2011.428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu, J., Wen, W., Zheng, Z., Shang, Y., Wei, Z., Xiao, Z., Pan, Z., Du, Q., Wang, W. and Zhang, M. (2011b). LGN/mInsc and LGN/NuMA complex structures suggest distinct functions in asymmetric cell division for the Par3/mInsc/LGN and Gαi/LGN/NuMA pathways. Mol. Cell 43, 418-431. 10.1016/j.molcel.2011.07.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu, J., Shang, Y., Chen, J. and Zhang, M. (2012). Structure and function of the guanylate kinase-like domain of the MAGUK family scaffold proteins. Front. Biol. 7, 379-396. 10.1007/s11515-012-1244-9 [DOI] [Google Scholar]
- Zhu, J., Shang, Y., Xia, Y., Zhang, R. and Zhang, M. (2016). An atypical MAGUK GK target recognition mode revealed by the interaction between DLG and KIF13B. Structure 24, 1876-1885. 10.1016/j.str.2016.08.008 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.