

Abstract
Programmed DNA double-strand breaks (DSBs) are necessary for meiosis in mammals. A sufficient number of DSBs ensure the normal pairing/synapsis of homologous chromosomes. Abnormal DSB repair undermines meiosis, leading to sterility in mammals. The DSBs that initiate recombination are repaired as crossovers and noncrossovers, and crossovers are required for correct chromosome separation. Thus, the placement, timing, and frequency of crossover formation must be tightly controlled. Importantly, mutations in many genes related to the formation and repair of DSB result in infertility in humans. These mutations cause nonobstructive azoospermia in men, premature ovarian insufficiency and ovarian dysgenesis in women. Here, we have illustrated the formation and repair of DSB in mammals, summarized major factors influencing the formation of DSB and the theories of crossover regulation.
Keywords: azoospermia, crossover, DSB, meiosis, recombination
INTRODUCTION
Meiosis is a specialized form of cell division, that generates haploid gametes from diploid cells, and it is essential for sexual reproduction and evolution.1 The programmed DNA double-strand breaks (DSBs) are critical in meiosis and the number of DSBs is tightly regulated. Excessive DSBs destabilize the genome, whereas insufficient DSBs impede the crossover (CO) formation, leading to erroneous separation of homologous chromosomes.2 Many proteins are involved in DSB repair, which generates COs and noncrossovers (NCOs).3 The CO repair pathway is critical for mammals. First, CO repair promotes the exchange of genetic material between homologous chromosomes. This exchange is essential for the genetic diversity that contributes to biological evolution and adaptation to environment changes.4,5 Second, COs establish physical connections between homologous chromosomes. These connections help maintain the balance of forces which ensure that the homologous chromosomes are correctly aligned on the equatorial plate, and pulled apart by the meiotic spindle.4 Therefore, it is crucial to understand the formation and repair of DSB in mammalian meiosis.
The formation and repair of meiotic DSB in mammals are complex and delicate. These processes include the formation of DSB, the resection of DSB ends, homology search, DNA strand invasion, as well as the stabilization and resolution of the double Holliday junctions (dHJs) as shown in Figure 1.
Figure 1.
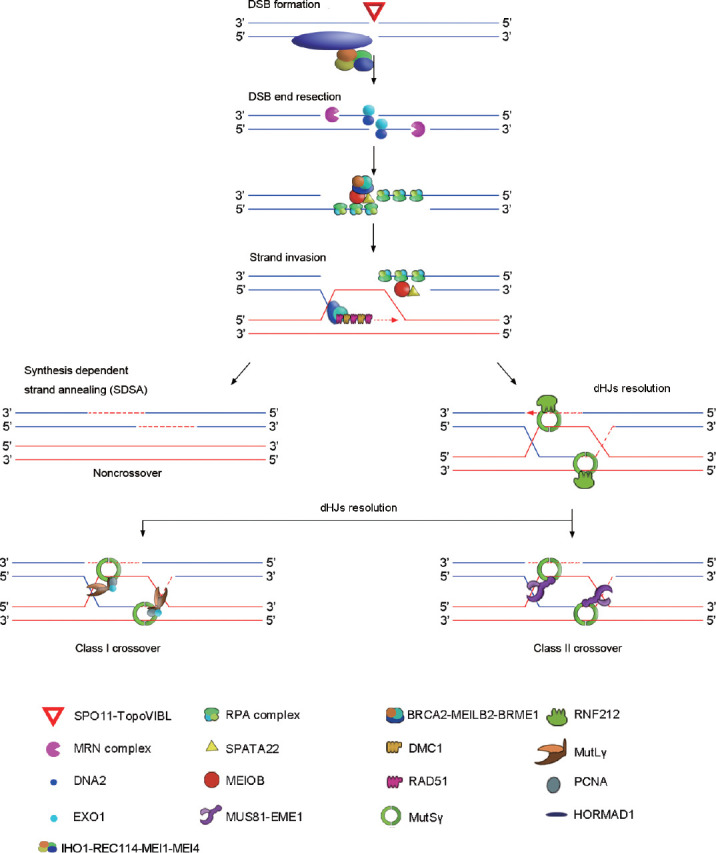
The formation and repair process of DSB in mammals. DSB: double-strand break.
THE FORMATION OF DSBS
Meiotic recombination is initiated by the programmed induction of DSBs by a topoisomerase-like enzyme SPO11. At the beginning of meiosis, SPO11 binds to DNA strands and generates DSBs.6 The SPO11 binding requires open chromatin, which is critically mediated by PRDM9, a meiosis-specific histone methyltransferase.7 PRDM9 contains a N-terminal KRAB domain involved in protein-protein interactions, a SSXRD domain that acts as a transcription repressor in SSX proteins, a SET domain with methyltransferase activity, and a zinc finger domain. This zinc finger domain contains a single proximal zinc finger separated from the rest of the terminal C2H2 zinc finger array comprised of 8 to over 20 fingers.8 PRDM9 methylates histone H3 at lysine 4 (H3K4) and lysine 36 (H3K36) at surrounding nucleosomes in vivo, hence H3K4me3/H3K36me3-double-positive histone modifications are iconic feature of the DSB hotspots.9 The terminal C2H2 zinc finger domain of PRDM9 binds to DNA motif and directs the positions of the DSB events that initiate meiotic recombination.10 The zinc finger array, encoded by a minisatellite repeat, is not only a variable among mouse strains, but also between humans and mice, which leads to the variability of DSB hotspots.11 PRDM9 binding sites, and thus hotspot locations are changed by the minisatellite sequence mutations such as duplications, deletions or rearrangements.11 The accessibility of PRDM9 to its special DNA motifs is enhanced by HELLS, which binds to PRDM9 as a pioneering factor and facilitates chromatin remodeling.12 Interestingly, the phenotype of Hells-deficient mice resembled those of Prdm9-knockout mice, and they both showed almost no changes in number of DSBs, but drastic changes in the locations of DSB on chromosomes.12,13 After binding to the DNA and modifying the nucleosomes, the methyltransferase PRDM9 somehow recruits SPO11 to cleave DNA at these meiotic DSB hotspots.14
SPO11 is an evolutionarily conserved topoisomerase-derived protein, and it is responsible for the formation of most DSBs with the assistance of TopoVIBL during meiosis.15 SPO11 contains two variants, SPO11α and SPO11β.16 SPO11α mediates DSB formation on sex chromosomes, while SPO11β mediates DSB formation on autosomes in mammals.17 SPO11β has an extra exon compared with SPO11α.16 TopoVIBL, a subunit of TopoVIB, which belongs to the topoisomerase type IIB family most likely regulates the cleavage of DNA through directly interacting with SPO11.15 Male Top6bl−/− mice were sterile and the formation of DSB was defective in spermatocytes.15 The activity of SPO11-TopoVIBL requires auxiliary protein complex, namely pre-DSB recombinosomes, including IHO1, REC114, MEI1, MEI4, and ANKRD31.18 HORMAD1 localizes along asynapsed axes and recruits IHO1 to establish an platform, on which focal pre-DSB recombinosomes assemble.18 The phenotypes of the Iho1, Mei1, Mei4, and Rec114-deficient mice were similar to those of the Spo11-deficient mice, showing abnormal DSB formation. Therefore, the pre-DSB recombinosomes are essential for meiotic DSB formation in mammals.6,19,20,21,22
It is worth noting that the regulatory mechanism of DSB formation on sex chromosomes is different from that on autosomes. ANKRD31 is highly and uniquely expressed in meiotic cells and appears between pre-leptotene and early-pachytene, which promotes DSB formation on the pseudo-autosomal region (PAR) of X and Y chromosome.23 ANKRD31 accumulates on the asynapsed chromosome axes, especially on the PAR of spermatocytes.23 The loss of ANKRD31 caused infertility in male mice and accelerated declining of fertility in female mice.23 Mo-2 arrays have been considered as one of the important factors responsible for regulating DSB formation on PAR.24 Mo-2 arrays are 20-kb minisatellites, with a 31-bp repeat localized on PAR of sex chromosomes and the noncentromeric ends of chromosomes 4, 9 and 13.24 Mo-2 arrays can recruit ANKRD31, which further recruits IHO1 and REC114 to DSB hotspot sites to form DSB.23,24 The formation of DSB mediated by Mo-2 arrays is independent of PRDM9, and heterochromatin histone modifications are enriched at the position of mo-2 arrays at the onset of meiosis.24
FACTORS INFLUENCING THE FORMATION OF DSBS
A sufficient number of DSBs are advantageous to ensure the pairing/synapsis of homologous chromosomes. However, excessive DSBs are harmful because irreparable DSBs undermine the integrity of the genome.2 Therefore, sophisticated machinery modulates the number of DSBs in mammals.
SPO11 dosage
The expression level of SPO11 determines the number of DSBs. SPO11β is mainly expressed in leptotene, and SPO11α is expressed in pachytene/diplotene spermatocytes.16 Doubled SPO11β expression increased the number of DSB by about 25%, whereas half SPO11β expression decreased the amount of DSBs by 20%–30%, indicating that the transgene dosage of SPO11 is critical for the number of DSBs.25,26
Chromosome length
Meiotic DSBs are not randomly distributed, as random distribution leads to a higher risk of mis-segregation for smaller chromosomes.27 Studies have shown that accessory DSB proteins, including REC114 and MER2, preferentially bind to short chromosomes and exist for a longer time to ensure that the short chromosomes have enough DSBs for faithful separation of homologous chromosomes in yeast.28 It should be noted that large chromosomes better tolerate fewer DSBs, and they are less vulnerable to entanglement than small chromosomes in mammals.26 Compared with the X chromosome, the Y chromosome seems to be relatively resistant to tangling.26
Synaptonemal complex
As a sufficient number of DSBs are needed to support the normal synapsis of chromosomes, vice versa the synapsed state of chromosomes affects DSB formation. Without synapsis, DSBs will persist on asynapsed segment of chromosomes until pachynema.26,29 This phenomenon may be related to HORMAD1 and pre-DSB recombinosomes. When homologous chromosomes are not fully synapsed or asynapsed, HORMAD1 serves as a platform to continuously recruit pre-DSB recombinosomes to the chromosome to promote DSB formation.18 When the homologous chromosomes are completely synapsed, HORMAD1 and pre-DSB recombinosomes dissociate from the chromosome to prevent overproduction of DSB.18 Moreover, synaptonemal complex promotes IHO1 depletion from synapsed chromosome axes.18
DNA damage-responsive phosphoinositide 3-kinase-related kinases
The DNA damage-responsive phosphoinositide 3-kinase (PI3K) related kinases including ATM, ATR and PRKDC, play important roles in the regulation of DSB in mammals. ATM, as a dimer with no biological activity in undamaged somatic cells, becomes active monomers through intermolecular auto-phosphorylation on serine 1981 after DSB formation.30 The phosphorylated ATM is recruited by the MRE11-RAD50-NBS1 (MRN) complex to the DSB sites and phosphorylates the MRN complex.30 Subsequently, ATM phosphorylates MDC1 and Serine 139 of H2AX.30 Phosphorylation of MDC1 recruits RING-finger ubiquitin ligases RNF8 and RNF168.30 RNF8 ubiquitinates γH2AX, which recruits 53bp1 and BRCA1, thereby spreading the DSB signal.30 ATM can also remove IHO1 from the chromosome axes by cooperating with ATR and PRKDC to inhibit DSBs in early pachynema, yet the specific molecular mechanism is still unclear.18
THE RESECTION OF DSB ENDS
The ends of DSB lack the 3′-hydroxyl or 5′-phosphate group after DSB formation.31 The MRN complex plays a crucial role in resecting the DSB ends, which is necessary for normal meiosis.32 MRE11 is a homodimer with the dual endonuclease and exonuclease activities.33 The MRN complex cleaves the 5’ strand away from the DSBs through CtIP and the endonuclease activity of MRE11.34 After cleavage, MRE11 generates a short single-stranded DNA (ssDNA) through its 3’ to 5’ exonuclease activity, while EXO1 and DNA2 further resect this end through their 5’ to 3’ exonuclease activity and finally produce a long single-stranded DNA end.32
RAD50, as a member of the ATP binding cassette (ABC) superfamily, interacts with two MRE11 monomers, and forms a globular domain that interacts with DNA.35 In the ATP-binding conformation, two catalytical RAD50s block the active sites of MRE11, whereas in the ATP-free conformation, RAD50 dissociates, allowing MRE11 to bind and hydrolyze the DNA phosphodiester backbone.35 NBS1 is the third subunit of the MRN complex and responds to the phosphorylation of CtIP through its FHA and BRCT domains, thereby stimulating the activity of the MRE11-RAD50 core complex through direct physical interactions with MRE11.35 Knockout of each component of MRN complex resulted in embryonic lethality in mice, and Nbs1 knockout in germ cells led to meiotic arrest and male infertility.34
HOMOLOGY SEARCH AND STRAND INVASION
The 3’ ssDNA overhangs are produced from resection of the ends of DSB and coated with RPA complex to protect ssDNA and remove secondary structure.36,37 RPA complex is comprised of three subunits of RPA1, RPA2, and RPA3.37 RPA1 is the largest and is mainly responsible for the DNA binding activity of the complex.38 The loss of RPA1 impaired loading of RAP2 and RPA3, and inhibited recruitment of DMC1 and RAD51 to the chromosome.38
MEIOB, a meiosis-specific protein containing the OB domain, forms a highly compact complex with SPATA22.36 The complex binds to ssDNA for recruiting the BRCA2-MEILB2-BRME1 complex.36,39,40 MEIOB and SPATA22 together facilitate the localization of BRCA2-MEILB2-BRME1 on ssDNA as shown that MEILB2 and BRME1 foci decreased substantially in Meiob−/− mice.41 The BRCA2-MEILB2-BRME1 complex directly recruits RAD51 and DMC1 for binding to ssDNA and replacing RPA.39
After recruited to ssDNA to replace RPA, DMC1 and RAD51 assemble with the ssDNA into nucleoprotein filaments to guide homologous searching and form D-loop structure together with the HOP2-MND1 heterodimer.42 It has been demonstrated that HOP2 and MND1 are two key accessory proteins to stimulate the synapsis phase of DNA strand exchange in vitro.43 Most DSBs were repaired in spermatocytes, with intact Hop2 but deleted Mnd1.44 However, DSBs were not repaired and spermatocytes were blocked at the pachytene-like stage in Hop2 knockout male mice.45 These findings indicate that HOP2 independently promotes strand invasion in vivo. The invading 3’ ssDNA overhang promotes the synthesis of new DNA; thereafter, the D-loop structure extends to the second end, allowing the capture of the 3’ ssDNA end of the other break. The DNA binding activity of MEIOB may play an essential role during this process.36 Meiob-deletion caused sterility in both genders due to meiotic arrest in zygotene or pachytene-like stage.36,46
THE STABILIZATION AND RESOLUTION OF THE DOUBLE HOLLIDAY JUNCTIONS
After the resection of 3’ ssDNA flap and end ligation, the dHJs are formed and maintained by multiple proteins.36 The DNA mismatch repair family proteins, MSH4 and MSH5, form the MutSγ complex, which stabilizes the dHJs.47 The MutSγ complex first appeared in zygonema as discrete foci on the chromosome axis, reached its maximum number in early pachynema, and declined in mid-pachynema.48,49 Msh4−/− mice and Msh5−/− mice were viable but sterile, with almost complete failure of homologous synapsis, as well as apoptosis of the meiotic cells before pachynema.48,49 The enrichment of MSH4 in the dHJs structure requires the participation of RNF212, which promotes the localization of MutSγ complex via SUMOylation.50
There are two main pathways in the resolution of dHJs based on its symmetry. When the dHJs are resolved in asymmetric manner by structure-specific endonucleases (MLH1-MLH3 and MUS81-EME), the COs are generated.3 In contrast, the symmetric resolution of dHJs results in the formation of NCOs. The formation of CO is regulated through class I or II mechanisms. In class I crossover, when the MutLγ complex formed by MLH1/MLH3 as heterodimer is recruited by the MutSγ complex on dHJs, it resolves the dHJs via the endonuclease activity.3 The endonuclease activity of MutLγ complex was abolished when the conserved metal binding DQHA(X)2E(X)4E motif of MLH3 was mutated.51 Besides, EXO1 and MutSγ can stimulate the endonuclease activity of MutLγ.51 Replication factor C (RFC) and the proliferating cell nuclear antigen (PCNA) can form a complex with MutLγ and MutSγ to preferentially cleave DNA with the Holliday junctions.51,52 On the other hand, class II crossover is independent of the MutLγ or MutSγ complex, and is produced by the structure-specific endonuclease MUS81 and EME1 in mammals.53 Mus81−/− mice were viable and fertile, but spermatocytes showed partial apoptosis in pachynema and metaphase.53
There are two speculative explanations for the regulatory mechanism of class I crossover and class II crossover. First, MSH4 and MSH5 bind to dHJs, and recruit MLH1 and MLH3 to the majority of the CO sites, while MUS81 is recruited to the remaining CO sites.53 However, the interaction between MSH4-MSH5 and MUS81 has not been validated. Second, others believe that MSH4-MSH5 and MUS81 belong to two distinct pathways, and MUS81 and MLH1-MLH3 antagonizes each other.53 It has been proved that human MUS81 can directly bind to dHJs in vitro.54 When occupying a subset of CO sites, MUS81 prevents MLH1-MLH3 from binding to these sites.53 These antagonistic effects disappear and MLH1-MLH3 occupies all CO sites in the absence of MUS81.53 These two models are reasonable hypotheses and need to be further investigated. Currently, BTBD12 has received attention on its potential role in homologous recombination in mammals. BTBD12 interacts with many of the key players such as BLM in both CO pathways.55 Importantly, MLH1-MLH3 focus numbers increased when BTBD12 was defective.55 Therefore, BTBD12 may functionally integrate the different CO pathways during mammalian meiosis.
The formation and repair of mammalian DSBs involve a large number of genes and proteins as summarized in Table 1. Knockout of these genes exerts important impact on meiosis.
Table 1.
Phenotypes of knockout male mice related to the formation and repair of double-strand breaks
| Gene name | Arrest stages | Male phenotype | Reference |
|---|---|---|---|
| Prdm9 | Pachytene-like | Infertility | Mihola et al.13 |
| Hells | Pachytene-like | Infertility | Spruce et al.12 |
| Hormad1 | Pachytene-like | Infertility | Stanzione et al.19 |
| Iho1 | Zygotene-pachytene | Infertility | Stanzione et al.19 |
| Mei1 | Zygotene-like | Infertility | Libby et al.22 |
| Mei4 | Zygotene-like | Infertility | Kumar et al.21 |
| Rec114 | Zygotene-like | Infertility | Kumar et al.20 |
| Ankrd31 | Pachytene-like | Infertility | Papanikos et al.23 |
| Spo11 | Zygotene-like | Infertility | Baudat et al.6 |
| Top6bl | Zygotene-like | Infertility | Robert et al.15 |
| Sycp1 | Pachytene | Infertility | de Vries et al.84 |
| Sycp2 | Zygotene-like | Infertility | Yang et al.85 |
| Sycp3 | Zygotene-like | Infertility | Yuan et al.86 |
| Scre | Late zygotene | Infertility | Liu et al.87 |
| Syce1 | Pachytene | Infertility | Bolcun-Filas et al.88 |
| Syce2 | Pachytene | Infertility | Bolcun-Filas et al.89 |
| Syce3 | Pachytene | Infertility | Schramm et al.90 |
| Tex11 | Anaphase | Infertility | Yang et al.91 |
| Tex12 | Pachytene | Infertility | Hamer et al.92 |
| Six6os1 | Pachytene-like | Infertility | Gomez et al.93 |
| Rpa1 | Metaphase to anaphase | Infertility | Shi et al.38 |
| Meilb2 | Zygotene | Infertility | Zhang et al.94 |
| Dmc1 | Early-zygotene | Infertility | Yoshida et al.95 |
| Spata22 | Late zygotene-like | Infertility | La Salle et al.96 |
| Meiok21 | Zygotene/pachytene | Infertility | Shang et al.97 |
| Hop2 | Pachytene-like | Infertility | Petukhova et al.45 |
| Meiob | Pachytene-like | Infertility | Luo et al.36 |
| Souquet et al.46 | |||
| Brme1 | Mid-late pachytene | Infertility | Takemoto et al.39 |
| Brca2 | Zygotene | Infertility | Zhang et al.94 |
| Msh4/5 | Pachytene-like | Infertility | Kneitz et al.48 |
| de Vries et al.49 | |||
| Rnf212 | Metaphase | Infertility | Reynolds et al.50 |
| Mlh1 | Pachytene | Infertility | Edelmann et al.100 |
| Mlh3 | Metaphase | Infertility | Lipkin et al.98 |
CHARACTERISTICS OF CO SITES IN MAMMALIAN MEIOSIS
DSBs are predominantly converted to NCOs, with only around 10% converted to COs.3 Why are there so many DSBs but only a small number of COs formed? Exploring the characteristics of CO sites elicits an important speculation that the chromatin state is more open on CO sites than NCO sites.56 COs formation requires a stable and permissive chromatin environment to recruit CO-related proteins, including the MutSγ complex and MutLγ complex.56 Thus, DSB hotspots with looser chromatin have a greater chance of forming COs. Second, the designated CO sites are easier to be bound by PRDM9.57 PRDM9 binding on the template chromosome increases the chance of DSBs being resolved as a CO.57 Third, CO sites have higher GC content, and DSB hotspots with higher GC content are more likely to be repaired by the CO pathway.57
THE THEORIES OF CO REGULATION
Meiotic bet-hedging strategy
Recombination is a “double-edged sword” which generates new alleles to adapt to environmental changes and increases the diversity of organisms.2 On the other hand, excessive recombination impairs genome stability in the organism.4,5 COs only happen when benefits outweigh the adverse effects. More COs in gametes are advantageous when the environment changes while fewer COs in gametes are beneficial during environmental stasis.5 Organisms effectively adapt to environment changes through delicate control of the number of COs.
CO assurance
CO formation is essential for proper chromosome segregation in meiosis. CO assurance refers to needing at least one CO (obligate CO) between each pair of homologous chromosomes.58 Previous data indicate that even a single DSB is converted to a CO with high efficiency in the Caenorhabditis elegans. However, the theory of CO assurance still needs to be validated in mammals.59
CO maturation inefficiency (CMI)
Organisms randomly generate and remove a small subset of COs, at the pre-designated CO sites, during maturation of COs to ensure that only one or two COs occur on a chromosome. This phenomenon is defined as CMI and is only observed in human females.60 When CMI is abnormally enhanced or diminished, aneuploidy segregation of chromosomes occurs.5 The existence of this mechanism seems counterintuitive but is of great significance. First, CMI increases the time between two consecutive pregnancies in young women, and reduces the birth rate to ensure that existing children have sufficient resources to survive.60 Second, CMI interacts with other factors to reduce the fertility of elderly women.60
CO interference
The phenomenon of CO interference of one CO on the chromosome will reduce the probability of another CO occurring within a certain interval nearby.58 The strength of this interference effect decreases with increasing distance.61 Interestingly, CO interference appears at the level of the whole chromosome, but is limited to specific domains, indicating that COs communicate at the level of the entire chromosome.62 The molecular mechanism of CO interference is unclear and needs to be studied.
CO homeostasis
The number of COs remains roughly constant as the number of DSBs change.25,58 Homeostatic control is enforced during at least two stages in mice, with one stage being after the formation of early recombination intermediates, and the other stage being the maturation of these intermediates toward COs,25 and the latter stage may be related with CMI.
CO patterning
CO patterning describes that the production of COs is inhibited on the telomeres and centromeres in many species including humans.63 However, the mouse chromosomes are acrocentric, and male mice have a greater number of COs near the telomeres.57 The differences may be explained by the sex chromosomes characteristics of mice. Because the PAR is located in the sub-telomere region in mice, there is an obligation for a CO in the PAR of male mice to ensure the normal separation of the X and Y chromosome.64
ABNORMAL FORMATION AND REPAIR OF DSBS CAUSE HUMAN INFERTILITY
The natality around the world including China is declining, and infertility is one of the main reasons. About 15% of couples have infertility problems, and half of the problems occur in men.65 So far, mutations in many genes involved the formation and repair of DSBs have been reported to be the cause of spermatocyte arrest.
Three genes involved in the formation of DSB have been reported to be related to human infertility, including PRDM9, SPO11 and MEI1. Two SNPs (G433V and T685R) of PRDM9 have been found to exist in patients with azoospermia, but not in fertile subjects.66 The results indicate that these mutations in PRDM9 may be specifically related to male azoospermia. The p.Glu186Lys variant of SPO11 caused meiotic arrest in two brothers from a family in the Middle East.67 Consistent with its function in mice, the homozygous missense mutation of MEI1 caused two brothers from a consanguineous family to suffer from nonobstructive azoospermia (NOA) and meiosis arrest.68
Most of the genes responsible for human infertility are involved in the repair of DSBs during meiosis. The deletion of a glutamate residue in HOP2 (Q201del) in humans damaged the estrogen signal transduction pathway, impaired the size of the follicular pool during fetal development, and caused follicular atresia during puberty, which eventually resulted in ovarian dysgenesis.69 A homozygous missense mutation (c.106G>A) in DMC1, which co-segregated with NOA and primary ovarian insufficiency (POI) phenotypes in a consanguineous Chinese family has been identified.70 This single amino acid substitution results in protein misfolding and malfunction of DMC1.70 STAG3 was first described as a POI gene in 2014, and a homozygous missense variant resulted in POI and NOA in human.71,72,73,74 The truncation of the C-terminal of the MEIOB (c.1218G>A) caused female POI.65 A nonsynonymous amino acid mutation (N64I) in MEIOB caused azoospermia.75 In the process of stabilizing dHJs, the homozygous mutation of MSH4 (P638L and S754L) caused arrest of spermatogenic maturation in human males.76 Studies have found that missense mutations in TEX11 (V748A) caused male infertility with NOA.77 It has also been reported that a homozygous RNF212 (c.111dupT) variant caused azoospermia in male siblings.73 In clinical patients, a homozygous frameshift variant in DNA mismatch repair gene MLH3 (c.3632delA) was also found to cause severe oligozoospermia, leading to male infertility.78
Mutations in related proteins of synaptonemal complex have also been reported to cause infertility in humans, including SYCP2, SYCP3 and SYCE1. Exome sequencing of infertile males revealed three heterozygous SYCE2 frameshift variants in patients with cryptozoospermia and azoospermia.79 A 1-bp deletion (643delA) in SYCP3 has been identified in two patients, and the mutation results in a premature stop codon and truncation of the C-terminal, coiled-coil-forming region of the SYCP3 protein, leading to azoospermia with meiotic arrest in human.80 Mutations in the SYCE1 gene have also been reported in humans; for example, a nonsense homozygous mutation results in POI, and homozygous mutations at two different splice sites cause NOA.81,82,83
Mutations in many genes related to the formation and repair of DSBs have been found to cause human infertility, and a summary is shown in (Table 2). Collectively, these data indicate that the establishment and repair of DSBs are indispensable for the maintenance of human fertility.
Table 2.
Meiosis-associated genes that cause human infertility
| Gene name | Clinical phenotype | Mutation type | Reference |
|---|---|---|---|
| PRDM9 | NOA | SNPs | Irie et al.66 |
| SPO11 | NOA | Missense | Fakhro et al.67 |
| MEI1 | NOA and ovarian dysgeneis | Missense | Ben Khelifa et al.68 |
| HOP2 | Ovarian dysgeneis | Deletion of one residue (Glu210) | Zangen et al.69 |
| DMC1 | NOA and POI | Missense | He et al.70 |
| MEIOB | NOA and POI | Frame-shift deletion and missense | Gershoni et al.75 |
| Gershoni et al.65 | |||
| RNF212 | NOA | Frame-shift insertion | Krausz et al.76 |
| TEX11 | NOA | Missense | Yang et al.77 |
| SYCP2 | NOA | Frame-shift deletion | Schilit et al.79 |
| SYCP3 | NOA | Frame-shift deletion | Miyamoto et al.80 |
| SYCE1 | NOA and POI | Nonsense and splice site homozygous mutation | de Vries et al.81 |
| Maor-Sagie et al.82 | |||
| Pashaei et al.83 | |||
| STAG3 | NOA and POI | Missense | Riera-Escamilla et al.73 |
| Jaillard et al.74 | |||
| MSH4 | NOA and POI | Missense and splice homozygous site mutation | Carlosama et al.99 |
| Krausz et al.76 | |||
| MLH3 | Oligozoospermia | Frame-shift deletion | Nawaz et al.78 |
NOA: nonobstructive azoospermia; POI: premature ovarian insufficiency; SNP: single-nucleotide polymorphisms
CONCLUSION
Although numerous studies have revealed the mechanism of formation and repair of DSBs in mammalian meiosis, there are still lots of unanswered questions, especially the mechanism for the regulation of COs. First, in addition to the open chromatin status of CO sites, there should be more potential factors that affect the locations of CO. Future experiments need to be carried out to identify the specificity of CO sites, especially the chromatin environment around them. Second, RNF212 is an important protein that marks CO sites during mid-pachytene.50 RNF212 contains the RING-finger domain, which is an E3 ubiquitin ligase that can catalyze the modification of proteins by ubiquitination of molecules.50 Therefore, the selection of CO sites may be affected by ubiquitination or SUMO modification. Second, considering that the DSB sites have a special motif to recruit PRDM9, whether the CO sites contain a specific motif which recruits CO-related proteins including MutSγ and MutLγ complexes is unknown in mammals. Third, most DSBs are converted to NCOs in meiotic cells.3 Whether this phenomenon is related to CO interference is still unclear. Moreover, as the number of DSBs formed in meiotic cells is relatively large compared to the demand for COs, it is interesting to explore the “threshold” of the number of DSBs.
AUTHOR CONTRIBUTIONS
WQ and MCL reviewed the literature, collected data, and wrote the manuscript. CL, YTX, and YMX collaborated in writing, revising, and editing the manuscript. MCL planned this article, designed the concept of this article, helped in literature research and writing the manuscript, and supervised this project. All authors read and approved the final manuscript.
COMPETING INTERESTS
All authors declare no competing interests.
ACKNOWLEDGMENTS
We apologize to those colleagues whose research is not included here because of limited space. This work is supported by National Key Research & Development Program of China (No. 2018YFC1003400), National Natural Science Foundation of China (No. 31771588) and the Strategic Collaborative Research Program of the Ferring Institute of Reproductive Medicine (No. FIRMC200509) to MCL.
REFERENCES
- 1.Parvanov ED, Petkov PM, Paigen K. Prdm9 controls activation of mammalian recombination hotspots. Science. 2010;327:835. doi: 10.1126/science.1181495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kim S, Peterson SE, Jasin M, Keeney S. Mechanisms of germ line genome instability. Semin Cell Dev Biol. 2016;54:177–87. doi: 10.1016/j.semcdb.2016.02.019. [DOI] [PubMed] [Google Scholar]
- 3.Toledo M, Sun X, Brieno-Enriquez MA, Raghavan V, Gray S, et al. A mutation in the endonuclease domain of mouse MLH3 reveals novel roles for MutLgamma during crossover formation in meiotic prophase I. PLoS Genet. 2019;15:e1008177. doi: 10.1371/journal.pgen.1008177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yadav VK, Claeys Bouuaert C. Mechanism and control of meiotic DNA double-strand break formation in S. cerevisiae. Front Cell Dev Biol. 2021;9:642737. doi: 10.3389/fcell.2021.642737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang S, Veller C, Sun F, Ruiz-Herrera A, Shang Y, et al. Per-nucleus crossover covariation and implications for evolution. Cell. 2019;177:326–38.e16. doi: 10.1016/j.cell.2019.02.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Baudat F, Manova K, Yuen JP, Jasin M, Keeney S. Chromosome synapsis defects and sexually dimorphic meiotic progression in mice lacking Spo11. Mol Cell. 2000;6:989–98. doi: 10.1016/s1097-2765(00)00098-8. [DOI] [PubMed] [Google Scholar]
- 7.Keeney S, Giroux CN, Kleckner N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell. 1997;88:375–84. doi: 10.1016/s0092-8674(00)81876-0. [DOI] [PubMed] [Google Scholar]
- 8.Imai Y, Baudat F, Taillepierre M, Stanzione M, Toth A, et al. The PRDM9 KRAB domain is required for meiosis and involved in protein interactions. Chromosoma. 2017;126:681–95. doi: 10.1007/s00412-017-0631-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Powers NR, Parvanov ED, Baker CL, Walker M, Petkov PM, et al. The meiotic recombination activator PRDM9 trimethylates both H3K36 and H3K4 at recombination hotspots in vivo. PLoS Genet. 2016;12:e1006146. doi: 10.1371/journal.pgen.1006146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sandovici I, Sapienza C. PRDM9 sticks its zinc fingers into recombination hotspots and between species. F1000 Biol Rep. 2010;2:37. doi: 10.3410/B2-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Davies BJ, Hatton E, Altemose N, Hussin JG, Pratto F, et al. Re-engineering the zinc fingers of PRDM9 reverses hybrid sterility in mice. Nature. 2016;530:171–6. doi: 10.1038/nature16931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Spruce C, Dlamini S, Ananda G, Bronkema N, Tian H, et al. HELLS and PRDM9 form a pioneer complex to open chromatin at meiotic recombination hot spots. Genes Dev. 2020;34:398–412. doi: 10.1101/gad.333542.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mihola O, Trachtulec Z, Vlcek C, Schimenti JC, Forejt J. A mouse speciation gene encodes a meiotic histone H3 methyltransferase. Science. 2009;323:373–5. doi: 10.1126/science.1163601. [DOI] [PubMed] [Google Scholar]
- 14.Paiano J, Wu W, Yamada S, Sciascia N, Callen E, et al. ATM and PRDM9 regulate SPO11-bound recombination intermediates during meiosis. Nat Commun. 2020;11:857. doi: 10.1038/s41467-020-14654-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Robert T, Nore A, Brun C, Maffre C, Crimi B, et al. The TopoVIB-Like protein family is required for meiotic DNA double-strand break formation. Science. 2016;351:943–9. doi: 10.1126/science.aad5309. [DOI] [PubMed] [Google Scholar]
- 16.Bellani MA, Boateng KA, McLeod D, Camerini-Otero RD. The expression profile of the major mouse SPO11 isoforms indicates that SPO11beta introduces double strand breaks and suggests that SPO11alpha has an additional role in prophase in both spermatocytes and oocytes. Mol Cell Biol. 2010;30:4391–403. doi: 10.1128/MCB.00002-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kauppi L, Barchi M, Baudat F, Romanienko PJ, Keeney S, et al. Distinct properties of the XY pseudoautosomal region crucial for male meiosis. Science. 2011;331:916–20. doi: 10.1126/science.1195774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dereli I, Stanzione M, Olmeda F, Papanikos F, Baumann M, et al. Four-pronged negative feedback of DSB machinery in meiotic DNA-break control in mice. Nucleic Acids Res. 2021;49:2609–28. doi: 10.1093/nar/gkab082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stanzione M, Baumann M, Papanikos F, Dereli I, Lange J, et al. Meiotic DNA break formation requires the unsynapsed chromosome axis-binding protein IHO1 (CCDC36) in mice. Nat Cell Biol. 2016;18:1208–20. doi: 10.1038/ncb3417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kumar R, Oliver C, Brun C, Juarez-Martinez AB, Tarabay Y, et al. Mouse REC114 is essential for meiotic DNA double-strand break formation and forms a complex with MEI4. Life Sci Alliance. 2018;1:e201800259. doi: 10.26508/lsa.201800259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kumar R, Bourbon HM, de Massy B. Functional conservation of Mei4 for meiotic DNA double-strand break formation from yeasts to mice. Genes Dev. 2010;24:1266–80. doi: 10.1101/gad.571710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Libby BJ, Reinholdt LG, Schimenti JC. Positional cloning and characterization of Mei1, a vertebrate-specific gene required for normal meiotic chromosome synapsis in mice. Proc Natl Acad Sci U S A. 2003;100:15706–11. doi: 10.1073/pnas.2432067100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Papanikos F, Clement JA, Testa E, Ravindranathan R, Grey C, et al. Mouse ANKRD31 regulates spatiotemporal patterning of meiotic recombination initiation and ensures recombination between X and Y sex chromosomes. Mol Cell. 2019;74:1069–85.e11. doi: 10.1016/j.molcel.2019.03.022. [DOI] [PubMed] [Google Scholar]
- 24.Acquaviva L, Boekhout M, Karasu ME, Brick K, Pratto F, et al. Ensuring meiotic DNA break formation in the mouse pseudoautosomal region. Nature. 2020;582:426–31. doi: 10.1038/s41586-020-2327-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cole F, Kauppi L, Lange J, Roig I, Wang R, et al. Homeostatic control of recombination is implemented progressively in mouse meiosis. Nat Cell Biol. 2012;14:424–30. doi: 10.1038/ncb2451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kauppi L, Barchi M, Lange J, Baudat F, Jasin M, et al. Numerical constraints and feedback control of double-strand breaks in mouse meiosis. Genes Dev. 2013;27:873–86. doi: 10.1101/gad.213652.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Keeney S, Lange J, Mohibullah N. Self-organization of meiotic recombination initiation: general principles and molecular pathways. Annu Rev Genet. 2014;48:187–214. doi: 10.1146/annurev-genet-120213-092304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Murakami H, Lam I, Huang PC, Song J, van Overbeek M, et al. Multilayered mechanisms ensure that short chromosomes recombine in meiosis. Nature. 2020;582:124–8. doi: 10.1038/s41586-020-2248-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cloutier JM, Mahadevaiah SK, ElInati E, Nussenzweig A, Toth A, et al. Histone H2AFX links meiotic chromosome asynapsis to prophase I oocyte loss in mammals. PLoS Genet. 2015;11:e1005462. doi: 10.1371/journal.pgen.1005462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Derheimer FA, Kastan MB. Multiple roles of ATM in monitoring and maintaining DNA integrity. FEBS Lett. 2010;584:3675–81. doi: 10.1016/j.febslet.2010.05.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Serrano-Benitez A, Cortes-Ledesma F, Ruiz JF. “An end to a means”: how DNA-end structure shapes the double-strand break repair process. Front Mol Biosci. 2019;6:153. doi: 10.3389/fmolb.2019.00153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ceppi I, Howard SM, Kasaciunaite K, Pinto C, Anand R, et al. CtIP promotes the motor activity of DNA2 to accelerate long-range DNA end resection. Proc Natl Acad Sci U S A. 2020;117:8859–69. doi: 10.1073/pnas.2001165117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lafrance-Vanasse J, Williams GJ, Tainer JA. Envisioning the dynamics and flexibility of Mre11-Rad50-Nbs1 complex to decipher its roles in DNA replication and repair. Prog Biophys Mol Biol. 2015;117:182–93. doi: 10.1016/j.pbiomolbio.2014.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zhang B, Tang Z, Li L, Lu LY. NBS1 is required for SPO11-linked DNA double-strand break repair in male meiosis. Cell Death Differ. 2020;27:2176–90. doi: 10.1038/s41418-020-0493-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Syed A, Tainer JA. The MRE11-RAD50-NBS1 complex conducts the orchestration of damage signaling and outcomes to stress in DNA replication and repair. Annu Rev Biochem. 2018;87:263–94. doi: 10.1146/annurev-biochem-062917-012415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Luo M, Yang F, Leu NA, Landaiche J, Handel MA, et al. MEIOB exhibits single-stranded DNA-binding and exonuclease activities and is essential for meiotic recombination. Nat Commun. 2013;4:2788. doi: 10.1038/ncomms3788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Crickard JB, Greene EC. Biochemical attributes of mitotic and meiotic presynaptic complexes. DNA Repair (Amst) 2018;71:148–57. doi: 10.1016/j.dnarep.2018.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Shi B, Xue J, Yin H, Guo R, Luo M, et al. Dual functions for the ssDNA-binding protein RPA in meiotic recombination. PLoS Genet. 2019;15:e1007952. doi: 10.1371/journal.pgen.1007952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Takemoto K, Tani N, Takada-Horisawa Y, Fujimura S, Tanno N, et al. Meiosis-specific C19orf57/4930432K21Rik/BRME1 modulates localization of RAD51 and DMC1 to DSBs in mouse meiotic recombination. Cell Rep. 2020;31:107686. doi: 10.1016/j.celrep.2020.107686. [DOI] [PubMed] [Google Scholar]
- 40.Ribeiro J, Dupaigne P, Petrillo C, Ducrot C, Duquenne C, et al. The meiosis-specific MEIOB-SPATA22 complex cooperates with RPA to form a compacted mixed MEIOB/SPATA22/RPA/ssDNA complex. DNA Repair (Amst) 2021;102:103097. doi: 10.1016/j.dnarep.2021.103097. [DOI] [PubMed] [Google Scholar]
- 41.Zhang J, Gurusaran M, Fujiwara Y, Zhang K, Echbarthi M, et al. The BRCA2-MEILB2-BRME1 complex governs meiotic recombination and impairs the mitotic BRCA2-RAD51 function in cancer cells. Nat Commun. 2020;11:2055. doi: 10.1038/s41467-020-15954-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Petukhova GV, Pezza RJ, Vanevski F, Ploquin M, Masson JY, et al. The Hop2 and Mnd1 proteins act in concert with Rad51 and Dmc1 in meiotic recombination. Nat Struct Mol Biol. 2005;12:449–53. doi: 10.1038/nsmb923. [DOI] [PubMed] [Google Scholar]
- 43.Pezza RJ, Camerini-Otero RD, Bianco PR. Hop2-Mnd1 condenses DNA to stimulate the synapsis phase of DNA strand exchange. Biophys J. 2010;99:3763–72. doi: 10.1016/j.bpj.2010.10.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tsubouchi H, Roeder GS. The Mnd1 protein forms a complex with hop2 to promote homologous chromosome pairing and meiotic double-strand break repair. Mol Cell Biol. 2002;22:3078–88. doi: 10.1128/MCB.22.9.3078-3088.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Petukhova GV, Romanienko PJ, Camerini-Otero RD. The Hop2 protein has a direct role in promoting interhomolog interactions during mouse meiosis. Dev Cell. 2003;5:927–36. doi: 10.1016/s1534-5807(03)00369-1. [DOI] [PubMed] [Google Scholar]
- 46.Souquet B, Abby E, Herve R, Finsterbusch F, Tourpin S, et al. MEIOB targets single-strand DNA and is necessary for meiotic recombination. PLoS Genet. 2013;9:e1003784. doi: 10.1371/journal.pgen.1003784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lahiri S, Li Y, Hingorani MM, Mukerji I. MutSgamma-induced DNA conformational changes provide insights into its role in meiotic recombination. Biophys J. 2018;115:2087–101. doi: 10.1016/j.bpj.2018.10.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kneitz B, Cohen PE, Avdievich E, Zhu L, Kane MF, et al. MutS homolog 4 localization to meiotic chromosomes is required for chromosome pairing during meiosis in male and female mice. Genes Dev. 2000;14:1085–97. [PMC free article] [PubMed] [Google Scholar]
- 49.de Vries SS, Baart EB, Dekker M, Siezen A, de Rooij DG, et al. Mouse MutS-like protein Msh5 is required for proper chromosome synapsis in male and female meiosis. Genes Dev. 1999;13:523–31. doi: 10.1101/gad.13.5.523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Reynolds A, Qiao H, Yang Y, Chen JK, Jackson N, et al. RNF212 is a dosage-sensitive regulator of crossing-over during mammalian meiosis. Nat Genet. 2013;45:269–78. doi: 10.1038/ng.2541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cannavo E, Sanchez A, Anand R, Ranjha L, Hugener J, et al. Regulation of the MLH1-MLH3 endonuclease in meiosis. Nature. 2020;586:618–22. doi: 10.1038/s41586-020-2592-2. [DOI] [PubMed] [Google Scholar]
- 52.Kulkarni DS, Owens SN, Honda M, Ito M, Yang Y, et al. PCNA activates the MutLgamma endonuclease to promote meiotic crossing over. Nature. 2020;586:623–7. doi: 10.1038/s41586-020-2645-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Holloway JK, Booth J, Edelmann W, McGowan CH, Cohen PE. MUS81 generates a subset of MLH1-MLH3-independent crossovers in mammalian meiosis. PLoS Genet. 2008;4:e1000186. doi: 10.1371/journal.pgen.1000186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Taylor ER, McGowan CH. Cleavage mechanism of human Mus81-Eme1 acting on Holliday-junction structures. Proc Natl Acad Sci U S A. 2008;105:3757–62. doi: 10.1073/pnas.0710291105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Svendsen JM, Harper JW. GEN1/Yen1 and the SLX4 complex: solutions to the problem of Holliday junction resolution. Genes Dev. 2010;24:521–36. doi: 10.1101/gad.1903510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chen Y, Lyu R, Rong B, Zheng Y, Lin Z, et al. Refined spatial temporal epigenomic profiling reveals intrinsic connection between PRDM9-mediated H3K4me3 and the fate of double-stranded breaks. Cell Res. 2020;30:256–68. doi: 10.1038/s41422-020-0281-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hinch AG, Zhang G, Becker PW, Moralli D, Hinch R, et al. Factors influencing meiotic recombination revealed by whole-genome sequencing of single sperm. Science. 2019;363:eaau8861. doi: 10.1126/science.aau8861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Saito TT, Colaiacovo MP. Regulation of crossover frequency and distribution during meiotic recombination. Cold Spring Harb Symp Quant Biol. 2017;82:223–34. doi: 10.1101/sqb.2017.82.034132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Rosu S, Libuda DE, Villeneuve AM. Robust crossover assurance and regulated interhomolog access maintain meiotic crossover number. Science. 2011;334:1286–9. doi: 10.1126/science.1212424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wang S, Liu Y, Shang Y, Zhai B, Yang X, et al. Crossover interference, crossover maturation, and human aneuploidy. Bioessays. 2019;41:e1800221. doi: 10.1002/bies.201800221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kleckner N, Zickler D, Jones GH, Dekker J, Padmore R, et al. A mechanical basis for chromosome function. Proc Natl Acad Sci U S A. 2004;101:12592–7. doi: 10.1073/pnas.0402724101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Libuda DE, Uzawa S, Meyer BJ, Villeneuve AM. Meiotic chromosome structures constrain and respond to designation of crossover sites. Nature. 2013;502:703–6. doi: 10.1038/nature12577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ottolini CS, Newnham L, Capalbo A, Natesan SA, Joshi HA, et al. Genome-wide maps of recombination and chromosome segregation in human oocytes and embryos show selection for maternal recombination rates. Nat Genet. 2015;47:727–35. doi: 10.1038/ng.3306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Burgoyne PS. Genetic homology and crossing over in the X and Y chromosomes of mammals. Hum Genet. 1982;61:85–90. doi: 10.1007/BF00274192. [DOI] [PubMed] [Google Scholar]
- 65.Gershoni M, Hauser R, Barda S, Lehavi O, Arama E, et al. A new MEIOB mutation is a recurrent cause for azoospermia and testicular meiotic arrest. Hum Reprod. 2019;34:666–71. doi: 10.1093/humrep/dez016. [DOI] [PubMed] [Google Scholar]
- 66.Irie S, Tsujimura A, Miyagawa Y, Ueda T, Matsuoka Y, et al. Single-nucleotide polymorphisms of the PRDM9 (MEISETZ) gene in patients with nonobstructive azoospermia. J Androl. 2009;30:426–31. doi: 10.2164/jandrol.108.006262. [DOI] [PubMed] [Google Scholar]
- 67.Fakhro KA, Elbardisi H, Arafa M, Robay A, Rodriguez-Flores JL, et al. Point-of-care whole-exome sequencing of idiopathic male infertility. Genet Med. 2018;20:1365–73. doi: 10.1038/gim.2018.10. [DOI] [PubMed] [Google Scholar]
- 68.Ben Khelifa M, Ghieh F, Boudjenah R, Hue C, Fauvert D, et al. A MEI1 homozygous missense mutation associated with meiotic arrest in a consanguineous family. Hum Reprod. 2018;33:1034–7. doi: 10.1093/humrep/dey073. [DOI] [PubMed] [Google Scholar]
- 69.Zangen D, Kaufman Y, Zeligson S, Perlberg S, Fridman H, et al. XX ovarian dysgenesis is caused by a PSMC3IP/HOP2 mutation that abolishes coactivation of estrogen-driven transcription. Am J Hum Genet. 2011;89:572–9. doi: 10.1016/j.ajhg.2011.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.He WB, Tu CF, Liu Q, Meng LL, Yuan SM, et al. DMC1 mutation that causes human non-obstructive azoospermia and premature ovarian insufficiency identified by whole-exome sequencing. J Med Genet. 2018;55:198–204. doi: 10.1136/jmedgenet-2017-104992. [DOI] [PubMed] [Google Scholar]
- 71.Caburet S, Arboleda VA, Llano E, Overbeek PA, Barbero JL, et al. Mutant cohesin in premature ovarian failure. N Engl J Med. 2014;370:943–9. doi: 10.1056/NEJMoa1309635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.van der Bijl N, Ropke A, Biswas U, Woste M, Jessberger R, et al. Mutations in the stromal antigen 3 (STAG3) gene cause male infertility due to meiotic arrest. Hum Reprod. 2019;34:2112–9. doi: 10.1093/humrep/dez204. [DOI] [PubMed] [Google Scholar]
- 73.Riera-Escamilla A, Enguita-Marruedo A, Moreno-Mendoza D, Chianese C, Sleddens-Linkels E, et al. Sequencing of a ‘mouse azoospermia’ gene panel in azoospermic men: identification of RNF212 and STAG3 mutations as novel genetic causes of meiotic arrest. Hum Reprod. 2019;34:978–88. doi: 10.1093/humrep/dez042. [DOI] [PubMed] [Google Scholar]
- 74.Jaillard S, McElreavy K, Robevska G, Akloul L, Ghieh F, et al. STAG3 homozygous missense variant causes primary ovarian insufficiency and male non-obstructive azoospermia. Mol Hum Reprod. 2020;26:665–77. doi: 10.1093/molehr/gaaa050. [DOI] [PubMed] [Google Scholar]
- 75.Gershoni M, Hauser R, Yogev L, Lehavi O, Azem F, et al. A familial study of azoospermic men identifies three novel causative mutations in three new human azoospermia genes. Genet Med. 2017;19:998–1006. doi: 10.1038/gim.2016.225. [DOI] [PubMed] [Google Scholar]
- 76.Krausz C, Riera-Escamilla A, Moreno-Mendoza D, Holleman K, Cioppi F, et al. Genetic dissection of spermatogenic arrest through exome analysis: clinical implications for the management of azoospermic men. Genet Med. 2020;22:1956–66. doi: 10.1038/s41436-020-0907-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Yang F, Silber S, Leu NA, Oates RD, Marszalek JD, et al. TEX11 is mutated in infertile men with azoospermia and regulates genome-wide recombination rates in mouse. EMBO Mol Med. 2015;7:1198–210. doi: 10.15252/emmm.201404967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Nawaz S, Ullah MI, Hamid BS, Nargis J, Nawaz M, et al. A loss-of-function variant in DNA mismatch repair gene MLH3 underlies severe oligozoospermia. J Hum Genet. 2021;66:725–30. doi: 10.1038/s10038-021-00907-z. [DOI] [PubMed] [Google Scholar]
- 79.Schilit SL, Menon S, Friedrich C, Kammin T, Wilch E, et al. SYCP2 translocation-mediated dysregulation and frameshift variants cause human male infertility. Am J Hum Genet. 2020;106:41–57. doi: 10.1016/j.ajhg.2019.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Miyamoto T, Hasuike S, Yogev L, Maduro MR, Ishikawa M, et al. Azoospermia in patients heterozygous for a mutation in SYCP3. Lancet. 2003;362:1714–9. doi: 10.1016/S0140-6736(03)14845-3. [DOI] [PubMed] [Google Scholar]
- 81.de Vries L, Behar DM, Smirin-Yosef P, Lagovsky I, Tzur S, et al. Exome sequencing reveals SYCE1 mutation associated with autosomal recessive primary ovarian insufficiency. J Clin Endocrinol Metab. 2014;99:E2129–32. doi: 10.1210/jc.2014-1268. [DOI] [PubMed] [Google Scholar]
- 82.Maor-Sagie E, Cinnamon Y, Yaacov B, Shaag A, Goldsmidt H, et al. Deleterious mutation in SYCE1 is associated with non-obstructive azoospermia. J Assist Reprod Genet. 2015;32:887–91. doi: 10.1007/s10815-015-0445-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Pashaei M, Rahimi Bidgoli MM, Zare-Abdollahi D, Najmabadi H, Haji-Seyed-Javadi R, et al. The second mutation of SYCE1 gene associated with autosomal recessive nonobstructive azoospermia. J Assist Reprod Genet. 2020;37:451–8. doi: 10.1007/s10815-019-01660-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.de Vries FA, de Boer E, van den Bosch M, Baarends WM, Ooms M, et al. Mouse Sycp1 functions in synaptonemal complex assembly, meiotic recombination, and XY body formation. Genes Dev. 2005;19:1376–89. doi: 10.1101/gad.329705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Yang F, De La Fuente R, Leu NA, Baumann C, McLaughlin KJ, et al. Mouse SYCP2 is required for synaptonemal complex assembly and chromosomal synapsis during male meiosis. J Cell Biol. 2006;173:497–507. doi: 10.1083/jcb.200603063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Yuan L, Liu JG, Zhao J, Brundell E, Daneholt B, et al. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol Cell. 2000;5:73–83. doi: 10.1016/s1097-2765(00)80404-9. [DOI] [PubMed] [Google Scholar]
- 87.Liu H, Huang T, Li M, Li M, Zhang C, et al. SCRE serves as a unique synaptonemal complex fastener and is essential for progression of meiosis prophase I in mice. Nucleic Acids Res. 2019;47:5670–83. doi: 10.1093/nar/gkz226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Bolcun-Filas E, Hall E, Speed R, Taggart M, Grey C, et al. Mutation of the mouse Syce1 gene disrupts synapsis and suggests a link between synaptonemal complex structural components and DNA repair. PLoS Genet. 2009;5:e1000393. doi: 10.1371/journal.pgen.1000393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Bolcun-Filas E, Costa Y, Speed R, Taggart M, Benavente R, et al. SYCE2 is required for synaptonemal complex assembly, double strand break repair, and homologous recombination. J Cell Biol. 2007;176:741–7. doi: 10.1083/jcb.200610027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Schramm S, Fraune J, Naumann R, Hernandez-Hernandez A, Hoog C, et al. A novel mouse synaptonemal complex protein is essential for loading of central element proteins, recombination, and fertility. PLoS Genet. 2011;7:e1002088. doi: 10.1371/journal.pgen.1002088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Yang F, Gell K, van der Heijden GW, Eckardt S, Leu NA, et al. Meiotic failure in male mice lacking an X-linked factor. Genes Dev. 2008;22:682–91. doi: 10.1101/gad.1613608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hamer G, Wang H, Bolcun-Filas E, Cooke HJ, Benavente R, et al. Progression of meiotic recombination requires structural maturation of the central element of the synaptonemal complex. J Cell Sci. 2008;121:2445–51. doi: 10.1242/jcs.033233. [DOI] [PubMed] [Google Scholar]
- 93.Gomez HL, Felipe-Medina N, Sanchez-Martin M, Davies OR, Ramos I, et al. C14ORF39/SIX6OS1 is a constituent of the synaptonemal complex and is essential for mouse fertility. Nat Commun. 2016;7:13298. doi: 10.1038/ncomms13298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Zhang J, Fujiwara Y, Yamamoto S, Shibuya H. A meiosis-specific BRCA2 binding protein recruits recombinases to DNA double-strand breaks to ensure homologous recombination. Nat Commun. 2019;10:722. doi: 10.1038/s41467-019-08676-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Yoshida K, Kondoh G, Matsuda Y, Habu T, Nishimune Y, et al. The mouse RecA-like gene Dmc1 is required for homologous chromosome synapsis during meiosis. Mol Cell. 1998;1:707–18. doi: 10.1016/s1097-2765(00)80070-2. [DOI] [PubMed] [Google Scholar]
- 96.La Salle S, Palmer K, O’Brien M, Schimenti JC, Eppig J, et al. Spata22, a novel vertebrate-specific gene, is required for meiotic progress in mouse germ cells. Biol Reprod. 2012;86:45. doi: 10.1095/biolreprod.111.095752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Shang Y, Huang T, Liu H, Liu Y, Liang H, et al. MEIOK21: a new component of meiotic recombination bridges required for spermatogenesis. Nucleic Acids Res. 2020;48:6624–39. doi: 10.1093/nar/gkaa406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Lipkin SM, Moens PB, Wang V, Lenzi M, Shanmugarajah D, et al. Meiotic arrest and aneuploidy in MLH3-deficient mice. Nat Genet. 2002;31:385–90. doi: 10.1038/ng931. [DOI] [PubMed] [Google Scholar]
- 99.Carlosama C, Elzaiat M, Patino LC, Mateus HE, Veitia RA, et al. A homozygous donor splice-site mutation in the meiotic gene MSH4 causes primary ovarian insufficiency. Hum Mol Genet. 2017;26:3161–6. doi: 10.1093/hmg/ddx199. [DOI] [PubMed] [Google Scholar]
- 100.Edelmann W, Cohen PE, Kane M, Lau K, Morrow B, et al. Meiotic pachytene arrest in MLH1-deficient mice. Cell. 1996;85:1125–34. doi: 10.1016/s0092-8674(00)81312-4. [DOI] [PubMed] [Google Scholar]