

Abstract
Pancreatic cancer (PC) is a lethal malignancy cancer, and its mortality rates have been increasing worldwide. Diagnosis of this cancer is complicated, as it does not often present symptoms, and most patients present an irremediable tumor having a 5-year survival rate after diagnosis. Regarding treatment, many concerns have also been raised, as most tumors are found at advanced stages. At present, anticancer compounds-rich foods have been utilized to control PC. Among such bioactive molecules, flavonoid compounds have shown excellent anticancer abilities, such as quercetin, which has been used as an adjunctive or alternative drug to PC treatment by inhibitory or stimulatory biological mechanisms including autophagy, apoptosis, cell growth reduction or inhibition, EMT, oxidative stress, and enhancing sensitivity to chemotherapy agents. The recognition that this natural product has beneficial effects on cancer treatment has boosted the researchers' interest towards more extensive studies to use herbal medicine for anticancer purposes. In addition, due to the expensive cost and high rate of side effects of anticancer drugs, attempts have been made to use quercetin but also other flavonoids for preventing and treating PC. Based on related studies, it has been found that the quercetin compound has significant effect on cancerous cell lines as well as animal models. Therefore, it can be used as a supplementary drug to treat a variety of cancers, particularly pancreatic cancer. This review is aimed at discussing the therapeutic effects of quercetin by targeting the molecular signaling pathway and identifying antigrowth, cell proliferation, antioxidative stress, EMT, induction of apoptotic, and autophagic features.
1. Introduction
Pancreatic cancer (PC) is an increasingly common cancer of the gastrointestinal tract (GIT), with survival rates less than 5% at 5 years after diagnosis, and about 50% of all patients die over 6 months of diagnosis. According to estimations in the United States, PC will become the second most common cause of cancer death in the next twenty to thirty years. However, patients' prognosis with localized and respectable tumors remains poor with only 20% survival rate after surgery [1]. In addition, in line with GLOBOCAN 2018 assessments, PC which accounts for approximately 459,000 new cases and 432,000 deaths is the seventh leading cause of global cancer death [2]. In Europe, it is assumed that PC will quickly exceed breast cancer as the third cause of cancer death after colorectal and lung cancers [3].
PC is characterized as a tumor of the exocrine pancreas and ductal adenocarcinoma; however, a minor subset of patients also represents neuroendocrine tumors. Indeed, pancreatic intraepithelial neoplasia or precursor lesions are the operative factors in the acquisition of genetic shifts which trigger discernible pancreatic ductal adenocarcinoma (PDA) [4]. Unfortunately, PC symptoms do not begin up to the advanced stages of cancer and are usually vague, including nausea, vomiting, severe abdominal pain, and weight loss. Besides, previous studies revealed that type 2 diabetes, family history, obesity, and tobacco usage are the major risk factors for PC [1–5]. Therefore, studies are headed for PC prevention. A broad range of recent studies have explored the anticancer features of phytochemicals and have indicated that polyphenols, flavonoids, and flavones can be occupied against diverse types of cancers [6]. Flavonoids are secondary metabolites of plants with pharmacological activities (Table 1). Hence, fruits or vegetables, such as cocoa and coffee, are valuable sources of flavonoids [7, 8]. Based on chemical structure, oxidation degree, and unsaturation of linking chain, flavonoids are categorized into 6 main classes: isoflavonoids, flavones, flavanols, flavanones, flavonols, and anthocyanidins [9]. Quercetin and kaempferol are some of the most frequently found flavonols [10]. Quercetin (C15H10O7) is called by IUPAC (International Union of Pure and Applied Chemistry) as follows: 3,3,4,5,7-pentahydroxyflavone and 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxychromen-4-one [11]. It has been documented that quercetin offers antifungal, antioxidant, cytotoxic, hepatoprotective, and anticancer activities [12]. Specifically, both quercetin and its derivatives can prevent cancer-related diseases by regulating cellular signaling pathways. However, the anti-inflammatory and antioxidant properties of quercetin are the main factors for its activity as cell cycle inhibitors, and the apoptosis-inducing effect of quercetin has a key anticancer role [13, 14]. Noteworthy, quercetin is a general phytochemical in the regular dietary program of people worldwide since it can be widely found in daily foods, like tea, coffee, different vegetables, nuts, and fruits [15]. Quercetin and its derivatives pose biological inhibitory effects on the progression of the cancerous cell cycle; therefore, the metabolic pathways of quercetin are deemed as a significant factor in the plants' adaptive reaction. A number of recent studies have focused on the quercetin content of fruits and vegetables for its therapeutic purposes [16–19]. Additionally, as mentioned by Harwood et al. [20], commercially accessible quercetin can be consumed orally at a dose of 1 g daily, which is absorbed up to 60% and safe enough. In this sense, this review aims to discuss the anticancer properties of quercetin against PC, considering its low cost in comparison to synthetic drugs. In addition, the latest trends on quercetin features and their molecular mechanisms in cancer therapy are also summarized. Therefore, different research studies have analyzed the probable mechanisms through which quercetin exerts its antitumor effects against pancreatic cancer cells. Since there is not any review article on this subject based on our searches, we aimed to discuss the therapeutic effects of quercetin against pancreatic cancer cells for the first time.
Table 1.
A number of pharmacological activities of quercetin reported in recent studies.
| Dose | Model | Administration | Activity | Ref. |
|---|---|---|---|---|
| 25 μM | MCF-7 and MDA-MB-231 | Direct treatment | Apoptosis induction and anticancer effect | [129] |
| 20 μM | Human umbilical vein endothelial cells (HUVECs) | Direct treatment | Autophagy and promoted cell survival | [130] |
| 5.7 μM (DPPH assay) | Erythrocytes | Direct treatment | ROS and free radical scavenging activity | [131] |
| 100 mg kg−1 | C57BL/6J mice on ethanol-containing Lieber De Carli liquid diets | Gavage | Suppressed autophagic flux, decreased liver injury by ethanol consumption | [132] |
| 100 mg kg−1 | Streptococcus suis infected mice | Subcutaneous | Antimicrobial effect against Streptococcus suis | [133] |
| 100 mg kg−1 | Chronic ethanol feeding C57BL/6J mice | Oral | Decreased fat accumulation in liver (ethanol induced) | [134] |
| 30 mg kg−1 | STZ-induced diabetic rats | Intraperitoneal | Higher insulin levels, improved dyslipidemia, reduced serum blood glucose levels, decreased oxidative stress | [135] |
| 100-200 mg kg−1 | STZ-induced diabetic Wistar rats | Oral | Controlled insulin resistance, reduced blood sugar, pancreatic cells protection | [136] |
| 5-20 mg kg−1 | STZ-induced diabetic rats | Oral | Controlled body weight and blood glucose, performance in the Morris water test | [137] |
| 40 mg kg−1 | STZ-induced diabetic mice in the Morris water maze task | Oral | Enhanced the time spent by mice in the target quadrant in the Morris water maze task | [138] |
2. Pancreatic Cancer (PC)
Currently with an average 5-year survival rate, PC is estimated to be the second cause of cancer-related deaths by 2030 in the United States [21–23]. The possibility of developing PC is about 1.5% in both genders [23], despite it mainly occurs in elderly, between 70 and 80 years, mostly in unlocalized and incurable forms [21, 24]. PC often remains undetectable until it transforms into a metastatic tumor [25]. While the etiology of PC has not been completely understood, several genetic and environmental risk factors are known to increase the risk, including smoking, obesity, diets rich in animal fat, cystic fibrosis, and genetic predispositions [26]. According to Huang et al. [27], the highest incidence and mortality of PC are in countries with very high human development index or age-standardized rates (ASRs) or the countries with a higher prevalence of alcohol drinking, smoking, hypertension, physical inactivity, obesity, and high cholesterol. The highest incidence rates of PC were reported in Western Europe (ASR, 8.3), North America (ASR, 7.6), and Central and Eastern Europe (ASR, 7.5). The incidence of PC has men to women ratio of 1.4 : 1.0. More detailed information about the PC incidence and mortality based on the region and sex are presented in the main reference [27].
An earlier diagnosis would be very helpful in the successful treatment of this malignancy, despite the scarce presence of symptoms among individuals. Regarding treatment, surgery, chemotherapy, and radiotherapy are the most common therapeutic strategies applied for PC treatment. Actually, the standard course of treatment is surgery following adjuvant therapy; however, the recurrence of 70-80% of resected tumors ultimately occurs. Patients who are eligible for surgical resection comprise merely almost 10-15% of all patients with advanced PC. With the majority of patients being diagnosed at later stages, chemotherapy remains as only treatment option for PC. 5-Flourouracil (5-FU) and gemcitabine (GMC), alone or in combination with radiation, are the standard chemotherapy regimen for PC's treatment, even though the response rate is usually below 31%. GMC has some advantages over 5-FU, such as the ability to relieve most disease's symptoms and having a modest survival advantage; however, it could not extend the average survival rate much beyond 6 months, like other chemotherapeutics [28, 29]. Hence, with the limited success of current standard therapies, the search for new and effective treatment strategies and agents is urgently needed.
3. Naturally Occurring Phytochemicals for Anticancer Purposes
Various observational and prospective studies have revealed an indirect association between fruit and vegetable consumption with the occurrence of some cancers and the great potential of natural compounds to change the natural history of carcinogenesis [30–32]. Plants with some bioactive nonnutrient compounds isolated, characterized, and identified as phytochemicals have been ever more searched for their ability to treat different diseases, especially cancer [33–41]. It seems that natural products still hold out the best options to find effective novel components in the treatment of human diseases [42]. In addition, the development of scientific technologies such as genome mining, genetic engineering, and using of nanoparticles as carriers [43] improve the discovery of new drugs in cancer therapy [44]. The word ‘phytochemical' refers to plant (phyto in Greek) chemicals. Many of these phytochemicals could regulate a wide range of cellular signaling pathways which are involved in oxidative stress, growth, proliferation, differentiation, and death [37, 45–48]. For example, they exhibit antioxidant properties by affecting Nrf2-Keap1 pathway, where upon activation, Nrf2 translocate into the nucleus, binds to ARE (antioxidant response elements) or EpREs (electrophile response elements) and increases the expression of ATP-dependent drug efflux pumps, detoxification enzymes, and endogenous antioxidants [49]. These events eventually lead to the protection of cells against ROS (reactive oxygen species) [50, 51]. Phytochemicals could also suppress tumor progression and induce apoptosis in pre-neoplastic or neoplastic cells by affecting cell cycle, JAK-STAT, NF-κB, and cytochrome C signaling pathways [52, 53]. One of the phytochemicals is garcinol, a Polyisoprenylated Benzophenone that can inhibit STAT-3 pathway by suppressing the upstream kinases (c-Src, JAK1, and JAK2) in HNSCC cells. Garcinol also inhibits NF-κB activation by the suppression of TGF-β and inhibitor of IκB kinase (IKK) activation in HNSCC cells [54]. In addition, Li et al. showed that garcinol prevents the growth of human HNSCC xenograft tumors in male athymic nu/nu mice [54]. It can be concluded that garcinol has potential antitumor effects in head and neck carcinoma through the suppression of multiple proinflammatory cascades. Activator protein 1 (AP-1) as a key transcription factor in the control of several cellular processes is involved in inflammatory disorders and cancer. Several natural compounds such as kaempferide, resveratrol, apigenin, isorhamnetin, citrifolinoside A, viscolin, curcumin, and quercetin can modulate AP-1-associated signaling pathways for cancer prevention and intervention [55].
Among the plethora of biologically active phytochemicals with anticancer potential, they are chemically categorized into phenolics, carotenoids, phytosterols, organosulfur compounds, and nitrogen-containing compounds [56, 57]. Phenolics are structurally characterized with one (phenolic acids) or more (polyphenols) aromatic rings with one or more hydroxyl (OH) groups [58]. Phenolic compounds can be divided into flavonoids and nonflavonoids [59]. Flavonoids, including glycosides, aglycone, and methylated derivatives, comprise half of phenolic compounds [60]; flavonoids are subgrouped into flavones, flavanones, flavanonols, flavanols, flavonols, isoflavones, chalcones, and anthocyanidins [61, 62]. Nonflavonoids also have several subgroups which include stilbenes, phenolic acids, lignans, coumarins, and tannins [63] (Figure 1).
Figure 1.
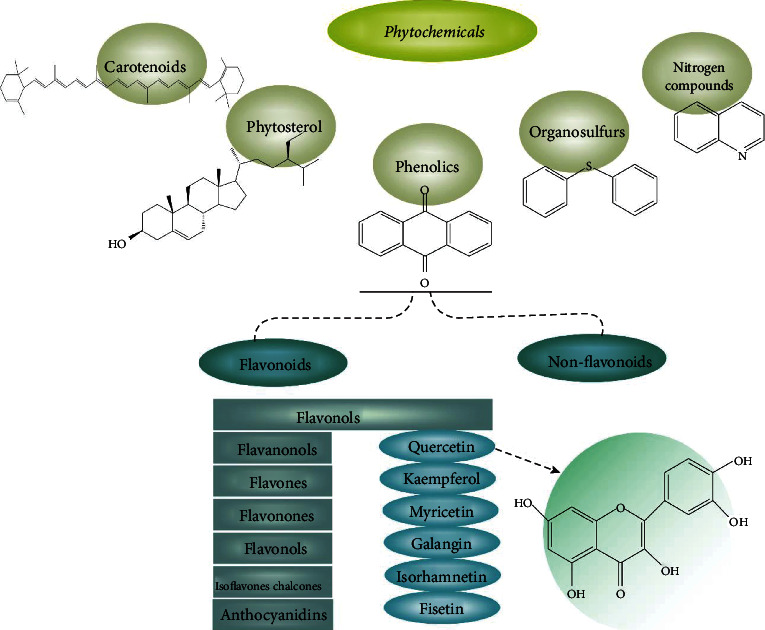
Classified phytochemicals with anticancer potential and their chemical structure.
4. Flavonoids and Anticancer Effects: Key Focus on Quercetin
Belonging to the class of polyphenolic flavonoid compounds and the subclass of flavonols, quercetin is ubiquitous in daily foods, including various plants, vegetables, nuts, seeds, fruits, tea, and red wine [64, 65]. However, fruits and plants are being studied as promising sources of quercetin [17, 66–68]. Quercetin comprises the characteristic structure of flavonoids (backbone C6-C3-C6) in which two benzene rings are bonded by a 3-carbone heterocyclic pyrone [69, 70]. Quercetin has two antioxidant pharmacophores in this structure, which allow to act as a free radical scavenging agent and join to transitional metal ions [69]. The ideal arrangement of the catechol and the OH group at C3, the position in quercetin structure, also adds to its free radical scavenging ability [69, 71]. The replacement of its various OH groups grants quercetin different biochemical and pharmacological functions [72]. It has been estimated that the average daily intake of quercetin could be about 25 mg [20]. The bioavailability of quercetin relies on its metabolic form in the food [73]. Quercetin may be found as free or aglycone state and conjugated forms, in which it interacts with several molecules, including lipids, carbohydrates, alcohols, and sulfate groups to form its derivatives, including prenylated quercetin, quercetin ethers, quercetin glycoside, and quercetin sulfate [72]. In plants, the form of quercetin is quercetin glucosides (quercetin-glucose conjugates). Quercetin glucosides undergo hydrolysis to form quercetin aglycone following the absorption in the apical membrane of the enterocytes. Then, enterocytic transferases metabolize quercetin aglycone to the glucuronidated, sulfonylated, and methylated forms [73]. These quercetin metabolites when transported to the liver undergo other conjugation processes to generate Que-3-glucuronide and quercetin-3′-sulfate [73–75]. The peak plasma concentration of quercetin varies from 3.5 to 5.0 μM once being absorbed in the form of glucosides. However, its peak plasma concentration is less than 0.33 μM when absorbed in the unconjugated form, showing less efficient absorption [76].
Quercetin has numerous benefits on human health, including anticancer, antioxidant, antidiabetic, antiulcer, anti-inflammatory, antiviral, antiallergic, antihypertension and anti-infection, cardioprotective, gastroprotective, and immune-modulatory effects [69, 77]. With the specific impact on tumor cells and without any impact on normal and nontransformed ones, quercetin has fascinated many researchers to investigate its potential as an adjuvant to suppress oxidative stress, proliferation, and metastasis [78]. Several studies showed the inhibitory impacts of quercetin against pancreatic, colorectal, prostate, lung, ovarian, nasopharyngeal, breast, and kidney cancers [79–85]. A number of recent clinical studies have scrutinized quercetin's effect on PC. In this regard, Liu et al. [86] have explored the anticancer effects and mechanistic actions of quercetin in GMC-resistant cancer cells. In this survey, BxPC-3, PANC-1 and HepG2, and Huh-7 cell lines were studied. Proliferation assays presented that quercetin had cytotoxic effects on GMC-resistant cell lines including HepG2 and PANC-1, and flow cytometric analysis specified a noteworthy proapoptotic effect on these cell lines. GMC treatment, along with quercetin, caused increased anticancer effects compared with GMC alone, and quercetin led to S phase arrest in resistant cell lines. Hoca et al. [5] investigated the effect of quercetin and resveratrol on epithelial-mesenchymal transition (EMT) of CD133+ and CD133− pancreatic cancer cells. CD133+ cells were obtained from the PANC-1 cells by the MiniMACS system. CD133+ and CD133− PANC-1 cells were treated with different concentrations of resveratrol and quercetin. Immunocytochemistry tests using antibodies such as TNF-α, ACTA-2, N-cadherin, IL-1β, and vimentin were applied for assessing the anticancer and antimetastatic properties of resveratrol and quercetin. Results revealed that the immunostaining intensity of CD133+ cells was stronger than that of CD133− cells. ACTA-2, N-cadherin, and IL-1β immunoreactivities were significantly decreased, whereas vimentin and TNF-α immunoreactivities increased in quercetin treated CD133+ cells. In addition, quercetin was more effective than resveratrol in inhibiting metastasis. Guo et al. [87] have studied the therapeutic potential of quercetin in targeting sonic hedgehog (SHH) signaling of PDA. The effects of quercetin on the apoptosis, migration, and invasion of pancreatic cancer cells (PCCs) were evaluated in PDA xenograft mouse models. According to the results, quercetin inhibited the PCC proliferation by downregulating c-Myc expression and suppressed the EMT by reducing TGF-β1 level, which inhibited the PCC migration and invasion. Quercetin treatment reduced the PDA growth and metastasis in nude mouse models by decreasing SHH activity. Additionally, SHH activated TGF-β1/Smad2/3 signaling and stimulated EMT by inducing the expression of Snail1and Zeb2 that instigated a partial reversal of quercetin-mediated inhibition of PCC migration and invasion.
4.1. Molecular Mechanisms Underlying Quercetin-Mediated Effects in Cancer
4.1.1. Effect in Autophagy and Apoptosis Induction
According to Pang et al. [88], quercetin can affect CD36 and decrease the death rate of PC by facilitating the uptake of fatty acids, improving the cell adhesion, stimulating immune response, and regulating thrombospondin-1. Furthermore, previous trends indicated that quercetin has proapoptotic activity in suppressing Bcl-2 protein and in upregulating the p53 gene; however, inhibition of Bcl-2 transcription could prevent the tumors development [12]. In an illustrative study, Serri et al. [89], have investigated the effect of GMC with biodegradable nanoparticles (NPs) loaded within quercetin on PC cell lines. The manufactured NPs decorated with hyaluronic acid (HA) and loaded with quercetin and GMC presented a developed cytotoxicity on PANC-1 and Mia-PaCa-2 cell lines when compared with the bare drugs and the NPs nondecorated with HA on the surface. The results indicated that, the NPs exposing HA may enhance the anti-inflammatory activity of Que, which led to a reduction of interleukin (IL) expression levels in cell lines and preliminarily increased with lipopolysaccharides (LPS). In another survey, Lan et al. [90] showed that quercetin accelerates cell death and chemosensitivity of human PC cells. The results showed that silencing of a receptor for advanced glycation end products (RAGE) by RAGE-specific siRNA intensified the autophagy and apoptosis through suppressing PI3K/AKT/mTOR axis in MIA Paca-2 and GMC-resistant cells (MIA Paca-2 GMCR cells). Moreover, quercetin reduced RAGE expression and facilitated the apoptosis, autophagy, and chemosensitivity to GMC in MIA Paca-2 GMCR cells, which suggests that further cytotoxicity has been achieved by the addition of quercetin in treatment with GMC. Yu et al. [91] showed that quercetin initiated inhibitory activities against PATU-8988 and PANC-1 cells and reduced the release of matrix metalloproteinase (MMP). In this study, they used STAT-3 and IL-6 activation to scrutinize the effects of quercetin treatment on cell malignancy. The MMP secretion and epithelial mesenchymal transition (EMT) stimulated the STAT-3 signaling pathway, while quercetin reversed IL-6-induced EMT and invasion. As main findings, this study showed that quercetin is an effective agent in PC treatment as it blocks the STAT-3 signaling pathway, leading to the suppression of EMT and metastasis. In addition, Nwaeburu et al. [92] have explored the effect of quercetin on miRNA expression in PC cells and concluded that quercetin treatment induced the expression of miR-200b-3p in AsPC1 cell lines, which has a crucial role in the irregular division of PDA cells by notch signaling regulation (Figure 2).
Figure 2.
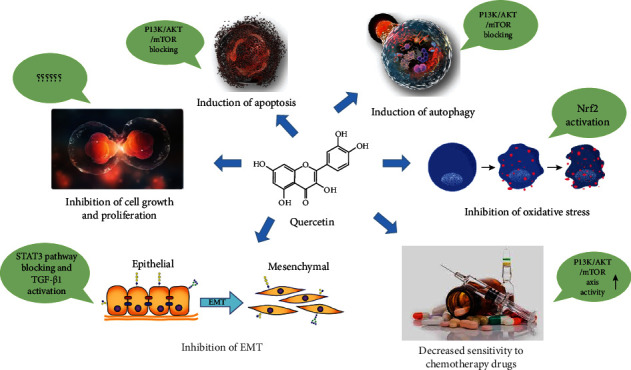
Schematic representation of the molecular structure of quercetin composition and its role in physiological conditions through signaling pathways. The flavonoid quercetin through inhibitory and stimulatory mechanisms performs functions such as inducing autophagy and apoptosis and reducing or inhibiting cell growth and proliferation, EMT, oxidative stress, and sensitivity to chemotherapy drugs.
4.1.2. Effect in Proliferation and Cell Growth
Inhibition of PC cell proliferation could signify a distinct mechanism of anticancer effects of quercetin (Table 2). In this way, Pham et al. [93] studied the effect of quercetin on dysregulated epigenetic readers, including bromodomain and extraterminal domain (BET) proteins, in in vitro and xenograft models of PC. According to the results, after treatment with BET inhibitors and quercetin the proliferation and sphere-forming ability of cancer cells was reduced, and apoptosis stimulated. In addition, quercetin diminished the nuclear protein hnRNPA1 which control mRNA translation and export of antiapoptotic proteins, in vivo and increased the BET inhibitors effects at suppressing cells proliferation and tumor growth. In another study, Nwaeburu et al. [94] explored the quercetin influence on PC cells proliferation by the activation of Notch-inhibitor Numbl as let-7c target gene. In vivo xenotransplantation of PDA cell and following IV injection of let-7c provoked a noteworthy reduction of tumor mass in the fertilized chick egg model. Immunohistochemistry analysis demonstrated that let-7c upregulated the Numbl and reserved Notch and progression markers. The findings illustrate that Que-induced let-7c declines cancer cell divisions and tumor growth.
Table 2.
Anticancer effects of quercetin against PC.
| Dose | In vitro/in vivo | Cell line | Effective mechanism | Ref. |
|---|---|---|---|---|
| 100 μM and 75 mg kg−1 | In vivo and in vitro | PANC-1 and Patu8988 | EMT suppression by reducing TGF-β1 level, inhibition of growth, invasion, and migration of cells, apoptosis of cancer cells by antagonizing TGF-β/Smad and SHH signaling pathways | [87] |
| 20 μM | In vitro | Mia-PaCa-2 and PANC-1 | Reduced IL-6 and IL-8 expressions and enhanced cytotoxicity against Mia-PaCa-2 and PANC-1 cell lines | [89] |
| 100 μM | In vitro | PANC-1 | Reduced immunoreactivities such as ACTA-2, IL-1β, and N-cadherin, increased TNF-α and vimentin, prevention of EMT | [139] |
| 20 μM and 40 mg kg−1 | In vivo and in vitro | PDAC | Improved effects of BET inhibitors at suppressing tumor development and reduced hnRNPA1 in vivo | [93] |
| 50-200 μM | In vitro | MIA Paca-2, BxPC-3, AsPC-1, HPAC and PANC-1 | Quercetin showed a RAGE silencing like effect that attenuate RAGE expression to accelerate apoptosis, autophagy, and chemosensitivity of MIA Paca-2 GEMR cells | [90] |
| 20-80 μM | In vitro | PANC-1 and PATU-8988 | Quercetin reversed IL-6-induced EMT by the stimulation of the STAT3 signaling pathway and prevented the migration | [91] |
| 50 μM | In vivo | AsPC-1 and PANC-1 | Upregulation of miR-200b-3p that promoted the Notch signaling pathway of daughter cells to turn into symmetric | [92] |
| 50 μM | In vitro | AsPC-1, CRL-4023, and PANC-1 | Notch inhibition by quercetin-induced let-7c and marker progression, upregulation of Numbl, and tumor development reduction | [94] |
| 100 nM | In vitro | CFPAC-1 and SNU-213 | Suppressed TGF-β- and VEGF-A-induced migratory activity induced at low dosages in CFPAC-1, but not in bFGF-activated SNU-213 cells | [140] |
4.1.3. Effect in Oxidative Stress
Redox homeostasis is very important for cell function and ROS have an essential role in cell signaling. However, the disturbance in the antioxidant system could lead to excessive intracellular ROS levels, such as hydroxy free radicals and H2O2 [95]. The excessive intracellular levels of ROS result in oxidative damages to many biological macromolecules which includes lipids, proteins, and genetic material, giving rise to pathological conditions, like cancer, inflammation, atherosclerosis, angiogenesis, as well as aging [96–100]. Therefore, helping cells to keep redox homeostasis is of great value, which can be provided by consuming natural nutritional components, such as Que.
With the numerous OH groups and conjugated π orbitals allowing it to donate hydrogen or electrons, and thus scavenge superoxide anion (•O2−) and H2O2. Quercetin is regarded as outstanding free-radical scavenging antioxidant [101]. Quercetin could generate the semi-quinone radical and H2O2 by reaction with •O2−, while also decreases H2O2 levels in the presence of peroxidases and keeps cells safe against H2O2 damages [64]. The semi-quinone is one of potentially harmful reactive oxidation products and undertakes a second oxidation reaction with Que, producing additional quinone (Que-Quinone; QQ) [64]. QQ is held culprit for lipid peroxidation as well as protein and DNA damages with higher affinity to react with lipids, proteins, and DNA [64, 102]. QQ with high reactivity towards thiols could arylate protein thiols, impairing several vital enzymes; however, it generates relatively stable glutathione (GSH)-oxidized adducts including 8-glutathionyl-quercetin (8-GSQ), 6-glutathionyl-quercetin (6-GSQ), and 2′-glutathionyl-quercetin (2′-GSQ) when reduced GSH exists [103, 104]; this reaction is reversible and glutathionyl-quercetin adducts could be constantly disassociated into QQ and GSH [105]. Consequently, high GSH concentrations within cells, oxidized quercetin forms GSQ by reaction with GSH, neutralizing the toxicity of QQ. Yet, oxidized quercetin reacts with protein thiols while lower concentrations of GSH exist within cells, showing the prooxidant effect of quercetin [105, 106]. Therefore, the GSH concentration within cells determines whether the antioxidant effect of quercetin could prevail over its prooxidant effect. Indeed, high levels of GSH limit quercetin cytotoxicity and permit it to show its antioxidant activity but not prooxidant activity [107]. Besides, it has been shown that quercetin induces GSH synthesis [108, 109]. Moreover, quercetin also exerts antioxidant activity by activating the nuclear factor erythroid 2-related factor 2 (Nrf2) as well as its downstream targets, which are vital for maintaining cell redox hemostasis [110, 111].
4.1.4. Effect in Epithelial-to-Mesenchymal Transition (EMT)
A physiological process, epithelial-to-mesenchymal transition (EMT), has an key function in mammalian embryonic development and cell and tissue balance; however, it has also an important role in tumorigenesis and tumor progression [112]. During EMT, epithelial cells undergo some changes including losing cellular polarity, disabling junctions between cells and adhesive connections, and obtaining penetration and migration capabilities [113, 114]. EMT can be monitored by protein markers, including E- and N-cadherin, Snail and Vimentin [114, 115]. Furthermore, MMPs closely related to EMT are recently introduced as EMT markers and as a predisposing factor for it, providing an appropriate condition for tumor infiltration and metastasis by degrading the extracellular matrix (ECM) and basement membrane (BM) nearby the tumor exterior [116, 117]. The impact of quercetin on EMT in PC cells have been investigated by a few studies. In one study, it has been shown that quercetin treatment could decrease EMT and MMP secretion in PATU-8688 PC cell line [91]. Quercetin reduced mRNA and protein expressions level of N-cadherin, Slug, Vimentin Zeb1, Twist, and Snail, indicating the potential of quercetin to reverse the EMT process; however, it increased E-cadherin expression [91]. Quercetin also inhibited MMP2 and MMP7 protein expressions [91]. Besides, it has been indicated that quercetin exerted its inhibitory impacts on EMT, invasion, and metastasis in PC cells through suppressing the STAT-3 signaling pathway [91]. Another study showed that quercetin repressed EMT by suppressing SHH and TGF-β/Smad signaling pathways, involved in promoting EMT by the induction of Zeb2 and Snail1 expressions [87]. Quercetin downregulated Vim (encoding vimentin) and Acta2 (encoding α-SMA) gene expressions, and upregulated Cdh1 (encoding E-cadherin) gene expression in PANC-1 and Patu8988 cells; upon quercetin treatment, the protein levels of type I collagen, N-cadherin, α-SMA and vimentin were reduced,; however, the protein level of E-cadherin was increased in cells [87]. Quercetin decreased TGF-β1 expression and that of EMT-TFs (EMT-inducing transcription factors) Snail1 and Zeb2 [87]. EMT-TFs (Snail1 and Zeb2) are the key downstream target of TGF-β1/Smad2/3 signaling pathway suppressing E-cadherin expression [118, 119]. Furthermore, the nuclear translocation and phosphorylation of Smad2 and Smad3 were also suppressed by quercetin [87]. It has been reported that upon activation by TGF-β1 and forming heteromeric complexes with Smad4, Smad2 and Smad3 translocate to the nucleus and induce EMT-TFs' expression [120]. It has also been indicated that quercetin may inhibit EMT in PC stem cells by suppressing the expression of N-cadherin [5]. Quercetin downregulated Twist2 expression, a protein involved in EMT, in PC stem cells [121], suggesting EMT inhibition by quercetin [122].
4.1.5. Effect in Chemo-Sensitivity
By the improved effectiveness in combination with other dietary agents, quercetin has been investigated as a promising adjuvant to increase the effectiveness of numerous chemotherapeutics [122, 123]. Lan, Chen, Kuo, Lu and Yen [90] showed that quercetin may decline cell viability, promote autophagy, and increase apoptosis by suppressing receptors for advanced glycation end products (RAGE) in GMC-resistant PC cells, with a greater impact once accompanied with GMC. The results revealed that RAGE silencing promoted GMC-induced cytotoxicity in MIA Paca-2 and MIA Paca-2 GEMR cell via the PI3K/AKT/mTOR axis [90]. As RAGE silencing, quercetin reduced the expression of RAGE, which led to cell cycle arrest, apoptosis, autophagy, and promoted GEM efficacy in MIA Paca-2 GEMR cells [90], proposing quercetin quercetin as enhancer of chemotherapy efficacy of drugs against PC. In another study, quercetin promoted tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis in TRAIL-resistant PC cells [124], and decreased cellular FLICE-like inhibitory protein (cFLIP) expression, while activated c-Jun N-terminal kinase (JNK), leading to the proteasomal degradation of cFLIP and eventually making PC cells more susceptible to TRAIL-induced apoptosis [124]. It has also been reported that quercetin decreased the viability of PC cell lines including PANC-1, MiaPaCa-2, and BxPC-3 [125, 126]. Once combined with other chemotherapeutics, such as GMC or 5-FU, quercetin could affect chemotherapy efficacy depending on cell lines applied, either to suppress proliferation of or have no impact on cancer cells [125, 126]. Borska et al. [127] indicated that quercetin induced apoptosis and suppressed cell proliferation in both daunorubicin sensitive EPP85-181P and resistant EPP85-181RDB PC cell lines. Quercetin had synergistic effects with daunorubicin in both sensitive and resistant cells [127]. They also showed that quercetin treatment could decrease P-glycoprotein expression [128].
5. Conclusion
Food consumption combined with therapeutic agents has been considered a key for the successful treatment of several diseases, including cancer. Conventional therapies like natural components besides other therapeutic methods due to their lower cost and side effects have been increasingly considered by researchers. Specifically, quercetin exerts an anticancer effect against PC cancer cells by mediating apoptosis, but recent studies have also indicated that quercetin affects various signal transduction pathways to reduce cancer progression. Quercetin suppresses the expression of N-cadherin, MMP-9, STAT-3 signaling pathways and potentially inhibits EMT, invasion, and metastasis. Quercetin enhances gemcitabine chemosensitivity in pancreatic cancer cells through the inhibitory effect on RAGE expression. Meanwhile, it has wide accessibility, efficacy and low toxicity comparing with other studied compounds, make it an appealing agent in cancer treatment. More recently, quercetin has been introduced and applied as a promising drug in the treatment of various cancers alone or in combination with other chemotherapeutic agents. Future well-designed clinical studies are needed to help the scientists to evaluate the safety and potential of quercetin against PC.
Acknowledgments
The authors acknowledge the Molecular Medicine Research Center, Bio-Medicine Institute, Tabriz University of Medical Sciences, and the Clinical Research Development Unit of Sina Educational, Research and Treatment Center, Tabriz University of Medical Sciences, Tabriz, Iran. This work was supported and funded by Tabriz University of Medical Sciences, Tabriz, Iran (grant number: 68344).
Contributor Information
Vahideh Tarhriz, Email: t.tarhriz@yahoo.com.
Hossein Ahangari, Email: ahangaryhossein.tbzmed73@gmail.com.
Javad Sharifi-Rad, Email: javad.sharifirad@gmail.com.
William C. Cho, Email: chocs@ha.org.hk.
Data Availability
The data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no competing interests.
Authors' Contributions
Parina Asgharian and Abbas Pirpour Tazehkand contributed equally to this work.
References
- 1.Mizrahi J. D., Surana R., Valle J. W., Shroff R. T. Pancreatic cancer. The Lancet . 2020;395(10242):2008–2020. doi: 10.1016/S0140-6736(20)30974-0. [DOI] [PubMed] [Google Scholar]
- 2.Bray F., Ferlay J., Soerjomataram I., Siegel R. L., Torre L. A., Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: a Cancer Journal for Clinicians . 2018;68(6):394–424. doi: 10.3322/caac.21492. [DOI] [PubMed] [Google Scholar]
- 3.Siegel R. L., Miller K. D., Jemal A. Cancer statistics, 2016. CA: a Cancer Journal for Clinicians . 2016;66(1):7–30. doi: 10.3322/caac.21332. [DOI] [PubMed] [Google Scholar]
- 4.Scarpa A., Real F. X., Luchini C. Genetic unrelatedness of co-occurring pancreatic adenocarcinomas and IPMNs challenges current views of clinical management. Gut . 2018;67(9):1561–1563. doi: 10.1136/gutjnl-2018-316151. [DOI] [PubMed] [Google Scholar]
- 5.Hoca M., Becer E., Kabadayı H., Yücecan S., Vatansever H. S. The effect of resveratrol and quercetin on epithelial-mesenchymal transition in pancreatic cancer stem cell. Nutrition and Cancer . 2020;72(7):1231–1242. doi: 10.1080/01635581.2019.1670853. [DOI] [PubMed] [Google Scholar]
- 6.Vafadar A., Shabaninejad Z., Movahedpour A., et al. Quercetin and cancer: new insights into its therapeutic effects on ovarian cancer cells. Cell & Bioscience . 2020;10:1–17. doi: 10.1186/s13578-020-00397-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nabavi S. M., Šamec D., Tomczyk M., et al. Flavonoid biosynthetic pathways in plants: versatile targets for metabolic engineering. Biotechnology Advances . 2020;38:p. 107316. doi: 10.1016/j.biotechadv.2018.11.005. [DOI] [PubMed] [Google Scholar]
- 8.Scarano A., Chieppa M., Santino A. Looking at flavonoid biodiversity in horticultural crops: a colored mine with nutritional benefits. Plants . 2018;7(4):p. 98. doi: 10.3390/plants7040098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Durazzo A., Lucarini M., Souto E. B., et al. Polyphenols: a concise overview on the chemistry, occurrence, and human health. Phytotherapy Research . 2019;33(9):2221–2243. doi: 10.1002/ptr.6419. [DOI] [PubMed] [Google Scholar]
- 10.Kopustinskiene D. M., Jakstas V., Savickas A., Bernatoniene J. Flavonoids as anticancer agents. Nutrients . 2020;12(2):p. 457. doi: 10.3390/nu12020457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dabeek W. M., Marra M. V. Dietary quercetin and kaempferol: bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients . 2019;11(10):p. 2288. doi: 10.3390/nu11102288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Davoodvandi A., Shabani Varkani M., Clark C. C., Jafarnejad S. Quercetin as an anticancer agent: focus on esophageal cancer. Journal of Food Biochemistry . 2020;44(9, article e13374) doi: 10.1111/jfbc.13374. [DOI] [PubMed] [Google Scholar]
- 13.Salehi B., Machin L., Monzote L., et al. Therapeutic potential of quercetin: new insights and perspectives for human health. ACS Omega . 2020;5(20):11849–11872. doi: 10.1021/acsomega.0c01818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Soofiyani S. R., Hosseini K., Forouhandeh H., et al. Quercetin as a novel therapeutic approach for lymphoma. Oxidative Medicine and Cellular Longevity . 2021;2021:15. doi: 10.1155/2021/3157867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shafabakhsh R., Asemi Z. Quercetin: a natural compound for ovarian cancer treatment. Journal of ovarian research . 2019;12(1):1–9. doi: 10.1186/s13048-019-0530-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bhagat S., Rathore M., Kachhwaha S., Sharma H. K. Phytochemical screening, determination of Total phenol content, Total flavonoid content and quantitative estimation of Rutin and quercetin using RP-HPLC in the fruits of Capparis decidua (Forsk.) Edgew. Ind. J. Pure app. Biosci . 2021;9:254–261. [Google Scholar]
- 17.Zeng Y., Li Y., Yang J., et al. Therapeutic role of functional components in alliums for preventive chronic disease in human being. Evidence-based Complementary and Alternative Medicine . 2017;2017:13. doi: 10.1155/2017/9402849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Górecki M., Hallmann E. The antioxidant content of coffee and its in vitro activity as an effect of its production method and roasting and brewing time. Antioxidants . 2020;9(4):p. 308. doi: 10.3390/antiox9040308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Amoussa A. M. O., Zhang L., Lagnika C., et al. Effects of preheating and drying methods on pyridoxine, phenolic compounds, ginkgolic acids, and antioxidant capacity of Ginkgo biloba nuts. Journal of Food Science . 2021;86(9):4197–4208. doi: 10.1111/1750-3841.15864. [DOI] [PubMed] [Google Scholar]
- 20.Harwood M., Danielewska-Nikiel B., Borzelleca J., Flamm G., Williams G., Lines T. A critical review of the data related to the safety of quercetin and lack of evidence of _in vivo_ toxicity, including lack of genotoxic/carcinogenic properties. Food and Chemical Toxicology . 2007;45(11):2179–2205. doi: 10.1016/j.fct.2007.05.015. [DOI] [PubMed] [Google Scholar]
- 21.Mateos R. S., Conlon K. C. Pancreatic cancer. Surgery (Oxford) . 2016;34(6):282–291. doi: 10.1016/j.mpsur.2016.03.011. [DOI] [Google Scholar]
- 22.DeSantis C. E., Ma J., Goding Sauer A., Newman L. A., Jemal A. Breast cancer statistics, 2017, racial disparity in mortality by state. CA: a Cancer Journal for Clinicians . 2017;67(6):439–448. doi: 10.3322/caac.21412. [DOI] [PubMed] [Google Scholar]
- 23.Rahib L., Smith B. D., Aizenberg R., Rosenzweig A. B., Fleshman J. M., Matrisian L. M. Projecting cancer incidence and deaths to 2030: the unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Research . 2014;74(11):2913–2921. doi: 10.1158/0008-5472.CAN-14-0155. [DOI] [PubMed] [Google Scholar]
- 24.Hidalgo M. Pancreatic cancer. New England Journal of Medicine . 2010;362(17):1605–1617. doi: 10.1056/NEJMra0901557. [DOI] [PubMed] [Google Scholar]
- 25.Maisonneuve P. Epidemiology and burden of pancreatic cancer. La Presse Médicale . 2019;48(3):e113–e123. doi: 10.1016/j.lpm.2019.02.030. [DOI] [PubMed] [Google Scholar]
- 26.Ramakrishnan P., Loh W. M., Gopinath S. C., et al. Selective phytochemicals targeting pancreatic stellate cells as new anti- fibrotic agents for chronic pancreatitis and pancreatic cancer. Acta Pharmaceutica Sinica B . 2020;10(3):399–413. doi: 10.1016/j.apsb.2019.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Huang J., Lok V., Ngai C. H., et al. Worldwide burden of, risk factors for, and trends in pancreatic cancer. Gastroenterology . 2021;160(3):744–754. doi: 10.1053/j.gastro.2020.10.007. [DOI] [PubMed] [Google Scholar]
- 28.Shore S., Raraty M., Ghaneh P., Neoptolemos J. Chemotherapy for pancreatic cancer. Alimentary Pharmacology & Therapeutics . 2003;18(11-12):1049–1069. doi: 10.1111/j.1365-2036.2003.01781.x. [DOI] [PubMed] [Google Scholar]
- 29.Boreddy S. R., Srivastava S. K. Pancreatic cancer chemoprevention by phytochemicals. Cancer Letters . 2013;334(1):86–94. doi: 10.1016/j.canlet.2012.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Manson M. M. Cancer prevention - the potential for diet to modulate molecular signalling. Trends in Molecular Medicine . 2003;9(1):11–18. doi: 10.1016/S1471-4914(02)00002-3. [DOI] [PubMed] [Google Scholar]
- 31.Knekt P., Kumpulainen J., Järvinen R., et al. Flavonoid intake and risk of chronic diseases. The American Journal of Clinical Nutrition . 2002;76(3):560–568. doi: 10.1093/ajcn/76.3.560. [DOI] [PubMed] [Google Scholar]
- 32.Bagheri Y., Barati A., Nouraei S., et al. Comparative study of gavage and intraperitoneal administration of gamma-oryzanol in alleviation/attenuation in a rat animal model of renal ischemia/reperfusion-induced injury. Iranian Journal of Basic Medical Sciences . 2021;24:p. 175. doi: 10.22038/IJBMS.2020.51276.11642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Temple N. J., Wilson T., Jacobs D. R., Jr. Nutritional Health: Strategies for Disease Prevention . Springer Science & Business Media; 2012. [Google Scholar]
- 34.Liu R. H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. The American Journal of Clinical Nutrition . 2003;78(3):517S–520S. doi: 10.1093/ajcn/78.3.517S. [DOI] [PubMed] [Google Scholar]
- 35.Pezzuto J. M. Plant-derived anticancer agents. Biochemical Pharmacology . 1997;53(2):121–133. doi: 10.1016/S0006-2952(96)00654-5. [DOI] [PubMed] [Google Scholar]
- 36.Aggarwal B. B., Kunnumakkara A. B., Harikumar K. B., Tharakan S. T., Sung B., Anand P. Potential of spice-derived phytochemicals for cancer prevention. Planta Medica . 2008;74(13):1560–1569. doi: 10.1055/s-2008-1074578. [DOI] [PubMed] [Google Scholar]
- 37.Surh Y.-J. Cancer chemoprevention with dietary phytochemicals. Nature Reviews Cancer . 2003;3(10):768–780. doi: 10.1038/nrc1189. [DOI] [PubMed] [Google Scholar]
- 38.Mans D. R., Da Rocha A. B., Schwartsmann G. Anti-cancer drug discovery and development in Brazil: targeted plant collection as a rational strategy to acquire candidate anti-cancer compounds. The Oncologist . 2000;5(3):185–198. doi: 10.1634/theoncologist.5-3-185. [DOI] [PubMed] [Google Scholar]
- 39.Wang J., Zhu R., Sun D., et al. Intracellular uptake of curcumin-loaded solid lipid nanoparticles exhibit anti-inflammatory activities superior to those of curcumin through the NF-κB signaling pathway. Journal of Biomedical Nanotechnology . 2015;11(3):403–415. doi: 10.1166/jbn.2015.1925. [DOI] [PubMed] [Google Scholar]
- 40.Fazel Nabavi S., Thiagarajan R., Rastrelli L., et al. Curcumin: a natural product for diabetes and its complications. Current Topics in Medicinal Chemistry . 2015;15(23):2445–2455. doi: 10.2174/1568026615666150619142519. [DOI] [PubMed] [Google Scholar]
- 41.Bagheri Y., Fathi E., Maghoul A., et al. Effects of Achillea tenuifolia Lam.hydro-alcoholic extract on anxiety-like behavior and reproductive parameters in rat model of chronic restraint stress. Human & Experimental Toxicology . 2021;1(article 09603271211026723) doi: 10.1177/09603271211026723. [DOI] [PubMed] [Google Scholar]
- 42.Newman D. J., Cragg G. M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. Journal of Natural Products . 2020;83(3):770–803. doi: 10.1021/acs.jnatprod.9b01285. [DOI] [PubMed] [Google Scholar]
- 43.Kashyap D., Tuli H. S., Yerer M. B., et al. Seminars in cancer biology . Academic Press; 2021. Natural product-based nanoformulations for cancer therapy: Opportunities and challenges; pp. 5–23. [DOI] [PubMed] [Google Scholar]
- 44.Atanasov A. G., Zotchev S. B., Dirsch V. M., Supuran C. T. Natural products in drug discovery: advances and opportunities. Nature Reviews Drug Discovery . 2021;20(3):200–216. doi: 10.1038/s41573-020-00114-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lee K. W., Bode A. M., Dong Z. Molecular targets of phytochemicals for cancer prevention. Nature Reviews Cancer . 2011;11(3):211–218. doi: 10.1038/nrc3017. [DOI] [PubMed] [Google Scholar]
- 46.Howes M.-J. R., Simmonds M. S. The role of phytochemicals as micronutrients in health and disease. Current Opinion in Clinical Nutrition & Metabolic Care . 2014;17:558–566. doi: 10.1097/MCO.0000000000000115. [DOI] [PubMed] [Google Scholar]
- 47.Vidya Priyadarsini R., Nagini S. Cancer chemoprevention by dietary phytochemicals: promises and pitfalls. Current Pharmaceutical Biotechnology . 2012;13(1):125–136. doi: 10.2174/138920112798868610. [DOI] [PubMed] [Google Scholar]
- 48.Rahimi M., Bagheri A., Bagheri Y., et al. Renoprotective effects of prazosin on ischemia-reperfusion injury in rats. Human & Experimental Toxicology . 2021;40(8):1263–1273. doi: 10.1177/0960327121993224. [DOI] [PubMed] [Google Scholar]
- 49.Tazehkand A. P., Salehi R., Velaei K., Samadi N. The potential impact of trigonelline loaded micelles on Nrf2 suppression to overcome oxaliplatin resistance in colon cancer cells. Molecular Biology Reports . 2020;47(8):5817–5829. doi: 10.1007/s11033-020-05650-w. [DOI] [PubMed] [Google Scholar]
- 50.Lee J. H., Khor T. O., Shu L., Su Z.-Y., Fuentes F., Kong A.-N. T. Dietary phytochemicals and cancer prevention: Nrf2 signaling, epigenetics, and cell death mechanisms in blocking cancer initiation and progression. Pharmacology & Therapeutics . 2013;137(2):153–171. doi: 10.1016/j.pharmthera.2012.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kumar H., Kim I.-S., More S. V., Kim B.-W., Choi D.-K. Natural product-derived pharmacological modulators of Nrf2/ARE pathway for chronic diseases. Natural Product Reports . 2014;31(1):109–139. doi: 10.1039/C3NP70065H. [DOI] [PubMed] [Google Scholar]
- 52.Seeram N. P., Adams L. S., Zhang Y., et al. Blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry extracts inhibit growth and stimulate apoptosis of human cancer cells in vitro. Journal of Agricultural and Food Chemistry . 2006;54(25):9329–9339. doi: 10.1021/jf061750g. [DOI] [PubMed] [Google Scholar]
- 53.Klein E. A., Thompson I. M., Tangen C. M., et al. Vitamin E and the risk of prostate cancer: the selenium and vitamin E cancer prevention trial (SELECT) JAMA . 2011;306(14):1549–1556. doi: 10.1001/jama.2011.1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Li F., Shanmugam M. K., Chen L., et al. Garcinol, a polyisoprenylated benzophenone modulates multiple proinflammatory signaling cascades leading to the suppression of growth and survival of head and neck carcinoma. Cancer Prevention Research . 2013;6(8):843–854. doi: 10.1158/1940-6207.CAPR-13-0070. [DOI] [PubMed] [Google Scholar]
- 55.Tewari D., Nabavi S. F., Nabavi S. M., et al. Targeting activator protein 1 signaling pathway by bioactive natural agents: possible therapeutic strategy for cancer prevention and intervention. Pharmacological Research . 2018;128:366–375. doi: 10.1016/j.phrs.2017.09.014. [DOI] [PubMed] [Google Scholar]
- 56.Mollakhalili Meybodi N., Mortazavian A. M., Bahadori Monfared A., Sohrabvandi S., Aghaei Meybodi F. Phytochemicals in cancer prevention: a review of the evidence. Iranian Journal of Cancer Prevention . 2017;In Press doi: 10.17795/ijcp-7219. [DOI] [Google Scholar]
- 57.Lee T.-Y., Tseng Y.-H. The potential of phytochemicals in oral cancer prevention and therapy: a review of the evidence. Biomolecules . 2020;10(8):p. 1150. doi: 10.3390/biom10081150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Arora I., Sharma M., Tollefsbol T. O. Combinatorial epigenetics impact of polyphenols and phytochemicals in cancer prevention and therapy. International Journal of Molecular Sciences . 2019;20(18):p. 4567. doi: 10.3390/ijms20184567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Frond A. D., Iuhas C. I., Stirbu I., et al. Phytochemical characterization of five edible purple-reddish vegetables: anthocyanins, flavonoids, and phenolic acid derivatives. Molecules . 2019;24(8):p. 1536. doi: 10.3390/molecules24081536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tungmunnithum D., Thongboonyou A., Pholboon A., Yangsabai A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: an overview. Medicine . 2018;5(3):p. 93. doi: 10.3390/medicines5030093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Shahidi F., Yeo J. Bioactivities of phenolics by focusing on suppression of chronic diseases: a review. International Journal of Molecular Sciences . 2018;19(6):p. 1573. doi: 10.3390/ijms19061573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Liskova A., Koklesova L., Samec M., et al. Flavonoids in cancer metastasis. Cancers . 2020;12(6):p. 1498. doi: 10.3390/cancers12061498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Costea T., Hudiță A., Ciolac O.-A., et al. Chemoprevention of colorectal cancer by dietary compounds. International Journal of Molecular Sciences . 2018;19(12):p. 3787. doi: 10.3390/ijms19123787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Metodiewa D., Jaiswal A. K., Cenas N., Dickancaité E., Segura-Aguilar J. Quercetin may act as a cytotoxic prooxidant after its metabolic activation to semiquinone and quinoidal product. Free Radical Biology and Medicine . 1999;26(1-2):107–116. doi: 10.1016/S0891-5849(98)00167-1. [DOI] [PubMed] [Google Scholar]
- 65.Hollman P. C., van Trijp J. M. P., Buysman M. N., et al. Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Letters . 1997;418(1-2):152–156. doi: 10.1016/S0014-5793(97)01367-7. [DOI] [PubMed] [Google Scholar]
- 66.Grzelak-Błaszczyk K., Milala J., Kołodziejczyk K., et al. Protocatechuic acid and quercetin glucosides in onions attenuate changes induced by high fat diet in rats. Food & Function . 2020;11(4):3585–3597. doi: 10.1039/C9FO02633A. [DOI] [PubMed] [Google Scholar]
- 67.Rolnik A., Żuchowski J., Stochmal A., Olas B. Quercetin and kaempferol derivatives isolated from aerial parts of _Lens culinaris_ Medik as modulators of blood platelet functions. Industrial Crops and Products . 2020;152:p. 112536. doi: 10.1016/j.indcrop.2020.112536. [DOI] [Google Scholar]
- 68.Bhatt S. K., Javagal R. M., Nanjarajurs M. S., Eligar S. M. In vitro anti-inflammatory property of a Quercetin-3-O-diglucoside-7-O-glucoside characterized from fresh leaves ofTrigonella foenum-graecumL. International Journal of Food Properties . 2021;24(1):1438–1452. doi: 10.1080/10942912.2021.1969946. [DOI] [Google Scholar]
- 69.Boots A. W., Haenen G. R., Bast A. Health effects of quercetin: from antioxidant to nutraceutical. European Journal of Pharmacology . 2008;585(2-3):325–337. doi: 10.1016/j.ejphar.2008.03.008. [DOI] [PubMed] [Google Scholar]
- 70.Khiari Z., Makris D. P. Stability and transformation of major flavonols in onion (Allium cepa) solid wastes. Journal of Food Science and Technology . 2012;49(4):489–494. doi: 10.1007/s13197-010-0201-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.D'Andrea G. Quercetin: a flavonol with multifaceted therapeutic applications? Fitoterapia . 2015;106:256–271. doi: 10.1016/j.fitote.2015.09.018. [DOI] [PubMed] [Google Scholar]
- 72.Panche A., Diwan A., Chandra S. Flavonoids: an overview. Journal of nutritional science . 2016;5 doi: 10.1017/jns.2016.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.O’Leary K. A., Day A. J., Needs P. W., Mellon F. A., O’Brien N. M., Williamson G. Metabolism of quercetin-7- and quercetin-3-glucuronides by an _in vitro_ hepatic model: the role of human β-glucuronidase, sulfotransferase, catechol- _O_ -methyltransferase and multi-resistant protein 2 (MRP2) in flavonoid metabolism. Biochemical Pharmacology . 2003;65(3):479–491. doi: 10.1016/S0006-2952(02)01510-1. [DOI] [PubMed] [Google Scholar]
- 74.Day A. J., Mellon F., Barron D., Sarrazin G., Morgan M. R., Williamson G. Human metabolism of dietary flavonoids: identification of plasma metabolites of quercetin. Free Radical Research . 2001;35(6):941–952. doi: 10.1080/10715760100301441. [DOI] [PubMed] [Google Scholar]
- 75.O'Leary K. A., Day A. J., Needs P. W., Sly W. S., O'Brien N. M., Williamson G. Flavonoid glucuronides are substrates for human liver β-glucuronidase. FEBS Letters . 2001;503(1):103–106. doi: 10.1016/S0014-5793(01)02684-9. [DOI] [PubMed] [Google Scholar]
- 76.Manach C., Williamson G., Morand C., Scalbert A., Rémésy C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. The American Journal of Clinical Nutrition . 2005;81(1):230S–242S. doi: 10.1093/ajcn/81.1.230S. [DOI] [PubMed] [Google Scholar]
- 77.Lakhanpal P., Rai D. K. Quercetin: a versatile flavonoid. Internet Journal of Medical Update . 2007;2(2):22–37. doi: 10.4314/ijmu.v2i2.39851. [DOI] [Google Scholar]
- 78.Lugli E., Ferraresi R., Roat E., et al. Quercetin inhibits lymphocyte activation and proliferation without inducing apoptosis in peripheral mononuclear cells. Leukemia Research . 2009;33(1):140–150. doi: 10.1016/j.leukres.2008.07.025. [DOI] [PubMed] [Google Scholar]
- 79.Li X., Zhou N., Wang J., et al. Quercetin suppresses breast cancer stem cells (CD44+/CD24−) by inhibiting the PI3K/Akt/mTOR-signaling pathway. Life Sciences . 2018;196:56–62. doi: 10.1016/j.lfs.2018.01.014. [DOI] [PubMed] [Google Scholar]
- 80.Baby B., Antony P., Vijayan R. Interactions of quercetin with receptor tyrosine kinases associated with human lung carcinoma. Natural Product Research . 2018;32(24):2928–2931. doi: 10.1080/14786419.2017.1385015. [DOI] [PubMed] [Google Scholar]
- 81.Huang D.-Y., Dai Z.-R., Li W.-M., Wang R.-G., Yang S.-M. Inhibition of EGF expression and NF-κB activity by treatment with quercetin leads to suppression of angiogenesis in nasopharyngeal carcinoma. Saudi journal of biological sciences . 2018;25(4):826–831. doi: 10.1016/j.sjbs.2016.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Polukonova N. V., Navolokin N. A., Bucharskaya A. B., et al. The apoptotic activity of flavonoid-containing Gratiola officinalis extract in cell cultures of human kidney cancer. Russian Open Medical Journal . 2018;7(4) doi: 10.15275/rusomj.2018.0402. [DOI] [Google Scholar]
- 83.Yang Y., Wang T., Chen D., et al. Quercetin preferentially induces apoptosis in KRAS-mutant colorectal cancer cells via JNK signaling pathways. Cell Biology International . 2019;43(2):117–124. doi: 10.1002/cbin.11055. [DOI] [PubMed] [Google Scholar]
- 84.Lee J., Han S.-I., Yun J.-H., Kim J. H. Quercetin 3-O-glucoside suppresses epidermal growth factor–induced migration by inhibiting EGFR signaling in pancreatic cancer cells. Tumor Biology . 2015;36(12):9385–9393. doi: 10.1007/s13277-015-3682-x. [DOI] [PubMed] [Google Scholar]
- 85.Liu Y., Gong W., Yang Z., et al. Quercetin induces protective autophagy and apoptosis through ER stress via the p-STAT3/Bcl-2 axis in ovarian cancer. Apoptosis . 2017;22(4):544–557. doi: 10.1007/s10495-016-1334-2. [DOI] [PubMed] [Google Scholar]
- 86.Liu Z. J., Xu W., Han J., et al. Quercetin induces apoptosis and enhances gemcitabine therapeutic efficacy against gemcitabine-resistant cancer cells. Anti-Cancer Drugs . 2020;31(7):684–692. doi: 10.1097/CAD.0000000000000933. [DOI] [PubMed] [Google Scholar]
- 87.Guo Y., Tong Y., Zhu H., et al. Quercetin suppresses pancreatic ductal adenocarcinoma progression via inhibition of SHH and TGF-β/Smad signaling pathways. Cell Biology and Toxicology . 2021;37(3):479–496. doi: 10.1007/s10565-020-09562-0. [DOI] [PubMed] [Google Scholar]
- 88.Pang B., Xu X., Lu Y., et al. Prediction of new targets and mechanisms for quercetin in the treatment of pancreatic cancer, colon cancer, and rectal cancer. Food & Function . 2019;10(9):5339–5349. doi: 10.1039/C9FO01168D. [DOI] [PubMed] [Google Scholar]
- 89.Serri C., Quagliariello V., Iaffaioli R. V., et al. Combination therapy for the treatment of pancreatic cancer through hyaluronic acid-decorated nanoparticles loaded with quercetin and gemcitabine: a preliminary in vitro study. Journal of Cellular Physiology . 2019;234(4):4959–4969. doi: 10.1002/jcp.27297. [DOI] [PubMed] [Google Scholar]
- 90.Lan C.-Y., Chen S.-Y., Kuo C.-W., Lu C.-C., Yen G.-C. Quercetin facilitates cell death and chemosensitivity through RAGE/PI3K/AKT/mTOR axis in human pancreatic cancer cells. Journal of Food and Drug Analysis . 2019;27(4):887–896. doi: 10.1016/j.jfda.2019.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Yu D., Ye T., Xiang Y., et al. Quercetin inhibits epithelial–mesenchymal transition, decreases invasiveness and metastasis, and reverses IL-6 induced epithelial–mesenchymal transition, expression of MMP by inhibiting STAT3 signaling in pancreatic cancer cells. Oncotargets and Therapy . 2017;Volume 10:4719–4729. doi: 10.2147/OTT.S136840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Nwaeburu C. C., Abukiwan A., Zhao Z., Herr I. Quercetin-induced miR-200b-3p regulates the mode of self-renewing divisions in pancreatic cancer. Molecular Cancer . 2017;16:1–10. doi: 10.1186/s12943-017-0589-8. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 93.Pham T. N., Stempel S., Shields M. A., et al. Quercetin enhances the anti-tumor effects of BET inhibitors by suppressing hnRNPA1. International Journal of Molecular Sciences . 2019;20(17):p. 4293. doi: 10.3390/ijms20174293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Nwaeburu C. C., Bauer N., Zhao Z., et al. Up-regulation of microRNA let-7c by quercetin inhibits pancreatic cancer progression by activation of Numbl. Oncotarget . 2016;7(36):58367–58380. doi: 10.18632/oncotarget.11122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Wang X., Michaelis E. K. Selective neuronal vulnerability to oxidative stress in the brain. Frontiers in Aging Neuroscience . 2010;2:p. 12. doi: 10.3389/fnagi.2010.00012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Buonocore G., Perrone S., Tataranno M. L. Seminars in Fetal and Neonatal Medicine . WB Saunders; 2010. Oxygen toxicity: chemistry and biology of reactive oxygen species; pp. 186–190. [DOI] [PubMed] [Google Scholar]
- 97.Cash T. P., Pan Y., Simon M. C. Reactive oxygen species and cellular oxygen sensing. Free Radical Biology and Medicine . 2007;43(9):1219–1225. doi: 10.1016/j.freeradbiomed.2007.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Gibellini L., Pinti M., Nasi M., et al. Interfering with ROS metabolism in cancer cells: the potential role of quercetin. Cancers . 2010;2(2):1288–1311. doi: 10.3390/cancers2021288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Fathi E., Farahzadi R., Javanmardi S., Vietor I. L-carnitine Extends the Telomere Length of the Cardiac Differentiated CD117+- Expressing Stem Cells. Tissue and Cell . 2020;67:p. 101429. doi: 10.1016/j.tice.2020.101429. [DOI] [PubMed] [Google Scholar]
- 100.Fathi E., Sanaat Z., Farahzadi R. Mesenchymal stem cells in acute myeloid leukemia: a focus on mechanisms involved and therapeutic concepts. Blood research . 2019;54(3):165–174. doi: 10.5045/br.2019.54.3.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Heijnen C., Haenen G., Van Acker F., Van der Vijgh W., Bast A. Flavonoids as peroxynitrite scavengers: the role of the hydroxyl groups. Toxicology In Vitro . 2001;15(1):3–6. doi: 10.1016/S0887-2333(00)00053-9. [DOI] [PubMed] [Google Scholar]
- 102.Yoshino M., Haneda M., Naruse M., Murakami K. Prooxidant activity of flavonoids: copper-dependent strand breaks and the formation of 8-hydroxy-2’-deoxyguanosine in DNA. Molecular Genetics and Metabolism . 1999;68(4):468–472. doi: 10.1006/mgme.1999.2901. [DOI] [PubMed] [Google Scholar]
- 103.Boots A. W., Kubben N., Haenen G. R., Bast A. Oxidized quercetin reacts with thiols rather than with ascorbate: implication for quercetin supplementation. Biochemical and Biophysical Research Communications . 2003;308(3):560–565. doi: 10.1016/S0006-291X(03)01438-4. [DOI] [PubMed] [Google Scholar]
- 104.Spencer J. P., Kuhnle G. G., Williams R. J., Rice-Evans C. Intracellular metabolism and bioactivity of quercetin and its in vivo metabolites. Biochemical Journal . 2003;372(1):173–181. doi: 10.1042/bj20021972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Boots A. W., Balk J. M., Bast A., Haenen G. R. The reversibility of the glutathionyl-quercetin adduct spreads oxidized quercetin-induced toxicity. Biochemical and Biophysical Research Communications . 2005;338(2):923–929. doi: 10.1016/j.bbrc.2005.10.031. [DOI] [PubMed] [Google Scholar]
- 106.Boots A. W., Li H., Schins R. P., et al. The quercetin paradox. Toxicology and Applied Pharmacology . 2007;222(1):89–96. doi: 10.1016/j.taap.2007.04.004. [DOI] [PubMed] [Google Scholar]
- 107.Cossarizza A., Ferraresi R., Troiano L., et al. Simultaneous analysis of reactive oxygen species and reduced glutathione content in living cells by polychromatic flow cytometry. Nature Protocols . 2009;4:1790–1797. doi: 10.1038/nprot.2009.189. [DOI] [PubMed] [Google Scholar]
- 108.Granado-Serrano A. B., Martín M. A., Bravo L., Goya L., Ramos S. Quercetin modulates Nrf2 and glutathione-related defenses in HepG2 cells: involvement of p38. Chemico-Biological Interactions . 2012;195(2):154–164. doi: 10.1016/j.cbi.2011.12.005. [DOI] [PubMed] [Google Scholar]
- 109.Kobori M., Takahashi Y., Akimoto Y., et al. Chronic high intake of quercetin reduces oxidative stress and induces expression of the antioxidant enzymes in the liver and visceral adipose tissues in mice. Journal of Functional Foods . 2015;15:551–560. doi: 10.1016/j.jff.2015.04.006. [DOI] [Google Scholar]
- 110.Jin Y., Huang Z. L., Li L., et al. Quercetin attenuates toosendanin-induced hepatotoxicity through inducing the Nrf2/GCL/GSH antioxidant signaling pathway. Acta Pharmacologica Sinica . 2019;40(1):75–85. doi: 10.1038/s41401-018-0024-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Hejazian S. M., Khatibi S. M. H., Barzegari A., et al. Nrf-2 as a therapeutic target in acute kidney injury. Life Sciences . 2021;264:p. 118581. doi: 10.1016/j.lfs.2020.118581. [DOI] [PubMed] [Google Scholar]
- 112.Nieto M. A., Huang R. Y.-J., Jackson R. A., Thiery J. P. EMT: 2016. Cell . 2016;166(1):21–45. doi: 10.1016/j.cell.2016.06.028. [DOI] [PubMed] [Google Scholar]
- 113.Thiery J. P. Epithelial-mesenchymal transitions in tumour progression. Nature Reviews Cancer . 2002;2(6):442–454. doi: 10.1038/nrc822. [DOI] [PubMed] [Google Scholar]
- 114.Agiostratidou G., Hulit J., Phillips G. R., Hazan R. B. Differential cadherin expression: potential markers for epithelial to mesenchymal transformation during tumor progression. Journal of Mammary Gland Biology and Neoplasia . 2007;12(2-3):127–133. doi: 10.1007/s10911-007-9044-6. [DOI] [PubMed] [Google Scholar]
- 115.Wong I. Y., Javaid S., Wong E. A., et al. Collective and individual migration following the epithelial-mesenchymal transition. Nature Materials . 2014;13(11):1063–1071. doi: 10.1038/nmat4062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Orlichenko L. S., Radisky D. C. Matrix metalloproteinases stimulate epithelial-mesenchymal transition during tumor development. Clinical & Experimental Metastasis . 2008;25(6):593–600. doi: 10.1007/s10585-008-9143-9. [DOI] [PubMed] [Google Scholar]
- 117.Nagase H., Visse R., Murphy G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovascular Research . 2006;69:562–573. doi: 10.1016/j.cardiores.2005.12.002. [DOI] [PubMed] [Google Scholar]
- 118.Thiery J. P., Acloque H., Huang R. Y., Nieto M. A. Epithelial-mesenchymal transitions in development and disease. Cell . 2009;139(5):871–890. doi: 10.1016/j.cell.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 119.Zheng X., Carstens J. L., Kim J., et al. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature . 2015;527(7579):525–530. doi: 10.1038/nature16064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Xu Y., Chang R., Peng Z., et al. Loss of polarity protein AF6 promotes pancreatic cancer metastasis by inducing snail expression. Nature Communications . 2015;6:1–12. doi: 10.1038/ncomms8184. [DOI] [PubMed] [Google Scholar]
- 121.Vesuna F., Lisok A., Kimble B., Raman V. Twist modulates breast cancer stem cells by transcriptional regulation of CD24 expression. Neoplasia . 2009;11(12):1318–1328. doi: 10.1593/neo.91084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Zhou W., Kallifatidis G., Baumann B., et al. Dietary polyphenol quercetin targets pancreatic cancer stem cells. International Journal of Oncology . 2010;37(3):551–561. doi: 10.3892/ijo_00000704. [DOI] [PubMed] [Google Scholar]
- 123.Appari M., Babu K. R., Kaczorowski A., Gross W., Herr I. Sulforaphane, quercetin and catechins complement each other in elimination of advanced pancreatic cancer by miR-let-7 induction and K-ras inhibition. International Journal of Oncology . 2014;45(4):1391–1400. doi: 10.3892/ijo.2014.2539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Kim J. H., Kim M. J., Choi K.-C., Son J. Quercetin sensitizes pancreatic cancer cells to TRAIL-induced apoptosis through JNK-mediated cFLIP turnover. The International Journal of Biochemistry & Cell Biology . 2016;78:327–334. doi: 10.1016/j.biocel.2016.07.033. [DOI] [PubMed] [Google Scholar]
- 125.Azimi H., Khakshur A. A., Abdollahi M., Rahimi R. Potential new pharmacological agents derived from medicinal plants for the treatment of pancreatic cancer. Pancreas . 2015;44(1):11–15. doi: 10.1097/MPA.0000000000000175. [DOI] [PubMed] [Google Scholar]
- 126.Lee J. H., Lee H.-B., Jung G. O., Oh J. T., Park D. E., Chae K. M. Effect of quercetin on apoptosis of PANC-1 cells. Journal of the Korean Surgical Society . 2013;85(6):249–260. doi: 10.4174/jkss.2013.85.6.249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Borska S., Drag-Zalesinska M., Wysocka T., et al. Antiproliferative and pro-apoptotic effects of quercetin on human pancreatic carcinoma cell lines EPP85-181P and EPP85-181RDB. Folia Histochemica et Cytobiologica . 2010;48(2):222–229. doi: 10.2478/v10042-08-0109-1. [DOI] [PubMed] [Google Scholar]
- 128.Borska S., Sopel M., Chmielewska M., Zabel M., Dziegiel P. Quercetin as a potential modulator of P-glycoprotein expression and function in cells of human pancreatic carcinoma line resistant to daunorubicin. Molecules . 2010;15(2):857–870. doi: 10.3390/molecules15020857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Kıyga E., Şengelen A., Adıgüzel Z., Uçar E. Ö. Investigation of the role of quercetin as a heat shock protein inhibitor on apoptosis in human breast cancer cells. Molecular Biology Reports . 2020;47(7):4957–4967. doi: 10.1007/s11033-020-05641-x. [DOI] [PubMed] [Google Scholar]
- 130.Rezabakhsh A., Rahbarghazi R., Malekinejad H., Fathi F., Montaseri A., Garjani A. Quercetin alleviates high glucose-induced damage on human umbilical vein endothelial cells by promoting autophagy. Phytomedicine . 2019;56:183–193. doi: 10.1016/j.phymed.2018.11.008. [DOI] [PubMed] [Google Scholar]
- 131.Lee K. S., Park S. N. Cytoprotective effects and mechanisms of quercetin, quercitrin and avicularin isolated from _Lespedeza cuneata_ G. Don against ROS-induced cellular damage. Journal of Industrial and Engineering Chemistry . 2019;71:160–166. doi: 10.1016/j.jiec.2018.11.018. [DOI] [Google Scholar]
- 132.Li Y., Chen M., Wang J., et al. Quercetin ameliorates autophagy in alcohol liver disease associated with lysosome through mTOR-TFEB pathway. Journal of Functional Foods . 2019;52:177–185. doi: 10.1016/j.jff.2018.10.033. [DOI] [Google Scholar]
- 133.Li G., Shen X., Wei Y., Si X., Deng X., Wang J. Quercetin reduces _Streptococcus suis_ virulence by inhibiting suilysin activity and inflammation. International Immunopharmacology . 2019;69:71–78. doi: 10.1016/j.intimp.2019.01.017. [DOI] [PubMed] [Google Scholar]
- 134.Zeng H., Guo X., Zhou F., et al. Quercetin alleviates ethanol-induced liver steatosis associated with improvement of lipophagy. Food and Chemical Toxicology . 2019;125:21–28. doi: 10.1016/j.fct.2018.12.028. [DOI] [PubMed] [Google Scholar]
- 135.Yang D. K., Kang H.-S. Anti-diabetic effect of cotreatment with quercetin and resveratrol in streptozotocin-induced diabetic rats. Biomolecules & Therapeutics . 2018;26(2):130–138. doi: 10.4062/biomolther.2017.254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Sharma G., Kumar S., Sharma M., Upadhyay N., Ahmed Z., Mahindroo N. Anti-diabetic, anti-oxidant and anti-adipogenic potential of quercetin rich ethyl acetate fraction of Prunus persica. Pharmacognosy Journal . 2018;10(3):463–469. doi: 10.5530/pj.2018.3.76. [DOI] [Google Scholar]
- 137.Nuckols T. K., Keeler E., Anderson L. J., et al. Economic evaluation of quality improvement interventions designed to improve glycemic control in diabetes: a systematic review and weighted regression analysis. Diabetes Care . 2018;41(5):985–993. doi: 10.2337/dc17-1495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Sundstrom J. M., Hernández C., Weber S. R., et al. Proteomic analysis of early diabetic retinopathy reveals mediators of neurodegenerative brain diseases. Investigative Ophthalmology & Visual Science . 2018;59(6):2264–2274. doi: 10.1167/iovs.17-23678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.El-kott A. F., Shati A. A., Ali Al-kahtani M., Alharbi S. A. The apoptotic effect of resveratrol in ovarian cancer cells is associated with downregulation of galectin-3 and stimulating miR-424-3p transcription. Journal of Food Biochemistry . 2019;43(12, article e13072) doi: 10.1111/jfbc.13072. [DOI] [PubMed] [Google Scholar]
- 140.Lee J., Lee J., Kim S. J., Kim J. H. Quercetin-3-O-glucoside suppresses pancreatic cancer cell migration induced by tumor-deteriorated growth factors in vitro. Oncology Reports . 2016;35(4):2473–2479. doi: 10.3892/or.2016.4598. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.