

Abstract
Adapting flexibly to recent events is essential in everyday life. A robust measure of such adaptive behavior is the congruency sequence effect (CSE) in the prime-probe task, which refers to a smaller congruency effect after incongruent trials than after congruent trials. Prior findings indicate that the CSE in the prime-probe task reflects control processes that modulate response activation after the prime onsets but before the probe appears. They also suggest that similar control processes operate even in a modified prime-probe task wherein the initial prime is a relevant target, rather than merely a distractor. Since adaptive behavior frequently occurs in the absence of irrelevant stimuli, the present study investigates the nature of the control processes that operate in this modified prime-probe task. Specifically, it investigates whether these control processes modulate only the response cued by the prime (response-specific control) or also other responses (response-general control). To make this distinction, we employed a novel variant of the modified prime-probe task wherein primes and probes are mapped to different responses (i.e., effectors), such that only response-general control processes can engender a CSE. Critically, we observed a robust CSE in each of two experiments. This outcome supports the response-general control hypothesis. More broadly, it suggests that the control processes underlying the CSE overlap with general mechanisms for adapting to sequential dependencies in the environment.
Keywords: conflict adaptation, congruency repetition, episodic retrieval, cognitive control
Introduction
Adapting flexibly to recent events is a crucial aspect of cognitive control (Miller & Cohen, 2001). To investigate adaptive behavior, researchers use distractor-interference tasks such as the flanker, Stroop, Simon, and prime-probe tasks (Botvinick, Braver, Barch, Carter, & Cohen, 2001; Gratton, Coles, & Donchin, 1992; Kunde & Wuhr, 2006; Ridderinkhof, 2002). For example, in each trial of the prime-probe task, a prime (or distractor) precedes a probe (or target) that participants are asked to identify. The prime cues the same response as the probe in congruent trials but a different response in incongruent trials. Participants usually respond more slowly in incongruent (vs. congruent) trials. However, this congruency effect is smaller after incongruent (vs. congruent) trials. Thus, mean probe response time (RT) depends jointly on all four stimuli over the past two trials in a pattern known as the congruency sequence effect (CSE). The CSE, which appears in many tasks, is thought to index control processes that modulate the processing of the distractor differently after incongruent (vs. congruent) trials (Gratton et al., 1992). In line with this view, a “control-driven” CSE emerges even without feature integration (e.g., stimulus or response repetition) or contingency learning (e.g., stimulus frequency) confounds (Jimenez & Mendez, 2014; Kim & Cho, 2014; Schmidt & Weissman, 2014).
An emerging view posits that the CSE depends critically on episodic retrieval (Dignath, Hommel, Johannsen, & Kiesel, 2019; Egner, 2014; Frings et al., 2020; Spape & Hommel, 2008; Weissman, Hawks, & Egner, 2016). Here, participants create an episodic memory of each trial that includes the stimuli and responses that occurred, various relationships between those stimuli and/or responses (e.g., congruent or incongruent), and the task sets and control settings that were employed. Critically, the repetition of a previous-trial feature, and/or the local temporal context, triggers the retrieval of an episodic memory of the previous trial. This biases control processes to prepare for an upcoming trial that resembles the previous one (Egner, 2014). Thus, performance is better when trial congruency repeats than when it alternates, leading to a CSE.
The episodic retrieval account is agnostic with regard to how control processes engender a CSE (Egner, 2014). Recently, however, we combined the episodic retrieval account with the view that control processes engender a CSE by modulating response activation (Ridderinkhof, 2002; Stürmer, Leuthold, Soetens, Schroter, & Sommer, 2002; Weissman, Egner, Hawks, & Link, 2015). Specifically, we proposed that, in the prime-probe task, control processes prepare for a trial whose congruency matches that of the previous trial by using the identity of the prime to modulate response activation. For example, control processes might inhibit the response cued by the prime after an incongruent trial or enhance (i.e., excite) it after a congruent trial.
Two recent findings are consistent with this proposal. First, when a long (i.e., 1000 ms) temporal interval separates the prime and probe, which eliminates the overall congruency effect, the CSE is associated with a negative congruency effect after incongruent trials (Weissman, Egner et al., 2015). This is more consistent with a modulation of response activation before the probe appears than with a shift of attention away from the prime. For example, inhibiting the response cued by the prime might slow the response to the probe when the current trial is congruent, relative to incongruent, thereby reversing the congruency effect (Ridderinkhof, 2002). In contrast, even shifting all of one’s attention away from the prime could eliminate the congruency effect but not reverse it (Schmidt, 2018). Second, the CSE in mean probe RT is larger when participants respond to both the prime (during the 1000 ms interval prior to probe onset) and the probe than when they respond only to the probe (Grant & Weissman, 2019). Since the prime is task-relevant (vs. irrelevant) in the former, modified prime-probe task, its relation to the probe (e.g., incongruent) is likely encoded and later retrieved more effectively than in the latter, distractor-interference version of the same task. Indeed, task-relevance increases the efficacy of episodic binding (Hommel, 2007) and retrieval (Huffman, Hilchey, Weidler, Mills, & Pratt, 2020), which should lead to a relatively large CSE in the modified prime-probe task.
The response-specific and response-general control hypotheses
The main aim of the present experiments is to distinguish between two hypotheses regarding the nature of the response modulation that engenders the CSE in the modified prime-probe task (Grant & Weissman, 2019). The response-specific control hypothesis posits that control processes engender a CSE solely by modulating the response cued by the prime. This hypothesis is motivated by data suggesting that greater suppression or inhibition of the response cued by the distractor after incongruent relative to congruent trials contributes to the CSE in distractor-interference tasks (Lee & Cho, 2013; Ridderinkhof, 2002; Stürmer et al., 2002; Weissman, Jiang, & Egner, 2014). The existence of such a response-specific control mechanism is normatively justifiable as a means to regulate low-level repetition priming effects. Such a mechanism may modulate the activation of the distractor response based on current expectations regarding whether this response will or will not predict the correct response to the upcoming target1. Although there is no distractor in the modified prime-probe task, after participants respond to the prime, control processes may enhance the prime response after congruent trials but inhibit the prime response after incongruent trials (Weissman, Colter, Grant, & Bissett, 2017). These modulations of the prime response would lead to a smaller (or reversed) congruency effect in mean probe RT after incongruent relative to congruent trials (i.e., a CSE). Notice, however, that this reasoning applies only if the prime and probe in congruent trials are mapped to the same response (i.e., effector), a point to which we will return shortly.
The response-general control hypothesis is motivated by data suggesting that control processes can use a distractor’s identity to prepare a congruent or incongruent response to an upcoming target (Logan, 1985; Logan & Zbrodoff, 1979). Here, the control processes underlying the CSE overlap with those that learn relatively complex dependencies in stimulus and response sequences (Jones, Curran, Mozer, & Wilder, 2013). In relation to the modified prime-probe task, this view posits that, after participants respond to the prime following a congruent trial, control processes enhance the response cued by the prime and/or inhibit the opposite response2. Following an incongruent trial, however, control processes inhibit the response cued by the prime and/or enhance the opposite response (Weissman et al., 2017). These effects would lead to a smaller (or reversed) congruency effect in mean probe RT after incongruent relative to congruent trials (i.e., a CSE). Critically, however, control processes would engender a CSE not only by modulating the response cued by the prime but also by modulating other responses.
Prior studies could not distinguish between the response-specific and response-general control hypotheses because the prime and probe in congruent trials were mapped to the same response. Consider a study of the modified prime-probe task that made use of force-sensitive keys (Weissman, 2019). Data from this study indicate that, just before the probe appears, participants exert more force on (a) the response key cued by the prime (i.e., the prime-congruent key) after congruent trials and (b) the opposite key (i.e., the prime-incongruent key) after incongruent trials. The latter result could indicate that control processes use the identity of the prime to enhance a prime-incongruent response to the probe after incongruent trials (response-general control). However, it could also reflect lateral disinhibition of the prime-incongruent response that is caused by inhibiting the prime-congruent response (response-specific control).
Experiment 1
The goal of Experiment 1 was to distinguish between the response-specific and response-general control hypotheses. To achieve this goal, we designed a novel variant of the modified prime-probe task wherein the prime and probe are mapped to different responses in both congruent and incongruent trials. Here, participants respond to the prime letters A, B, C, and D using the index, middle, ring, and pinkie fingers (respectively) of their left hand and to the probe letters A, B, C, and D using the corresponding fingers of their right hand. As we explain next, only the response-general control hypothesis predicts a CSE in this task.
The response-general control hypothesis predicts a CSE because it posits that control processes can modulate a response that differs from the one cued by the prime. Thus, after a congruent trial, participants could respond to the prime letter “A” with the left index finger and control processes could then prepare for a probe letter “A” by enhancing a corresponding right index finger response or inhibiting a non-corresponding right middle finger response3. Analogously after an incongruent trial, control processes could prepare for a probe letter “B” by inhibiting a corresponding right index finger response or enhancing a non-corresponding right middle finger response. Either, or both, of these effects would lead to faster probe RT when trial congruency repeats (vs. alternates), thereby engendering a CSE.
In contrast, the response-specific control hypothesis does not predict a CSE in the present task. This is because the probe requires a different response than the prime in both congruent and incongruent trials. For this reason, modulating (e.g., inhibiting) the response cued by the prime on the left hand should not differentially influence the ability to prepare a corresponding (i.e., congruent) versus non-corresponding (i.e., incongruent) probe response on the right hand. Further, any change in lateral disinhibition that arises from modulating (e.g., inhibiting) the response cued by the prime should apply equally to corresponding and non-corresponding probe responses on the opposite hand. Consequently, modulating the prime response in the present task should not influence the congruency effect or engender a CSE.
Finally, we note that the present study differs from prior studies that have investigated whether changing the effectors that participants use to respond in consecutive trials influences the CSE. These studies have shown that the CSE is sometimes smaller when participants switch between different hands (Kim & Cho, 2014; Lim & Cho, 2018) or between their hands and their feet (Braem, Verguts, & Notebaert, 2011; Janczyk & Leuthold, 2018) from one trial to the next than when they do not switch. However, the CSE in the distractor-interference version of the prime-probe task is not influenced by whether participants use the same or different hands to respond in consecutive trials (Weissman, Colter, Drake, & Morgan, 2015). Further, the hand that participants use to respond in the present study varies within each trial, rather than across trials as in the studies above. Thus, the present study tests a qualitatively different hypothesis than these prior studies. Specifically, it tests whether control processes can engender a CSE by modulating a response that differs from the one cued by the prime. Prior studies could not test this hypothesis because the prime and probe in congruent trials were always mapped to the same response.
Methods
Participants
Based on power analyses conducted with G*Power 3.1.9.2 (Faul, Erdfelder, Lang, & Buchner, 2007), we sought usable data from 24 healthy participants. These analyses employed an alpha level of 0.05 and the effect size for the CSE in mean probe RT from a prior study of the modified prime-probe task () (Grant & Weissman, 2019). The results showed that 24 participants would provide over 99% power for observing a CSE.
Thirty-three college students from the University of Michigan’s Psychology Department subject pool participated in the experiment. We excluded the data from four students who performed the task with less than 70% accuracy4 and from five students who self-reported a neuropsychiatric disorder. None of the remaining 24 participants (13 female, 11 male; mean age, 18.7 years; age range, 18–20 years; SD = 0.9 years) reported any history of neurological or neuropsychiatric disorders, seizures, head trauma, or uncorrected visual or hearing impairments. Participants received course credit in exchange for taking part in the study. The University of Michigan’s Behavioral Sciences Institutional Review Board approved the study.
Stimuli
A fixation cross (0.8°× 0.8°) appeared for two seconds at the beginning and end of each block. The primes (Arial Unicode MS, 154-point font) were the letters “A” (2.3°× 2.3°), “B” (1.9°× 2.3°), “C” (2.1°× 2.3°), and “D” (2.0°× 2.3°). The probes (Arial Unicode MS, 77-point font) were also the letters “A” (1.3°× 1.2°), “B” (0.8°× 1.2°), “C” (0.9°× 1.2°), and “D” (0.9°× 1.2°), but they were much smaller. All stimuli appeared in white on a black background.
Experimental Design
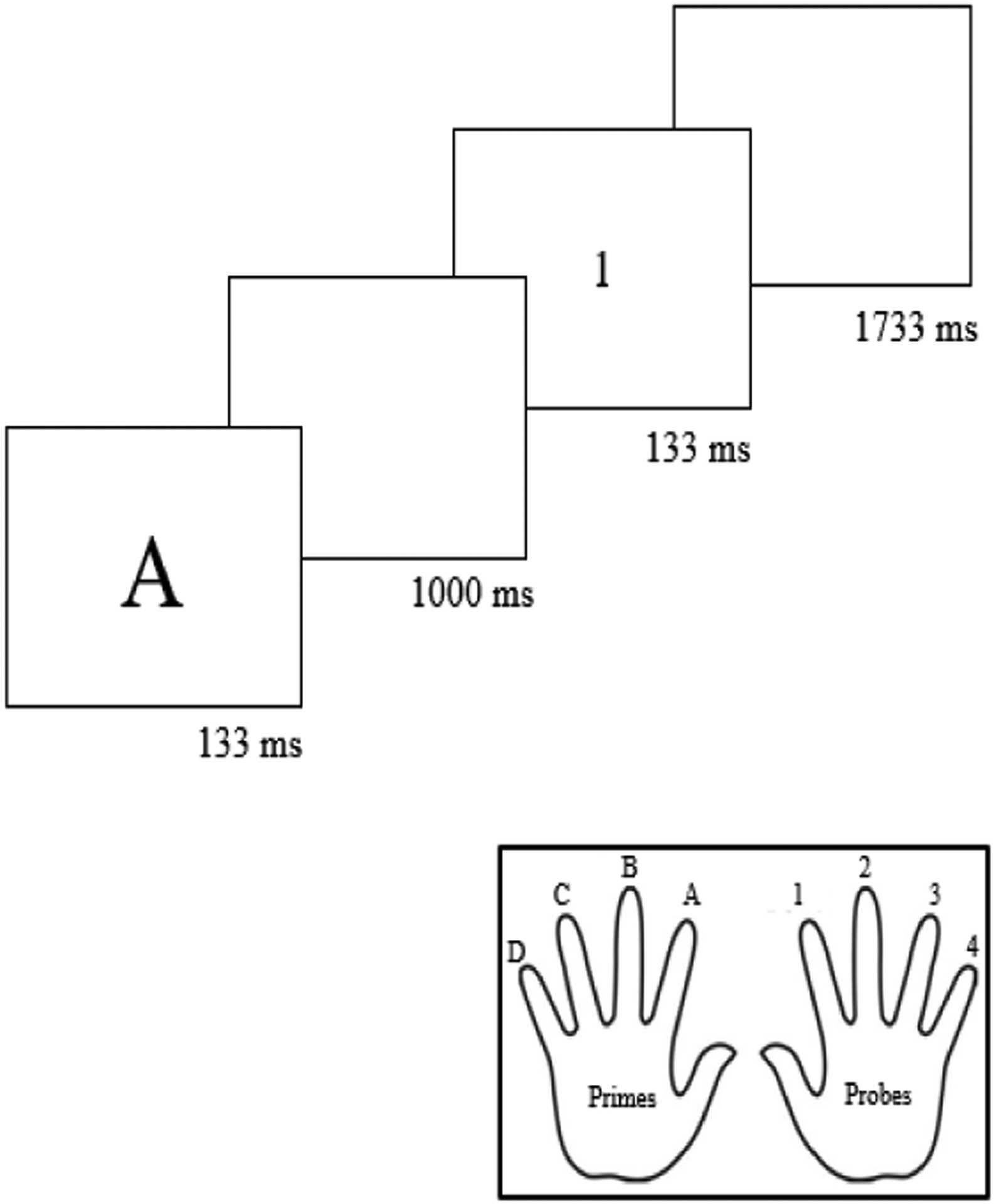
Each 3-second trial consisted of four sequential events: (1) a prime letter (duration, 133 ms), a blank screen (duration, 1000 ms), a probe letter (duration, 133 ms), and a second blank screen (duration, 1733 ms) (Fig. 1). In congruent trials, the prime and probe were the same letter (e.g., A). In incongruent trials, the prime and probe were different letters (e.g., A and B). There were four unique congruent pairs of prime and probe stimuli (A-A, B-B, C-C, D-D) and four unique incongruent pairs (A-B, B-A, C-D, D-C).
Figure 1.

The modified prime-probe task that we employed in Experiment 1. In each trial (top), participants responded to the prime (first letter) during the 1000 ms inter-stimulus interval (ISI) and to the probe (second letter) during the 1733 ms ISI. As shown at the bottom, participants used one of four fingers on their left hand to respond to the prime letter and one of four fingers on their right hand to respond to the probe letter. The figure provides an example of a congruent trial wherein participants were supposed to press their left index finger to identify the prime and their right index finger to identify the probe.
We instructed participants to respond to the prime and probe in each trial using a QWERTY keyboard. First, we asked participants to respond to A, B, C, and D prime letters during the first (1000 ms) blank screen by pressing the F (left index finger), D (left middle finger), S (left ring finger), and A (left pinkie finger) keys, respectively. Second, we asked participants to respond to A, B, C, and D probe letters during the second (1733 ms) blank screen by pressing the H (right index finger), J (right middle finger), K (right ring finger), and L (right pinkie finger) keys, respectively.
We imposed three constraints on the trial sequence for each 64-trial block. First, to avoid feature integration confounds, which can occur when stimuli and/or responses repeat in consecutive trials (Hommel, Proctor, & Vu, 2004), we presented the letters A and/or B in odd trials and the letters C and/or D in even trials. Second, to avoid contingency learning confounds (Schmidt & De Houwer, 2011), which occur when each distractor is associated with the congruent target more often than with the incongruent target, we presented each unique congruent and incongruent letter pair equally often. Third, we presented the trials in each block in a pseudo-randomized order. Specifically, we presented the trials such that congruent and incongruent letter pairs each preceded congruent and incongruent letter pairs equally often, separately for odd and even trials. Three of the resulting four successive-trial congruency sequences appeared 16 times per block. However, one appeared only 15 times because no trial preceded the first trial. The minority congruency sequence varied randomly across blocks. Table 1 provides examples of the four successive-trial congruency sequences in Experiment 1.
Table 1.
Examples of the four successive-trial congruency sequences in Experiment
| Previous Trial | Current Trial | |||
|---|---|---|---|---|
| Sequence | Stimulus pair | Response pair | Stimulus pair | Response pair |
| cC | A-A | left index, right index | D-D | left pinkie, right pinkie |
| cI | A-A | left index, right index | C-D | left ring, right pinkie |
| iC | A-B | left index, right middle | D-D | left pinkie, right pinkie |
| iI | A-B | left index, right middle | C-D | left ring, right pinkie |
Note: “Stimulus pair” and “Response pair” indicate the prime followed by the probe on each trial. cC: Previous trial congruent, current trial congruent; cI: Previous trial congruent, current trial incongruent; iC: Previous trial incongruent, current trial congruent; iI: Previous trial incongruent, current trial incongruent; I-C: Incongruent—Congruent; CSE: Congruency sequence effect, calculated as: (RTcI − RTcC) − (RTiI − RTiC).
Procedure
Participants completed a 64-trial practice block followed by nine 64-trial test blocks. We instructed participants to respond as quickly and as accurately as possible to the prime and probe letter in each trial as described earlier. If a participant responded incorrectly or did not respond within 900 ms of stimulus onset, the word “Error” appeared centrally on the computer screen for 200 ms. To present the stimuli and record participants’ responses, we employed the Psychophysics Toolbox (Brainard, 1997) running on a Windows 10 PC.
Data Analyses
We employed separate repeated-measured analyses of variance (ANOVAs) to analyze mean probe RT and mean probe error rate (ER). The ANOVAs included two factors: (1) previous trial congruency (congruent, incongruent) and (2) current trial congruency (congruent, incongruent). In the analysis of mean probe RT, we excluded outliers (trials with RTs more than three standard deviations from their condition means), errors (involving either the prime response or the probe response), and trials after errors. In the analysis of mean probe ER, we excluded outliers, trials after errors, and trials wherein participants responded incorrectly to the prime. Overall accuracy was relatively high (86.1%), and the percentage of outliers was relatively low (0.7%).
Results
Mean Probe RT
There were two significant main effects. First, there was a main effect of previous trial congruency, F(1, 23) = 89.20, p < 0.001, , because mean RT was slower after incongruent trials (446 ms) than after congruent trials (427 ms). Second, there was a main effect of current trial congruency, F(1, 23) = 73.42, p < 0.001, , because mean RT was longer in current incongruent trials (464 ms) than in current congruent trials (409 ms).
Critically, there was a significant interaction between previous trial congruency and current trial congruency (i.e., a CSE), F(1, 23) = 46.17, p < 0.001, . In line with the response-general control hypothesis, but not with the response-specific control hypothesis, the congruency effect was smaller after incongruent (vs. congruent) trials (37 ms vs. 74 ms) (Fig. 2).
Figure 2.
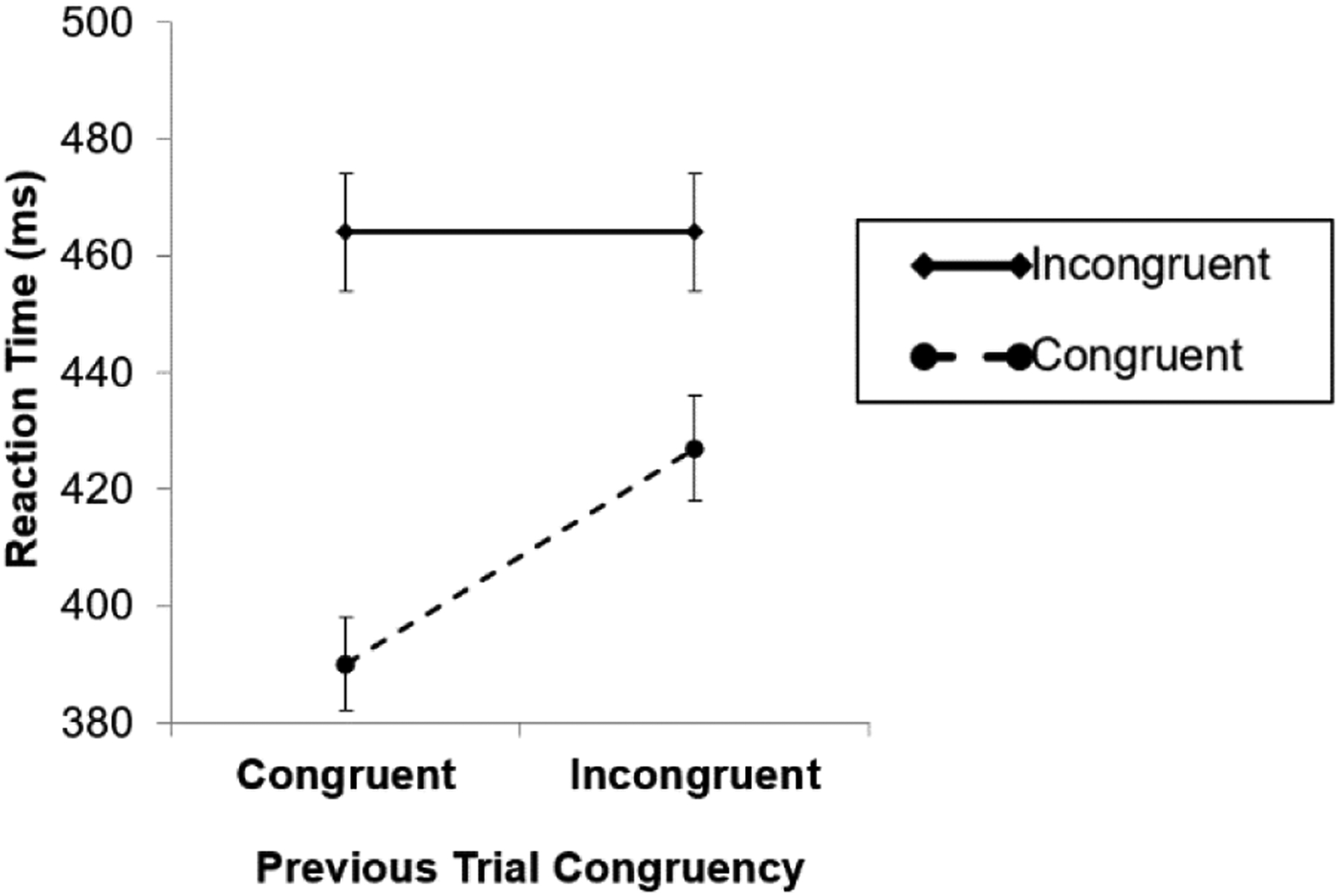
The congruency sequence effect (CSE) in Experiment 1. Consistent with the response-general control hypothesis, but not with the response-specific control hypothesis, there was a CSE even though participants responded to primes and probes using fingers on different hands. Error bars indicate ±1 standard error (across participants) from the condition mean.
Mean Probe ER
There was one significant effect. Specifically, there was a main effect of current trial congruency, F(1, 23) = 21.94, p < 0.001, , because mean ER was higher in current incongruent trials (6.3%) than in current congruent trials (3.0%). The main effect of previous trial congruency did not achieve conventional levels of significance, F(1, 23) = 3.61, p = 0.07, . However, mean ER was higher after incongruent trials (5.1%) than after congruent trials (4.2%), as we observed in the analysis of mean RT. The interaction between previous trial congruency and current trial congruency was not significant, F(1, 23) < 1.
Discussion
We observed both an overall congruency effect and a CSE in mean probe RT even though participants responded to primes and probes with different effectors. Critically, the latter finding supports the response-general control hypothesis, wherein control processes can engender a CSE in the present task by modulating a response other than the one cued by the prime. In particular, this finding suggests that control processes can engender a CSE by integrating a memory of previous trial congruency (i.e., congruent or incongruent) with the identity of the current-trial prime to prepare a congruent or incongruent probe response on the opposite hand. In contrast, this finding is not easily explained by the response-specific control hypothesis, wherein control processes can engender a CSE only by modulating the response cued by the prime. This is because, in our task, the prime and probe are mapped to different responses in both congruent and incongruent trials. Thus, even if control processes modulate (e.g., inhibit) the response cued by the prime after both of these trial types, this modulation should not differentially influence the ability to make a corresponding (i.e., congruent) versus non-corresponding (i.e., incongruent) probe response on the opposite hand. In other words, this modulation should neither influence the congruency effect nor engender a CSE. For these reasons, our findings are more consistent with the response-general control hypothesis than with the response-specific control hypothesis.
Experiment 2
The support we observed for the response-general control hypothesis in Experiment 1 concerns the output of control processes (i.e., the probe responses they act on). There is also the question of the input (i.e., features or representations) that control processes use to derive expectations of upcoming probe responses. In particular, the congruency relations that control processes use to engender a CSE in the present modified prime-probe task remain unclear. More specifically, it is unclear whether control processes use only perceptual congruency relations between the prime and probe to engender a CSE or also categorical congruency relations (Frings et al., 2020). In Experiment 2, we sought to distinguish between these possibilities.
The perceptual congruency hypothesis posits that control processes engender a CSE by preparing for a probe that has the same perceptual relation to the prime as in the previous trial and modulating response activation accordingly. More specifically, this hypothesis posits that control processes prepare for a probe that looks physically similar to the prime when the previous prime and probe were physically similar, and/or a probe that looks physically different from the prime when the previous prime and probe were physically different. In line with this hypothesis, the prime and the probe in Experiment 1 were the same letter in congruent trials but different letters in incongruent trials. Moreover, some findings suggest that the CSE in distractor-interference tasks is driven, at least partly, by the fact that the distractor and target look more similar in congruent relative to incongruent trials (Aarts & Roelofs, 2011; Compton, Huber, Levinson, & Zheutlin, 2012; Notebaert & Verguts, 2006; Schmidt & Weissman, 2015; Verbruggen, Notebaert, Liefooghe, & Vandierendonck, 2006).
The categorical congruency hypothesis posits that control processes engender a CSE by preparing for a probe that has the same categorical stimulus or categorical response relation to the prime as in the previous trial and modulating response activation accordingly. First, control processes may prepare for a probe stimulus that has the same categorical stimulus relation to the prime stimulus as in the previous trial. For example, they may use the ordinal (e.g., alphabetical) value of the prime (e.g., third for the letter C) to prepare for a probe that has the same ordinal value after congruent trials (e.g., third for the letter C) and/or a different ordinal value after incongruent trials (e.g., fourth for the letter D). Second, they may prepare for a probe that has the same categorical response relation to the prime as in the previous trial. After a congruent trial wherein the prime and probe required corresponding responses on opposite hands (e.g., left index finger and right index finger), control processes may use the current prime response on the left hand (e.g., left ring finger) to prepare for a corresponding probe response on the right hand (e.g., right ring finger). In contrast, after an incongruent trial wherein the prime and probe required non-corresponding responses on opposite hands (e.g., left index finger and right middle finger), control processes may use the current prime response on the left hand (e.g., left ring finger) to prepare for a non-corresponding probe response on the right hand (e.g., right pinkie finger).
It is important not to confuse the distinction we draw here, which concerns the stimulus or response relations that determine trial congruency, from the abstract nature of the CSE itself. The fact that a CSE appears in confound-minimized tasks shows that it arises from repetitions of trial congruency—an abstract relation between the prime and probe—rather than from repetitions of individual stimuli or responses. The question we ask here concerns which relations between stimuli or responses serve to define trial congruency in the present task. Specifically, we ask whether control processes define trial congruency solely in terms of perceptual relations between the prime and probe stimuli (i.e., whether these stimuli are perceptually similar or dissimilar in any given trial), or also with respect to categorical stimulus or categorical response relations between these stimuli (i.e., whether these stimuli have similar or dissimilar ordinal values or require responses with corresponding or non-corresponding fingers on the left and right hands).
Thus, the goal of Experiment 2 was to distinguish between the perceptual and categorical congruency hypotheses. To this end, we altered the task from Experiment 1 by replacing the four probe letters (A, B, C, D) with four probe digits (1, 2, 3, 4). In this task, the primes and probes do not match with regard to their perceptual features in either congruent trials (A-1, B-2, C-3, D-4) or incongruent trials (A-2, B-1, C-4, D-3), because the primes and probes always come from different stimulus categories (i.e., letters and digits). However, the primes and probes still correspond with regard to their categorical stimulus (i.e., alphabetical or numerical ordering) and categorical response (i.e., finger mapping) relations in congruent trials but not in incongruent trials. In other words, trial congruency is not meaningful if it is defined with respect to perceptual relations but is meaningful if it is defined with respect to categorical relations. Given these considerations, we reasoned that if control processes can use only perceptual congruency relations to engender a CSE, then we should not observe a CSE in this task. In contrast, if control processes can use categorical congruency relations to engender a CSE, as some prior findings from confound-minimized tasks suggest could be the case (Schmidt & Weissman, 2015), then we should observe a CSE.
Methods
Participants
Based on power analyses conducted with G*Power 3.1.9.2 (Faul et al., 2007), we sought usable data from 24 healthy participants. These analyses employed an alpha level of 0.05 and the effect size for the CSE in mean probe RT from Experiment 1 (). We found that 24 participants would provide over 99% power for observing a CSE.
Twenty-eight college students from the University of Michigan’s Psychology Department subject pool participated in the experiment. We excluded the data from three students who performed the task with less than 70% accuracy5 and from one student who self-reported a neuropsychiatric disorder. None of the remaining 24 participants (17 female, 7 male; mean age, 19.0 years; age range, 18–22 years; SD = 0.9 years) reported any history of neurological or neuropsychiatric disorders, seizures, head trauma, or uncorrected visual or hearing impairments. Participants received course credit in exchange for taking part in the study. The University of Michigan’s Behavioral Sciences Institutional Review Board approved the study.
Stimuli
The stimuli were the same as those in Experiment 1 with a single exception. The probes were the digits “1” (0.4°× 1.2°), “2” (0.8°× 1.2°), “3” (0.7°× 1.2°), and “4” (0.8°× 1.2°) (Fig. 3).
Figure 3.
The prime-probe task we employed in Experiment 2. In each trial (top), participants responded to the prime letter during the 1000 ms inter-stimulus interval (ISI) and to the probe digit during the 1733 ms ISI. As shown at the bottom, participants used one of four fingers on their left hand to respond to the prime letter and one of four fingers on their right hand to respond to the probe digit. The figure provides an example of a congruent trial wherein participants were supposed to press their left index finger to identify the prime and their right index finger to identify the probe.
Experimental Design
The experimental design was identical to that in Experiment 1 with one exception. We replaced the probe letters A, B, C, and D with the probe digits 1, 2, 3, and 4. To avoid stimulus and response repetition (i.e., feature integration) confounds, we presented prime As and Bs and probe 1s and 2s in odd trials and prime Cs and Ds and probe 3s and 4s in even trials. Table 2 provides examples of the four successive-trial congruency sequences in Experiment 2.
Table 2.
Examples of the four successive-trial congruency sequences in Experiment 2
| Previous Trial | Current Trial | |||
|---|---|---|---|---|
| Sequence | Stimulus pair | Response pair | Stimulus pair | Response pair |
| cC | A-1 | left index, right index | D-4 | left pinkie, right pinkie |
| cI | A-1 | left index, right index | C-4 | left ring, right pinkie |
| iC | A-2 | left index, right middle | D-4 | left pinkie, right pinkie |
| iI | A-2 | left index, right middle | C-4 | left ring, right pinkie |
Note: “Stimulus pair” and “Response pair” indicate the prime followed by the probe on each trial. cC: Previous trial congruent, current trial congruent; cI: Previous trial congruent, current trial incongruent; iC: Previous trial incongruent, current trial congruent; iI: Previous trial incongruent, current trial incongruent; I-C: Incongruent—Congruent; CSE: Congruency sequence effect, calculated as: (RTcI – RTcC) – (RTiI – RTiC).
Procedure
The procedure was identical to that in Experiment 1.
Data Analyses
The data analyses were identical to those in Experiment 1. On average, overall accuracy was relatively high (85.3%), and the percentage of outliers was relatively low (0.8%).
Results
Mean Probe RT
There was one significant main effect. Specifically, there was a main effect of current trial congruency, F(1, 23) = 45.76 p < 0.001, , because mean RT was longer in current incongruent trials (475 ms) than in current congruent trials (446 ms). The main effect of previous trial congruency was not significant (p > 0.10).
Critically, there was a significant interaction between previous trial congruency and current trial congruency, F(1, 23) = 11.67, p = 0.002, . Consistent with the categorical congruency hypothesis, the congruency effect was smaller after incongruent trials (23 ms) than after congruent trials (35 ms) (Fig. 4).
Figure 4.
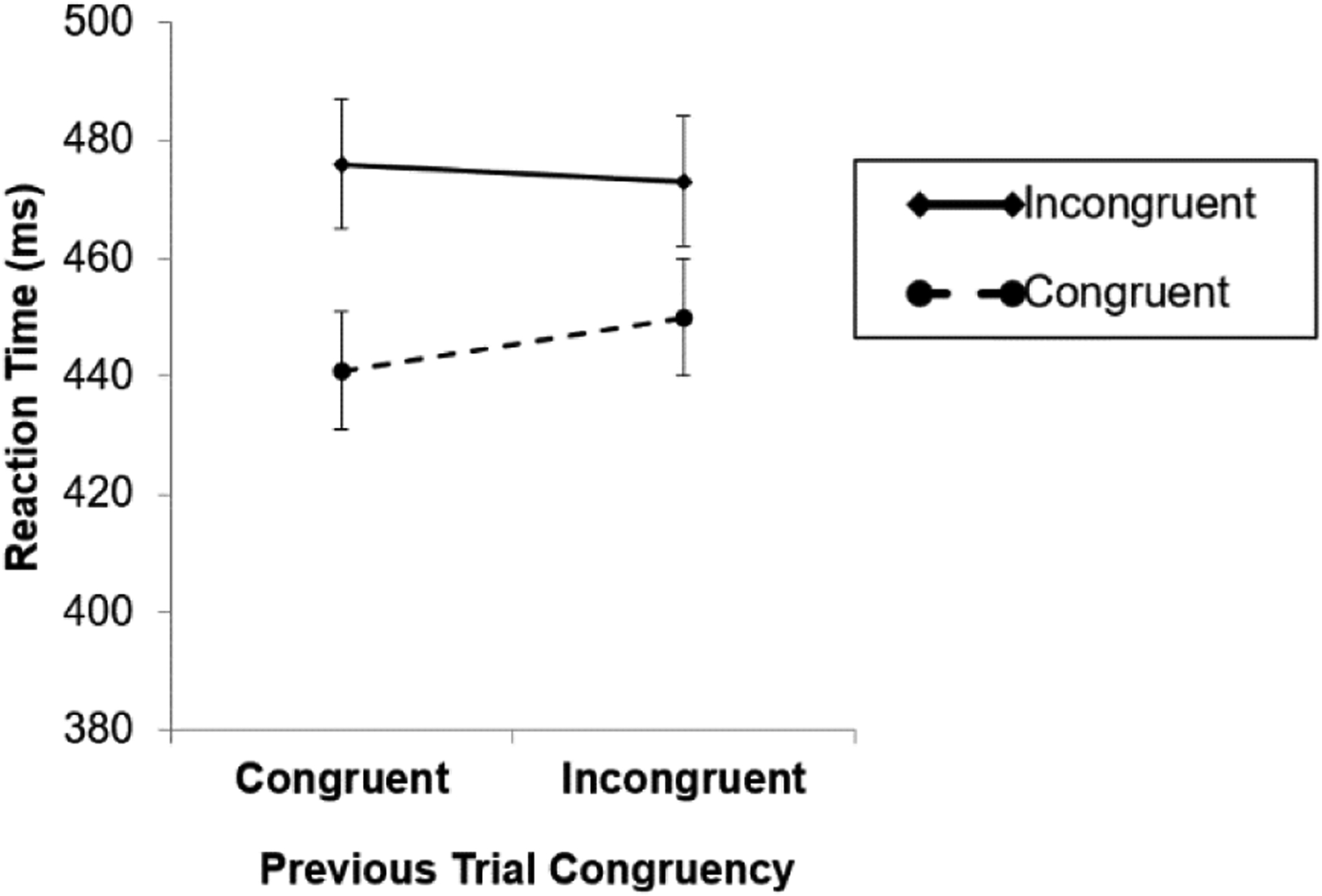
The congruency sequence effect (CSE) in Experiment 2. Consistent with the categorical congruency hypothesis, there was a CSE even though the prime and probe were associated with different stimulus categories and responses in both congruent and incongruent trials. Error bars indicate ±1 standard error (across participants) from the condition mean.
Mean Probe ER
There was a significant main effect of current trial congruency F(1, 23) = 34.94, p < 0.001, . As expected, mean ER was higher in incongruent trials (6.6%) than in congruent trials (3.2%). No other effects were significant (all p-values > 0.20).
Discussion
The results of Experiment 2 show that categorical congruency between the prime and probe (in the absence of perceptual congruency) influences both overall performance and the control processes underlying the CSE in the present modified prime-probe task. First, we observed a main effect of current trial congruency (i.e., a congruency effect), which indicates that categorical congruency influences overall performance in this task. Indeed, as we explained earlier, congruent and incongruent trials in this task differ with regard to categorical congruency but not with regard to perceptual congruency. Second, and most important, we observed a CSE. This indicates that control processes can prepare for a probe that has the same categorical relation to the prime as in the previous trial. These results provide novel support for the categorical congruency hypothesis.
Our findings in Experiment 2 also provide additional support for the response-general control hypothesis. Indeed, they conceptually replicate our main finding in Experiment 1, which is that a CSE appears even when primes and probes are mapped to responses on different hands. This outcome shows once again that the control processes underlying the CSE can modulate a response other than the one cued by the prime.
Finally, the results of Experiment 2 rule out an alternative, response-specific control account of our findings in Experiment 1. Recall that in Experiment 1 the prime letter always cued two responses, one via the prime stimulus-response mapping and one via the probe stimulus-response mapping. For example, the prime letter “A” cued both a left index finger response (as a prime) and a right index finger response (as a probe). Notably, the response cued by the prime according to the probe stimulus-response mapping (e.g., the right index finger response for a prime “A”) was incorrect only in incongruent trials, because the probe required this response in congruent trials. Thus, control processes may have inhibited the two responses cued by the prime only after incongruent trials. Any such inhibition could have engendered a CSE by slowing mean probe RT in subsequent congruent trials wherein the probe required one of these (now inhibited) responses, thereby reducing the congruency effect after incongruent trials. In this account, control is still specific to responses that are cued by the prime. However, control operates on all of the responses cued by the prime, rather than only the response that is cued by the prime stimulus-response mapping. Critically, contrary to this account, Experiment 2 revealed a CSE even though each prime cued a response only via the prime stimulus-response mapping (because the primes were letters mapped to the left hand while the probes were digits mapped to the right hand). This outcome provides further evidence for a response-general control account of the CSEs that we have observed.
Exploratory Across-Experiment Analysis
The results of Experiment 2 show that control processes use categorical congruency relations to engender a CSE. However, they do not reveal whether control processes also use perceptual congruency relations. The latter hypothesis predicts a larger CSE in Experiment 1 than in Experiment 2, because only in Experiment 1 were the prime and probe more perceptually similar in congruent trials than in incongruent trials. Consistent with this prediction, an exploratory across-experiment analysis revealed an interaction among experiment, previous trial congruency and current trial congruency, F(1, 46) = 14.219, p < 0.001, , because the CSE was larger in Experiment 1 (36 ms) than in Experiment 2 (12 ms). The interaction between experiment and current trial congruency was also significant, F(1,46) = 11.605, p = 0.001, , because the congruency effect was larger in Experiment 1 (55 ms) than in Experiment 2 (29 ms). Thus, the mechanism underlying the CSE appears to be relatively flexible, seizing on whatever sources of congruency are available to adapt to recent events.
General Discussion
In the present study, we investigated whether the CSE in the modified prime-probe task indexes response-specific or response-general control. To do so, we mapped primes and probes to responses on the left and right hands, respectively, such that only response-general control processes could engender a CSE. Consistent with the response-general control hypothesis, we observed a robust CSE in each of two experiments. Additionally, our second experiment revealed a CSE even though the prime and probe stimuli came from different categories (i.e., letters and digits). As we discuss next, these findings have important implications for our understanding of how cognitive control processes enable adaptive behavior.
Implications for the response-specific and response-general control hypotheses
To our knowledge, the present findings are the first to distinguish between the response-specific and response-general control accounts of the CSE in the modified prime-probe task, favoring the latter. This outcome indicates that the control processes underlying the CSE can modulate not only the response cued by the prime but also other responses. It also suggests that these control processes overlap with those that support the learning of relatively complex dependencies in stimulus and response sequences (Jones et al., 2013), rather than being limited to managing repetition priming and response conflict. For example, it suggests that these control processes may engender a CSE by integrating a memory of previous trial congruency (e.g., congruent) with the response cued by the current-trial prime (e.g., “left index finger”) to deduce, and then prepare, a response to the upcoming probe with a homologous finger on the opposite hand (e.g., “right index finger”). In contrast, the present findings diverge from the predictions of the response-specific control hypothesis, wherein control mechanisms merely suppress or enhance the low-level tendency to repeat the response cued by the prime.
Interestingly, both experiments revealed that the CSE was driven primarily by a simple effect (in the statistical sense) of previous trial congruency on mean probe RT in congruent trials, with little or no effect in incongruent trials6. This pattern may reflect the combined effect of two control processes, one responsible for the CSE and another that produces overall slowing following incongruent trials. Specifically, in addition to the response modulation mechanism we have focused on, there may be a second control process that increases response caution (e.g., raises the threshold for responding) following an incongruent trial. Critically, while the response modulation process speeds RTs in iI (vs. cI) trials, the threshold process has the opposite effect, potentially leading to no difference in mean RT between iI and cI trials. In contrast, these processes work in the same direction in congruent trials to speed RTs in cC (vs. iC) trials. The end result is a CSE as we observed. Notably, this account is analogous to the conflict monitoring account of the CSE in distractor-interference tasks, wherein the first control process shifts attention toward the target after incongruent trials (Ullsberger, Bylsma, & Botvinick, 2005). Thus, a two-process control mechanism may engender CSEs in a variety of tasks with the nature of the first process (e.g., response modulation, shift of attention, etc.) varying across tasks.
Our findings also indicate for the first time that response-general control processes can employ categorical congruency relations between the prime and probe to engender a CSE. That is, they indicate that control processes can engender a CSE using ordinal value relations between the prime and probe or anatomical correspondences between fingers on the left and right hands. This outcome indicates that control processes can employ relatively complex congruency relations to engender a CSE in the present modified prime-probe tasks. It is also consistent with the view that repeating a categorical feature can trigger the retrieval of a previous episode (Frings et al., 2020) and thereby engender a CSE. Future studies could identify more precisely which categorical relations are employed (stimulus, response, or both). They could also follow up on the results of our exploratory analysis, which suggest that the CSE arises from multiple representations (e.g., perceptual congruency and categorical congruency).
Broader implications
The present findings extend our understanding of the relations, or bindings, in episodic memory that control processes employ to engender a CSE. Prior work has established that bindings form between different stimuli (Hommel, 1998), between stimuli and responses (Frings, Rothermund, & Wentura, 2007; Hommel, 1998), between different responses (Moeller & Frings, 2019a, 2019b), and even between stimuli and control processes (i.e., stimulus-control associations) (Weissman et al., 2016; Whitehead, Pfeuffer, & Egner, 2020). Prior work has also shown that retrieving these sorts of bindings engenders the CSE (Hommel et al., 2004; Mayr, Awh, & Laurey, 2003; Weissman et al., 2016). To our knowledge, however, the present findings are the first to show that control processes employ categorical relations based on ordinal values or anatomical correspondences between fingers on the left and right hands to engender a CSE. These findings advance our understanding of the episodic memory representations that control processes employ to enable adaptive behavior. They also further our understanding of how control processes actually use those representations to engender a CSE (i.e., by modulating a response that potentially differs from the one cued by the prime).
One may wonder why control processes should prepare for repetitions of trial congruency given that congruency varies randomly in our experiments. One possibility is that, in randomly varying, real-world environments the previous event provides an estimate of the current environmental state and, hence, predicts to some degree the nature of the next event (Behrens, Woolrich, Walton, & Rushworth, 2007). Thus, preparing for repetitions of recent events may be adaptive. By extension, control processes may prepare for congruency repetitions because preparing for repetitions of relationships between elementary events (e.g., congruent or incongruent) is also adaptive (Hommel & Wiers, 2017). This possibility fits with the view that implicit expectations about upcoming trial congruency contribute to the CSE (Egner, 2014).
Limitations
It is reasonable to ask whether the present CSEs are limited to tasks wherein the prime and probe in congruent trials are mapped to homologous fingers on the left and right hands. To test this hypothesis, future studies could assess whether a CSE appears in a variant of Experiment 2 wherein the letters A–D are mapped to the four fingers of the left hand in a left-to-right fashion, rather than in a right-to-left fashion as in the present study. In this variant, each congruent trial involves two spatially corresponding, yet non-homologous responses (e.g., A-1 involves the left pinkie and right index fingers, B-2 involves the left ring and right middle fingers, etc.). Thus, observing a CSE would indicate that the present CSEs generalize to tasks wherein the prime and probe in congruent trials are mapped to non-homologous fingers on the left and right hands. Such an outcome would also suggest the possibility that control processes can employ spatial congruency relations between the prime and probe responses to engender CSEs in the present tasks. Future studies could investigate these hypotheses.
Finally, one may wonder whether the absence of a negative congruency effect after incongruent trials in our tasks argues against a response modulation account of the present CSEs. We cannot exclude this possibility, but a more likely explanation is that the overall congruency effect is larger in the present 8-choice tasks (42 ms on average) than in 4-choice versions of the modified prime-probe task (~ 0 ms) (Grant & Weissman, 2019). Thus, after incongruent trials, control processes that modulate response activation after the prime appears can reduce, but not eliminate, the congruency effect. Consistent with this explanation, a negative congruency effect after incongruent trials is typically observed only when there is no overall congruency effect. Moreover, the size of the CSE in the distractor-interference version of the prime-probe task does not vary with whether the overall congruency effect is large (78 ms) or absent (0 ms), suggesting that similar control processes (i.e., those that modulate response activation after the prime appears) engender the CSE in both conditions (Weissman, Egner, et al., 2015).
Conclusion
We sought to distinguish between response-specific and response-general control in a modified prime-probe task wherein all of the stimuli are task-relevant, and primes and probes are mapped to different responses in both congruent and incongruent trials. Our findings indicate that response-general control processes engender a CSE in this task. They also indicate that these control processes can employ categorical stimulus and/or response relations – not just physical identity relations – between the prime and probe to engender a CSE. Future variants of the modified prime-probe task may provide additional insights into how interactions among learning, memory, and cognitive control give rise to adaptive behavior.
Public significance statement.
Adapting flexibly to recent events is a crucial aspect of cognitive control. For example, after discovering that a passenger’s directions for reaching one destination are incorrect, a driver may become cautious about following the same passenger’s directions to a second destination. It remains unclear, however, exactly how control processes adapt flexibly to whether or not advance information (e.g., driving directions) was recently useful. More specifically, it remains unclear whether they adapt solely by modulating the response that advance information currently cues (e.g., by inhibiting a “turn left” response that a passenger suggests) or also by modulating a different response (e.g., by activating an alternative “turn right” response). Our findings support the latter possibility and thereby distinguish between competing accounts of adaptive control.
Acknowledgments
The authors thank Andrea Dai, Cathryn Goldman, Ceren Ege, Al-Amin Ali, Alexis Salinas, and Daphne Samuel for assisting with data collection. The raw data associated with the present manuscript are publicly available on the Open Science Framework. Interested parties can access these data freely via the following link: https://osf.io/j2wt7/?view_only=5219a945766d4f13a813ef5c5e689031
Funding:
This work was supported in part by National Institute of Mental Health Award R21 MH120741 to Matt Jones. It was also supported in part by the National Science Foundation Graduate Research Fellowship Program under Grant No. (DGE1256260). Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Footnotes
We use the term “expectation” to refer to an adaptive system that (a) utilizes prior input (experience) to derive predictions about future events and then (b) uses those predictions to guide output (behavior).
All tasks we consider involve binary responding, meaning there are two possible responses at each step of the task.
As in prior studies of the modified prime-probe task (Grant & Weissman, 2019; Weissman, 2019), each prime and each probe can be only one of two letters in a given trial (i.e., A or B in odd trials and C or D in even trials; see the Experimental Design section for more information). Therefore, it is possible for control processes to prepare a specific incongruent response to the probe.
Including these participants in the ANOVAs reported below does not alter which main effects and interactions are significant.
Including these participants in the ANOVAs produces the same main effects and interactions that we report in the Results with one exception: the main effect of previous trial congruency is significant, F(1, 23) = 6.643 p = 0.016, , rather than merely trending (p < 0.10). However, this main effect is not relevant to testing the present hypotheses.
Specifically, we observed a simple effect of previous trial congruency on mean RT in current congruent trials (Experiment 1: F(1,23) = 93.15, p < 0.001, ; Experiment 2: F(1,23) = 12.64, p < 0.002, ) but not in current incongruent trials (Experiment 1: F(1,23) < 1; Experiment 2: F(1,23) = 1.17, p > 0.28, ).
References
- Aarts E, & Roelofs A (2011). Attentional control in anterior cingulate cortex based on probabilistic cueing. J Cogn Neurosci, 23(3), 716–727. [DOI] [PubMed] [Google Scholar]
- Behrens TE, Woolrich MW, Walton ME, & Rushworth MF (2007). Learning the value of information in an uncertain world. Nat Neurosci, 10(9), 1214–1221. [DOI] [PubMed] [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, & Cohen JD (2001). Conflict monitoring and cognitive control. Psychol Rev, 108(3), 624–652. [DOI] [PubMed] [Google Scholar]
- Braem S, Verguts T, & Notebaert W (2011). Conflict adaptation by means of associative learning. J Exp Psychol Hum Percept Perform, 37(5), 1662–1666. [DOI] [PubMed] [Google Scholar]
- Brainard DH (1997). The Psychophysics Toolbox. Spat Vis, 10(4), 433–436. [PubMed] [Google Scholar]
- Compton RJ, Huber E, Levinson AR, & Zheutlin A (2012). Is “conflict adaptation” driven by conflict? Behavioral and EEG evidence for the underappreciated role of congruent trials. Psychophysiology, 49(5), 583–589. [DOI] [PubMed] [Google Scholar]
- Dignath D, Hommel B, Johannsen L, & Kiesel A (2019). Reconciling cognitive-control and episodic-retrieval accounts of sequential conflict modulation: Binding of control-states into event-files. JEP: HPP, 45(9), 1265–1270. [DOI] [PubMed] [Google Scholar]
- Egner T (2014). Creatures of habit (and control): a multi-level learning perspective on the modulation of congruency effects. Front Psychol, 5, 1247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faul F, Erdfelder E, Lang AG, & Buchner A (2007). G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav Res Methods, 39(2), 175–191. [DOI] [PubMed] [Google Scholar]
- Frings C, Hommel B, Koch I, Rothermund K, Dignath D, Giesen C, … Philipp A (2020). Binding and Retrieval in Action Control (BRAC). Trends Cogn Sci, 24(5), 375–387. [DOI] [PubMed] [Google Scholar]
- Frings C, Rothermund K, & Wentura D (2007). Distractor repetitions retrieve previous responses to targets. Q J Exp Psychol (Hove), 60(10), 1367–1377. [DOI] [PubMed] [Google Scholar]
- Grant LD, & Weissman DH (2019). Turning distractors into targets increases the congruency sequence effect. Acta Psychol (Amst), 192, 31–41. [DOI] [PubMed] [Google Scholar]
- Gratton G, Coles MGH, & Donchin E (1992). Optimizing the use of information: strategic control of activation and responses. JEP: General, 4, 480–506. [DOI] [PubMed] [Google Scholar]
- Hommel B (1998). Event Files: Evidence for Automatic Integration of Stimulus-Response Episodes. Visual Cognition, 5(1/2), 183–216. [Google Scholar]
- Hommel B (2007). Feature integration across perception and action: event files affect response choice. Psychol Res, 71(1), 42–63. [DOI] [PubMed] [Google Scholar]
- Hommel B, Proctor RW, & Vu KP (2004). A feature-integration account of sequential effects in the Simon task. Psychol Res, 68(1), 1–17. [DOI] [PubMed] [Google Scholar]
- Hommel B, & Wiers RW (2017). Towards a Unitary Approach to Human Action Control. Trends Cogn Sci, 21(12), 940–949. [DOI] [PubMed] [Google Scholar]
- Huffman G, Hilchey MD, Weidler BJ, Mills M, & Pratt J (2020). Does feature-based attention play a role in the episodic retrieval of event files? J Exp Psychol Hum Percept Perform, 46(3), 241–251. [DOI] [PubMed] [Google Scholar]
- Janczyk M, & Leuthold H (2018). Effector system-specific sequential modulations of congruency effects. Psychon Bull Rev, 25(3), 1066–1072. [DOI] [PubMed] [Google Scholar]
- Jimenez L, & Mendez A (2014). Even with time, conflict adaptation is not made of expectancies. Front Psychol, 5, 1042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones M, Curran T, Mozer MC, & Wilder MH (2013). Sequential effects in response time reveal learning mechanisms and event representations. Psychol Rev, 120(3), 628–666. [DOI] [PubMed] [Google Scholar]
- Kim S, & Cho YS (2014). Congruency sequence effect without feature integration and contingency learning. Acta Psychol (Amst), 149, 60–68. [DOI] [PubMed] [Google Scholar]
- Kunde W, & Wuhr P (2006). Sequential modulations of correspondence effects across spatial dimensions and tasks. Mem Cognit, 34(2), 356–367. [DOI] [PubMed] [Google Scholar]
- Lee J, & Cho YS (2013). Congruency sequence effect in cross-task context: evidence for dimension-specific modulation. Acta Psychol (Amst), 144(3), 617–627. [DOI] [PubMed] [Google Scholar]
- Lim CE, & Cho YS (2018). Determining the scope of control underlying the congruency sequence effect: roles of stimulus-response mapping and response mode. Acta Psychol (Amst), 190, 267–276. [DOI] [PubMed] [Google Scholar]
- Logan GD (1985). Executive control of thought and action. Acta Psychol (Amst), 60, 193–210. [Google Scholar]
- Logan GD, & Zbrodoff JN (1979). When it helps to be misled: Facilitative effects of increasing the frequency of conflicting stimuli in a Stroop-like task. Memory & Cognition, 7(3), 166–174. [Google Scholar]
- Mayr U, Awh E, & Laurey P (2003). Conflict adaptation effects in the absence of executive control. Nature Neuroscience, 6, 450–452. [DOI] [PubMed] [Google Scholar]
- Miller EK, & Cohen JD (2001). An integrative theory of prefrontal cortex function. Annual Review of Neuroscience, 24, 167–202. [DOI] [PubMed] [Google Scholar]
- Moeller B, & Frings C (2019a). From simple to complex actions: Response-response bindings as a new approach to action sequences. J Exp Psychol Gen, 148(1), 174–183. [DOI] [PubMed] [Google Scholar]
- Moeller B, & Frings C (2019b). Response-response binding across effector-set switches. Psychon Bull Rev, 26(6), 1974–1979. [DOI] [PubMed] [Google Scholar]
- Notebaert W, & Verguts T (2006). Stimulus conflict predicts conflict adaptation in a numerical flanker task. Psychonomic Bulletin & Review, 13(6), 1078–1084. [DOI] [PubMed] [Google Scholar]
- Ridderinkhof KR (2002). Micro- and macro-adjustments of task set: activation and suppression in conflict tasks. Psychol Res, 66(4), 312–323. [DOI] [PubMed] [Google Scholar]
- Schmidt JR (2018). Evidence against conflict monitoring and adaptation: An updated review. Psychon Bull Rev. [DOI] [PubMed] [Google Scholar]
- Schmidt JR, & De Houwer J (2011). Now you see it, now you don’t: controlling for contingencies and stimulus repetitions eliminates the Gratton effect. Acta Psychol (Amst), 138(1), 176–186. [DOI] [PubMed] [Google Scholar]
- Schmidt JR, & Weissman DH (2014). Congruency sequence effects without feature integration or contingency learning confounds. PLoS One, 9(7). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt JR, & Weissman DH (2015). Contingent attentional capture triggers the congruency sequence effect. Acta Psychol (Amst), 159, 61–68. [DOI] [PubMed] [Google Scholar]
- Spape MM, & Hommel B (2008). He said, she said: episodic retrieval induces conflict adaptation in an auditory Stroop task. Psychon Bull Rev, 15(6), 1117–1121. [DOI] [PubMed] [Google Scholar]
- Stürmer B, Leuthold H, Soetens E, Schroter H, & Sommer W (2002). Control over location-based response activation in the Simon task: behavioral and electrophysiological evidence. J Exp Psychol Hum Percept Perform, 28(6), 1345–1363. [DOI] [PubMed] [Google Scholar]
- Verbruggen F, Notebaert W, Liefooghe B, & Vandierendonck A (2006). Stimulus- and response-conflict-induced cognitive control in the flanker task. Psychonomic Bulletin & Review, 13(2), 328–333. [DOI] [PubMed] [Google Scholar]
- Weissman DH (2019). Let your fingers do the walking: Finger force distinguishes competing accounts of the congruency sequence effect. Psychon Bull Rev, 26(5), 1619–1626. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Colter K, Drake B, & Morgan C (2015). The congruency sequence effect transfers across different response modes. Acta Psychol (Amst), 161, 86–94. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Colter KM, Grant LD, & Bissett PG (2017). Identifying stimuli that cue multiple responses triggers the congruency sequence effect independent of response conflict. J Exp Psychol Hum Percept Perform, 43(4), 677–689. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Egner T, Hawks Z, & Link J (2015). The congruency sequence effect emerges when the distracter precedes the target. Acta Psychologica (Amst), 156, 8–21. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Hawks ZW, & Egner T (2016). Different levels of learning interact to shape the congruency sequence effect. J Exp Psychol Learn Mem Cogn, 42(4), 566–583. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Jiang J, & Egner T (2014). Determinants of congruency sequence effects without learning and memory confounds. J Exp Psychol Hum Percept Perform, 40(5), 2022–2037. [DOI] [PubMed] [Google Scholar]
- Whitehead PS, Pfeuffer CU, & Egner T (2020). Memories of control: One-shot episodic learning of item-specific stimulus-control associations. Cognition, 199, 104220. [DOI] [PMC free article] [PubMed] [Google Scholar]