

Abstract
Accumulating evidence indicates that specific strains of mucosa-associated Escherichia coli (E. coli) can influence the development of colorectal carcinoma. This study aimed to investigate the prevalence and characterization of mucosa-associated E. coli obtained from the colorectal cancer (CRC) patients and control group. At two referral university-affiliated hospitals in northwest Iran, 100 patients, 50 with CRC and 50 without, were studied over the course of a year. Fresh biopsy specimens were used to identify mucosa-associated E. coli isolates after dithiothreitol mucolysis. To classify the E. coli strains, ten colonies per sample were typed using enterobacterial repetitive intergenic consensus-based PCR (ERIC-PCR). The strains were classified into phylogroups using the quadruplex PCR method. The PCR method was used to examine for the presence of cyclomodulin, bfp, stx1, stx2, and eae-encoding genes. The strains were tested for biofilm formation using the microtiter plate assay. CRC patients had more mucosa-associated E. coli than the control group (p < 0.05). Enteropathogenic Escherichia coli (EPEC) was also found in 23% of CRC strains and 7.1% of control strains (p < 0.05). Phylogroup A was predominant in control group specimens, while E. coli isolates from CRC patients belonged most frequently to phylogroups D and B2. Furthermore, the frequency of cyclomodulin-encoding genes in the CRC patients was significantly higher than the control group. Around 36.9% of E. coli strains from CRC samples were able to form biofilms, compared to 16.6% E. coli strains from the control group (p < 0.05). Noticeably, cyclomodulin-positive strains were more likely to form biofilm in comparison to cyclomodulin-negative strains (p < 0.05). In conclusion, mucosa-associated E. coli especially cyclomodulin-positive isolates from B2 and D phylogroups possessing biofilm-producing capacity colonize the gut mucosa of CRC patients.
1. Introduction
Colorectal cancer (CRC) is the world's third most common cancer and the second leading cause of cancer death [1]. Because of its high morbidity and mortality rate, CRC is an important public health issue [2]. Mutations that occur in tumor suppressor genes, oncogenes, and genes related to DNA repair mechanisms can lead to CRC. Depending on the origin of the mutations, colorectal carcinomas are divided into three types: sporadic (70%), familial (25%), and inherited (5%) [3].
The human gut microbiota contains over more than 1,000 microbial species, adding together 1014 microorganisms [4] that play an essential role in many important physiological processes, such as food digestion, metabolism, and immune response [5]. Shifts in the composition of these resident microbiota, the so called microbial dysbiosis, have been found to cause various diseases, including CRC [6, 7], cardiovascular disease, inflammatory bowel disease, and diabetes mellitus [6]. Approximately 20% of all cancers in humans can be related to infectious agents. Gut bacteria may be involved in the initiation or progression of sporadic CRC via a variety of mechanisms, including inducing inflammation, generating reactive oxygen species, and producing genotoxins [8]. Published literature suggests that some bacterial species such as Bacteroides fragilis [9], Streptococcus bovis, Fusobacterium spp., Helicobacter pylori, and Clostridium septicum are associated with colorectal carcinogenesis [10]. Moreover, considering the fact that E. coli is the most common facultative anaerobic resident in human gut flora, several research studies have shown a strong link between mucosa-associated E. coli and CRC [8, 11–13]. Moreover, E. coli strains are categorized into phylogenetic groups (A, B1, B2, C, D, E, and F) based on the existence or absence of a variety of genes and DNA fragment [14]. Pathogenic E. coli strains are mostly found in the B2 or D phylogroups, while commensal strains are mostly found in groups A and B1 [15].
Interestingly, enteropathogenic Escherichia coli (EPEC) is commonly found in CRC patients, in contrast to their occasional presence in the control subjects [16]. Additionally, E. coli isolates that carry the eae gene (encoding for intimin protein) are able to attach the intestinal epithelium and are classed as attaching and effacing E. coli (AEEC). More noteworthy, EPEC is also the most widely found AEEC bacteria in humans. This bacterium suppresses the expression of key DNA mismatch repair proteins (MMR), which suggests that chronic mucosal EPEC infection can contribute to the development of CRC tumors [17]. Typical EPEC strains (tEPEC) have the eae and bfp (gene encoding for bundle-forming pili) genes but no Shiga toxin genes (stx). Often known as atypical EPEC, certain clinical isolates of EPEC lack the bfp gene [18].
Usually, pathogenic E. coli strains produce several toxins called cyclomodulins including the cytotoxic necrotizing factor (CNF), cycle inhibiting factor (Cif), colibactin, and cytolethal distending toxins (CDTs). The colibactin is a toxin produced by polyketide synthetase (pks) [8]. Cyclomodulins are attracting growing attention due to their ability to influence cellular differentiation, apoptosis, and cell proliferation by disrupting the eukaryotic cell cycle and/or promoting DNA damage [19]. For example, B2 E. coli strains carrying the cyclomodulin-encoding genes are more prevalent in colon tumor biopsies in comparison to control samples suggesting a possible role of these isolates in CRC carcinogenesis [12, 20].
In addition, recent studies have shown that bacterial biofilms are associated with human colon cancer [21, 22]. Mucus-invasive bacterial biofilms, for example, were found on the colon mucosa of CRC patients more frequently than in healthy subjects [23]. Biofilm formation causes an increase in epithelial junction permeability in both normal and cancerous colons, which can facilitate direct access to mutagens from bacteria to the colonic epithelial surface and encourage procarcinogenic tissue inflammation [24]. In general, E. coli appears to play a significant role in CRC carcinogenesis, but there are little details on the characterization of colonic mucosa-associated E. coli from CRC patients. Therefore, the aim of this analysis was to compare the presence and characteristics of mucosa-associated E. coli isolates in biopsy specimens from CRC patients and a control group.
2. Materials and Methods
2.1. Patients
Between June 2019 and June 2020, 100 patients were examined at two referral university-affiliated hospitals in northwest Iran (Table 1). Among these, 50 patients had CRC, while equal number of patients without CRC had piles (n = 26), pruritus ani (n = 12), irritable bowel syndrome (IBS) (n = 2), and no symptoms (refer for screening; n = 10) were taken as controls into the study.
Table 1.
Characteristics of CRC patients and control group subjects.
| Characteristics | CRC patients (n = 50) | Control group (n = 50) | P value |
|---|---|---|---|
| Age, median (range) | 64 (40–83) | 56 (31–82) | P > 0.05 |
| Gender, male/female | 27/23 | 17/33 | P > 0.05 |
| BMI (kg/m2) (mean ± SD) | 25.6 ± 3.1 | 26.1 ± 3.3 | P > 0.05 |
Clinical features (such as weight loss, rectal bleeding, abdominal pain, and bowel habit changes), a positive fecal occult blood test, colonoscopy, and pathology findings were used to diagnose CRC. Furthermore, IBS was diagnosed based on the presence of clinical indications and symptoms (Rome criteria).
In this study, noncancerous individuals without acute or chronic inflammation were included in the control group. In addition, inclusion criteria for CRC patients were a histological diagnosis of malignant tumor without any previous treatment. Moreover, patients who had taken antibiotics in the four weeks leading up to the endoscopy were also excluded from the study.
2.2. Sample Treatment
During colonoscopy, biopsy specimens were obtained from the colon and rectum using regular endoscopic forceps. The neoplastic characteristics of the biopsy samples were verified by pathological results. The mucosal biopsy specimens were placed in an Eppendorf tube containing normal saline and transferred to the lab [16]. The samples were washed three times in 5 mL PBS to remove the fecal bacteria [12]. Then, in order to remove the mucus layer, fresh biopsy specimens were transferred to a microtube containing 500 µL of 0.016% wt/vol dithiothreitol solution and rotated for 10 minutes [25]. The obtained supernatant was inoculated in nutrient broth (NB) and incubated at 37°C for 24 hours. After the specified period, broth culture was subcultured on MacConkey agar. E. coli isolates were identified by standard conventional biochemical tests.
2.3. ERIC-PCR Typing and Phylogenetic Classification
Ten colonies of E. coli from each positive sample were typed with enterobacterial repetitive intergenic consensus-based PCR (ERIC-PCR) as described by Versalovic et al. (1991) [26]. All E. coli strains were then divided to phylogroups based on the presence of the DNA fragment (TSPE4.C2) and arpA, chuA, trpA, and yjaA genes, which were detected by the quadruplex PCR method proposed by Clermont et al. [14].
2.4. Detection of Cyclomodulin Genes and Other Virulence Factors
The PCR method was performed using primers specific for the cyclomodulin (cif, cnf1, cnf2, cnf3, cdtB-I, cdtB-II, cdtB-III, cdtB-IV, cdtB-V, and pks genomic island), clbA, clbQ [27], bfp, stx1, stx2, and intimin (eae)-encoding genes [25].
The template DNA from E. coli isolates was extracted using the boiling water method, as described previously [27]. The PCR reaction was carried out in a 50 µL mixture containing 10–100 ng of the template DNA, 0.2 mM each dNTPs, 0.5 pmol of each primer, 3 mM MgCl2, and 1.0 U of DNA polymerase (Yekta Tajhiz Azma, Iran), in the corresponding reaction buffer.
2.5. Biofilm Production Assay
Biofilm formation assays were carried out by the microtiter plate method, as described before using 2% crystal violet. Optical density (OD) of each stained well was measured at 570 nm. The cutoff OD was defined as three standard deviations above the mean OD of the negative control. TSB medium without bacteria was considered as a negative reagent control. All isolates were classified into no or weak biofilm producers, moderate biofilm producers, or high biofilm producers [28].
2.6. Statistical Analysis
In this study, the results were analyzed by the chi-square test or Fisher's exact test using SPSS software for Windows (version 23 SPSS Inc., Chicago, IL, USA). A p value less than 0.05 was considered a statistically significant difference.
3. Results
3.1. E. coli Strains in CRC and Control Group Specimens
The investigation of the presence of E. coli isolates showed that the number of specimen without E. coli was remarkably higher in control specimens (20%, n = 10/50) than in those with CRC patients (4%, n = 2/50), P = 0.028. In this study, some specimens carried more than one E. coli strains. As some specimens carried more than one E. coli strains, a total of 65 E. coli strains from CRC patients and 42 E. coli strains from the control group were taken into the study (Figure 1).
Figure 1.
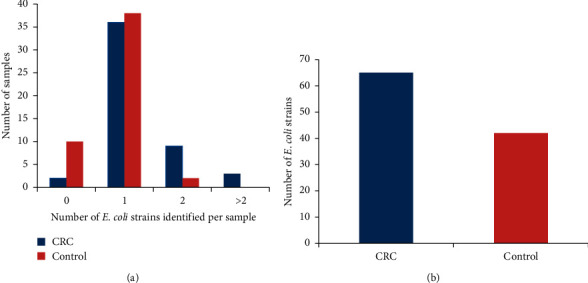
Number of E. coli strains obtained from 100 subjects comprising 50 CRC patients and the equivalent number of control subjects. (a) Some specimens did not carry any strain of E. coli and some specimens carried one or more than one E. coli strains. (b) A total of 65 E. coli strains from CRC patients and 42 E. coli strains from the control group identified.
3.2. Prevalence of EPEC in CRC and Control Group Samples
EPEC are defined as isolates that have the eae and bfp genes but no Shiga toxin genes. Often known as atypical EPEC, certain clinical isolates of EPEC lack the bfp gene.
The eae gene was detected in 23% E. coli isolated from clinical specimen obtained from the CRC group, but only in 7.1% E. coli isolated from the control group, the difference being statistically significant (p=0.036). Moreover, all eae-positive strains were negative for stx1 and stx2 genes. Interestingly, all EPEC strains were atypical EPEC (PCR negative for bfp) (Figure 2).
Figure 2.
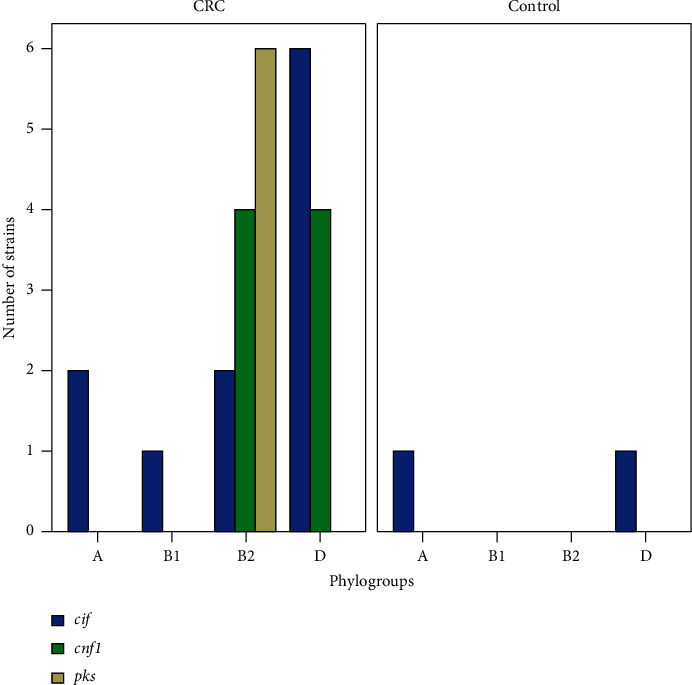
Distribution of cyclomodulin-positive E. coli in various phylogenetic groups.
3.3. Distribution of E. coli Phylogroups according to the Specimen Origin
Phylogenetic studies revealed that the prevalence of phylogroups in E. coli strains from CRC specimens and controls varied significantly. As given in Table 2, phylogroup D was the predominant type in E. coli obtained from CRC patients (41.5%), followed by phylogroups B2 (36.9%), A (13.9%), and B1 (7.7%). On the other hand, E. coli isolated in the control group most frequently belonged to phylogroup A (38.1%), followed by phylogroups B1 (19%), D (16.7%), B2 (11.9%), E (9.5%), and F (4.7%).
Table 2.
Phylogroups distribution of E. coli isolates according to specimen origins.
| Specimen types | Number (%) of E. coli strains in various phylogenetic groups | |||||
|---|---|---|---|---|---|---|
| A | B1 | B2 | D | E | F | |
| CRC patients∗ | 9 (13.9) | 5 (7.7) | 24 (36.9) | 27 (41.5) | 0 (0) | 0 (0) |
| Control group∗∗ | 16 (38.1) | 8 (19) | 5 (11.9) | 7 (16.7) | 4 (9.5) | 2 (4.7) |
∗ 65 E. coli strains were obtained from 50 CRC patients; ∗∗42 E. coli strains were obtained from 50 control group subjects.
3.4. Distribution of Cyclomodulin-Encoding Genes according to the Sample Origin and Phylogroups
As given in Table 3, the increased frequencies of cyclomodulin-encoding genes on CRC samples (30.7%, n = 20/65) compared to control tissues were statistically significant (4.7%, n = 2/42), p=0.001. It is worth noting that the cif gene was the only gene revealed (4.7%) among cyclomodulin-encoding genes in E. coli strains obtained from the control group. Among E. coli strains from CRC patients, the strains harboring cif, cnf1, and pks genes represented 16.9%, 12.3%, and 9.2% of the total strains isolated, respectively. Additionally, 7.7% (n = 5) E. coli strains from CRC patients carried more than one cyclomodulin-encoding genes. Moreover, cnf2, cnf3, and cdtB genes were not found in any of the strains from CRC specimens.
Table 3.
Distribution of cyclomodulin-positive E. coli according to phylogenetic groups.
| Percentage (number) of E. coli strains from CRC patients in diverse phylogenetic groups | |||||||
| A (n = 9) | B1 (n = 5) | B2 (n = 24) | D (n = 27) | E (n = 0) | F (n = 0) | All (n = 65) | |
|
| |||||||
| cif | 22.2 (2) | 20 (1) | 8.3 (2) | 22.2 (6) | 0 (0) | 0 (0) | 16.9 (11) |
| cnf 1 | 0 (0) | 0 (0) | 16.7 (4) | 14.8 (4) | 0 (0) | 0 (0) | 12.3 (8) |
| pks | 0 (0) | 0 (0) | 25 (6) | 0 (0) | 0 (0) | 0 (0) | 9.2 (6) |
| cdt | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Cyclomodulin-encoding genes | 22.2 (2) | 20 (1) | 50 (12)∗ | 37 (10)∗ | 0 (0) | 0 (0) | 30.7 (20)∗ |
|
| |||||||
| Percentage (number) of E. coli strains from the control group in diverse phylogenetic groups | |||||||
| A (n = 16) | B1 (n = 8) | B2 (n = 5) | D (n = 7) | E (n = 4) | F (n = 2) | All (n = 42) | |
|
| |||||||
| cif | 6.2 (1) | 0 (0) | 0 (0) | 14.3 (1) | 0 (0) | 0 (0) | 4.7 (2) |
| cnf | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| pks | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| cdt | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Cyclomodulin-encoding genes | 6.2 (1) | 0 (0) | 0 (0) | 14.3 (1) | 0 (0) | 0 (0) | 4.7 (2) |
∗ Some E. coli isolates carried more than one cyclomodulin-encoding genes.
All strains of patients in the CRC group harboring pks genomic island belonged to phylogroup B2. Moreover, 50% E. coli strains in this group harboring cnf1 gene belonged to phylogroup B2, while 50% E. coli strains belonged to phylogroup D. The distribution of phylogroups amongst E. coli strains of the CRC group possessing cif gene was as follows: 54.5% belonged to phylogroup D, followed by phylogroups A (18.2%), B2 (18.2%), and B1 (9.1%) (Table 3).
3.5. Biofilm-Forming Ability of E. coli Strains from the Control Group and CRC Specimens
As given in Table 4, in this study, there was a significant difference between the ability to form biofilms in the CRC group (36.92%, n = 24/65) and the control group (16.6%, n = 7/42), p=0.03. Amongst the biofilm-producing strains from the control group, all strains were weak biofilm producers. In contrast, amongst E. coli strains of the CRC group, 33.3% were observed as strong or moderate biofilm producers, while 66.6% were weak biofilm producers.
Table 4.
Percentage (number) of biofilm-positive isolates and their biofilm formation ability.
| Biofilm-positive strains | Biofilm formation category | ||
|---|---|---|---|
| Moderate/strong producers | Weak producers | ||
| Cyclomodulin-positive strains from CRC patients (n = 20) | 60 (12) | 50 (6) | 50 (6) |
| Cyclomodulin-negative strains from CRC patients (n = 45) | 26.6 (12) | 16.6 (2) | 83.3 (10) |
| CRC (n = 65) | 36.9 (24) | 33.3 (8) | 66.6 (16) |
| Cyclomodulin-positive strains from the control group (n = 2) | 50 (1) | 0 (0) | 100 (1) |
| Cyclomodulin-negative strains from the control group (n = 40) | 15 (6) | 0 (0) | 100 (6) |
| Control (n = 42) | 16.6 (7) | 0 (0) | 100 (7) |
The number of biofilm-positive strains was significantly higher in cyclomodulin-positive E. coli strains (60%, n = 12/2) compared to cyclomodulin-negative E. coli strains (26.6%, n = 12/45), p=0.014.
4. Discussion
CRC is one of the deadliest cancers in the world. Indeed, bacterial infection has long been known as key factor in the etiology of CRC [8]. More recently, accumulating evidence supports that mucosa-associated E. coli can affect the development of CRC [8, 29, 30]. However, there are relatively few data available about the prevalence and characterization of mucosa-associated E. coli from CRC specimens.
In our study, the colonic biopsy specimens from CRC and control patients indicated that CRC samples are more colonized by mucosa-associated E. coli strains in comparison to the control group. This result is consistent with the results of other studies [12, 31].
Additionally, EPEC was found to be more prevalent in CRC (23%) than in control samples in this study (7.1%). Magdy et al. have previously reported the presence of EPEC isolates in about 50% of CRC specimens and 20% healthy control [16]. Furthermore, recent evidence shows that chronic mucosal EPEC infection could promote molecular pathways involved in CRC carcinogenesis [17]. The higher colonization of biopsy specimens of CRC patients by mucosa-associated E. coli can be possibly explained that modifications of the colon mucosal properties in CRC lead to increased expression of adhesion and internalization of pathogens in the tumor microenvironment [8].
In the current study, the distribution of E. coli phylogroups in CRC patient was different from the control group. Most strains from CRC patients belonged to B2 and D phylogroups, while about 58% of the strains from the control group belonged to A and B1 phylogroups. Thus, it can be concluded that the intestinal mucosa of CRC patients is mainly colonized by E. coli strains that are more virulent. Buc et al. [12] and Raisch et al. [11] previously identified higher numbers of E. coli phylogroup B2 in CRC specimens than in controls in two studied. However, no major difference in the abundance of E. coli belonging to the phylogenetic group D was found between patients with CRC and diverticulosis in their studies. Group B2 E. coli strains can cause severe infections because their genetic background is favorable for obtaining the high number of virulence factors [32]. Additionally, macrophages are one of the main tumor-infiltrating immune cells which have a pivotal role in cancer progression. Interestingly, E. coli of the B2 phylogroup can survive and multiply within macrophages and promote protumoral activities of macrophages independently of colibactin production [33].
In the current research, CRC patients had a substantially higher frequency of cyclomodulin-positive E. coli than the control group, comparable to other studies performed by Bonnet et al. [8], Raisch et al. [11], and Buc et al. [12]. Overall, the findings indicated that cyclomodulin-positive E. coli isolates are implicated in the development of CRC.
We observed cyclomodulin-positive E. coli strains in only 4.7% of the control samples, while Buc et al. [12] and Bonnet et al. [8] observed these isolates in 19.4% and 26% of control samples, respectively. The high frequency of cyclomodulin-positive E. coli in their studies compared to ours may be due to they used diverticulosis patients as a control group and dysbiosis of gut microbiota has been reported in diverticulosis patients [34].
In our study, the prevalence of pks-positive E. coli isolated from CRC patients was about 10%, which in contrast to previous research studies is significantly lower. Other research studies have reported pks-positive E. coli isolated in CRC patients to range from 26% to 67% [8, 11, 12, 35]. pks + E. coli isolates are attracting attention because of their ability to induce DNA damage in epithelial cells, leading to genomic instability of mammalian cells [36–40] and thus promote development of colorectal cancer in animal models [35]. Over the course of 30 years, E. coli isolates from fecal samples of populations living in industrialized countries showed a phylogenetic shift from group A toward group B2. Moreover, about 30% of this commensal B2 E. coli isolates have acquired pks island by horizontal transfer [41].
Presence of cyclomodulin-encoding gene cif was revealed in 17% E. coli strains belonging to the CRC patients group. In addition, 12.3% of E. coli strains were positive for cnf gene. More noteworthy, amongst cyclomodulin-encoding genes, cif was the most common in our study, in contrast to other research studies whereby pks has been reported as the most common cyclomodulin-encoding gene [11, 12]. Thus, our study revealed that in addition to pks, other cyclomodulins especially cif and cnf might play an important role in development of CRC.
In this analysis, the number of biofilm-producing E. coli strains was higher in CRC samples than in control samples (p < 0.05). Furthermore, as compared to E. coli strains from healthy mucosa, CRC patients' E. coli isolates were high biofilm producers. Interestingly, we also found that cyclomodulin-positive E. coli isolates were more likely to generate biofilm than cyclomodulin-negative isolates in the current research. In this regard, Tomkovich et al. have previously reported the presence of mucus-invasive biofilms on the colon mucosa of about half CRC samples and in only 13% of control specimens [23]. In addition, Dejea et al. for the first time displayed that bacterial biofilms are associated with CRC. Presence of biofilms in tissue may correlate with bacterial invasion and changes in colon tissue biology with increased cellular proliferation [42].
The predominance of adhesin-encoding genes, particularly pili adhesins, that encourage colonization of carcinogenic strains in the intestinal tissue or colorectal tumor microenvironment was not detected in this investigation. It also does not examine into the cyclomodulin-producing strains' carcinogenic potential in animal studies.
As a result, we must examine the frequency and expression of adhesin-encoding genes in E. coli strains isolated from CRC patients and controls in the near future. In addition, we must investigate the ability of cyclomodulin-producing strains to interact with intestinal epithelial cells, as well as their potential carcinogenic effect in animal models.
5. Conclusion
Our study showed that E. coli isolates obtained from the CRC patients and controls are a heterogeneous group of isolates with variations in virulence factors, phylogroups, and biofilm-forming capacity. To summarize, CRC samples were more colonized by mucosa-associated E. coli, especially cyclomodulin-positive isolates from B2 and D phylogroups with the ability to form biofilms in comparison to the control group. This is the largest study of the prevalence and characterization of mucosa-associated E. coli isolated from CRC patients and a control group in Iran that we are aware of.
Acknowledgments
This study was supported by Infectious and Tropical Diseases Research Center, Tabriz University of Medical Sciences, Tabriz, Iran (62957).
Data Availability
The authors declare the data used to support the findings of this study are available from the corresponding author upon request.
Ethical Approval
This study was approved by the Research Ethics Committee (IR.TBZMED. REC.1398.409) at Tabriz University of Medical Sciences, Tabriz, Iran.
Disclosure
This is a report of a database from Ph.D. thesis of the first author registered in the Tabriz University of Medical Sciences.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Authors' Contributions
Roghayeh Nouri, Mohammad Ahangarzadeh Rezaee, and Alka Hasani conceptualized and designed the study. Kourosh Masnadi Shirazi, Bita Sepehri, Simin Sotoodeh, and Afshin Fattahzadeh involved in acquisition of data and sampling. Rohgayeh Nouri, Mohammad Reza Alivand, Babak Abdinia, and Fatemeh Hemmati analyzed and interpreted the study. Roghayeh Nouri and Mohammad Ahangarzadeh Rezaee reviewed the original draft. Mohammad Ahangarzadeh Rezaee and Alka Hasani supervised, reviewed, and edited the study. All authors accepted the final form of the manuscript.
References
- 1.Baidoun F., Elshiwy K., Elkeraie Y. Colorectal cancer epidemiology: recent trends and impact on outcomes. Current Drug Targets . 2020;2174 doi: 10.2174/1389450121999201117115717. [DOI] [PubMed] [Google Scholar]
- 2.Favoriti P., Carbone G., Greco M., Pirozzi F., Pirozzi R. E. M., Corcione F. Worldwide burden of colorectal cancer: a review. Updates in Surgery . 2016;68(1):7–11. doi: 10.1007/s13304-016-0359-y. [DOI] [PubMed] [Google Scholar]
- 3.Mármol I., Sánchez-de-Diego C., Pradilla Dieste A. Colorectal carcinoma: a general overview and future perspectives in colorectal cancer. International Journal of Molecular Sciences . 2017;18(1):p. 197. doi: 10.3390/ijms18010197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Rubinstein M. R., Wang X., Liu W., Hao Y., Cai G., Han Y. W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host & Microbe . 2013;14(2):195–206. doi: 10.1016/j.chom.2013.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ai D., Pan H., Han R., Li X., Liu G., Xia L. C. Using decision tree aggregation with random forest model to identify gut microbes associated with colorectal cancer. Genes . 2019;10(2):p. 112. doi: 10.3390/genes10020112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fan X., Jin Y., Chen G., Ma X., Zhang L. Gut microbiota dysbiosis drives the development of colorectal cancer. Digestion . 2021;102(4):508–515. doi: 10.1159/000508328. [DOI] [PubMed] [Google Scholar]
- 7.Genua F., Raghunathan V., Jenab M. The role of gut barrier dysfunction and microbiome dysbiosis in colorectal cancer development. Frontiers in Oncology . 2021;11(1016) doi: 10.3389/fonc.2021.626349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bonnet M., Buc E., Sauvanet P., et al. Colonization of the human gut by E. coli and colorectal cancer risk. Clinical Cancer Research . 2014;20(4):859–867. doi: 10.1158/1078-0432.ccr-13-1343. [DOI] [PubMed] [Google Scholar]
- 9.Haghi F., Goli E., Mirzaei B., Zeighami H. The association between fecal enterotoxigenic B. fragilis with colorectal cancer. BMC Cancer . 2019;19(1):879–884. doi: 10.1186/s12885-019-6115-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gagnière J., Raisch J., Veziant J., et al. Gut microbiota imbalance and colorectal cancer. World Journal of Gastroenterology . 2016;22(2):501–18. doi: 10.3748/wjg.v22.i2.501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Raisch J., Buc E., Bonnet M. Colon cancer-associated B2Escherichia colicolonize gut mucosa and promote cell proliferation. World Journal of Gastroenterology . 2014;20(21):p. 6560. doi: 10.3748/wjg.v20.i21.6560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Buc E., Dubois D., Sauvanet P., et al. High prevalence of mucosa-associated E. coli producing cyclomodulin and genotoxin in colon cancer. PLoS One . 2013;8(2):p. e56964. doi: 10.1371/journal.pone.0056964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bernier C., Gounon P., Le Bouguénec C. Identification of an aggregative adhesion fimbria (AAF) type III-encoding operon in enteroaggregative Escherichia coli as a sensitive probe for detecting the AAF-encoding operon family. Infection and Immunity . 2002;70(8):4302–4311. doi: 10.1128/iai.70.8.4302-4311.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Clermont O., Christenson J. K., Denamur E., Gordon D. M. The ClermontEscherichia coliphylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environmental Microbiology Reports . 2013;5(1):58–65. doi: 10.1111/1758-2229.12019. [DOI] [PubMed] [Google Scholar]
- 15.Kohoutova D., Smajs D., Moravkova P., et al. Escherichia coli strains of phylogenetic group B2 and D and bacteriocin production are associated with advanced colorectal neoplasia. BMC Infectious Diseases . 2014;14(1):733–738. doi: 10.1186/s12879-014-0733-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Magdy A., Elhadidy M., Abd Ellatif M. E., et al. Enteropathogenic Escherichia coli (EPEC): does it have a role in colorectal tumourigenesis? A prospective cohort study. International Journal of Surgery . 2015;18:169–173. doi: 10.1016/j.ijsu.2015.04.077. [DOI] [PubMed] [Google Scholar]
- 17.Maddocks O. D. K., Short A. J., Donnenberg M. S., Bader S., Harrison D. J. Attaching and effacing Escherichia coli downregulate DNA mismatch repair protein in vitro and are associated with colorectal adenocarcinomas in humans. PLoS One . 2009;4(5):p. e5517. doi: 10.1371/journal.pone.0005517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jomehzadeh N., Ahmadi K., Javaherizadeh H. The first evaluation relationship of integron genes and the multidrug-resistance in class A ESBLs genes in enteropathogenic Escherichia coli strains isolated from children with diarrhea in Southwestern Iran. Molecular Biology Reports . 2020;2020:1–7. doi: 10.1007/s11033-020-06047-5. [DOI] [PubMed] [Google Scholar]
- 19.Veziant J., Gagnière J., Jouberton E., et al. Association of colorectal cancer with pathogenicEscherichia coli: focus on mechanisms using optical imaging. World Journal of Clinical Oncology . 2016;7(3):p. 293. doi: 10.5306/wjco.v7.i3.293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yoshikawa Y., Tsunematsu Y., Matsuzaki N. Characterization of colibactin-producing Escherichia coli isolated from Japanese patients with colorectal cancer. Japanese Journal of Infectious Diseases . 2020;73:2020–2066. doi: 10.7883/yoken.JJID.2020.066. [DOI] [PubMed] [Google Scholar]
- 21.Tomkovich S., Gharaibeh R. Z. Human colon mucosal biofilms and murine host communicate via altered mRNA and microRNA expression during cancer. mSystems . 2020;5(1):e00451–e00419. doi: 10.1128/mSystems.00451-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Coleman O., Haller D. Microbe–mucus interface in the pathogenesis of colorectal cancer. Cancers . 2021;13(4):p. 616. doi: 10.3390/cancers13040616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tomkovich S., Dejea C. M., Winglee K. Human colon mucosal biofilms from healthy or colon cancer hosts are carcinogenic. Journal of Clinical Investigation . 2019;129(4):1699–1712. doi: 10.1172/jci124196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cheng Y., Ling Z., Li L. The intestinal microbiota and colorectal cancer. Frontiers in Immunology . 2020;11:p. 3100. doi: 10.3389/fimmu.2020.615056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Martin H. M., Campbell B. J., A Hart C. Enhanced Escherichia coli adherence and invasion in Crohn’s disease and colon cancer. Gastroenterology . 2004;127(1):80–93. doi: 10.1053/j.gastro.2004.03.054. [DOI] [PubMed] [Google Scholar]
- 26.Versalovic J., Koeuth T., Lupski R. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucleic Acids Research . 1991;9(24):6823–6831. doi: 10.1093/nar/19.24.6823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Dubois D., Delmas J., Cady A. Cyclomodulins in urosepsis strains of Escherichia coli. Journal of Clinical Microbiology . 2010;48(6):2122–2129. doi: 10.1128/jcm.02365-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Stepanović S., Vuković D., Hola V. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis . 2007;115(8):891–899. doi: 10.1111/j.1600-0463.2007.apm_630.x. [DOI] [PubMed] [Google Scholar]
- 29.Salesse L., Lucas C., Hoang M. H. Colibactin-producing Escherichia coli induce the formation of invasive carcinomas in a chronic inflammation-associated mouse model. Cancers . 2021;13(9):p. 2060. doi: 10.3390/cancers13092060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Iyadorai T., Mariappan V., Vellasamy K. Prevalence and association of pks+ Escherichia coli with colorectal cancer in patients at the University Malaya Medical Centre, Malaysia. PLoS One . 2020;15(1):p. e0228217. doi: 10.1371/journal.pone.0228217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Swidsinski A., Khilkin M., Kerjaschki D. Association between intraepithelial Escherichia coli and colorectal cancer. Gastroenterology . 1998;115(2):281–286. doi: 10.1016/s0016-5085(98)70194-5. [DOI] [PubMed] [Google Scholar]
- 32.Escobar-Páramo P., Clermont O., Blanc-Potard A.-B. A specific genetic background is required for acquisition and expression of virulence factors in Escherichia coli. Molecular Biology and Evolution . 2004;21(6):1085–1094. doi: 10.1093/molbev/msh118. [DOI] [PubMed] [Google Scholar]
- 33.Raisch J., Rolhion N., Dubois A. Intracellular colon cancer-associated Escherichia coli promote protumoral activities of human macrophages by inducing sustained COX-2 expression. Laboratory Investigation . 2015;95(3):296–307. doi: 10.1038/labinvest.2014.161. [DOI] [PubMed] [Google Scholar]
- 34.Barbara G., Scaioli E., Barbaro M. R. Gut microbiota, metabolome and immune signatures in patients with uncomplicated diverticular disease. Gut . 2017;66(7):1252–1261. doi: 10.1136/gutjnl-2016-312377. [DOI] [PubMed] [Google Scholar]
- 35.Arthur J. C., Perez-Chanona E., Mühlbauer M. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science . 2012;338(6103):120–123. doi: 10.1126/science.1224820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zeng H., Umar S., Rust B. Secondary bile acids and short chain fatty acids in the colon: a focus on colonic microbiome, cell proliferation, inflammation, and cancer. International Journal of Molecular Sciences . 2019;20(5):p. 1214. doi: 10.3390/ijms20051214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Secher T., Samba-Louaka A., Oswald E. Escherichia coli producing colibactin triggers premature and transmissible senescence in mammalian cells. PloS One . 2013;8(10):p. e77157. doi: 10.1371/journal.pone.0077157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Iftekhar A., Berger H., Bouznad N. Genomic aberrations after short-term exposure to colibactin-producing E. coli transform primary colon epithelial cells. Nature Communications . 2021;12(1):1–15. doi: 10.1038/s41467-021-21162-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dziubańska-Kusibab P., Berger H., Battistini F. Colibactin DNA-damage signature indicates mutational impact in colorectal cancer. Nature Medicine . 2020;26(7):1063–1069. doi: 10.1038/s41591-020-0908-2. [DOI] [PubMed] [Google Scholar]
- 40.Pleguezuelos-Manzano C., Puschhof J., Huber A. R. Mutational signature in colorectal cancer caused by genotoxic pks+ E. coli. Nature . 2020;580(7802):269–273. doi: 10.1038/s41586-020-2080-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Payros D., Secher T., Boury M. Maternally acquired genotoxic Escherichia coli alters offspring’s intestinal homeostasis. Gut Microbes . 2014;5(3):313–512. doi: 10.4161/gmic.28932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Dejea C. M., Wick E. C., Hechenbleikner E. M. Microbiota organization is a distinct feature of proximal colorectal cancers. Proceedings of the National Academy of Sciences . 2014;111(51):18321–18326. doi: 10.1073/pnas.1406199111. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors declare the data used to support the findings of this study are available from the corresponding author upon request.