

Abstract
Scientist and technologist have long sought to advance limb prostheses that connect directly to the peripheral nervous system, enabling a person with amputation to volitionally control synthetic actuators that move, stiffen and power the prosthesis, as well as to experience natural afferent sensations from the prosthesis. Recently, the agonist-antagonist myoneural interface (AMI) was developed, a mechanoneural transduction architecture and neural interface system designed to provide persons with amputation improved muscle-tendon proprioception and neuroprosthetic control. In this paper, we provide an overview of the AMI, including its conceptual framing and preclinical science, surgical techniques for its construction, and clinical efficacy related to pain mitigation, phantom limb range of motion, fascicle dynamics, central brain proprioceptive sensorimotor preservation, and prosthetic controllability. Following this broad overview, we end with a discussion of current limitations of the AMI and potential resolutions to such challenges.
Key Words: amputation, neuroprosthetic control, prosthesis
CONCEPTUAL FRAMING
The clinical standard-of-care for limb amputation surgery has changed little in over 2 millennia, and is not currently optimized to facilitate agonist-antagonist muscle dynamics and neural integration with powered limb prostheses.1–3 In a traditional amputation surgery, native muscle tissues are configured isometrically in a layered closure around the transected bone, forming a padded distal region for prosthetic socket usage.4,5 Such a traditional amputation disrupts natural agonist-antagonist dynamic interactions between native musculature within the amputated residuum. Furthermore, the traditional amputation fails to provide agonist-antagonist muscle end organs for transected nerves that once innervated muscles distal to the amputation site.3 These surgical characteristics limit the ability of spindle and Golgi tendon organs within affected musculature to communicate meaningful muscle-tendon length, speed and force information to the central nervous system (CNS).
Such a disruption of natural agonist-antagonist muscle dynamics is problematic for efferent control and musculotendinous proprioceptive signaling to and from an external limb prosthesis.3 The fundamental motor unit in humans is not a single muscle, but rather an agonist-antagonist muscle pair. Since a skeletal muscle can pull and not push, at least 2 opposing muscles are required to bi-directionally move and position a biological joint, wherein this positioning is largely dependent on the dynamic relationship between muscle spindle afferents in agonist-antagonist muscle groups acting simultaneously on the same degree-of-freedom.6,7 In addition to biological joint positioning, an opposing muscle-tendon pair can independently control joint impedance through the modulation of muscle co-activation and nonlinear tendon engagement.8 Thus, for a neuroprosthetic control system to enable such an independent controllability of prosthetic joint position and impedance while also providing natural musculotendinous proprioceptive sensations to the prosthetic wearer, we believe the limb amputation procedure should ideally maintain natural agonist-antagonist muscle dynamics for each external prosthetic degree-of-freedom to be controlled. Specifically, the amputation paradigm and neural interfacing approach should transmit the length, speed, and force of each agonist-antagonist muscle-tendon as an efferent command to the external prosthesis, and in turn, artificial proprioceptive-type signals from the external prosthesis, such as artificial joint position, speed and torque, should be transmitted through natural neural pathways to provide physiologically relevant musculotendinous proprioceptive afferents to the prosthetic wearer. Although substituting or recreating musculotendinous sensory perception through external vibrotactile or electrical nerve methods has been explored9–11, restoring natural proprioceptive sensations has not been broadly implemented in clinical practice. Natural musculotendinous proprioception is a critical component of motor feedback,12,13 and its restoration may improve prosthetic motor performance for persons with amputation.
During the Italian-Ethiopian war of 1896, an Italian physician named Giuliano Vanghetti imagined the first neuroprosthetic controller designed to achieve these goals. Vanghetti was the first scientist to conceive of the idea that native muscles within the amputated residuum could actually be mechanically linked to an external prosthesis to form a direct tissue-to-mechatronic mechanical coupling. The modern term for this neural interfacing technique is cineplasty.14–24 In this approach, skin-lined tendon or muscle tunnels are surgically created during the amputation procedure, and then post-amputation, the tunnels are mechanically connected to an external prosthetic joint through chains, cables, or straps. Such a direct muscle to prosthesis mechanical coupling allows patients to exert muscular forces directly onto the external prosthesis, and in principle, to experience muscle-tendon afferent feedback when the moving prosthetic joint stretches the opposing antagonistic muscle. Although some clinical benefits have been reported, cineplasty is no longer used in surgical and prosthetic practice because of its clinical challenges of tunnel skin irritation, infections, and muscle fatigue during the execution of prosthetic motor tasks.19,21 Further, cineplasty only harnesses the control of native musculature within the amputated residuum, providing no anatomical interface for muscle-tendon proprioceptive feedback and neuroprosthetic control for transected nerves that once innervated muscles distal to the amputation site.
As a resolution of these challenges, in 2014 the agonist-antagonist myonerural interface (AMI) was invented at the Massachusetts Institute of Technology.25,26 In distinction to cineplasty, the AMI framework does not directly connect muscle-tendon units to an external mechatronic prosthesis. Rather, an AMI is a surgically constructed agonist-antagonist muscle pair and neural interfacing strategy wherein muscle-tendon information from the pair is transmitted to the external prosthesis using artificial sensing and wired or wireless transmission technology. An illustration of the AMI is shown in Figure 1. When the AMI agonist is electrically depolarized either from a descending CNS efferent signal or from artificial muscle stimulations, the muscle contracts and mechanically stretches the surgically linked antagonist (Fig. 1B). The AMI muscle dynamics, as sensed by spindle and Golgi tendon organ afferents, provides the person with limb amputation natural proprioceptive sensations. Using various artificial sensing modalities, the AMI muscle dynamics are measured, and these sensory data are communicated using wired or wireless neural communications to an external prosthetic computer to inform a closed-loop powered prosthetic control. For every robotic degree-of-freedom to be controlled within the external prosthesis, at least one AMI muscle pair is constructed (Fig. 1C).
FIGURE 1.

The agonist-antagonist myoneural interface.
SYSTEM-LEVEL DESIGN
Figure 2 shows a control diagram of the proposed neuroprosthetic communications between AMI constructs and an external prosthesis. Contraction of the AMI agonist muscle occurs through the standard motor nerve efferent command, stretching the linked antagonist muscle. Artificial sensory signals of the AMI muscles’ lengths, speeds and forces are then transmitted to a biophysical controller that employs an intact biological limb model for its computations. In one proposed approach, magnetomicrometry is used for sensing AMI muscles’ lengths, speeds, and forces.27,28 Here two small magnetic beads are implanted into each AMI muscle, and tracked using an array of magnetometers positioned adjacent the AMI construct outside the body on the skin surface or attached to a prosthetic socket. A magnet tracking algorithm computes the distance between each magnet (muscle length), the time rate of change of that distance (muscle speed), and bead lateral vibrations (muscle force). The intact biological limb model converts these measured linear AMI muscle signals into desired prosthetic joint position, impedance, and torque signals. To position, stiffen and power the external prosthetic joints, the biophysical controller then applies a closed-loop control to servo to these desired values using computed errors between the desired signals and the actual measured sensory signals from the prosthesis. The AMI efferent agonist activation will simultaneously activate the native contractile mechanoreceptors in the Golgi tendon organ of the agonist muscle, as well as the native intrafusal muscle spindle stretch fibers of the mechanically coupled antagonist muscle, both of which will provide afferent proprioceptive signaling to the CNS. Subsequent volitional efferent activation of the AMI antagonist will stimulate a complimentary stretch on the AMI agonist. As such, the AMI construct is hypothesized to provide a more realistic agonist-antagonist mechanical coupling as compared with a traditional amputation paradigm, given its designed capacity to provide non-isometric fascicle strains and agonist-antagonist mechanoreceptor feedbacks to the CNS.
FIGURE 2.
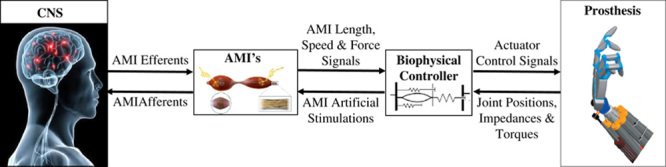
Bidirectional neural communication between agonist-antagonist myoneural interface (AMI) constructs and a powered external prosthesis.
The AMI framework is also designed to provide a closed-loop feedback from the prosthesis into the CNS (Fig. 2). Using electrodes placed on each AMI muscle, or on the nerve innervating each muscle, the biophysical controller applies functional electrical stimulation (FES) for prosthetic force and length feedback from the external prosthesis (alternatively, functional optical stimulation could be used to artificially stimulate AMI musculature for feedback control.29). By applying closed-loop FES on the antagonist as the agonist contracts, the force on the agonist is controlled by the biophysical controller based upon torque sensory information from the corresponding prosthetic joint. For example, when an upper extremity prosthetic user picks up a barbell weight and flexes her prosthetic wrist, the AMI corresponding to wrist flexors/extensors can be electrically stimulated so the user can experience the bar bell weight as a proprioceptive sensation. As the AMI agonist muscle contracts, with a motor nerve supply that once innervated the wrist flexors before limb amputation, FES artificial stimulations are applied to the AMI antagonist muscle, increasing the force borne by the agonist flexor. The magnitude of the FES stimulation is proportional to the force error between the measured AMI force and a scaled estimate of that force that would have been applied by the wrist flexors against the bar bell load before limb amputation.
Alternatively, a closed-loop artificial stimulation applied by the biophysical controller can exert a position control on the AMI using the measured AMI fascicle lengths and speeds. In the case where an external agent is positioning the external bionic joint, such positions would have to be reflected on the AMI muscles in order for the prosthetic user to receive accurate proprioceptive feedback. For example, if a person shakes hands with a person wearing a bionic arm, such a handshake may forcibly change the positions of the bionic joints. Bionic joint state sensory information would serve as control position and speed targets for the artificial stimulation applied to the AMI muscles by the bionic limb’s biophysical controller. For example, if the handshake flexed the bionic wrist, the biophysical controller would receive bionic wrist state information from a synthetic wrist sensor, and apply an electrical stimulation to the agonist AMI muscle proportional to the error between the measured bionic wrist position/speed and the measured position/speed from muscle fiber state magnetic sensors, causing the muscle to contract and the antagonist to stretch. The prosthetic user would then experience the position of their bionic wrist as imposed by the handshake through an afferent feedback to the CNS from muscle spindle receptors in the AMI muscle pair.
PRECLINICAL SCIENCE
Spanning the period from 2016 to 2019, the AMI was scientifically evaluated at Massachusetts Institute of Technology in preclinical experiments using murine and caprine animal models.30–33 In these experiments, both native30,31 and regenerative32,33 AMI constructs were investigated. A native AMI comprises 2 muscles with native vascularization and innervation that are surgically connected in series to form an agonist-antagonist pair.30,31 In distinction, a regenerative AMI is constructed onto tw2o transected nerves that, in the native intact anatomy, innervate muscles that move a joint in opposing directions (eg, flexion/extension, inversion/eversion). Here small devascularized and denervated muscle grafts are surgically placed within, or overlayed upon, the 2 transected nerves, and then are mechanically linked through a passive material (eg, tendon) in series to form an opposing muscle pair.32,33
For both native or regenerative AMI constructs, and for both murine and caprine animal models, these preclinical studies show that the AMI agonist muscle produces a contraction correlated with the electrical activation of its motor nerve, resulting in a graded agonist electromyography signal and a correlated physiological-level of stretch in the AMIs antagonist muscle. The studies also show that the AMI provides afferent neural feedback that monotonically increases with increasing antagonist muscle stretch caused by the contraction of the linked agonist muscle. These results suggest that the AMI has the potential to communicate meaningful musculotendinous proprioceptive feedback from a prosthetic limb by replicating the agonist-antagonist muscle dynamics that are fundamental to physiological proprioception.
SURGICAL IMPLEMENTATION IN HUMANS
In July 2016, bolstered by this foundational preclinical research, Brigham & Women’s Hospital (BWH) performed a first-in-human amputation employing an AMI surgical design.34,35 The first AMI subject was a 50-year-old male who, 2 years prior, sustained multiple injuries from a 50-foot fall while rock climbing. Following extensive screening and patient education, he was deemed appropriate to undergo the AMI surgery at the transtibial level. The patient’s operative procedure (subsequently branded the Ewing amputation) was uncomplicated and included construction of 2 native AMI constructs. For prosthetic ankle dorsi-plantarflexion control, a first AMI was constructed by linking the tibialis anterior (TA) to the lateral gastrocnemius, and for prosthetic subtalar inversion-eversion control, a second AMI was built by linking the peroneus longus to the tibialis posterior (Fig. 3). For both AMI constructs, a passive tendon connects each opposing muscle. To form a sliding pulley that enables low friction agonist-antagonist muscle movements, each tendon passes through a synovial canal. The synovial canals as well as embedded tendons for both AMIs were harvested from the lateral or medial tarsal tunnels in the discarded ankle.35 As of the writing of this paper, this transtibial AMI amputation procedure has been conducted on a total of 25 limbs in 22 patients.
FIGURE 3.

The transtibial agonist-antagonist myoneural interface amputation.
Later in 2018, BWH performed a first-in-human AMI amputation at the transfemoral level.36 In this procedure, native AMIs were constructed for the ankle and subtalar joints using the same muscle pairs as in the Ewing amputation; these were complimented, however, by the additional construction of a knee joint AMI comprised of the rectus femoris and lateral head of the biceps femoris. The ankle and subtalar AMI muscles were recruited through isolation and mobilization of their discrete neurovascular pedicles—in essence, requiring the establishment of 4 neurovascular muscle island flaps that were subsequently configured circumferentially around the distal thigh musculature. Tarsal tunnel recruitment was utilized for the 2 lower AMIs, as in the Ewing amputation, but provision of a gliding canal for the knee AMI was accomplished through utilization of ankle retinaculum and discarded muscle fascia.
In addition to these lower extremity acute amputation procedures, currently AMI constructs are being incorporated into the design of upper extremity amputations at both the transradial and transhumeral levels, as well as for revision procedures for lower extremity amputations.
CLINICAL EFFICACY
Reduced Pain, Enhanced Phantom Range of Motion, and Increased Fascicle Strains
In the study of Srinivasan et al37 the authors characterized the physiological outcomes of subjects with an AMI transtibial amputation (N=15) and compared the outcomes against those of matched controls (age and time-since-amputation matched) having a traditional transtibial amputation (N=7). For pain assessment, the PROMIS Sleep Disturbance 6a tool and Pain Interference 6a survey37,38 were administered orally to all subjects. AMI subjects reported on average less pain (12.25±4.9) as compared with the traditional cohort (17.29±10.22) (P=0.08), and 6 of the 15 subjects in the AMI cohort indicated zero pain in all categories.
For range of motion assessments of the phantom ankle-foot complex, in37 a 2 degree-of-freedom wireless goniometer was secured to the posterior aspect of the unaffected leg of the unilateral study subjects and used to synchronously measure ankle-foot kinematics for the unaffected limb. For these experiments, subjects were asked to emulate phantom movements onto the contralateral ankle-foot anatomy, mirroring the range of motion of the phantom joints through unaffected limb positioning. The demonstrated range of motion of the phantom limb of the ankle and subtalar joints in each direction (plantar flexion, dorsiflexion, inversion, eversion) were normalized as a percentage of the unaffected joints’ range of motion. AMI subjects experienced significantly greater range of motion than traditional subjects for plantar flexion, dorsiflexion, and eversion directions, but not for inversion (P<0.05).37
To assess fascicle dynamics, in Srinivasan et al37 ultrasound imaging was used for both AMI and traditional subjects. The AMI ultrasound data showed coupled agonist-antagonist motions within each ankle and subtalar AMI. With the exception of one traditional amputee study participant, who had undergone an Ertl osteomyoplastic amputation, dynamic coupling of antagonistic muscles was limited in traditional subjects. To quantify changes in muscle length, the fascicle strains from the antagonist muscle were measured during agonist contraction. Significantly greater fascicle strains were produced in AMI subjects (average strains ranging from 4% to 11%) as compared with traditional controls (average strains ranging from 1% to 5%) (P<0.05), validating the mechanistic goal of the AMI to enhance fascicle strains for agonist-antagonist muscle interactions.
Enhanced Central Brain Proprioceptive Signaling
The central brain undergoes adaptations in function after conventional limb amputation.39–41 In the study of Srinivasan et al,42 researchers compared results from the fMRI neuroimaging of individuals with the AMI amputation, a traditional amputation, and no amputation. Individuals with a traditional amputation demonstrated a significant decrease in proprioceptive activity, measured by activation of Brodmann area 3a, whereas functional activation in AMI subjects was not significantly different from no amputation controls (P<0.05). In this same study, Srinivasan et al42 the degree of proprioceptive activity in the central brain strongly correlated with fascicle strain levels in the peripheral muscles and performance on motor tasks (P<0.05), supporting the hypothesis that increased levels of agonist-antagonist muscle dynamics enhances musculotendinous proprioception and motor function. These results suggest that surgical techniques designed to restore anatomical musculotendenous agonist-antagonist dynamics result in desirable central sensorimotor plasticity.
Improved Neural Prosthetic Controllability
As noted earlier, the transtibial AMI surgical design comprises 2 native AMIs within the residual limb, one designated to control a prosthetic ankle joint and the second to control a prosthetic subtalar joint.34,35 One AMI, composed of the tibialis posterior and the peroneus longus, was designed to control the prosthetic subtalar joint responsible for prosthetic inversion and eversion movements. A second AMI, composed of the lateral gastrocnemius and the TA, was designed to control the prosthetic ankle joint, responsible for prosthetic plantar flexion and dorsiflexion movements. In the study of Clites et al34 surface electrodes or percutaneous needle electrodes were placed adjacent to, or within, each of the four AMI muscles, and these electromyography control signals were then transmitted to a 2-degree-of-freedom ankle-foot prosthesis comprising ankle and subtalar prosthetic degrees of freedom. In this preliminary study, an AMI subject displayed improved free-space controllability over his prosthesis compared with a group of 4 subjects having traditional amputation. The study also showed natural ankle-foot reflexive behaviors during stair ambulation in the AMI subject that was not demonstrated in the 4 subjects having traditional amputation. Furthermore, for the first time, the study of Clites et al34 demonstrated the capacity of the AMI system to apply a closed-loop joint torque control. In response to torque measured on the prosthesis, microprocessors on the prosthetic leg commanded artificial stimulations to the antagonist muscle within the AMI, controlling the force borne on the mechanically coupled agonist (Fig. 2). Specifically, when the AMI subject applied a volitional plantar flexion torque to compress a spring-loaded foot pedal, the subject could only compress the pedal at 4 significantly distinct levels when the onboard computer converted the measured prosthetic ankle torque into FES signals applied to the subject’s AMI TA muscle. When the FES muscle stimulations were turned off, the AMI subject could no longer control the ankle torque and compress the pedal to 4 significantly distinct torque levels (P<0.05). These results underscore the potential importance of a neural torque feedback from prosthetic sensors into the peripheral nervous system.
This preliminary control study34 motivated a broader study37 comprising a larger cohort of AMI patients. In the study of Srinivasan et al37 the authors characterized the control authority inherent to subjects with an AMI transtibial amputation (N=15) compared with that of matched control subjects with a traditional transtibial amputation (N=7). In a positional differentiation task, subjects were instructed to move their phantom to 0%, 25%, 50%, 75%, and 100% of the range of motion for both the ankle and subtalar joints without visual feedback, forcing each subject to use their musculotendinous proprioception from the affected limb. The authors hypothesized that subjects with greater control and proprioception would be able to position their phantom more distinctly and with greater precision. Statistical analyses demonstrated that AMI subjects had more significant discrimination in every category except between 75% and 100% plantar flexion as compared with traditional subjects. Further, AMI subjects demonstrated significantly less variance while performing the task when compared with traditional subjects. These results suggest that AMI subjects have more precise and distinct motor control of their residual muscles, likely informed by greater fascicle strains and proprioceptive afferents. Such an improved motor controllability may enable AMI subjects to have more distinct neuroprosthetic control over their external powered prostheses.
CURRENT LIMITATIONS OF THE AMI APPROACH
Although the AMI offers clinical advantages, there exist several important limitations. First, the only way to apply a controlled force onto the agonist is by artificially stimulating that agonists’ muscle antagonist, or vise versa. Since both muscles are innervated, such artificial activations cause afferent signaling of muscle length, speed and force of the artificially stimulated muscle, causing the user to feel the muscle activation and dynamics of both agonist-antagonist muscles simultaneously. In distinction, within an intact biological limb, forces are applied to muscle from many different sources, such as gravitation and inertia. For example, during a walking gait, after the heel impacts the ground surface, the ankle joint typically plantar flexes, causing the TA to stretch. Such a TA stretch is not caused by calf muscle activation and force generation; in fact, the calf muscle is typically not active during that phase of gait. To provide such a TA stretch using the AMI construct, the calf muscle linked to the TA would have to be artificially stimulated, but since the AMI calf muscle would be innervated in this case, the user would feel the calf muscle activation and dynamics, creating an artificial sensation to the prosthetic wearer.
A second limitation of the AMI approach relates to the inability to emulate the biological transmission coupling all the muscles that span any particular biological joint. In an intact limb, when a muscle contracts, and that contraction moves a biological joint or joints, such movement causes other muscles that span that same joint(s) to either be shortened or lengthened by varying degrees. Such a mechanical mapping from one muscle to the next is referred to herein as a joint or joints’ biological transmission. Each muscle’s origin and insertion locations, the load on each muscle-tendon, and the moment arms across which each muscle acts define the biological transmission. With the AMI approach, a muscle agonist is physically attached to a single antagonist using either a linear arrangement, such as shown in Figure 1, or across a pulley that re-directs the line of muscle action using, for example, a synovial canal.34,35 Once an AMI muscle pair is created, the transmission from the agonist to its single antagonist is fixed and is not likely to emulate the natural, intact limb transmission. Furthermore, even if multiple AMI muscle pairs are created for a single biological degree-of-freedom in order to capture natural muscle-tendon redundancy, an agonist contraction will still not create a natural level of muscle stretches in all biologically coupled antagonistic muscles spanning that joint, since there is only a mechanically linked coupling across each AMI, and not across multiple AMI muscle pairs. Given these difficulties, when an AMI agonist contracts, it causes an unnatural level of stretch across its antagonistic muscle(s), creating unnatural sensations for the person with amputation.
A third limitation of the AMI approach relates to difficulties associated with constructing a regenerative AMI.32 For a proximal limb amputation, either above-knee or above elbow, a 2-stage surgical procedure is required to surgically construct regenerative AMIs because of a lack of reliable knowledge regarding the discrete spatial organization of proximal nerves relative to their former muscle targets.33 In a first surgery, a large proximal nerve undergoes a fascicular split wherein the large nerve trunk is split into finer fascicles.3,33 At the transected end of each fascicle, a muscle graft is placed. After several months, each nerve fascicle regenerates into its respective graft, and the graft becomes fully vascularized. During the first surgery, the muscle grafts cannot be linked into agonist-antagonist muscle pairs because the surgeon does not know a priori which muscle corresponds to which joint movement direction of any particular joint (eg, ankle dorsiflexion/plantar flexion). Hence, a second stage surgery has to be performed after each muscle graft has been innervated, and each innervated muscle is electrophysiologically mapped to determine which muscle corresponds to each distal joint movement direction (eg, flexion, extension, inversion, eversion, etc.).
A fourth limitation of the AMI framework is that it fails to offer a broad spectrum of afferents for neuroprosthetic control, limited only to musculotendinous proprioception while not providing critical afferents such as cutaneous contact, pressure and shear.
MECHANONEURAL INTERFACES
As a possible resolution to these difficulties, a mechanoneural interface (MI) is proposed.43 The MI is a biological-synthetic interface designed to link a human peripheral nerve to a powered prosthesis. In this framework, a muscle actuator modulates under computer control the length, speed and force applied to an innervated tissue end organ, such as skin or muscle, to directly control the mechanoneural transduction into the CNS. The MI muscle actuator is mechanically coupled to its end organ and is computer controlled using a biophysical controller. The controller applies closed-loop artificial stimulations through an actuator muscle electrode using MI artificial sensory signals as feedback to directly modulate the length, speed and force applied to the end organ and thus the afferent signaling from a prosthetic appliance. There are 2 types of MI, namely the cutaneous mechanoneural interface (CMI) and the proprioceptive mechanoneural interface (PMI).
The CMI comprises a muscle actuator coupled to a natively-pedicled skin flap in a cuff-like architecture. Through neuroprosthetic electrical closed-loop stimulation, the muscle is actuated to induce strains or oscillatory vibrations on the skin flap, proportional to the contact or pressure measured by artificial sensors on an external prosthesis. Native dermal mechanotransducers generate the corresponding afferent signals, enabling a natural cutaneous sensory experience for the prosthetic user. Using regenerative, surgical and biomechatronic techniques, for each skin region that is to be represented in a prosthetic control system, one CMI is constructed within the affected limb. The CMI was recently evaluated in a preclinical study using a murine animal model.44 In the study, the CMI successfully elicited 4 levels of graded contact and eight distinct vibratory afferents that were insignificantly different from the analogous mechanical stimulation of intact skin, underscoring the CMIs capacity to recreate an array of cutaneous sensations.
The PMI comprises a muscle actuator connected in series to an innervated muscle end organ using a passive material connection for electrical isolation such as biological tendon, ligament, or a biocompatible material such as acellular dermal matrix. For each muscle that is to be represented in a prosthetic control system, one PMI is constructed within the affected limb. For the generalized PMI, the muscle end organ and series muscle actuator comprise an artificial electrode and sensors; an electrode is attached to the muscle actuator for FES control, and implantable sensors measure end organ length, speed and force for closed-loop feedback control using, for example, magnetomicrometry.27,28 Under closed-loop computer control, the muscle actuator applies controlled forces and displacements onto the innervated muscle organ to directly modulate proprioceptive afferents. In addition, the muscle end organ of each PMI provides efferent neural signaling for neuroprosthetic control.
For both the CMI and PMI, the muscle actuator is controlled through artificial stimulation without the human prosthetic wearer experiencing unnatural proprioceptive sensations caused by the actuation itself. In one approach, the muscle actuator is without an innervating nerve, and thus it can be activated through microprocessor-controlled artificial stimulations without an afferent signal. Here a stimulating electrode would apply electrical stimulations to elicit muscle contractions for the purpose of controlling the forces and displacements applied to the coupled end organ, as well as to maintain the mass and contractility of muscle actuator itself. Alternatively, the muscle actuator could be innervated using a cutaneous nerve taken from a denervated patch of skin; in this approach, artificial stimulations of the muscle actuator would then not induce proprioceptive signaling. In turn, the cutaneous nerve innervating the muscle actuator, in combination with artificial muscle stimulations, would serve to maintain its contractility and morphology.45 Finally, the muscle actuator could be innervated using a standard motor nerve wherein, during muscle actuator use, an electrical nerve stimulation enables a rapid, localized block of unnatural afferent conduction.46
Figure 4 shows a generalized architecture for the MI framework incorporating n PMIs for n neutrally linked muscle end organs, and mCMIs for m neurally-linked cutaneous skin regions. A critical feature of the agonist PMI is that its muscle actuator can apply forces onto its agonist muscle end organ independent of antagonist muscle activation, or vise versa. For example, consider the example of heel strike in a walking gait cycle when the TA is actively stretched within an intact limb during controlled plantar flexion. To emulate this phase of gait for a person with transtibial amputation, the active TA PMI within the amputated leg residuum would comprise the innervated TA as the muscle end organ and a muscle actuator attached in series with the TA. Using a neuromechanical biophysical limb model, the biophysical controller would estimate the length, speed and force that would be experienced by the TA if in fact the TA were physically coupled across the prosthetic ankle. Such an estimate could be achieved using the biophysical model with prosthetic sensory inputs such as prosthetic ankle position, speed, and torque, as well as ground reaction force and center-of-pressure. These biomimetic muscle dynamic parameters would then be used as targets for the TA PMI actuator control applied to the TA end organ. Through such a TA PMI control action, TA proprioceptive transduction would be controlled without antagonist calf muscle activations, providing a natural proprioceptive experience during the controlled plantar flexion phase of walking.
FIGURE 4.

Bidirectional neural communication between mechanoneural interface (MI) constructs and a powered external prosthesis. The MI framework comprises n proprioceptive mechanoneural interfaces (PMIs) and m cutaneous mechanoneural interfaces (CMIs) that provide efferent-afferent bidirectional control to the external prosthesis through a biophysical controller, providing the prosthetic wearer volitional efferent control as well as musculotendinous and cutaneous afferent sensations from the prosthesis. CSN indicates central nervous system.
In addition to these potential advantages, the PMI is designed to closely emulate the biological transmission coupling all the muscles that span any particular biological joint. In distinction to the AMI approach, agonist-antagonist muscle dynamics for the PMI system are not created by physically attaching muscles together. Rather, each PMI muscle end organ is virtually attached to all other PMI muscle end organs that span any particular virtual joint using an accurate, intact biophysical limb model of the missing limb. Thus, when an agonist PMI end organ contracts, the biophysical controller will command a natural set of fascicle strains for all its antagonistic end organ muscles, creating in software natural dynamic sensations of 3D biological joint movements for a person with amputation. Still further, since agonist-antagonist PMI end organs are not attached physically but through a biophysical model, regenerative PMI constructs can be created in a single stage surgery. Once PMI muscle graft innervation has occurred, end organ activity can be correlated to phantom limb movements (eg, flexion, extension, inversion, eversion, etc.) and the biophysical controller can then be updated to virtually link in software each agonist-antagonist pair for the creation of complex 3D biological joint movement afferents.
The MI framework is designed to mitigate current AMI limitations. However, for either the CMI or PMI approaches to be successful, high fidelity, closed-loop muscle actuation is required through artificial stimulation. To that end, for the full implementation of the MI approach, research progress has to be made in biological muscle sensing, artificial stimulation of muscle, closed-loop feedback controllers, and neural transmission. Research areas such as muscle magnetomicrometry,27,28 functional optical stimulation,29 neuroflexive control,47 and osseointegration48 may therefore prove critical to the efficacy of MIs.
SUMMARY
Society has long sought to advance brain controlled limb prostheses that enable a high fidelity volitional control with natural afferent percepts. In this paper, we provide an overview of the AMI, including its conceptual framing and preclinical science, surgical techniques for its construction, and clinical efficacy related to pain mitigation, phantom limb range of motion, fascicle dynamics, central brain proprioceptive sensorimotor preservation and prosthetic controllability. Following this overview, we provide a discussion of AMI limitations and potential solutions to such challenges. In the advancement of neuroprosthetic control systems, we feel that surgical, regenerative and neural interfacing technologies that harness biological mechanoneural transduction are of paramount importance.
Footnotes
H.H. and M.J.C. are currently receiving grant income from the Darpa Haptix Program (W911NF-17-2-0043), CDMRP (PRORP-CTA OR160165A; PRORP-CTA OR170384; PRORP-CTA OR180114), Defense Medical Research and Development Program (W81XWH-19-1-0151) and the NIH (1R01HD097135-01). The authors have 2 submitted patents on the Agonist-antagonist Myoneural Interface and its application to prosthetic control.
Contributor Information
Hugh Herr, Email: hherr@media.mit.edu.
Matthew J. Carty, Email: mcarty@bwh.harvard.edu.
REFERENCES
- 1.Tooms R.Crenshaw A. Amputations. Campbell’s Operative Orthopedics, 7th ed. St. Louis, MO: Mosby-Year Book; 1987:597–637. [Google Scholar]
- 2.Murdoch G, Wilson AJ. Amputation: Surgical Practice and Patient Management. St Louis, MO: Butterworth-Heinemann Medical; 1996. [Google Scholar]
- 3.Herr HM, Clites T, Srinivasan S, et al. Reinventing extremity amputation in the era of functional limb restoration. Ann Surg. 2021; 273:269–279. [DOI] [PubMed] [Google Scholar]
- 4.Eidt J, Kalapatapu V.Collins KA. Lower extremity amputation: techniques and results. Rutherford’s Vascular Surgery, 7th ed. London: Saunders Elsevier; 2010:135–161. [Google Scholar]
- 5.Ovadia SA, Askari M. Upper extremity amputations and prosthetics. Semin Plast Surg. 2015;29:55–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ribot-Ciscar E, Roll JP. Ago-antagonist muscle spindle inputs contribute together to joint movement coding in man. Brain Res. 1998;791:167–176. [DOI] [PubMed] [Google Scholar]
- 7.Dimitriou M. Human muscle spindle sensitivity reflects the balance of activity between antagonistic muscles. J Neurosci. 2014;34:13644–13655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hogan N. Adaptive control of mechanical impedance by coactivation of antagonist muscles. IEEE Trans Automat Control. 1984;29:681–690. [Google Scholar]
- 9.Goodwin GM, Mccloskey DI, Matthews PBC. The contribution of muscle afferents to kinaesthesia shown by vibration induced illusions of movement and by the effects of paralysing joint afferents. Brain. 1972;95:705–748. [DOI] [PubMed] [Google Scholar]
- 10.Goodwin GM, Mccloskey DI, Matthews PBC. Proprioceptive illusions induced by muscle vibration: contribution by muscle spindles to perception? Science. 1972;175:1382–1384. [DOI] [PubMed] [Google Scholar]
- 11.Tan DW, Schiefer MA, Keith MW, et al. A neural interface provides long-term stable natural touch perception. Sci Transl Med. 2014;6:257ra138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Riemann BL, Lephart SM. The sensorimotor system, part II: the role of proprioception in motor control and functional joint stability. J Athl Train. 2002;37:80–84. [PMC free article] [PubMed] [Google Scholar]
- 13.Kandel ER, Schwartz JH, Jessell TM, et al. Movement. Princ Neural Sci. 2013;5:1414–1434. [Google Scholar]
- 14.Alldredge RH. The cineplastic method in upper-extremity amputations. J Bone Joint Surg Am. 1948;30A:359–373. [PubMed] [Google Scholar]
- 15.Spittler AW, Rosen IE. Cineplastic muscle motors for prostheses of ARM amputees. J Bone Joint Surg Am. 1951;33:601–632. [DOI] [PubMed] [Google Scholar]
- 16.Spittler AW, Woodard GS. Cineplasty for arm amputees. Am J Nurs. 1953;53:802–805. [PubMed] [Google Scholar]
- 17.Kessler H, Gelb J. Pectoral cineplasty. Plast Reconstr Surg. 1954;13:10–19. [DOI] [PubMed] [Google Scholar]
- 18.Brav EA, Luscombe HB, Wi B, et al. Cineplasty: an end-result study. J Bone Joint Surg Am. 1957;39:59–76. [PubMed] [Google Scholar]
- 19.Mazet R. Cineplasty: historical review, present status, and critical evaluation of sixty-four patients. J Bone Joint Surg Am. 1958;40-A:1389–1400. [PubMed] [Google Scholar]
- 20.Brav EA, Macdonald WF, Woodard GH, et al. Follow-up notes on articles previously published in the journal. Cineplasty—ten years later. J Bone Joint Surg Am. 1964;46:1137–1138. [PubMed] [Google Scholar]
- 21.Weir RF. A century of the Sauerbruch-Lebsche-Vanghetti muscle cineplasty: the U.S. experience. 9thWorld Congr Int Soc Prosthetics Orthot. 1998;7:198–199. [Google Scholar]
- 22.Weir RF, Heckathorne CW, Childress DS. Cineplasty as a control input for externally powered prosthetic components. J Rehabil Res Dev. 2001;38:357–363. [PubMed] [Google Scholar]
- 23.Nambu S, Ikebuchi M, Taniguchi M, et al. Advantages of externally powered prosthesis with feedback system using pseudo-cineplasty. J Rehabil Res Dev. 2014;51:1095–1102. [DOI] [PubMed] [Google Scholar]
- 24.Tropea P, Mazzoni A, Micera S, et al. Giuliano Vanghetti and the innovation of “cineplastic operations”. Neurology. 2017;89:1627–1632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Herr H, Riso R, Song K, et al. Inventors; Peripheral neural interface via nerve regeneration to distal tissues. US patent US20160346099. 2016.
- 26.Herr H, Clites T, Maimon B, et al. Inventors; Method and system for providing proprioceptive feedback and functionality mitigating limb pathology. US patent 62/276422. 2019.
- 27.Taylor C, Abramson H, Herr H. Low-latency tracking of multiple permanent magnets. IEEE Sensors J. 2019;19:11458–11468. [Google Scholar]
- 28.PhD Thesis. Cameron Taylor. Massachusetts Institute of Technology. Media Arts & Technology. 2020.
- 29.Srinivasan S, Maimon B, Diaz M, et al. Closed-loop functional optogenetic stimulation. Nat Commun. 2018;9:5303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Clites T, Carty M, Srinivasan S, et al. A murine model of a novel surgical architecture for proprioceptive muscle feedback and its potential application to control of advanced limb prostheses. J Neural Eng. 2017;14:036002. [DOI] [PubMed] [Google Scholar]
- 31.Clites TR, Carty MJ, Srinivasan SS, et al. Caprine models of the agonist-antagonist myoneural interface implemented at the above- and below-knee amputation levels. Plast Reconstr Surg. 2019;144:218e–229e. [DOI] [PubMed] [Google Scholar]
- 32.Srinivasan SS, Carty M, Calvaresi PW, et al. On prosthetic control: a regenerative agonist antagonist myoneural interface. Sci Robotics. 2017;2:eaan2971. [DOI] [PubMed] [Google Scholar]
- 33.Srinivasan S, Diaz M, Carty M, et al. Towards functional restoration for persons with limb amputation: a dual-stage implementation of regenerative agonist-antagonist myoneural interfaces. Sci Rep. 2019;9:1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Clites T, Carty M, Ullauri J, et al. Proprioception from a neurally controlled lower extremity prosthesis. Sci Transl Med. 2018. doi: 10.1126/scitranslmed.aap8373. [DOI] [PubMed] [Google Scholar]
- 35.Clites T, Herr H, Srinivasan S, et al. The Ewing amputation: the first human implementation of the agonist-antagonist myoneural interface. Plast Reconstr Surg. 2018;6:e1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Srinivasan S, Herr H, Clites T, et al. Agonist-antagonist myoneural interfaces in above-knee amputation preserves distal joint function and perception. Ann Surg. 2021;273:e115–e118. [DOI] [PubMed] [Google Scholar]
- 37.Srinivasan S, Gutierrez-Arango S, Chia-En Teng A, et al. Neural interfacing architecture enables enhanced motor control and residual limb functionality postamputation. Proc Natl Acad Sci U S A. 2021;118:e2019555118. [Google Scholar]
- 38.PROMIS Scoring Manuals. 2021. Available at: https://www.healthmeasures.net/promis-scoring-manuals.
- 39.Bruurmijn M, Pereboom I, Vansteensel MJ, et al. Preservation of hand movement representation in the sensorimotor areas of amputees. Brain. 2017;140:3166–3178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bramati IE, Rodrigues EC, Simões EL, et al. Lower limb amputees undergo long-distance plasticity in sensorimotor functional connectivity. Sci Rep. 2019;9:1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhang J, Zhang Y, Wang L, et al. Brain functional connectivity plasticity within and beyond the sensorimotor network in lower-limb amputees. Front Hum Neurosci. 2018;12:403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Srinivasan S, Tuckute G, Zou J, et al. Agonist-antagonist myoneural interface amputation preserves proprioceptive sensorimotor neurophysiology in lower limbs. Sci Transl Med. 2020;12:eabc5926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Herr H, Song H, Srinivasan S. Inventors; Mechanoneural interfaces for prosthetic control. US patent provisional Ref. No.: 22366TE. 2020.
- 44.Srinivasan S, Herr H. A cutaneous mechanoneural interface for neuroprosthetic feedback. 2021. Available at: 10.1038/s41551-020-00669-7. [DOI] [PubMed] [Google Scholar]
- 45.Nghiem BT, Sando IC, Hu Y, et al. Sensory protection to enhance functional recovery following proximal nerve injuries: current trends. Plast Aesthet Res. 2015;2:202–207. [Google Scholar]
- 46.Patel YA, Butera RJ. Challenges associated with nerve conduction block using kilohertz electrical stimulation. J Neural Eng. 2018;15:031002. [DOI] [PubMed] [Google Scholar]
- 47.Geyer H, Herr H. A muscle-reflex model that encodes principles of legged mechanics produces human walking dynamics and muscle activities. IEEE Transac Neural Syst Rehabil Engg. 2010;18:263–273. [DOI] [PubMed] [Google Scholar]
- 48.Ortiz-Catalan M, Håkansson B, Brånemark R. An osseointegrated human-machine gateway for long-term sensory feedback and motor control of artificial limbs. Sci Transl Med. 2014;6:257re6. [DOI] [PubMed] [Google Scholar]