

ABSTRACT
Circular RNAs (circRNAs) are an evolutionarily conserved form of noncoding RNA with covalently closed loop structures. Initial studies established a functional role for circRNAs as potent microRNA sponges and many other studies have focussed solely on this. However, the biological functions of most circRNAs are still undetermined and other functional roles are gaining traction. These include protein sponges and regulators, and coding for proteins with an alternative mechanism of translation, potentially opening up a whole new transcriptome. The first step to gaining insight into circRNA function is accurate identification and various software platforms have been developed. Specialized detection software has now evolved into whole bioinformatics pipelines that can be used for detection, de novo identification, functional prediction, and validation of circRNAs. However, few cardiovascular circRNA studies have utilized these tools. This review summarizes current knowledge of circRNA biogenesis, bioinformatic detection tools and the emerging role of circRNAs in cardiovascular disease.
KEYWORDS: Circular RNA, circRNA, non-coding RNA, backsplicing, bioinformatics, cardiovascular disease
Introduction
The first endogenous circRNAs in humans were reported in the early 1990s as non-polyadenylated transcripts with a ‘scrambled’ exon structure (i.e. exons joined at consensus splice sites but in a different order to the primary pre-mRNA transcript) [1,2]. As most early transcriptomic studies used isolation methods that enriched for poly-A transcripts, circular RNAs, with their lack of poly-A tails, went undetected. However, with the evolution of next-generation sequencing kits that interrogated ribosomal-depleted total RNA (rather than poly-adenylated RNA), studies identifying and functionally characterizing circRNAs quickly proliferated. Fast forward 30 years and we now know that circRNAs can contain exonic and/or intronic sequences [3,4], are conserved across species and are associated with many different disease states, including cardiovascular disease [5–8]. Despite these advances, our understanding of their functional roles is still in its infancy and for many circRNAs, the functional mechanism has not been assigned.
CircRNA transcripts may be located in either the nucleus or the cytoplasm. Whereas exon-intron and intronic circRNAs remain in the nucleus and often appear to regulate the gene encoding them (parental gene) [9,10], exonic circRNAs are more commonly exported to the cytoplasm where they exert multiple functions [4]. CircRNAs are highly stable due to a lack of free ends that are vulnerable to exonuclease activity and can accumulate in non-proliferating tissues [11] and biofluids including plasma, saliva and urine [12–14].
This review is divided into four sections. The first section provides a brief summary of the current knowledge of circRNA biogenesis, focussing on the most recent developments in the field. The second section provides a comprehensive list of current bioinformatic detection and downstream in silico functional annotation tools. The third section describes circRNA functions, including roles as miRNA sponges, protein sponges, protein scaffolds and protein-coding transcripts. The final section discusses the emerging roles of circRNAs in cardiovascular disease, including as biomarkers and regulatory molecules in atherosclerosis, myocardial infarction/ischaemia-reperfusion injury, heart failure, cardiac hypertrophy, and cardiac fibrosis.
Biogenesis
Most human genes produce multiple circularized transcripts, a process termed alternative circularization [3,4,8]. In contrast to splicing of pre-mRNA to linear mRNA, exon-containing circRNAs are formed from non-canonical splicing of linear pre-mRNA, whereby a downstream 5ʹ splice site is joined to an upstream 3ʹ splice site in what is known as ‘back-splicing’. This results in a 3ʹ, 5ʹ phosphodiester bond at the back-splicing junction forming a covalently closed, circular transcript and a linear transcript with a skipped exon [15] (Figure 1(a)). Mutation of the canonical splice sites or inhibition of splicing by isoginkgetin (a general inhibitor of both the major and minor spliceosomes) diminishes circRNA production [16,17], suggesting that back-splicing of circRNAs utilizes the canonical spliceosome and the same splice sites as linear splicing.
Figure 1.
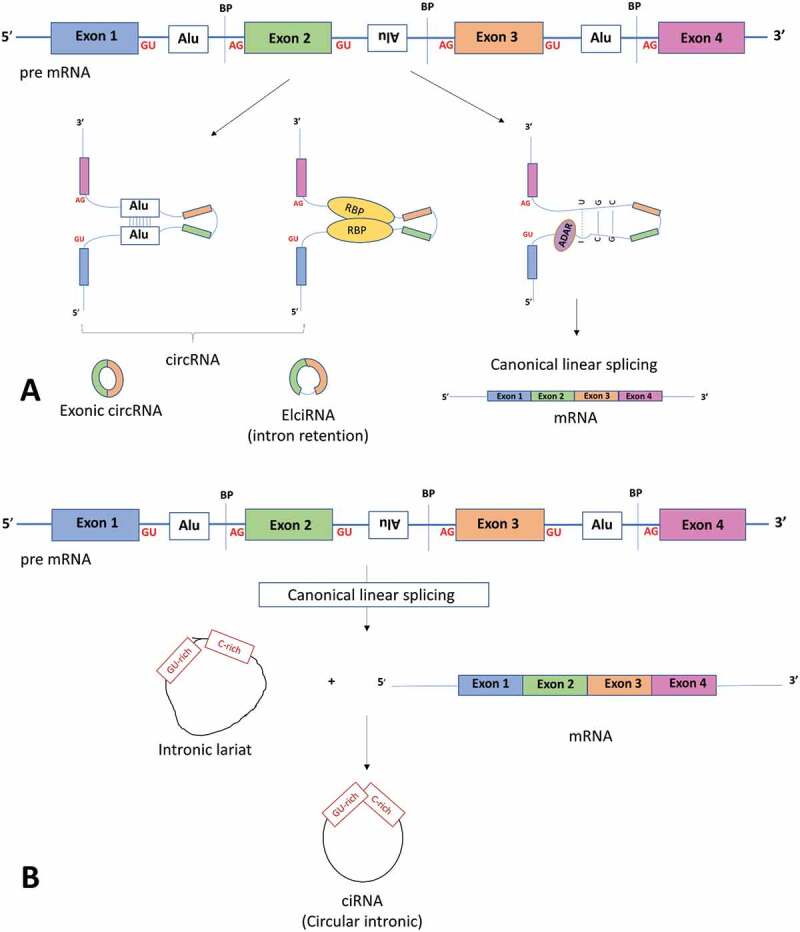
Components involved with linear mRNA or circular RNA biogenesis. (a) The top panel shows a pre-mRNA transcript with four exons. Long flanking introns containing inverted repeat elements such as Alu elements or trans-acting RBP proteins bring the downstream splice site into close proximity with the upstream splice site to favour back splicing and thus circRNA production. Conversely, introns bound by ADAR1 ‘melt’ RNA secondary structures by disrupting intronic base pairing to disrupt circularization. (b) Intronic circRNA biogenesis: Linear splicing produces lariat structures that are usually debranched and hydrolysed. However, GU rich and C rich intronic motifs escape debranching to form ciRNAs (intronic circRNAs). ADAR adenosine deaminases acting on RNA, BP: Branch points. RBP: RNA binding proteins. ALU: Alu repeat elements, mRNA messenger RNA
Certain genomic features appear to favour circRNA biogenesis. First, single exon circRNAs tend to have longer exons on average (~3-fold longer [18]), whereas multi-exon circRNAs are of more regular length. Second, exceptionally long introns that contain inverted repeat elements are significantly enriched in circRNA loci compared to linear controls, with nearly 90% of circRNAs in humans having ALU repeats in their flanking introns [19]. Third, conserved binding motifs for trans-acting RNA-binding proteins (RBPs) in flanking introns promote circularization by allowing RBP-associated intronic base pairing to facilitate the interaction of a downstream intron with an upstream intron (Figure 1(a)). Conversely, the RNA-editing enzyme, adenosine deaminase acting on RNA (ADAR) appears to inhibit circularization by ‘melting’ the secondary structures within the introns that facilitate circRNA biogenesis, thereby promoting the formation of linear transcripts (Figure 1(a)) [19]. Fourth, complementary binding of inverted repeat sequences or binding of RBP proteins promotes circularization by bringing the back-splice junctions into close proximity. This allows circularization of either the exon(s) only or, to a lesser extent, the exon with a retained intron (termed circRNAs containing exonic and intronic sequences, EIciRNAs, Figure 1(a)). Along with EIciRNAs, ciRNAs (circRNAs containing exclusively intronic sequences) are thought to reside in the nucleus and regulate their parental gene [9,10].
In contrast to exon-containing circRNAs, ciRNAs are generated during linear splicing when the 5ʹ splice site of the excised intron joins to an adenosine at the branch point, forming a lariat structure (Figure 1(b)). Lariat structures are usually removed by a debranching enzyme that hydrolyses the 2ʹ-5ʹ phosphodiester bond at the branch point to produce linear molecules. ciRNAs appear to be protected from the debranching process by consensus RNA motifs – a GU-rich sequence near the 5ʹ splice site and a C-rich sequence near the branch point (Figure 1(b)) – although the precise mechanism is unknown [10].
For export of exonic circRNAs from the nucleus to the cytoplasm the cell employs one of two pathways depending on circRNA length [20]. Initial work in Drosophila found that depletion of DExH/D box RNA helicase Hel25E resulted in accumulation of long circRNAs in the nucleus. Two human orthologues of this gene were found which shared 90% identity with Hel25E – UAP56 which regulates export of long (>800 nt) circRNAs and URH49 which regulates short circRNAs [20].
In summary, the current model of circRNA biogenesis is that back splicing is promoted by a looping of the intron(s) bringing the (back) splice sites into close proximity. This ‘looping’ appears to be facilitated by longer introns, inverted repeat sequences and RNA binding proteins [3,4,21]. Given that the same spliceosome and splice sites are used and that many circRNAs consist of exons, it may be tempting to speculate that circRNA biogenesis could be a regulator of mRNA production. However, accumulating evidence shows that circRNAs have their own function independent to that of the linear transcript.
CircRNA detection
CircRNA detection became possible as sequencing libraries evolved from poly-A enriched to ribosomal rRNA-depleted total RNA. Furthermore, circRNAs could be enriched by the addition of 3′-5′ exonuclease Ribonuclease R (RNAse R) which degrades linear RNA (although certain circRNAs are sensitive to RNAse R treatment [22,23]). However, RNAse R does not remove RNA with highly structured 3ʹ ends such as small nuclear RNAs (snRNAs) and histone mRNAs, or RNAs with G-rich G-quadruplex (G4) secondary structures, and circRNA library preparation methods were further refined, using RNAse R treatment followed by polyadenylation of transcripts and then poly(A)+ RNA depletion [24–26].
Following RNA sequencing, bioinformatic detection of circRNAs remains challenging. For a list of circRNA detection and functional prediction software see Table 1. While numerous programmes are freely available, there appears to be limited overlap in the circRNAs identified, suggesting a high rate of false positives and that combining outputs from several algorithms is recommended. The reader is directed to the following reviews for a detailed comparison between circRNA software programmes [27,28]. However, the field is rapidly evolving, and several programmes have been updated and new programmes developed since these articles were published.
Table 1.
Software for circular RNA detection and downstream applications
|
Software for Circular RNA detection | ||||
|---|---|---|---|---|
| Tool | Category | Aligner | Link | Reference |
| CIRCexplorer CIRCexplorer2 CIRCexplorer3/CLEAR |
Split alignment based | TopHat2, Bowtie, (STAR) |
https://pypi.org/project/CIRCexplorer/ https://circexplorer2.readthedocs.io/en/latest/ https://github.com/YangLab/CLEAR |
[10,36,37] |
| CIRI CIRI2 |
Split alignment based | BWA |
https://sourceforge.net/projects/ciri/ https://sourceforge.net/projects/ciri/files/CIRI2/ |
[38,39] |
| DCC | Split alignment based | STAR | https://github.com/dieterich-lab/DCC | [40] |
| find_circ | Split alignment based | Bowtie2 | https://github.com/marvin-jens/find_circ | [59] |
| MapSplice MapSplice2 |
Split alignment based | Bowtie | http://www.netlab.uky.edu/p/bioinfo/MapSplice2 | [41] |
| segemehl | Split alignment based | Segemehl | https://www.bioinf.uni-leipzig.de/Software/segemehl/ | [42] |
| UROBORUS | Split alignment based | TopHat2, Bowtie | https://github.com/WGLab/UROBORUS | [43] |
| ACFS | Pseudoreference based | BWA-MEM | https://github.com/arthuryxt/acfs | [22] |
| CircRNAFisher | Pseudoreference based | Bowtie2 | https://github.com/duolinwang/CircRNAFisher | [44] |
| KNIFE | Pseudoreference based | Bowtie, Bowtie2 | https://github.com/lindaszabo/KNIFE | [45] |
| BIQ | No alignment | Kmers | https://github.com/pmenzel/biq | |
| CircDBG | No alignment | Kmers | https://github.com/lxwgcool/CircDBG | [34] |
| CircMarker | No alignment | Kmers | https://github.com/lxwgcool/CircMarker | [46] |
| Software for validation and functional analysis of Circular RNAs | ||||
| ACValidator | In silico validation of circular RNAs | https://github.com/tgen/ACValidator | [47] | |
| CircCode | Identifying circRNA coding ability | https://github.com/PSSUN/CircCode | [48] | |
| CirComPara | Detect, quantify, and correlate expression of linear and circular RNAs from RNA-Seq data | https://github.com/egaffo/CirComPara | [49] | |
| CircInteractome | Explore potential interactions of circRNAs with RBPs, design specific divergent primers to detect circRNAs, study tissue- and cell-specific circRNAs, identify gene-specific circRNAs, explore potential miRNAs interacting with circRNAs, and design specific siRNAs to silence circRNAs | https://circinteractome.irp.nia.nih.gov/ | [26] | |
| CircPrimer | A software for annotating circRNAs and determining specificity of circRNA primers | http://www.bioinf.com.cn/ | [50] | |
| CircPro | Identification of circRNAs with protein-coding potential | http://bis.zju.edu.cn/CircPro/ | [51] | |
| CircRNAwrap | CircRNA identification, transcript prediction, and abundance estimation | https://github.com/liaoscience/circRNAwrap | [52] | |
| CircTools | CircRNA identification, RBP enrichment screenings, circRNA primer design, miRNA seed analysis and differential exon usage | https://github.com/dieterich-lab/circtools | [53] | |
| miARma | MiRNA, mRNA and circRNA analysis | http://miarmaseq.idoproteins.com/ | [54] | |
| NCLcomparator | Post-screening non-co-linear transcripts (circular, trans-spliced, or fusion RNAs) for high confidence selection | https://github.com/TreesLab/NCLcomparator | [55] | |
| ReCirc | Prediction of circRNA expression and function through probe reannotation of non-circRNA microarrays | http://licpathway.net:8080/ReCirc/ | [56] | |
| Ularcirc | Circular RNA detection independent of gene annotation, visualization of forward AND back-splice junctions recover predicted circRNA sequence, recover sequence of back-splice junctions and forward splice junctions, detect miRNA binding sites, detect putative open reading frame of circRNA |
https://github.com/VCCRI/Ularcirc | [57] | |
CircRNA detection software distinguishes circRNAs from their linear counterparts by exploiting the fact that circRNAs contain back-spliced junctions. Detection strategies can be split into three groups – those that use a multi-stage mapping approach, those that directly detect the back-spliced junction reads using split or chimeric reads and those that forgo the alignment step and assemble kmers (short sequences of nucleotides). In the multi-stage mapping approach, there is an initial mapping step where reads that align continuously to the reference genome are filtered out and only unmapped reads are further analysed. These ‘unmapped’ reads are then aligned to pseudo-sequences which are built around putative back-spliced junctions (BSJs) [29]. The newer aligners, including STAR2, HISAT or TopHat2 [30–32], are ‘splice aware’ (reads are split when aligning back to the reference to account for intronic sequences), which allows chimeric reads to be directly aligned to the human reference genome (Figure 2). Care must be taken here as chimeric reads that are in the opposite order can also be generated from other trans splicing instances in the genome such as intergenic (gene fusion) or intragenic (exon/gene duplications) [33]. These false positives can be reduced by selecting only those reads that occur in the same gene (to eradicate intergenic trans splicing and using biological replicates (to minimize intragenic trans splicing). Lastly, newer software packages are opting to forgo the mapping stage and instead use the reference and annotation to assemble and store all kmers that are located near exon boundaries. Each sequencing read is then examined for kmers which are matched to the stored kmers (for example, Circ DBG builds a De Bruijn graph from the kmers [34]). When two kmers from a single read are out of order compared to the reference, this suggests the presence of a circRNA. As this strategy assembles all possible kmers from the reference and annotation files, only circRNAs from annotated exons will be predicted, and no de novo circRNAs will be detected.
Figure 2.
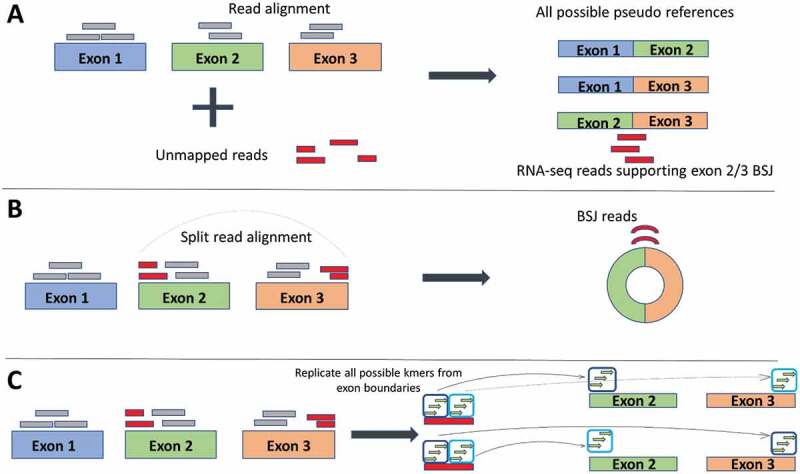
Three different strategies of identifying back-splice junction (BSJ) reads employed by circRNA detection software. (a) Pseudo reference strategy: Reads are first aligned to the reference (grey boxes) any unmapped reads (red boxes) are taken forward and aligned against pseudo references, which are all possible combinations of exon junctions. In this way, any reads that align identify the exons involved in the back-spliced junctions. (b) Split alignment strategy: Splice-aware aligners split the sequencing reads which allows direct alignment to the reference. Reads can be selected to choose the BSJ reads. (c) Kmer strategy: The actual step of read alignment is forgone. Instead, all possible kmers (short stretches of sequence) are created from the exon boundaries. Kmers are then matched to the reads. Matched kmers that are in sequence with the reference are considered linear spliced reads (top exons). Matched kmers that are out of sequence to the reference are considered circRNA, back-spliced junction reads (bottom exons)
Although circRNAs may align with multiple reads in an RNA Sequencing run, only the reads aligning to the back splice can be unambiguously assigned to the circRNA, as the other reads may originate from its linear mRNA counterparts. For this reason, the read counts for circRNAs are relatively low. Accordingly, caution must be taken when inferring the internal sequence of circular RNAs for in silico functional predictions (such as miRNA and RBP binding sites), and this is only possible for single exon circular RNAs. Unfortunately, this does not appear to be the case for several circRNA databases, which include many multi-exonic transcripts that have not been experimentally validated (currently 20 databases: 14 noncurated and six curated, containing 1953–1,223,114 noncurated or 249–3181 curated circRNAs, respectively) [35]. For an excellent review of the current state of circRNA databases see Vromman et al [35].
CircRNA functions
CircRNAs as microRNA sponges
The first papers to demonstrate functional roles for circRNAs found that the circRNAs miR-7 (ciRS-7) and sex-determining region Y (circSRY) bound and sequestered microRNAs miR-7 and miR-138 via 70 and 16 conserved binding sites, respectively [58,59]. Both circRNAs strongly suppressed the ability of the miRNA to bind to its target mRNA, leading to increased expression of the mRNA and suggesting a competing endogenous RNA regulation of the mRNA. This sponge-like mechanism caused great excitement and anticipation of a general functional role for circRNAs. However, bioinformatic analyses suggest that miRNA-binding sites are no more enriched on circRNA than would be expected by chance [60], and sequestering miRNAs appears to be just one of several emerging functional roles for circRNAs (Figure 3).
Figure 3.
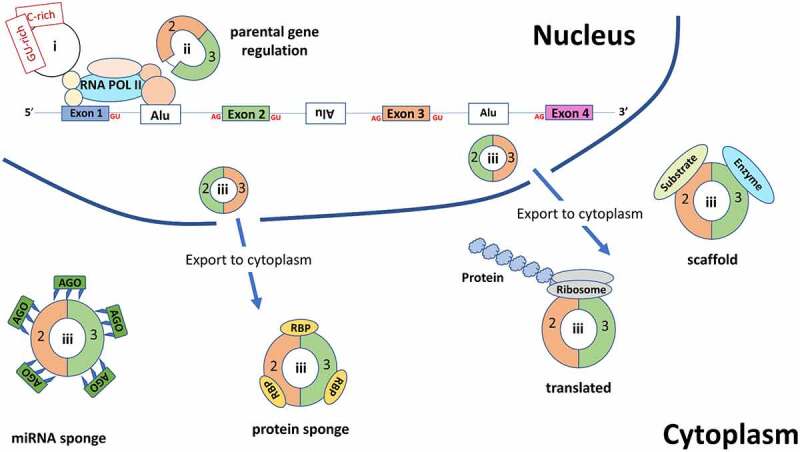
An overview of the various functions for circRNAs. Within the nucleus, intronic circRNAs (ciRNAs (I)) and exon and intron containing circRNAs (EIciRNAs (ii)) are involved in regulation of their parental gene. Exonic circRNAs(iii) are exported to the cytoplasm where they can either act as miRNA sponges (which also bind the Argonaute protein – an essential component of the RNA-induced silencing complex (RISC) which acts upon the targeted mRNA), RBP sponges, protein scaffolds or are translated. RBP: RNA binding protein, Ago: Argonaute protein, Alu: Alu elements
Three in vitro methods are typically used to demonstrate circRNA/miRNA interaction. First, luciferase reporter assays, which involve co-transfection of the circRNA sequence along with the miRNA of interest downstream of the luciferase gene [61], demonstrate miRNA binding to the circRNA sequence in vitro but not the actual circRNA in vivo. Second, biotin labelled probes can be used to co-precipitate or ‘pull down’ the circRNA/miRNA complex in vivo [62]. Third, RNA immunoprecipitation (RIP) or crosslinking-immunoprecipitation (CLIP) of the circRNA by an argonaute 2 protein (AGO2) antibody (part of the miRNA silencing machinery) demonstrates both circRNA/miRNA interaction and the RNA silencing mechanism. Each method should be validated as non-specific binding between circRNAs, and miRNAs can occur. For example, Lim et al [63] showed that biotin pull down of circSlc8a1-bound miRNAs identified 14 miRNAs which, after prioritization, led to five candidate miRNAs. Three of these showed significant and consistent inhibition of luciferase activity, with two having several in silico predicted binding sites. It was not until they performed an inverse pull-down assay to test for endogenous circSlc8a1 binding by using biotinylated miRNA mimics that they demonstrated only one of these miRNAs had a robust endogenous interaction to the circRNA.
CircRNAs as protein sponges, regulators, and scaffolds
CircRNAs can act as (i) protein sponges to block protein activity [64,65], (ii) positive regulators of polymerase II to regulate the transcription of their parental genes in the nucleus (examples include circEIF3J, circPAIP2 and ci-ankrd52 [9,10]), and (iii) protein scaffolds to colocalise enzymes and their substrates in the cytoplasm, thereby facilitating recruitment [66] and enzyme activity [67,68]. For a comprehensive review see Du et al [69].
CircRNAs as protein-coding transcripts
The notion of an alternative circRNA transcriptome is appealing given that circRNAs are predominantly located in the cytoplasm, contain exonic sequences and could potentially have an infinite open reading frame (ORF) because of their circular structure [70,71]. Using ribosome profiling, initial studies examined whether circRNA back spliced reads overlapped ribosome protected fragments (suggesting translation) but found no evidence [4,60]. Demonstrating circRNA translation via ribosomal footprinting is restricted by only being able to count the reads spanning the back spliced junction which is a fraction of total reads in the experiment which may account for the lack of success in early attempts.
An alternative to the canonical mechanism of translation initiation would be needed as circRNA lack the 5ʹ cap which is used for linear mRNA translation – circRNAs would need a cap-independent mechanism. Work on viruses and cellular physiological stress responses identified an alternative, 5ʹ cap-independent translation mechanism involving RNA structures known as internal ribosome entry sites (IRESs), which recruit ribosomes to an internal start codon to initiate translation [72]. It has been shown that circRNAs synthesized in vitro carrying engineered IRESs can be translated in this manner [73] but endogenous IRESs are rare. Recent bioinformatic analysis identified IRES-like AU-rich hexamers that were found to bind trans-acting factors and drive circRNA cap-independent translation in a green fluorescent protein (GFP) cell-based reporter system [74]. The authors predicted that sequences longer than 50 nucleotides would contain an IRES-like hexamer by chance. As >99% of circRNAs are >100 nucleotides in length, most circRNAs could potentially be translated by this mechanism [74].
In addition to IRESs, methylated adenosine residues (N6-methyladenosines (m6A), the most abundant base modification of RNA) in the 5ʹ UTR region, can directly recruit the eukaryotic initiation factor 3 (eIF3) and initiate translation in the absence of the cap-binding factor eIF4E [75]. N6-methyladenosine (m6A) consensus motifs are enriched in circRNAs and a single m6A site was found to be sufficient to drive translation in human cells [76]. For example, Legnini et al used a CRISPR/Cas9 system in mouse embryonic stem cells to produce constructs expressing tagged circ-ZNF609 transcripts which were translated into protein [77].
Importantly, additional to ribosomal footprinting, recent studies based on the use of mass spectrometry have allowed detection of several endogenous circRNA translated proteins. The first study by Pamudurti et al [78] detected protein encoded by circMbl which is generated from the muscleblind locus in Drosophila. Other studies to detect actual circRNA translated proteins via mass spectrometry are by Zhang et al [79] who showed that the circular form of the SNF2 histone linker PHD RING helicase (SHPRH) gene contained an open reading frame driven by IRES to encode a novel 146 amino acid which had tumour suppressor activity in human glioblastoma. Yang et al [80] demonstrated CircFBXW7 encodes a 185 amino acid protein with tumour suppressor activity also in human glioblastomas, while Liang et al [81] detected a 370 amino acids novel circβ-catenin isoform which promoted liver cancer cell growth, both in vitro and in vivo. Zheng et al [82] demonstrated circRNA hsa_circ_0000423 (termed as circPPP1R12A) encoded a 73 amino acid protein which promoted the tumour pathogenesis and metastasis in colon cancer tissues. And lastly, Xia et al [83] showed that Circ-AKT3 encodes a 174 amino acid (aa) novel protein, overexpression of which decreased the cell proliferation, radiation resistance and in vivo tumorigenicity of glioblastoma multiforme (GBM) cells.
In summary, from the literature, it would appear that the predominant functional mechanism of circRNAs is to sequester cytoplasmic miRNAs and participate in regulation of their downstream mRNA targets. However, this focus on miRNAs may reflect the fact that the first circRNAs to be discovered were potent miRNA sponges [58,59] and the relative technical ease of demonstrating this in vitro. By comparison, demonstrating circRNA protein translation is technically demanding. Early studies may have suffered from a lack of read depth to detect back spliced junctions for ribosomal footprinting and an inability to stimulate and detect circRNA protein expression owing to inadequate molecular tools. A recent paper by Mo et al [70] demonstrated an expression vector in mammalian cells that increased circRNA formation up to fivefold compared to previous methods, which revealed previously undetected translation of circRtn4. Furthermore, it is important to validate these circRNA encoded proteins in vivo and the use of highly sensitive and specific methods of protein detection, such as mass spectrometry are helping to overcome this technical challenge.
Circular RNAs in cardiovascular disease
Cardiovascular diseases (CVDs) are a leading cause of mortality and morbidity globally accounting for an estimated 17.9 million lives lost each year [84]. There is growing evidence that circRNAs have potential functional roles in cardiac pathologies including atherosclerosis, myocardial infarction, cardiac fibrosis, cardiac hypertrophy, and heart failure (as demonstrated in Tables 2 and 3). As we unravel the role of circRNAs in the development and progression of CVD, their potential as therapeutic targets to reverse pathophysiological remodelling or enhance protective mechanisms will become apparent [85].
Table 2.
circRNAs as potential biomarkers
| CircRNA | CVD category | Findings of the study | Methods | Reference |
|---|---|---|---|---|
| Hsa_circ_0001445 | Coronary Atherosclerosis | Hsa_circ_0001445 was remarkably stable in plasma. Plasma levels of hsa_circ_0001445 were proportional to coronary atherosclerotic burden and improved detection of coronary artery atherosclerosis. | RT-qPCR in plasma from 200 patients with suspected stable CAD. | Vilades et al 2020 [12] |
| Hsa_circ_0001879 and Hsa_circ_0004104 | CAD | Both significantly upregulated in CAD patients. Overexpression of hsa_circ_0004104 in vitro resulted in dysregulation of atherosclerosis-related genes | Microarray and RT-qPCR in whole blood from 24 CAD patients and 7 healthy controls. Further verified by qPCR in 412 CAD patients and 290 controls | Wang et al 2019 [98] |
| Hsa_circ_0124644 | CAD | First study to investigate the circRNA profile in the peripheral blood of CAD patients. Circulating levels of hsa_circ_0124644 improved detection of CAD. | Microarray in peripheral blood from 12 CAD patients and 12 healthy controls. qPCR of 5 circRNAs with highest fold change in 30 controls and 30 CAD patients. Validation in 137 CAD patients and 115 healthy controls | Zhao et al 2017 [101] |
| Hsa_circ_0005540 | CAD | Hsa_circ_0005540 was significantly associated with CAD. | RNA Sequencing of circRNA expression levels in plasma exosomes. CAD patients (n = 61) and non-CAD controls (n = 38) formed the profiling and internal validation phases. CAD (n = 47) patients and non-CAD controls (n = 51) for the validation phase. | Wu et al 2020 [99] |
| Myocardial Infarction-Associated Circular RNA (MICRA) | Myocardial Infarction | Circulating levels of MICRA were lower in MI patients. MICRA was a strong predictor of left ventricle (LV) dysfunction; patients with lower levels of MICRA were at higher risk of LV dysfunction | RT-qPCR in whole blood from 642 acute MI patients, 86 heathy controls | Vausort et al 2016 [96] |
| Myocardial Infarction-Associated Circular RNA (MICRA) | Myocardial Infarction | MICRA classified patients into ejection fraction (EF) groups. Improved risk classification after MI | RT-qPCR in whole blood from 472 acute MI patients classified into 3 groups according to ejection fraction | Salgado-Somoza et al 2017 [97] |
| CircHIPK3 | Myocardial Infarction | Exosomal circHIPK3 released from hypoxia-induced cardiomyocytes regulates cardiac angiogenesis after MI | Hypoxic cardiomyocytes released circHIPK3-rich exosomes to regulate oxidative stress damage in cardiac endothelial cells. Angiogenesis following MI in mice was promoted. | Wang et al 2020 [100] |
Table 3.
circRNAs associated with atherosclerosis
| CircRNA/miRNA interaction | |||||
|---|---|---|---|---|---|
| CircRNA | miRNA | Protein target | Findings of the study | Methods | Reference |
| CircRNA‑0044073 | miR‑107 | JAK/STAT | CircRNA‑0044073 is upregulated in atherosclerosis and promotes the proliferation and invasion of cells by targeting miR‑107 and activating the JAK/STAT signalling pathway | RT-qPCR in blood cells from patients with atherosclerosis and healthy controls (n = 20 total). RNA pulldown for miRNA interaction. | Shen et al 2019 [110] |
| Circ_CHFR | miR-370 | FOXO1 | Knock down of circ_CHFR inhibits the proliferation and migration of VSMCs in vitro | Microarray on human vascular smooth muscle cells (VSMCs). Luciferase reporter assay for miRNA interaction | Yang et al 2019 [106] |
| Circ_CHFR | miR-214-3p | Wnt3/β-catenin pathway | Circ_CHFR was up-regulated in atherosclerotic serum and ox-LDL-stimulated VSMC. Circ_CHFR regulated cell growth, migration, and inflammation via regulating the expression of Wnt3 as a competitive endogenous RNA (ceRNA) of miR-214 in ox-LDL-treated VSMCs | RT-qPCR in atherosclerotic (n = 32) and control patient’s serum (n = 32). Cell model of atherosclerosis in (VSMCs) with oxidized low-density lipoprotein. Luciferase reporter assay for miRNA interaction | Zhuang et al 2020 [107] |
| Circ-SATB2 | miR-939 | Stromal Interaction Molecule 1 (STIM1) | Circ-SATB2 and STIM1 up-regulated in proliferative VSMCs, miR-939 down-regulated. Circ-SATB2 can regulate VSMC phenotypic differentiation, proliferation, apoptosis and migration by promoting the expression of STIM1 | RT-qPCR in VSMCs. Dual luciferase assay for miRNA interaction | Mao et al 2018 [112] |
| Circ-Sirt1 | miR-132/212 | Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) | Circ-Sirt1 inhibits inflammatory phenotypic switching of VSMCs. Circ-Sirt1 controls NF-κB activation via enhancement of SIRT1 expression by binding to miR-132/212 in vascular smooth muscle cells | RT-qPCR and microarray in human arterial tissues (11 atherosclerotic, 14 control), human plasma (20 CAD, 20 control) and rat VSMCs. RNA pulldown for miRNA interaction | Kong et al 2019 [111] |
| Circ_0003204 | miR-370-3p | TGFβR2/phosph-SMAD3 | Circular RNA circ_0003204 inhibits proliferation, migration and tube formation of endothelial cell in atherosclerosis via miR-370-3p/TGFβR2/phosph-SMAD3 axis | RT-qPCR in human aorta endothelial cells (HAECs). Luciferase activity assay | Zhang et al 2019 [108] |
| Circ_RUSC2 | miR-661 | Tyrosine-protein kinase (SYK) | Circ_RUSC2 can promote the expression of SYK, a target gene of miR-661, and regulates VSMC proliferation, apoptosis, phenotypic modulation, and migration | RT-qPCR in human coronary artery smooth muscle cells. Dual luciferase assay | Sun et al 2019 [113] |
| CircRNA/Protein interaction | |||||
| CircRNA | Protein target | Findings of the study | Methods | Reference | |
| CircANRIL | Pescadillo Homologue 1 (PES1) | Binds pescadillo homologue 1 (PES1) to prevent rRNA maturation. This induces nucleolar stress and p53 activation which leads to apoptosis and proliferation inhibition. Atheroprotection is achieved by culling over-proliferating cell types in atherosclerotic plaques. | RT-qPCR for circRNA expression from peripheral blood mononuclear cell (PBMC), human primary tissues and cells. CAD patients (n = 2,280) undergoing coronary angiography. Also, human endarterectomy specimens (n = 218) collected from patients undergoing vascular surgery. GWAS for cells overexpressing circANRIL. λN-peptide-mediated pull-down of circANRIL-bound proteins | Holdt et al 2016 [65] | |
| Mechanism unknown | |||||
| CircRNA | Findings of the study | Methods | Reference | ||
| CircANRIL | Reduced circANRIL expression could prevent coronary AS by reducing vascular EC apoptosis and inflammatory factor expression | Atherosclerosis rat model (n = 60) 5 groups: control, model, empty vector group, over-expressed circANRIL group and low-expressed circANRIL group. serum lipids, apoptosis, protein and mRNA levels analysed in the 5 groups | Song et al 2017 [102] | ||
| Hsa_circ_0003204 | Hsa_circ_0003204 was notably upregulated in HUVECs treated with oxidized low-density lipoprotein indicating may be involved in the pathogenesis of atherosclerosis. Knockdown boosted the proliferation, migration, and invasion of oxLDL-induced HUVECs, suggesting that hsa_circ_0003204 may slow down the repair of vascular endothelial cell injury. | RT-PCR from (OxLDL) human umbilical vein endothelial cells (HUVECs) | Liu et al 2020 [109] | ||
Circular RNAs as novel biomarkers
Since the discovery of miRNAs in plasma in the mid 1990s, extracellular RNA in biofluids has emerged as an exciting source of biomarkers. Biofluids enable non-invasive, swift sampling, making them an attractive alternative to tissue biopsies.
CircRNAs have been reported in plasma, serum, saliva and urine [13,86–90] and are enriched in these compared to tissues [90,91]. In addition, circRNAs can be packaged into exosomes a subset of extracellular vesicles thought to function in neighbouring and distant cellular communication by transporting proteins, lipids and genetic material between cells [89]. Li et al [90] were the first to show that circRNAs are enriched in exosomes compared to the producer cells. They identified over 1000 circRNAs in human serum exosomes and were able to distinguish colon cancer patients from healthy controls thus demonstrating their value for biomarker application. After exosomes are released into the extracellular fluid they can deliver their internal cargo to initiate a variety of functional responses [92–94], strongly suggesting that there is molecular regulation on the packaging of circRNAs into the exosomes. Bioinformatic analysis showed that circRNAs containing the purine-rich motif 5ʹGMWGVWGRAG-3ʹ were enriched in exosomes compared to other cellular compartments and speculated that RNA binding proteins recognize and package circRNAs containing this motif into exosomes [95]. Due to their closed structure circRNAs are considerably more stable than linear mRNAs – in cells the half-life of circRNAs exceed 48 hours compared to <20 hours for linear RNA [4] – making them excellent candidate biomarkers and potential therapeutic targets for a range of cardiovascular diseases. Presently, only a few studies have demonstrated the use of circRNAs as biomarkers for coronary artery disease (CAD) and myocardial infarction (MI) (summarized in Table 2).
The first study to demonstrate circulating circRNAs as potential biomarkers for CVD was Vausort et al [96] who showed that levels of the circRNA myocardial infarction-associated circular RNA (MICRA) in peripheral blood were a strong predictor of left ventricular dysfunction in 642 MI patients discovery cohort 409 patients, validation cohort 233 patients compared to 86 healthy controls with an odds ratio of 0.53 (95% CI: 0.29 to 0.97). A second study by Salgado-Somoza et al [97] in 472 MI patients confirmed the association between MICRA in peripheral blood and the extent of left ventricular dysfunction and showed that MICRA levels could predict the development of heart failure and help risk stratify patients after MI. MICRA is notable as it is one of the few potential circRNA biomarkers for CVD to have been validated independently. MICRA is an 874-nucleotide long circRNA isoform of the gene ZNF609 (formed mainly from exon 1) [96]. To date, the function of MICRA is not known and further research is needed to determine whether a reduction in MICRA in peripheral blood cells (or other tissues) is directly involved in the development of left ventricular dysfunction and heart failure or forms part of the downstream inflammatory response.
Subsequently, Zhao et al identified circ_0124644 could predict CAD with a receiver operating characteristic area under the curve (ROC AUC) of 0.769 (95% confidence interval, CI, 0.710–0.827, p < 0.001) and a sensitivity and specificity of 0.861 and 0.626, respectively. Two further studies identified three other circRNAs from peripheral blood as potential biomarkers for CAD: Wang et al [98] established that hsa_circ_0001879 and hsa_circ_0004104 were significantly higher in CAD patients compared with controls (hsa_circ_0001879: ROC AUC 0.703 95% CI 0.656–0.750; p < 0.001; hsa_circ_0004104: ROC AUC 0.700 95% CI: 0.646–0.755; p < 0.001), whereas Vilades et al demonstrated hsa_circ_0001445 levels were proportional to coronary atherosclerotic burden in 200 patients with suspected stable CAD [12]. These associations have yet to be validated in independent studies.
Three studies have investigated exosomal circRNAs in cardiovascular disease [99–101]. In the first study to use RNA sequencing for circulating circRNA analysis, Wu et al [99] performed an initial screen of exosomal circRNA expression levels in 3 CAD patients and 3 controls and later validated their findings in two independent cohorts (58 CAD patients versus 35 controls and 47 CAD patients versus 51 controls). They found circ_0005540 was consistently associated with CAD (ROC AUC 0.853; 95% confidence interval = 0:799 − 0:906, P < 0:001 after adjustment for risk factors). In contrast, using cultured cells and a mouse MI model, Wang et al [100] demonstrated that hypoxic cardiomyocytes released circHIPK3 rich exosomes which reduced the infarct area by promoting angiogenesis at the infarct border. CircHIPK3 promoted cardiac endothelial cell proliferation, migration, and tube formation in vitro via miR-29a-mediated regulation of Vascular Endothelial Growth Factor A (VEGFA).
Although these studies highlight the potential of circRNAs as biomarkers, we should proceed with caution. Almost all studies suffer from small cohort sizes, a lack of consideration for confounding medications commonly used for the treatment of CAD and, except for MICRA, a lack of independent replication. Circulating circRNAs levels may be rapidly changing and so the timing of collection during the course of the disease needs to be considered. As yet, there is little consensus between studies as to which circRNAs are the lead candidates, and more work is needed to identify these. Moreover, except for the study by Wu et al [99], these studies have been carried out using targeted technologies of microarray or RT-qPCR, highlighting the potential for more discoveries to come with the use of more sensitive RNA sequencing coupled with specialized library preparation protocols for circRNA and exosome enrichment.
CircRNAs in atherosclerosis (AS)
Atherosclerosis is a silent, chronic inflammatory condition characterized by a narrowing of the arteries from the build-up of fats, cholesterol, and other substances, which can reduce blood flow to the heart muscle. Over the decades, our understanding of atherosclerosis has evolved from a model of passive cholesterol deposition to a dynamic, complex interplay of different cell types and cytokines (cell signalling molecules) resulting in a chronic inflammatory disease.
Table 3 summarizes current knowledge of the involvement of circRNAs in atherosclerosis. Of the 10 papers published to date, 8 studies have identified the mechanism underlying the association between the circRNA and atherosclerosis, with seven identifying in vitro circRNA/miRNA associations and one showing a circRNA/protein interaction. Notably, for three circRNAs (circANRIL, circCHFR and circ_0003204), associations with atherosclerosis have been reported by two independent studies. However, except for circ_0003204, the independent studies propose different functions or mechanisms of action for the circRNA.
CircANRIL [65,102] originates from chromosome 9p21, a region in the genome that has been identified by genome‐wide association studies (GWAS) as being strongly associated with cardiovascular disease, including coronary artery disease [103]. This locus is devoid of protein-coding genes but houses a long noncoding RNA called antisense noncoding RNA in the INK4 locus (ANRIL). Increased linear ANRIL is associated with increased atherosclerosis [104] and multiple linear isoforms as well as circular isoforms have been identified [65,105]. CircANRIL is expressed in human vascular tissue, smooth muscle cells and monocyte/macrophages, all of which play an important role in atherogenesis. A notable study by Holdt et al (add ref) using a large CAD cohort suggests circANRIL confers atheroprotection. However, a later study looking at the effect of circANRIL on the inflammatory response in a rat model of atherosclerosis by Song et al [102] suggests that expression of circANRIL was correlated with the expression of inflammatory factors in vascular endothelial cells (ECs). Over-expression of circANRIL exacerbated inflammation and promoted atherosclerosis. Further studies are needed to confirm whether circANRIL has a protective or antagonistic role in atherosclerosis [102].
The two studies involving Circ_CHFR suggested two different miRNA/protein interactions as its target. Yang et al [106] suggest circ_CHFR inhibits the proliferation and migration of vascular smooth muscle cells via acting as a sponge to miR-370 which targets the 3ʹUTR of FOXO1. Conversely, Zhuang et al [107] advocate that circ_CHFR promotes cell growth, migration and inflammation in oxidized low-density lipoprotein treated human vascular smooth muscle cells. In contrast, both studies by Zhang et al [108] and Liu et al [109] identifying circ_0003204 as a marker for atherosclerosis propose that it inhibits proliferation and angiogenesis in vascular endothelial cells.
Of the seven circRNA/miRNA studies, only two used the more compelling RNA pulldown method with the remaining studies using the less compelling luciferase method [110,111]). Shen et al [110] identified circ_0044073 as a potential modulator of JAK/STAT levels by binding miR-107 whereas Kong et al [111] identified that circSirt1 may alter nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) by binding miR-132/212 . Among the remaining five studies, Mao et al [112] proposed circSATB2 acts on Stromal Interaction Molecule 1 protein via miR-939, Sun et al [113] proposed circRUSC2 acts on Tyrosine-protein kinase via miR-661 and Zhang et al [108] who propose circ_0003204 inhibits proliferation, migration and tube formation of endothelial cells in atherosclerosis via a miR-370-3p/TGFβR2/phosph-SMAD3 axis. The remaining two studies to use the luciferase method suggest two different miRNAs involved with circCHFR [106,107]: Yang et al [108] who suggests circCHFR facilitates the proliferation and migration of vascular smooth muscle cells via miR-370/FOXO1/cyclin D1 pathway and Zhuang et al suggest circCHFR drives cell growth, migration and inflammation via the miR-214-3p/Wnt3/β-catenin pathway. Both studies used a cell model of atherosclerosis where human vascular smooth muscle cells were stimulated with oxidized low-density lipoprotein.
Together, these studies provide evidence to suggest circRNAs play a regulatory role in the development and progression of atherosclerosis. To date four studies [12,65,107,110] have reported altered levels of circRNAs in biofluids in patients with atherosclerosis, highlighting a potential role for these circRNAs (circRNA_0001445, circRNA-0044073, circCHFR, circANRIL) as potential biomarkers.
Myocardial infarction/ischaemia reperfusion injury
The coronary arteries are the major blood vessels supplying the heart muscle. Following the accumulation of atherosclerotic plaque in these arteries, the blood supply to the heart can become restricted and no longer meet the demands of the myocardium. Over time, this can lead to myocardial ischaemia, associated with a cascade of cellular, inflammatory and biochemical events [114], subclinical myocardial dysfunction and ultimately MI [115]. Cardiomyocytes have limited regenerative capacity and restoration of blood supply or reperfusion is the current clinical procedure to limit death of cardiomyocytes, but reperfusion itself can exacerbate injury to the site [116]. There is growing evidence that circRNAs are involved in cellular responses to MI and ischaemia reperfusion (IR), either through miRNA regulation or interaction with proteins in the cell cytoplasm.
In animal models or human cells in vitro, 18 circRNAs have been associated with MI/IR to date. Of these, 13 circRNAs have been suggested to regulate MI/IR-induced apoptosis in cardiomyocytes via interaction with miRNAs (Table 4) and represent potential therapeutic targets in this setting. Geng et al [61] were the first to suggest this mechanism, suggesting CDR1as acted as a sponge of miR-7 and led to an increase in apoptosis in cultured mouse cardiomyocytes. However, this study demonstrated no direct evidence for this interaction. Instead, overexpression of CDR1as through administration of an expression plasmid via intracardiac injection in a mouse model of MI led to an increase in infarct size which could be reversed by overexpression of miR-7a. A further three studies by Jin et al, Chen et al and Ji et al [117–119] provide no direct evidence for circRNA/miRNA interaction but instead investigated how overexpression of one affected expression of the other. These studies suggested that circ_0010729 modulates mir-145-5p/mTOR and MEK/ERK pathways [117], circDLPAG4 modulates the miR-143/HECTD1 axis [118] and circPAN3 modulates the miR-31-5p/Quaking axis [119]. Two recent studies have further demonstrated a role of circ_0010729 in hypoxic cardiomyocytes using dual-luciferase reporter assays, RNA immunoprecipitation assays and RNA pull-down assay [120,121]. Interestingly, they both suggest the target proteins are tumour necrosis factor receptor-associated factor (TRAF) but suggest these associate with different miRNAs. Lei et al suggest circ_0010729 acts on miR-27a-3p to regulate TRAF5 and Zhang et al suggest circ_0010729 acts on miR-370-3pto regulate TRAF6. Conversely, the study by Wang et al [121] was the first to use the more compelling method of AGO2 pulldown to not only demonstrate direct evidence of circMFACR (mitochondrial fission and apoptosis-related circRNA) interaction with miR-652-3p but also the AGO2 protein in vivo. A further three studies also provide evidence of circRNA/miRNA interaction with the AGO2 pulldown: Li et al [123] for the circ_ncx1/miR-133a-3p/CDIP1 axis; Gan et al [124] for the circ_101237/let-7a-5p/IGF2BP3 axis and Sun et al [125] for the circ_LAS1L/miR-125b/SFRP5 axis, which all demonstrated circRNA regulation of cardiomyocyte apoptosis. Beyond these, four recent studies have demonstrated cardioprotective roles of the circRNA via a circ/miRNA/protein axis. These are, Cai et al [126] who found circ-Ttc3 was markedly upregulated in the ischaemic myocardium and proposed circ-Ttc3 may bind miR-15b to increase ADP ribosylation factor like 2 (Arl2) expression; Zhao et al [127] who suggested circMACF1 may modulate miR-500b-5p which in turn inhibits epithelial membrane protein 1 (EMP1); Zhang et al [128] who demonstrated circ_0007623 promoted cardiac repair after acute myocardial ischaemia and suggest a miR-297/VEGFA axis and lastly Shao et al [129] who suggest circDENND2a binds to miR-34a which may modulate the β-catenin and Ras/Raf/MEK/ERK pathway.
Table 4.
circRNAs associated with myocardial infarction or ischaemia reperfusion injury
| CircRNA/miRNA interaction | |||||
|---|---|---|---|---|---|
| CircRNA | miRNA target | Protein target | Findings of the study | Methods | Reference |
| CDR1as | miR-7a | Poly (ADP-ribose) polymerase (PRP) and SP/KLF family of transcription factors (SP1) | Overexpression of CDR1 in vivo promoted apoptosis and increased cardiac infarct size; overexpression of miR-7a had the opposite effect | RT-qPCR from mouse cardiomyocytes. No direct evidence of circRNA/miRNA interaction. CDR1as induced apoptosis could be reverse by miR-7 overexpression | Geng et al 2016 [61] |
| Mitchondrial fission and apoptosis-related circRNA (MFACR) | miR-652-3p | MTP18 | Regulated mitochondrial fission and apoptosis in the heart | Mouse cardiomyocytes. AGO2 immunoprecipitation | Wang et al 2017 [122] |
| Ncx1 | miR-133a-3p | Pro-apoptotic gene cell death-inducing protein (CDIP1) | Increased in response to reactive oxygen species (ROS) and promotes cardiomyocyte apoptosis | H9c2 cells and neonatal rat cardiomyocytes. Biotinylated pulldown of miRNA-133 and circMFACR. AGO2 immunoprecipitation | Li et al 2018 [123] |
| CircHIPK3 | circHIPK3 by binding to miRNA-124-3p | Overexpression upregulated pro-apoptotic Bax, and downregulated anti-apoptotic Bcl-2 | Inhibited proliferative ability and induced apoptosis of cardiomyocytes after myocardial IR injury | Mouse cardiomyocytes. Dual-luciferase reporter gene assay | Bai et al 2019 [134] |
| Ttc3 | miR-15b | ADP ribosylation factor like 2 (Arl2) | Played a cardioprotective role after myocardial infarction. Overexpression counteracted hypoxia-induced ATP depletion and apoptotic death. | Rat cardiomyocytes and cardiac fibroblasts. Dual-luciferase reporter assay | Cai et al 2019 [126] |
| Circ_0010729 | miR-145-5p | mTOR and MEK/ERK pathways | Silencing circular RNA circ_0010729 protected human cardiomyocytes from oxygen-glucose deprivation-induced injury | 4 h oxygen-glucose-deprivation (OGD) human cardiomyocytes. No direct evidence for circRNA/miRNA interaction. Expression studies after overexpression of one against the other. | Jin et al 2019 [117] |
| Circ_0010729 | miR-27a-3p | Tumour necrosis factor receptor-associated factor 5 (TRAF5) | Circ_0010729 knockdown alleviated cell viability inhibition and cell apoptosis under hypoxic conditions. MiR-27a-3p was targeted by circ_0010729. MiR-27a-3p restoration enhanced cell viability, depleted cell apoptosis and promoted glycolysis of hypoxia-induced cells, while these effects were abolished by TRAF5 overexpression | miR-27a-3p/TRAF5 relationship was verified by dual-luciferase reporter assay, RNA immunoprecipitation (RIP) assay and RNA pull-down assay | Lei et al 2020 [120] |
| Circ_0010729 | miR-370-3p | Tumour necrosis factor receptor-associated factor 5 (TRAF5) | Circ_0010729 knockdown protected cardiomyocytes against hypoxic dysfunction. MiR-370-3p inhibition attenuated the protective effects of circ_0010729 knockdown on hypoxia-modulated cardiomyocyte dysfunction. | miR-370-3p/TRAF6 relationship was verified by dual-luciferase reporter assay, RNA immunoprecipitation (RIP) assay and RNA pull-down assay | Zhang et al 2020 [121] |
| CircMACF1 | miR-500b-5p | Epithelial membrane protein 1 (EMP1) | Attenuated Acute Myocardial Infarction | Mouse cardiomyocytes. Dual-luciferase reporter gene assay | Zhao et al 2020 [127] |
| Circular RNA_101237 | let‑7a‑5p | Insulin-like-growth factor 2 mRNA-binding protein 3 (IGF2BP3) | Mediated anoxia/reoxygenation injury | Mouse cardiomyocytes biotinylated miRNA pull-down. AGO2 immunoprecipitation |
Gan et al 2020 [124] |
| Circ_LAS1L | miR-125b | Secreted frizzled-related protein 5 (SFRP5) | Down-regulated in acute myocardial infarction (AMI). Regulates cardiac fibroblast activation, growth, and migration and apoptosis | Mouse cardiomyocytes. Luciferase reporter gene assay. Biotinylated RNA pull down. AGO2 immunoprecipitation |
Sun et al 2020 [125] |
| CircDLPAG4/HECTD1 | miR-143 | HECT Domain E3 Ubiquitin Protein Ligase 1 (HECTD1) | CircDLPAG4/HECTD1 played a vital role in apoptosis and cell migration in endothelial cells through ER stress in response to ischaemia/reperfusion injury | Primary Human Umbilical Vein Endothelial Cells (HUVEC)/mice. No direct evidence for circRNA/miRNA interaction. Expression studies after overexpression of one against the other. |
Chen et al 2020 [118] |
| Circ_ Pan3 | miR-31-5p | Quaking (QKI) | Silencing of miR-31-5p significantly alleviated the myocardial apoptosis induced by Doxorubicin (DOX) treatment. MiR-31-5p acts as a negative regulator of circPan3 by directly suppressing QKI both in vivo and in vitro | Mouse cardiomyocytes. No direct evidence for circRNA/miRNA interaction. Dual-luciferase reporter gene assay of QKI/miR-31-5p interaction. RNA immunoprecipitation of QKI/circPan3 interaction. Expression levels of circPan3 when miR-31-5p was inhibited | Ji et al 2020 [119] |
| Circ_0007623 | miR-297 | Vascular Endothelial Growth Factor A (VEGFA) | Hsa_circ_0007623 promoted cardiac repair after acute myocardial ischaemia, and protect cardiac function | Hypoxia-induced human umbilical vein endothelial cells (HUVECs). Dual luciferase reporter gene assay of circ_0007623/miR-297 | Zhang et al 2020 [128] |
| CircDENND2A | miR-34a | May work via β-catenin and Ras/Raf/MEK/ERK pathways | Overexpression of circDENND2A enhanced cell viability and migration but declined apoptosis under oxygen glucose deprivation (OGD) | Rat H9c2 cells. Luciferase activity assay For interaction of circDENND2A/miR-34a |
Shao et al 2020 [129] |
| CircRNA/Protein interaction | |||||
| CircRNA | Protein target | Findings of the study | Methods | Reference | |
| Autophagy-related circRNA (ACR) | DNA (cytosine-5-)-methyltransferase 3 beta (Dnmt3B) | Inhibition of autophagy – a type of cell death protects cardiomyocytes during ischaemia/reperfusion. CircRNA ACR was able to inhibit autophagy and cell death in cardiomyocytes to protect the heart from reducing I/R induced infarct sizes. ARC activated PTEN-induced kinase 1 (PINK1) expression (an autophagy associated gene) by directly binding to and blocking Dnmt3B-mediated methylation of the Pink1 promoter. | Microarray on mouse cardiomyocytes to look at differential expression in autophagy induced mouse hearts by I/R injury. Transcriptome microarray after ACR knockdown. RNA-binding protein immunoprecipitation (RIP) of ACR/Dnmt3B association. Chromatin immunoprecipitation (ChIP) assay showing Dnmt3B/Pink1 association | Zhou et al 2019 [131] | |
| CircNfix | Y-Box Binding Protein 1 (Ybx1) | Ybx1 is a transcription factor that is involved with cardiomyocyte differentiation. Interaction between circNfix and Ybx1 prevented nuclear translocation of Ybx1, causing cytoplasmic retention and degradation. Nfix also bound to miR-214, which has been demonstrated to be essential for cardiomyocyte proliferation. Loss of circNfix induced cardiac regeneration and angiogenesis and inhibited cardiomyocyte apoptosis after MI, which significantly restored cardiac function and improved the prognosis | Used RNA Sequencing data from another study [138] to filter for circRNAs differentially expressed in human, mouse and rat MI vs control/sham. RT-qPCR of circNfix. RNA pulldown assays and mass spectrometry analysis for Nfix/Ybx1 association. Luciferase, RNA pulldown for Nfix/mIR-214 association | Huang et al 2019 [132] | |
| CircFndc3b | Fused in Sarcoma (FUS) | CircFndc3b significantly downregulated in post MI mouse hearts and in human cardiac tissues of ischaemic cardiomyopathy patients. Overexpression in cardiac endothelial cells increases vascular endothelial growth factor-A (VEGF-A) expression which enhances angiogenic activity and reduces cardiomyocytes and endothelial cell apoptosis. CircFndc3b interacts with the RNA binding protein Fused in Sarcoma to regulate VEGF expression and signalling | Microarrays on sham or MI mouse hearts (n = 2) and ischaemic cardiomyopathy human heart tissue (n = 7), normal heart tissue (n = 4). RNA Immunoprecipitation (RIP) | Garikipati et al 2019 [133] | |
| CircFoxo3 | p21-CDK2 | Circ-Foxo3 highly expressed in the tissues of aged mice and patients. Ectopic expression of circFoxo3 induced cellular senescence of mouse embryonic fibroblasts (MEFs), where it interacted with the anti-senescence proteins ID1 and E2F1, and anti-stress proteins FAK and HIF1α. These interactions prevented nuclear translocation of these transcription factors, causing cytoplasmic retention and reduced function | Primary cardiomyocytes isolated from neonatal and 12-week heart mouse tissues (n = 20) RT-PCR for expression levels. Antibody pulldown for protein/circRNA association | Du et al 2017 [130] | |
| Mechanism unknown | |||||
| CircRNA | Protein target | Findings of the study | Methods | Reference | |
| CircRNA 010567 | May be related to the inhibition on the TGF-β1 signalling pathway |
Cardiac function and myocardial infarction (MI)-induced myocardial fibrosis (MF) improved, myocardial apoptosis was ameliorated in circRNA 010567 siRNA group compared with that in Model group. Regulatory mechanism may be related to the inhibition on the TGF-β1 signalling pathway |
Rat MI model by ligation of the left anterior descending coronary artery. Model rats were randomly divided into circRNA 010567 siRNA group and Model group, with sham operation group as Control group (n = 30). The effects of circRNA 010567 on myocardial infarction (MI)-induced myocardial fibrosis (MF), myocardial apoptosis, mRNA, and protein expression levels of TGF-β1 and Smad3 in heart tissues of MI rats were detected using the small animal ultrasound system, Masson staining, terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling (TUNEL) staining, reverse transcription-polymerase chain reaction (RT-qPCR), and Western blotting |
Bai et al 2020 [135] | |
In addition to circRNA/miRNA interaction, four studies have demonstrated circRNA/protein binding mechanisms involved in MI/IR injury. The first was Du et al [130] who used antibody pulldown for circFoxo3/p21-CDK2 association. This study was followed by three studies in the same year. Zhou et al [131] demonstrating Autophagy-related circRNA (ACR) was able to inhibit autophagy and cell death in cardiomyocytes to protect the heart from reducing I/R induced infarct sizes. The authors demonstrated circ_ACR blocks DNA methyltransferase 3 beta (Dnmt3B) protein methylation of PTEN-induced kinase 1 (Pink1) to activate Pink1 expression – an autophagy associated gene. Huang et al [132] showed interaction between circNfix and the transcription factor Y-Box Binding Protein 1 (Ybx1) which is involved in cardiomyocyte differentiation. Inhibition of circNfix induced cardiac regeneration and angiogenesis and inhibited cardiomyocyte apoptosis after MI, which significantly restored cardiac function and improved the prognosis. Finally, Garikipati et al [133] found that circFndc3b was significantly downregulated in post-MI mouse hearts and in human cardiac tissues of ischaemic cardiomyopathy patients. They used RNA Immunoprecipitation to show circFndc3b interaction with Fused in Sarcoma (FUS) protein to regulate vascular endothelial growth factor-A expression and signalling. Interestingly and perhaps surprisingly, it was not until the study by Huang et al [132] in 2019 that RNA Sequencing was used to identify CircNfix.
As well as circHIPK3 rich exosomes being shown to regulate oxidative stress damage by Wang et al (Table 2) circHIPK3 was first shown to aggravate myocardial ischaemia-reperfusion injury by binding to miRNA-124-3p. Bai et al [134] determined that circHIPK3 induced apoptosis in cardiomyocytes after MI by upregulating the oxidative stress regulators Bax and Bcl-2. Another study by the same author [135] showed inhibition of circ_010567 with siRNA improved cardiac function, alleviated myocardial fibrosis, and had a significantly lower apoptosis rate of myocardial cells. They suggest the regulatory mechanism may be related to the inhibition on the TGF-β1 signalling pathway.
It is encouraging to see the numerous studies in recent years demonstrating various circRNAs associated with MI and I/R injury offering several potential therapeutic targets with circHIPK3 being independently validated to act in oxidative stress. However, further independent replication in large, well-controlled studies and cohorts to prioritize circRNA candidates for future study will be needed.
CircRNAs in heart failure/hypertrophy/cardiac fibrosis
Once the damage has occurred through MI or chronic hypertension, the heart undergoes macroscopic and microscopic remodelling [136,137]. An early infiltrative inflammatory response is followed by replacement of infarcted myocardium with non-elastic fibrotic tissue. Diffuse interstitial fibrosis may also affect areas of the heart remote from the initial injury. The architecture of the left ventricle exhibits mechanically disadvantageous changes, including overall ventricular dilatation and alteration from an efficient elliptical shape to a spherical chamber morphology. In vulnerable
patients, these processes can lead to subclinical myocardial dysfunction and subsequent symptomatic heart failure (HF) [136,137,139]. Cardiac fibrosis can be caused by abnormal deposition of extracellular matrix in the myocardium. It can be the result of scarring after MI but can be more widespread and common in heart failure. The scarring is detrimental by either stiffening the myocardium thereby reducing the pumping ability of the heart by impairing electrical conductance [140].
Table 5 represents nine studies on circRNAs that have been demonstrated to act in hypertrophy and cardiac fibrosis contributing to heart failure. All nine studies suggest a circRNA-miRNA-protein axis. Only one study conducted by Han et al [141] was carried out in humans, with the remaining studies carried out in mice. Using RNA pulldown experiments, Han et al showed circ_0097435 could bind multiple miRNAs and promote apoptosis although the protein targets remain unknown. They then extracted exosomes from patients with heart failure and controls and showed that hsa_circ_0097435 expression was significantly higher in patients suggesting a biomarker role. Interestingly, two studies suggest yet a further role of the circHIPK3. CircHIPK3 was associated with two different miRNA/protein axes – miR-17-3p/Adenylyl cyclase type 6 (ADCY6) by Deng et al [142] with downregulation of circHIPK3 reducing cardiac fibrosis and maintaining cardiac function post MI, and miR-29b-3p/α-smooth muscle actin (α-SMA), Collagen type I alpha I (COL1A1, COL3A1) by Ni et al [143], who saw reduced cardiac fibroblast proliferation with circHIPK3 silencing. However, they did not show direct evidence for a circHIPK3/miRNA interaction and only demonstrated cytoplasmic co-localization by RNA FISH. Neither miRNA replicated the previous circHIPK3 study by Bai et al [134].
Table 5.
circRNAs associated with heart failure/hypertrophy and cardiac fibrosis
| CircRNA | miRNA target | Protein target | Action | Cell/Tissue type | Reference |
|---|---|---|---|---|---|
| Heart-related circRNA (HRCR) | miR-223 | Apoptosis repressor with CARD domain (ARC) | MiR-223 acted as a positive regulator of cardiac hypertrophy. Increased expression of HRCR sequesters free miR‐223 | RT-PCR of heart related circRNAs responsive to saline- vs. isoproterenol-infused mice. (n = 7) and transverse aortic constriction or to sham treatment (n = 8) Biotinylated RNA pulldown. AGO2 immunoprecipitation | Wang et al 2016 [148] |
| CircRNA_000203 | miR-26b-5p | Collagen alpha 2(I) (Col1a2) and connective tissue growth factor (CTGF) | Over-expression of circRNA_000203 could eliminate the anti-fibrosis effect of miR-26b-5p in cardiac fibroblasts | CircRNA expression analysis with microarray from mouse myocardium (n = 8 model, n = 8 control Biotinylated RNA pull-down of circRNA/miRNA. Luciferase assay for miRNA/mRNA targets | Tang et al 2017 [144] |
| CircRNA_010567 | miR-141 | TGF-β1 | CircRNA_010567 promoted myocardial fibrosis via suppressing miR-141 by targeting TGF-β1 | Mouse cardiac fibroblasts. CircRNA expression with microarray confirmed with RT-PCR. Luciferase assay for circRNA/miRNA association. | Zhou et al 2017 [150] |
| Circ-HIPK3 | miR-17-3p | Adenylyl cyclase type 6 (ADCY6) | Demonstrated an increase of Adenylate cyclase type 6 (ADCY6) caused by circ-HIPK3 which was ameliorated by miR-17-3p overexpression and vice versa, implicates a circ-HIPK3 – miR-17-3p – ADCY6 axis. Downregulation of circ-HIPK3 can alleviate fibrosis and maintain cardiac function post MI in mice | 24 mice divided into 4 groups – normal group (without surgery), control group (without ligation), NC (negative control) group and experiment group. Dual luciferase expression vectors with circ-HIPK3 co-transfected with miR-17-3p | Deng et al 2019 [142] |
| CircSlc8a1 | miR-133a | Adenovirus (AAV9)-mediated RNAi knockdown of circSlc8a1 attenuates cardiac hypertrophy from pressure-overload, whereas forced cardiomyocyte specific overexpression of circSlc8a1 resulted in heart failure | Mouse cardiomyocytes. Biotin-based pull-down of circRNA/miRNA association then qPCR of miRNA and luciferase assay | Lim et al 2019 [62] | |
| CircNFIB (nuclear factor 1 B-type) | miR-433 | AZIN1 and JNK1 | CircNFIB overexpression reduced cardiac fibroblast proliferation (a process involved with fibrosis) based on TGF-β stimulation), while inhibition of circNFIB promoted fibroblast proliferation. Action via miR-433. | RT-PCR of candidate circRNA expression levels in MI fibrosis mice (n = 5 control, n = 6 MI) and cardiac fibroblasts. miRNA association with dual luciferase reporter assay system. | Zhu et al 2019 [151] |
| CircHIPK3 | miR-29b-3p | α-smooth muscle actin(α-SMA), Collagen type I alpha I (COL1A1, COL3A1 | CircHIPK3 silencing reduced cardiac fibroblast proliferation, migration and the upregulation of a-SMA expression levels induced by Ang II in vitro | Mouse cardiac fibroblasts (CFs). No direct evidence for circRNA/miRNA interaction RT-PCR for circRNA expression level. RNA FISH to determine whether circHIPK3 and miR-29b-3p colocalise in CFs. | Ni et al 2019 [143] |
| CircRNA_000203 | miR26b-5p and miR-140-3p | Gata4 | CircRNA_000203 was found to be upregulated in the myocardium of cardiac hypertrophy induced mice. Demonstrated that circRNA_000203 sponged miR-26b-5p, −140-3p, abolished the suppression of Gata4 by miR-26b-5p, −140-3p, resulting in the increase of GATA4 in NMVCs | Neonatal mouse ventricular cardiomyocytes (NMVCs). Dual-luciferase assays of circRNA_000203/miR-26b-5p, and miR-140-3p Biotinylated RNA pull-down assay between circRNA_000203/miR-26b-5p,and miR-140-3pincubated | Li et al 2020 [145] |
| Hsa_circ_0097435 | hsa_miR_6799_5P, hsa_miR_5000_5P, hsa_miR_609 and hsa_miR_1294 | Unknown | Overexpression of circ_0097435 promoted cardiomyocyte apoptosis, silencing hsa_circ_0097435 inhibited apoptosis | RNA-Seq heart failure patients (n = 5) vs control (n = 4) Dysregulated expression confirmed with qPCR 40 patients with heart failure. RNA-pulldown experiments and AGO2-immunoprecipitation experiments revealed that hsa_circ_0097435 sponged multiple microRNAs | Han et al 2020 [141] |
Two studies suggest the same circRNA, circ_000203, exacerbates cardiac fibrosis and hypertrophy. Tang et al [144] showed circRNA_000203 inhibiting the anti-fibrosis effect of miR-26b-5p in cardiac fibroblasts via collagen alpha 2(I) (Col1a2) and connective tissue growth factor (CTGF); Li et al [142] showed circRNA_000203 could sponge miR-26b-5p along with miR-140-3p in mouse ventricular cardiomyocytes which enhances Gata4 levels to exacerbate cardiac hypertrophy. This is the first time two studies implicate the same circRNA acting upon the same miRNA, albeit with differing downstream targets.
Lim et al [63] built on earlier studies that demonstrated a role of miR-133a and cardiac hypertrophy and heart failure – inhibition of miR-133a induces whereas overexpression inhibits cardiac hypertrophy [146,147]. Lim et al demonstrated an interaction between circSlc8a1 and miR-133a and undertook both knockdown and overexpression of circSlc8a1 in vivo and showed that reducing endogenous circSlc8a1 (thereby increasing miR-133a levels) inhibited hypertrophy and forced cardiomyocyte overexpression resulted in heart failure thereby proposing inhibition of circSlc8a1 as a novel therapeutic target.
Lastly, a study by Want et al [148] suggested a cardioprotective of a circRNA role against fibrosis and hypertrophy. They showed that the circRNA they called heart-related circRNA (HRCR) acted as a sponge to the positive regulator of cardiac hypertrophy miR-223. By sequestering miR-223 they found less degradation of the protein apoptosis repressor with CARD domain (ARC).
Again, encouraging to see the numerous studies implicating various circRNAs in cardiac hypertrophy which are potentially therapeutic targets. It is also encouraging to start to see independent validation of the same circRNA such as circ_000203 implicated in the same disease phenotype, albeit with different mechanisms. The same circRNA may act upon more than one downstream target which is being suggested by the different studies. Further evidence for a significant cardiovascular circRNA – circHIPK3 is also provided. Again, a need for further studies with large sample sizes to support these findings is needed.
Summary
Growing evidence supports an important role of circRNAs in normal physiology and a range of disease states, including CVDs. The majority of studies have focussed on the circRNA/miRNA/mRNA axis which may be due to the first circRNAs being demonstrated as potent miRNA sponges and the relative ease of the in vitro methods that demonstrated this function. Technically challenging in vitro techniques demonstrating circRNA translation may have hindered initial studies, with ribosomal footprinting only detecting the fraction of reads that cross the back spliced junction. With circRNA library enrichment and the validation of proteins with mass spectrometry other circRNA functions are now emerging and whilst translated circRNAs have not been associated with CVD, evidence for circRNA translation in cancer biology [81,82] suggests it is only a matter of time before protein-coding cardiovascular circRNAs are identified as the technical challenges are overcome.
Several circRNA/miRNA/mRNA studies suffer from low cohort numbers and some show no direct evidence of the proposed circRNA/miRNA interaction. Many other studies use luciferase assays, which demonstrate an interaction with circRNA/miRNA mimics in vitro but these should be validated with more robust experimental methods such as circRNA/AGO2 immunoprecipitation.
As yet, there is a shortage of experimental replication confirming involvement of the same circRNA and consequently, there is little overlap of circRNAs within or between cardiovascular aetiologies. However, recent studies are beginning to validate the same circRNA involved in the same or different aetiologies. CircHIPK3 is commonly expressed in diverse tissues, such as the lung, heart, stomach, colon, brain and has been demonstrated to be a key circRNA in a variety of human cancers [149]. Several studies highlighted here suggest a role in both myocardial ischaemia and cardiac hypertrophy for circHIPK3. It is encouraging that other circRNAs are being independently replicated such as MICRA, circ_000203 and circ_0010729 as we begin to clarify the lead candidate biomarkers and therapeutic targets for cardiovascular disease among this relatively newly identified class of non-coding RNA. It is surprising that so few studies have employed RNA sequencing for circRNA detection with most cardiovascular studies using microarray or RT-qPCR technologies, limiting detection of novel circRNAs. With the development of methods for circRNA enrichment in biofluids and more user-friendly software and bioinformatics pipelines future uptake of RNA sequencing technologies detection may be encouraged. Additionally, the only in silico tools for circRNA analysis in the studies appear to be for predicting circRNA/miRNA binding sites and again this may be due to the focus on this mechanism of function. Table 1 highlights the bioinformatic tools that are available such as predicting potential interactions of circRNAs with RNA binding proteins as well as identification of circRNAs with protein-coding potential. The utilization of these bioinformatic tools may help to broaden the lens and validate other functions for circRNAs.
In recent years the physiological and pathological roles of cardiovascular circRNAs have begun to be identified. Additionally, several circRNAs have been found to be promising diagnostic and prognostic biomarkers as well as novel therapeutic targets. Precise mechanisms are still being elucidated but adding to traditional in vitro methods emerging biological and informatic techniques are advancing our understanding will be increased and clinical application of circRNAs will enhance our treatment for cardiovascular disease.
Funding Statement
This work was supported by the Heart Foundation of New Zealand, the Health Research Council of New Zealand. ZW holds a Heart Foundation of New Zealand PhD Scholarship.
Disclosure statement
The authors declare no conflict of interest.
References
- [1].Nigro JM, Cho KR, Fearon ER, et al. Scrambled exons. Cell. 1991;64(3):607–613. [DOI] [PubMed] [Google Scholar]
- [2].Cocquerelle C, Daubersies P, Majérus MA, et al. Splicing with inverted order of exons occurs proximal to large introns. Embo J. 1992;11(3):1095–1098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Zhang XO, Wang H-B, Zhang Y, et al. Complementary sequence-mediated exon circularization. Cell. 2014;159(1):134–147. [DOI] [PubMed] [Google Scholar]
- [4].Jeck WR, Sorrentino JA, Wang K, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. Rna. 2013;19(2):141–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Yuan C, Luo X, Zhan X, et al. EMT related circular RNA expression profiles identify circSCYL2 as a novel molecule in breast tumor metastasis. Int J Mol Med. 2020;45(6):1697–1710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Fu X, Zhang J, He X, et al. Circular RNA MAN2B2 promotes cell proliferation of hepatocellular carcinoma cells via the miRNA-217/MAPK1 axis. J Cancer. 2020;11(11):3318–3326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Akhter R. Circular RNA and Alzheimer’s disease. Adv Exp Med Biol. 2018;1087:239–243. [DOI] [PubMed] [Google Scholar]
- [8].Rybak-Wolf A, Stottmeister C, Glažar P, et al. Circular RNAs in the mammalian brain are highly abundant, conserved, and dynamically expressed. Mol Cell. 2015;58(5):870–885. [DOI] [PubMed] [Google Scholar]
- [9].Li Z, Huang C, Bao C, et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol. 2015;22(3):256–264. [DOI] [PubMed] [Google Scholar]
- [10].Zhang Y, Zhang X-O, Chen T, et al. Circular intronic long noncoding RNAs. Mol Cell. 2013;51(6):792–806. [DOI] [PubMed] [Google Scholar]
- [11].Bachmayr-Heyda A, Reiner AT, Auer K, et al. Correlation of circular RNA abundance with proliferation–exemplified with colorectal and ovarian cancer, idiopathic lung fibrosis, and normal human tissues. Sci Rep. 2015;5:8057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Vilades D, Martínez‐Camblor P, Ferrero‐Gregori A, et al. Plasma circular RNA hsa_circ_0001445 and coronary artery disease: performance as a biomarker. Faseb J. 2020;34(3):4403–4414. [DOI] [PubMed] [Google Scholar]
- [13].Song Z, Zhang Q, Zhu J, et al. Identification of urinary hsa_circ _0137439 as potential biomarker and tumor regulator of bladder cancer. Neoplasma. 2020;67(1):137–146. [DOI] [PubMed] [Google Scholar]
- [14].Fanale D, et al. Circular RNA in exosomes. Adv Exp Med Biol. 2018;1087:109–117. [DOI] [PubMed] [Google Scholar]
- [15].Zhang Y, Xue W, Li X, et al. The biogenesis of nascent circular RNAs. Cell Rep. 2016;15(3):611–624. [DOI] [PubMed] [Google Scholar]
- [16].Starke S, Jost I, Rossbach O, et al. Exon circularization requires canonical splice signals. Cell Rep. 2015;10(1):103–111. [DOI] [PubMed] [Google Scholar]
- [17].Ashwal-Fluss R, Meyer M, Pamudurti N, et al. circRNA biogenesis competes with pre-mRNA splicing. Mol Cell. 2014;56(1):55–66. [DOI] [PubMed] [Google Scholar]
- [18].Jeck WR, Sharpless NE.. Detecting and characterizing circular RNAs. Nat Biotechnol. 2014;32(5):453–461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Ivanov A, Memczak S, Wyler E, et al. Analysis of intron sequences reveals hallmarks of circular RNA biogenesis in animals. Cell Rep. 2015;10(2):170–177. [DOI] [PubMed] [Google Scholar]
- [20].Huang C, Liang D, Tatomer DC, et al. A length-dependent evolutionarily conserved pathway controls nuclear export of circular RNAs. Genes Dev. 2018;32(9–10):639–644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Conn SJ, Pillman K, Toubia J, et al. The RNA binding protein quaking regulates formation of circRNAs. Cell. 2015;160(6):1125–1134. [DOI] [PubMed] [Google Scholar]
- [22].You X, Conrad TOF. Acfs: accurate circRNA identification and quantification from RNA-Seq data. Sci Rep. 2016;6(1):38820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Szabo L, Salzman J. Detecting circular RNAs: bioinformatic and experimental challenges. Nat Rev Genet. 2016;17(11):679–692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Pandey PR, Rout PK, Das A, et al. RPAD (RNase R treatment, polyadenylation, and poly(A)+ RNA depletion) method to isolate highly pure circular RNA. Methods. 2019;155:41–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Xiao MS, Wilusz JE. An improved method for circular RNA purification using RNase R that efficiently removes linear RNAs containing G-quadruplexes or structured 3ʹ ends. Nucleic Acids Res. 2019;47(16):8755–8769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Panda AC, De S, Grammatikakis I, et al. High-purity circular RNA isolation method (RPAD) reveals vast collection of intronic circRNAs. Nucleic Acids Res. 2017;45(12):e116–e116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Hansen TB, Venø MT, Damgaard CK, et al. Comparison of circular RNA prediction tools. Nucleic Acids Res. 2016;44(6):e58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Zeng X, Lin W, Guo M, et al. A comprehensive overview and evaluation of circular RNA detection tools. PLoS Comput Biol. 2017;13(6):e1005420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Gao Y, Zhao F. Computational strategies for exploring circular RNAs. Trends Genet. 2018;34(5):389–400. [DOI] [PubMed] [Google Scholar]
- [30].Dobin A, Davis CA, Schlesinger F, et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics. 2013;29(1):15–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Kim D, Langmead B, Salzberg SL. HISAT: a fast spliced aligner with low memory requirements. Nat Methods. 2015;12(4):357–360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Kim D, Pertea G, Trapnell C, et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013;14(4):R36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Horiuchi T, Aigaki T. Alternative trans-splicing: a novel mode of pre-mRNA processing. Biol Cell. 2006;98(2):135–140. [DOI] [PubMed] [Google Scholar]
- [34].Li X, Wu Y. Detecting circular RNA from high-throughput sequence data with de Bruijn graph. BMC Genomics. 2020;21(1):749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Vromman M, Vandesompele J, Volders PJ. Closing the circle: current state and perspectives of circular RNA databases. Brief Bioinform. 2021 Jan 18;22(1):288-297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Zhang XO, Dong R, Zhang Y, et al. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016;26(9):1277–1287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Ma XK, Wang M-R, Liu C-X, et al. CIRCexplorer3: a CLEAR pipeline for direct comparison of circular and linear RNA expression. Genomics Proteomics Bioinformatics. 2019;17(5):511–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Gao Y, Wang J, Zhao F. CIRI: an efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015;16:4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Gao Y, Zhang J, Zhao F. Circular RNA identification based on multiple seed matching. Brief Bioinform. 2018;19(5):803–810. [DOI] [PubMed] [Google Scholar]
- [40].Cheng J, Metge F, Dieterich C. Specific identification and quantification of circular RNAs from sequencing data. Bioinformatics. 2016;32(7):1094–1096. [DOI] [PubMed] [Google Scholar]
- [41].Wang K, Singh D, Zeng Z, et al. MapSplice: accurate mapping of RNA-seq reads for splice junction discovery. Nucleic Acids Res. 2010;38(18):e178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Hoffmann S, Otto C, Doose G, et al. A multi-split mapping algorithm for circular RNA, splicing, trans-splicing and fusion detection. Genome Biol. 2014;15(2):R34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Song X, Zhang N, Han P, et al. Circular RNA profile in gliomas revealed by identification tool UROBORUS. Nucleic Acids Res. 2016;44(9):e87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Jia GY, Wang D-L, Xue M-Z, et al. CircRNAFisher: a systematic computational approach for de novo circular RNA identification. Acta Pharmacol Sin. 2019;40(1):55–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Szabo L, Morey R, Palpant NJ, et al. Statistically based splicing detection reveals neural enrichment and tissue-specific induction of circular RNA during human fetal development. Genome Biol. 2015;16:126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Li X, Chu C, Pei J, et al. CircMarker: a fast and accurate algorithm for circular RNA detection. BMC Genomics. 2018;19(S6):572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Sekar S, Geiger P, Adkins J, et al. ACValidator: a novel assembly-based approach for in silico verification of circular RNAs. Biol Methods Protoc. 2020;5(1):bpaa010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Sun P, Li G. CircCode: a powerful tool for identifying circRNA coding ability. Front Genet. 2019;10(981). DOI: 10.3389/fgene.2019.00981 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Gaffo E, Bonizzato A, Kronnie GT, et al. CirComPara: a multi-method comparative bioinformatics pipeline to detect and study circRNAs from RNA-seq data. Noncoding RNA. 2017;3(1). DOI: 10.3390/ncrna3010008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Zhong S, Wang J, Zhang Q, et al. CircPrimer: a software for annotating circRNAs and determining the specificity of circRNA primers. BMC Bioinformatics. 2018;19(1):292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Meng X, Chen Q, Zhang P, et al. CircPro: an integrated tool for the identification of circRNAs with protein-coding potential. Bioinformatics. 2017;33(20):3314–3316. [DOI] [PubMed] [Google Scholar]
- [52].Li L, Bu D, Zhao Y. CircRNAwrap - a flexible pipeline for circRNA identification, transcript prediction, and abundance estimation. FEBS Lett. 2019;593(11):1179–1189. [DOI] [PubMed] [Google Scholar]
- [53].Jakobi T, Uvarovskii A, Dieterich C. circtools-a one-stop software solution for circular RNA research. Bioinformatics. 2019;35(13):2326–2328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Andrés-León E, Núñez-Torres R, Rojas AM. miARma-Seq: a comprehensive tool for miRNA, mRNA and circRNA analysis. Sci Rep. 2016;6(1):25749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Chen CY, Chuang TJ. NCLcomparator: systematically post-screening non-co-linear transcripts (circular, trans-spliced, or fusion RNAs) identified from various detectors. BMC Bioinformatics. 2019;20(1):3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Zhao J, Li X, Guo J, et al. ReCirc: prediction of circRNA expression and function through probe reannotation of non-circRNA microarrays. Mol Omics. 2019;15(2):150–163. [DOI] [PubMed] [Google Scholar]
- [57].Humphreys DT, et al. Ularcirc: visualization and enhanced analysis of circular RNAs via back and canonical forward splicing. Nucleic Acids Res. 2019;47(20):e123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495(7441):333–338. [DOI] [PubMed] [Google Scholar]
- [59].Hansen TB, Jensen TI, Clausen BH, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495(7441):384–388. [DOI] [PubMed] [Google Scholar]
- [60].Guo JU, Agarwal V, Guo H, et al. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014;15(7):409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Geng HH, Li R, Su Y-M, et al. The circular RNA Cdr1as promotes myocardial infarction by mediating the regulation of miR-7a on its target genes expression. PLoS One. 2016;11(3):e0151753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Han D, Li J, Wang H, et al. Circular RNA circMTO1 acts as the sponge of microRNA-9 to suppress hepatocellular carcinoma progression. Hepatology. 2017;66(4):1151–1164. [DOI] [PubMed] [Google Scholar]
- [63].Lim TB, et al. Targeting the highly abundant circular RNA circSlc8a1 in cardiomyocytes attenuates pressure overload induced hypertrophy. Cardiovasc Res. 2019;115(14):1998–2007. [DOI] [PubMed] [Google Scholar]
- [64].Xia P, et al. A circular RNA protects dormant hematopoietic stem cells from DNA sensor cGAS-mediated exhaustion. Immunity. 2018;48(4):688–701.e7. [DOI] [PubMed] [Google Scholar]
- [65].Holdt LM, Stahringer A, Sass K, et al. Circular non-coding RNA ANRIL modulates ribosomal RNA maturation and atherosclerosis in humans. Nat Commun. 2016;7:12429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Chen N, et al. A novel FLI1 exonic circular RNA promotes metastasis in breast cancer by coordinately regulating TET1 and DNMT1. Genome Biol. 2018;19(1):218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Zeng Y, Du WW, Wu Y, et al. A circular RNA binds to and activates AKT phosphorylation and nuclear localization reducing apoptosis and enhancing cardiac repair. Theranostics. 2017;7(16):3842–3855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Du WW, Fang L, Yang W, et al. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ. 2017;24(2):357–370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Du WW, Zhang C, Yang W, et al. Identifying and characterizing circRNA-protein interaction. Theranostics. 2017;7(17):4183–4191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Stagsted LV, Nielsen KM, Daugaard I, et al. Noncoding AUG circRNAs constitute an abundant and conserved subclass of circles. Life Sci Alliance. 2019;2(3):e201900398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Mo D, Li X, Raabe CA, et al. A universal approach to investigate circRNA protein coding function. Sci Rep. 2019;9(1):11684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [72].Godet AC, David F, Hantelys F, et al. IRES trans-acting factors, key actors of the stress response. Int J Mol Sci. 2019;20(4):924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Chen CY, Sarnow P. Initiation of protein synthesis by the eukaryotic translational apparatus on circular RNAs. Science. 1995;268(5209):415. [DOI] [PubMed] [Google Scholar]
- [74].Wang Y, Wang Z. Efficient backsplicing produces translatable circular mRNAs. Rna. 2015;21(2):172–179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Meyer KD, Patil D, Zhou J, et al. 5ʹ UTR m(6)A promotes cap-independent translation. Cell. 2015;163(4):999–1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Yang Y, Fan X, Mao M, et al. Extensive translation of circular RNAs driven by N(6)-methyladenosine. Cell Res. 2017;27(5):626–641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Legnini I, Di Timoteo G, Rossi F, et al. Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis. Mol Cell. 2017;66(1):22–37.e9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Pamudurti NR, Bartok O, Jens M, et al. Translation of CircRNAs. Mol Cell. 2017;66(1):9–21.e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Zhang M, Huang N, Yang X, et al. A novel protein encoded by the circular form of the SHPRH gene suppresses glioma tumorigenesis. Oncogene. 2018;37(13):1805–1814. [DOI] [PubMed] [Google Scholar]
- [80].Yang Y, Gao X, Zhang M, et al. Novel role of FBXW7 circular RNA in repressing glioma tumorigenesis. JNCI. 2017;110(3):304–315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Liang W-C, Wong C-W, Liang -P-P, et al. Translation of the circular RNA circβ-catenin promotes liver cancer cell growth through activation of the Wnt pathway. Genome Biol. 2019;20(1):84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Zheng X, Chen L, Zhou Y, et al. A novel protein encoded by a circular RNA circPPP1R12A promotes tumor pathogenesis and metastasis of colon cancer via Hippo-YAP signaling. Mol Cancer. 2019;18(1):47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Xia X, Li X, Li F, et al. A novel tumor suppressor protein encoded by circular AKT3 RNA inhibits glioblastoma tumorigenicity by competing with active phosphoinositide-dependent Kinase-1. Mol Cancer. 2019;18(1):131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].World Health Organization . Cardiovascular diseases; 2020. Available from: https://www.who.int/health-topics/cardiovascular-diseases/#tab=tab_1
- [85].Santer L, Bär C, Thum T. Circular RNAs: a novel class of functional RNA molecules with a therapeutic perspective. Mol Ther. 2019;27(8):1350–1363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Zhang Y, Chen Y, Yao H, et al. Elevated serum circ_0068481 levels as a potential diagnostic and prognostic indicator in idiopathic pulmonary arterial hypertension. Pulm Circ. 2019;9(4):2045894019888416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Chen C, Shen H, Huang Q, et al. The circular RNA CDR1as regulates the proliferation and apoptosis of human cardiomyocytes through the miR-135a/HMOX1 and miR-135b/HMOX1 axes. Genet Test Mol Biomarkers. 2020;24(9):537–548. [DOI] [PubMed] [Google Scholar]
- [88].Bahn JH, Zhang Q, Li F, et al. The landscape of microRNA, Piwi-interacting RNA, and circular RNA in human saliva. Clin Chem. 2015;61(1):221–230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Wang Y, Liu J, Ma J, et al. Exosomal circRNAs: biogenesis, effect and application in human diseases. Mol Cancer. 2019;18(1):116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Li Y, Zheng Q, Bao C, et al. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. 2015;25(8):981–984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Hulstaert E, et al. Charting extracellular transcriptomes in the human biofluid RNA atlas. Cell Rep. 2020:33(13):108552. [DOI] [PubMed] [Google Scholar]
- [92].Zhang H, Zhu L, Bai M, et al. Exosomal circRNA derived from gastric tumor promotes white adipose browning by targeting the miR-133/PRDM16 pathway. Int J Cancer. 2019;144(10):2501–2515. [DOI] [PubMed] [Google Scholar]
- [93].Baruah J, Wary KK. Exosomes in the regulation of vascular endothelial cell regeneration. Front Cell Dev Biol. 2019;7:353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [94].Rios-Colon L, Arthur E, Niture S, et al. The role of exosomes in the crosstalk between adipocytes and liver cancer cells. Cells. 2020;9(9):1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Zhang J, Zhang X, Li C, et al. Circular RNA profiling provides insights into their subcellular distribution and molecular characteristics in HepG2 cells. RNA Biol. 2019;16(2):220–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [96].Vausort M, Salgado-Somoza A, Zhang L, et al. Myocardial infarction-associated circular rna predicting left ventricular dysfunction. J Am Coll Cardiol. 2016;68(11):1247–1248. [DOI] [PubMed] [Google Scholar]
- [97].Salgado-Somoza A, Zhang L, Vausort M, et al. The circular RNA MICRA for risk stratification after myocardial infarction. IJC Heart Vasculature. 2017;17:33–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Wang L, Shen C, Wang Y, et al. Identification of circular RNA Hsa_circ_0001879 and Hsa_circ_0004104 as novel biomarkers for coronary artery disease. Atherosclerosis. 2019;286:88–96. [DOI] [PubMed] [Google Scholar]
- [99].Wu WP, Pan Y-H, Cai M-Y, et al. Plasma-derived exosomal circular RNA hsa_circ_0005540 as a novel diagnostic biomarker for coronary artery disease. Dis Markers. 2020;2020:3178642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Wang Y, et al. Exosomal CircHIPK3 released from hypoxia-induced cardiomyocytes regulates cardiac angiogenesis after myocardial infarction. Oxid Med Cell Longev. 2020;2020:8418407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Zhao Z, Li X, Gao C, et al. Peripheral blood circular RNA hsa_circ_0124644 can be used as a diagnostic biomarker of coronary artery disease. Sci Rep. 2017;7(1):39918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Song CL, Wang J-P, Xue X, et al. Effect of circular ANRIL on the inflammatory response of vascular endothelial cells in a rat model of coronary atherosclerosis. Cell Physiol Biochem. 2017;42(3):1202–1212. [DOI] [PubMed] [Google Scholar]
- [103].Holdt LM, Teupser D. Recent studies of the human chromosome 9p21 locus, which is associated with atherosclerosis in human populations. Arterioscler Thromb Vasc Biol. 2012;32(2):196–206. [DOI] [PubMed] [Google Scholar]
- [104].Holdt LM, Beutner F, Scholz M, et al. ANRIL expression is associated with atherosclerosis risk at chromosome 9p21. Arterioscler Thromb Vasc Biol. 2010;30(3):620–627. [DOI] [PubMed] [Google Scholar]
- [105].Burd CE, Jeck WR, Liu Y, et al. Expression of linear and novel circular forms of an INK4/ARF-associated non-coding RNA correlates with atherosclerosis risk. PLoS Genet. 2010;6(12):e1001233–e1001233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [106].Yang L, Yang F, Zhao H, et al. Circular RNA circCHFR facilitates the proliferation and migration of vascular smooth muscle via miR-370/FOXO1/Cyclin D1 pathway. Mol Ther Nucleic Acids. 2019;16:434–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [107].Zhuang JB, Li T, Hu X-M, et al. Circ_CHFR expedites cell growth, migration and inflammation in ox-LDL-treated human vascular smooth muscle cells via the miR-214-3p/Wnt3/beta-catenin pathway. Eur Rev Med Pharmacol Sci. 2020;24(6):3282–3292. [DOI] [PubMed] [Google Scholar]
- [108].Zhang S, Song G, Yuan J, et al. Circular RNA circ_0003204 inhibits proliferation, migration and tube formation of endothelial cell in atherosclerosis via miR-370-3p/TGFbetaR2/phosph-SMAD3 axis. J Biomed Sci. 2020;27(1):11. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- [109].Liu H, Ma X, Mao Z, et al. Circular RNA has_circ_0003204 inhibits oxLDL-induced vascular endothelial cell proliferation and angiogenesis. Cell Signal. 2020;70:109595. [DOI] [PubMed] [Google Scholar]
- [110].Shen L, Hu Y, Lou J, et al. CircRNA0044073 is upregulated in atherosclerosis and increases the proliferation and invasion of cells by targeting miR107. Mol Med Rep. 2019;19(5):3923–3932. [DOI] [PubMed] [Google Scholar]
- [111].Kong P, Yu Y, Wang L, et al. circ-Sirt1 controls NF-κB activation via sequence-specific interaction and enhancement of SIRT1 expression by binding to miR-132/212 in vascular smooth muscle cells. Nucleic Acids Res. 2019;47(7):3580–3593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [112].Mao YY, Wang J-Q, Guo -X-X, et al. Circ-SATB2 upregulates STIM1 expression and regulates vascular smooth muscle cell proliferation and differentiation through miR-939. Biochem Biophys Res Commun. 2018;505(1):119–125. [DOI] [PubMed] [Google Scholar]
- [113].Sun J, Zhang Z, Yang S. Circ_RUSC2 upregulates the expression of miR-661 target gene SYK and regulates the function of vascular smooth muscle cells. Biochem Cell Biol. 2019;97(6):709–714. [DOI] [PubMed] [Google Scholar]
- [114].Michiels C. Physiological and pathological responses to hypoxia. Am J Pathol. 2004;164(6):1875–1882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Konstantinidis K, Whelan RS, Kitsis RN. Mechanisms of cell death in heart disease. Arterioscler Thromb Vasc Biol. 2012;32(7):1552–1562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [116].Turer AT, Hill JA. Pathogenesis of myocardial ischemia-reperfusion injury and rationale for therapy. Am J Cardiol. 2010;106(3):360–368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [117].Jin Q, Chen Y. Silencing circular RNA circ_0010729 protects human cardiomyocytes from oxygen-glucose deprivation-induced injury by up-regulating microRNA-145-5p. Mol Cell Biochem. 2019;462(1–2):185–194. [DOI] [PubMed] [Google Scholar]
- [118].Chen L, Luo W, Zhang W, et al. circDLPAG4/HECTD1 mediates ischaemia/reperfusion injury in endothelial cells via ER stress. RNA Biol. 2020;17(2):240–253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [119].Ji X, Ding W, Xu T, et al. MicroRNA-31-5p attenuates doxorubicin-induced cardiotoxicity via quaking and circular RNA Pan3. J Mol Cell Cardiol. 2020;140:56–67. [DOI] [PubMed] [Google Scholar]
- [120].Lei D, Wang Y, Zhang L, et al. Circ_0010729 regulates hypoxia-induced cardiomyocyte injuries by activating TRAF5 via sponging miR-27a-3p. Life Sci. 2020;262:118511. [DOI] [PubMed] [Google Scholar]
- [121].Zhang J, Gao C, Zhang J, et al. Circ_0010729 knockdown protects cardiomyocytes against hypoxic dysfunction via miR-370-3p/TRAF6 axis. Excli J. 2020;19:1520–1532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [122].Wang K, Gan T-Y, Li N, et al. Circular RNA mediates cardiomyocyte death via miRNA-dependent upregulation of MTP18 expression. Cell Death Differ. 2017;24(6):1111–1120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [123].Li M, Ding W, Tariq MA, et al. A circular transcript of ncx1 gene mediates ischemic myocardial injury by targeting miR-133a-3p. Theranostics. 2018;8(21):5855–5869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [124].Gan J, Yuan J, Liu Y, et al. Circular RNA_101237 mediates anoxia/reoxygenation injury by targeting let‑7a‑5p/IGF2BP3 in cardiomyocytes. Int J Mol Med. 2020;45(2):451–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [125].Sun LY, Zhao J-C, Ge X-M, et al. Circ_LAS1L regulates cardiac fibroblast activation, growth, and migration through miR-125b/SFRP5 pathway. Cell Biochem Funct. 2020;38(4):443–450. [DOI] [PubMed] [Google Scholar]
- [126].Cai L, Qi B, Wu X, et al. Circular RNA Ttc3 regulates cardiac function after myocardial infarction by sponging miR-15b. J Mol Cell Cardiol. 2019;130:10–22. [DOI] [PubMed] [Google Scholar]
- [127].Zhao B, Li G, Peng J, et al. CircMACF1 attenuates acute myocardial infarction through miR-500b-5p-EMP1 axis. J Cardiovasc Transl Res. 2020. DOI: 10.1007/s12265-020-09976-5 [DOI] [PubMed] [Google Scholar]
- [128].Zhang Q, Sun W, Han J, et al. The circular RNA hsa_circ_0007623 acts as a sponge of microRNA-297 and promotes cardiac repair. Biochem Biophys Res Commun. 2020;523(4):993–1000. [DOI] [PubMed] [Google Scholar]
- [129].Shao Y, Zhong P, Sheng L, et al. Circular RNA circDENND2A protects H9c2 cells from oxygen glucose deprivation-induced apoptosis through sponging microRNA-34a. Cell Cycle. 2020;19(2):246–255. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- [130].Du WW, Yang W, Chen Y, et al. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur Heart J. 2017;38(18):1402–1412. [DOI] [PubMed] [Google Scholar]
- [131].Zhou LY, Zhai M, Huang Y, et al. The circular RNA ACR attenuates myocardial ischemia/reperfusion injury by suppressing autophagy via modulation of the Pink1/FAM65B pathway. Cell Death Differ. 2019;26(7):1299–1315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [132].Huang S, Li X, Zheng H, et al. Loss of super-enhancer-regulated circRNA Nfix induces cardiac regeneration after myocardial infarction in adult mice. Circulation. 2019;139(25):2857–2876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [133].Garikipati VNS, Verma SK, Cheng Z, et al. Circular RNA CircFndc3b modulates cardiac repair after myocardial infarction via FUS/VEGF-A axis. Nat Commun. 2019;10(1):4317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [134].Bai M, Pan C-L, Jiang G-X, et al. CircHIPK3 aggravates myocardial ischemia-reperfusion injury by binding to miRNA-124-3p. Eur Rev Med Pharmacol Sci. 2019;23(22):10107–10114. [DOI] [PubMed] [Google Scholar]
- [135].Bai M, Pan C-L, Jiang G-X, et al. CircRNA 010567 improves myocardial infarction rats through inhibiting TGF-β1. Eur Rev Med Pharmacol Sci. 2020;24(1):369–375. [DOI] [PubMed] [Google Scholar]
- [136].Burchfield JS, Xie M, Hill JA. Pathological ventricular remodeling: mechanisms: part 1 of 2. Circulation. 2013;128(4):388–400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [137].Wu QQ, et al. Mechanisms contributing to cardiac remodelling. Clin Sci (Lond). 2017;131(18):2319–2345. [DOI] [PubMed] [Google Scholar]
- [138].Werfel S, Nothjunge S, Schwarzmayr T, et al. Characterization of circular RNAs in human, mouse and rat hearts. J Mol Cell Cardiol. 2016;98:103–107. [DOI] [PubMed] [Google Scholar]
- [139].Cohn JN, Ferrari R, Sharpe N. Cardiac remodeling–concepts and clinical implications: a consensus paper from an international forum on cardiac remodeling. Behalf of an international forum on cardiac remodeling. J Am Coll Cardiol. 2000;35(3):569–582. [DOI] [PubMed] [Google Scholar]
- [140].Hinderer S, Schenke-Layland K. Cardiac fibrosis - A short review of causes and therapeutic strategies. Adv Drug Deliv Rev. 2019;146:77–82. [DOI] [PubMed] [Google Scholar]
- [141].Han J, Zhang L, Hu L, et al. Circular RNA-expression profiling reveals a potential role of Hsa_circ_0097435 in heart failure via sponging multiple MicroRNAs. Front Genet. 2020;11:212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [142].Deng Y, Wang J, Xie G, et al. Circ-HIPK3 strengthens the effects of adrenaline in heart failure by MiR-17-3p - ADCY6 axis. Int J Biol Sci. 2019;15(11):2484–2496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [143].Ni H, Li W, Zhuge Y, et al. Inhibition of circHIPK3 prevents angiotensin II-induced cardiac fibrosis by sponging miR-29b-3p. Int J Cardiol. 2019;292:188–196. [DOI] [PubMed] [Google Scholar]
- [144].Tang CM, Zhang M, Huang L, et al. CircRNA_000203 enhances the expression of fibrosis-associated genes by derepressing targets of miR-26b-5p, Col1a2 and CTGF, in cardiac fibroblasts. Sci Rep. 2017;7:40342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [145].Li H, Xu J-D, Fang X-H, et al. Circular RNA circRNA_000203 aggravates cardiac hypertrophy via suppressing miR-26b-5p and miR-140-3p binding to Gata4. Cardiovasc Res. 2020;116(7):1323–1334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [146].Danowski N, Manthey I, Jakob HG, et al. Decreased expression of miR-133a but Not of miR-1 is associated with signs of heart failure in patients undergoing coronary bypass surgery. Cardiology. 2013;125(2):125–130. [DOI] [PubMed] [Google Scholar]
- [147].Carè A, Catalucci D, Felicetti F, et al. MicroRNA-133 controls cardiac hypertrophy. Nat Med. 2007;13(5):613–618. [DOI] [PubMed] [Google Scholar]
- [148].Wang K, Long B, Liu F, et al. A circular RNA protects the heart from pathological hypertrophy and heart failure by targeting miR-223. Eur Heart J. 2016;37(33):2602–2611. [DOI] [PubMed] [Google Scholar]
- [149].Wen J, Liao J, Liang J, et al. Circular RNA HIPK3: a Key Circular RNA in a Variety of Human Cancers. Front Oncol. 2020;10(773). DOI: 10.3389/fonc.2020.00773 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [150].Zhou B, Yu JW. A novel identified circular RNA, circRNA_010567, promotes myocardial fibrosis via suppressing miR-141 by targeting TGF-β1. Biochem Biophys Res Commun. 2017;487(4):769–775. [DOI] [PubMed] [Google Scholar]
- [151].Zhu Y, Pan W, Yang T, et al. Upregulation of circular RNA CircNFIB attenuates cardiac fibrosis by sponging miR-433. Front Genet. 2019;10:564. [DOI] [PMC free article] [PubMed] [Google Scholar]