

Abstract
Although most human papillomavirus (HPV) infections are harmless, persistent infection with high-risk types of HPV is known to be the leading cause of cervical cancer. Following the infection of the epithelium and integration into the host genome, the oncogenic proteins E6 and E7 disrupt cell cycle control by inducing p53 and retinoblastoma (Rb) degradation. Despite the FDA approval of prophylactic vaccines, there are still issues with cervical cancer treatment; thus, many therapeutic approaches have been developed to date. Due to strong immunogenicity, a high capacity for packaging foreign DNA, safety, and the ability to infect a myriad of cells, adenoviruses have drawn attention of researchers. Adenovirus vectors have been used for different purposes, including as oncolytic agents to kill cancer cells, carrier for RNA interference to block oncoproteins expression, vaccines for eliciting immune responses, especially in cytotoxic T lymphocytes (CTLs), and gene therapy vehicles for restoring p53 and Rb function.
Keywords: adenovirus, cervical Cancer, HPV Infection, HPV Oncoproteins, human Papillomavirus, vaccine
Introduction
Cervical cancer is the fourth most common cancer in women and the seventh most common cancer worldwide [1]. According to the global cancer observatory (GLOBOCAN) report in 2018, there were an estimated 570,000 new cases and 311,000 deaths from cervical cancer [2]. Unfortunately, cervical cancer affects young women and is the most common gynecologic malignancy in women under 35 years of age [3]. Although the incidence and mortality rates of cervical cancer have declined over the last few decades [2], several studies have indicated that socioeconomic levels play an essential role in the incidence, survival, and mortality rates of cervical cancer [4]. Approximately 90% of cervical cancer deaths occur in low-income and middle-income countries [5] due to several factors, including ineffective and inadequate treatment, poor hygiene conditions [6–8], and the absence of preventive [9] and screening programs [10].
Infection with human papillomavirus (HPV) is the most common cause of cervical cancer [11]. According to the International HPV Reference Center, the HPV family consists of 202 different genotypes [12]. HPV genotypes are categorized into two groups based on their ability to lead to malignancy: low-risk types, including HPV-6, 11, 40, 42, 43, 44, 54, 61, and 72, and high-risk types, including HPV16, 18, 31, 35, 39, 45, 51, 52, 56, 58, 66, and 68. While low-risk types are responsible for genital warts, high-risk types are associated with 99.7% of cervical cancer cases [13]. It has been demonstrated that HPV16 and 18 are the most carcinogenic genotypes [14]. Due to the carcinogenesis of HPV, various approaches have been developed for the prevention and treatment of HPV-induced cervical cancer. During the last two decades, adenovirus vectors have been developed as an effective tool in cancer therapy, and many clinical trials have been conducted or are in progress. In this review, we will discuss the tumorigenic mechanisms of HPV, therapeutic strategies for cervical cancer, and finally, how adenoviruses (Ads) are applied in cervical cancer therapy.
HPV structure and cervical cancer pathology
HPV genome and life cycle
HPVs are small (50–60 nm), nonenveloped, double-stranded DNA viruses with a genome size of ~8 kb belonging to the family Papillomaviridae [15]. The genome of HPV includes three regions: (a) the early region, which encodes three regulatory proteins (E1, E2, and E4) and three oncoproteins (E5, E6, and E7); (b) the late region, which encodes two structural proteins (L1 and L2); and (c) regulatory regions for HPV DNA replication and transcription, which are controlled by a long control region [16].
Although most sexually active women are infected by HPV, the majority of HPV infections are transitory and cleared by the immune system. However, HPV infection can persist in some women and develop into cervical intraepithelial neoplasia (CIN) and cancer [17–19]. CIN is a precancerous lesion, and depending on its grade, it is classified as CIN1, CIN2, and CIN3 [20]. Although CIN1 is considered a low-grade squamous intraepithelial lesion and 70%–80% of CIN1 lesions spontaneously become undetectable or regress without treatment [21, 22], CIN2 and CIN3 are high-grade SILs that can lead to invasive cervical cancer [23].
Following microwounds in the skin, oropharyngeal areas, and genital organs, HPV virions attach to basal epithelial cell receptors via capsid proteins [24]. The initial attachment of the capsid protein L1 to a cell surface receptor, heparan sulfate proteoglycans facilitate conformational changes of the capsid protein L2 [25, 26], subsequently resulting in L2 cleavage at the furin cleavage site, which is conserved among all HPVs [27]. After binding to the receptor, HPV virions are internalized into cells by clathrin/caveolar-mediated endocytosis [28, 29]. Once the viral genome enters the nucleus, viral DNA replication starts and generates a low-copy-number (50–100 copies/cell) genome. Following the differentiation of daughter cells, the viral genome is amplified concomitant with increasing E1 and E2 replication protein levels [30]. L1 and L2 capsid proteins are expressed in the terminally differentiated layer of the epithelium, viral particles are assembled and released, and the life cycle is repeated. Although the integration of HPV DNA into the host cell is not a part of the normal life cycle of the virus, high-risk HPV DNA is often incorporated into the host cell genome [31–33]. Integration of high-risk HPV DNA may result in the mutation and/or deletion of both host and viral genes [34–36]. Since the viral E2 protein acts as a transcriptional repressor of E6 and E7 and viral DNA integration disrupts the E2 sequence, and the deletion of E2 leads to increased expression of E6 and E7 [37, 38]. The overexpression of the E6 and E7 oncoproteins causes the deregulation of the cell cycle and several pathways, leading to carcinogenesis.
HPV oncoproteins
HPV E5 oncoprotein
Although E6 and E7 are known as the main oncoproteins of high-risk HPVs, it has been demonstrated that E5 also contributes to tumor progression and can augment the transforming activities of E6 and E7 [39]. Figure 1 shows the carcinogenic properties of the HPV E5 oncoprotein related to enhanced proliferation signaling. HPV E5 is a small (83 amino acids) and highly hydrophobic protein with unclear subcellular localization [40]. Due to its involvement in channel formation, HPV E5 is classified as a family of channel-forming viral membrane proteins called viroporins, which modulate ion homeostasis, virion production, vesicle trafficking, and viral genome entry [41]. It has been demonstrated that the transforming ability of HPV E5 is related to the modulation of epidermal growth factor receptor (EGFR) signaling. HPV E5 binds and inhibits the 16K subunit activity of the vacuolar H+-ATPase (v-ATPase), which alters the endosomal acidification process and degradation of EGFR, leading to EGFR recycling to the plasma membrane and increased EGFR expression [42]. HPV E5 can also delay and inhibit the degradation of EGFR by interfering with fusion between early and late endosomes [43] and preventing the ubiquitination of activated EGFR [44]. Furthermore, HPV E5 activates mitogen-activated protein kinase (MAPK) signaling in EGF-dependent and EGF-independent manners [45]. Once activated, MAPKs translocate into the nucleus and elicit many alterations in the expression profile of cells by phosphorylating different transcription factors [40]. HPV E5 also mediates the immune evasion of the virus by downregulating major histocompatibility complex class I (MHC I), which reduces viral epitope recognition by CD8+ T cells [46, 47]. Moreover, HPV E5 can modulate apoptosis induced by Fas ligand (FasL) and TRAIL [48]. Figure 2 shows the carcinogenic properties of HPV oncoproteins through impairing apoptosis and enhancing the progression of the cell cycle.
Fig. 1.

Carcinogenesis property of HPV E5 oncoprotein through enhancing proliferation signaling.
Fig. 2.

The carcinogenesis properties of HPV oncoproteins through impairing apoptosis and enhancing progression of the cell cycle.
HPV E6 oncoprotein
HPV E6 protein is a 160-amino acid protein with two zinc-binding motifs that is associated with HPV-related malignancies [49]. The most important role of the HPV E6 oncoprotein is the degradation of p53, a tumor suppressor protein. The E6 oncoprotein binds to a short leucine consensus sequence (LxxLL) within ubiquitin ligase E6-Associated Protein (E6-AP), and the E6/E6AP heterodimer targets p53 for proteasomal degradation [50]. The HPV E6 oncoprotein can also interfere with cellular apoptosis by accelerating the degradation of the proapoptotic protein procaspase 8 (CASP8) [51, 52], preventing the entrance of BAK proteins into the mitochondria [53], blocking the Fas/FasL pathway [54], binding to tumor necrosis factor receptor 1 (TNFR1) and inhibiting TNFR1-mediated apoptosis [55]. Furthermore, the HPV E6 oncoprotein can deregulate the cell cycle by overcoming its checkpoints. For example, the E6-AP-E6-p53 pathway downregulates miR-34a expression, resulting in p18Ink4c upregulation and entrance into S phase [56, 57]. E6 can inactivate the p300/CBP complex, a transcriptional coactivator, which is crucial for regulating differentiation and cell cycle progression [58, 59]. To persist in the host epithelium, the E6 oncoprotein decreases the immune response against HPV by downregulating the responses of Interferon α [60], Interferon β [61], and Interferon κ [62]. Another transforming ability of the E6 protein is related to inducing unlimited cell proliferation by enforcing the expression of human telomerase (hTERT) via the degradation of X-box binding 1–91 (NFX1-91), an hTERT transcriptional repressor [63], and the activation of the hTERT promoter [64]. Moreover, E6 contains a PDZ-binding domain (PBM) that mediates its binding to cellular proteins containing PDZ domains, such as Dlg, MAGI-1, and Scribble, leading to their proteolytic degradation and enhancing cell invasion [65].
HPV E7 oncoprotein
The HPV E7 protein is a small (~100 amino acids) acidic protein [66]. The N-terminal sequence of the E7 protein is similar to the domains of the adenovirus E1A protein and simian vacuolating virus 40 large tumor antigen (T Ag) and is designated conserved regions 1 and 2 (CR1 and CR2) [67]. The major transforming characteristic of HPV E7 is its interaction and the inactivation of the retinoblastoma (Rb) tumor suppressor protein pRB by its conserved Leu-X-Cys-X-Glu (LXCXE) motif in the CR2 domain [68]. Following the E7 interaction, pRB is degraded by cullin 2-containing E3 ubiquitin ligases, resulting in the release of the E2F transcription factor and enhancing cell proliferation [69]. The HPV E7 oncoprotein also induces unchecked cell proliferation by inactivating the cyclin inhibitors p21 and p27, proteins that tightly regulate cellular proliferation [70]. Similar to the E6 protein, E7 can interfere with interferon signaling [71, 72]. It has been shown that both the E6 and E7 oncoproteins promote angiogenesis by downregulating the expression of maspin and thrombospondin-1, acting as angiogenic inhibitors, and upregulating the expression of IL-8 and vascular endothelial growth factor (VEGF) [73]. Angiogenesis is a cancer hallmark that provides nutrients and oxygen to tumor cells [74–76].
Therapeutic strategies against HPV
There are three approved HPV preventive vaccines based on HPV L1-derived virus-like particles: Gardasil, Gardasil 9, and Cervarix [77]. Moreover, prophylactic vaccines based on L2, especially based on the conserved N-terminal region, are being developed. For example, adenovirus vectors have been designed to carry L2 protein epitopes from different HPV types as a preventive vaccine to induce neutralizing antibodies (NABs) against HPV infection [78, 79]. Due to the low immunogenicity of L2-based vaccines, using adjuvants such as monophosphoryl lipid A and alum can enhance immune responses [79]. Although approved prophylactic vaccines have shown great success and reduced HPV infections and HPV-related diseases [77, 80], they are insufficient in cases of existing infection and are not very cost-effective [77, 81]. Thus, several therapeutic approaches have been developed for the control and eradication of established HPV infections. The majority of therapeutic vaccines are based on oncoproteins E6 and E7 to deliver them to antigen-presenting cells (APCs) to activate HPV antigen-specific CD8+ cytotoxic T cells (CTLs) or CD4+ helper T cells [82].
Biotherapeutic approaches against HPV are mainly based on peptides/proteins, nucleic acids, whole cells (dendritic cells (DCs) and tumor cells), and live vector vaccines. Peptide-/protein-based vaccines are peptide segments containing epitopes or antigenic proteins themselves. Although these vaccines are safe, stable, and easy to produce, they require adjuvants and have reduced immunogenicity [83]. Most protein-based vaccines are presented through MHC II, which induces the production of antibodies instead of CTL responses [84]. In nucleic acid-based vaccines, DNA- or RNA-encoding sequences of target antigens are inserted into disabled vectors. Compared to DNA-based approaches, RNA-based vaccines have lower stability [82]. While both DNA- and RNA-based approaches lead to sustained antigen expression for a longer duration, RNA-based vaccines are safer due to the low risk of chromosomal integration [85]. In the case of whole cell-based vaccines, DCs are loaded with HPV antigens ex vivo and subsequently delivered to the infected host [86–88]. Although this approach is expensive and labor intensive, it induces robust immune responses as DCs act as natural adjuvants [89]. In tumor cell-based vaccines, tumor cells are isolated and manipulated to express immune-modulatory molecules [90, 91]. In the vector platform, live vector-based vaccines are classified as bacterial and viral vectors. Although live vectors produce strong humoral and cellular immune responses [92], preexisting immunity against vectors and safety risks for immunocompromised individuals are the main challenges in their application in the area of cancer treatment [93, 94].
Adenovirus structure, biology, and vectors
Adenoviruses (Ads) are large (~150 nm), nonenveloped, double-stranded DNA viruses with a genome size of ~36 kb [95] that encode four early (E1, E2, E3, and E4) and five late (L1-L5) genes [96]. The capsid of Ads comprises three major proteins (hexon, penton base, and fiber) and four minor proteins (IIIa, VI, VIII, and IX) [97]. There are 57 different serotypes of human Ad (HAd) divided into seven species (A–G) [98]. Coxsackievirus-Ad receptor (CAR) is the main receptor for the A, C, E, and F species, while desmoglein-2, CD46, CD80, and CD86 are alternative receptors for the B and D species [96]. To infect cells, the fiber knob domain of Ads interacts with cell surface receptors, and the virus internalizes into the cell through the binding of the arginine-glycine-aspartic acid (RGD) motif on the viral penton base to cellular integrins, mainly αvß3 and αvß5 [99]. Following the internalization and degradation of the viral protein coat, the early and late genes of Ad are transcribed in the host cell nucleus, leading to virus assembly and release from the cell.
One of the most important disadvantages of using Ads is preexisting antibodies against them due to their prevalence in the general population [100], hindering the utility of Ads derived from prevalent serotypes [101]. Different approaches have been developed to address this challenge, including coating Ads with polymers and using cellular carriers [102]. For example, O’Riordan et al. reported that shielding Ad with polyethylene glycol (PEG) could protect Ads from NABs while retaining infectivity [103]. However, there are significant advantages to using adenovirus vectors; their strong immunogenicity makes them as ideal candidates for applications in vaccination and oncolysis [104]. In addition, since most human cell primary receptors for Ads are integrin receptors, they can transduce and infect different cell types, including both dividing and nondividing cells [105, 106]. In terms of safety, Ad vectors are maintained as episomal DNA and do not integrate into the host genome [106]. Although common Ad vectors can deliver transgenes up to 8 kb in size [107], the newer generation of Ad vectors has the capacity for the delivery of foreign genes up to ~37 kb [108]. Figure 3 shows the structure of different adenovirus vectors. These advantages led to the approval of two Ad-based vectors by the Chinese state FDA (Food and Drug Administration) for head and neck malignancy, including Gendicine, a recombinant Ad expressing p53 (Ad5RSV-p53) [109], and Oncorine (H101), an oncolytic Ad vector (E1B-55K/E3-deleted) [110].
Fig. 3. Genome structure of adenovirus serotype 5 and different generations of adenovirus vectors.

The adenovirus genome encodes early (E) and late (L) genes in which the E1A protein, an immediate early protein, induces the expression of delayed early proteins, including E1B, E2A, E2B, E3, and E4. In first-generation adenovirus vectors, the E1 and E3 regions were deleted, and in the second generation, E1, E2, E3, and/or E4 were deleted. Helper-dependent adenovirus vectors only contain inverted terminal repeats (ITRs) and packaging signals. Oncolytic vectors are another class of adenovirus vectors. For example, Delta-24 vectors with a deletion in the E1A region replicate in and lyse tumor cells with a defect in the pRB pathway.
Using adenoviruses in cervical cancer therapy
Today, several clinical trials are ongoing for the treatment of cervical cancer using various therapeutic agents. For example, VB10.16, a potent therapeutic DNA plasmid vaccine, was designed to deliver HPV16 E7/E6 to patients with HPV 16-positive cervical cancer to induce strong immune responses (NCT04405349). Although there are no clinical trials regarding the use of adenovirus vectors in the treatment of cervical cancer, we discuss different strategies for using these vectors in the treatment of HPV-induced cervical cancer below, including the use of oncolytic Ads, gene therapy for restoring p53 and Rb, eliciting immune responses (especially CTLs), and carriers for RNA interference to block oncoproteins expression (Fig. 4).
Fig. 4. Different strategies for using adenovirus vectors in the treatment of HPV-induced cervical cancer.
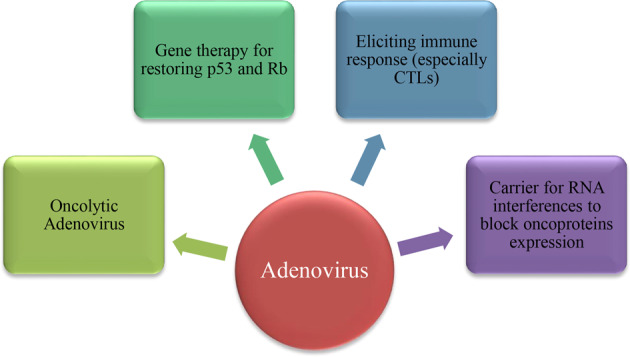
Oncolytic Ads infect tumor cells and lyse them, leading to cell disruption and the release of molecules that induce immune responses against cancer. Ads have also been used to deliver tumor suppressor genes, such as p53 and Rb, to restore their normal function in cancer cells. E6- and E7-expressing Ads can enhance immune responses, especially CTLs, against these oncoproteins. The incorporation of antisense RNAs and microRNAs (miRNAs) against oncoproteins in Ad vectors and introduction into cervical cancer cells can block the expression of the HPV oncoproteins E6 and E7.
Oncolytic adenovirus
Oncolytic Ads are engineered to replicate in and kill cancer cells selectively. Once the infected tumor cell is lysed, released virus progenies infect neighboring cancer cells, and the bloodstream may also help virions reach distant metastases [111]. Oncolytic Ads also induce immune responses against tumor cells by releasing tumor-related antigens [99]. Bauerschmitz et al. constructed an Ad5‐Δ24RGD oncolytic virus, which could selectively replicate in cervical cancers defective in the Rb‐p16 pathway and infect integrin-upregulated cancer cells. They indicated that the intratumoral injection (IT) of oncolytic Ads significantly reduced tumor growth in a murine model of cervical cancer. They also demonstrated the therapeutic benefit of the intravenous (IV) delivery of Ad5‐Δ24RGD. Although biodistribution analysis detected viral tropism toward the liver, spleen, lungs, heart, and kidney 18 h after IV delivery, there was no replication of Ad5‐Δ24RGD in human peripheral blood mononuclear cells, revealing its safety [112]. Oncolytic Ad expressing a p53 variant, which is resistant to degradation by the E6 oncoprotein, enhances the efficacy of oncolytic Ads against cervical carcinoma cells and HPV-immortalized keratinocytes up to 1000-fold [113]. Oncolytic Ads can also enhance the therapeutic efficacy of conventional cancer therapy agents. Yoon et al. investigated Ad-ΔE1B55 and Ad-ΔE1B19/55 therapeutic effects in combination with cisplatin. They showed that in vitro cell cytotoxicity, cell killing, and apoptosis induction with the combination of the double-deleted Ad Ad-ΔE1B19/55 along with cisplatin was greater than that of the single-deleted Ad Ad-ΔE1B55 plus cisplatin. The IT injection of oncolytic Ads and intraperitoneal administration of cisplatin 24 h later in a human cervical xenograft model led to a reduction in tumor size; the greatest effect was reported in the Ad-ΔE1B19/55 plus cisplatin group with an ~96% reduction in tumor size, and after 60 days, all of the animals in the Ad-ΔE1B19/55 plus cisplatin group were alive. These beneficial effects could be related to the induction of apoptosis by chemotherapy agents that have augmented viral distribution [114]. Mechanistically, Ad particles are included in or associated with apoptotic bodies due to treatment with a chemotherapy agent, which are phagocytosed by neighboring cells; thus, the phagocytosis of apoptotic bodies augment viral spread and distribution [115]. In another study, E1A-deleted and HPV16 E2-expressing oncolytic Ad (M5) was able to deplete HPV16 E6 and E7 oncoproteins in cervical cancer cells and an in vivo murine model due to negative effects of E2 on the transcription of E6 and E7 oncogenes and proapoptotic characteristics of the HPV E2 protein. The IT injection of M5 also inhibited tumor growth better than the IT injection of Adv5/dE1A. A combination of M5 with radiation led to synergic antitumor effects both in vitro and in vivo. Radiation increased viral yields by enhancing Ad uptake [116], which might have been due to inducing changes at the cell surface, cytoplasm, and nucleus [117]. M6, an oncolytic Ad carrying an antisense sequence against HPV 16 E6/E7, significantly improved the survival of tumor-bearing mice in combination with radiotherapy, inhibited the expression of the E6 and E7 oncogenes and induced apoptosis in HPV 16-positive cervical cancer cells [118].
Targeting the expression of oncoproteins
As discussed above, the oncoproteins E5, E6, and E7 are the major players in HPV-induced cervical tumorigenesis through different mechanisms. Thus, they are the most effective targets in the therapeutic approach. Antisense RNAs, as a type of noncoding RNA (ncRNA), are small (19–23 nucleotides) DNA transcripts that complement mRNA and are used for regulating gene expression at multiple levels [119]. Hamada et al. used a recombinant adenoviral vector to introduce antisense transcripts of the E6 and E7 oncogenes of HPV16 into cervical cancer cells. They detected HPV 16 E6/E7 antisense RNA for 14 days in Ad5CMV-HPV 16 AS-infected CaSki cells. Ad5CMV-HPV 16 AS inhibited the growth of cervical cancer cells, decreased E6 and E7 expression, and increased p53 and pRb expression. Morphological changes, as well as TUNEL assays and flow cytometry analysis, demonstrated that the infection of SiHa cells with the Ad5CMV-HPV 16 AS at an MOI of ten induced apoptosis. Ad5CMV-HPV 16 AS also inhibited tumor growth by 84% in a tumor-bearing nude mouse [120].
MicroRNAs (miRNAs) are other types of ncRNAs consisting of 21–25 nucleotides and regulate gene expression by blocking mRNA translation [80]. Bonetta et al. directed artificial miRNAs against HPV16/18 E6 using recombinant Ad vectors (Ad16 and Ad18). They observed E6 knockdown in HeLa and SiHa cell lines after treatment with Ad18 and Ad16, respectively. Furthermore, the level of the p53 tumor suppressor was increased, and the expression of miRNAs against E6 led to cell death in both cell lines. Mechanistically, E6 knockdown through miRNA-expressing Ads induced cell death by activating the intrinsic pathway of apoptosis. Moreover, the daily IT injection of 5 × 109 pfu miRNA-expressing Ad (Ad18) in HeLa-bearing nude mice for 5 days resulted in a significantly reduced tumor size and improved the survival of mice 8 weeks postinjection [121]. This group also developed a high-affinity bivalent ligand expressing Ad (Ad-PDZ-LxxL) to specifically target E6 of all high-risk HPVs. The construct was able to induce the death of HPV-positive cells, increase levels of p53 and its transcriptional target p21, and induce apoptosis by restoring p53 [122].
Inducing immune responses (with a vaccine)
Several strategies have been developed to induce robust immune responses against E6 and E7. The majority of immunotherapies aim to efficiently deliver E6 and E7 oncoproteins to APCs to activate HPV antigen-specific CTLs or CD4+ helper T cells, which can reduce and prevent tumor growth [81]. The goal of most approaches to induce immune responses is activating and enhancing CD8+ T cell immunity, which plays an essential role in the clearance of virus-infected cells.
Since both E6 and E7 oncoproteins are dominant at different stages of tumor development, most therapeutic approaches have targeted both oncoproteins. Cuburu et al. evaluated the efficacy of Ad vectors expressing HPV16 E6 and E7 fusion oncoproteins in inducing CD8+ T cell responses following prime-boost immunization via homologous or heterologous routes combining the intravaginal (Ivag) and intramuscular (IM) routes. Due to the induction of NABs against the Ad capsid, they used alternative Ad26 and Ad35 serotypes to allow the comparison of the route of injection without NAB interference. They indicated that the most promising administration order for inducing higher circulating and cervicovaginal-resident CD8+ T cell responses was IM prime followed by Ivag boost, whereas the order of serotype usage did not significantly affect the magnitude of CD8+ T cell responses. Furthermore, immunization with Ad35 IM prime/Ad26 Ivag boost induced E6- and E7-specific polyfunctional cytokine-secreting CD8+ T cells [123]. In contrast to HPV pseudovirions that require the disruption and permeabilization of the epithelium to infect basal cells [124], Ad vectors are able to transduce the intact cervicovaginal epithelium, which might be due to the expression of attachment factors and Ad-specific receptors [123] and can lead to the disruption of tight junctions and subsequent spread of viral particles [125]. In addition, it has been shown that E6- and E7-expressing Ads do not exhibit systematic toxic responses in primates [126].
Because of its better characterized immunological properties, therapeutic vaccines have been developed to elicit immune responses against E7 than against E6. Jin et al. used E7- and IL-12-expressing Ads (AdE7 and AdIL-12) to measure their antitumor effects against the TC-1 tumor model in mice. The IT coadministration of AdE7 and AdIL-12 showed significant antitumor effects, including reduced tumor size, higher levels of INF-γ and E7-specific antibodies, and more T helper cell proliferation than single treatment with either AdE7 or AdIL-12. Since only the AdE7 and AdIL-12 coadministration groups exhibited CTL responses, it could suggest that the majority of tumor suppression effects were mediated by CD8+ T cells [127]. In another study, Ahn et al. investigated the utility of AdIL-12 as an adjuvant for the HPV16 E7 subunit vaccine in an HPV16 E6/E7-associated cervical cancer mouse model. Combined injection using AdIL-12 plus E7 significantly reduced tumor size and enhanced IFN-γ production from E7-specific CD8+ T cells, suggesting that AdIL-12 plus E7 enhanced immune responses against tumors by increasing CTL expansion [128]. The enhanced immune response through coadministration might be due to IL-12 effects on the maturation of TH1 cells and CTLs [129] and the promotion of IFN-γ production by NK cells [130]. Gumez-Gutierrez et al. designed an Ad vector expressing a calreticulin (CRT)-HPV16 E7 fusion protein (Ad-CRT/E7) to test its prophylactic and therapeutic effects in an E7-expressing mouse cervical cancer model. In the prophylactic strategy, challenging mice with TC-1 cells one week after immunization with Ad-CRT/E7 led to complete protection against tumor growth, whereas immunization with AdE7 only protected 20% of mice. In the therapeutic strategy, vaccination with Ad-CRT/E7 significantly increased E7-specific T cell proliferation, IFN-γ production, and cytotoxic activity [131]. It has been demonstrated that CRT enhances MHC I presentation by delivering peptides into the endoplasmic reticulum [132, 133]. In addition, to enhance the efficacy of DC-based vaccination against HPV16-induced cervical cancer, Tillman et al. used bispecific antibodies to retarget AdE7 toward CD40 on DCs due to the CAR deficiency of DCs. They indicated that targeting DCs with AdE7 efficiently initiated antigen-specific immunity and CD8+ T cell responses toward HPV16 E7-expressing tumors. Furthermore, targeted DCs significantly reduced tumor growth and extended the survival of mice [134].
In addition to the E6 and E7 oncoproteins, other HPV proteins have also been applied to increase immune responses against HPV-induced cervical cancer. For example, it has been reported that the IM injection of HPV16 E5-expressing Ads reduces tumor growth by eliciting CD8+ T cell but not CD4+ T cell responses [135]. Furthermore, E1/E2 from cottontail rabbit papillomavirus (CRPV) expressed by an Ad vector has been shown to induce strong immunity against CRPV infection in rabbits [136].
Gene therapy
Gene therapy is the delivery of genetic material into target cells to express it correct abnormalities and have a therapeutic effect. One approach for the gene therapy of cancer cells is delivering tumor suppressor genes to restore their normal function. Since two tumor suppressors, p53 and Rb, are degraded by HPV oncoproteins, delivering wild-type p53 and Rb or their homologs may lead to tumor regression. It has been shown that introducing wild-type p53 into HPV-positive cervical cancer cells with Ad inhibited tumor cell growth by inducing apoptosis and arresting the cell cycle [137–139]. In addition, it has been shown that Ad-p53 enhanced the chemosensitization and radiosensitization of tumor cells. Inhibitory effects of the combination of paclitaxel and Ad-53 on HeLa cells were greater than those of monotherapy, and this combinatory treatment increased tumor cell apoptosis and reduced VEGF expression [140]. Xiao et al. investigated the efficacy and safety of Ad-p53 combined with chemotherapy (cisplatin + vincristine + bleomycin) in 40 patients with locally advanced cervical cancer. The intravenous administration of chemotherapy combined with the IT injection of 1 × 1012 VP Ad-p53 reduced tumor size by 15.25 ± 4.00 cm2, whereas tumor size reduction in the chemotherapy group was 11.42 ± 2.78 cm2. The combination regimen also significantly reduced the expression of VEGF, mutant p53, and microvessel density, without significant adverse effects [141]. Moreover, a combination of the IT injection of Ad-p53 with radiotherapy for the treatment of cervical cancer patients was safe and enhanced the antitumor efficacy of radiotherapy, including the 5-year overall survival rate and 5-year progression-free survival rate [142]. Although introducing wild-type p53 with Ad has shown effective antitumor activity, HPV E6-expressing cervical cancer cells are moderately resistant to the antitumor efficacy of exogenous p53 [143]. Since the p53 homologue p73 is resistant to HPV E6-inducing degradation and performs similar functions to p53 [144–146], Das et al. developed an Ad vector carrying p73β (Ad-p73) and evaluated its antitumor effects on HPV 16 E6-expressing cells. The infection of SW480 cells with Ad-p73 at an MOI of 20 increased the expression of p73β and its target gene p21WAF1/CIP1. Ad-p73 also inhibited tumor cell growth, arrested the cell cycle in the G1 phase, reduced DNA synthesis in cells, and induced apoptosis. Furthermore, Ad-p73 inhibited the proliferation of HeLa and HPV 16 E6-expressing cells more effectively than Ad-p53 [147].
In addition to p53, Rb is the other tumor suppressor protein that is degraded in cervical cancer development. Rb blocks cells entering S phase of the cell cycle and prevents aberrant proliferation. In some studies, Rb has been introduced into cancerous cells to restore its tumor suppressive function. Bourgo et al. activated the Rb pathway using the ectopic expression of an active allele of RB (PSM-RB) or overexpression of the p16ink4a tumor suppressor with an Ad vector in HPV-positive cervical cancer cells. Although p16ink4a expression did not affect cell cycle progression, the expression of PSM-RB induced cell cycle arrest and downregulated E2F gene expression in cervical cancer cells. The sustained expression of PSM-RB prevented the proliferation of SiHa cells and led to cell death and cell cycle inhibition [148]. However, it has been shown that Ad-Rb reduces p53-induced apoptosis in cervical cancer cells; thus, Rb may not be an ideal candidate for gene therapy in cervical cancer [149].
Conclusions
HPV is the leading cervical cancer-causing infectious agent. In addition to traditional treatments for cancer, such as chemotherapy and radiotherapy, prophylactic vaccines have also been developed to reduce the incidence of HPV-induced cervical cancer. The failure of prophylactic vaccines to treat existing cases as well as the high cost of these vaccines have made the development of therapeutic vaccines and tumor-limiting approaches a goal for most gynecology and oncology scientists. Because of several advantages, adenovirus vectors have been investigated through different approaches to control and treat HPV-induced cervical cancer. Although using Ad in cancer therapy has been considered, including targeting to the tumor site, enhancing IT spread, and avoiding liver and lung tropism, the well-characterized structure and genome of Ads allow scientists to modify and adapt it to increase their selectivity, potency of transduction, and antitumor effects. Genetic and chemical modifications of the Ad structure can be applied for targeting Ads toward specific tumors and reduce their tropism toward the liver and lung, such as by constructing chimeric Ads, shielding Ads with polymers, and incorporating tumor-specific promoters. Moreover, using fusogenic proteins, junction-opening peptides and extracellular matrix degrading enzymes can improve Ad IT spread. Therefore, given the solutions to the concerns raised about Ads and the advantages of these vectors, including their versatility, ability to be produced in large volumes, and ease of construction, adenovirus vectors are a valuable candidate for the design of prophylactic and therapeutic vaccines. Since most clinical trials are in the early phases, using Ad vectors as a safe and efficient standard therapeutic agent needs more investigation. In addition, it has become clear that monotherapy using Ads may be insufficient as a therapeutic agent; thus, shifting from monotherapy toward combinational therapy with other therapeutic agents, such as immune checkpoint inhibitors or CAR-T cells, will enhance antitumor activities.
Competing interests
The authors declare no competing interests.
Contributor Information
Babak Negahdari, Email: b-negahdari@sina.tums.ac.ir.
Hami Kaboosi, Email: h.kaboosi@iauamol.ac.ir.
References
- 1.LaVigne K, Leitao MM. Cervical cancer prevention. Fundam Cancer Prev Fourth Ed. 2019;13:629–52. [Google Scholar]
- 2.Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. doi: 10.3322/caac.21492. [DOI] [PubMed] [Google Scholar]
- 3.Pelkofski E, Stine J, Wages NA, Gehrig PA, Kim KH, Cantrell LA. Cervical Cancer in Women Aged 35 Years and Younger. Clin Ther. 2016;38:459–66. doi: 10.1016/j.clinthera.2016.01.024. [DOI] [PubMed] [Google Scholar]
- 4.Momenimovahed Z, Salehiniya H. Incidence, mortality and risk factors of cervical cancer in the world. Biomed Res Ther. 2017;4:1795. [Google Scholar]
- 5.Cohen PA, Jhingran A, Oaknin A, Denny L. Cervical cancer. Lancet. 2019;393:169–82. doi: 10.1016/S0140-6736(18)32470-X. [DOI] [PubMed] [Google Scholar]
- 6.Benard VB, Johnson CJ, Thompson TD, Roland KB, Sue ML, Cokkinides V, et al. Examining the association between socioeconomic status and potential human papillomavirus-associated cancers. Cancer. 2008;113:2910–8. doi: 10.1002/cncr.23742. [DOI] [PubMed] [Google Scholar]
- 7.Ganesan S, Michael JC, Subbiah V. Associated factors with cervical pre-malignant lesions among the married fisher women community at Sadras, Tamil Nadu. Asia-Pac J Oncol Nurs. 2015;2:42–50. doi: 10.4103/2347-5625.146223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rossi PG, Baldacchini F, Ronco G. The possible effects on socio-economic inequalities of introducing HPV testing as primary test in cervical cancer screening programs. Front Oncol. 2014;4:20. doi: 10.3389/fonc.2014.00020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chidyaonga-Maseko F, Chirwa ML, Muula AS. Underutilization of cervical cancer prevention services in low and middle income countries: A review of contributing factors. Pan Afr Med J. 2015;21:231. doi: 10.11604/pamj.2015.21.231.6350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Leinonen MK, Campbell S, Klungsøyr O, Lönnberg S, Hansen BT, Nygård M. Personal and provider level factors influence participation to cervical cancer screening: A retrospective register-based study of 1.3 million women in Norway. Prev Med (Balt) 2017;94:31–9. doi: 10.1016/j.ypmed.2016.11.018. [DOI] [PubMed] [Google Scholar]
- 11.Walboomers JMM, Jacobs MV, Manos MM, Bosch FX, Kummer JA, Shah KV, et al. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J Pathol. 1999;189:12–9. doi: 10.1002/(SICI)1096-9896(199909)189:1<12::AID-PATH431>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 12.Bzhalava D, Eklund C, Dillner J. International standardization and classification of human papillomavirus types. Virology. 2015;476:341–4. doi: 10.1016/j.virol.2014.12.028. [DOI] [PubMed] [Google Scholar]
- 13.Chabeda A, Yanez RJR, Lamprecht R, Meyers AE, Rybicki EP, Hitzeroth II. Therapeutic vaccines for high-risk HPV-associated diseases. Papillomavirus Res. 2018;5:46–58. doi: 10.1016/j.pvr.2017.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Guan P, Howell-Jones R, Li N, Bruni L, De Sanjosé S, Franceschi S, et al. Human papillomavirus types in 115,789 HPV-positive women: a meta-analysis from cervical infection to cancer. Int J Cancer. 2012;131:2349–59. doi: 10.1002/ijc.27485. [DOI] [PubMed] [Google Scholar]
- 15.Wright TC. Natural history of HPV infections. J Fam Pract. 2009;58:S3–7. [PubMed] [Google Scholar]
- 16.Chan CK, Aimagambetova G, Ukybassova T, Kongrtay K, Azizan A. Human papillomavirus infection and cervical cancer: epidemiology, screening, and vaccination - review of current perspectives. J Oncol. 2019;2019:3257939. doi: 10.1155/2019/3257939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Stanley M. Pathology and epidemiology of HPV infection in females. Gynecol Oncol. 2010;117:S5–10. doi: 10.1016/j.ygyno.2010.01.024. [DOI] [PubMed] [Google Scholar]
- 18.Rachel Skinner S, Wheeler CM, Romanowski B, Castellsagué X, Lazcano-Ponce E, Rowena Del Rosario-Raymundo M, et al. Progression of HPV infection to detectable cervical lesions or clearance in adult women: analysis of the control arm of the VIVIANE study. Int J Cancer. 2016;138:2428–38. doi: 10.1002/ijc.29971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mirabello L, Clarke MA, Nelson CW, Dean M, Wentzensen N, Yeager M, et al. The intersection of HPV epidemiology, genomics and mechanistic studies of HPV-mediated carcinogenesis. Viruses. 2018;10:80. doi: 10.3390/v10020080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Georgescu SR, Mitran CI, Mitran MI, Caruntu C, Sarbu MI, Matei C, et al. New insights in the pathogenesis of HPV infection and the associated carcinogenic processes: The role of chronic inflammation and oxidative stress. J Immunol Res. 2018;2018:5315816. doi: 10.1155/2018/5315816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Insinga RP, Dasbach EJ, Elbasha EH. Epidemiologic natural history and clinical management of Human Papillomavirus (HPV) Disease: A critical and systematic review of the literature in the development of an HPV dynamic transmission model. BMC Infect Dis. 2009;9:119. doi: 10.1186/1471-2334-9-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Moscicki AB, Ma Y, Wibbelsman C, Darragh TM, Powers A, Farhat S, et al. Rate of and risks for regression of cervical intraepithelial neoplasia 2 in adolescents and young women. Obstet Gynecol. 2010;116:1373–80. doi: 10.1097/AOG.0b013e3181fe777f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Baldwin P, Laskey R, Coleman N. Translational approaches to improving cervical screening. Nat Rev Cancer. 2003;3:217–26. doi: 10.1038/nrc1010. [DOI] [PubMed] [Google Scholar]
- 24.Wang X, Huang X, Zhang Y. Involvement of human papillomaviruses in cervical cancer. Front Microbiol. 2018;9:2896. doi: 10.3389/fmicb.2018.02896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Day PM, Lowy DR, Schiller JT. Heparan sulfate-independent cell binding and infection with Furin-precleaved papillomavirus capsids. J Virol. 2008;82:12565–8. doi: 10.1128/JVI.01631-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Horvath CA, Boulet GA, Renoux VM, Delvenne PO, Bogers JPJ. Mechanisms of cell entry by human papillomaviruses: An overview. Virol J. 2010;7:11. doi: 10.1186/1743-422X-7-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Richards RM, Lowy DR, Schiller JT, Day PM. Cleavage of the papillomavirus minor capsid protein, L2, at a furin consensus site is necessary for infection. Proc Natl Acad Sci USA. 2006;103:1522–7. doi: 10.1073/pnas.0508815103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Day PM, Lowy DR, Schiller JT. Papillomaviruses infect cells via a clathrin-dependent pathway. Virology. 2003;307:1–11. doi: 10.1016/s0042-6822(02)00143-5. [DOI] [PubMed] [Google Scholar]
- 29.Smith JL, Campos SK, Ozbun MA. Human papillomavirus type 31 uses a caveolin 1- and dynamin 2-mediated entry pathway for infection of human keratinocytes. J Virol. 2007;81:9922–31. doi: 10.1128/JVI.00988-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Narisawa-Saito M, Kiyono T. Basic mechanisms of high-risk human papillomavirus-induced carcinogenesis: roles of E6 and E7 proteins. Cancer Sci. 2007;98:1505–11. doi: 10.1111/j.1349-7006.2007.00546.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cullen AP, Reid R, Campion M, Lörincz AT. Analysis of the physical state of different human papillomavirus DNAs in intraepithelial and invasive cervical neoplasm. J Virol. 1991;65:606–12. doi: 10.1128/jvi.65.2.606-612.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pirami L, Giachè V, Becciolini A. Analysis of HPV16, 18, 31, and 35 DNA in pre-invasive and invasive lesions of the uterine cervix. J Clin Pathol. 1997;50:600–4. doi: 10.1136/jcp.50.7.600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Badaracco G, Venuti A, Sedati A, Marcante ML. HPV16 and HPV18 in genital tumors: Significantly different levels of viral integration and correlation to tumor invasiveness. J Med Virol. 2002;67:574–82. doi: 10.1002/jmv.10141. [DOI] [PubMed] [Google Scholar]
- 34.Bodelon C, Untereiner ME, Machiela MJ, Vinokurova S, Wentzensen N. Genomic characterization of viral integration sites in HPV-related cancers. Int J Cancer. 2016;139:2001–11. doi: 10.1002/ijc.30243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hu Z, Zhu D, Wang W, Li W, Jia W, Zeng X, et al. Genome-wide profiling of HPV integration in cervical cancer identifies clustered genomic hot spots and a potential microhomology-mediated integration mechanism. Nat Genet. 2015;47:158–63. doi: 10.1038/ng.3178. [DOI] [PubMed] [Google Scholar]
- 36.Schmitz M, Driesch C, Jansen L, Runnebaum IB, Dürst M. Non-random integration of the HPV genome in cervical cancer. PLoS ONE. 2012;7:e39632. doi: 10.1371/journal.pone.0039632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zur Hausen H. Papillomaviruses and cancer: from basic studies to clinical application. Nat Rev Cancer. 2002;2:342–50. doi: 10.1038/nrc798. [DOI] [PubMed] [Google Scholar]
- 38.Doorbar J. Model systems of human papillomavirus-associated disease. J Pathol. 2016;238:166–79. doi: 10.1002/path.4656. [DOI] [PubMed] [Google Scholar]
- 39.Maufort JP, Shai A, Pitot HC, Lambert PF. A role for HPV16 E5 in cervical carcinogenesis. Cancer Res. 2010;70:2924–31. doi: 10.1158/0008-5472.CAN-09-3436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Müller M, Prescott EL, Wasson CW, MacDonald A. Human papillomavirus E5 oncoprotein: function and potential target for antiviral therapeutics. Future Virol. 2015;10:27–39. [Google Scholar]
- 41.Wetherill LF, Holmes KK, Verow M, Muller M, Howell G, Harris M, et al. High-risk human papillomavirus E5 oncoprotein displays channel-forming activity sensitive to small-molecule inhibitors. J Virol. 2012;86:5341–51. doi: 10.1128/JVI.06243-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Venuti A, Paolini F, Nasir L, Corteggio A, Roperto S, Campo MS, et al. Papillomavirus E5: the smallest oncoprotein with many functions. Mol Cancer. 2011;10:140. doi: 10.1186/1476-4598-10-140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Thomsen P, Van Deurs B, Norrild B, Kayser L. The HPV16 E5 oncogene inhibits endocytic trafficking. Oncogene. 2000;19:6023–32. doi: 10.1038/sj.onc.1204010. [DOI] [PubMed] [Google Scholar]
- 44.Zhang B, Srirangam A, Potter DA, Roman A. HPV16 E5 protein disrupts the c-Cbl-EGFR interaction and EGFR ubiquitination in human foreskin keratinocytes. Oncogene. 2005;24:2585–8. doi: 10.1038/sj.onc.1208453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gu Z, Matlashewski G. Effect of human papillomavirus type 16 oncogenes on MAP kinase activity. J Virol. 1995;69:8051–6. doi: 10.1128/jvi.69.12.8051-8056.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ashrafi GH, Haghshenas MR, Marchetti B, O’Brien PM, Campo MS. E5 protein of human papillomavirus type 16 selectively downregulates surface HLA class I. Int J Cancer. 2005;113:276–83. doi: 10.1002/ijc.20558. [DOI] [PubMed] [Google Scholar]
- 47.Campo MS, Graham SV, Cortese MS, Ashrafi GH, Araibi EH, Dornan ES, et al. HPV-16 E5 down-regulates expression of surface HLA class I and reduces recognition by CD8 T cells. Virology. 2010;407:137–42. doi: 10.1016/j.virol.2010.07.044. [DOI] [PubMed] [Google Scholar]
- 48.Kabsch K, Alonso A. The human papillomavirus type 16 E5 protein impairs TRAIL- and FasL-mediated apoptosis in HaCaT cells by different mechanisms. J Virol. 2002;76:12162–72. doi: 10.1128/JVI.76.23.12162-12172.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Howie HL, Katzenellenbogen RA, Galloway DA. Papillomavirus E6 proteins. Virology. 2009;384:324–34. doi: 10.1016/j.virol.2008.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Martinez-Zapien D, Ruiz FX, Poirson J, Mitschler A, Ramirez J, Forster A, et al. Structure of the E6/E6AP/p53 complex required for HPV-mediated degradation of p53. Nature. 2016;529:541–5. doi: 10.1038/nature16481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Filippova M, Johnson MM, Bautista M, Filippov V, Fodor N, Tungteakkhun SS, et al. The large and small isoforms of human papillomavirus type 16 E6 bind to and differentially affect procaspase 8 stability and activity. J Virol. 2007;81:4116–29. doi: 10.1128/JVI.01924-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Garnett TO, Filippova M, Duerksen-Hughes PJ. Accelerated degradation of FADD and procaspase 8 in cells expressing human papilloma virus 16 E6 impairs TRAIL-mediated apoptosis. Cell Death Differ. 2006;13:1915–26. doi: 10.1038/sj.cdd.4401886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Thomas M, Banks L. Human papillomavirus (HPV) E6 interactions with Bak are conserved amongst E6 proteins from high and low risk HPV types. J Gen Virol. 1999;80:1513–7. doi: 10.1099/0022-1317-80-6-1513. [DOI] [PubMed] [Google Scholar]
- 54.Filippova M, Parkhurst L, Duerksen-Hughes PJ. The human papillomavirus 16 E6 protein binds to Fas-associated death domain and protects cells from Fas-triggered apoptosis. J Biol Chem. 2004;279:25729–44. doi: 10.1074/jbc.M401172200. [DOI] [PubMed] [Google Scholar]
- 55.Filippova M, Song H, Connolly JL, Dermody TS, Duerksen-Hughes PJ. The human papillomavirus 16 E6 protein binds to tumor necrosis factor (TNF) R1 and protects cells from TNF-induced apoptosis. J Biol Chem. 2002;277:21730–9. doi: 10.1074/jbc.M200113200. [DOI] [PubMed] [Google Scholar]
- 56.Wang X, Meyers C, Guo M, Zheng ZM. Upregulation of p18Ink4c expression by oncogenic HPV E6 via p53-miR-34a pathway. Int J Cancer. 2011;129:1362–72. doi: 10.1002/ijc.25800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Zhu Y, Han Y, Tian T, Su P, Jin G, Chen J, et al. MiR-21-5p, miR-34a, and human telomerase RNA component as surrogate markers for cervical cancer progression. Pathol Res Pract. 2018;214:374–9. doi: 10.1016/j.prp.2018.01.001. [DOI] [PubMed] [Google Scholar]
- 58.Patel D, Huang SM, Baglia LA, McCance DJ. The E6 protein of human papillomavirus type 16 binds to and inhibits co-activation by CBP and p300. EMBO J. 1999;18:5061–72. doi: 10.1093/emboj/18.18.5061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Xie X, Piao L, Bullock BN, Smith A, Su T, Zhang M, et al. Targeting HPV16 E6-p300 interaction reactivates p53 and inhibits the tumorigenicity of HPV-positive head and neck squamous cell carcinoma. Oncogene. 2014;33:1037–46. doi: 10.1038/onc.2013.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Li S, Labrecque S, Gauzzi MC, Cuddihy AR, Wong AHT, Pellegrini S, et al. The human papilloma virus (HPV)-18 E6 oncoprotein physically associates with Tyk2 and impairs Jak-STAT activation by interferon-α. Oncogene. 1999;18:5727–37. doi: 10.1038/sj.onc.1202960. [DOI] [PubMed] [Google Scholar]
- 61.Shah M, Anwar MA, Park S, Jafri SS, Choi S. In silico mechanistic analysis of IRF3 inactivation and high-risk HPV E6 species-dependent drug response. Sci Rep. 2015;5:1–14. doi: 10.1038/srep13446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rincon-Orozco B, Halec G, Rosenberger S, Muschik D, Nindl I, Bachmann A, et al. Epigenetic silencing of interferon-κ in human papillomavirus type 16-positive cells. Cancer Res. 2009;69:8718–25. doi: 10.1158/0008-5472.CAN-09-0550. [DOI] [PubMed] [Google Scholar]
- 63.Gewin L, Myers H, Kiyono T, Galloway DA. Identification of a novel telomerase repressor that interacts with the human papillomavirus type-16 E6/E6-AP complex. Genes Dev. 2004;18:2269–82. doi: 10.1101/gad.1214704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Oh ST, Kyo S, Laimins LA. Telomerase activation by human papillomavirus type 16 E6 protein: induction of human telomerase reverse transcriptase expression through Myc and GC-Rich Sp1 binding sites. J Virol. 2001;75:5559–66. doi: 10.1128/JVI.75.12.5559-5566.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Hoppe-Seyler K, Bossler F, Braun JA, Herrmann AL, Hoppe-Seyler F. The HPV E6/E7 oncogenes: key factors for viral carcinogenesis and therapeutic targets. Trends Microbiol. 2018;26:158–68. doi: 10.1016/j.tim.2017.07.007. [DOI] [PubMed] [Google Scholar]
- 66.McLaughlin-Drubin ME, Münger K. The human papillomavirus E7 oncoprotein. Virology. 2009;384:335–44. doi: 10.1016/j.virol.2008.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Phelps WC, Yee CL, Münger K, Howley PM. The human papillomavirus type 16 E7 gene encodes transactivation and transformation functions similar to those of adenovirus E1A. Cell. 1988;53:539–47. doi: 10.1016/0092-8674(88)90570-3. [DOI] [PubMed] [Google Scholar]
- 68.Munger K, Werness BA, Dyson N, Phelps WC, Harlow E, Howley PM. Complex formation of c-myc papillomavirus E7 proteins with the retinoblastoma tumor suppressor gene product. EMBO J. 1989;8:4099–105. doi: 10.1002/j.1460-2075.1989.tb08594.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Hwang SG, Lee D, Kim J, Seo T, Choe J. Human papillomavirus type 16 E7 binds to E2F1 and activates E2F1-driven transcription in a retinoblastoma protein-independent manner. J Biol Chem. 2002;277:2923–30. doi: 10.1074/jbc.M109113200. [DOI] [PubMed] [Google Scholar]
- 70.Fischer M, Uxa S, Stanko C, Magin TM, Engeland K. Human papilloma virus E7 oncoprotein abrogates the p53-p21-DREAM pathway. Sci Rep. 2017;7:1–11. doi: 10.1038/s41598-017-02831-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Songock WK. Kim S man, Bodily JM. The human papillomavirus E7 oncoprotein as a regulator of transcription. Virus Res. 2017;231:56–75. doi: 10.1016/j.virusres.2016.10.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ivashkiv LB, Donlin LT. Regulation of type I interferon responses. Nat Rev Immunol. 2014;14:36–49. doi: 10.1038/nri3581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Toussaint-Smith E, Donner DB, Roman A. Expression of human papillomavirus type 16 E6 and E7 oncoproteins in primary foreskin keratinocytes is sufficient to alter the expression of angiogenic factors. Oncogene. 2004;23:2988–95. doi: 10.1038/sj.onc.1207442. [DOI] [PubMed] [Google Scholar]
- 74.Goradel NH, Ghiyami-Hour F, Jahangiri S, Negahdari B, Sahebkar A, Masoudifar A, et al. Nanoparticles as new tools for inhibition of cancer angiogenesis. J Cell Physiol. 2018;233:2902–10. doi: 10.1002/jcp.26029. [DOI] [PubMed] [Google Scholar]
- 75.Goradel NH, Asghari MH, Moloudizargari M, Negahdari B, Haghi-Aminjan H, Abdollahi M. Melatonin as an angiogenesis inhibitor to combat cancer: mechanistic evidence. Toxicol Appl Pharmacol. 2017;335:56–63. doi: 10.1016/j.taap.2017.09.022. [DOI] [PubMed] [Google Scholar]
- 76.Goradel NH, Mohammadi N, Haghi-Aminjan H, Farhood B, Negahdari B, Sahebkar A. Regulation of tumor angiogenesis by microRNAs: State of the art. J Cell Physiol. 2019;234:1099–110. doi: 10.1002/jcp.27051. [DOI] [PubMed] [Google Scholar]
- 77.Gupta G, Glueck R, Patel PR. HPV vaccines: Global perspectives. Hum Vaccin Immunother. 2017;13:1421–4. doi: 10.1080/21645515.2017.1289301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Vujadinovic M, Khan S, Oosterhuis K, Uil TG, Wunderlich K, Damman S, et al. Adenovirus based HPV L2 vaccine induces broad cross-reactive humoral immune responses. Vaccine. 2018;36:4462–70. doi: 10.1016/j.vaccine.2018.06.024. [DOI] [PubMed] [Google Scholar]
- 79.Wu WH, Alkutkar T, Karanam B, Roden RB, Ketner G, Ibeanu OA. Capsid display of a conserved human papillomavirus L2 peptide in the adenovirus 5 hexon protein: A candidate prophylactic HPV vaccine approach. Virol J. 2015;12:140. doi: 10.1186/s12985-015-0364-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Arbyn M, Xu L, Simoens C, Martin-Hirsch PPL. Prophylactic vaccination against human papillomaviruses to prevent cervical cancer and its precursors. Cochrane Database Syst Rev. 2018;2018:CD009069. doi: 10.1002/14651858.CD009069.pub3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Cordeiro MN, De Lima R de CP, Paolini F, Melo AR da S, Campos APF, Venuti A, et al. Current research into novel therapeutic vaccines against cervical cancer. Expert Rev Anticancer Ther. 2018;18:365–76. doi: 10.1080/14737140.2018.1445527. [DOI] [PubMed] [Google Scholar]
- 82.Yang A, Farmer E, Wu TC, Hung CF. Perspectives for therapeutic HPV vaccine development. J Biomed Sci. 2016;23:1–19. doi: 10.1186/s12929-016-0293-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Liu T-Y, Hussein WM, Toth I, Skwarczynski M. Advances in peptide-based human papillomavirus therapeutic vaccines. Curr Top Med Chem. 2012;12:1581–92. doi: 10.2174/156802612802652402. [DOI] [PubMed] [Google Scholar]
- 84.Su JH, Wu A, Scotney E, Ma B, Monie A, Hung CF, et al. Immunotherapy for cervical cancer: research status and clinical potential. BioDrugs. 2010;24:109–29. doi: 10.2165/11532810-000000000-00000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Cheng MA, Farmer E, Huang C, Lin J, Hung CF, Wu TC, Therapeutic DNA. Vaccines for human papillomavirus and associated diseases. Hum Gene Ther. 2018;29:971–96. doi: 10.1089/hum.2017.197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Benencia F, Courrèges MC, Coukos G. Whole tumor antigen vaccination using dendritic cells: comparison of RNA electroporation and pulsing with UV-irradiated tumor cells. J Transl Med. 2008;6:21. doi: 10.1186/1479-5876-6-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kim JH, Kang TH, Noh KH, Bae HC, Kim SH, Yoo YD, et al. Enhancement of dendritic cell-based vaccine potency by anti-apoptotic siRNAs targeting key pro-apoptotic proteins in cytotoxic CD8+ T cell-mediated cell death. Immunol Lett. 2009;122:58–67. doi: 10.1016/j.imlet.2008.12.006. [DOI] [PubMed] [Google Scholar]
- 88.Peng S, Kim TW, Lee JH, Yang MU, Liangmei HE, Hung CF, et al. Vaccination with dendritic cells transfected with BAK and BAX siRNA enhances antigen-specific immune responses by prolonging dendritic cell life. Hum Gene Ther. 2005;16:584–93. doi: 10.1089/hum.2005.16.584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Santin A, Bellone S, Roman J, Burnett A, Cannon M, Pecorelli S. Therapeutic vaccines for cervical cancer: dendritic cell-based immunotherapy. Curr Pharm Des. 2005;11:3485–500. doi: 10.2174/138161205774414565. [DOI] [PubMed] [Google Scholar]
- 90.Chang EY, Chen CH, Ji H, Wang TL, Hung K, Lee BP, et al. Antigen-specific cancer immunotherapy using a GM-CSF secreting allogeneic tumor cell-based vaccine. Int J Cancer. 2000;86:725–30. doi: 10.1002/(sici)1097-0215(20000601)86:5<725::aid-ijc19>3.0.co;2-k. [DOI] [PubMed] [Google Scholar]
- 91.Mikysková R, Indrová M, Símová J, Jandlová T, Bieblová J, Jinoch P, et al. Treatment of minimal residual disease after surgery or chemotherapy in mice carrying HPV16-associated tumours: cytokine and gene therapy with IL-2 and GM-CSF. Int J Oncol. 2004;24:161–7. [PubMed] [Google Scholar]
- 92.Yang A, Jeang J, Cheng K, Cheng T, Yang B, Wu TC, et al. Current state in the development of candidate therapeutic HPV vaccines. Expert Rev Vaccines. 2016;15:989–1007. doi: 10.1586/14760584.2016.1157477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Lin K, Roosinovich E, Ma B, Hung CF, Wu TC. Therapeutic HPV DNA vaccines. Immunol Res. 2010;47:86–112. doi: 10.1007/s12026-009-8141-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Miles B, Safran HP, Monk BJ. Therapeutic options for treatment of human papillomavirus-associated cancers - novel immunologic vaccines: ADXS11–001. Gynecol Oncol Res Pract. 2017;4:10. doi: 10.1186/s40661-017-0047-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Reddy VS, Nemerow GR. Structures and organization of adenovirus cement proteins provide insights into the role of capsid maturation in virus entry and infection. Proc Natl Acad Sci USA. 2014;111:11715–20. doi: 10.1073/pnas.1408462111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Abudoureyimu M, Lai Y, Tian C, Wang T, Wang R, Chu X. Oncolytic adenovirus—a nova for gene-targeted oncolytic viral therapy in HCC. Front Oncol. 2019;9:1182. doi: 10.3389/fonc.2019.01182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Dai X, Wu L, Sun R, Zhou ZH. Atomic structures of minor proteins VI and VII in human adenovirus. J Virol. 2017;91:e00850–17. doi: 10.1128/JVI.00850-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Nattress CB, Halldén G. Advances in oncolytic adenovirus therapy for pancreatic cancer. Cancer Lett. 2018;434:56–69. doi: 10.1016/j.canlet.2018.07.006. [DOI] [PubMed] [Google Scholar]
- 99.Goradel NH, Mohajel N, Malekshahi ZV, Jahangiri S, Najafi M, Farhood B, et al. Oncolytic adenovirus: a tool for cancer therapy in combination with other therapeutic approaches. J Cell Physiol. 2019;234:8636–46. doi: 10.1002/jcp.27850. [DOI] [PubMed] [Google Scholar]
- 100.Davison AJ, Benko M, Harrach B. Genetic content and evolution of adenoviruses. J Gen Virol. 2003;84:2895–908. doi: 10.1099/vir.0.19497-0. [DOI] [PubMed] [Google Scholar]
- 101.Vannucci L, Lai M, Chiuppesi F, Ceccherini-Nelli L, Pistello M. Viral vectors: a look back and ahead on gene transfer technology. N Microbiol. 2013;36:1–22. [PubMed] [Google Scholar]
- 102.Goradel NH, Baker AT, Arashkia A, Ebrahimi N, Ghorghanlu S, Negahdari B. Oncolytic virotherapy: challenges and solutions. Curr Probl Cancer. 2021;45:100639. doi: 10.1016/j.currproblcancer.2020.100639. [DOI] [PubMed] [Google Scholar]
- 103.O’Riordan CR, Lachapelle A, Delgado C, Parkes V, Wadsworth SC, Smith AE, et al. PEGylation of adenovirus with retention of infectivity and protection from neutralizing antibody in vitro and in vivo. Hum Gene Ther. 1999;10:1349–58. doi: 10.1089/10430349950018021. [DOI] [PubMed] [Google Scholar]
- 104.Choi IK, Yun CO. Recent developments in oncolytic adenovirus-based immunotherapeutic agents for use against metastatic cancers. Cancer Gene Ther. 2013;20:70–6. doi: 10.1038/cgt.2012.95. [DOI] [PubMed] [Google Scholar]
- 105.Crystal RG. Adenovirus: the first effective in vivo gene delivery vector. Hum Gene Ther. 2014;25:3–11. doi: 10.1089/hum.2013.2527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Wold W, Toth K. Adenovirus vectors for gene therapy, vaccination and cancer gene therapy. Curr Gene Ther. 2014;13:421–33. doi: 10.2174/1566523213666131125095046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Vorburger SA, Hunt KK. Adenoviral gene therapy. Oncologist. 2002;7:46–59. doi: 10.1634/theoncologist.7-1-46. [DOI] [PubMed] [Google Scholar]
- 108.Brunetti-Pierri N, Ng P. Helper-dependent adenoviral vectors. In: Adenoviral vectors for gene therapy: second edition. Elsevier Inc.; 2016. p. 423–450. 10.1016/B978-0-12-800276-6.00017-6.
- 109.Zhang WW, Li L, Li D, Liu J, Li X, Li W, et al. The first approved gene therapy product for cancer Ad-p53 (Gendicine): 12 years in the clinic. Hum Gene Ther. 2018;29:160–79. doi: 10.1089/hum.2017.218. [DOI] [PubMed] [Google Scholar]
- 110.Liang M. Oncorine, the world first oncolytic virus medicine and its update in China. Curr Cancer Drug Targets. 2018;18:171–6. doi: 10.2174/1568009618666171129221503. [DOI] [PubMed] [Google Scholar]
- 111.Pesonen S, Kangasniemi L, Hemminki A. Oncolytic adenoviruses for the treatment of human cancer: focus on translational and clinical data. Mol Pharmacol. 2011;8:12–28. doi: 10.1021/mp100219n. [DOI] [PubMed] [Google Scholar]
- 112.Bauerschmitz GJ, Kanerva A, Wang M, Herrmann I, Shaw DR, Strong TV, et al. Evaluation of a selectively oncolytic adenovirus for local and systemic treatment of cervical cancer. Int J Cancer. 2004;111:303–9. doi: 10.1002/ijc.20217. [DOI] [PubMed] [Google Scholar]
- 113.Heideman DAM, Steenbergen RDM, van der Torre J, Scheffner M, Alemany R, Gerritsen WR, et al. Oncolytic adenovirus expressing a p53 variant resistant to degradation by HPV E6 protein exhibits potent and selective replication in cervical cancer. Mol Ther. 2005;12:1083–90. doi: 10.1016/j.ymthe.2005.06.443. [DOI] [PubMed] [Google Scholar]
- 114.Yoon AR, Kim JH, Lee YS, Kim H, Yoo JY, Sohn JH, et al. Markedly enhanced cytolysis by E1B-19kD-deleted oncolytic adenovirus in combination with cisplatin. Hum Gene Ther. 2006;17:379–90. doi: 10.1089/hum.2006.17.379. [DOI] [PubMed] [Google Scholar]
- 115.Mi J, Li ZY, Ni S, Steinwaerder D, Lieber A. Induced apoptosis supports spread of adenovirus vectors in tumors. Hum Gene Ther. 2001;12:1343–52. doi: 10.1089/104303401750270995. [DOI] [PubMed] [Google Scholar]
- 116.Wang W, Xia X, Wang S, Sima N, Li Y, Han Z, et al. Oncolytic adenovirus armed with human papillomavirus E2 gene in combination with radiation demonstrates synergistic enhancements of antitumor efficacy. Cancer Gene Ther. 2011;18:825–36. doi: 10.1038/cgt.2011.53. [DOI] [PubMed] [Google Scholar]
- 117.Chu RL, Post DE, Khuri FR, Van Meir EG. Use of replicating oncolytic adenoviruses in combination therapy for cancer. Clin Cancer Res. 2004;10:5299–312. doi: 10.1158/1078-0432.CCR-0349-03. [DOI] [PubMed] [Google Scholar]
- 118.Wang W, Sima N, Kong D, Luo A, Gao Q, Liao S, et al. Selective targeting of HPV-16 E6/E7 in cervical cancer cells with a potent oncolytic adenovirus and its enhanced effect with radiotherapy in vitro and vivo. Cancer Lett. 2010;291:67–75. doi: 10.1016/j.canlet.2009.09.022. [DOI] [PubMed] [Google Scholar]
- 119.zhong XJ, lan ZJ, Guo ZW. Antisense RNA: the new favorite in genetic research. J Zhejiang Univ Sci B. 2018;19:739–49. doi: 10.1631/jzus.B1700594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Hamada K, Shirakawa T, Gotoh A, Roth JA, Follen M. Adenovirus-mediated transfer of human papillomavirus 16 E6/E7 antisense RNA and induction of apoptosis in cervical cancer. Gynecol Oncol. 2006;103:820–30. doi: 10.1016/j.ygyno.2006.06.035. [DOI] [PubMed] [Google Scholar]
- 121.Bonetta AC, Mailly L, Robinet E, Travé G, Masson M, Deryckere F. Artificial microRNAs against the viral E6 protein provoke apoptosis in HPV positive cancer cells. Biochem Biophys Res Commun. 2015;465:658–64. doi: 10.1016/j.bbrc.2015.07.144. [DOI] [PubMed] [Google Scholar]
- 122.Ramirez J, Poirson J, Foltz C, Chebaro Y, Schrapp M, Meyer A, et al. Targeting the two oncogenic functional sites of the HPV E6 oncoprotein with a high-affinity bivalent ligand. Angew Chem Int Ed Engl. 2015;54:7958–62. doi: 10.1002/anie.201502646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Çuburu N, Khan S, Thompson CD, Kim R, Vellinga J, Zahn R, et al. Adenovirus vector-based prime-boost vaccination via heterologous routes induces cervicovaginal CD8+ T cell responses against HPV16 oncoproteins. Int J Cancer. 2018;142:1467–79. doi: 10.1002/ijc.31166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Roberts JN, Buck CB, Thompson CD, Kines R, Bernardo M, Choyke PL, et al. Genital transmission of HPV in a mouse model is potentiated by nonoxynol-9 and inhibited by carrageenan. Nat Med. 2007;13:857–61. doi: 10.1038/nm1598. [DOI] [PubMed] [Google Scholar]
- 125.Walters RW, Freimuth P, Moninger TO, Ganske I, Zabner J, Welsh MJ. Adenovirus fiber disrupts CAR-mediated intercellular adhesion allowing virus escape. Cell. 2002;110:789–99. doi: 10.1016/s0092-8674(02)00912-1. [DOI] [PubMed] [Google Scholar]
- 126.Wu J, Chen G, Zhuang FC, Gao M, Wu CD, He ZL, et al. Long-term toxicity, pharmacokinetics and immune effects of a recombinant adenovirus vaccine expressing human papillomavirus 16 E6 and E7 proteins (HPV16 E6E7-Ad5 Vac) in primates. Am J Transl Res. 2018;10:1539–51. [PMC free article] [PubMed] [Google Scholar]
- 127.Jin HS, Park EK, Lee JM, Namkoong SE, Kim DG, Lee YJ, et al. Immunization with adenoviral vectors carrying recombinant IL-12 and E7 enhanced the antitumor immunity to human papillomavirus 16-associated tumor. Gynecol Oncol. 2005;97:559–67. doi: 10.1016/j.ygyno.2005.01.046. [DOI] [PubMed] [Google Scholar]
- 128.Ahn WS, Bae SM, Kim TY, Kim TG, Lee JM, Namkoong SE, et al. A therapy modality using recombinant IL-12 adenovirus plus E7 protein in a human papillomavirus 16 E6/E7-associated cervical cancer animal model. Hum Gene Ther. 2003;14:1389–99. doi: 10.1089/104303403769211619. [DOI] [PubMed] [Google Scholar]
- 129.Germann T, Gately MK, Schoenhaut DS, Lohoff M, Mattner F, Fischer S, et al. Interleukin‐12/T cell stimulating factor, a cytokine with multiple effects on T helper type 1 (Th1) but not on Th2 cells. Eur J Immunol. 1993;23:1762–70. doi: 10.1002/eji.1830230805. [DOI] [PubMed] [Google Scholar]
- 130.Kobayashi M, Fitz L, Ryan M, Hewick RM, Clark SC, Chan S, et al. Identification and purification of natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologic effects on human lymphocytes. J Exp Med. 1989;170:827–45. doi: 10.1084/jem.170.3.827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Gomez-Gutierrez JG, Elpek KG, Montes De Oca-Luna R, Shirwan H, Sam Zhou H, McMasters KM. Vaccination with an adenoviral vector expressing calreticulin-human papillomavirus 16 E7 fusion protein eradicates E7 expressing established tumors in mice. Cancer Immunol Immunother. 2007;56:997–1007. doi: 10.1007/s00262-006-0247-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Basu S, Srivastava PK. Calreticulin, a peptide-binding chaperone of the endoplasmic reticulum, elicits tumor- and peptide-specific immunity. J Exp Med. 1999;189:797–802. doi: 10.1084/jem.189.5.797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Sadasivan B, Lehner PJ, Ortmann B, Spies T, Cresswell P. Roles for calreticulin and a novel glycoprotein, tapasin, in the interaction of MHC class I molecules with TAP. Immunity. 1996;5:103–14. doi: 10.1016/s1074-7613(00)80487-2. [DOI] [PubMed] [Google Scholar]
- 134.Tillman BW, Hayes TL, DeGruijl TD, Douglas JT, Curiel DT. Adenoviral vectors targeted to CD40 enhance the efficacy of dendritic cell-based vaccination against human papillomavirus 16-induced tumor cells in a murine model. Cancer Res. 2000;60:5456–63. [PubMed] [Google Scholar]
- 135.Liu D-W, Tsao Y-P, Hsieh C-H, Hsieh J-T, Kung JT, Chiang C-L, et al. Induction of CD8 T cells by vaccination with recombinant adenovirus expressing human papillomavirus type 16 E5 gene reduces tumor growth. J Virol. 2000;74:9083–9. doi: 10.1128/jvi.74.19.9083-9089.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Brandsma JL, Shlyankevich M, Zhang L, Slade MD, Goodwin EC, Peh W, et al. Vaccination of rabbits with an adenovirus vector expressing the papillomavirus E2 protein leads to clearance of papillomas and infection. J Virol. 2004;78:116–23. doi: 10.1128/JVI.78.1.116-123.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Ahn WS, Han YJ, Bae SM, Kim TH, Rho MS, Lee JM, et al. Differential suppression of human cervical cancer cell growth by adenovirus delivery of p53 in vitro: arrest phase of cell cycle is dependent on cell line. Jpn J Cancer Res. 2002;93:1012–9. doi: 10.1111/j.1349-7006.2002.tb02478.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Hamada K, Alemany R, Zhang WW, Hittelman WN, Lotan R, Roth JA, et al. Adenovirus-mediated transfer of a wild-type p53 gene and induction of apoptosis in cervical cancer. Cancer Res. 1996;56:3047–54. [PubMed] [Google Scholar]
- 139.Hamada K, Zhang WW, Alemany R, Wolf J, Roth JA, Mitchell MF. Growth inhibition of human cervical cancer cells with the recombinant adenovirus p53 in vitro. Gynecol Oncol. 1996;60:373–9. doi: 10.1006/gyno.1996.0057. [DOI] [PubMed] [Google Scholar]
- 140.Liu YL, Zheng XL, Liu FM. The mechanism and inhibitory effect of recombinant human P53 adenovirus injection combined with paclitaxel on human cervical cancer cell HeLa. Eur Rev Med Pharmacol Sci. 2015;19:1037–42. [PubMed] [Google Scholar]
- 141.Xiao J, Zhou J, Fu M, Liang LI, Deng Q, Liu X, et al. Efficacy of recombinant human adenovirus-p53 combined with chemotherapy for locally advanced cervical cancer: a clinical trial. Oncol Lett. 2017;13:3676–80. doi: 10.3892/ol.2017.5901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Su X, Chen WJ, Xiao SW, Li XF, Xu G, Pan JJ, et al. Effect and safety of recombinant adenovirus-p53 transfer combined with radiotherapy on long-term survival of locally advanced cervical cancer. Hum Gene Ther. 2016;27:1008–14. doi: 10.1089/hum.2016.043. [DOI] [PubMed] [Google Scholar]
- 143.Prabhu NS, Blagosklonny MV, Zeng YX, Wu GS, Waldman T, El-Deiry WS. Suppression of cancer cell growth by adenovirus expressing p21(WAF1/CIP1) deficient in PCNA interaction. Clin Cancer Res. 1996;2:1221–9. [PubMed] [Google Scholar]
- 144.Jost CA, Marin MC, Kaelin WG. P73 is a human p53-related protein that can induce apoptosis. Nature. 1997;389:191–4. doi: 10.1038/38298. [DOI] [PubMed] [Google Scholar]
- 145.Kaghad M, Bonnet H, Yang A, Creancier L, Biscan JC, Valent A, et al. Monoallelically expressed gene related to p53 at 1p36, a region frequently deleted in neuroblastoma and other human cancers. Cell. 1997;90:809–19. doi: 10.1016/s0092-8674(00)80540-1. [DOI] [PubMed] [Google Scholar]
- 146.Prabhu N, Somasundaram K, Satyamoorthy K, Herlyn M, El-Deiry WS. p73ß, unlike p53, suppresses growth and induces apoptosis of human papillomavirus E6-expressing cancer cells. Int J Oncol. 1998;13:5–9. doi: 10.3892/ijo.13.1.5. [DOI] [PubMed] [Google Scholar]
- 147.Das S, El-Deiry WS, Somasundaram K. Efficient growth inhibition of HPV 16 E6-expressing cells by an adenovirus-expressing p53 homologue p73β. Oncogene. 2003;22:8394–402. doi: 10.1038/sj.onc.1206908. [DOI] [PubMed] [Google Scholar]
- 148.Bourgo RJ, Braden WA, Wells SI, Knudsen ES. Activation of the retinoblastoma tumor suppressor mediates cell cycle inhibition and cell death in specific cervical cancer cell lines. Mol Carcinog. 2009;48:45–55. doi: 10.1002/mc.20456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Ip SM, Huang TG, Yeung WSB, Ngan HYS. pRb-expressing adenovirus Ad5-Rb attenuates the p53-induced apoptosis in cervical cancer cell lines. Eur J Cancer. 2001;37:2475–83. doi: 10.1016/s0959-8049(01)00308-2. [DOI] [PubMed] [Google Scholar]