

ABSTRACT
Glucagon-like peptide 1 (GLP-1) is an incretin hormone produced in the intestine that is secreted in response to nutrient exposure. GLP-1 potentiates glucose-dependent insulin secretion from the pancreatic β cells and promotes satiety. These important actions on glucose metabolism and appetite have led to widespread interest in GLP-1 receptor agonism. Typically, this involves pharmacological GLP-1 mimetics or targeted inhibition of dipeptidyl peptidase-IV, the enzyme responsible for GLP-1 degradation. However, nutritional strategies provide a widely available, cost-effective alternative to pharmacological strategies for enhancing hormone release. Recent advances in nutritional research have implicated the combined ingestion of protein and calcium with enhanced endogenous GLP-1 release, which is likely due to activation of receptors with high affinity and/or sensitivity for amino acids and calcium. Specifically targeting these receptors could enhance gut hormone secretion, thus providing a new therapeutic option. This narrative review provides an overview of the latest research on protein- and calcium-mediated GLP-1 release with an emphasis on human data, and a perspective on potential mechanisms that link potent GLP-1 release to the co-ingestion of protein and calcium. In light of these recent findings, potential future research directions are also presented.
Keywords: protein, calcium, GLP-1, extracellular calcium-sensing receptor, metabolic regulation, type 2 diabetes
Statement of Significance: Emerging evidence suggests the co-ingestion of protein and calcium can potently stimulate GLP-1 release.
Introduction
The obesity and type 2 diabetes (T2D) epidemics are major health concerns, whereby 13% and ∼6.3% of the global adult population (18 y or over) had obesity (1) and T2D (2) in 2016 and 2014, respectively. Furthermore, T2D is predicted to cost the UK National Health Service ∼£17 billion per year by 2035 (3), and obesity already costs >$2 trillion per year, globally (4) The current most effective long-term approach to treating obesity is bariatric surgery (5). Roux-en-Y gastric bypass surgery (RYGB) produces ∼63% excess body weight loss (difference between initial and final BMI) after 2 y (6), and T2D remission rates of >40% at 12 mo and at the time of latest follow-up (median, 23 mo) (7). Despite this, surgery is unlikely to be economically viable at the population level due to its cost of ≤£61,000 per quality-adjusted life year (8). Nevertheless, by establishing the mechanisms by which surgery can enhance weight loss and remission from diabetes, potential surgery “mimetics” can be explored to harness the weight-loss benefits of surgery, without the costs or the need for invasive procedures. A reported key mechanism for the surgery-induced weight loss and diabetes remission is greater nutrient delivery to intestinal epithelial L-cells and the associated enhancement of gut hormone availability (9–12). This suggests that targeting gut hormones released from intestinal epithelial cells holds promise as an obesity/T2D therapy.
Within the intestinal epithelium, enteroendocrine cells represent <1% of all gastrointestinal (GI) cells, and are a key component of the gut-brain-pancreas axis (13, 14). These enteroendocrine cells are categorized into a number of classes depending on their location along the GI tract, receptor expression, and secretory profile (15). Glucagon-like peptide 1 (GLP-1) is a peptide hormone produced and secreted by intestinal L-cells, which are located along the length of the GI epithelium, starting at the jejunum and then increasing in density along the small and large intestine down to the colon (10). GLP-1 has received a lot of attention because it is classified as an incretin hormone (hormones that stimulate glucose-dependent insulin secretion from the pancreas) along with glucose-dependent insulinotropic polypeptide (GIP). Additionally, GLP-1 also regulates appetite in a similar manner to hormones such as peptide tyrosine-tyrosine (PYY) and leptin, to promote fullness and reduce energy intake (16, 17). These collective actions have led to widespread interest in GLP-1 as a potential target for obesity and diabetic management and therapy.
Subsequently, agonists for the GLP-1 receptor have been a popular therapeutic option. Clinical trials that have involved GLP-1 receptor agonists such as exenatide and liraglutide have led to weight loss of ≤3 kg, and improvements in glycated hemoglobin of 2%, over 15–30 wk (18). More recently, 2.4 mg subcutaneous semaglutide administered once weekly for 68 wk (alongside a lifestyle intervention) resulted in 14.9% weight loss from baseline in obese individuals (19). Despite this, using a nutritional intervention to target L-cells and endogenous gut hormone release is still relevant as an alternative or combined treatment, due to a number of possible advantages over GLP-1 agonists. Firstly, nutritional intervention minimizes the potential adverse side effects, including GI discomfort and nausea, sometimes observed when using agonists (20, 21). In a review of the associated adverse side effects, the incidence of nausea, vomiting, and diarrhea was reasonably common (1 in 10) with ≤50% of patients affected by nausea (22), resulting in discontinuation rates of ≤10% in clinical practice (23). Secondly the vagal/neural dependency of part of GLP-1 action might not be accessible for GLP-1 enhancers/analogs (24, 25). For example, pharmacological GLP-1 administration may reduce appetite primarily via direct interaction with the brain (26), whereas endogenous GLP-1 effects on appetite might be exerted locally via receptors on the vagus nerve, and indirectly through gastric emptying (27, 28). Theoretically, continuous activation of the GLP-1 receptor via drug agonists could lead to receptor downregulation, and the natural rise and fall of nutrient-mediated GLP-1 secretion could be important for sustaining receptor sensitivity. Lastly, nutrient delivery might lead to increased secretion of colocalizing hormones including PYY, GIP, and oxyntomodulin (29) providing an additive effect on appetite suppression and blood glucose regulation.
The release of the incretin hormones, GLP-1 and GIP, and other gut hormones, including PYY and oxyntomodulin, is dependent on nutrient delivery to different enteroendocrine cells within the intestinal epithelium. Intestinal L-cells express several receptors and transporters selective to different macro and micronutrients (30). Because of this, whereas the extent of GLP-1 secretion is principally mediated by meal size (in order to reach the distal gut), the nutrient composition of a meal might also be influential for maximizing L-cell receptor activation and transporter utilization. Previously, carbohydrate, fat, and protein ingestion, as well as SCFAs produced by the gut microbiota, have been found to independently stimulate GLP-1 release (31–33). Emerging evidence suggests that the synergy of protein and calcium could also act as a potent secretagogue of GLP-1 (34, 35). In the most recent human study (35), this synergy produced some of the highest reported concentrations of GLP-1 following physiological ingestion to date. This is particularly interesting given that calcium does not provide energy, and therefore GLP-1 release is relatively greater despite no additional energy provision.
Previous reviews regarding the nutritional regulation of GLP-1 have discussed the specific molecular mechanisms underpinning fat-, carbohydrate-, and protein-mediated GLP-1 release (10, 36, 37). To our knowledge, only 1 review has addressed the effects of different nutrients on GLP-1 secretion (15); however, the effect of protein ingestion in humans was not discussed, as well as particular reference to protein forms and interaction with calcium, which is crucial for practical application. Therefore, this narrative review focuses on the incretin hormone GLP-1, and the research surrounding protein- and/or calcium-mediated GLP-1 secretion in humans. The potential mechanisms by which the combination of protein and calcium might enhance GLP-1 secretion are also discussed, alongside potential avenues for future research.
Secretion and Metabolism of GLP-1
GLP-1 is a peptide hormone produced by intestinal L-cells by differential processing of the proglucagon (GCG) gene (38). GLP-1 exists in 2 equipotent molecular active forms, GLP-17–37 and GLP-17–36amide, with the later representing ∼100% of circulating GLP-1 in humans (39, 40). Following the release of active GLP-1, only ∼10–15% enters the systemic circulation due to rapid degradation by dipeptidyl peptidase-IV (DPP4) in the splanchnic bed (38). Further degradation is likely to occur in the circulation by soluble DPP4, including DPP4 released from adipose tissue (41, 42). DPP4 cleaves 2 N-terminal amino acids of GLP-17–37 and GLP-17–36amide to produce GLP-19–37 and GLP-19–36amide, respectively, which act as low-affinity ligands for the GLP-1 receptor (43). Additionally, both the active and degraded forms of GLP-1 are cleared from the circulation via the kidneys (44). For the purpose of this review GLP-17–36amide will refer to the measurement of the receptor-active form of GLP-1, and GLP-1TOTAL will incorporate GLP-17–36amide as well as the downstream metabolite, GLP-19–36amide. Although plasma concentrations of GLP-17–36amide are the most convenient to sample, given such rapid degradation by DPP4, GLP-17–36amide concentrations recorded within lymph might be more sensitive to meal-induced GLP-1 secretion (45, 46). Lymphatic sampling is, however, currently challenging for human studies, and therefore plasma GLP-1 concentrations will be the main focus of this review.
GLP-1 secretion is regulated through meal intake, whereby GLP-1TOTAL plasma concentrations are very low in the fasted state (5–15 pmol/L) and rise following the ingestion of food by 2- to 4-fold (15, 38, 47). GLP-1 release has been demonstrated to fit either a monophasic (48, 49) or biphasic pattern (50, 51), reflecting a gradual rise to peak, or an initial rapid rise ∼10–15 min postmeal, followed by a secondary steady rise to peak, respectively. Peak GLP-1 concentrations occur ∼45–60 min after a meal, before a gradual decrease until the next prandial episode (38, 50). The early GLP-1 response was thought to occur via neural activation involving the vagus nerve (31, 52) and/or direct stimulation of the proximal jejunum L-cells, with the majority of GLP-1 stimulation occurring through direct interaction with distal L-cells (38). However, more recently it has been recognized that the density of L-cells in the duodenum is enough to account for at least part, if not most, of the early response (53).
GLP-1 has several known actions including glucose-dependent stimulation of insulin release, and inhibition of glucagon secretion, gastric emptying, and food intake (38, 54, 55). These actions are coordinated via the GLP-1 receptor, a G-protein–coupled receptor widely expressed in the brain, heart, pancreatic islets, and the GI tract (38, 56). Despite these actions contributing to improved metabolic control, GLP-1 has a half-life of only ∼2 min (10). This suggests that its effects are not solely mediated through direct activation of local GLP-1 receptors but largely through paracrine receptor activation on vagal afferent nerve fibers innervating target organs (27, 38, 57). Consequently, nutritional interventions targeting enhanced and prolonged GLP-1 release are of high interest.
Current Status of Knowledge
Protein-mediated GLP-1 release
Protein is considered the most satiating of the macronutrients (58, 59), and is often associated with weight loss (60), which could be mediated in part by protein-induced stimulation of appetite regulatory hormones, including GLP-1. Traditionally, fat and carbohydrate were thought to be the most potent stimulants of GLP-1 release (61), but this has subsequently been challenged by research comparing protein ingestion with carbohydrate and fat in humans.
Direct comparison between isocaloric macronutrients
A limited number of studies have compared GLP-1 release following ingestion of calorie- and/or volume-matched macronutrients. A comparison of the effect of high-protein compared with high-fat meals on GLP-1 release was studied in 12 healthy males who ingested milk and egg protein at 2 g/kg on one occasion and 0.88 g/kg oleic acid on the other (volume- and calorie-matched) (62). Following both meals, GLP-17–36amide and GLP-1TOTAL increased proportionally but did not differ in magnitude between the 2 conditions. Another study compared GLP-1 release following the ingestion of whey protein with that of maltodextrin (45 g) in 18 healthy weight women (BMI 19–25 kg/m2) (63). GLP-17–36amide incremental area under the curve (iAUC) was not significantly different between conditions; however, there was a trend for greater GLP-17–36amide iAUC following the whey protein test meal. Furthermore, the acute effects of 3 isocaloric (375 kcal) test meals, high in carbohydrate (100 g glucose), fat (84 mL double cream), or protein (352 g grilled turkey), resulted in similar peak GLP-17–36amide, although different times to peak (31). This might be explained by the different forms in which the 3 macronutrients were ingested, and the effect this could have on gastric emptying and subsequent delivery to the intestine. Although it is challenging to completely isolate one macronutrient from another, these studies provide some rationale to suggest that the ingestion of specific macronutrients largely results in comparable GLP-1 release. However, the measurement of GLP-1TOTAL would have provided greater sensitivity to detect changes in GLP-1 secretion between conditions in some of these studies.
Isocaloric meals high in protein, fat, or carbohydrate
A more applied approach would incorporate the interactions of different macronutrients, common to the typical diet. Ten healthy normal-weight and 10 overweight males were fed isocaloric pasta and dessert meals high in fat (65%), carbohydrate (66%), or protein (65%), with the remaining energy requirement shared equally between the other 2 macronutrients (∼66%:17%:17%). Satiety was greatest following the high-protein meal, despite no differences in GLP-17–36amide concentrations following each meal (64). Instead, this might be explained by the greater increase in PYY3-36 (the active form of a cosecreted L-cell–derived hormone) in the high-protein condition compared with the high-fat and high-carbohydrate conditions. In a similar study design, gut hormone responses of 8 healthy volunteers were measured following pancake breakfasts that provided 60% of energy from either protein, fat, or carbohydrate (65). The high-protein meal stimulated the greatest GLP-1TOTAL and PYY3-36 secretion; however, this did not translate into substantial changes in subsequent food intake measured during an ad libitum lunch meal between conditions. Although, this might be due to the timing of the ad libitum lunch 4 h after the test meal, which coincided with a gradual rise of ghrelin concentrations in each condition. In order to assess the influence of relative protein dose on GLP-1 release, 25 overweight men were fed isocaloric test meals (adjusted at the expense of carbohydrate) of normal protein (14% energy from protein), medium-high protein (25%), and high protein (50%). A dose-dependent increase in mean GLP-17–36amide and PYY3-36 concentrations was recorded following meals with increasing protein intake (66). This dose-dependent increase in GLP-1 could have been due to greater protein delivery to more distal parts of the intestine with increasing protein load (38).
The short-term effects of high-protein meals on GLP-1 release and appetite seem to persist over longer time frames. For example, 12 healthy women were provided a high-protein diet (30% energy from protein, 40% carbohydrate, and 30% fat) compared with an adequate-protein diet (10% energy from protein, 60% carbohydrate, and 30% fat) for 4 d, in a randomized crossover design (67). GLP-17–36amide was measured for a 24-h period on day 4 of each respective diet. GLP-17–36amide was significantly greater 15 min following dinner, and tended to be greater following breakfast, after the high-protein compared with the adequate-protein diet. Accordingly, the 24-h AUCs for hunger and satiety were lower and greater, respectively, for the high-protein diet. These studies provide some evidence to suggest that, in both healthy and overweight individuals, a (mixed-macronutrient) diet with a high relative protein composition can enhance GLP-1 release to a greater extent than diets with a high relative proportion of carbohydrate or fat.
Type and form of protein
Given that nutrient delivery and sensing are key mechanisms that regulate gut hormone secretion, it is important to consider the type and form of protein in the context of GLP-1 release.
Solid compared with liquid meals
It is well established that liquid meals empty the stomach at a faster rate than solid meals (68–70). Subsequently, faster nutrient delivery (following a liquid compared with a solid meal) could reduce the time window for intestinal absorption, thereby resulting in greater exposure of nutrients to more distal parts of the intestine, which could elevate GLP-1 release (38). Indeed, in 6 healthy lean males and females, GLP-17–36amide iAUC was significantly greater following a mixed macronutrient liquid compared with a solid meal, matched for energy content and volume (52% energy from carbohydrate, 34% energy from fat, 15% energy from protein) (71). Equally, GLP-17–36amide iAUC was elevated to a greater extent following a liquid mixed meal (345 kcal) compared with a solid mixed meal (362 kcal) in 6 participants who underwent recent surgical or medical weight loss within 1 y (72). Similar findings were also observed in a larger study incorporating 32 participants following RYGB surgery (73). GLP-1TOTAL iAUC was greater following a mixed macronutrient liquid compared with a solid meal (matched for nutrient composition and energy content), although this occurred despite no differences in gastric pouch emptying time between liquid and solid meals. This suggests that other factors such as osmolarity could also be responsible for GLP-1 differences following solid and liquid meals (10, 74). Studies are required to compare GLP-1 release following the ingestion of matched protein meals in solid and liquid form to see whether effects of meal form are modulated by specific macronutrients.
Whey compared with casein
Whey and casein proteins are the major constituents of milk protein, representing 20% and 80% of milk protein, respectively. Previous research has provided inconclusive evidence about whether one protein is more satiating than the other; however, due to faster gastric emptying following whey compared with casein ingestion, whey has been considered more satiating in the short term (< ∼180 min), and casein more satiating in the long term (> ∼180 min) (75). Considering these differences, GLP-1 responses have been variable. Eight healthy females and 1 male ingested whey protein on one occasion and casein protein on another, in the form of a 48-g liquid preload (76). The whey protein preload resulted in a 65% greater GLP-17–36amide iAUC compared with casein over 90 min. Despite this, a longer measurement period might have been more appropriate considering the slower rate of GI transit following casein consumption. Following a longer measurement period of 180 min, there were no significant differences in mean postprandial GLP-17–36amide concentrations between whey and casein protein preloads (30 g) in 24 overweight/obese males and females (77). Moreover, to determine GLP-1 release independent of gastric emptying, 6 healthy males ingested whey and casein in both intact and hydrolyzed (partially digested) forms on separate occasions. There were no significant differences in mean postprandial GLP-17–36amide responses or gastric emptying half-times between conditions over 120 min (78).
Other protein sources
The ingestion of other sources of protein, including gluten, soy, and cod, has been shown to stimulate GLP-1 secretion, although the magnitude of these responses does not appear to differ substantially between source (79–81). This appears consistent in individuals with healthy normal weight (79), overweight/obesity (79, 80), and T2D (81). In summary, findings suggest that ingestion of liquid compared with solid mixed-meals (matched for nutrient composition and caloric content) results in greater GLP-1 secretion. Whey and casein, when emptied from the stomach at similar rates, have comparable GLP-1 responses. Finally, ingestion of other protein forms including soy and gluten results in similar GLP-1 responses to whey and casein in terms of magnitude. Isolating specific amino acids/peptides (available following protein digestion/absorption) could provide some indication as to what conditions are optimal for protein-mediated GLP-1 release.
Amino acids
Individual amino acids are capable of stimulating GLP-1 release, and glutamine (82, 83), phenylalanine (34, 84), arginine (85–87), and tryptophan (88) have been shown to be some of the most effective. In one study, different amino acids (all 10 mM) were perfused through isolated loops of rat small intestine to stimulate GLP-1 release (34). Results suggested that phenylalanine was the most potent amino acid to stimulate GLP-1 secretion, followed by arginine > glutamine ∼ tryptophan > asparagine. Conversely, 10 g l-phenylalanine did not enhance GLP-1 concentrations in comparison with placebo capsules in healthy humans (89). In another study, l-tryptophan (0.15 kcal/min), l-phenylalanine (0.45 kcal/min), and l-glutamine (0.45 kcal/min) were infused intraduodenally for 90 min before a buffet-style test meal in healthy normal weight men (90). The lower rate of infusion of l-tryptophan was due to poor tolerance of higher doses. GLP-1TOTAL ΔiAUCs were comparable between amino acids, although food intake was reduced to a greater extent following l-tryptophan administration compared with l-phenylalanine and l-glutamine. More recently, each amino acid (apart from l-tyrosine due to solubility) was perfused into the lumen or vascular side of isolated rat small intestine (91). Luminal administration showed that l-valine, l-phenylalanine, and alanyl-l-glutamine (stable dipeptide isoform of l-glutamine) were the most powerful stimulators of GLP-1 secretion. Interestingly, vascular but not luminal administration of l-arginine and l-tryptophan resulted in 2.9- and 2.7-fold increases in GLP-1 secretion compared with baseline, which suggests that amino acid–mediated GLP-1 secretion occurs via absorptive and postabsorptive mechanisms (91). The potential to combine multiple amino acids/peptides to stimulate GLP-1 secretion is an exciting avenue for future nutritional interventions.
Summary
There is no conclusive evidence to suggest that proteins, fed in isolation, are more or less potent at stimulating GLP-1 secretion compared with other isolated macronutrients. However, the ingestion of mixed meals with high compared with low relative protein composition could be more effective at enhancing GLP-1 secretion. Protein feeding/administration alone is sufficient to enhance gut hormone release, and therefore identifying the most potent amino acids for stimulating gut hormone release could be beneficial for the design of protein forms/supplements to target enhanced gut hormone availability.
Calcium effects on appetite and GLP-1 concentrations
Calcium intake has been inversely associated with BMI and body fat content (92–94). Moreover, chronic calcium supplementation has also been associated with weight loss (95, 96). Possible mechanisms that could be responsible for this include a reduction in fat absorption (97), increased fat utilization (98), and/or a direct calcium effect on appetite (99, 100). However, the difference in fat absorption reported by Christensen et al. (97) between calcium and control supplements (increase in 1.6–8.8 g/d fecal fat) relates to 15.6–86 kcal/d and therefore is probably not meaningful for energy balance (101). Furthermore, subsequent research has found no evidence of calcium supplementation enhancing fat utilization, at rest or during physical activity (102, 103).
The effect of calcium on appetite could be related to its influence on appetite regulatory hormones. Following the ingestion of a high-calcium compared with a low-calcium mixed macronutrient breakfast there was a 22% increase in GLP-17–36amide iAUC over 120 min in healthy weight individuals (103). This was mirrored by a 19% increase in insulin iAUC for the high-calcium compared with the low-calcium condition. Conversely, in healthy overweight individuals, no differences in postprandial gut hormone concentrations were reported over 420 min in response to isocaloric low-, medium-, or high-calcium meals, or calcium carbonate supplement (104). There was, however, a lower triglyceride response in the medium- and high-calcium conditions compared with the calcium supplement, potentially suggesting decreased fat absorption. Nevertheless, there were no differences in appetite sensation between conditions.
Chronic calcium supplementation might also enhance GLP-1 availability. In a randomized crossover design, participants were provided with a defined diet including bread supplemented with 1 g calcium phosphate per day on one occasion and a placebo bread on the other (separated by a 2-wk washout) (105). GLP-1 responses were measured following a single administration and following the 3-wk intervention. No differences occurred following the single administration between conditions; however, plasma GLP-1TOTAL and GLP-17–36amide AUC were significantly greater following the 3-wk intervention with calcium phosphate compared with the 3-wk placebo (105).
With a relatively small body of literature and different study designs, more research is required to determine whether isolated calcium ingestion can specifically enhance GLP-1 availability, or whether calcium is in fact reliant on the co-ingestion of other macronutrients, including protein.
Protein and calcium co-ingestion enhances GLP-1 release
Foods containing both protein and calcium, for example, dairy, have been shown to stimulate GLP-1 release. Forty-nine overweight men and women were randomly assigned to either a high-dairy diet (∼1400 mg/d dairy) or a control diet (∼700 mg/d dairy) for 12 wk (106). The change in GLP-17–36amide concentrations between baseline and week 12 was significantly greater in the high-dairy condition, although this did not translate into any differences in weight loss between the 2 groups. More recently, gut peptide responses to cheese were measured using a murine intestinal STC-1 cell line (107). Nine of 10 water-soluble extracts of Irish cheddar cheeses stimulated GLP-1 secretion compared with the vehicle control, and all 10 cheeses significantly inhibited DPP4 compared with the buffer alone. In addition, 12 healthy participants were served test meals of reconstituted milk, cheese, whey protein, cod, and wheat gluten (108). There were no differences in GLP-1TOTAL AUC over 60 min between any conditions. A longer measurement period with greater resolution might, however, have been necessary for measuring the complete GLP-1 response.
Although these aforementioned studies utilized meals high in calcium and protein, further research has specifically isolated protein and calcium and demonstrated a potent synergy on GLP-1 release. The first of these studies, previously alluded to, used loops of rat small intestine to perfuse different amino acids in the absence or presence of calcium (34). In the presence of extracellular Ca2+, GLP-1 was secreted following the perfusion of each amino acid. However, the absence of extracellular Ca2+ during perfusion of amino acids completely abolished GLP-1 secretion. Phenylalanine was reported to be the most potent amino acid for stimulating GLP-1 release and was therefore perfused for 90 min in a Ca2+-deplete buffer. Ca2+ concentration was then progressively increased in 0.1, 0.3, 1, 3, to 10 mM steps every 15 min. Phenylalanine-mediated GLP-1 release was augmented by increasing Ca2+ concentrations (Figure 1). This phenylalanine-calcium synergy was also apparent for the release of PYY and GIP.
FIGURE 1.
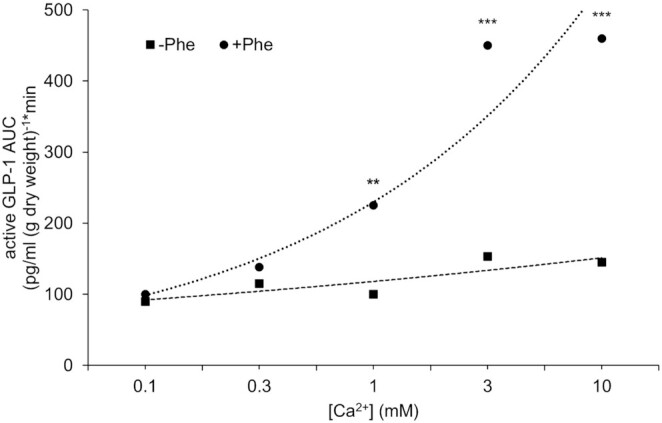
GLP-1 release in response to phenylalanine and accumulative calcium perfusion. Rat small intestine was perfused with Krebs–Henseleit buffer deplete of calcium ± 10 mM phenylalanine. At 20 min, Ca2+ was reintroduced cumulatively into the perfusate every 15 min over 90 min. The figure depicts the extracellular Ca2+–GLP-1 response relation using AUC for each Ca2+ addition. Student unpaired t tests were used to determine significance between control and phenylalanine: **P < 0.01, ***P < 0.001. GLP-1, glucagon-like peptide 1. Reproduced from reference 34 with permission.
The effect of this synergy on gut hormone release was subsequently explored in humans. Following an overnight fast and habitual diet and activity standardization, participants were fed either a control (CON, ∼104 mg calcium and 4 g protein), high-protein (PRO, ∼104 mg calcium and 29 g protein), high-calcium (CAL, ∼1170 mg calcium and 5 g protein), or a high-protein and high-calcium (PROCAL, ∼1170 mg calcium and 29 g protein) preload (109). Blood samples were collected every 15 min for 1 h. Whereas GLP-17–36amide iAUC was higher following PRO and PROCAL compared with CON, there were no clear differences between PRO and PROCAL. However, it could be that the collection period was not long enough to capture the entire response, or that the measurement of venous, as opposed to arterialized, GLP-17–36amide concentrations was problematic for detecting differences between conditions in terms of rate of secretion (110). Interestingly, ad libitum energy intake was overcompensated for with CAL and PROCAL compared with PRO, suggesting these conditions suppressed appetite to a greater extent than protein alone.
In a series of acute experiments, Chen et al. (35) provided further evidence of a synergy between calcium and protein. In the first study, preloads (matched for calcium content—1000 mg) consisted of 4380 mg calcium citrate, 3745 mg milk minerals high in calcium, and 2050 mg milk minerals high in calcium with 50 g whey protein hydrolysate. Milk minerals and protein increased GLP-1TOTAL iAUC by 9-fold compared with calcium citrate and milk minerals alone. The second study aimed to assess whether the addition of milk minerals rich in calcium to whey protein hydrolysate enhanced GLP-1 secretion compared with whey protein hydrolysate alone. Plasma GLP-1TOTAL iAUC was ∼25% greater following milk minerals and protein ingestion compared with protein alone, although no differences were reported for GIP and PYY iAUC. Importantly, the GLP-1TOTAL concentrations measured in response to the co-ingestion of protein and milk minerals were some of the highest ever reported following meal ingestion in humans. Peak plasma GLP-1TOTAL concentrations reached 91 ± 20 pmol/L for milk minerals and protein, compared with 43 ± 12 pmol/L and 78 ± 20 pmol/L for milk minerals alone and protein conditions alone, respectively.
Summary
Animal models and human studies have provided initial evidence of a potent synergy between protein and calcium on GLP-1 secretion. However, understanding the mechanisms behind this synergy are fundamental for optimizing nutrition to maximize GLP-1 availability.
Putative mechanisms underpinning potential protein- and calcium-mediated GLP-1 release
Nutrients that pass through the intestinal lumen are sensed by open-type enteroendocrine cells, which in turn leads to the release of peptide hormones from secretory granules into the lamina propria, before being taken up by blood capillaries or lymphatic vessels (38, 111, 112). Membrane-bound receptors and transporters are therefore pivotal for the detection and absorption of these nutrients, and many macronutrient-specific receptors have been distinguished. Notable receptors/transporters involved in amino acid sensing include: G-protein–coupled receptor class C 6A (GPRC6A) (113), G-protein–coupled receptor 142 (114), type 1 taste receptors (T1Rs) (115), and sodium-dependent neutral amino acid transporter (B AT1) (30). Receptors for peptides/oligopeptides include: G-protein–coupled receptor 93 (GPR93) (116) and peptide transporter 1 (PEPT1) (117). Additionally, the extracellular calcium-sensing receptor (CaSR) is receptive to both amino acids and peptides (30, 118, 119). Amongst these, CaSR, GPRC6A, and PEPT1 are all reportedly selective/sensitive to calcium.
The CaSR is present in the GI tract and responds to extracellular amino acids and calcium
CaSR is a class C G-protein–coupled receptor (GPCR) that responds to calcium as its principal physiological ligand (120, 121). However, some agonists are able to modify the receptor's affinity via allosteric actions (type 2 agonists). Amino acids are type 2 agonists that can bind to the extracellular domain of the CaSR and modulate its activity (122). Furthermore, the CaSR binds extracellular Ca2+ over a concentration range of 0.5–10 mM (122). Therefore, in the presence of these concentrations of calcium, CaSR can act as an l-amino acid receptor. Importantly, CaSR operates in a reciprocal fashion. While being directly activated by Ca2+, occupancy of the l-amino acid binding site enhances the sensitivity of CaSR to calcium (120, 123). Therefore, during a meal in which calcium and protein are in abundance, calcium and amino acids could act as coagonists of the CaSR (120). However, the CaSR is not responsive to every amino acid. Aromatic amino acids, including phenylalanine and tryptophan, and aliphatic amino acids are the most efficient agonists of CaSR. Charged amino acids are significantly less effective, with branched-chain amino acids almost ineffective (120).
The CaSR has been localized through immunostaining on both the apical and basolateral membranes of the small and large intestine villus and surface cells in both humans (124) and rats (125, 126). However, evidence appears inconclusive as to which site is most capable of sensing amino acids and initiating GLP-1 release. Using isolated perfused rat small intestine, calindol, an allosteric modulator of CaSR, strongly stimulated GLP-1 release when infused intravascularly but had minimal effects when administered intraluminally (127). Additionally, recent work utilizing the same model demonstrated that vascular inhibition of CaSR with NPS2143 (a CaSR antagonist) significantly reduced amino acid–stimulated GLP-1 release (91). This suggests that amino acid/peptide absorption is important, whereby amino acid delivery to the vasculature through basolateral transporters then activates CaSR located on the basolateral membrane (91, 127). Despite this, other reports suggest that CaSR can sense luminal nutrients (122), and that GLP-1, unlike GIP, does not necessarily require intestinal absorption for it to be stimulated (128).
The CaSR in particular, appears to be an important receptor for calcium- and protein-mediated GLP-1 release. Calhex 231 (another CaSR inhibitor) significantly inhibited amino acid–induced stimulation of GLP-1 by ∼70% in the presence of extracellular Ca2+, using isolated loops of rat small intestine (34). Equally, Calhex 231 and NPS2143 both significantly lowered peptone-triggered GLP-1 secretion from primary colonic cultures (117). Furthermore, unlike many other receptors, CaSR undergoes agonist-driven insertional signaling, making it highly resistant to functional desensitization (118, 129, 130) and therefore potentially capable of sustaining prolonged GLP-1 release in response to amino acids and calcium.
GPRC6A contains a calcium-binding site and could play a minor role in protein-and calcium-mediated GLP-1 release
GPRC6A is also a class C GPCR that shares 32% amino acid identity with CaSR, suggesting they could share significant functional properties (131, 132). GPRC6A is reportedly activated by a variety of ligands: osteocalcin, testosterone, basic amino acids, and divalent and trivalent cations (133, 134). Additionally, GPRC6A possesses a calcium-binding site, albeit with a weaker affinity than CaSR (135), although it is thought that this binding site allows direct activation of GPRC6A by calcium (136). When cells expressing GPRC6A were preincubated in physiological saline solution of low Ca2+ concentration (0.5 mM) and then switched to high Ca2+ (5 mM), several amino acids demonstrated intracellular Ca2+-mobilizing responses that were not evident under control conditions (137). The enhanced Ca2+-mobilizing responses could theoretically lead to greater GLP-1 exocytosis.
GPRC6A is activated by basic amino acids, which trigger GLP-1 secretion in GLUTag cell lines (derived from colonic tumors of transgenic mice expressing T antigen), but not in primary murine intestinal cultures (113, 134). For instance, l-ornithine administration significantly and dose-dependently increased intracellular Ca2+ concentrations, which correlated with increased GLP-1 secretion in a GLUTag cell line compared with unstimulated control cells (113). However, in the presence of a GPRC6A antagonist, calindol, l-ornithine–induced calcium elevations and subsequent GLP-1 release were suppressed. The authors also transfected GPRC6A small interfering RNAs into GLUTag cells and successfully reduced the expression of GPRC6A. Accordingly l-ornithine–induced GLP-1 release was significantly decreased. Despite this, utilizing a rodent GPRC6A whole-body knockout model, oral administration of both l-arginine and l-ornithine significantly increased plasma GLP-1TOTAL to a comparable magnitude in GPRC6A knockout and wild-type mice (138). However, as extensively discussed by Pi et al. (133, 139), caution should be taken when analyzing the reported effects. GPRC6A mouse knockout models are challenging due to variations in metabolic phenotype (140, 141), which are reportedly influenced by environmental challenges and genetic differences in mouse strains (139). In addition, the translational validity of such studies is questioned because GPRC6A (in humans) presents an evolutionarily divergent genotype characterized by a gain-of-function polymorphism (GPRC6AKGKY), which might alter the role of this receptor (133). The research so far suggests that GPRC6A likely plays a role in the coordination of GLP-1 release but is not essential for it. Further studies in humans are necessary to fully understand the contribution of this receptor to GLP-1 secretion and metabolic health.
GPCR signaling involves 2 major pathways leading to GLP-1 exocytosis
GPCRs, including CaSR and GPRC6A, represent the largest group of cell surface receptors. Following receptor binding, intracellular G-proteins are activated and couple to 2 main signaling pathways: the cAMP pathway and the phosphatidylinositol pathway (130, 142), illustrated in Figure 2. The former involves the modulation of cAMP, which activates downstream targets Epac2 (exchange protein directly activated by cAMP 2) and protein kinase A, influential for GLP-1 exocytosis (142–144). The latter pathway involves the activation of phospholipase C, which in turn mediates the production of diacylglycerol and inositol 1,4,5-triphosphate. These lead to the activation of protein kinase C and mobilization of Ca2+ from the endoplasmic reticulum, respectively, leading to GLP-1 exocytosis (142, 145, 146).
FIGURE 2.
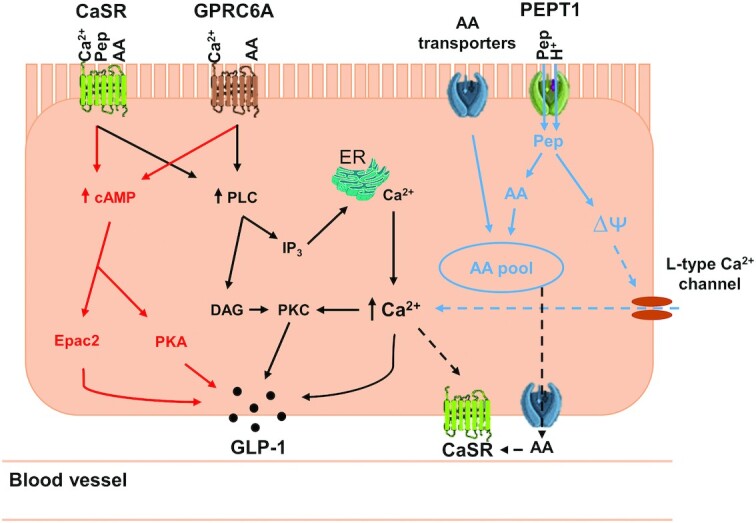
The potential putative mechanisms of calcium and protein synergy inducing GLP-1 secretion. Amino acids and peptides are sensed by CaSR (amino acids and peptides) and GPRC6A (amino acids only). The ability of CaSR to bind both peptides and amino acids is dependent on the presence of calcium, whereas GPRC6A also contains a calcium-binding site. This binding leads to 2 downstream signaling pathways involving cAMP (red) and phosphatidylinositol (black), leading to GLP-1 exocytosis. PEPT1 mediates the transport of peptides into the cell and subsequently causes membrane depolarization (potentially leading to calcium influx via L-type Ca2+ channels—blue dashed arrows) and ultimately GLP-1 exocytosis. Amino acids that enter the cell via amino acid transporters, including B˚AT1, and peptides that are broken down, join the amino acid intracellular pool. Intracellular amino acids and peptides transported out of the cell might also be sensed by CaSR localized on the basolateral membrane (black dashed arrows), which similarly triggers the signaling pathways highlighted in red and black. AA, amino acids; B˚AT1, sodium-dependent neutral amino acid transporter; CaSR, extracellular calcium-sensing receptor; DAG, diacylglycerol; Epac2, exchange protein directly activated by cAMP 2; ER, endoplasmic reticulum; GLP-1, glucagon-like peptide 1; GPRC6A, G-protein–coupled receptor class C 6A; IP3, inositol 1,4,5-triphosphate; Pep, peptides; PEPT1, peptide transporter 1; PKA, protein kinase A; PKC, protein kinase C; PLC, phospholipase C; ∆ᴪ, membrane depolarization.
Peptide transporter 1 provides intestinal transport for di-/tripeptides and might involve voltage-gated Ca2+ channel opening
The oligopeptide transporter, peptide transporter 1 (PEPT1), is located in the intestinal brush border membrane and has an important role in protein absorption in the intestine (147). PEPT1 is selective for dipeptides and tripeptides, but not for free amino acids or peptides containing ≥4 amino acids (148). PEPT1 is a member of the Slc15 family of proton co-transporters and its unique feature of using an inwardly directed proton gradient enables peptides to enter the cell even against a concentration gradient (148). This leads to membrane depolarization, subsequent calcium entry, and enteroendocrine hormone secretion (142). PEPT1-mediated transport of dipeptides has been shown to elicit hormone secretion in PEPT1-transfected STC-1 cells (149). The authors used human growth hormone (GH) co-transfection with PEPT1 as a reporter for PEPT1-mediated hormone secretion. They reported that 10 mM glycine-glycine (Gly-Gly) and 10 mM glycine-sarcosine (Gly-Sar) increased GH secretion in PEPT1-transfected STC-1 cells 3.8- and 4-fold, respectively. Moreover, using primary cultures from murine colon, the PEPT1 substrate cefalexin enhanced GLP-1 secretion 1.4-fold above baseline, suggesting a key coupling between PEPT1 and L-cell activation (117). In support, the GLP-1 secretory response to the non-hydrolysable Gly-Sar was suppressed by the PEPT1 antagonist 4-AMBA (117). Gly-Sar–mediated GLP-1 secretion was also sensitive to calcium, whereby Gly-Sar–induced GLP-1 secretion was completely inhibited in the absence of extracellular calcium or by the L-type calcium channel blocker nifedipine. Equally, peptone-mediated GLP-1 secretion was inhibited in a rat intestine perfusion model by 4-AMBA (127). However, amino acid–mediated GLP-1 secretion was also suppressed by 4-AMBA, suggesting the observed antagonism might not be fully specific for PEPT1. In contrast to the previously mentioned study, opening of L-type Ca2+ channels was not essential for peptone-mediated GLP-1 secretion, because nifedipine only blocked GLP-1 secretion following the luminal perfusion of glucose and not peptones (127). These findings suggest that calcium influx could be important for PEPT1-mediated hormone secretion in L-cells. Absorption of di-/tri-peptides into the cell could also be important for the basolateral CaSR sensing of amino acids/peptides when they are transported out of the cell.
Summary
The speculated mechanisms behind calcium- and protein-mediated GLP-1 release are summarized in Figure 2. CaSR is the key component of this working model, where both CaSR and GPRC6A receptors demonstrate a key coupling of amino acids/protein and calcium to trigger GLP-1 release. The potential localization of CaSR on both the luminal and vascular membrane of the L-cell could mediate GLP-1 release through luminal sensing of peptides and amino acids, and also basolateral sensing of amino acids in the vasculature. Basolateral sensing relies on luminal absorption of di- and tri-peptides/amino acids into the cell through PEPT1 and amino acid transporters including BoAT1, and then autocrine action following movement of amino acids out of the cell via basolateral transporters.
Areas for Future Research
As highlighted in this narrative review there is a limited number of human studies that have investigated the effect of protein and calcium co-ingestion on the secretion of endogenous GLP-1, and further studies are required to support the encouraging findings reported so far (35). Whereas many studies have investigated the effects of manipulating the type and amount of protein on gut hormone responses, less is known about the optimal dose of calcium to maximize protein-stimulated GLP-1 release. Although almost all dietary calcium is absorbed from the upper intestine (150), it might be possible that calcium can reach more distal areas of the intestine following a large dose. This could lead to increased GPCR activation and calcium absorption into the intracellular Ca2+ pool, and subsequently greater exocytosis of GLP-1. Further studies are also required to confirm the mechanisms speculated in the previous section. In particular, it is important to determine whether basolateral or apical localization of CaSR is more significant for gut hormone release, or whether they are equally vital. This would provide information on whether CaSR-mediated GLP-1 secretion is more dependent on nutrient contact, nutrient absorption, or both.
Currently, the only research conducted in humans using a protein-calcium design has sampled healthy, lean participants and is acute in nature (35, 109). Studies in individuals with overweight/obesity and T2D are required to determine if this effect is present in clinical populations requiring nutritional interventions for the regulation of blood glucose and weight loss. It is also important to determine whether this heightened response can be sustained chronically, potentially by an intervention diet high in protein and calcium, or by manipulating certain meals of the day, for example, breakfast, to feed a high-protein/high-calcium drink/supplement. Chronically, it might also be possible to influence the enteroendocrine cell population via nutritional intervention considering the rapid turnover of enteroendocrine cells of ∼5 d (151). If a high-protein/high-calcium meal intervention could upregulate intestinal L-cell growth, or equally the abundance of GPCRs at the cell membrane, this could be hugely important for gut hormone availability and associated blood glucose and appetite regulation.
Lastly, given that a number of receptors are selective to a range of different amino acids and peptides, the design of certain amino acid/peptide mixtures to maximize multiple receptor stimulation should be tested in vitro, and then, following optimization, in human participants. This could lead to the possibility of evoking a substantial GLP-1 response without the need to feed calorie-dense meals. This could improve weight loss via 2 main mechanisms: 1) by suppressing hunger through a potent peptide hormone response; and 2) the low-caloric nature of the nutrient stimulation.
Conclusion
Emerging research provides evidence of a potential synergy between protein and calcium on GLP-1 secretion. This synergistic effect has been substantiated by findings in cell cultures, animal models, and in human studies. The collective activation of different receptors/transporters as highlighted in our working model provides some insight into how this synergy might work on a mechanistic level. Future studies are required to substantiate this model and provide evidence of elevated GLP-1 responses translating to appetite suppression and blood glucose control, given the great potential to provide this nutritional stimulus in low-caloric doses.
ACKNOWLEDGEMENTS
The authors’ responsibilities were as follows—All authors were responsible for the design, writing, and final content; and all authors read and approved the final manuscript.
Notes
JDW is supported by a PhD studentship from Arla Foods Ingredients (DK). FK thanks MRC (UK) for funding (MR/P002927/1). JTG has received research funding from Arla Foods Ingredients (DK), LucozadeRibena Suntory Ltd (UK), and Kenniscentrum Suiker & Voeding (NL); and has acted as a consultant to PepsiCo (UK). These funders had no role in the design, implementation, analysis, or interpretation of this manuscript.
Author disclosures: The authors report no conflicts of interest.
Abbreviations used: B˚AT1, sodium-dependent neutral amino acid transporter; CaSR, extracellular calcium-sensing receptor; DPP4, dipeptidyl peptidase-IV; GH, growth hormone; GI, gastrointestinal; GIP, glucose-dependent insulinotropic polypeptide; GLP-1, glucagon-like peptide 1; GPCR, G-protein–coupled receptor; GPRC6A, G-protein–coupled receptor class C 6A; iAUC, incremental area under the curve; PEPT1, peptide transporter 1; PYY, peptide tyrosine-tyrosine; RYGB, Roux-en-Y gastric bypass surgery; Sar, sarcosine; T1R, type 1 taste receptor; T2D, type 2 diabetes.
Contributor Information
Jonathan D Watkins, Centre for Nutrition, Exercise and Metabolism, Department for Health, University of Bath, Bath, United Kingdom.
Françoise Koumanov, Centre for Nutrition, Exercise and Metabolism, Department for Health, University of Bath, Bath, United Kingdom.
Javier T Gonzalez, Centre for Nutrition, Exercise and Metabolism, Department for Health, University of Bath, Bath, United Kingdom.
References
- 1. Ritchie H, Roser M. Obesity. [Internet]. 2017; [cited 2021 Feb 3]. Available from: https://ourworldindata.org/obesity. [Google Scholar]
- 2. Khan MAB, Hashim MJ, King JK, Govender RD, Mustafa H, Al Kaabi J. Epidemiology of type 2 diabetes—global burden of disease and forecasted trends. J Epidemiol Glob Health. 2020;10(1):107–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Diabetes UK . The cost of diabetes report. [Internet]. 2014; [cited 2021 Feb 3]. Available from: https://www.diabetes.org.uk/resources-s3/2017-11/diabetes%20uk%20cost%20of%20diabetes%20report.pdf. [Google Scholar]
- 4. McKinsey Global Institute . Overcoming obesity: an initial economic analysis. [Internet]. 2014; [cited 2021 Feb 3]. Available from: https://www.mckinsey.com/∼/media/McKinsey/Business%20Functions/Economic%20Studies%20TEMP/Our%20Insights/How%20the%20world%20could%20better%20fight%20obesity/MGI_Overcoming_obesity_Full_report.ashx. [Google Scholar]
- 5. Pais R, Gribble FM, Reimann F. Stimulation of incretin secreting cells. Ther Adv Endocrinol Metab. 2016;7(1):24–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Buchwald H, Estok R, Fahrbach K, Banel D, Jensen MD, Pories WJ, Bantle JP, Sledge I. Weight and type 2 diabetes after bariatric surgery: systematic review and meta-analysis. Am J Med. 2009;122(3):248–56.e5. [DOI] [PubMed] [Google Scholar]
- 7. Pournaras DJ, Aasheim ET, Sovik TT, Andrews R, Mahon D, Welbourn R, Olbers T, le Roux CW. Effect of the definition of type II diabetes remission in the evaluation of bariatric surgery for metabolic disorders. Br J Surg. 2011;99(1):100–3. [DOI] [PubMed] [Google Scholar]
- 8. Picot J, Jones J, Colquitt JL, Gospodarevskaya E, Loveman E, Baxter L, Clegg AJ. The clinical effectiveness and cost-effectiveness of bariatric (weight loss) surgery for obesity: a systematic review and economic evaluation. Health Technol Assess (Rockv). 2009;13(41):1–190., 215–357, iii–iv. [DOI] [PubMed] [Google Scholar]
- 9. Ionut V, Bergman RN. Mechanisms responsible for excess weight loss after bariatric surgery. J Diabetes Sci Technol. 2011;5(5):1263–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Spreckley E, Murphy KG. The L-cell in nutritional sensing and the regulation of appetite. Front Nutr. 2015;2:23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Meek CL, Lewis HB, Reimann F, Gribble FM, Park AJ. The effect of bariatric surgery on gastrointestinal and pancreatic peptide hormones. Peptides. 2016;77:28–37. [DOI] [PubMed] [Google Scholar]
- 12. Dirksen C, Jorgensen NB, Bojsen-Moller KN, Kielgast U, Jacobsen SH, Clausen TR, Worm D, Hartmann B, Rehfeld JF, Damgaard Met al. Gut hormones, early dumping and resting energy expenditure in patients with good and poor weight loss response after Roux-en-Y gastric bypass. Int J Obes. 2013;37(11):1452–9. [DOI] [PubMed] [Google Scholar]
- 13. Sternini C, Anselmi L, Rozengurt E. Enteroendocrine cells: a site of ‘taste’ in gastrointestinal chemosensing. Curr Opin Endocrinol Diabetes Obes. 2008;15(1):73–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Gunawardene AR, Corfe BM, Staton CA. Classification and functions of enteroendocrine cells of the lower gastrointestinal tract. Int J Exp Pathol. 2011;92(4):219–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Bodnaruc AM, Prud'homme D, Blanchet R, Giroux I. Nutritional modulation of endogenous glucagon-like peptide-1 secretion: a review. Nutr Metab (Lond). 2016;13(1):92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Holst JJ. Incretin hormones and the satiation signal. Int J Obes. 2013;37(9):1161–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Holst JJ, Albrechtsen NJW, Rosenkilde MM, Deacon CF. Physiology of the incretin hormones, GIP and GLP-1-regulation of release and posttranslational modifications. Compr Physiol. 2019;9(4):1339–81. [DOI] [PubMed] [Google Scholar]
- 18. Drucker DJ, Nauck MA. The incretin system: glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes. Lancet. 2006;368(9548):1696–705. [DOI] [PubMed] [Google Scholar]
- 19. Wilding JPH, Batterham RL, Calanna S, Davies M, Van Gaal LF, Lingvay I, McGowan BM, Rosenstock J, Tran MTD, Wadden TAet al. Once-weekly semaglutide in adults with overweight or obesity. N Engl J Med. 2021;384(11):989–1002. [DOI] [PubMed] [Google Scholar]
- 20. Gentilella R, Pechtner V, Corcos A, Consoli A. Glucagon-like peptide-1 receptor agonists in type 2 diabetes treatment: are they all the same?. Diabetes Metab Res Rev. 2019;35(1):e3070. [DOI] [PubMed] [Google Scholar]
- 21. Tran KL, Park YI, Pandya S, Muliyil NJ, Jensen BD, Huynh K, Nguyen QT. Overview of glucagon-like peptide-1 receptor agonists for the treatment of patients with type 2 diabetes. Am Health Drug Benefits. 2017;10(4):178–88. [PMC free article] [PubMed] [Google Scholar]
- 22. Filippatos TD, Panagiotopoulou TV, Elisaf MS. Adverse effects of GLP-1 receptor agonists. Rev Diabet Stud. 2014;11(3–4):202–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Prasad-Reddy L, Isaacs D. A clinical review of GLP-1 receptor agonists: efficacy and safety in diabetes and beyond. Drugs Context. 2015;4:212283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Steinert RE, Beglinger C. Nutrient sensing in the gut: interactions between chemosensory cells, visceral afferents and the secretion of satiation peptides. Physiol Behav. 2011;105(1):62–70. [DOI] [PubMed] [Google Scholar]
- 25. Holst JJ, Deacon CF. Glucagon-like peptide-1 mediates the therapeutic actions of DPP-IV inhibitors. Diabetologia. 2005;48(4):612–5. [DOI] [PubMed] [Google Scholar]
- 26. Sisley S, Gutierrez-Aguilar R, Scott M, D'Alessio DA, Sandoval DA, Seeley RJ. Neuronal GLP1R mediates liraglutide's anorectic but not glucose-lowering effect. J Clin Invest. 2014;124(6):2456–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Krieger J-P, Arnold M, Pettersen KG, Lossel P, Langhans W, Lee SJ. Knockdown of GLP-1 receptors in vagal afferents affects normal food intake and glycemia. Diabetes. 2016;65(1):34. [DOI] [PubMed] [Google Scholar]
- 28. Krieger JP, Langhans W, Lee SJ. Vagal mediation of GLP-1’s effects on food intake and glycemia. Physiol Behav. 2015;152(Pt B):372–80. [DOI] [PubMed] [Google Scholar]
- 29. Svendsen B, Pedersen J, Albrechtsen NJ, Hartmann B, Torang S, Rehfeld JF, Poulsen SS, Holst JJ. An analysis of cosecretion and coexpression of gut hormones from male rat proximal and distal small intestine. Endocrinology. 2015;156(3):847–57. [DOI] [PubMed] [Google Scholar]
- 30. Reimann F, Gribble FM. Mechanisms underlying glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1 secretion. J Diabetes Investig. 2016;7(Suppl 1):13–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Elliott RM, Morgan LM, Tredger JA, Deacon S, Wright J, Marks V. Glucagon-like peptide-1 (7-36)amide and glucose-dependent insulinotropic polypeptide secretion in response to nutrient ingestion in man: acute post-prandial and 24-h secretion patterns. J Endocrinol. 1993;138(1):159–66. [DOI] [PubMed] [Google Scholar]
- 32. Herrmann C, Goke R, Richter G, Fehmann HC, Arnold R, Goke B. Glucagon-like peptide-1 and glucose-dependent insulin-releasing polypeptide plasma levels in response to nutrients. Digestion. 1995;56(2):117–26. [DOI] [PubMed] [Google Scholar]
- 33. Chambers ES, Preston T, Frost G, Morrison DJ. Role of gut microbiota-generated short-chain fatty acids in metabolic and cardiovascular health. Curr Nutr Rep. 2018;7(4):198–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Mace OJ, Schindler M, Patel S. The regulation of K- and L-cell activity by GLUT2 and the calcium-sensing receptor CasR in rat small intestine. J Physiol. 2012;590(12):2917–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Chen Y-C, Smith HA, Hengist A, Chrzanowski-Smith OJ, Mikkelsen UR, Carroll HA, Betts JA, Thompson D, Saunders J, Gonzalez JT. Co-ingestion of whey protein hydrolysate with milk minerals rich in calcium potently stimulates glucagon-like peptide-1 secretion: an RCT in healthy adults. Eur J Nutr. 2020;59:2449–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Tolhurst G, Reimann F, Gribble FM. Nutritional regulation of glucagon-like peptide-1 secretion. J Physiol. 2009;587(1):27–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Martin AM, Sun EW, Keating DJ. Mechanisms controlling hormone secretion in human gut and its relevance to metabolism. J Endocrinol. 2019;244(1):R1–R15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Holst JJ. The physiology of glucagon-like peptide 1. Physiol Rev. 2007;87(4):1409–39. [DOI] [PubMed] [Google Scholar]
- 39. Orskov C, Rabenhoj L, Wettergren A, Kofod H, Holst JJ. Tissue and plasma concentrations of amidated and glycine-extended glucagon-like peptide I in humans. Diabetes. 1994;43(4):535–9. [DOI] [PubMed] [Google Scholar]
- 40. Kuhre RE, Albrechtsen NW, Windeløv JA, Svendsen B, Hartmann B, Holst JJ. GLP-1 amidation efficiency along the length of the intestine in mice, rats and pigs and in GLP-1 secreting cell lines. Peptides. 2014;55:52–7. [DOI] [PubMed] [Google Scholar]
- 41. Lamers D, Famulla S, Wronkowitz N, Hartwig S, Lehr S, Ouwens DM, Eckardt K, Kaufman JM, Ryden M, Müller Set al. Dipeptidyl peptidase 4 is a novel adipokine potentially linking obesity to the metabolic syndrome. Diabetes. 2011;60(7):1917–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Sell H, Bluher M, Kloting N, Schlich R, Willems M, Ruppe F, Knoefel WT, Dietrich A, Fielding BA, Arner Pet al. Adipose dipeptidyl peptidase-4 and obesity: correlation with insulin resistance and depot-specific release from adipose tissue in vivo and in vitro. Diabetes Care. 2013;36(12):4083–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Deacon CF, Johnsen AH, Holst JJ. Degradation of glucagon-like peptide-1 by human plasma in vitro yields an N-terminally truncated peptide that is a major endogenous metabolite in vivo. J Clin Endocrinol Metab. 1995;80(3):952–7. [DOI] [PubMed] [Google Scholar]
- 44. Müller TD, Finan B, Bloom SR, D'Alessio D, Drucker DJ, Flatt PR, Fritsche A, Gribble F, Grill HJ, Habener JFet al. Glucagon-like peptide 1 (GLP-1). Mol Metab. 2019;30:72–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Lu WJ, Yang Q, Sun W, Woods SC, D'Alessio D, Tso P. The regulation of the lymphatic secretion of glucagon-like peptide-1 (GLP-1) by intestinal absorption of fat and carbohydrate. Am J Physiol Gastrointest Liver Physiol. 2007;293(5):G963–71. [DOI] [PubMed] [Google Scholar]
- 46. Arnold M, Dai Y, Tso P, Langhans W. Meal-contingent intestinal lymph sampling from awake, unrestrained rats. Am J Physiol Regul Integr Comp Physiol. 2012;302(12):R1365–R71. [DOI] [PubMed] [Google Scholar]
- 47. Orskov C, Poulsen SS, Moller M, Holst JJ. Glucagon-like peptide I receptors in the subfornical organ and the area postrema are accessible to circulating glucagon-like peptide I. Diabetes. 1996;45(6):832–5. [DOI] [PubMed] [Google Scholar]
- 48. Vilsbøll T, Krarup T, Deacon CF, Madsbad S, Holst JJ. Reduced postprandial concentrations of intact biologically active glucagon-like peptide 1 in type 2 diabetic patients. Diabetes. 2001;50(3):609–13. [DOI] [PubMed] [Google Scholar]
- 49. Vollmer K, Holst JJ, Baller B, Ellrichmann M, Nauck MA, Schmidt WE, Meier JJ. Predictors of incretin concentrations in subjects with normal, impaired, and diabetic glucose tolerance. Diabetes. 2008;57(3):678–87. [DOI] [PubMed] [Google Scholar]
- 50. Lim GE, Brubaker PL. Glucagon-like peptide-1 secretion by the l-Cell. The View From Within. 2006;55(Supplement 2):S70–S7. [Google Scholar]
- 51. Rask E, Olsson T, Söderberg S, Johnson O, Seckl J, Holst JJ, Ahrén B, Northern Sweden Monitoring of Trends and Determinants in Cardiovascular Disease (MONICA). Impaired incretin response after a mixed meal is associated with insulin resistance in nondiabetic men. Diabetes Care. 2001;24(9):1640–5. [DOI] [PubMed] [Google Scholar]
- 52. Rocca AS, Brubaker PL. Role of the vagus nerve in mediating proximal nutrient-induced glucagon-like peptide-1 secretion. Endocrinology. 1999;140(4):1687–94. [DOI] [PubMed] [Google Scholar]
- 53. Theodorakis MJ, Carlson O, Michopoulos S, Doyle ME, Juhaszova M, Petraki K, Egan JM. Human duodenal enteroendocrine cells: source of both incretin peptides, GLP-1 and GIP. Am J Physiol Endocrinol Metab. 2006;290(3):E550–9. [DOI] [PubMed] [Google Scholar]
- 54. Verdich C, Flint A, Gutzwiller JP, Naslund E, Beglinger C, Hellstrom PM, Long SJ, Morgan LM, Holst JJ, Astrup A. A meta-analysis of the effect of glucagon-like peptide-1 (7-36) amide on ad libitum energy intake in humans. J Clin Endocrinol Metab. 2001;86(9):4382–9. [DOI] [PubMed] [Google Scholar]
- 55. MacDonald PE, El-Kholy W, Riedel MJ, Salapatek AM, Light PE, Wheeler MB. The multiple actions of GLP-1 on the process of glucose-stimulated insulin secretion. Diabetes. 2002;51(Suppl 3):S434–42. [DOI] [PubMed] [Google Scholar]
- 56. Mayo KE, Miller LJ, Bataille D, Dalle S, Goke B, Thorens B, Drucker DJ. International Union of Pharmacology. XXXV. The glucagon receptor family. Pharmacol Rev. 2003;55(1):167–94. [DOI] [PubMed] [Google Scholar]
- 57. Hayes MR. Neuronal and intracellular signaling pathways mediating GLP-1 energy balance and glycemic effects. Physiol Behav. 2012;106(3):413–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Rolls BJ, Hetherington M, Burley VJ. The specificity of satiety: the influence of foods of different macronutrient content on the development of satiety. Physiol Behav. 1988;43(2):145–53. [DOI] [PubMed] [Google Scholar]
- 59. Vandewater K, Vickers Z. Higher-protein foods produce greater sensory-specific satiety. Physiol Behav. 1996;59(3):579–83. [DOI] [PubMed] [Google Scholar]
- 60. Leidy HJ, Clifton PM, Astrup A, Wycherley TP, Westerterp-Plantenga MS, Luscombe-Marsh ND, Woods SC, Mattes RD. The role of protein in weight loss and maintenance. Am J Clin Nutr. 2015;101(6):1320S–9S. [DOI] [PubMed] [Google Scholar]
- 61. Layer P, Holst JJ, Grandt D, Goebell H. Ileal release of glucagon-like peptide-1 (GLP-1). Association with inhibition of gastric acid secretion in humans. Dig Dis Sci. 1995;40(5):1074–82. [DOI] [PubMed] [Google Scholar]
- 62. Carr RD, Larsen MO, Winzell MS, Jelic K, Lindgren O, Deacon CF, Ahren B. Incretin and islet hormonal responses to fat and protein ingestion in healthy men. Am J Physiol Endocrinol Metab. 2008;295(4):E779–84. [DOI] [PubMed] [Google Scholar]
- 63. Chungchunlam SM, Henare SJ, Ganesh S, Moughan PJ. Dietary whey protein influences plasma satiety-related hormones and plasma amino acids in normal-weight adult women. Eur J Clin Nutr. 2015;69(2):179–86. [DOI] [PubMed] [Google Scholar]
- 64. Batterham RL, Heffron H, Kapoor S, Chivers JE, Chandarana K, Herzog H, Le Roux CW, Thomas EL, Bell JD, Withers DJ. Critical role for peptide YY in protein-mediated satiation and body-weight regulation. Cell Metab. 2006;4(3):223–33. [DOI] [PubMed] [Google Scholar]
- 65. van der Klaauw AA, Keogh JM, Henning E, Trowse VM, Dhillo WS, Ghatei MA, Farooqi IS. High protein intake stimulates postprandial GLP1 and PYY release. Obesity. 2013;21(8):1602–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Belza A, Ritz C, Sorensen MQ, Holst JJ, Rehfeld JF, Astrup A. Contribution of gastroenteropancreatic appetite hormones to protein-induced satiety. Am J Clin Nutr. 2013;97(5):980–9. [DOI] [PubMed] [Google Scholar]
- 67. Lejeune M, Westerterp KR, Adam TCM, Luscombe-Marsh ND, Westerterp-Plantenga MS. Ghrelin and glucagon-like peptide 1 concentrations, 24-h satiety, and energy and substrate metabolism during a high-protein diet and measured in a respiration chamber. Am J Clin Nutr. 2006;83(1):89–94. [DOI] [PubMed] [Google Scholar]
- 68. Little TJ, Pilichiewicz AN, Russo A, Phillips L, Jones KL, Nauck MA, Wishart J, Horowitz M, Feinle-Bisset C. Effects of intravenous glucagon-like peptide-1 on gastric emptying and intragastric distribution in healthy subjects: relationships with postprandial glycemic and insulinemic responses. J Clin Endocrinol Metab. 2006;91(5):1916–23. [DOI] [PubMed] [Google Scholar]
- 69. Russo A, Stevens JE, Chen R, Gentilcore D, Burnet R, Horowitz M, Jones KL. Insulin-induced hypoglycemia accelerates gastric emptying of solids and liquids in long-standing type 1 diabetes. J Clin Endocrinol Metab. 2005;90(8):4489–95. [DOI] [PubMed] [Google Scholar]
- 70. Kelly KA. Gastric emptying of liquids and solids: roles of proximal and distal stomach. Am J Physiol. 1980;239(2):G71–6. [DOI] [PubMed] [Google Scholar]
- 71. Brynes AE, Frost GS, Edwards CMB, Ghatei MA, Bloom SR. Plasma glucagon-like peptide-1 (7-36) amide (GLP-1) response to liquid phase, solid phase, and meals of differing lipid composition. Nutrition. 1998;14(5):433–6. [DOI] [PubMed] [Google Scholar]
- 72. Lee CJ, Brown TT, Cheskin LJ, Choi P, Moran TH, Peterson L, Matuk R, Steele KE. Effects of meal composition on postprandial incretin, glucose and insulin responses after surgical and medical weight loss. Obes Sci Pract. 2015;1(2):104–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Stano S, Alam F, Wu L, Dutia R, Ng SN, Sala M, McGinty J, Laferrère B. Effect of meal size and texture on gastric pouch emptying and glucagon-like peptide 1 after gastric bypass surgery. Surg Obes Relat Dis. 2017;13(12):1975–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Petersen B, Christiansen J, Holst JJ. A glucose-dependent mechanism in jejunum inhibits gastric acid secretion: a response mediated through enteroglucagon?. Scand J Gastroenterol. 1985;20(2):193–7. [DOI] [PubMed] [Google Scholar]
- 75. Bendtsen LQ, Lorenzen JK, Bendsen NT, Rasmussen C, Astrup A. Effect of dairy proteins on appetite, energy expenditure, body weight, and composition: a review of the evidence from controlled clinical trials. Adv Nutr. 2013;4(4):418–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Hall WL, Millward DJ, Long SJ, Morgan LM. Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite. Br J Nutr. 2003;89(2):239–48. [DOI] [PubMed] [Google Scholar]
- 77. Bendtsen LQ, Lorenzen JK, Gomes S, Liaset B, Holst JJ, Ritz C, Reitelseder S, Sjödin A, Astrup A. Effects of hydrolysed casein, intact casein and intact whey protein on energy expenditure and appetite regulation: a randomised, controlled, cross-over study. Adv Nutr. 2014;112(8):1412–22. [DOI] [PubMed] [Google Scholar]
- 78. Calbet JA, Holst JJ. Gastric emptying, gastric secretion and enterogastrone response after administration of milk proteins or their peptide hydrolysates in humans. Eur J Nutr. 2004;43(3):127–39. [DOI] [PubMed] [Google Scholar]
- 79. Bowen J, Noakes M, Clifton PM. Appetite regulatory hormone responses to various dietary proteins differ by body mass index status despite similar reductions in ad libitum energy intake. J Clin Endocrinol Metab. 2006;91(8):2913–9. [DOI] [PubMed] [Google Scholar]
- 80. Holmer-Jensen J, Mortensen LS, Astrup A, de Vrese M, Holst JJ, Thomsen C, Hermansen K. Acute differential effects of dietary protein quality on postprandial lipemia in obese non-diabetic subjects. Nutr Res. 2013;33(1):34–40. [DOI] [PubMed] [Google Scholar]
- 81. Mortensen LS, Hartvigsen ML, Brader LJ, Astrup A, Schrezenmeir J, Holst JJ, Thomsen C, Hermansen K. Differential effects of protein quality on postprandial lipemia in response to a fat-rich meal in type 2 diabetes: comparison of whey, casein, gluten, and cod protein. Am J Clin Nutr. 2009;90(1):41–8. [DOI] [PubMed] [Google Scholar]
- 82. Samocha-Bonet D, Chisholm DJ, Holst JJ, Greenfield JR. L-glutamine and whole protein restore first-phase insulin response and increase glucagon-like peptide-1 in type 2 diabetes patients. Nutrients. 2015;7(4):2101–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Reimann F, Williams L, da S, Xavier G, Rutter GA, Gribble FM. Glutamine potently stimulates glucagon-like peptide-1 secretion from GLUTag cells. Diabetologia. 2004;47(9):1592–601. [DOI] [PubMed] [Google Scholar]
- 84. Alamshah A, Spreckley E, Norton M, Kinsey-Jones JS, Amin A, Ramgulam A, Cao Y, Johnson R, Saleh K, Akalestou Eet al. l-Phenylalanine modulates gut hormone release and glucose tolerance, and suppresses food intake through the calcium-sensing receptor in rodents. Int J Obes. 2017;41(11):1693–701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Amin A, Neophytou C, Thein S, Martin NM, Alamshah A, Spreckley E, Bloom SR, Murphy KG. l-Arginine increases postprandial circulating GLP-1 and PYY levels in humans. Obesity. 2018;26(11):1721–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Clemmensen C, Smajilovic S, Smith EP, Woods SC, Brauner-Osborne H, Seeley RJ, D'Alessio DA, Ryan KK. Oral L-arginine stimulates GLP-1 secretion to improve glucose tolerance in male mice. Endocrinology. 2013;154(11):3978–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Alamshah A, McGavigan AK, Spreckley E, Kinsey-Jones JS, Amin A, Tough IR, O'Hara HC, Moolla A, Banks K, France Ret al. l-Arginine promotes gut hormone release and reduces food intake in rodents. Diabetes Obes Metab. 2016;18(5):508–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Acar I, Cetinkaya A, Lay I, Ileri-Gurel E. The role of calcium sensing receptors in GLP-1 and PYY secretion after acute intraduodenal administration of L-tryptophan in rats. Nutr Neurosci. 2020;23:481–9. [DOI] [PubMed] [Google Scholar]
- 89. Amin A, Frampton J, Liu Z, Franco-Becker G, Norton M, Alaa A, Li JV, Murphy KG. Differential effects of L- and D-phenylalanine on pancreatic and gastrointestinal hormone release in humans: a randomized crossover study. Diabetes Obes Metab. 2021;23(1):147–57. [DOI] [PubMed] [Google Scholar]
- 90. Steinert RE, Ullrich SS, Geary N, Asarian L, Bueter M, Horowitz M, Feinle-Bisset C. Comparative effects of intraduodenal amino acid infusions on food intake and gut hormone release in healthy males. Physiol Rep. 2017;5(21):e13492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Modvig IM, Kuhre RE, Jepsen SL, Xu S, Engelstoft MS, Egerod KL, Schwartz TW, Ørskov C, Rosenkilde MM, Holst JJ. Amino acids differ in their capacity to stimulate GLP-1 release from the perfused rat small intestine and stimulate secretion by different sensing mechanisms. Am J Physiol Endocrinol Metab. 2021;320:E874–85. [DOI] [PubMed] [Google Scholar]
- 92. Tidwell DK, Valliant MW. Higher amounts of body fat are associated with inadequate intakes of calcium and vitamin D in African American women. Nutr Res. 2011;31(7):527–36. [DOI] [PubMed] [Google Scholar]
- 93. Zemel MB, Shi H, Greer B, Dirienzo D, Zemel PC. Regulation of adiposity by dietary calcium. FASEB J. 2000;14(9):1132–8. [PubMed] [Google Scholar]
- 94. Mirmiran P, Esmaillzadeh A, Azizi F. Dairy consumption and body mass index: an inverse relationship. Int J Obes. 2005;29(1):115–21. [DOI] [PubMed] [Google Scholar]
- 95. Zemel MB, Thompson W, Milstead A, Morris K, Campbell P. Calcium and dairy acceleration of weight and fat loss during energy restriction in obese adults. Obes Res. 2004;12(4):582–90. [DOI] [PubMed] [Google Scholar]
- 96. Onakpoya IJ, Perry R, Zhang J, Ernst E. Efficacy of calcium supplementation for management of overweight and obesity: systematic review of randomized clinical trials. Nutr Rev. 2011;69(6):335–43. [DOI] [PubMed] [Google Scholar]
- 97. Christensen R, Lorenzen JK, Svith CR, Bartels EM, Melanson EL, Saris WH, Tremblay A, Astrup A. Effect of calcium from dairy and dietary supplements on faecal fat excretion: a meta-analysis of randomized controlled trials. Obes Rev. 2009;10(4):475–86. [DOI] [PubMed] [Google Scholar]
- 98. Gonzalez JT, Stevenson EJ. New perspectives on nutritional interventions to augment lipid utilisation during exercise. Br J Nutr. 2012;107(3):339–49. [DOI] [PubMed] [Google Scholar]
- 99. Ping-Delfos WC, Soares M. Diet induced thermogenesis, fat oxidation and food intake following sequential meals: influence of calcium and vitamin D. Clin Nutr. 2011;30(3):376–83. [DOI] [PubMed] [Google Scholar]
- 100. Gilbert JA, Joanisse DR, Chaput JP, Miegueu P, Cianflone K, Almeras N, Tremblay A. Milk supplementation facilitates appetite control in obese women during weight loss: a randomised, single-blind, placebo-controlled trial. Br J Nutr. 2011;105(1):133–43. [DOI] [PubMed] [Google Scholar]
- 101. Jeukendrup AE, Wallis GA. Measurement of substrate oxidation during exercise by means of gas exchange measurements. Int J Sports Med. 2005;26(Suppl 1):S28–37. [DOI] [PubMed] [Google Scholar]
- 102. Gonzalez JT, Rumbold PL, Stevenson EJ. Appetite sensations and substrate metabolism at rest, during exercise, and recovery: impact of a high-calcium meal. Appl Physiol Nutr Metab. 2013;38(12):1260–7. [DOI] [PubMed] [Google Scholar]
- 103. Gonzalez JT, Stevenson EJ. Calcium co-ingestion augments postprandial glucose-dependent insulinotropic peptide 1–42, glucagon-like peptide-1 and insulin concentrations in humans. Eur J Nutr. 2014;53(2):375–85. [DOI] [PubMed] [Google Scholar]
- 104. Lorenzen JK, Nielsen S, Holst JJ, Tetens I, Rehfeld JF, Astrup A. Effect of dairy calcium or supplementary calcium intake on postprandial fat metabolism, appetite, and subsequent energy intake. Am J Clin Nutr. 2007;85(3):678–87. [DOI] [PubMed] [Google Scholar]
- 105. Trautvetter U, Jahreis G. Effect of supplementary calcium phosphate on plasma gastrointestinal hormones in a double-blind, placebo-controlled, cross-over human study. Br J Nutr. 2014;111(2):287–93. [DOI] [PubMed] [Google Scholar]
- 106. Jones KW, Eller LK, Parnell JA, Doyle-Baker PK, Edwards AL, Reimer RA. Effect of a dairy- and calcium-rich diet on weight loss and appetite during energy restriction in overweight and obese adults: a randomized trial. Eur J Clin Nutr. 2013;67(4):371–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Kondrashina A, Seratlic S, Kandil D, Treguier N, Kilcawley K, Schellekens H, Beresford T, Giblin L. Irish Cheddar cheese increases glucagon-like peptide-1 secretion in vitro but bioactivity is lost during gut transit. Food Chem. 2018;265:9–17. [DOI] [PubMed] [Google Scholar]
- 108. Nilsson M, Stenberg M, Frid AH, Holst JJ, Bjorck IM. Glycemia and insulinemia in healthy subjects after lactose-equivalent meals of milk and other food proteins: the role of plasma amino acids and incretins. Am J Clin Nutr. 2004;80(5):1246–53. [DOI] [PubMed] [Google Scholar]
- 109. Gonzalez JT, Green BP, Brown MA, Rumbold PL, Turner LA, Stevenson EJ. Calcium ingestion suppresses appetite and produces acute overcompensation of energy intake independent of protein in healthy adults. J Nutr. 2015;145(3):476–82. [DOI] [PubMed] [Google Scholar]
- 110. Chen Y-C, Edinburgh RM, Hengist A, Smith HA, Walhin J-P, Betts JA, Thompson D, Gonzalez JT. Venous blood provides lower glucagon-like peptide-1 concentrations than arterialized blood in the postprandial but not the fasted state: consequences of sampling methods. Exp Physiol. 2018;103(9):1200–5. [DOI] [PubMed] [Google Scholar]
- 111. Cummings DE, Overduin J. Gastrointestinal regulation of food intake. J Clin Invest. 2007;117(1):13–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. D'Alessio D, Lu W, Sun W, Zheng S, Yang Q, Seeley R, Woods SC, Tso P. Fasting and postprandial concentrations of GLP-1 in intestinal lymph and portal plasma: evidence for selective release of GLP-1 in the lymph system. Am J Physiol Regul Integr Comp Physiol. 2007;293(6):R2163–9. [DOI] [PubMed] [Google Scholar]
- 113. Oya M, Kitaguchi T, Pais R, Reimann F, Gribble F, Tsuboi T. The G protein-coupled receptor family C group 6 subtype A (GPRC6A) receptor is involved in amino acid-induced glucagon-like peptide-1 secretion from GLUTag cells. J Biol Chem. 2013;288(7):4513–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Rudenko O, Shang J, Munk A, Ekberg JP, Petersen N, Engelstoft MS, Egerod KL, Hjorth SA, Wu M, Feng Yet al. The aromatic amino acid sensor GPR142 controls metabolism through balanced regulation of pancreatic and gut hormones. Mol Metab. 2019;19:49–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Jang H-J, Kokrashvili Z, Theodorakis MJ, Carlson OD, Kim B-J, Zhou J, Kim HH, Xu X, Chan SL, Juhaszova Met al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc Natl Acad Sci U S A. 2007;104(38):15069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Choi S, Lee M, Shiu AL, Yo SJ, Aponte GW. Identification of a protein hydrolysate responsive G protein-coupled receptor in enterocytes. Am J Physiol Gastrointest Liver Physiol. 2007;292(1):G98–G.112. [DOI] [PubMed] [Google Scholar]
- 117. Diakogiannaki PR, Tolhurst G, Parker HE, Horscroft J, Rauscher B, Zietek T, Daniel H, Gribble FM, Reimann F. Oligopeptides stimulate glucagon-like peptide-1 secretion in mice through proton-coupled uptake and the calcium-sensing receptor. Diabetologia. 2013;56(12):2688–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Norton M, Murphy KG. Targeting gastrointestinal nutrient sensing mechanisms to treat obesity. Curr Opin Pharmacol. 2017;37:16–23. [DOI] [PubMed] [Google Scholar]
- 119. Wang M, Yao Y, Kuang D, Hampson DR. Activation of family C G-protein-coupled receptors by the tripeptide glutathione. J Biol Chem. 2006;281(13):8864–70. [DOI] [PubMed] [Google Scholar]
- 120. Conigrave AD, Quinn SJ, Brown EM. l-Amino acid sensing by the extracellular Ca2+-sensing receptor. Proc Natl Acad Sci U S A. 2000;97(9):4814–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Conigrave AD, Franks AH, Brown EM, Quinn SJ. l-Amino acid sensing by the calcium-sensing receptor: a general mechanism for coupling protein and calcium metabolism?. Eur J Clin Nutr. 2002;56(11):1072–80. [DOI] [PubMed] [Google Scholar]
- 122. Ojha U. Protein-induced satiation and the calcium-sensing receptor. Diabetes Metab Syndr Obes. 2018;11:45–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Wellendorph P, Brauner-Osborne H. Molecular basis for amino acid sensing by family C G-protein-coupled receptors. Br J Pharmacol. 2009;156(6):869–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Sheinin Y, Kallay E, Wrba F, Kriwanek S, Peterlik M, Cross HS. Immunocytochemical localization of the extracellular calcium-sensing receptor in normal and malignant human large intestinal mucosa. J Histochem Cytochem. 2000;48(5):595–602. [DOI] [PubMed] [Google Scholar]
- 125. Chattopadhyay N, Cheng I, Rogers K, Riccardi D, Hall A, Diaz R, Hebert SC, Soybel DI, Brown EM. Identification and localization of extracellular Ca(2+)-sensing receptor in rat intestine. Am J Physiol. 1998;274(1):G122–30. [DOI] [PubMed] [Google Scholar]
- 126. Cheng SX, Okuda M, Hall AE, Geibel JP, Hebert SC. Expression of calcium-sensing receptor in rat colonic epithelium: evidence for modulation of fluid secretion. Am J Physiol Gastrointest Liver Physiol. 2002;283(1):G240–50. [DOI] [PubMed] [Google Scholar]
- 127. Modvig IM, Kuhre RE, Holst JJ. Peptone-mediated glucagon-like peptide-1 secretion depends on intestinal absorption and activation of basolaterally located calcium-sensing receptors. Physiol Rep. 2019;7(8):e14056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Diakogiannaki E, Gribble FM, Reimann F. Nutrient detection by incretin hormone secreting cells. Physiol Behav. 2012;106(3):387–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Grant MP, Stepanchick A, Cavanaugh A, Breitwieser GE. Agonist-driven maturation and plasma membrane insertion of calcium-sensing receptors dynamically control signal amplitude. Sci Signal. 2011;4(200):ra78. [DOI] [PubMed] [Google Scholar]
- 130. Conigrave AD, Ward DT. Calcium-sensing receptor (CaSR): pharmacological properties and signaling pathways. Best Pract Res Clin Endocrinol Metab. 2013;27(3):315–31. [DOI] [PubMed] [Google Scholar]
- 131. Conigrave AD, Hampson DR. Broad-spectrum L-amino acid sensing by class 3 G-protein-coupled receptors. Trends Endocrinol Metab. 2006;17(10):398–407. [DOI] [PubMed] [Google Scholar]
- 132. Conigrave AD, Hampson DR. Broad-spectrum amino acid-sensing class C G-protein coupled receptors: molecular mechanisms, physiological significance and options for drug development. Pharmacol Ther. 2010;127(3):252–60. [DOI] [PubMed] [Google Scholar]
- 133. Pi M, Nishimoto SK, Quarles LD. GPRC6A: jack of all metabolism (or master of none). Mol Metab. 2017;6(2):185–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Wellendorph P, Hansen KB, Balsgaard A, Greenwood JR, Egebjerg J, Brauner-Osborne H. Deorphanization of GPRC6A: a promiscuous L-alpha-amino acid receptor with preference for basic amino acids. Mol Pharmacol. 2005;67(3):589–97. [DOI] [PubMed] [Google Scholar]
- 135. Pi M, Faber P, Ekema G, Jackson PD, Ting A, Wang N, Fontilla-Poole M, Mays RW, Brunden KR, Harrington JJet al. Identification of a novel extracellular cation-sensing G-protein-coupled receptor. J Biol Chem. 2005;280(48):40201–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Pi-Sunyer X, Astrup A, Fujioka K, Greenway F, Halpern A, Krempf M, Lau DC, le Roux CW, Violante Ortiz R, Jensen CBet al. A randomized, controlled trial of 3.0 mg of liraglutide in weight management. N Engl J Med. 2015;373(1):11–22. [DOI] [PubMed] [Google Scholar]
- 137. Kuang D, Yao Y, Lam J, Tsushima RG, Hampson DR. Cloning and characterization of a family C orphan G-protein coupled receptor. J Neurochem. 2005;93(2):383–91. [DOI] [PubMed] [Google Scholar]
- 138. Clemmensen C, Jorgensen CV, Smajilovic S, Brauner-Osborne H. Robust GLP-1 secretion by basic L-amino acids does not require the GPRC6A receptor. Diabetes Obes Metab. 2017;19(4):599–603. [DOI] [PubMed] [Google Scholar]
- 139. Pi M, Nishimoto SK, Darryl Quarles L. Explaining divergent observations regarding osteocalcin/GPRC6A endocrine signaling. Endocrinology. 2021;162(4):bqab011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Pi M, Chen L, Huang MZ, Zhu W, Ringhofer B, Luo J, Christenson L, Li B, Zhang J, Jackson PDet al. GPRC6A null mice exhibit osteopenia, feminization and metabolic syndrome. PLoS One. 2008;3(12):e3858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Wellendorph P, Johansen LD, Jensen AA, Casanova E, Gassmann M, Deprez P, Clément-Lacroix P, Bettler B, Bräuner-Osborne H. No evidence for a bone phenotype in GPRC6A knockout mice under normal physiological conditions. J Mol Endocrinol. 2009;42(3):215–23. [DOI] [PubMed] [Google Scholar]
- 142. Tolhurst G, Reimann F, Gribble FM. Intestinal sensing of nutrients. Handb Exp Pharmacol. 2012(209):309–35. [DOI] [PubMed] [Google Scholar]
- 143. Simpson AK, Ward PS, Wong KY, Collord GJ, Habib AM, Reimann F, Gribble FM. Cyclic AMP triggers glucagon-like peptide-1 secretion from the GLUTag enteroendocrine cell line. Diabetologia. 2007;50(10):2181–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. Adriaenssens AE, Reimann F, Gribble FM. Distribution and stimulus secretion coupling of enteroendocrine cells along the intestinal tract. Compr Physiol. 2018;8(4):1603–38. [DOI] [PubMed] [Google Scholar]
- 145. Rey O, Young SH, Jacamo R, Moyer MP, Rozengurt E. Extracellular calcium sensing receptor stimulation in human colonic epithelial cells induces intracellular calcium oscillations and proliferation inhibition. J Cell Physiol. 2010;225(1):73–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Wauson EM, Lorente-Rodriguez A, Cobb MH. Minireview: nutrient sensing by G protein-coupled receptors. Mol Endocrinol. 2013;27(8):1188–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Adibi SA. The oligopeptide transporter (Pept-1) in human intestine: biology and function. Gastroenterology. 1997;113(1):332–40. [DOI] [PubMed] [Google Scholar]
- 148. Daniel H. Molecular and integrative physiology of intestinal peptide transport. Annu Rev Physiol. 2004;66:361–84. [DOI] [PubMed] [Google Scholar]
- 149. Matsumura K, Miki T, Jhomori T, Gonoi T, Seino S. Possible role of PEPT1 in gastrointestinal hormone secretion. Biochem Biophys Res Commun. 2005;336(4):1028–32. [DOI] [PubMed] [Google Scholar]
- 150. Peacock M. Calcium metabolism in health and disease. Clin J Am Soc Nephrol. 2010;5(Suppl 1):S23. [DOI] [PubMed] [Google Scholar]
- 151. Cheng H, Leblond CP. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine. III. Entero-endocrine cells. Am J Anat. 1974;141(4):503–19. [DOI] [PubMed] [Google Scholar]