

Abstract
Retinal damage in the adult zebrafish induces Müller glia reprogramming to produce neuronal progenitor cells that proliferate and differentiate into retinal neurons. Notch signaling, which is a fundamental mechanism known to drive cell-cell communication, is required to maintain Müller glia in a quiescent state in the undamaged retina, and repression of Notch signaling is necessary for Müller glia to reenter the cell cycle. The dynamic regulation of Notch signaling following retinal damage also directs proliferation and neurogenesis of the Müller glia-derived progenitor cells in a robust regeneration response. In contrast, mammalian Müller glia respond to retinal damage by entering a prolonged gliotic state that leads to additional neuronal death and permanent vision loss. Understanding the dynamic regulation of Notch signaling in the zebrafish retina may aid efforts to stimulate Müller glia reprogramming for regeneration of the diseased human retina. Recent findings identified DeltaB and Notch3 as the ligand-receptor pair that serves as the principal regulators of zebrafish Müller glia quiescence. In addition, multiomics datasets and functional studies indicate that additional Notch receptors, ligands, and target genes regulate cell proliferation and neurogenesis during the regeneration time course. Still, our understanding of Notch signaling during retinal regeneration is limited. To fully appreciate the complex regulation of Notch signaling that is required for successful retinal regeneration, investigation of additional aspects of the pathway, such as post-translational modification of the receptors, ligand endocytosis, and interactions with other fundamental pathways is needed. Here we review various modes of Notch signaling regulation in the context of the vertebrate retina to put recent research in perspective and to identify open areas of inquiry.
Key Words: differentiation, gliosis, Müller glia, neuronal progenitor cell, Notch signaling, proliferation, quiescence, retinal development, retinal regeneration, zebrafish
Introduction
The vertebrate retina is a highly organized neural tissue consisting of six major types of neurons (rod and cone photoreceptors, horizontal cells, bipolar cells, amacrine cells, and ganglion cells) and three major classes of non-neuronal support cells (retinal pigment epithelial cells, Müller glia, and microglia). Retinal cell bodies are arranged in three distinct nuclear layers (outer nuclear layer, inner nuclear layer, and the ganglion cell layer) that are separated by two plexiform layers (outer plexiform layer and inner plexiform layer), which consist of the axonal and dendritic processes that form synaptic connections. This laminated structure results in efficient detection and pre-processing of sensory information that relies on the intimate cell-cell contacts between neurons, as well as between neurons and Müller glia, which represent the radial glia in the retina (Amini et al., 2017). Retinal damage leads to compromised neuronal function, cell death, disrupted cell-cell contacts, and impaired vision (Hoon et al., 2014). In response to damage, the mammalian retina enters a state of reactive gliosis, in which Müller glia hypertrophy disrupts retinal homeostasis. This leads to secondary neuronal cell death, glial scarring, and vision loss (Bringmann and Wiedemann, 2012). Vision impairment, which affects an estimated 2.2 billion people worldwide (World Health Organization, 2019), often negatively affects quality of life beyond vision loss because of increased economic burden and mental health problems, including depression and loneliness, especially in the aging population (Gordois et al., 2012; Demmin and Silverstein, 2020). Therefore, there is great need for strategies that will heal the damaged retina and restore visual function.
The ultimate strategy is to replace the missing neurons in the damaged retina and restore a healthy, functioning retina. Zebrafish (Danio rerio) possess the ability to regenerate lost neurons from an endogenous source, the Müller glia (Figure 1A). Unlike in mammals, adult zebrafish Müller glia respond to retinal damage by transitioning to a transient gliotic state before entering a reprogrammed state (Thomas et al., 2016; Hoang et al., 2020). A reprogrammed Müller glial cell divides asymmetrically to maintain the Müller glia and to produce a neuronal progenitor cell (NPC) (Nagashima et al., 2013; Lahne et al., 2015). The NPC continues to proliferate to produce a cluster of multipotent progenitors that differentiate into all retinal cell types with a bias towards the cells lost to damage (Powell et al., 2016; Lahne et al., 2021). The regenerated neurons develop normal morphologies, form stereotypical connections, and restore circuitry for visual function (D’Orazi et al., 2016; McGinn et al., 2018, 2019).
Figure 1.
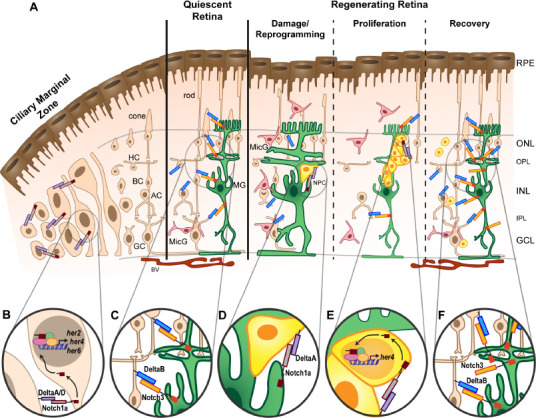
Dynamic Notch signaling during retinal regeneration.
(A) Schematic of the adult zebrafish retina. The retina is organized into three nuclear layers: ganglion cell layer (GCL), inner nuclear layer (INL), and outer nuclear layer (ONL), which are separated by the inner plexiform layer (IPL) and outer plexiform layer (OPL) where synaptic connections are located. Müller glia (MG) cell bodies reside in the INL, but their projections extend throughout all three nuclear layers. MG are mostly quiescent in the intact retina, except for occasional proliferation to give rise to rod photoreceptor progenitors. In the intact, quiescent retina, resting microglia (MicG, pink) reside in the plexiform layers with ramified morphology. The inner, basal surface of the retina is vascularized (blood vessels, BV), and the outer, apical surface is lined with the retinal pigment epithelium (RPE). The ciliary marginal zone (CMZ) at the periphery of the retina is an area of persistent neurogenesis with proliferative multipotent retinal progenitors that give rise to ganglion cells (GC), amacrine cells (AC), bipolar cells (BC), horizontal cells (HC), and cone photoreceptors. (B) The CMZ is marked by expression of notch1a and notch1b receptor genes, deltaA and deltaD ligand genes, and downstream target genes her2, her4, and her6 (see text). (C) MG express Notch3, which when activated with the DeltaB ligand, maintains MG quiescence. Upon damage, neurons degenerate, and MG hypertrophy, followed by reprogramming and asymmetric cell division to retain a MG and generate a neuronal progenitor cell (NPC). MicG migrate to areas of damage and adopt an amoeboid morphology. (D) Notch3 expression is downregulated in the proliferating retina, but other Notch signaling components are necessary for continued proliferation of the NPCs, in particular Notch1a and DeltaA. (E) Continued NPC proliferation produces a cluster of NPCs in the regenerating retina. This process requires the Notch downstream target, her4. (F) The NPCs migrate to the areas of damage and differentiate into the lost neurons. Notch3 expression is reestablished in the quiescent MG.
A complex network of signaling pathways is necessary as retinal damage breaks cell-cell contacts and new contacts are formed through remodeling, proliferation, and differentiation. Notch signaling is an obvious candidate as a major regulator of retinal regeneration because of its well-characterized role in retinal progenitor cell maintenance and Müller glia specification during development (Furukawa et al., 2000; Jadhav et al., 2006a, b). Additionally, Notch signaling relies on contact-mediated interactions between neighboring cells, which means the loss of cell contacts during retinal damage and the generation of different cell contacts as NPCs are generated, proliferate, and commit to neuronal fates are opportunities for Notch involvement. Notch signaling is activated when the Notch receptor expressed on the signal-receiving cell binds a ligand expressed on the signal-sending cell. A series of proteolytic cleavages are induced to release the Notch intracellular domain (NICD) for translocation into the nucleus of the signal-receiving cell where it effects gene expression changes (Henrique and Schweisguth, 2019). The outcomes of Notch signaling are highly context dependent with respect to specific tissue and to time, so despite the absence of signaling intermediates or second messenger amplification, there are abundant modes of Notch signaling regulation. These include post-translational modification during receptor production, ligand-receptor expression patterning, transendocytosis within the signal-sending cell, receptor cleavage events, NICD-dependent regulation of target gene transcription, and interactions with other signaling pathways (Figure 2).
Figure 2.
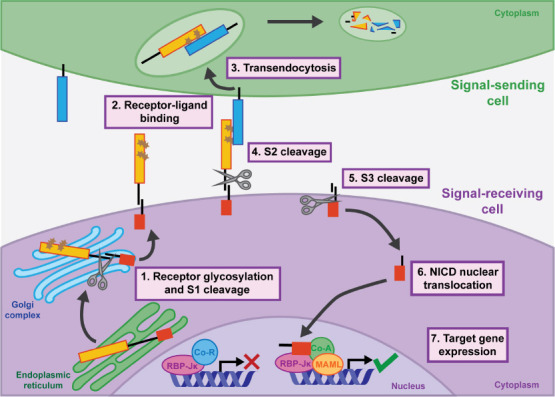
Core components of the Notch signaling pathway.
Schematic of the Notch signaling pathway highlighting points of possible regulation during retinal regeneration. 1: Notch receptors are post-translationally modified by glycosylation and Furin-dependent S1 cleavage. 2: Notch receptors on the signal-receiving cell bind ligands on the signalsending cell. 3: Transendocytosis pulls the ligand-NECD complex into the signal-sending cell. 4: S2 site is available for metalloprotease-dependent cleavage. 5: γ-Secretase-dependent S3 cleavage releases NICD from the membrane. 6: NICD translocates into the nucleus of the signal-receiving cell and forms a complex with CSL transcription factor RBP-Jκ, MAML, and co-activators. 7: Notch binding of the chromatin ultimately induces expression changes of NICD target genes. Co-A: Co-activator protein; Co-R: co-repressor protein; CSL: (CBF1/RBP-Jκ, Su(H), LAG-1); MAML: Mastermind-like; NECD: Notch extracellular domain; NICD: Notch intracellular domain; S1: cleavage site 1; S2: cleavage site 2; S3: cleavage site 3.
Within zebrafish retinal regeneration, we largely understand the roles of Notch signaling through the use of γ-secretase inhibitors that prevent NICD cleavage and nuclear translocation (Conner et al., 2014), a Notch reporter line that allows for visualization of NICD activity (Wan and Goldman, 2017), and a Notch1a NICD transgenic line that can overexpress NICD via a heat shock promoter (Wan et al., 2012). Recent results have identified DeltaB and Notch3 as the ligand-receptor pair that maintains Müller glia quiescence (Campbell et al., 2021) and have linked Notch signaling to the reprogramming factors Ascl1a and Lin28a (Elsaeidi et al., 2018), the epigenetic factor Hdac1 (Mitra et al., 2018), and the Tgfβ signaling pathway (Lee et al., 2020; Sharma et al., 2020). Here we aim to review these recent findings in the context of Notch signaling regulation, but also review the potential modes of regulation that have not yet been explored in zebrafish retinal regeneration (Table 1). Understanding these regulatory mechanisms will guide future inquiry into the mechanism by which the zebrafish retina responds to disruptions in cell-cell contact to elicit the regenerative response.
Table 1.
Zebrafish and mammalian orthologs of the core components in Notch signaling
| Component | Mammals | Danio rerio (Zebrafish) |
|---|---|---|
| Receptors | ||
| NOTCH1 | Notch1a, Notch1b | |
| NOTCH2 | Notch2 | |
| NOTCH3 | Notch3 | |
| NOTCH4 | ||
| Receptor proteases | ||
| S1 cleavage (furin-dependent) | FURIN | Furina, Furinb |
| S2 cleavage (metalloprotease-dependent) | ADAM10 | Adam10a, Adam10b |
| S3 cleavage (γ-secretase dependent) | Presenilin (PSEN1, PSEN2) | Psen1, Psen2 |
| Nicastrin (NCSTN) | Ncstn | |
| APH1 | Aph1b | |
| Presenilin enhancer (PSENEN) | Psenen | |
| Glycosyltransferase modifiers | ||
| O-glucosyl-transferase | POGLUT1 | Poglut1 |
| POGLUT2 | Poglut2 | |
| POGLUT3 | Poglut3 | |
| O-fucosyl-transferase | POFUT1 | Pofut1 |
| EGF domain-specific O-GlcNAc transferase | EOGT | Eogt |
| β1,3-GlcNAc transferase | Lunatic Fringe (LFNG) | Lfng |
| Manic Fringe (MFNG) | Mfng | |
| Radical Fringe (RFNG) | Rfng | |
| Ligands | ||
| DLL1 | DeltaA, DeltaD | |
| DLL3 | DeltaB, DeltaC | |
| DLL4 | Dll4 | |
| Jagged1 (JAG1) | Jag1a, Jag1b | |
| Jagged2 (JAG2) | Jag2a, Jag2b | |
| Endosomal sorting/membrane trafficking regulators | ||
| Ring finger E3 ubiquitin ligase (ligand endocytosis) | Mindbomb (MIB1), Skeletrophin (MIB2) | Mindbomb (Mib1 and Mib2) |
| Ring finger E3 ubiquitin ligase (receptor endocytosis) | Deltex (DTX1, DTX2, DTX3, DTX4) | Dtx1, Dtx2, Dtx3, Dtx4a, Dtx4b |
| HECT domain E3 ubiquitin ligase (receptor endocytosis) | ITCH | Itcha, Itchb, Nedd4 |
| Negative regulators | Numb, Numb-like | Numb, Numbl |
| Nuclear effectors | ||
| CSL DNA-binding transcription factor | RBPJ | Rbpja, Rbpjb |
| Transcriptional coactivator | Mastermind-like (MAML1, MAML2, MAML3) | Maml1, Maml2, Maml3 |
| Transcriptional corepressors | NCOR1, NCOR2 (SMRT) | Ncor1, Ncor2 |
| SPEN (SHARP/MINT) | Spen | |
| Target genes | ||
| bHLH repressors | Hairy/Enhancer of Split and related genes (HES/HEY) | her and hey genes |
| Hes1 | her6 | |
| Hes5 | her2, her4 |
Search Strategy and Selection Criteria
Literature cited in this review was published from 1997 to 2021 and was searched on PubMed and Google Scholar using combinations of keywords and citation tracking on PubMed. Search terms included: Notch signaling, receptor, ligand, target genes, retina, development, regeneration, zebrafish, Müller glia, and stem cells. Literature was also searched through ZFIN within each of the gene pages for the zebrafish factors listed in Table 1.
Elements of Notch Signaling in Context with Zebrafish Retinal Regeneration
Notch receptor synthesis and modification
Notch receptors are a highly conserved family of single-pass transmembrane proteins. The zebrafish genome contains four Notch receptor genes: notch1a, notch1b, notch2, and notch3. Each gene encodes a receptor that is processed into a heterodimer following ligand-independent, Furin-dependent S1 cleavage during transit through the Golgi network (Logeat et al., 1998). Zebrafish possess two Furin paralogs, furina and furinb. Following S1 cleavage, the resulting Notch extracellular domain (NECD) dimerizes with the Notch transmembrane and intracellular domain (TMIC) through noncovalent interactions. The NECD is the ligand-binding domain and consists of 29–36 epidermal growth factor (EGF)-like repeats, a negative regulatory region, and cysteine-rich LIN-12/Notch repeats. The TMIC contains the NICD, which consists of a RAM domain, ankyrin repeats flanked by two nuclear localization signals, a transcriptional activation domain, and a C-terminal PEST domain (Kopan and Ilagan, 2009). The S1 cleavage event is necessary for biological activity of the Notch receptors (Lake et al., 2009), and therefore has the potential to serve as a regulatory point for Notch signaling in the signal-receiving cell. Following ligand binding, two additional proteolytic cleavage events must occur at the S2 and S3 cleavage sites in Notch to generate the NICD (Steinbuck and Winandy, 2018).
Notch receptor activity is highly regulated by glycosylation. Prior to the S1 cleavage event, O-linked glycan modifications can be added to each EGF-like repeat within the NECD (Saiki et al., 2021). O-glucosyltransferases add O-glucose to the serine residues within an EGF-like repeat. In mammals this is carried out by POGLUT1 (Rumi in flies) between the first two cysteines of an EGF-like repeat (consensus sequence C1-X-S-X-(P/A)-C2) and by POGLUT2 and POGLUT3 within the ligand-binding domains of NOTCH1 and NOTCH3 (consensus sequence C3-X-N-T-X-G-S-F-X-C4; Takeuchi et al., 2018). O-fucosyltransferases add O-fucose to the serine or threonine residue between the second and third cysteines (consensus sequence C2-X-X-X-X-(S/T)-C3) (Moloney et al., 2000), and in mammals, this is achieved with POFUT1 (Shi and Stanley, 2003). O-linked-β-N-acetylglucosamine modification is added to the serine or threonine residue between the fifth and sixth cysteines (consensus sequence C5-X-X-G-X-(S/T)-G-X-X-C6) by the EGF domain-specific O-linked N-acetylglucosamine (GlcNAc) transferase EOGT in mammals (Sakaidani et al., 2012). While they do exist, little research has been reported on Notch signaling regulation with the orthologs of these enzymes in zebrafish.
Further fine-tuning of the O-fucose modifications leads to another layer of Notch regulation. This is achieved by the Fringe proteins, which catalyze the addition of GlcNAc to O-fucose (Rana and Haltiwanger, 2011). In mammals and zebrafish, this includes Lunatic fringe (Lfng), Manic fringe (Mfng), and Radical fringe (Rfng). In Drosophila, where there is a single Fringe protein, classic studies demonstrated that Fringe is restricted to the ventral half of the developing eye (Cho and Choi, 1998), where it inhibits Notch activity with the Serrate ligand and enhances Notch activity with the Delta ligand, thus creating a dorsoventral boundary that directs eye morphogenesis (Panin et al., 1997).
In the developing mouse retina, Lfng influences angiogenic sprouting by enhancing Dll4 ligand binding and reducing Jag1 ligand binding to Notch (Benedito et al., 2009). In zebrafish, Lfng maintains progenitor cells in the developing hindbrain by promoting lateral inhibition of neurogenesis through closely regulated Notch activation (Nikolaou et al., 2009), and in both heart and fin regeneration, Lfng is upregulated to increase Notch activation (Grotek et al., 2013; Münch et al., 2017). Recent single-cell RNA-seq data suggest that lfng is expressed in both activated and resting Müller glia in the zebrafish retina, while mfng is expressed in the retinal immune cells, the microglia (Hoang et al., 2020), which have been shown to play a role in regulating the Müller glia-derived regeneration response (Lahne et al., 2020; Iribarne, 2021). Investigations of the Fringe proteins in the zebrafish retina may help elucidate ligand-receptor relationships and the dynamic role of Notch signaling during regeneration.
Ligand-receptor interactions
The Notch ligands belong to the DSL (Delta/Serrate/Lag) family of transmembrane proteins (D’Souza et al., 2008). Zebrafish have five Delta-like ligands: DeltaA, DeltaB, DeltaC, DeltaD, and Delta-like4 (Dll4), as well as four Jagged ligands: Jag1a, Jag1b, Jag2a, and Jag2b. The ligands all have an extracellular domain composed of several EGF-like repeats and a cysteine-rich N-terminal DSL domain that is required for interaction with Notch. Similar to the NECD, the EGF-like repeats of DSL ligands can be glycosylated, however it is unclear if these types of modifications affect ligand activity (Panin et al., 2002). Instead, Notch ligands are regulated by expression pattern, ubiquitylation, and endocytosis.
Notch receptor and ligand expression patterns influence the Notch regulation of cell fate by lateral specification (or lateral inhibition), which is the mechanism by which the fate of one cell influences the fate of its neighboring cells (Bray, 2016). This is accomplished through a negative feedback loop where the ligand-expressing cell induces the receptor-expressing cell to express Notch target genes, which are transcriptional repressors of the ligand genes, thereby repressing ligand expression in the signal-receiving cell. Within the damaged and regenerating retina, the mechanism of lateral specification may indicate to surviving cells, in particular the Müller glia, the state of the neighboring cells, as well as direct the fate of Müller glia-derived progenitor cells. Notch3 protein, which is expressed in Müller glia of the undamaged retina, is downregulated as Müller glia proliferate, and is reestablished in Müller glia of the recovering retina (Campbell et al., 2021). Alternatively, deltab mRNA is expressed throughout the undamaged retina and more highly in neurons than in the Müller glia, suggesting a complementary pattern to notch3 expression (Campbell et al., 2021). DeltaB-Notch3 interaction is required to maintain the Müller glia in a quiescent, non-proliferative state in the intact retina (Figure 1C and F; Campbell, et al., 2021). In addition, expression of dll4 is downregulated following injury and can be increased by forced expression of Fgf8a or Tgfb3, both of which inhibit injury-dependent proliferation, suggesting that Dll4 may also be a candidate ligand for Notch3 (Wan and Goldman, 2017; Lee et al., 2020). However, morpholino-mediated knockdown of Dll4 expression reduced Müller glia proliferation during light damage unlike the knockdowns of Notch3 and DeltaB, which both increased proliferation during light damage (Campbell et al., 2021).
Alternatively, the deltaA and deltaD ligands and the notch1a receptor are expressed in the developing zebrafish retina with expression gradually restricted over time to the ciliary marginal zone (Taylor et al., 2015), an area of persistent neurogenesis throughout the life of the fish (Figure 1B) (Hitchcock et al., 2004). Data from single-cell RNA-seq studies suggest that, similar to the developing retina, deltaA, deltaD, and notch1a are expressed in reprogrammed/activated Müller glia at the time of peak Müller glia proliferation (36 hours light treatment), with deltaA, deltaC, and deltaD also expressed in Müller glia-derived progenitors (Figure 1D; Hoang et al., 2020). Since Notch1 activity is known to maintain retinal progenitors in a proliferative state in the developing retina (Jadhav et al., 2006a), then the outcomes of Notch activation during regeneration may be dictated, in part, by specific ligand-receptor expression patterns.
Ubiquitylation and endocytic trafficking
In addition to receptor glycosylation and ligand-receptor expression patterns, Notch activity is also regulated by ubiquitylation and endocytosis. Ubiquitylation and endocytosis of the ligand serves two purposes: 1) to regulate the amount of Notch ligand at the cell surface and 2) to generate force on the ligand-receptor complex to expose the S2 cleavage site in the negative regulatory region of the Notch receptor (Weinmaster and Fischer, 2011). Mindbomb (Mib) and Neuralized (Neur) are E3 ubiquitin ligases that ubiquitylate the Notch ligands, although it appears that only Mib is required for vertebrate Notch signaling, while both Mib and Neur function in Notch signaling in Drosophila (Weinmaster and Fischer, 2011). The mib1 mutant was first discovered in zebrafish and led to the description that Mib1 binds and ubiquitylates Delta ligands (Itoh et al., 2003). A functionally redundant Mib2 was identified in zebrafish and possesses an overlapping and distinct substrate specificity for the Notch ligands relative to Mib1 (Zhang et al., 2007a, b). During zebrafish development, the mib1 mutant retina has decreased expression of the Notch target gene her4 and increased expression of the neuronal differentiation marker huC, indicating that Mib is essential for increased Notch target gene expression, as well as neuronal differentiation during retinogenesis (Yoo et al., 2017). Furthermore, the Ran-binding protein family member RanBP9 was discovered to bind and antagonistically regulate Mib. Morpholino-mediated knockdown of RanBP9 increased her4 and decreased huC expression and also decreased Müller glia differentiation (Yoo et al., 2017). The endocytic regulation of Notch ligands has significant impact on Notch signaling, so ligand gene expression data alone may not fully reflect Notch activity in the signal-receiving cell.
Endocytosis of the Notch receptor within the signal-receiving cell can also affect pathway activity by reducing Notch receptor levels at the cell surface or by allowing ligand-independent receptor activation (Schnute et al., 2018). Several E3 ubiquitin ligases have been implicated in this process, including Deltex, Itch, and Nedd4. In zebrafish, the developing nervous system expresses deltex1, which Notch signaling suppresses through the downstream targets Her2 and Her8a (Cheng et al., 2015). Overexpression of the complete deltex1 coding sequence induced neuronal and glial cell differentiation, while expression of deltex1 coding sequence lacking the essential RING finger motif reduced neuronal and glial cell differentiation (Cheng et al., 2015). Numb and Numb-like also negatively regulate Notch activity. Numb antagonizes Notch signaling by recruiting E3-ligases, such as Itch, to ubiquitylate the NICD, marking it for degradation and signaling for Numb endocytic activity (Steinbuck and Winandy, 2018). In the mouse retina, Numb antagonizes Notch signaling in late-stage retinal progenitor cells to drive the production of terminal asymmetric divisions (Kechad et al., 2012). Therefore, in the regenerating zebrafish retina, Numb may be required in the later stages of NPC amplification to transition the Müller glia-derived NPCs from proliferation to differentiation.
Metalloprotease-dependent cleavage
Prior to ligand-binding, the Notch receptor’s S2 cleavage site is masked by the negative regulatory region. With ligand binding, the force of transendocytosis of the ligand-bound NECD into the signal-sending cell unfolds the negative regulatory region to allow cleavage of S2 by a metalloprotease (Gordon et al., 2007). Members of the ADAM (a disintegrin and metalloprotease) family of membrane-bound sheddases facilitate this event (Hsia et al., 2019). In multiple species, both ADAM10 and ADAM17 have been implicated in the S2 Notch cleavage, however only ADAM10 has been shown to be necessary for Notch function under physiological conditions (Alabi et al., 2021). In the developing retina, Adam10 conditional knockout mice exhibit lamination defects and abnormal neurogenesis due to a reduction of proliferating progenitor cells, which is characteristic of a deficiency in Notch signaling. Constitutive activation of Notch signaling via the Notch1 ICD rescues the Adam10 conditional knockout, confirming that Adam10 is essential for Notch signaling in the developing retina (Toonen et al., 2016).
Zebrafish have two Adam10 paralogs, Adam10a and Adam10b, and according to single-cell RNA-seq data, adam10a is expressed in all retinal cell types, including both resting and activated Müller glia following retinal damage (Hoang et al., 2020). Zebrafish adam17a is also expressed in all retinal cell types in response to damage (Hoang et al., 2020). While it may not play a significant role in Notch signaling, its expression is noted since a primary target of ADAM17 is tumor necrosis factor alpha (Tnfα) (Hsia et al., 2019), a factor that is produced by dying neurons and microglia and required for Müller glia proliferation during retinal regeneration (Nelson et al., 2013; Iribarne et al., 2019; Hoang et al., 2020). In the undamaged zebrafish retina, the combination of Tnfα and Notch repression is sufficient to activate a large percentage of Müller glia to proliferate and produce progenitor cells that commit to neuronal lineages in the absence of retinal damage (Conner et al., 2014). How Tnfα and Notch signaling interact to induce this response is still an open area of inquiry.
In addition, secreted frizzled-related proteins (Sfrp), which are described as multifunctional regulators of Wnt signaling (Esteve et al., 2011b), can also regulate Notch signaling through ADAM10 during mouse retinal neurogenesis (Esteve et al., 2011a). While loss of Sfrp1/2 expression in the Sfrp1–/–; Sfrp2–/– mouse retina results in overactivation of Notch signaling, ADAM10 inhibition rescues the phenotype (Esteve et al., 2011a). Furthermore, Sfrp1 was shown to physically interact with ADAM10 to inhibit processing of Notch (Esteve et al., 2011a). In zebrafish, sfrp1a and sfrp5 are both expressed in the developing retina and are implicated in patterning along the dorsoventral axis (Holly et al., 2014). The convergence of Notch, Tnfα, and Wnt signaling pathways on ADAM activity suggests that the regulation of Notch signaling for successful retinal regeneration may hinge, in part, on the regulation of Adam10 activity.
γ-Secretase-dependent cleavage
The S2 cleavage of ligand-activated Notch results in two products: the shed NECD polypeptide and a membrane-bound form called the Notch Extracellular Truncation, which is short-lived as it contains the S3 cleavage site and serves as the substrate for γ-secretase (Mumm et al., 2000). The cleavage by γ-secretase releases NICD from the membrane, allowing it to translocate directly to the nucleus (De Strooper et al., 1999). The γ-secretase complex is composed of four multi-pass membrane proteins that include the catalytic component Presenilin (Psen), Nicastrin, Anterior pharynx-defective 1, and Presenilin enhancer 2 (Wolfe, 2019). Many type-1 transmembrane proteins are γ-secretase substrates, although the most extensively studied are amyloid-β precursor protein and the Notch receptors (Hitzenberger et al., 2020). In zebrafish, morpholino-mediated knockdown of Psen1 results in abnormal eye morphology (Nery et al., 2017), and psen2 is highly expressed in the developing eye (Groth et al., 2002).
Inhibitors of the γ-secretase complex are often used to investigate Notch signaling during retinal regeneration. Two commonly used inhibitors are DAPT, which binds near the catalytic site on Presenilin, and RO4929097, which also binds and inhibits with a much stronger potency (Ran et al., 2017). Repression of Notch signaling with RO4929097 was sufficient to induce Müller glia proliferation in the absence of retinal damage and to increase proliferation during the light damage paradigm (Conner et al., 2014). When combined with TNFα exposure, Notch inhibition through RO4929097 induced a robust proliferative response in the undamaged zebrafish retina and produced Müller glia-derived progenitors that committed to the neuronal fate and differentiated into most types of retinal neurons (Conner et al., 2014). Similarly, DAPT exposure expanded the zone of proliferating Müller glia around the site of retinal poke injury where ascl1a expression was also increased (Wan et al., 2012). Expression of the Notch1a ICD with the Tg(hsp70l:gal4); Tg(UAS:myc-notch1a-intra) transgenic line reversed the effect of γ-secretase inhibition, indicating that induced and increased Müller glia proliferation was regulated by the inhibition of Notch signaling (Wan et al., 2012; Conner et al., 2014).
In the poke-injured retina with DAPT-mediated Notch inhibition, the pool of Müller glia-derived progenitor cells was biased to adopt the Müller glia fate, and alternatively, the limited number of progenitors produced in the injured retina with Notch1a ICD overexpression tended to differentiate into photoreceptors (Wan et al., 2012). This was surprising given that the opposite effect is observed in the developing mouse retina, where Notch inhibition enhances photoreceptor production (Jadhav et al., 2006b), and Notch1 activity maintains the progenitor state and promotes the glial cell fate (Furukawa et al., 2000; Jadhav et al., 2006a). These results may indicate that Notch signaling acts differently in the injured adult retina where dedifferentiation and proliferation are restricted for the purpose of repair within the fully formed retina, rather than for the formation of the entire retinal tissue from a mass of progenitor cells as in development. Alternatively, the differences in these results may reflect the methods used to modulate Notch signaling activity. Notch repression with γ-secretase inhibitor broadly inhibits all Notch receptor activity and forced NICD expression induces NICD activity in all cells, regardless of endogenous Notch expression or activity, presumably at a high level. Instead, targeting individual Notch signaling components in specific cell types may reveal unique roles for Notch signaling components that are hidden by broad inhibition or high levels of forced expression. For example, morpholino-mediated knockdowns of the Notch receptors in the light-damaged retina revealed that Müller glia quiescence is specifically maintained by Notch3, while Notch1a, Notch 1b, and Notch2 are each necessary for proliferation during regeneration (Campbell et al., 2021). While the available tools, whether pharmacological inhibition, knockdown techniques, or forced transgenic expression, are each useful in specific ways, we must consider how each tool influences Notch activity levels, as current modeling proposes that high and low Notch signaling activity influence different cell fate outcomes (Ivanov, 2019).
NICD-dependent transcriptional regulation
Following γ-secretase-dependent cleavage, the NICD translocates directly to the nucleus, where it forms a complex with the DNA-binding transcription factor CSL (CBF1/RBP-Jκ in mammals, Su(H) in Drosophila, and LAG-1 in C. elegans) and the co-activating factor Mastermind (Zema et al., 2020). In zebrafish, orthologs of these factors include the RBP-Jκ paralogs Rbpja and Rbpjb and the Mastermind-like homologs Maml1, Maml2, and Maml3.
The original model for transcriptional control of Notch target genes was that RBP-Jκ binds to CSL-binding sites statically. In the absence of NICD, co-repressors form a complex with RBP-Jκ to repress gene expression, and when Notch signaling is activated, the NICD displaces the co-repressors and activates transcription (Kao et al., 1998). However, this model was challenged by whole-genome binding profiles for RBP-Jκ and NICD, which revealed that CSL-binding sites in the enhancers and promoters of Notch target genes are dynamically bound by RBP-Jκ (Castel et al., 2013). In the absence of NICD, these sites are weakly bound by RBP-Jκ, and Notch signaling activation recruits the RBP-Jκ-NICD-MAML complex to these sites. Furthermore, for a set of binding sites, RBP-Jκ does not recruit NICD, suggesting that RBP-Jκ can function independently of NICD to directly regulate transcriptional targets (Castel et al., 2013). In the developing mouse retina, the proneural factor Atoh7 is regulated, in part, by Notch signaling. Four CSL binding sites were identified within the Atoh7 enhancer region that are regulated by RBP-Jκ using both Notch-dependent and Notch-independent mechanisms. It is suggested that levels of RBP-Jκ protein influence the RBP-Jκ-bound state of the Atoh7 locus (Miesfeld et al., 2018). In light of these and other data, it is suggested that instead of a switch model where NICD displaces co-repressors from RBP-Jκ, a dynamic exchange of the entire complex occurs and levels of the components, including RBP-Jκ and NICD, influence regulation of target transcription (Bray, 2016).
Further evidence that Notch signaling outcomes are dependent on protein levels comes from experiments where the Notch1 NICD was swapped with the Notch2 NICD and no abnormal phenotype was detected, indicating that the N1ICD and the N2ICD are functionally equivalent (Liu et al., 2015). Instead, Notch signaling outcomes appear to be dependent on the level of Notch signal. In the adult zebrafish telencephalic ventricular zone, the level of Notch signaling is suggested to influence radial glia oscillation between quiescent and neurogenic states (Chapouton et al., 2010). Cells with low levels of Notch activity express high levels of proliferation markers, whereas Notch1a ICD overexpression induced cell cycle exit and promoted a reversible quiescent state, such that cells could reenter the cell cycle after peak NICD overexpression. This led to the proposal that endogenous oscillation of Notch levels promotes a quiescent radial glial state when Notch signaling is active and induces radial glia proliferation when Notch levels are low. In a lateral inhibition-like feedback mechanism, proliferating progenitors activate Notch in their neighboring cells to maintain the pool of quiescent and proliferating progenitors during adulthood (Chapouton et al., 2010). In the zebrafish retina, Notch3 maintains Müller glia quiescence, whereas the other Notch receptors are required for sufficient Müller glia and NPC proliferation following damage (Campbell et al., 2021). Therefore, it is surprising that the Notch1a ICD can reduce Müller glia proliferation following Notch repression with γ-secretase inhibitor (Conner et al., 2014). In light of the evidence for functional equivalence of NICDs and the proposal that Notch levels dictate progenitor states, the simple explanation for these results is that induced overexpression of Notch1a ICD does not recapitulate endogenous Notch1a expression, but instead increases Notch signaling activity to a level that rescues the γ-secretase inhibition and promotes a return to Müller glia quiescence. Based on this model, however, additional investigation is required to determine how a specific level of Notch signaling induces Müller glia quiescence, while another level promotes proliferation and yet another level induces neuronal cell commitment and differentiation.
A recent integrated model of retinal neurogenesis proposes that the Notch-mediated lateral specification mechanism induces an oscillation of Hes1 expression, which drives expression of target gene products that not only oscillate, but also accumulate within a progenitor cell. The levels of these factors influence whether the cell differentiates to a neuronal phenotype, which is determined by the levels of transcription factors that have accumulated, or maintains a progenitor state and divides to give daughter cells that inherit the accumulated factors (Ivanov, 2019). This model further proposes that low Notch activity is required to promote the transition of progenitors into various retinal cell fates while high, sustained Notch activity promotes the Müller glia phenotype. Therefore, Notch1a ICD overexpression experiments must be interpreted carefully since the resulting reduction in Müller glia and NPC proliferation may be the result of high Notch signaling activity, which does not exclude the possibility that Notch1a (or Notch1b or Notch2) may be required for proliferation and progenitor transitions during regeneration.
Target gene expression
The principal transcriptional targets of Notch are members of the hairy and enhancer of split (Hes) and Hes related family of genes. The homologs in zebrafish are the her and hey genes, and they encode basic helix-loop-helix transcriptional repressors that play essential roles in maintaining progenitor cell populations and regulating cell fate decisions during tissue development (Kageyama et al., 2007). Two known Notch targets, Hes1 and Hes5, are required for development of the mouse optic vesicles and play roles in retinal development (Hatakeyama and Kageyama, 2004). During retinogenesis, the Dll1/Dll4-Notch1-Rbpj-Hes1 axis promotes retinal progenitor cell proliferation and inhibits retinal ganglion cell neurogenesis through suppression of Atoh7 expression (Riesenberg et al., 2009; Maurer et al., 2014). Hes5 is expressed in retinal progenitor cells once retinal neurogenesis begins, where it co-localizes with developing retinal ganglion cells. In addition, it is expressed in Müller glia of the adult mouse retina (Riesenberg et al., 2018).
The zebrafish ortholog of Hes1 is her6 and the orthologs of Hes5 are her2 and her4 (including the five tandem repeats her4.1-4.5). In the zebrafish retina, her6 is expressed in Müller glia and progenitor cells in the CMZ (Bernardos et al., 2005). Zebrafish her4 is also expressed in the CMZ (Figure 1B), as well as in slowly proliferating Müller glia in the adult retina, which is the source of newly generated rod photoreceptors throughout the life of the fish. In response to light damage, her4 expression is upregulated in proliferating cells in the outer and inner nuclear layers, which likely corresponds to the Müller glia-derived NPCs (Figure 1E) (Wilson et al., 2016). Inhibition of Notch signaling in the undamaged retina via the γ-secretase inhibitor RO4929097 decreased her4 expression but induced proliferation, giving further confirmation that her4 is a target of Notch signaling but suggesting that her4 is not required for Müller glia reprogramming (Conner et al., 2014). Instead, Her4 is suggested to regulate Müller glia reprogramming and proliferation through direct repression of lin28a since examination of the lin28a promoter revealed two Her4 binding sites, and morpholino-mediated knockdown of Her4.1 increased lin28a expression following poke injury (Mitra et al., 2018).
The functional investigation of downstream targets of Notch signaling during retinal regeneration has been limited to only a few target genes. However, it is likely that dynamic expression of combinations of her genes and other Notch targets contributes to the regulation of Müller glia quiescence, reprogramming, proliferation, and cell fate determination for successful regeneration. For example, the Hes1 ortholog her6, which promotes retinal progenitor cell proliferation and inhibits retinal ganglion cell neurogenesis, is broadly expressed in resting and activated Müller glia following light damage and NMDA-induced damage (Hoang et al., 2020), suggesting that it is another Notch target that regulates reprogramming and proliferation during regeneration. Alternatively, hey1 expression is transiently downregulated following retinal injury and can be upregulated with forced expression of quiescence factors Tgfb3 or Fgf8a, suggesting that hey1 may be a target of Notch3 (Wan and Goldman, 2017; Lee et al., 2020). To fully understand the dynamics of Notch signaling, it will be essential to explore the contributions of multiple Notch targets.
Network interactions
Damage disrupts the retinal landscape and, in response, a complex network of signaling pathways must be employed for successful regeneration (Lahne et al., 2020; Gao et al., 2021). To fully understand the dynamic role of Notch signaling for successful retinal regeneration, we must understand the interactions of Notch within the broader network of signaling at each stage of regeneration.
Recent studies demonstrate that Notch signaling maintains Müller glia quiescence in concert with Tgfb signaling and Fgf8a signaling (Wan and Goldman, 2017; Lee et al., 2020). Quiescent Müller glia in the zebrafish retina express tgfb3, and tgfb3 suppression is required for injury-dependent Müller glia proliferation. Furthermore, Tgfb3 suppresses reprogramming and proliferation and acts upstream of Notch (Lee et al., 2020). This result contrasts with another recent study that reports a pro-proliferative function of Tgfb signaling (Sharma et al., 2020), however the pro-proliferative function results from TGF-β1 application, not Tgfb3. At later stages of regeneration, Tgfb signaling is required for terminating NPC proliferation through Notch (Sharma et al., 2020). Müller glia also express fgf8a in the uninjured retina, which is almost immediately downregulated following injury. Fgf8a suppresses Müller glia proliferation through Notch activation. However, suppression of Müller glia proliferation by Fgf8a only applies to the older Müller glia of the central retina where Fgf8a increases Notch signaling. In the peripheral retina, where persistent neurogenesis is constantly producing young retinal tissue, Fgf8a acts through Notch inhibition to stimulate Müller glia proliferation (Wan and Goldman, 2017). Together, these studies suggest that additional signaling pathways act with Notch signaling to maintain quiescence in a context-dependent manner depending on age or on stage of regeneration.
Upon damage, rapid gene expression changes are induced in Müller glia as they enter a transient reactive state and progress into a reprogrammed/proliferative state (Hoang et al., 2020). The expression of notch3 is rapidly and drastically downregulated within four hours of light treatment, while notch1a and notch1b expression increases by 20 hours (Campbell et al., 2021), but it is unclear what signals are upstream of damage-induced changes in notch expression. A proposed mechanism involves the neurotransmitter γ-aminobutyric acid (GABA). As photoreceptors die, the horizontal cells are no longer stimulated to release GABA, which is detected by Müller glia. Inhibition of GABA signaling induces Müller glia proliferation and activation of Notch target her4 (Rao et al., 2017). Müller glia also respond to damaged-induced inflammatory signals such as TNFα, which is produced by dying neurons and enhances the Notch inhibition proliferative response (Nelson et al., 2013; Conner et al., 2014), and HB-EGF, which positively regulates deltaA, notch1a, and her4 expression through the reprogramming factor Ascl1a (Wan et al., 2012).
The upregulation of Notch signaling components by Ascl1a appears contradictory given that Notch inhibition is required for proliferation. However, it supports a dual role for Notch signaling in both maintaining Müller glia quiescence through Notch3 as well as being necessary for Müller glia reprogramming and proliferation through a combination of Notch1a, Notch1b, and Notch2 (Campbell et al., 2021). This is similar to what is seen in the mouse and chick retinas. During retinal development in mice, Notch signaling regulates retinal progenitor cell proliferation (Jadhav et al., 2006a), but promotes gliogenesis at later stages (Nelson et al., 2011; de Melo et al., 2016). In the injured postnatal chick retina, Notch signaling is required for Müller glia dedifferentiation and proliferation but later inhibits differentiation of newly generated progenitor cells (Hayes et al., 2007; Ghai et al., 2010). Recent studies indicate that Notch signaling is regulated during the proliferative phase by multiple factors, including Hdac1, which is secluded from NPCs and works with Mycb to inhibit her4.1 in the cells that neighbor NPCs (Mitra et al., 2018, 2019).
The role of Notch signaling to maintain Müller glia quiescence in the undamaged retina is well established, due to multiple studies with γ-secretase inhibitors and with the identification of Notch3 as the specific receptor that maintains quiescence (Wan et al., 2012; Conner et al., 2014; Campbell et al., 2021). Additionally, reports of crosstalk between Notch signaling and other major pathways as well as Notch signaling studies during retinal development provide a solid framework for our understanding of the dynamic role of Notch signaling during retinal regeneration (Table 2). However, the mechanisms by which Notch3 maintains quiescence and by which another Notch receptor or combination of receptors function in reprogramming and NPC proliferation is unclear. Given what it known about Notch signaling, its regulation may occur at any point from the post-translational modifications that bias ligand-receptor binding to the cell-specific ligand-receptor expression patterning to the oscillations and combinations of target genes that are induced upon Notch signaling activation. Further investigation is needed to address these points of regulation so we can work towards inducing endogenous repair of retinal damage in mammals.
Table 2.
Key Notch signaling findings in retinal development, growth, and regeneration
| Signaling component | Organism | Key finding | Reference |
|---|---|---|---|
| Development | |||
| Notch1 | Mouse | Maintains progenitor cell state and promotes cell proliferation | Furukawa et al., 2000 |
| Promotes MG formation | Jadhav et al., 2006a | ||
| Inhibits photoreceptor fate | Jadhav et al., 2006b | ||
| Dll4/Jagged1 and Fringe proteins | Mouse | Fringe glycosylation of Notch enhances Dll4 binding and weakens Jagged1 binding to regulate retinal angiogenesis | Benedito et al., 2009 |
| Mindbomb | Zebrafish | RanBP9 antagonistically regulates Mindbomb, which influences her4 expression | Yoo et al., 2017 |
| Numb | Mouse | Promotes early-stage cell-cycle progression and late-stage terminal asymmetric cell divisions | Kechad et al., 2012 |
| ADAM10 | Mouse | Activates Notch signaling to promote progenitor cell proliferation | Toonen et al., 2016 |
| Sfrp1 and Sfrp2 bind and downregulate ADAM10 activity | Esteve et al., 2011a | ||
| Psen1 | Zebrafish | Knockdown reduces eye size | Nery et al., 2017 |
| Rbpj | Mouse | Interacts with four CSL binding sites to repress Atoh7 expression in Notch-dependent and independent pathways | Miesfeld et al., 2018 |
| Hes1 | Mouse | Hes1 expressed in all progenitor cells | Riesenberg et al., 2018 |
| Hes1 suppresses Hes5 transcription | Nelson et al., 2011 | ||
| Hes5 | Hes5 expressed in subsets of progenitors and in differentiating ganglion cells | ||
| Hes5 expressed in adult MG | |||
| Hes5 | Mouse | Hes5-dependent gliogenesis requires Lhx2 | de Melo et al., 2016 |
| Dll1/Dll4-Notch1-Rbpj-Hes1 | Mouse | Promotes retinal progenitor cell proliferation and inhibits retinal ganglion cell neurogenesis | Riesenberg et al., 2009 Maurer et al., 2014 |
| Ciliary Marginal Zone (persistent neurogenesis) | |||
| Notch1a, Notch1b, DeltaA, DeltaD, Her2, Her4, Her6 | Zebrafish | Expression restricted to proliferative CMZ cells during retinal development and in the adult retina | Bernardos et al., 2005 Raymond et al., 2006 Taylor et al., 2015 |
| Regeneration | |||
| Notch3, DeltaB | Zebrafish | Notch3 expressed in MG, deltaB expressed in neurons; knockdown of either stimulates MG proliferation | Campbell et al., 2021 |
| γ-Secretase | Zebrafish | γ-Secretase inhibition increased/induced MG proliferation, which was reversed by Notch1a ICD expression | Wan et al., 2012 |
| MG proliferation induced by γ-secretase inhibition was enhanced by TNFα | Conner et al., 2014 | ||
| Notch1 and γ-secretase | Postnatal chick | Notch1 and signaling components upregulated after retinal damage | Hayes et al., 2007 |
| γ-Secretase inhibition prohibited MG dedifferentiation and proliferation at early regeneration stages and increased newly generated neurons at late regeneration stages | |||
| Notch1 expression is upregulated in proliferating MG in insulin/FGF2-treated retinas | Ghai et al., 2010 | ||
| γ-Secretase inhibition blocked FGF2-induced proliferation | |||
| Notch signaling/Fgf8a | Zebrafish | Forced Fgf8a expression increases Notch signaling and suppresses MG proliferation in older tissue but inhibits Notch and stimulates MG proliferation in young tissue | Wan and Goldman, 2017 |
| Dll4 | dll4 and hey1 expression is suppressed after injury, but forced Fgf8a after injury increases expression of both | ||
| Hey1 | |||
| Notch signaling/Tgfb signaling | Zebrafish | Tgfb3 acts upstream of Notch signaling to suppress injury-induced MG reprogramming and proliferation | Lee et al., 2020 |
| Dll4 | Forced Tgfb3 increases dll4 and hey1 expression | ||
| Hey1 | |||
| Her4 | Pharmacological inhibition of Tgfb increases her4.1 expression and decreases NPC proliferation at late stages | Sharma et al., 2020 | |
| Her4 | Zebrafish | her4 expressed in slowly proliferating MG of undamaged adult retina and in proliferating cells in ONL & INL following light damage | Wilson et al., 2016 |
| GABA inhibition induces MG proliferation and her4:RFP expression | Rao et al., 2017 | ||
| Represses lin28a expression; two Her4 binding sites on lin28a promoter | Mitra et al., 2018 | ||
| Late inhibition of her4.1 increases NPC proliferation | |||
| Mycb inhibits her4.1 expression with Hdac1 in cells neighboring NPCs | Mitra et al., 2019 | ||
| Stimulation of Regeneration in Mammals | |||
| Notch1, Delta1, Hes1 | Rat | Expression increased in neurotoxin-damaged retina | Das et al., 2006 |
| γ-Secretase | γ-Secretase inhibition decreased cell cycle regulators cyclinA and cyclinD1 | ||
| Jag1 | Mouse | Jag1-activation of Notch signaling with Wnt activation induced MG cell cycle reentry and rod photoreceptor differentiation in culture, retinal explant, and in S334ter rat in vivo | Del Debbio et al., 2010 |
| Hes1 | Rat | Notch activates MG by inhibiting p27Kip1 expression | Del Debbio et al., 2016 |
| γ-Secretase | Mouse | γ-Secretase inhibition with forced Ascl1a and Lin28 expression inefficiently stimulated MG proliferation in mouse retina | Elsaeidi et al., 2018 |
ADAM: A disintegrin and metalloprotease 10; CMZ: ciliary marginal zone; Dll: delta-like; Her: hairy-related; Hes: hairy and enhancer of split; Hey: Hes-related; Jag: jagged; MG: Müller glia; Psen: presenilin.
Concluding Remarks: Implications for Mammalian Retinal Regeneration
The goal of retinal regeneration research is to discover mechanisms to induce endogenous stem cells to produce neurons that are lost in human disease. In response to damage, Müller glia in the mouse retina enter a reactive state that is molecularly similar to Müller glia in the zebrafish and chick. However, while the zebrafish and chick Müller glia pass through this reactive phase to reprogram and proliferate, the mouse Müller glia pass through the reactive phase, arrest, and return to quiescence (Hoang et al., 2020). Forced expression of Ascl1 together with HDAC inhibition can reprogram mouse Müller glia with limited neurogenic potential (Jorstad et al., 2017), and Müller glia-specific deletion of the NFI factors, Nfia/b/x, can induce proliferation and formation of retinal neurons (Hoang et al., 2020). Therefore, the mammalian retina possesses the potential to undergo a regenerative response following damage, but the dynamic network of endogenous signals is not fully employed as in the highly regenerative zebrafish retina or in the limited-regenerative postnatal chick retina.
How does the Notch pathway function in the damaged mammalian retina, and is it acting dynamically like in the zebrafish retina? Expression studies in mice confirm that Notch signaling remains active in postmitotic Müller glia after they begin to differentiate (Nelson et al., 2011) and that Notch pathway genes, such as Notch1, Notch2, Hes1, and Hes5, are expressed in resting and activated Müller glia in the adult (Hoang et al., 2020). In the damaged mouse retina, Notch1, Delta1, and Hes1 expression increases (Das et al., 2006). Furthermore, activation of Notch and Wnt signaling together can induce limited Müller glia proliferation and differentiation into rod photoreceptors, both in mouse retinal explants and in vivo in the S334ter rat mutant that exhibits rod photoreceptor degeneration (Del Debbio et al., 2010, 2016). However, whereas forced expression of ascl1a and lin28a in combination with γ-secretase inhibition can stimulate widespread Müller glia proliferation and neuronal generation in the zebrafish retina, a similar forced expression and Notch inhibition in the mouse retina results in limited and inefficient Müller glia proliferation (Elsaeidi et al., 2018). This suggests that additional endogenous molecular mechanisms limit Müller glia activation and robust proliferation in the mammalian retina. Identification of those mechanisms is a necessary first step towards inducing endogenous repair.
The Hippo pathway was recently identified as an essential molecular mechanism that restrains Müller glia reprogramming and proliferation (Hamon et al., 2019; Rueda et al., 2019). Following retinal damage in mice, the Hippo pathway effector Yes-associated protein (Yap) is upregulated, which signals the upregulation of cell-cycle genes for Müller glia activation. However, sustained activity of Yap in the damaged retina is prevented by active Hippo signaling that phosphorylates Yap, marking it for degradation and preventing its interaction with the TEAD transcription factor. Bypassing Hippo signaling through Yap overexpression or with a phosphodeficient form of Yap results in spontaneous and robust Müller glia reprogramming and proliferation, although neurogenic competence is not induced (Hamon et al., 2019; Rueda et al., 2019). In the zebrafish retina, yap1 is upregulated in reactive Müller glia and knockdown of Yap1 or pharmacological inhibition of the Yap1/TEAD interaction was shown to reduce Müller glia proliferation in the light-damaged retina, indicating a conserved role for Yap as an essential driver of Müller glia proliferation (Hoang et al., 2020). Recent studies have shown that the Hippo pathway interacts with other major signaling pathways through its role as a central mediator of cellular signals (Wang and Martin, 2017). Epidermal stem cell fate, for example, is promoted by Notch inhibition through mechano-activation of YAP/TAZ (Totaro et al., 2017), however, the specific crosstalk between the Hippo and Notch pathways to regulate Müller glia activation requires further exploration.
Recent research describes a role for Notch signaling primarily as a positive regulator of Müller glia quiescence and inhibitor of proliferation, however knockdown of the individual Notch receptors in the regenerating zebrafish retina suggests that Notch signaling may be dynamic in its effects during regeneration. This leads to many outstanding questions about how Notch signaling is dynamically regulated during quiescence and in response to retinal damage, which include:
What are the cell-specific expression patterns and functions of Notch receptors and ligands throughout the stages of retinal regeneration? How do specific ligand-receptor binding pairs elicit different responses during regeneration? What levels of Notch signaling activity are required for the progression through regenerative stages?
What is upstream of Notch signaling that regulates ligand expression and promotes stability of cell-cell contact with regard to ligand-receptor binding in the intact retina? Does the death of specific cell types as a result of different damage methods influence the dynamics of Notch signaling?
How do Notch regulatory factors such as the Fringe proteins, Mindbomb, Numb, and ADAM10, all which have identified roles in retinal development, influence Notch signaling at the different phases of zebrafish retinal regeneration?
What are the transcriptional targets of Notch signaling in the retina and how do combinations of Notch receptors and targets interact with the network of signaling pathways during the regeneration time course?
Addressing these and other gaps in our understanding of dynamic Notch signaling in the regenerating retina will inform our efforts in employing potential Notch-based therapeutics to stimulate endogenous repair and restore vision in retinal damage and disease.
Additional file: Open peer review reports 1 (88.1KB, pdf) and 2 (84.4KB, pdf) .
Acknowledgments:
We thank members of the Hyde Lab for thoughtful discussions.
Footnotes
Conflicts of interest: The authors declare no conflicts of interest.
Financial support: This work was supported by National Eye Institute R01-EY024519 and U01-EY027267 (to DRH) and the Center for Zebrafish Research, University of Notre Dame.
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewers: Mariana S. Silveira, Federal University of Rio de Janeiro, Brazil; Lucia Poggi, University of Trento, Italy.
Funding: This work was supported by National Eye Institute R01-EY024519 and U01-EY027267 (to DRH) and the Center for Zebrafish Research, University of Notre Dame.
References
- 1.Alabi RO, Lora J, Celen AB, Maretzky T, Blobel CP. Analysis of the conditions that affect the selective processing of endogenous notch1 by ADAM10 and ADAM17. Int J Mol Sci. 2021;22:1846. doi: 10.3390/ijms22041846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Amini R, Rocha-Martins M, Norden C. Neuronal migration and lamination in the vertebrate retina. Front Neurosci. 2017;11:742. doi: 10.3389/fnins.2017.00742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Benedito R, Roca C, Sorensen I, Adams S, Gossler A, Fruttiger M, Adams RH. The notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis. Cell. 2009;137:1124–1135. doi: 10.1016/j.cell.2009.03.025. [DOI] [PubMed] [Google Scholar]
- 4.Bernardos RL, Lentz SI, Wolfe MS, Raymond PA. Notch-Delta signaling is required for spatial patterning and Müller glia differentiation in the zebrafish retina. Dev Biol. 2005;278:381–395. doi: 10.1016/j.ydbio.2004.11.018. [DOI] [PubMed] [Google Scholar]
- 5.Bray SJ. Notch signalling in context. Nat Rev Mol Cell Biol. 2016;17:722–735. doi: 10.1038/nrm.2016.94. [DOI] [PubMed] [Google Scholar]
- 6.Bringmann A, Wiedemann P. Müller glial cells in retinal disease. Ophthalmologica. 2012;227:1–19. doi: 10.1159/000328979. [DOI] [PubMed] [Google Scholar]
- 7.Campbell LJ, Hobgood JS, Jia M, Boyd P, Hipp RI, Hyde DR. Notch3 and DeltaB maintain Müller glia quiescence and act as negative regulators of regeneration in the light-damaged zebrafish retina. Glia. 2021;69:546–566. doi: 10.1002/glia.23912. [DOI] [PubMed] [Google Scholar]
- 8.Castel D, Mourikis P, Bartels SJ, Brinkman AB, Tajbakhsh S, Stunnenberg HG. Dynamic binding of RBPJ is determined by Notch signaling status. Genes Dev. 2013;27:1059–1071. doi: 10.1101/gad.211912.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chapouton P, Skupien P, Hesl B, Coolen M, Moore JC, Madelaine R, Kremmer E, Faus-Kessler T, Blader P, Lawson ND, Bally-Cuif L. Notch activity levels control the balance between quiescence and recruitment of adult neural stem cells. J Neurosci. 2010;30:7961–7974. doi: 10.1523/JNEUROSCI.6170-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cheng YC, Huang YC, Yeh TH, Shih HY, Lin CY, Lin SJ, Chiu CC, Huang CW, Jiang YJ. Deltex1 is inhibited by the Notch-Hairy/E(Spl) signaling pathway and induces neuronal and glial differentiation. Neural Dev. 2015;10:28. doi: 10.1186/s13064-015-0055-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cho KO, Choi KW. Fringe is essential for mirror symmetry and morphogenesis in the Drosophila eye. Nature. 1998;396:272–276. doi: 10.1038/24394. [DOI] [PubMed] [Google Scholar]
- 12.Conner C, Ackerman KM, Lahne M, Hobgood JS, Hyde DR. Repressing notch signaling and expressing TNFα are sufficient to mimic retinal regeneration by inducing Müller glial proliferation to generate committed progenitor cells. J Neurosci. 2014;34:14403–14419. doi: 10.1523/JNEUROSCI.0498-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.D’Orazi FD, Zhao XF, Wong RO, Yoshimatsu T. Mismatch of synaptic patterns between neurons produced in regeneration and during development of the vertebrate retina. Curr Biol. 2016;26:2268–2279. doi: 10.1016/j.cub.2016.06.063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.D’Souza B, Miyamoto A, Weinmaster G. The many facets of Notch ligands. Oncogene. 2008;27:5148–5167. doi: 10.1038/onc.2008.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Das AV, Mallya KB, Zhao X, Ahmad F, Bhattacharya S, Thoreson WB, Hegde GV, Ahmad I. Neural stem cell properties of Müller glia in the mammalian retina: regulation by Notch and Wnt signaling. Dev Biol. 2006;299:283–302. doi: 10.1016/j.ydbio.2006.07.029. [DOI] [PubMed] [Google Scholar]
- 16.de Melo J, Zibetti C, Clark BS, Hwang W, Miranda-Angulo AL, Qian J, Blackshaw S. Lhx2 is an essential factor for retinal gliogenesis and notch signaling. J Neurosci. 2016;36:2391–2405. doi: 10.1523/JNEUROSCI.3145-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.De Strooper B, Annaert W, Cupers P, Saftig P, Craessaerts K, Mumm JS, Schroeter EH, Schrijvers V, Wolfe MS, Ray WJ, Goate A, Kopan R. A presenilin-1-dependent gamma-secretase-like protease mediates release of Notch intracellular domain. Nature. 1999;398:518–522. doi: 10.1038/19083. [DOI] [PubMed] [Google Scholar]
- 18.Del Debbio CB, Balasubramanian S, Parameswaran S, Chaudhuri A, Qiu F, Ahmad I. Notch and Wnt signaling mediated rod photoreceptor regeneration by Müller cells in adult mammalian retina. PLoS One. 2010;5:e12425. doi: 10.1371/journal.pone.0012425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Del Debbio CB, Mir Q, Parameswaran S, Mathews S, Xia X, Zheng L, Neville AJ, Ahmad I. Notch signaling activates stem cell properties of müller glia through transcriptional regulation and Skp2-mediated degradation of p27Kip1. PLoS One. 2016;11:e0152025. doi: 10.1371/journal.pone.0152025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Demmin DL, Silverstein SM. Visual impairment and mental health: unmet needs and treatment options. Clin Ophthalmol. 2020;14:4229–4251. doi: 10.2147/OPTH.S258783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Elsaeidi F, Macpherson P, Mills EA, Jui J, Flannery JG, Goldman D. Notch suppression collaborates with Ascl1 and Lin28 to unleash a regenerative response in fish retina but not in mice. J Neurosci. 2018;38:2246–2261. doi: 10.1523/JNEUROSCI.2126-17.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Esteve P, Sandonis A, Cardozo M, Malapeira J, Ibanez C, Crespo I, Marcos S, Gonzalez-Garcia S, Toribio ML, Arribas J, Shimono A, Guerrero I, Bovolenta P. SFRPs act as negative modulators of ADAM10 to regulate retinal neurogenesis. Nat Neurosci. 2011a;14:562–569. doi: 10.1038/nn.2794. [DOI] [PubMed] [Google Scholar]
- 23.Esteve P, Sandonis A, Ibanez C, Shimono A, Guerrero I, Bovolenta P. Secreted frizzled-related proteins are required for Wnt/β-catenin signalling activation in the vertebrate optic cup. Development. 2011b;138:4179–4184. doi: 10.1242/dev.065839. [DOI] [PubMed] [Google Scholar]
- 24.Furukawa T, Mukherjee S, Bao ZZ, Morrow EM, Cepko CL. rax, Hes1, and notch1 promote the formation of müller glia by postnatal retinal progenitor cells. Neuron. 2000;26:383–394. doi: 10.1016/s0896-6273(00)81171-x. [DOI] [PubMed] [Google Scholar]
- 25.Gao H, A L, Huang X, Chen X, Xu H. Müller glia-mediated retinal regeneration. Mol Neurobiol. 2021;58:2342–2361. doi: 10.1007/s12035-020-02274-w. [DOI] [PubMed] [Google Scholar]
- 26.Ghai K, Zelinka C, Fischer AJ. Notch signaling influences neuroprotective and proliferative properties of mature Müller glia. J Neurosci. 2010;30:3101–3112. doi: 10.1523/JNEUROSCI.4919-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gordois A, Cutler H, Pezzullo L, Gordon K, Cruess A, Winyard S, Hamilton W, Chua K. An estimation of the worldwide economic and health burden of visual impairment. Glob Public Health. 2012;7:465–481. doi: 10.1080/17441692.2011.634815. [DOI] [PubMed] [Google Scholar]
- 28.Gordon WR, Vardar-Ulu D, Histen G, Sanchez-Irizarry C, Aster JC, Blacklow SC. Structural basis for autoinhibition of Notch. Nat Struct Mol Biol. 2007;14:295–300. doi: 10.1038/nsmb1227. [DOI] [PubMed] [Google Scholar]
- 29.Grotek B, Wehner D, Weidinger G. Notch signaling coordinates cellular proliferation with differentiation during zebrafish fin regeneration. Development. 2013;140:1412–1423. doi: 10.1242/dev.087452. [DOI] [PubMed] [Google Scholar]
- 30.Groth C, Nornes S, McCarty R, Tamme R, Lardelli M. Identification of a second presenilin gene in zebrafish with similarity to the human Alzheimer’s disease gene presenilin2. Dev Genes Evol. 2002;212:486–490. doi: 10.1007/s00427-002-0269-5. [DOI] [PubMed] [Google Scholar]
- 31.Hamon A, García-García D, Ail D, Bitard J, Chesneau A, Dalkara D, Locker M, Roger JE, Perron M. Linking YAP to müller glia quiescence exit in the degenerative retina. Cell Rep. 2019;27:1712–1725. doi: 10.1016/j.celrep.2019.04.045. [DOI] [PubMed] [Google Scholar]
- 32.Hatakeyama J, Kageyama R. Retinal cell fate determination and bHLH factors. Semin Cell Dev Biol. 2004;15:83–89. doi: 10.1016/j.semcdb.2003.09.005. [DOI] [PubMed] [Google Scholar]
- 33.Hayes S, Nelson BR, Buckingham B, Reh TA. Notch signaling regulates regeneration in the avian retina. Dev Biol. 2007;312:300–311. doi: 10.1016/j.ydbio.2007.09.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Henrique D, Schweisguth F. Mechanisms of Notch signaling: a simple logic deployed in time and space. Development. 2019;146:172148. doi: 10.1242/dev.172148. [DOI] [PubMed] [Google Scholar]
- 35.Hitchcock P, Ochocinska M, Sieh A, Otteson D. Persistent and injury-induced neurogenesis in the vertebrate retina. Prog Retin Eye Res. 2004;23:183–194. doi: 10.1016/j.preteyeres.2004.01.001. [DOI] [PubMed] [Google Scholar]
- 36.Hitzenberger M, Gotz A, Menig S, Brunschweiger B, Zacharias M, Scharnagl C. The dynamics of γ-secretase and its substrates. Semin Cell Dev Biol. 2020;105:86–101. doi: 10.1016/j.semcdb.2020.04.008. [DOI] [PubMed] [Google Scholar]
- 37.Hoang T, Wang J, Boyd P, Wang F, Santiago C, Jiang L, Yoo S, Lahne M, Todd LJ, Jia M, Saez C, Keuthan C, Palazzo I, Squires N, Campbell WA, Rajaii F, Parayil T, Trinh V, Kim DW, Wang G, et al. Gene regulatory networks controlling vertebrate retinal regeneration. Science. 2020;370:8598. doi: 10.1126/science.abb8598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Holly VL, Widen SA, Famulski JK, Waskiewicz AJ. Sfrp1a and Sfrp5 function as positive regulators of Wnt and BMP signaling during early retinal development. Dev Biol. 2014;388:192–204. doi: 10.1016/j.ydbio.2014.01.012. [DOI] [PubMed] [Google Scholar]
- 39.Hoon M, Okawa H, Della Santina L, Wong RO. Functional architecture of the retina: development and disease. Prog Retin Eye Res. 2014;42:44–84. doi: 10.1016/j.preteyeres.2014.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hsia HE, Tüshaus J, Brummer T, Zheng Y, Scilabra SD, Lichtenthaler SF. Functions of ‘A disintegrin and metalloproteases (ADAMs)’ in the mammalian nervous system. Cell Mol Life Sci. 2019;76:3055–3081. doi: 10.1007/s00018-019-03173-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Iribarne M. Inflammation induces zebrafish regeneration. Neural Regen Res. 2021;16:1693–1701. doi: 10.4103/1673-5374.306059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Iribarne M, Hyde DR, Masai I. TNFα induces müller glia to transition from non-proliferative gliosis to a regenerative response in mutant zebrafish presenting chronic photoreceptor degeneration. Front Cell Dev Biol. 2019;7:296. doi: 10.3389/fcell.2019.00296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Itoh M, Kim CH, Palardy G, Oda T, Jiang YJ, Maust D, Yeo SY, Lorick K, Wright GJ, Ariza-McNaughton L, Weissman AM, Lewis J, Chandrasekharappa SC, Chitnis AB. Mind bomb is a ubiquitin ligase that is essential for efficient activation of notch signaling by delta. Dev Cell. 2003;4:67–82. doi: 10.1016/s1534-5807(02)00409-4. [DOI] [PubMed] [Google Scholar]
- 44.Ivanov D. Notch signaling-induced oscillatory gene expression may drive neurogenesis in the developing retina. Front Mol Neurosci. 2019;12:226. doi: 10.3389/fnmol.2019.00226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Jadhav AP, Cho SH, Cepko CL. Notch activity permits retinal cells to progress through multiple progenitor states and acquire a stem cell property. Proc Natl Acad Sci U S A. 2006a;103:18998–19003. doi: 10.1073/pnas.0608155103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jadhav AP, Mason HA, Cepko CL. Notch 1 inhibits photoreceptor production in the developing mammalian retina. Development. 2006b;133:913–923. doi: 10.1242/dev.02245. [DOI] [PubMed] [Google Scholar]
- 47.Jorstad NL, Wilken MS, Grimes WN, Wohl SG, VandenBosch LS, Yoshimatsu T, Wong RO, Rieke F, Reh TA. Stimulation of functional neuronal regeneration from Müller glia in adult mice. Nature. 2017;548:103–107. doi: 10.1038/nature23283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kageyama R, Ohtsuka T, Kobayashi T. The Hes gene family: repressors and oscillators that orchestrate embryogenesis. Development. 2007;134:1243–1251. doi: 10.1242/dev.000786. [DOI] [PubMed] [Google Scholar]
- 49.Kao HY, Ordentlich P, Koyano-Nakagawa N, Tang Z, Downes M, Kintner CR, Evans RM, Kadesch T. A histone deacetylase corepressor complex regulates the Notch signal transduction pathway. Genes Dev. 1998;12:2269–2277. doi: 10.1101/gad.12.15.2269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kechad A, Jolicoeur C, Tufford A, Mattar P, Chow RWY, Harris WA, Cayouette M. Numb is required for the production of terminal asymmetric cell divisions in the developing mouse retina. J Neurosci. 2012;32:17197–17210. doi: 10.1523/JNEUROSCI.4127-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kopan R, Ilagan MX. The canonical Notch signaling pathway: unfolding the activation mechanism. Cell. 2009;137:216–233. doi: 10.1016/j.cell.2009.03.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lahne M, Brecker M, Jones SE, Hyde DR. The Regenerating adult zebrafish retina recapitulates developmental fate specification programs. Front Cell Dev Biol. 2021;8:617923. doi: 10.3389/fcell.2020.617923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lahne M, Li J, Marton RM, Hyde DR. Actin-cytoskeleton- and rock-mediated INM are required for photoreceptor regeneration in the adult zebrafish retina. J Neurosci. 2015;35:15612–15634. doi: 10.1523/JNEUROSCI.5005-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lahne M, Nagashima M, Hyde DR, Hitchcock PF. Reprogramming Müller glia to regenerate retinal neurons. Annu Rev Vis Sci. 2020;6:171–193. doi: 10.1146/annurev-vision-121219-081808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lake RJ, Grimm LM, Veraksa A, Banos A, Artavanis-Tsakonas S. In vivo analysis of the Notch receptor S1 cleavage. PLoS One. 2009;4:e6728. doi: 10.1371/journal.pone.0006728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lee MS, Wan J, Goldman D. Tgfb3 collaborates with PP2A and notch signaling pathways to inhibit retina regeneration. Elife. 2020;9:e55137. doi: 10.7554/eLife.55137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Liu Z, Brunskill E, Varnum-Finney B, Zhang C, Zhang A, Jay PY, Bernstein I, Morimoto M, Kopan R. The intracellular domains of Notch1 and Notch2 are functionally equivalent during development and carcinogenesis. Development. 2015;142:2452–2463. doi: 10.1242/dev.125492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Logeat F, Bessia C, Brou C, LeBail O, Jarriault S, Seidah NG, Israel A. The Notch1 receptor is cleaved constitutively by a furin-like convertase. Proc Natl Acad Sci U S A. 1998;95:8108–8112. doi: 10.1073/pnas.95.14.8108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Maurer KA, Riesenberg AN, Brown NL. Notch signaling differentially regulates Atoh7 and Neurog2 in the distal mouse retina. Development. 2014;141:3243–3254. doi: 10.1242/dev.106245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.McGinn TE, Galicia CA, Leoni DC, Partington N, Mitchell DM, Stenkamp DL. Rewiring the regenerated zebrafish retina: reemergence of bipolar neurons and cone-bipolar circuitry following an inner retinal lesion. Front Cell Dev Biol. 2019;7:95. doi: 10.3389/fcell.2019.00095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.McGinn TE, Mitchell DM, Meighan PC, Partington N, Leoni DC, Jenkins CE, Varnum MD, Stenkamp DL. Restoration of dendritic complexity functional connectivity and diversity of regenerated retinal bipolar neurons in adult zebrafish. J Neurosci. 2018;38:120–136. doi: 10.1523/JNEUROSCI.3444-16.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Miesfeld JB, Moon MS, Riesenberg AN, Contreras AN, Kovall RA, Brown NL. Rbpj direct regulation of Atoh7 transcription in the embryonic mouse retina. Sci Rep. 2018;8:10195. doi: 10.1038/s41598-018-28420-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Mitra S, Sharma P, Kaur S, Khursheed MA, Gupta S, Ahuja R, Kurup AJ, Chaudhary M, Ramachandran R. Histone deacetylase-mediated müller glia reprogramming through Her4 1-Lin28a axis is essential for retina regeneration in zebrafish. iScience. 2018;7:68–84. doi: 10.1016/j.isci.2018.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mitra S, Sharma P, Kaur S, Khursheed MA, Gupta S, Chaudhary M, Kurup AJ, Ramachandran R. Dual regulation of lin28a by Myc is necessary during zebrafish retina regeneration. J Cell Biol. 2019;218:489–507. doi: 10.1083/jcb.201802113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Moloney DJ, Shair LH, Lu FM, Xia J, Locke R, Matta KL, Haltiwanger RS. Mammalian Notch1 is modified with two unusual forms of O-linked glycosylation found on epidermal growth factor-like modules. J Biol Chem. 2000;275:9604–9611. doi: 10.1074/jbc.275.13.9604. [DOI] [PubMed] [Google Scholar]
- 66.Mumm JS, Schroeter EH, Saxena MT, Griesemer A, Tian X, Pan DJ, Ray WJ, Kopan R. A ligand-induced extracellular cleavage regulates γ-secretase-like proteolytic activation of Notch1. Mol Cell. 2000;5:197–206. doi: 10.1016/s1097-2765(00)80416-5. [DOI] [PubMed] [Google Scholar]
- 67.Münch J, Grivas D, Gonzalez-Rajal A, Torregrosa-Carrion R, de la Pompa JL. Notch signalling restricts inflammation and serpine1 expression in the dynamic endocardium of the regenerating zebrafish heart. Development. 2017;144:1425–1440. doi: 10.1242/dev.143362. [DOI] [PubMed] [Google Scholar]
- 68.Nagashima M, Barthel LK, Raymond PA. A self-renewing division of zebrafish Müller glial cells generates neuronal progenitors that require N-cadherin to regenerate retinal neurons. Development. 2013;140:4510–4521. doi: 10.1242/dev.090738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Nelson BR, Ueki Y, Reardon S, Karl MO, Georgi S, Hartman BH, Lamba DA, Reh TA. Genome-wide analysis of Müller glial differentiation reveals a requirement for Notch signaling in postmitotic cells to maintain the glial fate. PLoS One. 2011;6:e22817. doi: 10.1371/journal.pone.0022817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Nelson CM, Ackerman KM, O’Hayer P, Bailey TJ, Gorsuch RA, Hyde DR. Tumor necrosis factor-alpha is produced by dying retinal neurons and is required for Müller glia proliferation during zebrafish retinal regeneration. J Neurosci. 2013;33:6524–6539. doi: 10.1523/JNEUROSCI.3838-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Nery LR, Silva NE, Fonseca R, Vianna MRM. Presenilin-1 targeted morpholino induces cognitive deficits increased brain Abeta1-42 and decreased synaptic marker PSD-95 in zebrafish larvae. Neurochem Res. 2017;42:2959–2967. doi: 10.1007/s11064-017-2327-4. [DOI] [PubMed] [Google Scholar]
- 72.Nikolaou N, Watanabe-Asaka T, Gerety S, Distel M, Koster RW, Wilkinson DG. Lunatic fringe promotes the lateral inhibition of neurogenesis. Development. 2009;136:2523–2533. doi: 10.1242/dev.034736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Panin VM, Papayannopoulos V, Wilson R, Irvine KD. Fringe modulates Notch-ligand interactions. Nature. 1997;387:908–912. doi: 10.1038/43191. [DOI] [PubMed] [Google Scholar]
- 74.Panin VM, Shao L, Lei L, Moloney DJ, Irvine KD, Haltiwanger RS. Notch ligands are substrates for protein O-fucosyltransferase-1 and Fringe. J Biol Chem. 2002;277:29945–29952. doi: 10.1074/jbc.M204445200. [DOI] [PubMed] [Google Scholar]
- 75.Powell C, Cornblath E, Elsaeidi F, Wan J, Goldman D. Zebrafish Müller glia-derived progenitors are multipotent exhibit proliferative biases and regenerate excess neurons. Sci Rep. 2016;6:24851. doi: 10.1038/srep24851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Ran Y, Hossain F, Pannuti A, Lessard CB, Ladd GZ, Jung JI, Minter LM, Osborne BA, Miele L, Golde TE. γ-Secretase inhibitors in cancer clinical trials are pharmacologically and functionally distinct. EMBO Mol Med. 2017;9:950–966. doi: 10.15252/emmm.201607265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Rana NA, Haltiwanger RS. Fringe benefits: functional and structural impacts of O-glycosylation on the extracellular domain of Notch receptors. Curr Opin Struct Biol. 2011;21:583–589. doi: 10.1016/j.sbi.2011.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Rao MB, Didiano D, Patton JG. Neurotransmitter-regulated regeneration in the zebrafish retina. Stem Cell Reports. 2017;8:831–842. doi: 10.1016/j.stemcr.2017.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Raymond P, Barthel L, Bernardos R, Perkowski J. Molecular characterization of retinal stem cells and their niches in adult zebrafish. BMC Dev Biol. 2006;6:36. doi: 10.1186/1471-213X-6-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Riesenberg AN, Conley KW, Le TT, Brown NL. Separate and coincident expression of Hes1 and Hes5 in the developing mouse eye. Dev Dyn. 2018;247:212–221. doi: 10.1002/dvdy.24542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Riesenberg AN, Liu Z, Kopan R, Brown NL. Rbpj cell autonomous regulation of retinal ganglion cell and cone photoreceptor fates in the mouse retina. J Neurosci. 2009;29:12865–12877. doi: 10.1523/JNEUROSCI.3382-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Rueda EM, Hall BM, Hill MC, Swinton PG, Tong X, Martin JF, Poché RA. The hippo pathway blocks mammalian retinal müller glial cell reprogramming. Cell Rep. 2019;27:1637–1649. doi: 10.1016/j.celrep.2019.04.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Saiki W, Ma C, Okajima T, Takeuchi H. Current views on the roles of O-glycosylation in controlling notch-ligand interactions. Biomolecules. 2021;11:309. doi: 10.3390/biom11020309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sakaidani Y, Ichiyanagi N, Saito C, Nomura T, Ito M, Nishio Y, Nadano D, Matsuda T, Furukawa K, Okajima T. O-linked-N-acetylglucosamine modification of mammalian Notch receptors by an atypical O-GlcNAc transferase Eogt1. Biochem Biophys Res Commun. 2012;419:14–19. doi: 10.1016/j.bbrc.2012.01.098. [DOI] [PubMed] [Google Scholar]
- 85.Schnute B, Troost T, Klein T. Endocytic trafficking of the notch receptor. Adv Exp Med Biol. 2018;1066:99–122. doi: 10.1007/978-3-319-89512-3_6. [DOI] [PubMed] [Google Scholar]
- 86.Sharma P, Gupta S, Chaudhary M, Mitra S, Chawla B, Khursheed MA, Saran NK, Ramachandran R. Biphasic role of Tgf-β signaling during müller glia reprogramming and retinal regeneration in zebrafish. iScience. 2020;23:100817. doi: 10.1016/j.isci.2019.100817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Shi S, Stanley P. Protein O-fucosyltransferase 1 is an essential component of Notch signaling pathways. Proc Natl Acad Sci U S A. 2003;100:5234–5239. doi: 10.1073/pnas.0831126100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Steinbuck MP, Winandy S. A review of notch processing with new insights into ligand-independent notch signaling in T-cells. Front Immunol. 2018;9:1230. doi: 10.3389/fimmu.2018.01230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Takeuchi H, Schneider M, Williamson DB, Ito A, Takeuchi M, Handford PA, Haltiwanger RS. Two novel protein O-glucosyltransferases that modify sites distinct from POGLUT1 and affect Notch trafficking and signaling. Proc Natl Acad Sci U S A. 2018;115:E8395–8402. doi: 10.1073/pnas.1804005115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Taylor SM, Alvarez-Delfin K, Saade CJ, Thomas JL, Thummel R, Fadool JM, Hitchcock PF. The bHLH transcription factor NeuroD governs photoreceptor genesis and regeneration through delta-notch signaling. Invest Ophthalmol Vis Sci. 2015;56:7496–7515. doi: 10.1167/iovs.15-17616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Thomas JL, Ranski AH, Morgan GW, Thummel R. Reactive gliosis in the adult zebrafish retina. Exp Eye Res. 2016;143:98–109. doi: 10.1016/j.exer.2015.09.017. [DOI] [PubMed] [Google Scholar]
- 92.Toonen JA, Ronchetti A, Sidjanin DJ. A disintegrin and metalloproteinase10 (ADAM10) regulates NOTCH signaling during early retinal development. PLoS One. 2016;11:e0156184. doi: 10.1371/journal.pone.0156184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Totaro A, Castellan M, Battilana G, Zanconato F, Azzolin L, Giulitti S, Cordenonsi M, Piccolo S. YAP/TAZ link cell mechanics to Notch signalling to control epidermal stem cell fate. Nat Commun. 2017;8:15206. doi: 10.1038/ncomms15206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Wan J, Goldman D. Opposing actions of Fgf8a on Notch signaling distinguish two müller glial cell populations that contribute to retina growth and regeneration. Cell Rep. 2017;19:849–862. doi: 10.1016/j.celrep.2017.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Wan J, Ramachandran R, Goldman D. HB-EGF is necessary and sufficient for Müller glia dedifferentiation and retina regeneration. Dev Cell. 2012;22:334–347. doi: 10.1016/j.devcel.2011.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Wang J, Martin JF. Hippo pathway: an emerging regulator of craniofacial and dental development. J Dent Res. 2017;96:1229–1237. doi: 10.1177/0022034517719886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Weinmaster G, Fischer JA. Notch ligand ubiquitylation: what is it good for. Dev Cell. 2011;21:134–144. doi: 10.1016/j.devcel.2011.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Wilson SG, Wen W, Pillai-Kastoori L, Morris AC. Tracking the fate of her4 expressing cells in the regenerating retina using her4:Kaede zebrafish. Exp Eye Res. 2016;145:75–87. doi: 10.1016/j.exer.2015.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wolfe MS. Structure and function of the γ-secretase complex. Biochemistry. 2019;58:2953–2966. doi: 10.1021/acs.biochem.9b00401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.World Health Organization (2019) World report on vision. www.who.int/publications-detail/world-report-on-vision.
- 101.Yoo KW, Thiruvarangan M, Jeong YM, Lee MS, Maddirevula S, Rhee M, Bae YK, Kim HG, Kim CH. Mind bomb-binding partner RanBP9 plays a contributory role in retinal development. Mol Cells. 2017;40:271–279. doi: 10.14348/molcells.2017.2308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Zema S, Pelullo M, Nardozza F, Felli MP, Screpanti I, Bellavia D. A Dynamic role of mastermind-like 1: A journey through the main (path)ways between development and cancer. Front Cell Dev Biol. 2020;8:613557. doi: 10.3389/fcell.2020.613557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Zhang C, Li Q, Jiang YJ. Zebrafish Mib and Mib2 are mutual E3 ubiquitin ligases with common and specific delta substrates. J Mol Biol. 2007a;366:1115–1128. doi: 10.1016/j.jmb.2006.11.096. [DOI] [PubMed] [Google Scholar]
- 104.Zhang C, Li Q, Lim CH, Qiu X, Jiang YJ. The characterization of zebrafish antimorphic mib alleles reveals that Mib and Mind bomb-2 (Mib2) function redundantly. Dev Biol. 2007b;305:14–27. doi: 10.1016/j.ydbio.2007.01.034. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.