

Abstract
The interferon pathway, a key antiviral defense mechanism, is being considered as a therapeutic target in COVID‐19. Both, substitution of interferon and JAK/STAT inhibition to limit cytokine storms have been proposed. However, little is known about possible abnormalities in STAT signaling in immune cells during SARS‐CoV‐2 infection. We investigated downstream targets of interferon signaling, including STAT1, STAT2, pSTAT1 and 2, and IRF1, 7 and 9 by flow cytometry in 30 patients with COVID‐19, 17 with mild, and 13 with severe infection. We report upregulation of STAT1 and IRF9 in mild and severe COVID‐19 cases, which correlated with the IFN‐signature assessed by Siglec‐1 (CD169) expression on peripheral monocytes. Interestingly, Siglec‐1 and STAT1 in CD14+ monocytes and plasmablasts showed lower expression among severe cases compared to mild cases. Contrary to the baseline STAT1 expression, the phosphorylation of STAT1 was enhanced in severe COVID‐19 cases, indicating a dysbalanced JAK/STAT signaling that fails to induce transcription of interferon stimulated response elements (ISRE). This abnormality persisted after IFN‐α and IFN‐γ stimulation of PBMCs from patients with severe COVID‐19. Data suggest impaired STAT1 transcriptional upregulation among severely infected patients may represent a potential predictive biomarker and would allow stratification of patients for certain interferon‐pathway targeted treatments.
Keywords: STAT1, IRF9, Type I interferon, COVID‐19, pSTAT1
Anomalous upregulation of STAT1 and IRF9 (key components of IFN signaling) in B and T cells and monocytes from patients with severe COVID‐19, with absence of pSTAT1 upregulation upon IFN restimulation. Mild COVID‐19 group had strong STAT1 and IRF9 upregulation.
Introduction
Following viral infection, a complex regulatory system of innate and adaptive immune mechanisms is initiated to defend against viral invasion. One of many responses to viral infection is the induction of the pleiotropic cytokine IFN [1]. It acts as a key link between the innate and adaptive immune system. IFN‐α (type I IFN) is mainly secreted by plasmacytoid DCs, while IFN‐γ (type II IFN) is predominantly produced by NK cells and by certain T cells and macrophages. Both, type I and type II IFN, have diverse but complementary antiviral effects such as induction of apoptosis and activation of macrophages, NK cells, as well as B and T lymphocytes [2, 3].
Regarding severe acute respiratory syndrome coronavirus type 2 (SARS‐CoV‐2) infection, IFN‐related antiviral response attracted attention since inborn errors of type I IFN and the presence of autoantibodies against type I IFN were found to be associated with a severe course of the disease [4, 5]. Roughly, 10% of coronavirus disease 2019 (COVID‐19) patients with severe pneumonia produce neutralizing autoantibodies against IFN‐α, IFN‐ω, or both, while patients with no or mild disease have no detectable autoantibodies [4]. Further, it has been shown that patients with severe COVID‐19 have a highly impaired IFN type I signature, with reduced IFN‐α production and activity [6].
Type I IFNs signal through JAK and STAT pathway and thereby stimulate gene expression. After specific binding to the IFN‐α receptor (IFNAR), consisting of two chains (IFNAR1 and IFNAR2), autophosphorylation of the newly formed complex leads to phosphorylation of the receptor‐associated JAK1 and Tyrosine Kinase 2 (TYK2) [7, 8]. Subsequently, further phosphorylation of cytoplasmatic STAT1 and STAT2 induces its dimerization and interaction with IFN regulatory factor 9 (IRF9), a strong enhancer of translocation to the nucleus [9, 10, 11]. The activation of STAT1 is achieved by tyrosine phosphorylation on Y701 that is followed by nuclear accumulation [12].
The formation of a complex called IFN‐stimulated gene factor 3 (ISGF3) binding to IFN‐stimulated response elements (ISRE) finally results in stimulation of gene expression [12]. Likewise, the same sequence of biochemical processes leads to the transcriptional influence of IFN‐γ (type II IFN), involving the IFN‐γ receptor, JAK1 and JAK2, and dimerization of the homodimer STAT1 binding to IFN‐γ‐activated site (GAS) [1]. Figure 1A schematically depicts JAK/STAT signaling. Although it has been shown that the virus proteins NSP5, ORF7a, N, and ORF6 are able to directly interfere with JAK/STAT signaling by inhibiting translocation of STATs or inhibiting their phosphorylation in SARS‐CoV‐2‐infected cultured cells [13, 14], it is not fully understood how JAK/STAT signaling is altered in immune cells and if a dysbalance in JAK/STAT signaling may contribute to disease severity.
Figure 1.
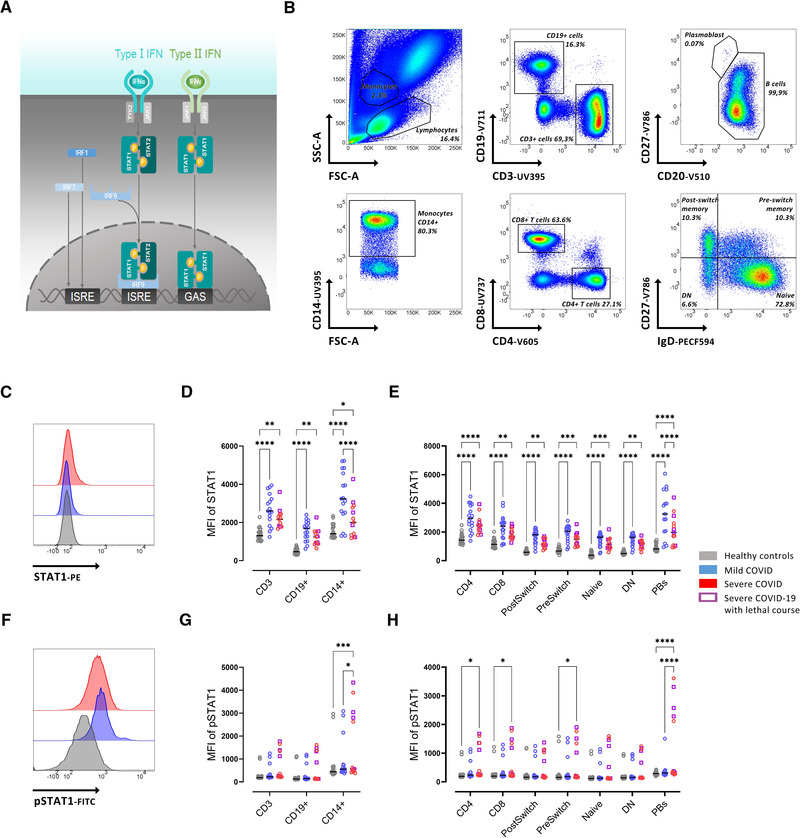
Reduced STAT1 expression in severe COVID‐19 patients. (A) Schematic depiction of JAK/STAT signaling. (B) Gating strategy on whole blood flow cytometry for IgD+CD27‐ (Naïve), IgD+CD27+ (PreSwitch), IgD‐CD27+ (PostSwitched), and IgD‐CD27‐ (Double Negative, DN) as well as CD4+ and CD8+ T cells. (C) Representative histograms of baseline expression of STAT1 on B cells from healthy controls (grey), mild (blue), and severe (red) COVID‐19 patients. (D) Median fluorescence intensity (MFI) of STAT1 in CD3+, CD19+, and CD14+ cells. (E) MFI of STAT1 in T‐ and B‐ cell subsets (as described in B). (F) Representative histograms of baseline expression of pSTAT1 on B cells from healthy controls (grey), mild (blue), and severe (red) COVID‐19 patients. (G) MFI of STAT1 in CD3+, CD19+, and CD14+ cells. (H) MFI of pSTAT1 in T‐ and B‐cell subsets (as described in B). Median and data from healthy controls (n = 20), mild COVID‐19 (n = 17), and severe COVID‐19 (n = 13) patients. (B‐H) Data shown are representative from nine independent experiments. Two‐way ANOVA with Sidack post‐test. *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Deceased patients are indicated as purple quadrats
Here, we report an increased STAT1 expression in mild and severe COVID‐19 patients compared to controls. Severe COVID cases showed lower STAT1 expression than mild patients, accompanied by elevated phosphorylation of STAT1 at the pY701 phosphosite suggesting a disturbance of signal transduction related to impaired STAT1 transcription, which is not surmountable by additional IFN stimulation.
Results
Cohort characteristics
In the current cohort, we included 17 outpatients with mild COVID‐19 during their quarantine (WHO 8‐point ordinal scale 1 and 2) and 13 hospitalized patients treated for severe COVID‐19 pneumonia on an Intensive Care Unit (ICU) (WHO 8‐point ordinal scale ≥ 4) (Table 1) [15]. Age and days post‐symptom onset were not significantly different between both groups, while most patients with mild COVID‐19 were female. Among patients with severe disease manifestation, six patients subsequently died because of COVID‐19.
Table 1.
Patient characteristics
| Mild COVID (n = 17) | Severe COVID (n = 13) | Healthy controls (n = 20) | |
|---|---|---|---|
| Age (median, IQR) | 47.4 (34.0; 57.0) | 61 (53.5; 69.5) | 35 (29.0;47.0) |
| Female | 7 | 1 | 14 |
| Days post‐symptom onset | 7 (10; 5.5) | 10 (8.5; 15) | |
| Concomitant antibiotic treatment | 0 | 6 | |
| Immunosuppressive treatment before COVID | 4 | 1 | |
| Dexamethasone | 0 | 12 | |
| Tocilizumab | 0 | 1 | |
| Remdesivir | 0 | 1 | |
| CRP mg/dL | n.a. | 116 (85.75; 188) | |
| WHO Ordinal scale 1 | 2 | ||
| WHO Ordinal scale 2 | 15 | ||
| WHO Ordinal scale 5 | 6 | ||
| WHO Ordinal scale 6 | 2 | ||
| WHO Ordinal scale 8 | 5 |
Reduced STAT1 expression in patients with severe COVID‐19
Since severe COVID‐19 has a highly impaired IFN type I signature, with especially reduced IFN‐α production and activity [16], we initially asked how the major effector downstream targets of type I IFN are affected among patients with COVID‐19. We found a significantly increased STAT1 protein expression in all analyzed cell subsets from whole blood analysis, including T cells (CD4+ and CD8+), B cells (IgD+CD27− (Naïve), IgD+CD27+ (PreSwitched), IgD−CD27+ (PostSwitched), and IgD−CD27− (double negative), plasmablasts (CD20lowCD27high), and monocytes (CD14+) from all patients with COVID‐19 compared to healthy controls (Fig. 1C‐E). Most notably, reduced STAT1 was observed in severe COVID‐19 cases compared to mild COVID‐19 cases, in particular, in CD14+ monocytes and plasmablasts, respectively (Fig 1D and E). We also observed an increased STAT2 expression in CD14+, CD4+ cells, and all B cell subsets from patients with mild COVID‐19 compared to controls (Supporting information Fig. S1A). STAT2 expression in plasmablasts and preSwitch B cell was reduced in patients with severe COVID‐19 compared to patients with mild COVID‐19 (Supporting information Fig. S1A).
Since STAT1 signaling is mainly regulated through phosphorylation, we were interested in the phosphorylated form of STAT1. Baseline pSTAT1 was significantly increased in monocytes, CD4+ and CD8+ T cells, preswitched B cells (IgD+CD27+), and plasmablasts from severe COVID‐19 patients compared to healthy controls (Fig. 1F‐H).
To better understand the relation between the expression of full protein STAT1 and phosphorylated protein (pSTAT1), the ratio of pSTAT1/STAT1 was obtained. A reduced ratio of pSTAT1/STAT1 was observed in plasmablasts and CD14+ cells from COVID‐19 patients with mild disease compared to those with severe disease (Supporting information Fig. S1B). No differences in pSTAT2 levels were observed among study groups (Supporting information Fig. S1C). These results suggest that mildly affected COVID‐19 patients increased their STAT1/STAT2 expression, but not their detectable levels of phosphorylation, while severely affected COVID‐19 patients showed a greater increase of phosphorylation in relation to the increase of total STAT1 expression. Thus, there is a substantial impairment to increase STAT1 transcription in severe COVID‐19 that especially affects CD14+ monocytes and plasmablasts.
Reduced IRF9 expression in patients with severe COVID‐19
STAT1 is part of IFN‐mediated viral response, representing a key component of complexes like ISGF3 and GAS, responsible for amplified IFN‐mediated signals [12]. To better understand the nature of STAT1 alteration in patients with COVID‐19, IRF9 expression (one component of ISGF3 and GAS complexes) was evaluated. An increased IRF9 expression was found in CD8+ T cells, IgD+CD27−, IgD+CD27+, IgD−CD27+, and IgD−CD27− B cells, PB and CD14+ cells from patients with mild COVID‐19 compared to healthy controls (Fig. 2B, C). Similar to STAT1, severe COVID‐19 was characterized by reduced IRF9 expression compared to mild COVID‐19, in PB and CD14+ cells (Fig. 2 A‐C). Patients with severe COVID‐19 had lower IRF9 expression compared to mild cases, consistent with a reduced STAT1 expression.
Figure 2.
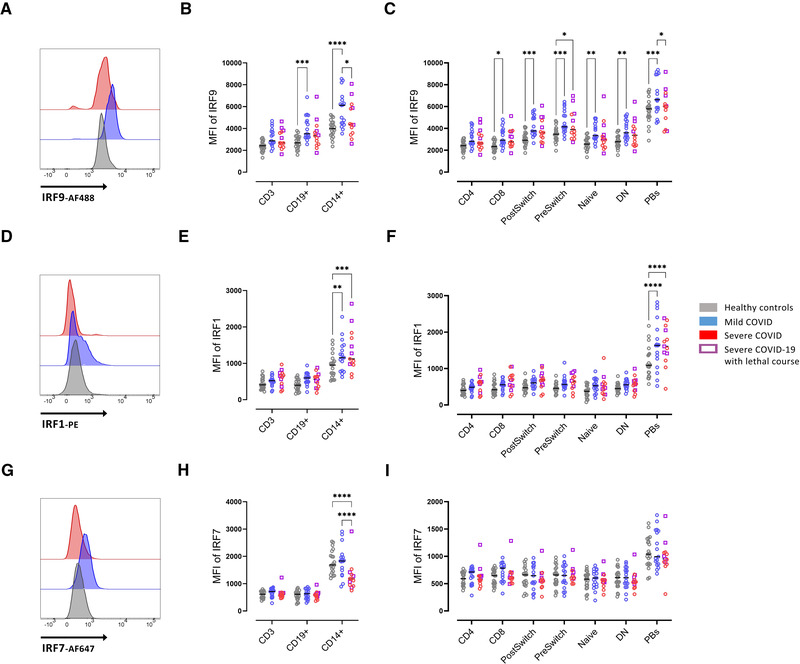
Enhanced intracellular IRF9 expression in severe COVID‐19. (A) Representative histograms of baseline expression of IRF9 on B cells from healthy controls (grey), mild (blue), and severe (red) COVID‐19 patients. (B) Median fluorescence intensity (MFI) of IRF9 in CD3+, CD19+, and CD14+ cells. (C) MFI of IRF9 in T‐ and B‐cell subsets (as described in Fig. 1B). (D) Representative histograms of baseline expression of IRF1 on B cells healthy controls (grey), mild (blue) and severe (red) COVID‐19 patients. (E) MFI of IRF1 in CD3+, CD19+, and CD14+ cells. (F) MFI of IRF1 in T‐ and B‐cell subsets (as described in Fig. 1B). (G) Representative histograms of baseline expression of IRF7 on B cells from healthy controls (grey), mild (blue), and severe (red) COVID‐19 patients. (H) MFI of IRF7 in CD3+, CD19+, and CD14+ cells. (I) MFI of IRF7 in T‐ and B‐cell subsets (as described in Fig. 1B). Median and data from healthy controls (n = 20), mild COVID‐19 (n = 17), and severe COVID‐19 (n = 13) patients. (A‐I) Data shown are representative from nine independent experiments. Two‐way ANOVA with Sidack post‐test. *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Deceased patients are indicated as purple quadrats
Regarding the expression of IRF1, a molecule well known to directly bind and impact the ISRE [12], we found no difference between healthy controls and patients with COVID‐19 irrespective of severity in T cells, and IgD+CD27−, IgD+CD27+, IgD−CD27+, and IgD−CD27− B cells while in CD14+ monocytes and plasmablasts, IRF1 was significantly increased among patients with mild or severe COVID‐19 compared to healthy controls (Fig. 2D‐F).
Expression of intracellular phosphorylated (pS477/pS479) IRF7 did not significantly differ between all groups studied and between the different T‐ and B‐cell populations (Fig. 2G‐I). Only in monocytes, IRF7 was reduced in severely affected COVID‐19 patients compared to mild cases and healthy controls.
Correlation of STAT1 with IFN signature
We wondered how these findings interrelate especially with regard to the IFN‐signature, which was assessed by the expression of the surrogate marker Siglec‐1 (CD169) on monocytes [16]. In our cohort, we observed a significant reduction of Siglec‐1 expression in severely affected COVID‐19 patients (Fig. 3A). Siglec‐1 and STAT1 showed a significant correlation among CD3+ T cells, CD19+ B cells, and monocytes (Fig. 3B), while pSTAT1 did not significantly correlate with Siglec‐1 as a surrogate for IFN signature or age (Fig. 3C). Also, STAT1 and IRF9 both components of the ISGF3 did not correlate (Fig. 3C).
Figure 3.
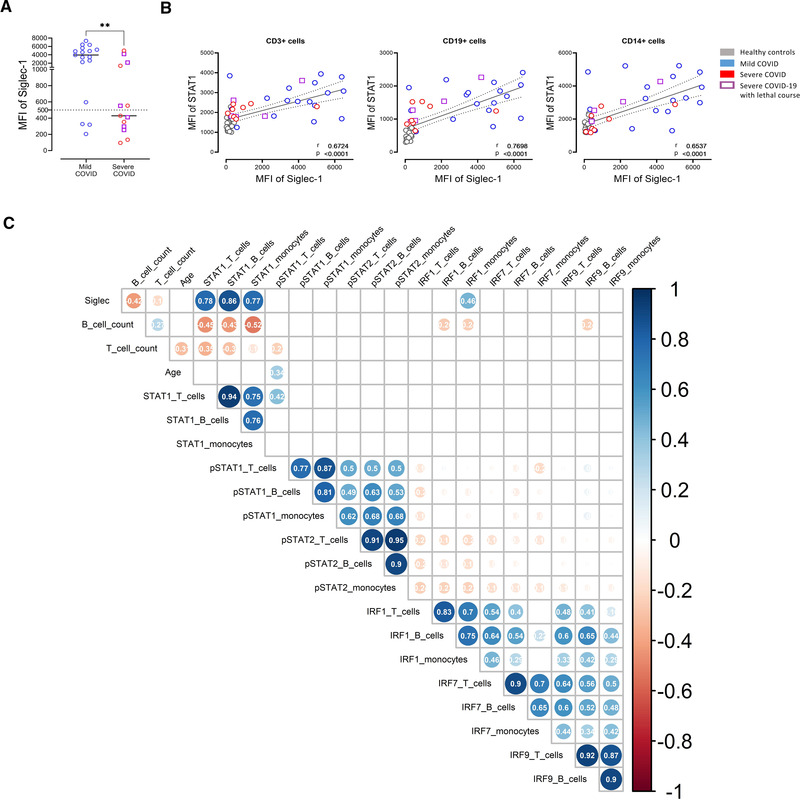
Significant correlation of STAT1 and Siglec‐1 (CD169). (A) Siglec‐1 (CD169) expression on CD14+ monocytes in mild and severe COVID‐19 patients. (B) Correlation of Siglec‐1 expression on the surface of CD14+ monocytes with intracellular STAT1 expression in CD3+ T cells, CD19+ B cells, or CD14 + monocytes. Each point represents a donor. Mann–Whitney U test. *p < 0.05, **p < 0.01. (C) Spearman´s correlation matrix showing the correlation of all investigated parameters (STAT1, pSTAT1, pSTAT2, IRF1, IRF7, and IRF9) in relation to the analyzed cell populations (B cells, T cells, and monocytes). Corresponding correlations are represented by red (negative) or blue (positive) circles. Size and intensity of color refer to the strength of correlation. Data from healthy controls (n = 20), mild COVID‐19 (n = 17), and severe COVID‐19 (n = 13) patients. (A‐C) Data shown are representative from nine independent experiments. Only correlations with p ≤ 0.05 are indicated. Deceased patients are indicated as purple quadrats
Reduced STAT1 response upon IFN stimulation in severe COVID‐19
To further evaluate the functionality of type I and II IFN signaling in the context of our finding of reduced baseline whole protein expression of STAT1, we stimulated PBMCs of healthy controls, mildly and severely affected COVID‐19 patients with low doses of either IFN‐α or IFN‐γ for 48 h. As a control unstimulated cells (RPMI) of each group were used. Stimulation with IFN‐α led to a transcriptional upregulation of STAT1 in CD3+ T cells and CD19+ B cells in healthy controls and in mild COVID‐19 cases in CD19+ B cells (Fig 4A). In severe COVID‐19 cases, a tendency of upregulation of STAT1 was observed (Fig. 4A). The phosphorylation of STAT1 after low‐dose IFN‐α or IFN‐γ stimulation for 48 h remained unchanged in healthy controls and mildly affected COVID‐19 patients, while it was enhanced in severely affected COVID‐19 patients already in unstimulated cells and without further increase after 48 h incubation with IFN‐α or IFN‐γ (Fig. 4B).
Figure 4.
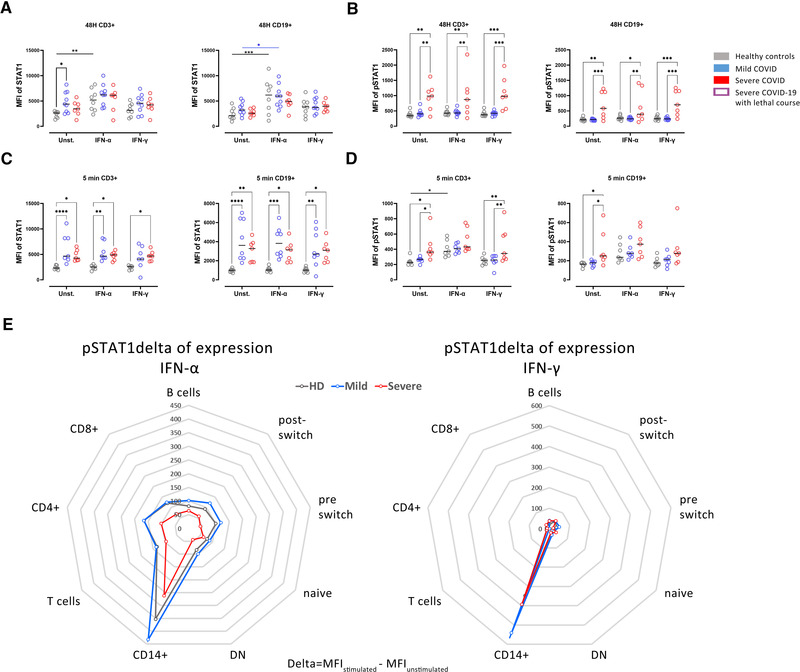
Attenuated pSTAT1 response upon IFN type I and II stimulation in severe COVID‐19. (A) STAT1 and (B) pSTAT1 expression in CD3+ T cells and CD19+ B cells in culture of PBMCs from healthy controls (grey, n = 8), mild (blue, n = 9), or severe (red, n = 7) COVID‐19 patients. Cells were stimulated for 48 h with either IFN‐α (5 ng/mL) or IFN‐γ (5 ng/mL), or only RPMI as a control. (C) STAT1 and (D) pSTAT1 expression in PBMCs of the same donors as in (A) stimulated with IFN‐α (100 ng/mL) or IFN‐γ (100 ng/mL) for 5 min. (E) Delta of pSTAT1 expression in cell populations after stimulation with IFN‐α (left) or IFN‐γ (right) for 5 min in the three study groups. Untreated control values of pSTAT1 were subtracted to show the individual increase of STAT1 phosphorylation. Data are presented as radar diagrams. Two‐way ANOVA with Sidack post‐test. *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001
As expected, a persistent increase of STAT1 expression in CD3+ and CD19+ cells from COVID‐19 patients (both mild and severe) compared to controls was observed in short‐term culture (5 min, Fig. 4C). The short‐term stimulation with higher doses of IFN‐α was able to increase pSTAT1 in CD3+ from controls but not in COVID‐19 cases (Fig. 4D). Also, higher pSTAT1 expression was seen in patients with severe COVID‐19 compared to patients with mild COVID‐19 and controls in unstimulated cultures and IFN‐γ stimulated cultures but not in IFN‐α stimulated cultures in CD3+ cells (Fig. 4D). IFN‐γ stimulation did not impact STAT1 phosphorylation (Fig. 4D, E).
Discussion
COVID‐19 is characterized by excessive production of multiple proinflammatory cytokines [17, 18] and patients with severe COVID‐19 have a highly impaired IFN type I signature, in particular, reduced IFN‐α production and activity [6]. A common signaling route of cytokines including IFNs is the JAK/STAT signaling pathway [19]. JAKs and STATs provide a highly complex but orchestrated system of heterogeneous molecules with specific signaling through defined receptor complexes including recruitment of different STATs and resulting in specific downstream transcription [7].
In this study, we describe an upregulation of STAT1 and IRF9 in mildly and severely affected COVID‐19 patients, which correlated with the IFN‐signature reflected by Siglec‐1 surface expression. Both Siglec‐1 and STAT1 were lower among severely affected COVID‐19 patients compared to mildly affected COVID‐19 patients, especially in plasmablasts and monocytes. This is of particular interest since, these cells are considered the main players of pathogenesis in severe COVID‐19 [20, 21]. The data suggest that certain viral factors may limit proper STAT1 and IRF9 function in severe COVID‐19 patients in these cells and that the inhibition of translocation of STATs is more pronounced than the inhibition of phosphorylation [13, 14]. An alternative explanation is that some individuals react differently to the same viral load, resulting in the detrimental alterations of pSTAT1/IRF9. These results are in accordance with other authors, showing Siglec‐1 expression correlates with viral load in mild COVID patients, but not in severe COVID patients [22].
Increased levels of STAT1 in B cells were previously reported in SLE patients that correlate with Siglec‐1 expression on CD14+ monocytes [23]. In a similar way, SLE patients have increased levels of the STAT1 protein in CD4+ T cells, alteration associated with perturbed homeostasis of Treg and disease severity [24]. Discordant to the whole protein levels, the phosphorylation of STAT1 is enhanced in severely affected COVID‐19 patients suggesting a dysbalanced JAK/STAT signaling that fails to induce transcription of the ISRE. An increased pSTAT1 is also present without stimulation in cultured PBMCS (here CD19+ B cells and CD3+ cells) and no further increase by IFN‐α or IFN‐γ stimulation is achieved. Further, this intervention could not demonstrate a transcriptional increase of STAT1 which would indicate reversibility of this condition. Interestingly, high expression of unphosphorylated STAT2 and IRF9 leads to enhanced IL‐6 expression in response to NF‐κB activators [25], which could be relevant for proper response in COVID‐19, compromised in severe patients.
Interestingly, phosphorylation of STAT1 in response to IFN stimulation can persist for hours, but newly synthesized unphosphorylated STAT1 induced by pSTAT1 can persist for several days [26]. This suggests high levels of unphosphorylated STAT1 in COVID patients were synthesized in response to persistent type I IFN signaling. It is worth to note that cells treated with lentivirus which exhibited an increase in STAT1, did not completely phosphorylate in response to IFN stimulation [27]. The role of STAT1 is not completely dependent on phosphorylation [26, 27], which suggests a role of alternative unphosphorylated STAT1‐mediated pathways in COVID‐19.
The reduced Siglec‐1 expression on the surface of CD14+ monocytes of patients suffering from severe COVID‐19 compared to patients with mild COVID‐19 is consistent with published data [28, 29, 30]. As a result of this finding and efficient analysis of Siglec‐1 on CD14+ cells, we started to routinely measure Siglec‐1 in all patients with COVID‐19 admitted to ICUs to distinguish between patients with high and low IFN signature to test the hypothesis of an association of IFN signature and outcome of the patients.
The absence of correlation between STAT1 and pSTAT1 with IRF9, suggests that an alternative IRF9 independent signaling [12] could have a role in COVID‐19. Previously, it was reported for IRF9 knockout mice that type I IFN mediates a potent inflammatory response associated with a more severe neurological disease [31]. Varicella zoster virus prevents type I IFN response reducing IRF9 and inhibition of STAT2 phosphorylation in infected cells [32]. IRF9 also prevents exhaustion of CD8+ T cells in lymphocytic choriomeningitis infection [33]. Interestingly, SARS‐CoV‐2 spike transfected cells secrete miR‐148a and ‐590 via exosomes which induce degradation of IRF9 in human microglia [34].
Currently, optimal treatment for patients with COVID‐19 is still uncertain and both interventions, blocking of IFN signaling by JAK/STAT inhibition [35, 36] or the use of different types of IFN as substitutions, have been suggested as a treatment for COVID‐19 and showed efficacy in small clinical trials [37, 38]. Thus, the key question raises which intervention is appropriate for which patient? In patients with high IFN signaling that are likely in an early stage of the disease, inhibition strategies appear to be attractive especially during the phase of cytokine storm. On the other hand, patients with an inability to increase antiviral response are likely in severe stage of disease, IFN substitution may hold more promise than further inhibition.
The current data indicate severe impairment of STAT1 and IRF9 in severely infected COVID‐19 patients consistent with inappropriate type IFN upregulation as a potential mechanism for enhanced disease severity. It appears of utmost importance to understand the underlying mechanisms in more detail as patients with diminished type IFN response may benefit from targeted therapies. In this regard, JAK/STAT inhibition may hold promise for patients with higher STAT1 expression (mild cases), while patients with low Siglec‐1 and unable inability to increase antiviral response may benefit from IFN substitution.
Methods
Study participants
Peripheral blood samples (EDTA anticoagulated vaccutainer system, Becton Dickinson Biosciences, BD, NJ, USA) from 20 healthy controls and 30 COVID patients were analyzed, 17 with mild (WHO 8‐point ordinal scale 1 and 2) and 13 with a severe course of the disease (WHO 8‐point ordinal scale ≥ 4) [15]. Donor information is summarized in Table 1. Severe cases were participants in the CAPSID trial that investigated convalescent plasma therapy (EudraCT2020‐001210‐38; NCT04433910I). Samples for this study were collected at baseline prior to administration of the investigation drug. All participants or their legal representatives gave written informed consent according to the approval of the ethics committee at the Charité University Hospital, Berlin (EA2/066/20 Pa‐Covid‐19) and University of Ulm (CAPSID trial (115/20 and 488/20) [39, 40].
Analytical methods and flow cytometry
All procedures adhered to guidelines for the use of flow cytometry and cell sorting in immunological studies [41]. Intracellular phenotyping of STAT1, pSTAT1, pSTAT2, IRF1, and IRF7 levels in B and T cells was conducted as previously published [23]. Briefly, peripheral whole blood (500 μL) was lysed and fixed the same day of acquisition using 5 mL BD phosflow lyse/fix buffer (BD, 10 min, 37°C) considering the manufacturer's protocol (mix of 1:5 in distillated aqua). After two washing steps using ice‐cold PBS Dulbecco (Biochrom GMBH, Berlin, Germany) (530 g, 8 min, 4°C), permeabilization was performed using 200 μL BD Perm Buffer II (BD, 12h, −20°C), followed by overnight storage at −20°C. Next, cells were washed twice with PBS Dulbecco containing 0.5% BSA/EDTA (530 × g, 8 min, 4°C) and resuspended in 50 μL of PBS with 20% Brilliant buffer (BD). After incubation with 2.5 μL of FcR blocking reagent (Miltenyi Biotec, NRW, Germany) for 5 min, staining for 1 h and subsequent flow cytometry analysis were carried out on the same day.
All flow cytometry analyses were performed using a BD FACS Fortessa (BD). To ensure comparable MFIs overtime of the analyses, Cytometer Setup and Tracking beads (BD) and Rainbow Calibration Particles (BD) were used. For flow cytometric analysis, the following fluorochrome‐labeled antibodies were used: BUV395 anti‐CD14 (BD, clone M5E2, 1:50), PE‐Cy7 anti‐CD3 (BD, clone UCHT1, 1:100), BV510 anti‐CD4 (BD, clone SK3, 1:50), BUV737 anti‐CD8 (BD, clone SK1, 1:500), BV711 anti‐CD19 (BD, clone SJ25C1,1:25), BV421 anti‐CD20 (BD, clone 2H7, 1:25), BV786 anti‐CD27 (BD, clone L128, 1:50), PE‐CF594 anti‐IgD (Biolegend, CA, USA, clone IA6‐2, 1:500) for basic immunophenotyping of cell populations of interest. Quantitative analysis was done using the following intracellular fluorochrome‐labeled antibodies: PE anti‐STAT1 (BD, clone 1/Stat1, 1:10), FITC anti‐pSTAT1 (BD, clone 4a, 3:20), AF647 anti‐pSTAT2 (R&D Systems, MN, USA clone 1021D, 1:5), PE anti‐IRF1 (BD, clone 20/IRF‐1, 1:20), AF647 anti‐IRF7 (BD, clone K47‐671, 1:10). For IRF9 and STAT2 analysis, an unconjugated IRF9 antibody (Thermo Fisher, OH, USA isotype rabbit IgG, clone 14H9L22, 1:100) or STAT2 antibody (Cell signaling, PA, USA) was applied. After staining and washing cells, a secondary antibody was used for specific binding of IRF9 or STAT2 (Jackson Immunoresearch, PA, USA, isotype donkey anti‐rabbit IgG, polyclonal, 1:100). Number of absolute B cells was measured with Trucount (BD) and samples were processed according to the manufacturer's instruction. As a control, at least one healthy control sample was processed simultaneously with patients’ samples. Siglec‐1 (CD169) expression analysis on CD14+ monocytes was performed at baseline as previously described [42].
Isolation of PBMCs
PBMCs were obtained by density gradient centrifugation using Ficoll–Paque PLUS (GE Healthcare Bio‐Sciences, IL, USA).
Functional analysis of IFN‐α and IFN‐γ signaling pathways
To evaluate the functional cellular responsiveness upon IFN stimuli, stimulation experiments were carried out as previously described [23]. In brief, we suspended isolated PBMCs in RPMI medium (GlutaMAX, Life Technologies, Paisley, UK) and stimulated with IFN‐α2a (100 ng/mL for 5 min and 5 ng/mL 48 h) (Recombinant Human) or IFN‐γ1b (100 ng/mL for 5 min and 5 ng/mL 48 h) (Recombinant Human, Milteny). Cells were then harvested, washed, lysed, permeabilized, and stained using the same staining as mentioned above followed by flow cytometry.
Statistical analysis
Flow cytometry data were analyzed using FACSDiva software (BD) and FlowJo vX (TreeStar, OR, USA). For graphical and statistical analysis, GraphPad Prism (version 7.00, GraphPad Software, CA, USA) was used. Mann–Whitney tests were used for the comparison of two groups. For multiple group comparison, two‐way ANOVA with Šidák's post‐test for multiple comparison. Spearman correlation coefficient was calculated to detect possible associations between parameters or disease activity, respectively. p‐values less than 0.05 were considered significant. Correlation matrix was calculated using base R and corrplot package (R Foundation for Statistical Computing) using Spearman method.
Ethics Statement
This study was carried out in accordance with the recommendations of the ethics' committees at the Charité University Hospital Berlin and University of Ulm with written informed consent from all subjects.
Patient consent statement
All subjects gave written informed consent in accordance with the Declaration of Helsinki, according to the authorization of the local Ethical Committee.
Conflict of Interest
The authors declare no conflict of interest.
Author Contributions
ES, AA, JR, DK, DZ, and LS enrolled patients and collected samples. HR, ES, FSZ, LS, and AA analyzed the data. SK and HS provided patients within the clinical trial. KUE, TD, AL, ES, and HS supervised the work and acquired funding. All authors developed, read, and approved the current manuscript.
Peer review
The peer review history for this article is available at https://publons.com/publon/10.1002/eji.202149575.
Abbreviations
- COVID‐19
coronavirus disease 2019
- GAS
IFN‐γ‐activated site
- ICU
Intensive care unit
- IFNAR
IFN‐α receptor
- IRF
IFN regulatory factor
- ISGF3
interferon‐stimulated gene factor 3
- ISRE
interferon stimulated response elements
- pSTAT
phosphorylated signal transducer and activator of transcription
- RPMI
Roswell Park Memorial Institute
- SARS‐CoV‐2
severe acute respiratory syndrome coronavirus type 2
- TYK2
Tyrosine Kinase 2
Supporting information
Figure S1 Reduced STAT2 expression in severe COVID‐19 patients. (A) Median fluorescence intensity (MFI) of STAT2 in CD3+, CD19 and CD14+ cells, as well in T and B cell subsets. Median from healthy controls (n = 3), mild COVID‐19 (n = 7) and severe COVID‐19 (n = 5) patients. (B) MFI of pSTAT2 in in CD3+, CD19 and CD14+ cells, as well in T and B cell subsets. Median from healthy controls (n = 20), mild COVID‐19 (n = 17) and severe COVID‐19 (n = 13) patients. (C) Ratio pSTAT/STAT1 in CD3+, CD19 and CD14+ cells, as well in T and B cell subsets. Median from healthy controls (n = 20), mild COVID‐19 (n = 17) and severe COVID‐19 (n = 13) patients. Two way ANOVA with Sidack post‐test. *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001. Deceased patients are indicated as purple quadrats.
Acknowledgements
ES was funded by the Federal Ministry of Education and Research (BMBF) Grant BCOVIT, 01KI20161. ES received a grant by the Berlin Institute of Health with the Charité Clinician Scientist Program funded by the Charité –Universitätsmedizin Berlin and the Berlin Institute of Health. ALS is funded by a scholarship of the German Society of Rheumatology. TD is grantholder of the Deutsche Forschungsgemeinschaft Grants KO 2270/7 1, KO‐2270/4‐1 (KK); Do491/7‐5, 11‐1, Transregio 130 TP24. HRA holds a scholarship of the COLCIENCIAS scholarship No. 727, 2015. The CAPSID trial was funded by the German Federal Ministry of Health (Bundesministerium für Gesundheit) to HS and SK.
Open access funding enabled and organized by Projekt DEAL.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
- 1. Platanias, L. C. , Mechanisms of type‐I‐ and type‐II‐interferon‐mediated signalling. Nat. Rev. Immunol. 2005. 5: 375–386. [DOI] [PubMed] [Google Scholar]
- 2. Vivier, E. , Tomasello, E. , Baratin, M. , Walzer, T. and Ugolini, S. , Functions of natural killer cells. Nat. Immunol. 2008. 9: 503–510. [DOI] [PubMed] [Google Scholar]
- 3. Bonjardim, C. A. , Interferons (IFNs) are key cytokines in both innate and adaptive antiviral immune responses–and viruses counteract IFN action. Microbes Infect. 2005. 7: 569–578. [DOI] [PubMed] [Google Scholar]
- 4. Bastard, P. , Rosen, L. B. , Zhang, Q. , Michailidis, E. , Hoffmann, H. H. , Zhang, Y. , Dorgham, K. et al., Autoantibodies against type I IFNs in patients with life‐threatening COVID‐19. Science 2020. 370: eabd4585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Zhang, Q. , Bastard, P. , Liu, Z. , Le Pen, J. , Moncada‐Velez, M. , Chen, J. , Ogishi, M. et al., Inborn errors of type I IFN immunity in patients with life‐threatening COVID‐19. Science 2020. 370: eabd4570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Hadjadj, J. , Yatim, N. , Barnabei, L. , Corneau, A. , Boussier, J. , Smith, N. , Pere, H. et al., Impaired type I interferon activity and inflammatory responses in severe COVID‐19 patients. Science 2020. 369: 718–724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Shuai, K. , Ziemiecki, A. , Wilks, A. F. , Harpur, A. G. , Sadowski, H. B. , Gilman, M. Z. and Darnell, J. E. , Polypeptide signalling to the nucleus through tyrosine phosphorylation of JAK and STAT proteins. Nature 1993. 366: 580–583. [DOI] [PubMed] [Google Scholar]
- 8. Darnell, J. E., Jr. , Kerr, I. M. and Stark, G. R. , JAK‐STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 1994. 264: 1415–1421. [DOI] [PubMed] [Google Scholar]
- 9. Banninger, G. and Reich, N. C. , STAT2 nuclear trafficking. J. Biol. Chem. 2004. 279: 39199–39206. [DOI] [PubMed] [Google Scholar]
- 10. Paul, A. , Tang, T. H. and Ng, S. K. , Interferon regulatory factor 9 structure and regulation. Front. Immunol. 2018. 9: 1831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Platanitis, E. , Demiroz, D. , Schneller, A. , Fischer, K. , Capelle, C. , Hartl, M. , Gossenreiter, T. et al., A molecular switch from STAT2‐IRF9 to ISGF3 underlies interferon‐induced gene transcription. Nat. Commun. 2019. 10: 2921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Michalska, A. , Blaszczyk, K. , Wesoly, J. and Bluyssen, H. A. R. , A positive feedback amplifier circuit that regulates interferon (IFN)‐stimulated gene expression and controls Type I and Type II IFN responses. Front. Immunol. 2018. 9: 1135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Wu, Y. , Ma, L. , Zhuang, Z. , Cai, S. , Zhao, Z. , Zhou, L. , Zhang, J. et al., Main protease of SARS‐CoV‐2 serves as a bifunctional molecule in restricting type I interferon antiviral signaling. Signal Transduct Target Ther. 2020. 5: 221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Lei, X. , Dong, X. , Ma, R. , Wang, W. , Xiao, X. , Tian, Z. , Wang, C. et al., Activation and evasion of type I interferon responses by SARS‐CoV‐2. Nat. Commun. 2020. 11: 3810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Dodd, L. E. , Follmann, D. , Wang, J. , Koenig, F. , Korn, L. L. , Schoergenhofer, C. , Proschan, M. et al., Endpoints for randomized controlled clinical trials for COVID‐19 treatments. Clin. Trials 2020. 17: 472–482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Biesen, R. , Demir, C. , Barkhudarova, F. , Grun, J. R. , Steinbrich‐Zollner, M. , Backhaus, M. et al., Sialic acid‐binding Ig‐like lectin 1 expression in inflammatory and resident monocytes is a potential biomarker for monitoring disease activity and success of therapy in systemic lupus erythematosus. Arthritis Rheum. 2008. 58: 1136–1145. [DOI] [PubMed] [Google Scholar]
- 17. Lucas, C. , Wong, P. , Klein, J. , Castro, T. B. R. , Silva, J. , Sundaram, M. , Ellingson, M. K. et al., Longitudinal analyses reveal immunological misfiring in severe COVID‐19. Nature 2020. 584: 463–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Mehta, P. , McAuley, D. F. , Brown, M. , Sanchez, E. , Tattersall, R. S. , Manson, J. J. and HLH Across Speciality Collaboration, U.K. , COVID‐19: consider cytokine storm syndromes and immunosuppression. Lancet 2020. 395: 1033–1034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Salas, A. , Hernandez‐Rocha, C. , Duijvestein, M. , Faubion, W. , McGovern, D. , Vermeire, S. , Vetrano, S. et al., JAK‐STAT pathway targeting for the treatment of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020. 17: 323–337. [DOI] [PubMed] [Google Scholar]
- 20. Ferreira‐Gomes, M. , Kruglov, A. , Durek, P. , Heinrich, F. , Tizian, C. , Heinz, G. A. , Pascual‐Reguant, A. et al., SARS‐CoV‐2 in severe COVID‐19 induces a TGF‐beta‐dominated chronic immune response that does not target itself. Nat. Commun. 2021. 12: 1961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Kosyreva, A. , Dzhalilova, D. , Lokhonina, A. , Vishnyakova, P. and Fatkhudinov, T. , The role of macrophages in the pathogenesis of SARS‐CoV‐2‐associated acute respiratory distress syndrome. Front. Immunol. 2021. 12: 682871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Doehn, J. M. , Tabeling, C. , Biesen, R. , Saccomanno, J. , Madlung, E. , Pappe, E. , Gabriel, F. et al., CD169/SIGLEC1 is expressed on circulating monocytes in COVID‐19 and expression levels are associated with disease severity. Infection 2021. 49: 757–762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Aue, A. , Szelinski, F. , Weissenberg, S. Y. , Wiedemann, A. , Rose, T. , Lino, A. C. and Dorner, T. , Elevated STAT1 expression but not phosphorylation in lupus B cells correlates with disease activity and increased plasmablast susceptibility. Rheumatology (Oxford) 2020. 59: 3435–3442. [DOI] [PubMed] [Google Scholar]
- 24. Goropevsek, A. , Gorenjak, M. , Gradisnik, S. , Dai, K. , Holc, I. , Hojs, R. , Krajnc, I. et al., Increased levels of STAT1 protein in blood CD4 T cells from systemic lupus erythematosus patients are associated with perturbed homeostasis of activated CD45RA(‐)FOXP3(hi) regulatory subset and follow‐up disease severity. J. Interferon Cytokine Res. 2017. 37: 254–268. [DOI] [PubMed] [Google Scholar]
- 25. Nan, J. , Wang, Y. , Yang, J. and Stark, G. R. , IRF9 and unphosphorylated STAT2 cooperate with NF‐kappaB to drive IL6 expression. Proc. Natl. Acad. Sci. U. S. A. 2018. 115: 3906–3911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Lehtonen, A. , Matikainen, S. and Julkunen, I. , Interferons up‐regulate STAT1, STAT2, and IRF family transcription factor gene expression in human peripheral blood mononuclear cells and macrophages. J Immunol. 1997. 159: 794–803. [PubMed] [Google Scholar]
- 27. Cheon, H. and Stark, G. R. , Unphosphorylated STAT1 prolongs the expression of interferon‐induced immune regulatory genes. Proc. Natl. Acad. Sci. U. S. A. 2009. 106: 9373–9378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Ortillon, M. , Coudereau, R. , Cour, M. , Rimmele, T. , Godignon, M. , Gossez, M. , Yonis, H. et al., Monocyte CD169 expression in COVID‐19 patients upon intensive care unit admission. Cytometry A 2021. 99: 466–471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Bourgoin, P. , Soliveres, T. , Barbaresi, A. , Loundou, A. , Belkacem, I. A. , Arnoux, I. , Bernot, D. et al., CD169 and CD64 could help differentiate bacterial from CoVID‐19 or other viral infections in the Emergency Department. Cytometry A 2021. 99: 435–445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Bedin, A. S. , Makinson, A. , Picot, M. C. , Mennechet, F. , Malergue, F. , Pisoni, A. , Nyiramigisha, E. et al., Monocyte CD169 expression as a biomarker in the early diagnosis of coronavirus disease 2019. J. Infect. Dis. 2021. 223: 562–567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Hofer, M. J. , Li, W. , Lim, S. L. and Campbell, I. L. , The type I interferon‐alpha mediates a more severe neurological disease in the absence of the canonical signaling molecule interferon regulatory factor 9. J Neurosci. Off. J. Soc. Neurosci. 2010. 30: 1149–1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Verweij, M. C. , Wellish, M. , Whitmer, T. , Malouli, D. , Lapel, M. , Jonjic, S. , Haas, J. G. et al., Varicella viruses inhibit interferon‐stimulated JAK‐STAT signaling through multiple mechanisms. PLoS Pathog. 2015. 11: e1004901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Huber, M. , Suprunenko, T. , Ashhurst, T. , Marbach, F. , Raifer, H. , Wolff, S. , Strecker, T. et al., IRF9 prevents CD8(+) T cell exhaustion in an extrinsic manner during acute lymphocytic choriomeningitis virus infection. J. Virol. 2017. 91: e01219‐17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Mishra, R. and Banerjea, A. C. , SARS‐CoV‐2 spike targets USP33‐IRF9 axis via exosomal miR‐148a to activate human microglia. Front. Immunol. 2021. 12: 656700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Hayek, M. E. , Mansour, M. , Ndetan, H. , Burkes, Q. , Corkren, R. , Dulli, A. , Hayek, R. , Parvez, K. and Singh, S. , Anti‐inflammatory treatment of COVID‐19 pneumonia with tofacitinib alone or in combination with dexamethasone is safe and possibly superior to dexamethasone as a single agent in a predominantly African American cohort. Mayo. Clin. Proc. Innov. Qual. Outcomes 2021. 5: 605–613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Wijaya, I. , Andhika, R. , Huang, I. , Purwiga, A. , Budiman, K. Y. , Bashari, M. H. , Reniarti, L. and Roesli, R. M. A. , The use of Janus Kinase inhibitors in hospitalized patients with COVID‐19: systematic review and meta‐analysis. Clin. Epidemiol. Glob. Health 2021. 11: 100755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Hung, I. F. , Lung, K. C. , Tso, E. Y. , Liu, R. , Chung, T. W. , Chu, M. Y. , Ng, Y. Y. et al., Triple combination of interferon beta‐1b, lopinavir‐ritonavir, and ribavirin in the treatment of patients admitted to hospital with COVID‐19: an open‐label, randomised, phase 2 trial. Lancet 2020. 395: 1695–1704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Andreakos, E. and Tsiodras, S. , COVID‐19: lambda interferon against viral load and hyperinflammation. EMBO Mol. Med. 2020. 12: e12465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Kurth, F. , Roennefarth, M. , Thibeault, C. , Corman, V. M. , Muller‐Redetzky, H. , Mittermaier, M. , Ruwwe‐Glosenkamp, C. et al., Studying the pathophysiology of coronavirus disease 2019: a protocol for the Berlin prospective COVID‐19 patient cohort (Pa‐COVID‐19). Infection 2020. 48: 619–626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Körper, S. , Weiss, M. , Zickler, D. , Wiesmann, T. , Zacharowski, K. , Corman, V. M. , Grüner, B. et al., High dose convalescent plasma in COVID‐19: results from the randomized trial CAPSID. medRxiv 2021. 10.1101/2021.05.10.21256192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Cossarizza, A. , Chang, H. D. , Radbruch, A. , Acs, A. , Adam, D. , Adam‐Klages, S. , Agace, W. W. et al., Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019. 49: 1457–1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Rose, T. , Szelinski, F. , Lisney, A. , Reiter, K. , Fleischer, S. J. , Burmester, G. R. , Radbruch, A. et al., SIGLEC1 is a biomarker of disease activity and indicates extraglandular manifestation in primary Sjogren's syndrome. RMD Open 2016. 2: e000292. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1 Reduced STAT2 expression in severe COVID‐19 patients. (A) Median fluorescence intensity (MFI) of STAT2 in CD3+, CD19 and CD14+ cells, as well in T and B cell subsets. Median from healthy controls (n = 3), mild COVID‐19 (n = 7) and severe COVID‐19 (n = 5) patients. (B) MFI of pSTAT2 in in CD3+, CD19 and CD14+ cells, as well in T and B cell subsets. Median from healthy controls (n = 20), mild COVID‐19 (n = 17) and severe COVID‐19 (n = 13) patients. (C) Ratio pSTAT/STAT1 in CD3+, CD19 and CD14+ cells, as well in T and B cell subsets. Median from healthy controls (n = 20), mild COVID‐19 (n = 17) and severe COVID‐19 (n = 13) patients. Two way ANOVA with Sidack post‐test. *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001. Deceased patients are indicated as purple quadrats.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.