

Abstract
We reviewed the scientific literature on non-floodplain wetlands (NFWs), freshwater wetlands typically located distal to riparian and floodplain systems, to determine hydrological, physical, and chemical functioning and stream and river network connectivity. We assayed the literature for source, sink, lag, and transformation functions, as well as factors affecting connectivity. We determined NFWs are important landscape components, hydrologically, physically, and chemically affecting downstream aquatic systems. NFWs are hydrologic and chemical sources for other waters, hydrologically connecting across long distances and contributing compounds such as methylated mercury and dissolved organic matter. NFWs reduced flood peaks and maintained baseflows in stream and river networks through hydrologic lag and sink functions, and sequestered or assimilated substantial nutrient inputs through chemical sink and transformative functions. Landscape-scale connectivity of NFWs affects water and material fluxes to downstream river networks, substantially modifying the characteristics and function of downstream waters. Many factors determine the effects of NFW hydrological, physical, and chemical functions on downstream systems, and additional research quantifying these factors and impacts is warranted. We conclude NFWs are hydrologically, chemically, and physically interconnected with stream and river networks though this connectivity varies in frequency, duration, magnitude, and timing.
Keywords: connectivity, function, non-floodplain, river networks, wetlands
INTRODUCTION
Wetlands are landscape elements existing along a continuum of connectivity, climatically and hydrologically interacting with other landscape elements through space and time (Ward 1989; Cohen et al. 2016). Instream, riparian, and floodplain wetlands experience fast and/or near-continuous hydrologically mediated connections and interactions. Wetlands are also connected through extremely slow or diffuse pathways (e.g., deep groundwater or atmospherically connected; Rains et al. 2015; Cohen et al. 2016). Researchers are using increasingly advanced and novel methods to quantify wetland locations along this conceptual connectivity continuum (e.g., Jones 2015; Klaus et al. 2015; Vanderhoof et al. 2016; Brooks et al. 2018; Thorslund et al., unpublished data).
Wetlands are functionally part of the landscape, performing hydrological, chemical, and biological functions affecting downstream systems (e.g., Marton, Creed, et al. 2015; Rains et al. 2015; Cohen et al. 2016; Biggs et al. 2017; Schofield et al. 2018). Paradoxically, some functional contributions of wetlands might be inversely related to their rate of connectivity or position along the connectivity continuum (i.e., slower connections could result in larger effects). For instance, wetlands that intercept storm runoff store water and process entr materials (e.g., nutrients). While the subsequent connection with downstream waters may be slow or diffuse (e.g., via groundwater or atmospheric), the functional effect of retaining nutrients and stormwater may be great.
However, functional connections for wetlands are challenging to quantify, requiring elucidation of both the connection pathway and the variable of interest. Descriptors of connectivity include the frequency, duration, magnitude, timing, and rate of change of fluxes to, and exchanges with, downstream waters (Poff et al. 1997; Poff et al. 2007). Current estimates are that ~45 million wetland hectares exist in the conterminous United States (U.S.) (Dahl 2011), and each hectare exists along a continuum of connectivity. There is thus a need to analyze and synthesize the available peer-reviewed scientific literature on the connectivity and quantifiable functional effects of wetlands on other waters to establish a baseline understanding of the interchange of materials, organisms, and energy among and between landscape elements (e.g., Hynes 1975; Forman 1995). This information, the connectivity and effects of wetlands on other waters, provides management agencies and stakeholders with information germane to the sustainable use and management of aquatic resources. Armed with sufficiently robust scientific information, federal, state, and local agencies can endeavor to establish and promulgate guidelines and regulations designed to safeguard aquatic resource integrity while concurrently providing guidance on wise management and growth.
To address this data need, the USEPA (2015) analyzed and synthesized over 1,300 peer-reviewed publications in a 408-page report, summarizing the state of science on the connectivity between streams, wetlands, and downgradient systems, which is the topic of this featured collection (Alexander et al. 2018). Connectivity as used by USEPA (2015, A-2; Leibowitz et al. 2018) and maintained here in this updated review refers to “[t]he degree to which components of a river system are joined by various transport mechanisms; … determined by the characteristics of both the physical landscape and the biota of the specific system.” The primary transport mechanisms considered in this expanded and updated review are surface water and shallow groundwater flows and the transport and transformation of physical and chemical materials, all of which connect watersheds in four dimensions (i.e., longitudinal, lateral, vertical, and temporal, Ward 1989; see also Covino 2017).
This contribution, as well as others (e.g., Alexander et al. 2018; Fritz et al. 2018; Goodrich et al. 2018; Leibowitz et al. 2018; Schofield et al. 2018), builds on the findings of USEPA (2015) with updated literature and scientific advances in quantifying aquatic system connectivity. While USEPA (2015) addresses wetlands in riparian and floodplain areas as well as wetlands not so situated, this review is specifically focused on hydrological, physical, and chemical connectivity of non-floodplain wetlands (NFWs), freshwater wetlands not spatially located within riparian areas or floodplains of river networks, further described below. In the U.S., NFWs may cover more than 6.6 million hectares and comprise >16% of the total freshwater wetland acreage (Lane and D’Amico 2016; Figure 1). Biological connectivity and functions of NFWs are covered in a separate review (Schofield et al. 2018). The review is organized into the following sections: (1) Definitions and Conceptual Approach; (2) Hydrological, Physical, and Chemical Functions of NFWs; and (3) Synthesis and Implications.
FIGURE 1.

Spatial distribution (heat map of ha/km2) of NFWs (or “geographically isolated wetlands,” GIWs in Lane and D’Amico 2016) derived from geographic information system analyses of a floodplain proxy (Lane and D’Amico 2016). Approximately 8.4 million NFWs covering more than 6.6 million ha were reported by Lane and D’Amico (2016). Source: Figure reproduced from Lane and D’Amico (2016). State abbreviations are given in the figure.
EFINITIONS AND CONCEPTUAL APPROACH
Floodplain and Non-Floodplain Wetlands: Definitions
All rivers and streams have riparian areas, defined as transitional zones between terrestrial and aquatic ecosystems distinguished by gradients in biophysical conditions, ecological processes, and organisms. However, not all portions of river and stream networks — the hierarchical, interconnected populations of perennial, intermittent, and ephemerally flowing channels that drain water to a river — have floodplains. Floodplains are level areas bordering stream or river channels that are formed by sediment deposition from those channels under present climatic conditions. Floodplain and associated river channel forms (e.g., meandering, braided, anastomosing) are determined by interacting fluvial factors, including sediment size and supply, channel gradient, and streamflow (Church 2002, 2006). As stream channels increase in size and decrease in slope, a mixture of erosion and deposition processes usually is at work. At some point in the lower portions of river networks, sediment deposition becomes the dominant process and floodplains form. Floodplains are inundated during moderate to high water events (Leopold 1994; Osterkamp 2008).
Like riparian areas, wetlands are transitional areas between terrestrial and aquatic ecosystems. Wetlands include areas such as swamps, bogs, fens, marshes, ponds, and pools (Mitsch et al. 2009). According to Cowardin et al. (1979), an area is classified as a wetland if it has one or more of the following three attributes: (1) the area supports predominantly hydrophytes (i.e., water-loving plants) at least periodically; (2) the land has substrate that is predominantly undrained hydric soil; or (3) the land has nonsoil substrate that is saturated with water or covered by shallow water at some time during the growing season of each year. Note that the Cowardin et al. (1979) definition requires only one of these characteristics, in contrast to the U.S. federal regulatory definition, which requires all three (33 Code of Federal Regulations 328.3(b); see also USACE 1987). Thus, as used in this review, a wetland need not meet the U.S. federal regulatory definition.
Floodplain wetlands are portions of floodplains that meet the aforementioned Cowardin et al. (1979) definition of a wetland. Floodplain wetlands have hydrologic connections to streams and rivers that are predominantly bidirectional (i.e., from wetlands to streams and rivers and vice versa) through lateral movement of surface water and groundwater between the channel and floodplain areas, either through overbank flooding (i.e., lateral expansion of the network) or hyporheic flow (USEPA 2015).
NFWs are found in landscape settings where the potential exists for predominantly unidirectional, lateral hydrologic flows from wetlands (sensu Cowardin et al. 1979) to the river network through surface water or groundwater. Such a setting would typically include upgradient areas such as hillslopes or upland areas outside of the floodplain (e.g., Lane et al. 2017). Any wetland setting where water could normally only flow from the wetland toward a river network (i.e., unidirectionally) would be considered a non-floodplain setting, regardless of the magnitude and duration of flows and of travel times (sensu Ward 1989).
Thus, the terms “floodplain” and “non-floodplain” are meant to describe the landscape setting in which wetlands occur and do not refer to a particular Cowardin et al. (1979) wetland type or class. Many wetland types occur in both settings. For example, a palustrine emergent wetland (Cowardin et al. 1979) could be located outside a floodplain, or it could be located within a floodplain and subject to bidirectional hydrologic flows. A wetland that is classified as depressional in the hydrogeomorphic approach could have any combination of inlets and outlets or none at all (Smith et al. 1995). Such a wetland would be grouped with floodplain wetlands if it had both an input and output channel, since it would have bidirectional flow. A depressional wetland with a surface outlet channel and no inlet or with no outlets and inlets, however, would be considered non-floodplain because water could flow downgradient only from the wetland to the river network, and not from a stream to the wetland. For instance, wetlands that serve as origins for streams are grouped with NFWs because they have unidirectional flow through their outlet channel.
Hence, the hydrologic connections with river networks fundamentally differ for floodplain wetlands and NFWs. Hydrologic connections between NFWs and river networks originate via surface water spillage or merging, or near-surface groundwater flow when water inputs exceed outputs (e.g., evapotranspiration, deep groundwater loss) and available storage. A major consequence of the two different landscape settings is that while waterborne materials can be transported only from the NFW to the river network, waterborne materials can be transported both from the floodplain wetland to the river network and from the river network to the floodplain wetland (see Fritz et al. 2018). In the latter case, there is a mutual, interacting effect on the structure and function of both the floodplain wetland and river network. In contrast, a NFW can materially affect a river through the transport of waterborne materials, but the opposite is generally not true. Note that we limit our use of floodplain and non-floodplain landscape settings to the direction of hydrologic flow; the terms cannot be used to describe directionality of geochemical or biological flows. For example, mobile organisms can move between a stream and NFW and vice versa (e.g., Subalusky et al. 2009a, b; Mushet et al. 2013). In Alaska, transport of live salmon or their carcasses from streams to riparian areas by brown bears (Ursus arctos) accounts for more than 20% of riparian nitrogen budgets (Helfield and Naiman 2006). Although this example is in a floodplain setting, it shows how geochemical fluxes can be decoupled from hydrologic flows. Furthermore, we recognize that there may be exceptions to our unidirectional flow paradigm. For instance, flooding events may create conditions that belie the typical unidirectional flow regime. We consider the “normal” or typical flow patterning in determining the flow regime, though that, too, is replete with qualifiers.
Wetland Functions in This Review
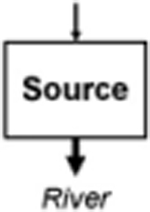
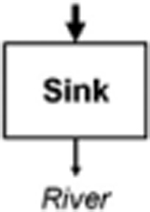
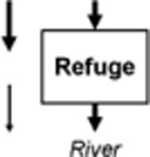
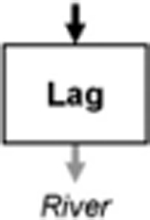
Leibowitz et al. (2008) identified three functions by which wetlands (and streams) influence material fluxes into downstream waters: source, sink, and refuge. USEPA (2015) expanded on this framework to include two additional functions: lag and transformation (Leibowitz et al. 2018). These five functions (summarized and defined in Table 1) provide the conceptual framework for understanding how physical and chemical connections between NFWs and downstream waters influence river systems. This rubric similarly provides a framework to link other landscape elements (Forman 1995) within watersheds (i.e., providing a means to compare functions between streams, floodplain wetlands, and NFWs; USEPA 2015). This review focused on four of the five functions: source, sink, transformation, and lag in NFWs. Schofield et al. (2018) analyze biological connectivity and the refuge function of streams, floodplain wetlands, and NFWs.
TABLE 1.
Functions by which NFWs affect material and energy fluxes to downstream waters.
| Function | Definition | Wetland examples |
|---|---|---|
|
Net increase in a material or energy flux (exports > imports) | Phytoplankton production from floodplain (Schemel et al. 2004; Lehman et al. 2008) |
|
Net decrease in a material or energy flux (exports < imports) | Sediment deposition, denitrification (Johnson 1991) |
|
Avoidance of a nearby sink function, thereby preventing a net decrease in material or energy flux (exports = imports) | Riparian wetlands as aquatic refuges in dryland rivers (Leigh et al. 2010) |
|
Temporary storage and subsequent release of materials or energy without affecting cumulative flux (exports = imports); delivery is delayed and can be prolonged | Flood attenuation (Bullock and Acreman 2003) |
|
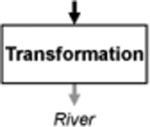
Conversion of a material or energy into a different form; the amount of the base material or energy is unchanged (base exports = base imports), but its composition (e.g., mass of the different forms) can vary | Mercury methylation (Galloway and Branfireun 2004; Selvendiran et al. 2008) |
Source: Table reproduced from USEPA (2015).
Note: This review focused on source, sink, lag, and transformation functions; see Schofield et al. (2018) for refuge functions.
Literature Review Approach
We focused entirely on peer-reviewed, publicly accessible sources of information about surface water and shallow (or near-surface) groundwater connections and interactions of NFWs that influence the function and condition of downstream surface waters. We conducted keyword searches using terms including the types of waters, connections, and downstream effects of interest (e.g., [wetland* AND [river* OR stream*] AND [connect* OR isolat*]]). We used science citation databases and search engines available through Web of Science and Google Scholar to search primary (original research) and secondary (review) literature. These searches included references citing or cited in relevant publications obtained through initial searches. We additionally reviewed and included citations provided by peer-review panels and in public comments on drafts of the USEPA (2015) report, which we co-authored. In addition, we expanded and updated the findings from USEPA (2015) by incorporating recently published material in this review. Finally, as most authors do not use the aforementioned term “non-floodplain wetland” in describing their study objects, we ascribed wetland typology and setting based on information in the paper and/or our knowledge of the study areas available in the peer-reviewed literature.
HYDROLOGICAL, PHYSICAL, AND CHEMICAL FUNCTIONS OF NFWS
We examined the NFW literature and herein report findings demonstrating source, sink, transformation, and lag functions with potential links to downstream systems. We then review the hydrologic and physical pathways by which NFWs may affect downstream waters. Finally, we couple the two, providing updated analyses and syntheses of our findings from the literature on the individual and cumulative effects of NFWs on downstream waters.
NFW Source Function
Hydrologic Source.
An NFW functions as a hydrologic source to downgradient ecosystems when total water inputs minus outputs (e.g., evapotranspiration, groundwater recharge) exceed the storage capacity of the wetland basin and subsequent outflow connects it via surface water to another aquatic system. This occurs when water inputs to the wetland — contributing area precipitation (including snowmelt) and saturation excess flow or near-surface groundwater discharge — fill the available soil pore space and topographic volume of NFWs. Subsequent inputs minus outputs exceed total wetland storage capacity and flow through a surface or near-surface water pour-point downgradient in what has been termed “fill-and-spill” hydrology (Spence and Woo 2003; Tromp-van Meerveld and McDonnell 2006).
NFWs can serve as the hydrologic sources or origins of headwater streams (Fergus et al. 2017; Fritz et al. 2018) when the unidirectional excess flow continues directly into an adjacent, downgradient stream system (USEPA 2015; Figure 2a). These surface connections can be perennial, intermittent, ephemeral, or episodic. White and Crisman (2016) reported that 90% of headwater streams analyzed across Florida originated from wetlands and seeps (a type of NFW). Perennial streams emanated from wetlands more frequently than impermanent streams, while the opposite was found for seeps (White and Crisman 2016). Seeps, springs, and some wetlands are expressions of groundwater at the surface (e.g., Winter and Rosenberry 1995; Euliss et al. 2004; Hayashi et al. 2016). Morley et al. (2011) reported that NFWs primarily supplied by groundwater discharge (i.e., seeps) provided 40%–80% of the water volume for the streams to which they were connected via perennial surface flow. Tufford (2011) monitored four headwater seepage wetlands in the South Carolina Coastal Plain and found that the water table dynamics varied seasonally and annually, intermittently creating hydrologic sources that connected NFW seeps to a downgradient floodplain through swales (i.e., landscape features lacking defined beds and banks or channels that convey flow especially during storm events or snowmelt; USEPA 2015; e.g., Figure 2b).
FIGURE 2.

Types of hydrologic connections between NFWs and streams or rivers. (a) A wetland connected to a river by surface flow through a headwater stream channel. (b) A wetland connected to a river by surface flow through a nonchannelized swale. (c) A NFW connected to a river by groundwater flow (flowpath can be local, intermediate, or regional). (d) A NFW that is hydrologically isolated from a river. Note that in a–c, flows connecting the wetland and river may be perennial, intermittent, or ephemeral. Source: Figure reproduced from USEPA (2015).
In addition to a wetland filling and spilling and reaching the stream network, wetlands can also fill-and-merge within a basin (Huang et al. 2011; Barton et al. 2013; Leibowitz et al. 2016; Wu and Lane 2016), creating NFW complexes that frequently exchange materials within and among the surface water–connected wetland systems (Figure 3; Wu and Lane 2016; Calhoun et al. 2017). Like fill-and-spill wetlands, fill-and-merge wetlands are hydrologic sources when the maximum capacity of the bounding basin is exceeded and the wetland complex connects via continuous surface water to another aquatic system (Shaw et al. 2012; Wu and Lane 2017). That is, fill-and-merge wetlands become fill-and-spill systems when the internal storage is exceeded and no additional merging can occur within a given basin (see also the discussion on NFW Hydrologic Lag Functions, below).
FIGURE 3.

Conceptual model of simple and complex depressions that merge and spill: (a) profile view, (b) plan view, (c) hierarchical view. A schematic example of fill-and-merge behavior where simple depressions (e.g., Simple depression A, Simple depression B) merge with increasing inundation to form Complex depression D. Additional inundation merges Simple depression C with Complex depression D to create Complex depression E, which “spills” at spill elevation #3 (SE3) to another water body. Source: Wu and Lane (2016); reproduced with permission of Springer.
While part of the hydrologic cycle (see Cohen et al. 2016), water in these wetland complexes may or may not eventually connect as surface water or near-surface groundwater into a stream network. For instance, Wilcox et al. (2011) found that nearly 20% of precipitation that fell on a Texas Gulf Coast complex of depressional wetlands spilled into a nearby water body, the Armand Bayou, over a four-year study period. By contrast, Vanderhoof and Alexander (2016) examined two nonadjacent Landsat imagery path/row combinations in the Prairie Pothole Region (PPR) across both drought and deluge conditions over a 21-year period and reported that under wet conditions, 21% of the wetlands merged with at least one other wetland and 7% of wetlands connected with lakes and streams, most via connecting to other “stepping-stone” wetlands (Vanderhoof and Alexander 2016). Vanderhoof et al. (2016) and Vanderhoof et al. (2017) showed that filling and merging of wetlands, and of wetlands and streams, was a function of landscape position relative to other aquatic systems as well as antecedent and prevailing hydrologic conditions. Brooks et al. (2018) conducted a water isotope analysis of surface water origins in a watershed within the PPR, determining that NFWs cumulatively contributed water to streamflow in 11 of 12 sampling events (based on the surface water evaporative enrichment signal in the stream). They furthermore determined that the surface (or near-surface) water connections between the NFWs and the stream networks occurred year-round and that landscape storage and subsequent downgradient flow sustained flow in the stream network.
The scientific literature we found focused primarily on wetland-to-wetland connectivity, not wetland-to-stream. Nevertheless, the literature on wetland-to-wetland connectivity demonstrates NFWs exhibiting surface water connections with other water bodies, while illustrating the variable nature of these hydrologic connections. For instance, Leibowitz and Vining (2003) reported that fill-and-spill dynamics affected 28% of NFWs within a 40-km2 area in the PPR of North America, including 22 wetland complexes (i.e., two or more connected wetlands) during a wetter-than-average year. Conversely, none of the connections observed by Leibowitz and Vining (2003) existed in a drier year. Bowling et al. (2003) analyzed a 471 km2 Alaskan watershed using synthetic aperture radar and reported inundation and connectivity of wetlands and tundra varied between 15% and 67% over two years, depending on available antecedent surface water storage. Leibowitz et al. (2016) explored longer term connectivity in the PPR, determining that some wetlands were filling and spilling to other wetlands over days to weeks, whereas others connected via fill-and-merge hydrology were surficially connected over decades. Kahara et al. (2009) analyzed typological changes in the abundance of different NFW basins, reporting that wetland merging increased with increasing annual precipitation and this changed the abundance of different NFW types (e.g., temporary, seasonal, and semipermanently inundated or saturated wetlands). For instance, one area of the PPR showed decreases in the areal extent of temporary (−47%) and seasonal (−25%) NFWs with increased precipitation, but these losses were offset by concomitant increases in semipermanent wetland area (+185%).
Hydrologically connected wetlands have different physical and hydrologic source properties than relatively unconnected systems. Cook and Hauer (2007) found wetlands that merged due to snowmelt in a Montana prairie setting had longer hydroperiods and volume, as well as higher specific conductance and primary productivity, than nonmerged wetlands. Similarly, Leibowitz et al. (2016) reported differences in dissolved ions, water levels, and macroinvertebrates between merged wetlands and those that did not merge but connected over shorter durations via fill-and-spill hydrology. These field-based measures may inform future connectivity assessments. For instance, Ali et al. (2017) recently developed an electrical conductivity (EC) indicator of potential groundwater-driven wetland-to-stream connectivity. Similarly, Thorslund et al. (unpublished data) analyzed stream and NFW chloride concentrations across several North American study sites; a mass-balance analysis demonstrated NFWs were active sites of catchment-scale flow-generation, though with abundant local to regional heterogeneity.
NFWs can recharge near-surface and deeper groundwater (see Figure 2c), or function simply as an aquatic system embedded within an upland or terrestrial area and not as a vertical or lateral hydrologic “source” for any other waters (e.g., see Figure 2d, Hayashi et al. 2016; but see Cohen et al. 2016). For instance, Hayashi et al. (1998a, b) demonstrated that a NFW received 30%–60% of snowmelt runoff from its basin, but that summer evapotranspiration at the wetland margin effectively removed an abundance of the runoff into the wetland, leaving little water for deep-groundwater recharge (see NFW Sink Function, below).
Much of the literature on NFWs as hydrologic sources has focused on stream baseflow. Groundwater discharge wetlands connected via overland/near-surface flow to streams, such as fens or seeps (e.g., Tufford 2011), are important sources of stream baseflow (Morley et al. 2011). Moreover, as noted, wetlands can be focal points for groundwater recharge, which may contribute to baseflow. Rains (2011), for example, found that perched and flow-through wetland ponds in southwestern Alaska were sites of net groundwater recharge. Given the high prevalence of ponds on the landscape (Rains 2011), these wetlands could substantially affect stream baseflow via groundwater inputs (see Groundwater Connectivity below).
In summary, NFWs are hydrologically dynamic, and are frequently sources of surface waters to other wetlands and other aquatic bodies. NFWs serve as the hydrologic origins or sources of headwater streams (White and Crisman 2016; Fritz et al. 2018). An abundance of research in the North American PPR has demonstrated fill-and-spill and fill-and-merge behaviors, as well as surface water connections to stream and lake systems (e.g., Vanderhoof and Alexander 2016; Vanderhoof et al. 2016; Vanderhoof et al. 2017; Brooks et al. 2018). NFW fill-and-spill behavior occurs in other physiographic regions (e.g., California vernal pools, Rains et al. 2006; Delmarva Ponds, Jones et al. 2018) suggesting similar controlling landscape and climatological factors. Wetlands that merge or connect with other systems were found to substantially affect the physical and chemical characteristics of the receiving water (e.g., Cook and Hauer 2007; Leibowitz et al. 2016; Ali et al. 2017; Brooks et al. 2018).
Chemical Source.
NFWs can be major chemical sources to other waters, contributing dissolved organic compounds (e.g., Mulholland and Kuenzler 1979; Urban et al. 1989; Eckhardt and Moore 1990; Koprivnjak and Moore 1992; Kortelainen 1993; Clair et al. 1994; Hope et al. 1994; Dillon and Molot 1997; Gergel et al. 1999), methylated mercury (MeHg) (St. Louis et al. 1994; Ullrich et al. 2001), nutrients (e.g., phosphorus, Flaig and Reddy 1995; Lane and Autrey 2016), and salts (e.g., Hayashi et al. 1998a, b; Euliss et al. 2004; Heagle et al. 2013; Nachshon et al. 2014; Ali et al. 2017). For instance, boreal forest basins composed of NFWs in central Ontario, Canada, exported between 11.4 and 31.5 kg C/ha/yr as dissolved organic carbon to downstream waters (Creed et al. 2003). While dissolved organic materials such as carbon are important sources of energy for aquatic communities (Hobbie 1992; Reddy and DeLaune 2008), they can have negative effects on downstream waters because contaminants, such as MeHg (see below) can adsorb to dissolved organic matter (Thurman 1985; Driscoll et al. 1995) and under some conditions dissolved organic compound exports can increase the acidity of downstream waters. For instance, Gorham et al. (1986) addressed watershed factors associated with lake and forest acidification in Nova Scotia, Canada. In addition to atmospheric deposition of acid precipitates, they found that the ratio of non-floodplain muskeg peatlands to lakes was significantly correlated with lake acidification, as muskeg wetland–dominated watersheds exported high-molecular-weight organic acids via either overland or shallow groundwater flow. Further linking NFWs to lakes, Gorham et al. (1986) reported that even small amounts of dissolved organic carbon can greatly affect lake water pH; the pH of waters with a dissolved organic carbon value of 4.5 mg/L (the log-normal mean) was 100 times more acidic than waters with a dissolved organic carbon value of <1 mg/L (the minimum concentration).
Wetland soils are important sources of methylmercury, a particularly toxic and mobile form of the element frequently entrained and transferred downgradient with dissolved organic compounds (Grigal 2002). Mercury enters the global atmosphere primarily through waste incineration and coal combustion; it can directly enter wetland systems or can be deposited on terrestrial areas and then transported into riparian areas and wetlands via rainfall and runoff (St. Louis et al. 1994). Ullrich et al. (2001) noted that methylmercury production was linked to low pH, low salinity, and presence of decomposable organic matter in reducing environments, factors that are present in wetlands. The redox conditions found in the presence of a fluctuating water table are thought to be a strong driver of mercury methylation (Heyes et al. 2000; Branfireun and Roulet 2002; Branfireun 2004). Once formed through microbial or other processes in wetlands, mercury and methylmercury export is controlled by the export of organic matter, such as dissolved organic compounds and humic and fulvic acids (Linqvist et al. 1991; Mierle and Ingram 1991; Driscoll et al. 1995). For example, Branfireun et al. (1996) reported 58% of MeHg-laden peat porewater leaving a headwater catchment study area occurred during stormflow, 41% during baseflow, and 1% transported via groundwater. St. Louis et al. (1994) found that boreal forest catchments in Minnesota with NFWs had lower total mercury concentrations than catchments lacking NFWs, but had yields of MeHg from wetlands that were 26–79 times higher than upland areas. This yielded 1.84–5.55 mg MeHg/ha/yr to streams in the Great Lakes basin, where mercury could be incorporated into lake-wide food webs. Porvari and Verta (2003) found that bioaccumulating methylmercury export from non-floodplain peatlands to downstream waters ranged from 0.03 to 3.8 ng MeHg/L, and that catchments with greater wetland abundances had greater methylmercury export.
Nutrients that are mobilized into wetlands via near-surface runoff may create high concentrations in the wetland soils (Flaig and Reddy 1995; Reddy and DeLaune 2008; Hoffmann et al. 2009). The nutrients may stay absorbed to (or bonded with) the soils until a rainfall or precipitation event creates a concentration gradient in the wetland such that the nutrients, such as phosphorus, are released from the soil into the water column (see Chemical Sink function, below). For example, a suggested agricultural nutrient runoff management practice to decrease nutrient loading to Lake Okeechobee, Florida, suggested by Flaig and Reddy (1995) is to use NFWs for phosphorus assimilation. However, they note that the capacity is finite and, furthermore, that the NFWs could become nutrient sources if loading rates are high enough and the wetlands are subsequently connected (e.g., via ditches or major precipitation events). Lane and Autrey (2016, 45) analyzed phosphorus (P) concentrations in NFWs across Florida and concluded that NFWs “… have high potential to retain P, but that the entrained P may be remobilized … depending on storm and groundwater input P concentrations.” Hoos et al. (2013) found nitrate-nitrogen concentrations in streams to be positively associated with the abundance of a likely NFW type (East Gulf Coastal Plain Near-Coast Pine Flatwoods) in their application of the SPARROW (Spatially Referenced Regression on Watershed Attributes) model. They hypothesized that the short-term hydroperiod of the NFWs created aerobic conditions for nitrate formation in wetland soils, and (similar to phosphorus example above) the nutrients were mobilized during precipitation events that connected the wetland to the stream (including shallow groundwater flows).
NFWs can also be sources of elevated salts which, like mercury, can originate in wetland catchments and be entrained or dissolved and hydrologically transported to NFWs. Hayashi et al. (1998a, b) noted that snowmelt in a PPR non-floodplain basin transported 4–5 kg/yr of chloride from the surrounding area to the wetland, suggesting a sink function. However, with growing-season evapotranspiration, the solute also moved from the wetland into the surrounding upland; mass balance equations suggested that 0.1–0.6 kg/yr of chloride was lost to deep groundwater. Nachshon et al. (2014) analyzed 20 years of surface and near-surface salinity data in an upland/NFW setting in Canada’s PPR. They found that NFWs existed across a continuum of salinities (e.g., Euliss et al. 2004), ranging from diluted freshwater systems with EC of <500 μS/cm to brackish systems (EC between 500 and 5,000 μS/cm) to brackish-saline wetlands (EC > 5,000 μS/cm). Summer rainfall flushed accumulated salts from the landscape to NFWs along surface and near-surface flowpaths. Some NFWs became salinity sources to other wetlands when fill-and-spill dynamics (noted above) or near-surface potentiometric heads forced high-density waters downgradient; terminal or receiving wetlands could be considered a chemical sink for high-salinity waters (see Chemical Sink functions, below). These findings suggest that NFWs can be salinity sinks at the landscape scale, but at the local or wetland scale, they may be sources of elevated chloride to the proximal upland areas (Hayashi et al. 1998a, b; Nachshon et al. 2014) or to other systems if they merge (as noted above; see Cook and Hauer 2007; Vanderhoof et al. 2016; Vanderhoof et al. 2017). Interestingly, Nachshon et al. (2014, 1261) noted that if a series of extremely wet years in the PPR was followed by extreme drought, “… salt concentrations in the upper soil horizons of the wetlands [could be found] at concentrations that were never measured before …” with potential impacts on other waters if or when connectivity subsequently occurred through merging or filling.
In summary, biogeochemical characteristics (e.g., high microbial activity in anoxic soils with available carbon) and physical characteristics (e.g., evapotranspiration) in NFWs create conditions conducive to chemical source functions. Examples from the literature on MeHg and salinity, hydrologically exported through dissolved organic matter entrainment or dissolution, demonstrate some of the mechanisms by which processes and functions internal to NFWs may affect other systems (see Factors Affecting Hydrologic Connectivity of NFWs, below). Conditions such as drainage ditches or subsurface tiling that expedite the delivery of NFW-sourced compounds can have negative consequences on receiving systems (e.g., Nair et al. 2015).
NFW Sink Function
Hydrologic Sink.
In addition to serving as a hydrologic source, NFWs can serve as hydrologic sinks when the storage capacity is not exceeded by net inflows (e.g., surface water and groundwater) minus outflows from evaporation or transpiration (e.g., Haan and Johnson 1968; Boelter and Verry 1977; Bullock and Acreman 2003). Storage volumes in NFWs can vary greatly. Field-measured storage volume in 10 NFWs in southern Florida ranged from 900 to 20,488 m3 and averaged 6,836 ± 7,163 m3 (Haag et al. 2005). Lane and D’Amico (2010) used light detection and ranging (LIDAR) data and calculated that over 8,500 NFWs in central Florida had a median storage value of 876 m3 (1,619 m3 average), and that this varied by Cowardin et al. (1979) wetland type. However, Evenson et al. (2016) calculated that ~50% of the >24,000 NFWs in the ~1,800 km2 Pipestem Creek, North Dakota watershed had small storage capacities (<100 m3). Wu and Lane (2016) used LIDAR collected during drought conditions to identify over 12,000 NFW depressions in a North Dakota watershed. They found that surface area in NFWs with standing water averaged over 2 ha and these wetlands currently stored, on average, ~32,000 m3 of water despite a drought during the data collection period. Dry wetlands (74% of the total) averaged two orders of magnitude less storage per wetland, ~366 m3 (Wu and Lane 2016).
Rainfall and snowmelt runoff can flow into NFWs where high evaporation and low discharge rates, coupled with high surface storage capacity, can result in large hydrologic sink functioning at the watershed scale (Shaw et al. 2012). The proportion of NFW area to wetland catchment area (Aw/Ac) is a proxy measure to account for NFW hydrologic sink functions: higher Aw/Ac values suggest greater potential hydrologic sink functioning as more precipitation would be routed to NFWs. Wu and Lane (2017) reported Aw/Ac of ~14% in a study of over 30,000 depressions in the PPR using LIDAR. A large-scale analysis by Watmough and Schmoll (2007) reported Aw/Ac of 8% in the Canadian PPR, similar to the 9% reported by Hayashi et al. (2016). NFWs stored from 11% to 20% of the precipitation in their respective watersheds in a PPR region study by Gleason et al. (2007). Hayashi et al. (2004, 95) analyzed isotopic signatures in a 152 km2 basin in the PPR and found that, “… the majority of [over-winter precipitation storage] is probably stored in wetlands (i.e. fens and bogs) covering 30% of the basin …” Bowling et al. (2003) reported that 24%–42% of snowpack in an Alaskan watershed could be maintained in surface storage (i.e., stored in NFWs).
Evaporation and transpiration losses in particular can be substantial in NFWs. For instance, Lide (1997) estimated that evaporation accounted for 78% of water losses from a wetland system; Duever et al. (1994) noted that evapotranspiration can account for 70%–90% of input precipitation. Van der Kamp et al. (1999) reported that ~75% of input precipitation to a studied Canadian NFW was lost to evapotranspiration at the wetland margins. Bowling et al. (2003) estimated ~50% of snowmelt in an Alaskan basin was evaporated from open water areas (i.e., NFWs).
It is the aboveground volume in NFW wetlands that makes them so effective at landscape water storage. Wetlands effectively store water because their aboveground portions frequently contain a largely empty volume for water storage, in contrast to belowground water storage which is only partly available for water storage due to the presence of soil particles (Johnson 1967; McLaughlin et al. 2014). As noted, stored surface water is acted upon by evaporation, transpiration, and deep groundwater recharge sink functions that result in the water being, in essence, removed from the local — and landscape — hydrologic processes. Among the benefits of water storage in NFWs is that this reduces streamflow (Vining 2002) and the downstream flooding associated with peak streamflow (e.g., Hubbard and Linder 1986; Evenson et al. 2015, 2016; Fossey and Rousseau 2016; Evenson et al., in press). Hubbard and Linder (1986), for example, calculated the water retention capacity of more than 200 non-floodplain prairie pothole wetlands in northeastern South Dakota. They observed that a large amount of snowmelt and precipitation could be cumulatively held by many small wetlands, reducing the potential for flooding at downstream locations. Jones et al. (2018) analyzed NFWs in the Delmarva region (Delaware, Maryland, and Virginia) and identified that 48% of 102,000 depressional NFWs were affected by some sort of ditch; plugging or filling those ditches could increase landscape-scale water storage capacity by 80%. Vanderhoof et al. (in press) analyzed Landsat imagery between 1985 and 2015 covering over 300,000 km2 of the PPR (North and Northwest Drift Plains) and the Northwestern Great Plains (NGP). The NFWs of the PPR had 2.6 times the surface water extent of the NGP (under median climatic conditions), demonstrating that the PPR wetlands were serving, “… a surface water storage function during wet periods, reducing the amount of water contributing to downstream flooding.” Evenson et al. (in press) modeled the hydrologic effects of 13,000 NFWs in the PPR and found that NFWs attenuated peak flows; increased flooding likelihood came from wetland area (and concomitant volume) losses.
Hayashi et al. (1998a, b) determined that NFWs received 30%–60% of snowmelt runoff in their Canadian study area. Similarly, a U.S. Geological Survey study in the PPR found that wetlands, including both depressional (including NFWs) and nondepressional types, stored about 11%–20% of the precipitation that fell in a given watershed, and that storage could be increased by wetland restoration (Gleason et al. 2007). Evenson et al. (2015) simulated the effects of 280 NFWs on watershed hydrology in a North Carolina watershed, finding that NFWs decreased peak flows through the lag function (see below). Vining (2002) concluded that wetland storage in the Starkweather Coulee Subbasin of North Dakota likely resulted in decreased streamflow. Fossey and Rousseau (2016) modeled both riparian and NFW effects, finding NFWs decreased peak flows by 7%–16% depending on the severity of flow (2-, 20-, or 100-year return interval stormflows); NFWs were found to maintain baseflows under drought conditions. Rovansek et al. (1996) found snowmelt to be the most important source of water for wetlands and ponds in the Alaskan Arctic Coastal Plain, and that these wetlands and ponds functioned as surface storage, thereby removing water from snowmelt floods. In contrast, both Roulet and Woo (1986) and Ford and Bedford (1987) found surface storage in cold-weather regions to be limited by frozen soils and ice, limiting the lag (and sink) function in wetlands.
Regression equations developed to predict peak flows during flooding events frequently use lake and wetland storage areas as variables. Using this approach for Wisconsin watersheds, Novitzki (1979) estimated that peak flood flows were only 20% as large in watersheds with 40% lake and wetland area relative to watersheds without lakes or wetlands. Johnston et al. (1990) found that small losses of wetlands in watersheds with <10% wetlands could have major effects on flood flow in basins around Minneapolis, Minnesota. Wang et al. (2010) modeled the influence of wetlands on hydrologic processes in Manitoba and Minnesota and found that the loss of 10%–20% of the wetlands in the study basins would increase peak discharge by 40%. Similarly, Yang et al. (2010) calculated restoration of 600 ha of wetlands in a ~25,000-ha watershed would decrease peak stream discharge by 23%. Peak streamflows were shown to be negatively correlated with lake and wetland storage in Minnesota (Jacques and Lorenz 1988), although a later study found peak flows to be correlated with lake storage only and not wetland storage (Lorenz et al. 2010).
Though NFWs affect floodwaters through hydrologic sink functions, there is a finite capacity to this function. Shaw et al. (2012, 3148) observed, “[h]ighfrequency, low runoff volume [precipitation or snowmelt] events … can be completely or partially attenuated through impoundment by landscape depressions ….” However, Shaw et al. (2012) continued and noted that once a NFW and soil macropore storage volume capacity threshold is exceeded by precipitation and downgradient connections are created by runoff, the contributing area of the basin is substantially increased and storage in wetland depressions has little effect on downstream flood severity. Shaw et al. (2012) found that only 39% of NFWs in a PPR study area contributed to the basin pour-point flow during snowmelt; the remaining 61% of NFWs acted as hydrologic sinks. In a modeling analysis, Evenson et al. (in press) found larger NFWs served as gate-keepers (i.e., wetlands with a high water storage capacity threshold), storing surface runoff and attenuating peak flows in a downstream network. Modeled loss of the larger systems by Evenson et al. (in press) led to larger contributing areas affecting streamflow. Smaller wetlands (<3.0 ha) in Evenson et al. (in press) comprised 95% of the study wetlands but only 35% of the cumulative storage capacity. The modeled effects of losses in small wetland landscape storage resulted in a 90% decrease in landscape residence time, as surface flow following precipitation or snowmelt events drained across the landscape essentially unimpeded by depression water storage.
In summary, the literature regarding hydrologic sink functioning of NFWs is abundant (e.g., Shaw et al. 2012; Shook et al. 2015; Vanderhoof et al., in press). With seasonally dependent high evapotranspiration rates and basin storage, NFWs perform hydrologic sink functions that affect downstream systems, particularly by reducing peak flows. The effects of this function are similar to the lag function (below), which also affects water flow to downgradient pour points. Structural limitations, such as the maximum storage capacity of a NFW or NFW complex, create thresholds affecting hydrologic sink functioning (e.g., Roulet and Woo 1986; Ford and Bedford 1987; Shaw et al. 2012; Evenson et al., in press).
Chemical Sink.
NFWs retain compounds through chemical (and physical) processes such as denitrification, ammonia volatilization, microbial and plant biomass assimilation, sedimentation, sorption and precipitation reactions, biological uptake, and long-term storage in plant detritus (Reddy et al. 1999; Reddy and DeLaune 2008). Marton, Creed, et al. (2015) reviewed the literature on sediment, carbon, phosphorus, and nitrogen storage and transformation in geographically isolated (i.e., non-floodplain) wetlands, concluding (and titling their review) that, “Geographically isolated wetlands are important biogeochemical reactors on the landscape.” Storage or sink rates for NFWs reported from the literature by Marton, Creed, et al. (2015) were: sediment (230–3,600 g/m2/yr), carbon (21–317 g/m2/yr), and phosphorus (0.01–5.0 g/m2/yr) (see NFW Chemical Transformation Function, below for nitrogen findings). The substantial rate variation reported by Marton, Creed, et al. (2015; see also Marton, Chowdhury, et al. 2015) was ascribed to spatial (i.e., size, shape, landscape position) and temporal (i.e., temperature, soil moisture, redox, etc.) variables.
Many studies have addressed chemical sink functions of wetlands, with an abundance of papers focusing on nutrients, namely nitrogen and phosphorus. NFWs inundated with human sewage were shown to remove more than 95% of the phosphorus, nitrate, ammonium, and total nitrogen (Dierberg and Brezonik 1984; Ewel and Odum 1984 and chapters therein). Craft and Chiang (2002) determined that wetland soils stored a disproportionately large share of nitrogen compared with upland soils in spite of uniform soil organic matter across the landscape. A bog NFW in Massachusetts was reported to sequester nearly 80% of the system’s various nitrogen inputs, including precipitation that had a range of 1.2–1.9 mg N/L (Hemond 1983). Hoos et al. (2013) modeled nitrogen and phosphorus dynamics in the southeast U.S. using the SPARROW model and reported the abundance of two typical NFW types (Atlantic Coastal Plain Peatland Pocosin and Southern Coastal Plain Nonriverine Cypress Dome) were factors significantly associated with decreased transport of those nutrients. According to Heagle et al. (2013), the repeated wet–dry cycling of closed-basin NFWs in their PPR study area resulted in substantial subsurface accumulation of sulfate; up to 107 kg was estimated in the soils of a 5-ha PPR NFW. Craft and Chiang (2002) quantified sediment accumulation rates between floodplain and non-floodplain wetlands in southwestern Georgia (951 and 1,289 g/m2, respectively); no difference was found between accumulation rates for organic carbon, total nitrogen, or total phosphorus. Craft et al. (2017) summarized published mean nutrient and sediment storage functions from nine NFW studies and 17 floodplain wetlands; NFW storage of organic carbon was 152% that of floodplain wetlands, but 12%–14% of nitrogen and phosphorous and ~25% that of sediment, though no meta-analysis statistics were conducted. Lane and Autrey (2017) reported that nitrogen and carbon soil accumulation rates in NFWs, measured using cesium-137 radioisotope concentrations in three ecoregions in the U.S., averaged 3.1 ± 3.1 g N/m2/yr and 43.4 ± 39.0 g organic C/m2/yr, respectively. Sediment accretion averaged 0.6 ± 0.4 mm/yr and did not differ by ecoregion, whereas phosphorus accumulation in NFWs was significantly greater in the Erie Drift Plain (0.10 ± 0.10 g P/m2/yr) vs. the Middle Atlantic Coastal Plain (0.01 ± 0.01 g P/m2/yr) or Southern Coastal Plain wetlands (0.04 ± 0.04 g P/m2/yr) (Lane and Autrey 2017). Lane and Autrey (2016) found that phosphorus-sorption capacity varied by Cowardin et al. (1979) wetland type, with forested NFWs having three times the sorption capacity of emergent marsh NFWs (1,275 mg P/kg soil and 418 mg P/kg soil, respectively). In another example, phosphorus retention in NFW marshes of the lower Lake Okeechobee, Florida Basin ranged from 0.3 to 8.0 mg soluble reactive P/m2/day (Dunne et al. 2006). This retention represents a sizeable amount of phosphorus removal, because only about 7% of the watershed was comprised of NFWs. Similarly, NFWs in the Lake Okeechobee Basin were found to have greater storage of total phosphorus than the uplands in which they were embedded (236 kg/ha and 114 kg/ha, respectively; Cheesman et al. 2010). Marton et al. (2014) found that NFWs in Ohio had greater phosphorus-sorption capacities than uplands (297 mg P/kg soil and 86 mg P/kg soil, respectively). These findings were echoed by Dunne et al. (2007), who reported that more phosphorus was stored in NFW plant biomass and soil than in corresponding upland compartments, with wetland surface soils (0–10 cm) representing the largest phosphorus reservoir (>87%) and soil organic matter accounting for >69% of the soil total phosphorus variability. They further suggested that restoring 5%–20% of the NFW area in priority basins draining to Lake Okeechobee, Florida could increase phosphorus storage in NFWs by up to 13 kg P/ha (Dunne et al. 2007). However, Bhadha et al. (2011) found that NFW phosphorus losses from infiltration to groundwater accounted for 14% of phosphorus flux, which suggests that near-surface flow dynamics are also important to managing phosphorus at the landscape level (and reiterating that NFWs may be chemical and hydrologic sources to other systems; Nair et al. 2015). Marton et al. (2014) reported that natural NFWs outperformed both restored and riparian systems in phosphorus retention. However, other studies have also shown that floodplain wetlands have a higher sink capacity than NFWs for phosphorus retention (Craft and Casey 2000; Cohen et al. 2007). Wolf et al. (2013) found greater sedimentation and nitrogen sedimentation in floodplain wetlands vs. NFWs, but reported phosphorus mineralization did not differ between types in floodplain and non-floodplain settings.
Cheng and Basu (2017) conducted a meta-analysis using over 600 published reports on wetland, lake, and reservoir nutrient retention rates. Smaller wetlands were more reactive (i.e., greater retention rates) than larger wetlands and the ratio of sediment contact area to volume was critical to these findings. Cohen et al. (2016, 1980) reported that NFWs are “unambiguously small” and that each order of magnitude increase in wetland size decreases the likelihood that a wetland is a NFW by a factor of three to eight times depending on the ecoregional setting. McCauley et al. (2015) noted that consolidation drainage in the PPR results in landscape changes from many, smaller wetlands to fewer, larger wetlands. Van Meter and Basu (2015), Serran and Creed (2016), and Serran et al. (2017) found that smaller wetlands have been disproportionately lost across the Prairie Pothole landscape. Evenson et al. (in press) found that small NFW removal resulted in decreased inundation time at the landscape scale. Integrating the findings from Marton, Creed, et al. (2015), McCauley et al. (2015), Cohen et al. (2016), Cheng and Basu (2017), and Evenson et al. (in press) suggests biogeochemical functions are affected by the loss of NFWs.
Other pollutants and compounds can be mitigated by NFW sink processes (and/or transformation; see below). For example, microbial methanogenesis completely removed the pesticide atrazine from a mountainous bog in North Carolina (Kao et al. 2002). The environmental contaminants cobalt (Co) and nickel (Ni) were remediated by wetland plants common in forested NFWs; plant concentrations were found to range from 1 to 530 mg Co/kg and up to 250 mg Ni/kg (Brooks et al. 1977). An extensively studied NFW bog in Massachusetts annually stored 54 mg/m2 magnesium, 36 mg/m2 potassium, and 46 mg/m2 lead; the bog also provided acid rain buffering for downstream waters (Hemond 1980).
In summary, the literature is replete with examples of substantive chemical sink functions performed by NFWs. The studies reviewed focused on nutrient compounds (i.e., nitrogen and phosphorus) describing the rates and associated mechanisms across a wide range of NFW types and ecoregions. Factors affecting processing rates include source concentrations and biogeochemical conditions (frequently determined by temporal and spatial conditions and characteristics; e.g., Cheng and Basu 2017; Evenson et al., in press). The collective implication of these studies is that the sink function of NFWs is significant. For instance, in a meta-analysis of NFW biogeochemical functions, Marton, Creed, et al. (2015, 415) estimated that NFW losses in the PPR have negatively affected downstream systems due to lost sink functionality, resulting in “… an increase of between 5 and 140 Tg per year (1 Tg = 1012 g) of sediment entering surface waters and decreases of 0.84–13 Tg per year C sequestration, 0.00040–0.20 Tg per year P storage, and 0.032–0.21 Tg per year [in lost] denitrification potential.” However, intact NFWs, such as the >1.2 million ha identified by Lane et al. (2012) in the southeastern U.S., “… have the potential to sequester 0.25–3.8 Tg of organic C each year … 1.4–42.7 Tg of sediment, [and] 0.00012–0.059 Tg of P …” (Marton, Creed, et al. 2015, 415).
NFW Chemical Transformation Function
Freshwater wetlands and peatlands are active areas for microbially mediated transformations (Boon 2000, 2006) such as the aforementioned mercury methylation and reactive nitrogen (Nr) transformation to N2 through denitrification. Galloway and Cowling (2002, 71) define Nr as, “… biologically active, photochemically reactive, and radiatively active N compounds in the atmosphere and biosphere … [including] inorganic reduced forms of N (e.g., NH3, NH4+), inorganic oxidized forms (e.g., NOx, HNO3, N2O, NO3−), and organic compounds (e.g., urea, amines, proteins).” Transformations of Nr to N2 are prevalent in wetlands that are exposed to Nr and have anoxic conditions, labile carbon, and facultative microbial communities (Reddy and DeLaune 2008). Jordan et al. (2011) reported ~20% of the Nr load reaching wetlands in the contiguous U.S. was removed through plant uptake, denitrification, absorption, burial, and anaerobic ammonium oxidation. As noted, Cohen et al. (2016) determined that wetlands typifying non-floodplain systems (e.g., vernal pools, prairie potholes, etc.) were typically small. With this small size, they found, came a high perimeter length per unit area; since wetland edges are areas of most active biogeochemical processing, these wetland systems are of particular importance in landscapelevel functioning (such as transformative processes; see Cohen et al. 2016; Holgerson and Raymond 2016). Indeed, Ghermandi et al. (2010) found that water quality exported from wetlands was inversely related to size. Cheng and Basu (2017) constructed a nutrient-removal model in wetland systems based on over 600 published studies and found that 50% of the total nitrogen load was removed by wetlands smaller than ~0.03 ha (~300 m2) in size. Losing smaller wetlands, they conclude, “… will lead to a greater fraction of the landscape removal potential lost because of the higher reactivity of smaller wetlands” (5051).
Marton, Creed, et al. (2015) reviewed the literature on biogeochemical processes in NFWs, reporting that denitrification rates ranged from 0.8 to 2.0 g/m2/yr and varied with spatial and temporal variables; input NO3− concentrations were found to also control transformation rates. Racchetti et al. (2011) reported that 60%–100% of NO3− inputs to geographically isolated (i.e., non-floodplain) wetlands were transformed. NFWs in Michigan were found by Whitmire and Hamilton (2008) to remove nitrate-nitrogen (NO3-N) and sulfate (SO42−) at rates of 0.04–0.55 mg NO3-N/L/ha and 0.06–0.30 mg SO42−/L/ha. These rates are significant, considering that nitrate concentration in Michigan groundwater was reported to average 0.5 mg NO3-N/L (Whitmire and Hamilton 2008), though groundwater concentrations >3 mg NO3-N/L are considered contaminated (Madison and Brunett 1985). Prairie pothole NFWs in the upper Midwest removed >80% of the nitrate load via denitrification (Moraghan 1993). A large marsh NFW removed 86% of nitrate, 78% of ammonium, and 20% of phosphate through assimilation and sedimentation, sorption, and other mechanisms, including transformation (Davis et al. 1981). In a study of NFWs of North Carolina and Florida, Lane et al. (2015) reported average denitrification rates of 6.89 ± 5.02 μg N/kg soil dry weight (DW)/h; differences were found based on vegetation structure, with emergent marsh NFWs denitrifying at almost triple the rate of forested NFWs. This was hypothesized to result from the greater abundance of labile carbon in the marsh wetlands (Lane et al. 2015). Marton, Chowdhury, et al. (2015) analyzed NFWs in Indiana and found denitrification rates of 88.8 μg N/kg DW/h. NFWs studied in Maryland and Delaware had microbially mediated denitrification rates of 0.06–0.76 mg N/kg soil/day (Jordan et al. 2007). Because these NFWs comprised >70% of the wetland area in the study basin, this value indicates a significant watershed denitrification capacity. Marton, Chowdhury, et al. (2015) found that NFWs denitrified at twice the rate of upland systems, 12.3 ± 4.5 ng N/g soil/h vs. 5.3 ± 1.7 ng N/g soil/h. NFW samples amended with carbon, nitrogen, or both have been found to dramatically increase the denitrification rate (e.g., Lane et al. 2015; Marton, Chowdhury, et al. 2015) suggesting that NFWs have substantial chemical transformation functioning that may be limited by inputs (see also Ullah and Faulkner 2006; Dodla et al. 2008). Using spatial data from Lane et al. (2012), Marton, Creed, et al. (2015) estimated that the intact NFWs of the southeastern U.S. collectively transform, or denitrify, 0.0095–0.063 Tg N/yr.
In summary, NFWs perform substantial chemical transformation functions, the rates of which frequently depend on factors such as temperature, available energy sources, soil characteristics (e.g., anaerobic or aerobic conditions), and chemical compound concentration (e.g., Marton, Creed, et al. 2015; Marton, Chowdhury, et al. 2015). Much of the literature we found focuses on transformation of nitrogen species to N2, though other compounds were also discussed (e.g., SO42−; MeHg in the chemical source functions, above).
NFW Hydrologic Lag Function
While hydrologic sinks discussed above affect the volume of water on the landscape due to processes such as evapotranspiration, hydrologic lags delay water delivery without substantially affecting quantity through temporary storage or by routing water through flowpaths with long travel times. The lag, sink, and source functions work in conjunction with one another. In essence, the lag function can be viewed as the time-integrated sink and source functions. That is, water initially stored by wetlands can be released over time, reducing peak flows and increasing low or baseflows. Fossey and Rousseau (2016) modeled both riparian and NFW effects, finding NFWs decreased peak flows by 7%–16% depending on the severity of flow (2-, 20-, or 100-year return interval stormflows), while maintaining baseflows under drought conditions. Fossey and Rousseau (2016) concluded that, “… at any event scale any type of wetlands [e.g., floodplain and non-floodplain wetlands] plays a role in reducing peak flows … and sustaining low flows.” As noted above, Evenson et al. (2015) simulated the effects of 280 NFWs on the hydrology in a North Carolina watershed, finding that NFWs decreased peak flows through the lag function.
The lag function is a result of two mechanisms affecting flow. First, a lag occurs when surface and near-surface groundwater inputs to a NFW fill available space below a spill elevation. With lag functions (vs. sink functions), the wetland storage capacity is ultimately exceeded resulting in surface runoff or near-surface flow. When storage capacity is exceeded by storm events during otherwise dry periods, watersheds containing extensive wetlands can require more time for water discharge to rise and fall in response to storm events due to the lag function (Lindsay et al. 2004). Second, a lag occurs through the routing of water to near-surface groundwater and then to stream systems. Water routed through near-surface groundwater-to-stream systems, as opposed to movement via surface water, would be expected to take longer (i.e., to lag) due to the tortuous pathways required by soil particles (and rocks, roots, etc.) — see Groundwater Connectivity, below.
The ability of wetlands to reduce flooding via storage and the lag function varies, with topography exerting an influence (e.g., slope). Using stable hydrogen and oxygen isotopes of water, McEachern et al. (2006) found that snowmelt in boreal forests was discharged rapidly in a sloped watershed. In contrast, in a lowland watershed, much of the snowmelt was stored by wetlands, particularly by bog NFWs with headwater stream channel outlets. In northern Canada, stream runoff was positively correlated with slope and the presence of channel fens, but negatively correlated with lowland depressional NFWs (Quinton et al. 2003). A literature review of wetlands having no direct inlets or outlets to a river system concluded that those wetlands reduced or delayed flooding (Bullock and Acreman 2003). Findings by Bullock and Acreman (2003) were more varied for slope wetlands with direct connectivity to a river: 26 of 62 studies found reduced flooding, while 27 of the 62 studies concluded that those wetlands increased flooding.
Antecedent moisture conditions and currently available wetland storage also affect the lag function. Boelter and Verry (1977) noted that two storms of nearly equal volume and intensity produced different runoff responses from the same peatland. One storm occurring in the spring at a time of already high water tables led to runoff. The other, in midsummer at a time of low water tables, increased the water depth in the NFW but did not exceed the wetland’s water storage capacity, precluding runoff. This mechanism has been observed in simulations of prairie pothole wetland hydrology, in which wetlands reduced streamflow until storage capacity was exceeded (Haan and Johnson 1968; Shaw et al. 2012). Thus, NFWs can function as a sink in dry periods if storage capacity is not exceeded and evaporation rates surpass groundwater recharge (see Hydrologic Sink function, above). These same wetlands can function as sources in wet periods if NFW storage capacity has been exceeded. Finally, the same wetlands can create a lag during the filling process in advance of the spilling event by initially storing inputs but producing surface runoff if their storage capacity is exceeded. This phenomenon explains in part how wetlands can both increase and decrease streamflows (Bullock and Acreman 2003) — see also Factors Affecting Hydrologic Connectivity of NFWs, below.
Thus, NFWs provide hydrologic lag functions when depressions fill with precipitation, snowmelt, or other surface and near-surface inputs. The lag functional is dependent on a NFW basin spill threshold; after such a point is passed, the contributing area transitions from terminating at the NFW to affecting flow in downstream systems (e.g., Boelter and Verry 1977; Shaw et al. 2012; Jones et al. 2018; Brooks et al. 2018; Evenson et al., in press). Hydrologic models have demonstrated that when contributing area flow terminates at a NFW, near-surface and groundwater recharge functions maintain baseflow in downgradient streams (e.g., Evenson et al. 2015; Fossey and Rousseau 2016). Similarly, basin filling from overland or near-surface inputs creates a temporal lag that can decouple stormflow events, even in systems where pervasive overland flow connects landscape elements within watersheds to a downgradient pour-point (Bullock and Acreman 2003; Brooks et al. 2018). However, when NFW storage volume is at capacity, the lag function is effectively bypassed (Shaw et al. 2012).
Factors Affecting Hydrologic Connectivity of NFWs
Surface Water Connectivity.
Multiple factors affect whether a potential surface connection exists between a NFW and another water body and, if and when actualized, the duration, frequency, and magnitude of that connection. These factors can be organized into three major categories: (1) those characteristic of the intervening area between the NFW and the receiving water body; (2) those intrinsic to the NFW or NFW complex itself; and (3) exogenous forcing factors, such as climate. In the first category, intervening distance is a first-order factor (Cohen et al. 2016). Le and Kumar (2014) analyzed NFWs in five study areas across the U.S. and found that potential hydrologic connectivity, as determined by nearest-neighbor distances, followed a universal power law distribution, with most wetlands connecting over short distances. The short intervening distances between wetlands can cause rapid increases in hydrologic connectivity during precipitation/snowmelt events as different parts of the watershed connect (or fill, merge, and spill) and the watershed contributing area increases quickly (Le and Kumar 2014). Neff and Rosenberry (2017) found that the quantity of modeled NFW groundwater provided to the stream network decreased with increasing distance from the network. Vanderhoof et al. (2016) and Vanderhoof and Alexander (2016) found that hydrologic connections between NFWs and streams were positively correlated with stream density, suggesting that connections increase with decreasing distances between wetlands and streams. However, with wet conditions, wetlands were observed connecting to streams over long distances (up to 37 km; Vanderhoof and Alexander 2016). Ameli and Creed (2017) found that both surface water and groundwater inputs from NFWs contributed to river flows, with surface inputs affecting flows up to 8 km from the river system (and groundwater flows affecting the river system up to 30 km away). Human activity can alter this relationship. Historical loss of NFWs in Iowa has increased distances between wetlands that remain (e.g., McCauley et al. 2015; Van Meter and Basu 2015). For example, human activities have also increased the mean distance of Iowa and Nebraska NFWs to streams due to relatively higher rates of drainage and loss of upland-embedded NFWs farther away from the stream network (Uden et al. 2014; Van Meter and Basu 2015).
Other characteristics of the intervening area between NFWs and the stream network include topography, vegetation, soil, and human alterations, all of which can affect flow characteristics. For instance, slope has been positively correlated with streamflow (McEachern et al. 2006), as areas of lower slope typically have a lower potentiometric head. Quinton et al. (2003) reported that stream runoff was negatively correlated with the abundance of lowland bog NFWs. Precipitation events on soils with low infiltration capacity are more likely to result in overland flow whereas soils with higher infiltration capacity are more likely to result in groundwater flowpaths. In their literature review, Bullock and Acreman (2003) found that studies of NFWs with no direct surface water connectivity reduced or delayed flooding downstream. However, 42% (i.e., 26 of 62) of the slope (NFW) wetland studies that were reviewed resulted in reduced flooding and 44% (i.e., 27 of 62) of those increased downstream flooding, which suggest antecedent moisture conditions, wetland storage, and characteristics of the connection between the NFWs and another system (e.g., vegetation, roughness, pore space, hydraulic conductance, etc.) affect connectivity dynamics.
Storage is a function intrinsic to NFWs, affecting connectivity via the fill-and-spill or fill-and-merge of surface waters. As described above, spilling or runoff from a wetland occurs when wetland storage minus losses (via evaporation or groundwater recharge) are exceeded by inputs (via precipitation, groundwater discharge, or overland flow). The inputs and losses are driven in part by climate. Everything being equal, under wetter conditions, wetland storage capacity is more likely to be exceeded. In the PPR, temporary overland connectivity between NFWs has been observed in wet years (e.g., Leibowitz and Vining 2003; Cook and Hauer 2007; Leibowitz et al. 2016). Brooks et al. (2018) found that on average, 33% of the Missouri Coteau (part of the PPR) was connected using Landsat imagery analyses, and determined that this was mostly via the fill-and-merge process between NFWs as the stream network was poorly developed in their study area. Although these studies focused on NFW-to-NFW connections, their findings illustrate that connectivity is a dynamic process, and that NFWs can exhibit temporary surface water connections with other water bodies during wet periods (e.g., Wilcox et al. 2011; Vanderhoof and Alexander 2016; Vanderhoof et al. 2016; Brooks et al. 2018).
Research into the frequency, duration, magnitude, and timing of surface connections of NFWs to river networks has received less attention than those of riparian and floodplain wetlands (see Fritz et al. 2018), though this has changed markedly in recent years through advances in hydrologic modeling, statistical approaches, and remote sensing and geostatistical analyses, especially in the North American PPR and portions of the Atlantic Coastal Plain (e.g., Wilcox et al. 2011; Lang et al. 2012; Golden et al. 2014; Golden et al. 2015; Huang et al. 2014; Evenson et al. 2015; Shook et al. 2015; Fossey and Rousseau 2016; Vanderhoof et al. 2016; Vanderhoof et al. 2017; Ameli and Creed 2017; Jin et al. 2017; Thorslund et al., unpublished data). Where connections have been reported, the duration of a connection is more often described (i.e., whether the connection is perennial, intermittent, or ephemeral). The frequency of a connection, in the case of intermittent or ephemeral flows, or the magnitude of that connection (i.e., the amount of flow between a wetland and a river network) has been less empirically studied or reported due in part to the difficulties in attributing source waters (but see McDonough et al. 2015; Vanderhoof et al. 2016; Brooks et al. 2018; Thorslund et al., unpublished data). A combination of small-scale field-based studies coupled with larger scale remote sensing applications and modeling will effectively quantify surface water connectivity of non-floodplain systems (Golden et al. 2017; see Brooks et al. 2018).
Groundwater Connectivity.
In addition to surface water connections, groundwater flow can connect NFWs with other water bodies, potentially over great distances (e.g., 30 km; Ameli and Creed 2017). Studies have shown NFWs can receive groundwater discharge, contribute to groundwater recharge, or connect via both pathways (Lide et al. 1995; Devito et al. 1996; Matheney and Gerla 1996; Rosenberry and Winter 1997; Pyzoha et al. 2008). For example, a study of four North Dakota prairie NFWs by Arndt and Richardson (1989) demonstrated groundwater connections as one wetland recharged groundwater, one was a flow-through wetland, and one was a groundwater discharge system (Euliss et al. 2004). Using stable isotopes, Matheney and Gerla (1996) concluded that although most of the water in a depressional NFW came from precipitation, groundwater connections accounted for the high salinity of the wetland soil. High salinity can be indicative of net groundwater discharge to a wetland (Brinson 1993; Ali et al. 2017). Min et al. (2010) reported that 38% of rainfall that entered four NFWs in Florida was recharged to groundwater. Bullock and Acreman (2003) found 69 studies making reference to groundwater recharge from wetlands; of these, 32 observed groundwater recharge from a wetland whereas 18 did not. Rosenberry and Winter (1997) determined that groundwater mounding may prevent connections that would otherwise follow topography. Neff and Rosenberry (2017) conducted a model analysis suggesting that groundwater mounding could be short-circuited by the presence of sand layers promoting lateral conductivity or drought conditions decreasing the presence of the water table mounds.
Groundwater flow-through wetlands are sites of both groundwater discharge and recharge, in essence a surface expression of the groundwater system (Richardson et al. 1992; Kehew et al. 1998; Ferone and Devito 2004). This dynamic has been shown in many locations, including prairie pothole NFWs (Richardson et al. 1992), NFWs in glacially formed landscapes in southwest Michigan (Kehew et al. 1998), Alaskan pond NFWs (Rains 2011), Florida cypress dome NFW systems (Sun et al. 1995), and small Wisconsin lakes (Born et al. 1979). The lakes and NFWs of the Nebraska Sand Hills are also predominantly flow-through and an expression of a large regional groundwater system (Winter 1999).
Like surface water, groundwater connections can vary in duration, frequency, and magnitude, and these connections can be affected by a number of factors. Whether a wetland recharges groundwater, is a site of groundwater discharge, or both, is determined by topography, geology, soil features, and seasonal position of the water table relative to the wetland (Neff and Rosenberry 2017). Shedlock et al. (1993), for example, concluded that groundwater discharged into a NFW bog along Lake Michigan through a breach in the sediments underlying the wetland. In dry periods when water tables are low, water tends to move from wetlands into the groundwater, while in wetter periods with higher water tables, water can flow in the opposite direction from shallow groundwater into the wetlands (Phillips and Shedlock 1993; Pyzoha et al. 2008; McLaughlin et al. 2014). Lide et al. (1995) observed both groundwater flow into and from a Carolina bay NFW, with discharge to the wetland when the water table was high and recharge to the groundwater when the water table was low. Sun et al. (1995) observed similar phenomena in a Florida cypress dome NFW. This exchange and temporary storage of water represents a lag function (see NFW Hydrologic Lag Function, above; see also McLaughlin et al. 2014) that can make NFWs particularly important for groundwater recharge during dry periods. Rosenberry and Winter (1997) indicated that groundwater discharge to a wetland often alternates with flow from the wetland to groundwater, and the direction of flow is controlled by the balance of recent precipitation with current evapotranspiration demands.
The magnitude and transit time of groundwater flow from a wetland to other surface waters depends on the intervening distance and the properties of the rock or unconsolidated sediments between the water bodies (i.e., the hydraulic conductivity of the material). In some carbonate or volcanic rocks, for example, groundwater can flow relatively freely through large openings; while in unconsolidated material, such as gravel, sand, silt, or clay, the spaces between particles determine the time required for water to flow a given distance (Winter et al. 2003). In porous material, such as gravel, water can travel a kilometer in a few days; in fine-textured materials, such as silt or clay, hundreds to thousands of years might be required for water to travel the same distance (Winter and LaBaugh 2003).
In agricultural regions, the transit time of subsurface flows is increased substantially by artificial subsurface drainage pipes, known as tile drains (Schiller et al. 2012). Wetlands in these areas are sometimes fitted with inlets that connect directly to tile drains, quickly moving temporarily ponded water through the subsurface and to outlets that discharge directly to ditches or streams (Tomer et al. 2010). Ditching wetlands can not only make them into sources of water but also sources of nutrients and ions from their legacy performing sink functions on the landscape (e.g., Brunet and Westbrook 2012; Nair et al. 2015).
In summary, NFWs can recharge groundwater, receive discharging groundwater, or be flow-through wetlands, and sometimes all three in a given water-year (Sun et al. 1995). Impermeable layers underlying some NFWs may create perched systems with low groundwater connectivity to other systems; these layers may also be breached or be discontinuous underneath larger wetlands, permitting connections to near-surface or deeper groundwater. NFW connections to groundwater encompass sink, source, and lag functions affecting downgradient base and stormflows; groundwater connections can be relatively quick, or ponderously slow. Further examinations into the NFW groundwater connections and factors affecting these interactions and subsequent connections to downstream systems require additional analyses and research, especially in areas outside of the North American PPR.
SYNTHESIS AND IMPLICATIONS: HYDROLOGICAL, PHYSICAL, AND CHEMICAL EFFECTS OF NFWS ON DOWNSTREAM WATERS
NFWs lack well-defined surface water inlets and/or outlets to other water bodies and may include depressions, slopes, flats, and other similar wetland types, as well as regionally described systems such as some Prairie Potholes, playa lakes, vernal pools, and Carolina bays (e.g., Tiner 2003). NFWs are abundant aquatic systems throughout the U.S.; Lane and D’ (2016) conducted a coterminous U.S. spatial analysis using a geospatial floodplain proxy to identify more than 8.4 million potential NFWs covering more than 6.6 million hectares (see Figure 1). Hydrologic flows through NFWs are predominantly unidirectional, in contrast to bidirectional flows that occur in riparian and floodplain wetlands. Much of the updated literature we examined on NFWs indicates that these systems have important hydrologic and water quality functions that affect downstream waters and rivers (Creed et al. 2017). In USEPA (2015), the literature did not sufficiently support a definitive conclusion regarding non-floodplain connectivity of particular groups or classes of wetlands, though USEPA (2015) acknowledged that NFWs which intersect surficial and near-surface runoff can perform substantive chemical sink and transformative functions. Recent scientific contributions to the literature, additional sources now included in this updated review, and advances in hydrologic modeling (e.g., Evenson et al. 2015, 2016; Fossey and Rousseau 2016; Ameli and Creed 2017; Cheng and Basu 2017; Evenson et al., in press; Thorslund et al., unpublished data), remote sensing analyses (e.g., Vanderhoof et al. 2016; Vanderhoof et al. 2017; Jones et al. 2018), field-based observations (e.g., Shaw et al. 2012; Ali et al. 2017), and emerging coupled field and remote sensing studies (e.g., Brooks et al. 2018) have further informed our analyses and synthesis of the literature such that we now conclude all NFWs are interconnected with stream and river networks (Cohen et al. 2016), though this connectivity varies in frequency, duration, magnitude, and timing (Ward 1989; Covino 2017). This complex landscape-scale connectivity, in turn, affects water and material fluxes — the resultants of substantial hydrological, physical, and chemical functioning in NFWs — that modify the characteristics and function of downstream waters (e.g., Marton, Creed, et al. 2015; Rains et al. 2015; Cohen et al. 2016; Calhoun et al. 2017).
It is evident that NFWs that are connected to the river network through surface water are hydrologic sources and will have an influence on downstream waters, regardless of whether the outflow is permanent, intermittent, ephemeral, or episodic (e.g., Wilcox et al. 2011; Rains et al. 2015; Calhoun et al. 2017; Brooks et al. 2018). Such NFWs include wetlands that are the origins of streams or are connected downstream to the river network through ditches, as well as those connected through swales. It is the quality (i.e., concentrations of entrained or dissolved compounds) and quantity of surface water exports from NFWs that informs this conclusion. The finding may have important research and management implications. For instance, White and Crisman (2016) concluded that NFWs are headwaters to ~90% of mapped streams they studied in Florida, and Nadeau and Rains (2007) estimated that up to 60% of stream km in the conterminous U.S. may be first- or second-order systems (i.e., headwaters). However, the most highly refined spatial stream data available for the U.S., the National Hydrography Dataset (NHD; 1:24,000), does not typically identify stream systems of <1.6 km, and there are significant omission errors. For instance, Lang et al. (2012) reported that the NHD identified 66% of stream length compared with a product developed using higher resolution LIDAR data. Thus, it is evident that NFWs serve as abundant stream origins in certain parts of the country, though the origins of the vast majority of streams in the U.S. are unmapped and hence unreported.
NFWs that do not connect to the river network through surface water can still be a source of water (or be connected) through local, intermediate, or regional groundwater flows, often covering great distances (e.g., over 30 km, Ameli and Creed 2017) and providing meaningful hydrologic inputs that maintain baseflows (e.g., 313 m3/day modeled to provide to river flow from groundwater recharging NFWs; Ameli and Creed 2017). Connectivity between these NFWs and downstream waters will vary within a watershed as a function of local factors (e.g., position, topography, and soil characteristics) as well as time, as the river network and water table expand and contract in response to local climate (e.g., Vanderhoof et al. 2016; Vanderhoof et al. 2017).
NFWs that lack a surface or near-surface/groundwater hydrologic connection to other water bodies also influence downstream systems by acting as sinks, creating lags, and/or transforming material. These wetlands can effectively reduce peak flow and flooding, while potentially increasing low flows. The physical storage of water in NFWs, dependent on available volume (e.g., not affected by volume storage reductions due to frozen soils or ice), is incontrovertibly and empirically occurring across the landscape (e.g., Boelter and Verry 1977; Shaw et al. 2012; Brooks et al. 2018). However, once a NFW (or complex) has reached capacity, the storage capacity and impact on downgradient flow mitigation is effectively negligible.
Chemical sink and transformation functions of NFWs have effects on the water quality of downstream waters when these wetlands intersect the flowpath between pollutant source and downstream waters. Wetland chemical sink functions are likely to be greatest when the wetland is located downgradient from pollutant sources and upgradient from a stream or river. The ability of NFWs to perform sink functions is such that maintaining, restoring, and protecting these wetlands have been promulgated as part of watershed nutrient management plans (e.g., Zhang et al. 2009). The literature demonstrates that transformation and sink functions in NFWs provide significant removal of certain pollutants (e.g., transformation of nitrogen species; sink of phosphorus species; but see MeHg studies noted above). The rates depend on pollutant source-area contributions, modes of transportation (i.e., entrained or dissolved), as well as hydrology, temperature, available carbon, and other factors (Marton, Creed, et al. 2015; Marton, Chowdhury, et al. 2015; Cohen et al. 2016; Cheng and Basu 2017).
The updated literature we reviewed provides ample evidence that NFWs can and do provide hydrologic and chemical functions that affect material fluxes to other waters, including other wetlands within a basin, streams and lakes, and groundwater. These results suggest the cumulative influence of many individual NFWs within watersheds can strongly affect the spatial scale, magnitude, frequency, and duration of hydrologic and chemical fluxes or transfers of water and materials to downstream waters.
Caveats associated with this review include the fact that the term “non-floodplain wetland” was infrequently used in the literature, and our designation as such was frequently based on the context, study location, or other information available in the published research that may have resulted in errors of commission. Errors of omission occurred in this review, as the welcome increase in scientific literature on NFW functions portends a better understanding of their downstream effects. Increasing the spatial distribution of NFW studies to encompass the full breadth of ecoregions and wetland typologies will result in increased certainty of the conclusions. Contributions were dominated by studies from researchers of the North American PPR; our understanding will be further improved by additional studies in other areas and ecoregions.
The updated literature supports the following conclusions in this review on the hydrological, physical, and chemical functions of NFWs and their effects on downstream systems:
NFWs exist across a gradient of connectivity from poorly connected systems to well-connected wetlands and wetland complexes.
All NFWs are therefore interconnected with stream and river networks, though this connectivity varies in frequency, duration, magnitude, and timing. This complex landscape-scale connectivity affects water and material fluxes — the resultants of substantial hydrological, physical, and chemical functioning in NFWs — that modify the characteristics and function of downstream waters.
NFWs can be hydrologically connected directly to river networks through natural or constructed channels, nonchannelized surface flows, or subsurface flows, the latter of which can travel long distances to affect downstream waters. Through this connection, NFWs act as a source of water and other materials to downstream waters.
Water storage by NFWs well outside of riparian or floodplain areas affects streamflow through the sink and lag functions.
NFWs act as sinks, sources, and transformers for various pollutants, especially nutrients.
The connectivity of NFWs with other systems, as well as the relative size of their contributing area, may vary in space and time along with their functions that affect watershed-scale hydrodynamics and chemical fluxes.
There are a number of factors that influence the magnitude, frequency, and duration of connections between wetlands and streams. These include spatial proximity, wetland storage capacity, climate, and characteristics of intervening area between wetlands and the stream (e.g., soil permeability).
NFWs are abundant landscape elements important for multiple functions taking place across temporal and spatial scales that contribute to watershed integrity (e.g., Creed et al. 2017). The connectivity of NFWs with other systems, as well as the relative size of their contributing area, may vary in space and time along with their functions that affect watershed-scale hydrodynamics and chemical fluxes. We agree with Shook et al. (2015), as well as Cohen et al. (2016) who in particular note that, “[w]atershed discharge integrates the entire continuum of hydrologic connectivity, not just rapid or surface-connected flow paths.” A corollary to this conclusion is that as hydrology mediates and facilitates much of the physical and chemical functions of NFWs, the material concentration of watershed discharge reflects the time and space-varying contributions and effects of all landscape elements, including NFWs (cf. Hynes 1975).
ACKNOWLEDGMENTS
This review benefited from discussions held at the “North American Analysis and Synthesis on the Connectivity of ‘Geographically Isolated Wetlands’ to Downstream Waters” Working Group supported by the John Wesley Powell Center for Analysis and Synthesis, funded by the U.S. Geological Survey and the U.S. Environmental Protection Agency (USEPA) Office of Research and Development, National Exposure Research Laboratory. We greatly appreciate the technical reviews conducted by Rose Kwok of the USEPA Office of Water. The views expressed in this manuscript are solely those of the authors and do not necessarily reflect the views or policies of the USEPA. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Footnotes
Paper No. JAWRA-17-0073-P of the Journal of the American Water Resources Association (JAWRA).
DISCLOSURE
Former JAWRA Editor Kenneth J. Lanfear served as acting editor in chief for all articles in this featured collection. Parker J. Wigington, Jr., an author on some of the collection papers and who was JAWRA editor in chief at the time the collection was submitted, had no role in the review or editorial decisions for any part of the collection.
Contributor Information
Charles R. Lane, National Exposure Research Laboratory, U.S. Environmental Protection Agency, Cincinnati, Ohio, USA
Scott G. Leibowitz, National Health and Environmental Effects Research Laboratory, U.S. Environmental Protection Agency, Corvallis, Oregon, USA
Bradley C. Autrey, National Exposure Research Laboratory, U.S. Environmental Protection Agency, Cincinnati, Ohio, USA
Stephen D. LeDuc, National Center for Environmental Assessment, U.S. Environmental Protection Agency, Washington, D.C., USA
Laurie C. Alexander, National Center for Environmental Assessment, U.S. Environmental Protection Agency, Washington, D.C., USA.
LITERATURE CITED
- Alexander LC, Fritz KM, Schofield KA, Autrey BC, DeMeester JE, Golden HE, Goodrich DC, Kepner WG, Kiperwas HR, Lane CR, LeDuc SD, Leibowitz SG, McManus MG, Pollard AI, Ridley CE, Vanderhoof MK, and Wigington PJ Jr. 2018. “Featured Collection Introduction: Connectivity of Streams and Wetlands to Downstream Waters.” Journal of the American Water Resources Association 54 (2): 287–297. 10.1111/1752-1688.12630. [DOI] [Google Scholar]
- Ali G, Haque A, Basu NB, and Badiou P. 2017. “Groundwater-Driven Wetland-Stream Connectivity in the Prairie Pothole Region: Inferences Based on Electrical Conductivity Data.” Wetlands 37 (4): 773–85. 10.1007/s13157-017-0913-5. [DOI] [Google Scholar]
- Ameli AA, and Creed IF. 2017. “Quantifying Hydrologic Connectivity of Wetlands to Surface Water Systems.” Hydrology and Earth System Sciences 21: 1791–808. 10.5194/hess-21-1791-2017. [DOI] [Google Scholar]
- Arndt JL, and Richardson JL. 1989. “Geochemistry of Hydric Soil Salinity in a Recharge-Throughflow-Discharge Prairie-Pothole Wetland System.” Soil Science Society of America Journal 53 (3): 848–55. 10.2136/sssaj1989.03615995005300030037x. [DOI] [Google Scholar]
- Barton AB, Herczeg AL, Dahlhaus PG, and Cox JW. 2013. “A Geochemical Approach to Determining the Hydrological Regime of Wetlands in a Volcanic Plain, South-Eastern Australia.” In Groundwater and Ecosystems, edited by Ribeiro L, Stigter TY, Chambel A, de Melo Condesso MT, Monteiro JP, and Medeiros A, 69–79. Boca Raton, FL: CRC Press. [Google Scholar]
- Bhadha JH, Jawitz JW, and Min J. 2011. “Phosphorus Mass Balance and Internal Load in an Impacted Subtropical Isolated Wetland.” Water, Air, & Soil Pollution 218 (1–4): 619–32. 10.1007/s11270-010-0673-9. [DOI] [Google Scholar]
- Biggs J, von Fumetti S, and Kelly-Quinn M. 2017. “The Importance of Small Waterbodies for Biodiversity and Ecosystem Services: Implications for Policy Makers.” Hydrobiologia 793 (1): 3–9. 10.1007/s10750-016-3007-0. [DOI] [Google Scholar]
- Boelter D, and Verry ES. 1977. “Peatland and Water in the Northern Lake States.” U.S. Department of Agriculture, U.S. Forest Service, North Central Forest Experiment Station Report General Technical Report NC-31, St. Paul, MN. [Google Scholar]
- Boon P 2000. “Bacterial Biodiversity in Wetlands.” In Biodiversity in Wetlands: Assessment, Function, and Conservation, edited by Gopal B, Junk WJ, and Davis JA, 281–310. Leiden, The Netherlands: Backhuys Publishers. [Google Scholar]
- Boon P 2006. “Biogeochemistry and Bacterial Ecology of Hydrologically Dynamic Wetlands.” In Ecology of Freshwater and Estuarine Wetlands, edited by Batzer DP and Sharitz RA, 115–76. Berkeley, CA: University of California Press. 10.1525/california/9780520247772.001.0001. [DOI] [Google Scholar]
- Born SM, Smith SA, and Stephenson DA. 1979. “Hydrogeology of Glacial-Terrain Lakes, with Management and Planning Applications.” Journal of Hydrology 43: 7–43. 10.1016/0022-1694(79)90163-X. [DOI] [Google Scholar]
- Bowling LC, Kane DL, Gieck RE, Hinzman LD, and Lettenmaier DP. 2003. “The Role of Surface Storage in a Low-Gradient Arctic Watershed.” Water Resources Research 39 (4): 1087. 10.1029/2002wr001466. [DOI] [Google Scholar]
- Branfireun B 2004. “Does Microtopography Influence Subsurface Pore-Water Chemistry? Implications for the Study of Methylmercury in Peatlands.” Wetlands 24: 207–11. 10.1672/0277-5212(2004)024. [DOI] [Google Scholar]
- Branfireun BA, Heyes A, and Roulet NT. 1996. “The Hydrology and Methylmercury Dynamics of a Precambrian Shield Headwater Peatland.” Water Resources Research 32 (6): 1785–94. 10.1029/96WR00790. [DOI] [Google Scholar]
- Branfireun BA, and Roulet NT. 2002. “Controls on the Fate and Transport of Methylmercury in a Boreal Headwater Catchment, Northwestern Ontario, Canada.” Hydrology and Earth System Sciences 6 (4): 785–94. 10.5194/hess-6-785-2002. [DOI] [Google Scholar]
- Brinson MM 1993. “Hydrogeomorphic Classification for Wetlands.” Final Report. Washington, D.C.: U.S. Army Corps of Engineers. [Google Scholar]
- Brooks JR, Mushet DM, Vanderhoof MK, Leibowitz SG, Christensen JR, Neff BP, Rosenberry DO, Rugh WD, and Alexander LC. 2018. “Estimating Wetland Connectivity to Streams in the Prairie Pothole Region: An Isotopic Approach and Remote Sensing Approach.” Water Resources Research 54 (2): 955–977. 10.1002/2017WR021016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooks RR, McCleave JA, and Schofield EK. 1977. “Cobalt and Nickel Uptake by the Nyssaceae.” Taxon 26: 197–201. [Google Scholar]
- Brunet NN, and Westbrook CJ. 2012. “Wetland Drainage in the Canadian Prairies: Nutrient, Salt and Bacteria Characteristics.” Agriculture, Ecosystems & Environment 146: 1–12. 10.1016/j.agee.2011.09.010. [DOI] [Google Scholar]
- Bullock A, and Acreman M. 2003. “The Role of Wetlands in the Hydrological Cycle.” Hydrology and Earth System Sciences 7 (3): 358–89. 10.5194/hess-7-358-2003. [DOI] [Google Scholar]
- Calhoun AJK, Mushet DM, Alexander LC, DeKeyser ES, Fowler L, Lane CR, Lang MW, Rains MC, Richter SC, and Walls SC. 2017. “The Significant Surface-Water Connectivity of ‘Geographically Isolated Wetlands’.” Wetlands 37 (4): 801–06. 10.1007/s13157-017-0887-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheesman AW, Dunne EJ, Turner BL, and Reddy KR. 2010. “Soil Phosphorus Forms in Hydrologically Isolated Wetlands and Surrounding Pasture Uplands.” Journal of Environmental Quality 39 (4): 1517–25. 10.2134/jeq2009.0398. [DOI] [PubMed] [Google Scholar]
- Cheng FY, and Basu NB. 2017. “Biogeochemical Hotpots: Role of Small Water Bodies in Landscape Nutrient Processing.” Water Resources Research 53: 5038–56. 10.1002/2016WR020102. [DOI] [Google Scholar]
- Church M 2002. “Geomorphic Thresholds in Riverine Landscapes.” Freshwater Biology 47 (4): 541–57. 10.1046/j.1365-2427.2002.00919.x. [DOI] [Google Scholar]
- Church M 2006. “Bed Material Transport and the Morphology of Alluvial River Channels.” Annual Review of Earth and Planetary Sciences 34: 325–54. 10.1146/annurev.earth.33.092203.122721. [DOI] [Google Scholar]
- Clair TA, Pollock TL, and Ehrman JM. 1994. “Exports of Carbon and Nitrogen from River Basins in Canada’s Atlantic Provinces.” Global Biogeochemical Cycles 8: 441–50. 10.1029/94GB02311. [DOI] [Google Scholar]
- Cohen MJ, Creed IF, Alexander LC, Basu N, Calhoun A, Craft CB, D’Amico E, DeKeyser S, Fowler L, Golden HE, Jawitz JW, Kalla P, Kirkman LK, Lane CR, Lang MW, Leibowitz SG, Lewis DB, Marton JM, McLaughlin DL, Mushet D, Raanan-Kiperwas H, Rains MC, Smith L, and Walls S. 2016. “Do Geographically Isolated Wetlands Influence Landscape Functions?” Proceedings of the National Academy of Sciences of the United States of America 113 (8): 1978–86. 10.1073/pnas.1512650113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen MJ, Paris J, and Clark MW. 2007. “P-Sorption Capacity Estimation in Southeastern USA Wetland Soils Using Visible/Near-Infrared (VNIR) Reflectancy Spectroscopy.” Wetlands 27 (4): 1098–111. 10.1672/0277-5212(2007)27[1098:PCEISU]2.0.CO;2. [DOI] [Google Scholar]
- Cook BJ, and Hauer FR. 2007. “Effects of Hydrologic Connectivity on Water Chemistry, Soils, and Vegetation Structure and Function in an Intermontane Depressional Wetland Landscape.” Wetlands 27 (3): 719–38. 10.1672/0277-5212(2007)27[719:EOHCOW]2.0.CO;2. [DOI] [Google Scholar]
- Covino T 2017. “Hydrologic Connectivity as a Framework for Understanding Biogeochemical Flux Through Watersheds and Along Fluvial Networks.” Geomorphology 277: 133–44. 10.1016/j.geomorph.2016.09.030. [DOI] [Google Scholar]
- Cowardin LM, Carter V, Goulet FC, and LaRoe ET. 1979. Classification of Wetlands and Deepwater Habitats of the United States. Washington, D.C.: U.S. Fish and Wildlife Service. [Google Scholar]
- Craft CB, and Casey WP. 2000. “Sediment and Nutrient Accumulation in Floodplain and Depressional Freshwater Wetlands of Georgia, USA.” Wetlands 20 (2): 323–32. 10.1672/0277-5212(2000)020[0323:SANAIF]2.0.CO;2. [DOI] [Google Scholar]
- Craft CB, and Chiang C. 2002. “Forms and Amounts of Soil Nitrogen and Phosphorus Across a Longleaf Pine-Depressional Wetland Landscape.” Soil Science Society of America Journal 66 (5): 1713–21. 10.2136/sssaj2002.1713. [DOI] [Google Scholar]
- Craft CB, Vymazal J, and Kröpfelová L. 2017. “Carbon Sequestration and Nutrient Accumulation in Floodplain and Depressional Wetlands.” Ecological Engineering. 10.1016/j.ecoleng.2017.06.034. [DOI] [Google Scholar]
- Creed IF, Lane CR, Alexander L, Basu N, Calhoun AJK, Cohen MJ, Craft C, D’Amico E, DeKeyser E, Fowler L, Golden HE, Jawitz JW, Kalla P, Kirkman LK, Lang MW, Leibowitz SG, Lewis DB, Marton J, McLaughlin DL, Raanan-Kiperwas H, Rains MC, Smith L, and Serran JN. 2017. “Enhancing Protection for Vulnerable Waters.” Nature Geoscience 10: 809–15. 10.1038/ngeo3041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Creed IF, Sanford SE, Beall FD, Molot LA, and Dillon PJ. 2003. “Cryptic Wetlands: Integrating Hidden Wetlands in Regression Models of the Export of Dissolved Organic Carbon from Forested Landscapes.” Hydrological Processes 17 (18): 3629–48. 10.1002/hyp.1357. [DOI] [Google Scholar]
- Dahl TE 2011. Status and Trends of Wetlands in the Conterminous United States 2004 to 2009. Washington, D.C.: U.S. Department of the Interior, Fish and Wildlife Service. [Google Scholar]
- Davis CB, Baker JL, van der Valk AG, and Beer CE. 1981. “Prairie Pothole Marshes as Traps for Nitrogen and Phosphorus in Agricultural Runoff.” In Proceedings of the Midwest Conference on Wetland Values and Management, edited by Richardson B, 153–63. Navarre, MN: Freshwater Society. [Google Scholar]
- Devito KJ, Hill AR, and Roulet N. 1996. “Groundwater-Surface Water Interactions in Headwater Forested Wetlands of the Canadian Shield.” Journal of Hydrology 181: 127–47. 10.1016/0022-1694(95)02912-5. [DOI] [Google Scholar]
- Dierberg FE, and Brezonik PL. 1984. “The Effect of Wastewater on the Surface Water and Groundwater Quality of Cypress Domes.” In Cypress Swamps, edited by Ewel KC and Odum HT, 83–101. Gainesville, FL: University of Florida Press. [Google Scholar]
- Dillon PJ, and Molot LA. 1997. “Effects of Landscape Form on Export of Dissolved Organic Carbon, Iron, and Phosphorus from Forested Stream Catchments.” Water Resources Research 33: 2591–600. 10.1029/97WR01921. [DOI] [Google Scholar]
- Dodla SK, Wang JJ, DeLaune RD, and Cook RL. 2008. “Denitrification Potential and Its Relation to Organic Carbon Quality in Three Coastal Wetland Soils.” Science of The Total Environment 407 (1): 471–80. 10.1016/j.scitotenv.2008.08.022. [DOI] [PubMed] [Google Scholar]
- Driscoll CT, Blette V, Yan C, Schofield CL, Munson R, and Holsapple J. 1995. “The Role of Dissolved Organic Carbon in the Chemistry and Bioavailability of Mercury in Remote Adirondack Lakes.” Water, Air, & Soil Pollution 80: 499–508. 10.1007/BF01189700. [DOI] [Google Scholar]
- Duever MJ, Meeder JF, Meeder LC, and McCollom JM. 1994. “The Climate of South Florida and Its Role in Shaping the Everglades Ecosystem.” In Everglades: The Ecosystem and Its Restoration, edited by Davis SM and Ogden JC, 225–48. Delray Beach, FL: St. Lucie Press. [Google Scholar]
- Dunne EJ, Reddy KR, and Clark MW. 2006. “Phosphorus Release and Retention by Soils of Natural Isolated Wetlands.” International Journal of Environment and Pollution 28 (3): 496–516. 10.1504/IJEP.2006.011225. [DOI] [Google Scholar]
- Dunne EJ, Smith J, Perkins DB, Clark MW, Jawitz JW, and Reddy KR. 2007. “Phosphorus Storages in Historically Isolated Wetland Ecosystems and Surrounding Pasture Uplands.” Ecological Engineering 31 (1): 16–28. 10.1016/j.ecoleng.2007.05.004. [DOI] [Google Scholar]
- Eckhardt BW, and Moore TR. 1990. “Controls on Dissolved Organic Carbon Concentrations in Streams of Southern Quebec.” Canadian Journal of Fisheries and Aquatic Sciences 47: 1537–44. [Google Scholar]
- Euliss N, LaBaugh J, Frederickson L, Mushet D, Laubhan M, Swanson G, Winter T, Rosenberry D, and Nelson R. 2004. “The Wetland Continuum: A Conceptual Framework for Interpreting Biological Studies.” Wetlands 24 (2): 448–58. 10.1672/0277-5212(2004)024[0448:twcacf]2.0.co;2. [DOI] [Google Scholar]
- Evenson GR, Golden HE, Lane CR, and D’Amico E. 2015. “Geographically Isolated Wetlands and Watershed Hydrology: A Modified Model Analysis.” Journal of Hydrology 529: 240–56. 10.1016/j.jhydrol.2015.07.039. [DOI] [Google Scholar]
- Evenson GR, Golden HE, Lane CR, and D’Amico E. 2016. “An Improved Representation of Geographically Isolated Wetlands in a Watershed-Scale Hydrologic Model.” Hydrological Processes 30: 4168–84. 10.1002/hyp.10930. [DOI] [Google Scholar]
- Evenson GR, Golden HE, Lane CR, McLaughlin DL, and D’Amico E. in press. “Depressional Wetlands Affect Watershed Hydrological, Biogeochemical, and Ecological Functions.” Ecological Applications. 10.1002/eap.1701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ewel KC, and Odum HT. 1984. “Ecological Patterns in Cypress Swamps.” In Cypress Swamps, edited by Ewel KC and Odum HT, 5–6. Gainesville, FL: University of Florida Press. [Google Scholar]
- Fergus CE, Lapierre JF, Oliver SK, Skaff NK, Cheruvelil KS, Webster K, Scott C, and Soranno P. 2017. “The Freshwater Landscape: Lake, Wetland, and Stream Abundance and Connectivity at Macroscales.” Ecosphere 8 (8): e01911. 10.1002/ecs2.1911. [DOI] [Google Scholar]
- Ferone JM, and Devito KJ. 2004. “Shallow Groundwater-Surface Water Interactions in Pond-Peatland Complexes Along a Boreal Plains Topographic Gradient.” Journal of Hydrology 292: 75–95. 10.1016/j.jhydrol.2003.12.032. [DOI] [Google Scholar]
- Flaig EG, and Reddy KR. 1995. “Fate of Phosphorus in the Lake Okeechobee Watershed, Florida, USA: Overview and Recommendations.” Ecological Engineering 5 (2–3): 127–42. 10.1016/0925-8574(95)00021-6. [DOI] [Google Scholar]
- Ford J, and Bedford BL. 1987. “The Hydrology of Alaskan Wetlands, USA: A Review.” Arctic and Alpine Research 19 (3): 209–29. [Google Scholar]
- Forman RTT 1995. “Some General Principles of Landscape and Regional Ecology.” Landscape Ecology 10 (3): 133–42. [Google Scholar]
- Fossey MR, and Rousseau AN. 2016. “Can Isolated and Riparian Wetlands Mitigate the Impact of Climate Change on Watershed Hydrology? A Case Study Approach.” Journal of Environmental Management 184 (Part 2): 327–39. 10.1016/j.jenvman.2016.09.043. [DOI] [PubMed] [Google Scholar]
- Fritz KM, Schofield KA, Alexander LC, McManus MG, Golden HE, Lane CR, Kepner WG, LeDuc SD, DeMeester JE, and Pollard AI. 2018. “Physical and Chemical Connectivity of Streams and Riparian Wetlands to Downstream Waters: A Synthesis.” Journal of the American Water Resources Association 54 (2): 323–345. 10.1111/1752-1688.12632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galloway JN, and Cowling EB. 2002. “Reactive Nitrogen and the World: 200 Years of Change.” AMBIO: A Journal of the Human Environment 31 (2): 64–71. 10.1579/0044-7447-31.2.64. [DOI] [PubMed] [Google Scholar]
- Galloway ME, and Branfireun BA. 2004. “Hydrological and Biogeochemical Controls on Mercury Fate and Transport in a Southern Ontario Forested Wetland.” The Science of Total Environment 325: 239–54. 10.1016/j.scitotenv.2003.11.010. [DOI] [PubMed] [Google Scholar]
- Gergel SE, Turner MG, and Kratz TK. 1999. “Dissolved Organic Carbon as an Indicator of the Scale of Watershed Influence on Lakes and Rivers.” Ecological Applications 9: 1377–90. 10.1890/1051-0761(1999)009[1377:DOCAAI]2.0.CO;2. [DOI] [Google Scholar]
- Ghermandi A, van den Bergh JCJM, Brander LM, de Groot HLF, and Nunes PALD. 2010. “Values of Natural and Human-Made Wetlands: A Meta-Analysis.” Water Resources Research 46 (12): W12516. 10.1029/2010wr009071. [DOI] [Google Scholar]
- Gleason RA, Tangen BA, Laubhan MK, Kermes KE, and Euliss NH. 2007. “Estimating Water Storage Capacity of Existing and Potentially Restorable Wetland Depressions in a Subbasin of the Red River of the North.” U.S. Geological Survey Open File Report 2007–1159, United States Geological Survey, Reston, VA. [Google Scholar]
- Golden HE, Creed IF, Ali G, Basu NB, Neff B, Rains MC, McLaughlin DL, Alexander LC, Ameli AA, Christensen JR, Evenson GR, Jones CN, Lane CR, and Lang MW. 2017. “Integrating Geographically Isolated Wetlands into Land Management Decisions.” Frontiers in Ecology and the Environment 15 (6): 319–27. 10.1002/fee.1504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golden HE, Lane CR, Amatya DM, Bandilla KW, Raanan-Kiperwas H, Knightes CD, and Ssegane H. 2014. “Hydrologic Connectivity between Geographically Isolated Wetlands and Surface Water Systems: A Review of Select Modeling Methods.” Environmental Modelling & Software 53: 190–206. 10.1016/j.envsoft.2013.12.004. [DOI] [Google Scholar]
- Golden HE, Sander HA, Lane CR, Zhao C, Price K, D’Amico E, and Christensen JR. 2015. “Relative Effects of Geographically Isolated Wetlands on Streamflow: A Watershed-Scale Analysis.” Ecohydrology 9 (1): 21–38. 10.1002/eco.1608. [DOI] [Google Scholar]
- Goodrich DC, Kepner WG, Levick LR, and Wigington PJ. 2018. “Southwestern Intermittent and Ephemeral Stream Connectivity. Journal of the American Water Resources Association 54 (2): 400–422. 10.1111/1752-1688.12636. [DOI] [Google Scholar]
- Gorham E, Underwood JK, Martini FB, and Ogden JG. 1986. “Natural and Anthropogenic Causes of Lake Acidification in Nova Scotia.” Nature 324 (6096): 451–53. 10.1038/324451a0. [DOI] [Google Scholar]
- Grigal DF 2002. “Inputs and Outputs of Mercury from Terrestrial Watersheds: A Review.” Environmental Reviews 10 (1): 1–39. 10.1139/a01-013. [DOI] [Google Scholar]
- Haag KH, Lee TM, and Herndon DC. 2005. “Bathymetry and Vegetation in Isolated Marsh and Cypress Wetlands in the Northern Tampa Bay Area, 2000–2004.” U.S. Geological Survey Scientific Investigations Report 2005–5109, 49 pp. [Google Scholar]
- Haan CT, and Johnson HP. 1968. “Hydraulic Model of Runoff from Depressional Areas. Part I: General Considerations.” Transactions of the American Society of Agricultural Engineers 11: 364–67. 10.13031/2013.39411. [DOI] [Google Scholar]
- Hayashi M, Quinton WL, Pietroniro A, and Gibson JJ. 2004. “Hydrologic Functions of Wetlands in a Discontinuous Permafrost Basin Indicated by Isotopic and Chemical Signatures.” Journal of Hydrology 296 (1–4): 81–97. 10.1016/j.jhydrol.2004.03.020. [DOI] [Google Scholar]
- Hayashi M, van der Kamp G, and Rosenberry DO. 2016. “Hydrology of Prairie Wetlands: Understanding the Integrated Surface-Water and Groundwater Processes.” Wetlands 36 (2): 237–54. 10.1007/s13157-016-0797-9. [DOI] [Google Scholar]
- Hayashi M, van der Kamp G, and Rudolph DL. 1998a. “Water and Solute Transfer Between a Prairie Wetland and Adjacent Uplands, 1. Water Balance.” Journal of Hydrology 207 (1–2): 42–55. 10.1016/s0022-1694(98)00098-5. [DOI] [Google Scholar]
- Hayashi M, van der Kamp G, and Rudolph DL. 1998b. “Water and Solute Transfer Between a Prairie Wetland and Adjacent Uplands, 2. Chloride Cycle.” Journal of Hydrology 207 (1–2): 56–67. 10.1016/S0022-1694(98)00099-7. [DOI] [Google Scholar]
- Heagle D, Hayashi M, and van der Kamp G. 2013. “Surface–Subsurface Salinity Distribution and Exchange in a Closed-Basin Prairie Wetland.” Journal of Hydrology 478: 1–14. 10.1016/j.jhydrol.2012.05.054. [DOI] [Google Scholar]
- Helfield JM, and Naiman RJ. 2006. “Keystone Interactions: Salmon and Bear in Riparian Forests of Alaska.” Ecosystems 9 (2): 167–80. 10.1007/s10021-004-0063-5. [DOI] [Google Scholar]
- Hemond HF 1980. “Biogeochemistry of Thoreau’s Bog, Concord, Massachusetts.” Ecological Monographs 50: 507–26. [Google Scholar]
- Hemond HF 1983. “The Nitrogen Budget of Thoreau’s Bog.” Ecology 64: 99–109. [Google Scholar]
- Heyes A, Moore TR, Rudd JWM, and Dugoua JJ. 2000. “Methyl Mercury in Pristine and Impounded Boreal Peatlands, Experimental Lakes Area, Ontario, Canada.” Canadian Journal of Fisheries and Aquatic Sciences 57: 2211–22. 10.1139/f00-197. [DOI] [Google Scholar]
- Hobbie JE 1992. “Microbial Control of Dissolved Organic Carbon in Lakes: Research for the Future.” Hydrobiologia 229: 169–80. [Google Scholar]
- Hoffmann CC, Kjaergaard C, Uusi-Kämppä J, Hansen HCB, and Kronvang B. 2009. “Phosphorus Retention in Riparian Buffers: Review of Their Efficiency.” Journal of Environmental Quality 38: 1942–55. 10.2134/jeq2008.0087. [DOI] [PubMed] [Google Scholar]
- Holgerson MA, and Raymond PA. 2016. “Large Contribution to Inland Water CO2 and CH4 Emissions from Very Small Ponds.” Nature Geosciences 9 (3): 222–26. 10.1038/ngeo2654. [DOI] [Google Scholar]
- Hoos AB, Moore RB, Garcia AM, Noe GB, Terziotti SE, Johnston CM, and Dennis RL. 2013. “Simulating Stream Transport of Nutrients in the Eastern United States, 2002, Using a Spatially-Referenced Regression Model and 1:100,000-Scale Hydrography.” U.S. Geological Survey Scientific Investigations Report 2013–5102, 33 pp. http://pubs.usgs.gov/sir/2013/5102/. [Google Scholar]
- Hope D, Billet MF, and Cresser MS. 1994. “A Review of the Export of Carbon in River Water: Fluxes and Processes.” Environmental Pollution 84: 301–24. 10.1016/0269-7491(94)90142-2. [DOI] [PubMed] [Google Scholar]
- Huang C, Peng Y, Lang MW, Yeo I, and McCarty G. 2014. “Wetland Inundation Mapping and Change Monitoring Using Landsat and Airborne LiDAR Data.” Remote Sensing of Environment 141: 231–42. 10.1016/j.rse.2013.10.020. [DOI] [Google Scholar]
- Huang S, Young C, Feng M, Heidemann K, Cushing M, Mushet DM, and Liu S. 2011. “Demonstration of a Conceptual Model for Using LiDAR to Improve the Estimation of Floodwater Mitigation Potential of Prairie Pothole Region Wetlands.” Journal of Hydrology 405: 417–26. 10.1016/j.jhydrol.2011.05.040. [DOI] [Google Scholar]
- Hubbard DE, and Linder RL. 1986. “Spring Runoff Retention in Prairie Pothole Wetlands.” Journal of Soil and Water Conservation 41: 122–25. [Google Scholar]
- Hynes HBN 1975. “The Stream and Its Valley.” Verhandlungen des Internationalen Verein Limnologie 19: 1–15. [Google Scholar]
- Jacques J, and Lorenz DL. 1988. “Techniques for Estimating the Magnitude and Frequency of Floods of Ungauged Streams in Minnesota.” USGS Water-Resources Investigations Report 84–4170, U.S. Department of the Interior, U.S. Geological Survey, Washington, D.C. [Google Scholar]
- Jin H, Huang C, Lang MW, Yeo I, and Stehman SV. 2017. “Monitoring of Wetland Inundation Dynamics in the Delmarva Peninsula Using Landsat Time-Series Imagery from 1985 to 2011.” Remote Sensing of Environment 190: 26–41. 10.1016/j.rse.2016.12.001. [DOI] [Google Scholar]
- Johnson AI 1967. “Specific Yield — Compilation of Specific Yields for Various Materials.” USGS Water-Supply Paper 1662-D, U.S. Department of the Interior, U.S. Geological Survey, Washington, D.C. [Google Scholar]
- Johnson CA 1991. “Sediment and Nutrient Retention by Freshwater Wetlands: Effects on Surface Water Quality.” Critical Reviews in Environmental Control 21 (5, 6): 491–565. [Google Scholar]
- Johnston C, Detenbeck N, and Niemi G. 1990. “The Cumulative Effect of Wetlands on Stream Water Quality and Quantity. A Landscape Approach.” Biogeochemistry 10: 105–41. [Google Scholar]
- Jones CN, Evenson GR, McLaughlin DM, Vanderhoof MK, Lang MW, McCarty GW, Golden HE, Lane CR, and Alexander LC. 2018. “Estimating Restorable Wetland Water Storage at Landscape Scales.” Hydrological Processes 32 (2): 305–13. 10.1002/hyp.11405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones J 2015. “Efficient Wetland Surface Water Detection and Monitoring Via Landsat: Comparison with In Situ Data from the Everglades Depth Estimation Network.” Remote Sensing 7 (9): 12503–38. 10.3390/rs70912503. [DOI] [Google Scholar]
- Jordan S, Stoffer J, and Nestlerode J. 2011. “Wetlands as Sinks for Reactive Nitrogen at Continental and Global Scales: A Meta-Analysis.” Ecosystems 14 (1): 144–55. 10.1007/s10021-010-9400-z. [DOI] [Google Scholar]
- Jordan TE, Andrews MP, Szuch RP, Whigham DF, Weller DE, and Jacobs AD. 2007. “Comparing Functional Assessments of Wetlands to Measurements of Soil Characteristics and Nitrogen Processing.” Wetlands 27 (3): 479–97. 10.1672/0277-5212(2007)27[479:CFAOWT]2.0.CO;2. [DOI] [Google Scholar]
- Kahara SN, Mockler RM, Higgins KF, Chipps SR, and Johnson RR. 2009. “Spatiotemporal Patterns of Wetland Occurrence in the Prairie Pothole Region of Eastern South Dakota.” Wetlands 29 (2): 678–89. 10.1672/07-09.1. [DOI] [Google Scholar]
- Kao CM, Wang JY, Chen KF, Lee HY, and Wu MJ. 2002. “Non-Point Source Pesticide Removal by a Mountainous Wetland.” Water Science and Technology 46 (6–7): 199–206. [PubMed] [Google Scholar]
- Kehew AE, Passero RN, Krishnamurthy RV, Lovett CK, Betts MA, and Dayharsh BA. 1998. “Hydrogeochemical Interaction between a Wetland and an Unconfined Glacial Drift Aquifer, Southwestern Michigan.” Ground Water 36: 849–56. 10.1111/j.1745-6584.1998.tb02204.x. [DOI] [Google Scholar]
- Klaus J, McDonnell JJ, Jackson CR, Du E, and Griffiths NA. 2015. “Where Does Streamwater Come from in Low-Relief Forested Watersheds? A Dual-Isotope Approach.” Hydrology and Earth System Sciences 19 (1): 125–35. 10.5194/hess-19-125-2015. [DOI] [Google Scholar]
- Koprivnjak J-F, and Moore TR. 1992. “Sources, Sinks, and Fluxes of Dissolved Organic Carbon in Subarctic Fen Catchments.” Arctic and Alpine Research 24: 204–10. [Google Scholar]
- Kortelainen P 1993. “Content of Total Organic Carbon in Finnish Lakes and Its Relationship to Catchment Characteristics.” Canadian Journal of Fisheries and Aquatic Sciences 50: 1477–83. [Google Scholar]
- Lane CR, and Autrey BC. 2016. “Phosphorus Retention of Forested and Emergent Marsh Depressional Wetlands in Differing Land Uses in Florida, USA.” Wetlands Ecology and Management 24 (1): 45–60. 10.1007/s11273-015-9450-2. [DOI] [Google Scholar]
- Lane CR, and Autrey BC. 2017. “Sediment Accretion and Accumulation of P, N, and Organic C in Depressional Wetlands of Three Ecoregions of the United States.” Marine and Freshwater Research 68: 2253–65. 10.1071/MF16372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lane CR, Autrey BC, Jicha T, Lehto L, Elonen C, and Seifert-Monson L. 2015. “Denitrification Potential in Geographically Isolated Wetlands of North Carolina and Florida, USA.” Wetlands 35 (3): 459–71. 10.1007/s13157-015-0633-7. [DOI] [Google Scholar]
- Lane CR, and D’Amico E. 2010. “Calculating the Ecosystem Service of Water Storage in Isolated Wetlands Using LiDAR in North Central Florida, USA.” Wetlands 30 (5): 967–77. 10.1007/s13157-010-0085-z. [DOI] [Google Scholar]
- Lane CR, and D’Amico E. 2016. “Identification of Putative Geographically Isolated Wetlands of the Conterminous United States.” Journal of the American Water Resources Association 52 (3): 705–22. 10.1111/1752-1688.12421. [DOI] [Google Scholar]
- Lane CR, D’Amico E, and Autrey BC. 2012. “Isolated Wetlands of the Southeastern United States: Abundance and Expected Condition.” Wetlands 32 (4): 753–67. 10.1007/s13157-012-0308-6. [DOI] [Google Scholar]
- Lane CR, Hall A, D’Amico E, Sangwan N, and Merwade V. 2017. “Characterizing the Extent of Spatially Integrated Floodplain and Wetland Systems in the White River, Indiana, USA.” Journal of the American Water Resources Association 53: 774–90. 10.1111/1752-1688.12531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang MW, McDonough O, McCarty G, Oesterling R, and Wilen B. 2012. “Enhanced Detection of Wetland-Stream Connectivity Using LiDAR.” Wetlands 32: 461–79. 10.1007/s13157-012-0279-7. [DOI] [Google Scholar]
- Le PVV, and Kumar P. 2014. “Power-Law Scaling of Topographic Depressions and Their Hydrologic Connectivity.” Geophysical Research Letters 41: 1553–59. 10.1002/2013gl059114. [DOI] [Google Scholar]
- Lehman PW, Sommer T, and Rivard L. 2008. “The Influence of Floodplain Habitat on the Quantity and Quality of Riverine Phytoplankton Carbon Produced during the Flood Season in San Francisco Estuary.” Aquatic Ecology 42: 363–78. 10.1007/s10452-007-9102-6. [DOI] [Google Scholar]
- Leibowitz SG, Mushet DM, and Newton WE. 2016. “Intermittent Surface Water Connectivity: Fill and Spill Vs. Fill and Merge Dynamics.” Wetlands 36 (2): 323–42. 10.1007/s13157-016-0830-z. [DOI] [Google Scholar]
- Leibowitz SG, and Vining KC. 2003. “Temporal Connectivity in a Prairie Pothole Complex.” Wetlands 23 (1): 13–25. 10.1672/0277-5212(2003)023[0013:tciapp]2.0.co;2. [DOI] [Google Scholar]
- Leibowitz SG, Wigington PJ, Rains MC, and Downing DM. 2008. “Non-Navigable Streams and Adjacent Wetlands: Addressing Science Needs Following the Supreme Court’s Rapanos Decision.” Frontiers in Ecology and the Environment 6 (7): 366–73. 10.1890/070068. [DOI] [Google Scholar]
- Leibowitz SG, Wigington PJ Jr., Schofield KA, Alexander LC, Vanderhoof MK, and Golden HE. 2018. “Connectivity of Streams and Wetlands to Downstream Waters: An Integrated Systems Framework.” Journal of the American Water Resources Association 54 (2). 10.1111/1752-1688.12631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leigh C, Sheldon F, Kingsford RT, and Arthington AH. 2010. “Sequential Floods Drive ‘Booms’ and Wetland Persistence in Dryland Rivers: A Synthesis.” Marine and Freshwater Research 61: 896–908. 10.1071/MF10106. [DOI] [Google Scholar]
- Leopold LB 1994. A View of the River. Cambridge, MA: Harvard University Press. [Google Scholar]
- Lide RF 1997. “When is a Depression Wetland a Carolina Bay?” Southeastern Geographer 36: 90–98. 10.1353/sgo.1997.0005. [DOI] [Google Scholar]
- Lide RF, Meentemeyer VG, Pinder JE, and Beatty LM. 1995. “Hydrology of a Carolina Bay Located on the Upper Coastal-Plain of Western South-Carolina.” Wetlands 15 (1): 47–57. [Google Scholar]
- Lindsay JB, Creed IF, and Beall FD. 2004. “Drainage Basin Morphometrics for Depressional Landscapes.” Water Resources Research 40 (9): W09307. 10.1029/2004WR003322. [DOI] [Google Scholar]
- Linqvist O, Johansson K, Aastrup M, Andersson A, Bringmark L, Hovsenius G, Hakanson L, Iverfeldt A, Meili M, and Timm B. 1991. “Mercury in the Swedish Environment — Recent Research on Causes, Consequences, and Remedial Measures.” Water, Air, & Soil Pollution 55 (1–2): 1–261. 10.1007/BF00542429. [DOI] [Google Scholar]
- Lorenz DL, Sanocki CA, and Kocian MJ. 2010. “Techniques for Estimating the Magnitude and Frequency of Peak Flows on Small Streams in Minnesota Based on Data through Water Year 2005.” USGS Scientific Investigations Report 2009–5250, U.S. Department of the Interior, U.S. Geological Survey, in cooperation with the Minnesota Department of Transportation and the Minnesota Pollution Control Agency, Reston, VA. [Google Scholar]
- Madison RJ, and Brunett JO. 1985. “Overview of the Occurrence of Nitrate in Ground Water of the United States. National Water Summary 1984-Hydrologic Events, Selected Water-Quality Trends, and Ground-Water Resources.” U.S. Geological Survey Water-Supply Paper 2275, pp. 93–105. [Google Scholar]
- Marton JM, Chowdhury RR, and Craft CB. 2015. “A Comparison of the Spatial Variability of Denitrification and Related Soil Properties in Restored and Natural Depression Wetlands in Indiana, USA.” International Journal of Biodiversity Science, Ecosystem Services & Management 11 (1): 36–45. 10.1080/21513732.2014.950981. [DOI] [Google Scholar]
- Marton JM, Creed IF, Lewis D, Lane CR, Basu N, Cohen MJ, and Craft C. 2015. “Geographically Isolated Wetlands are Important Biogeochemical Reactors on the Landscape.” BioScience 65 (4): 408–18. 10.1093/biosci/biv009. [DOI] [Google Scholar]
- Marton JM, Fennessy MS, and Craft CB. 2014. “USDA Conservation Practices Increase Carbon Storage and Water Quality Improvement Functions: An Example from Ohio.” Restoration Ecology 22 (1): 117–24. 10.1111/rec.12033. [DOI] [Google Scholar]
- Matheney RK, and Gerla PJ. 1996. “Environmental Isotopic Evidence for the Origins of Ground and Surface Water in a Prairie Discharge Wetland.” Wetlands 16: 109–20. [Google Scholar]
- McCauley LA, Anteau MJ, van der Burg MP, and Wiltermuth MT. 2015. “Land Use and Wetland Drainage Affect Water Levels and Dynamics of Remaining Wetlands.” Ecosphere 6 (6): art92. 10.1890/es14-00494.1. [DOI] [Google Scholar]
- McDonough OT, Lang MW, Hosen JD, and Palmer MA. 2015. “Surface Hydrologic Connectivity between Delmarva Bay Wetlands and Nearby Streams Along a Gradient of Agricultural Alteration.” Wetlands 35: 41–53. 10.1007/s13157-014-0591-5. [DOI] [Google Scholar]
- McEachern P, Prepas EE, and Chanasyk DS. 2006. “Landscape Control of Water Chemistry in Northern Boreal Streams of Alberta.” Journal of Hydrology 323: 303–24. 10.1016/j.jhydrol.2005.09.016. [DOI] [Google Scholar]
- McLaughlin DL, Kaplan DA, and Cohen MJ. 2014. “A Significant Nexus: Geographically Isolated Wetlands Influence Landscape Hydrology.” Water Resources Research 50: 7153–66. 10.1002/2013WR015002. [DOI] [Google Scholar]
- Mierle G, and Ingram R. 1991. “The Role of Humic Substances in the Mobilization of Mercury from Watersheds.” Water, Air, & Soil Pollution 56: 349–57. [Google Scholar]
- Min J-H, Perkins DB, and Jawitz JW. 2010. “Wetland-Groundwater Interactions in Subtropical Depressional Wetlands.” Wetlands 30 (5): 997–1006. 10.1007/s13157-010-0043-9. [DOI] [Google Scholar]
- Mitsch WJ, Gosselink JG, Anderson C, and Zhang L. 2009. Wetland Ecosystems (First Edition). Hoboken, NJ: John Wiley & Sons. [Google Scholar]
- Moraghan JT 1993. “Loss and Assimilation of 15N-Nitrate Added to a North Dakota Cattail Marsh.” Aquatic Botany 46: 225–34. [Google Scholar]
- Morley TR, Reeve AS, and Calhoun AJK. 2011. “The Role of Headwater Wetlands in Altering Streamflow and Chemistry in a Maine, USA Catchment.” Journal of the American Water Resources Association 47: 337–49. 10.1111/j.1752-1688.2010.00519.x. [DOI] [Google Scholar]
- Mulholland PJ, and Kuenzler EJ. 1979. “Organic Carbon Export from Upland and Forested Wetland Watersheds.” Limnology and Oceanography 24: 960–66. [Google Scholar]
- Mushet DM, Euliss NH, Chen Y, and Stockwell CA. 2013. “Complex Spatial Dynamics Maintain Northern Leopard Frog (Lithobates pipiens) Genetic Diversity in a Temporally Varying Landscape.” Herpetological Conservation and Biology 8 (1): 163–75. [Google Scholar]
- Nachshon U, Ireson A, van der Kamp G, Davies SR, and Wheater HS. 2014. “Impacts of Climate Variability on Wetland Salinization in the North American Prairies.” Hydrology and Earth System Sciences 18 (4): 1251–63. 10.5194/hess-18-1251-2014. [DOI] [Google Scholar]
- Nadeau T-L, and Rains MC. 2007. “Hydrological Connectivity between Headwater Streams and Downstream Waters: How Science Can Inform Policy.” Journal of the American Water Resources Association 43 (1): 118–33. 10.1111/j.1752-1688.2007.00010.x. [DOI] [Google Scholar]
- Nair VD, Clark MW, and Reddy KR. 2015. “Evaluation of Legacy Phosphorus Storage and Release from Wetland Soils.” Journal of Environmental Quality 44: 1956–64. 10.2134/jeq2015.03.0154. [DOI] [PubMed] [Google Scholar]
- Neff B, and Rosenberry DO. 2017. “Groundwater Connectivity in Upland-Embedded Wetlands of the Prairie Pothole Region.” Wetlands: 1–13. 10.1007/s13157-017-0956-7. [DOI] [Google Scholar]
- Novitzki RP 1979. “Hydrologic Characteristics of Wisconsin’s Wetlands and Their Influence on Floods.” In Wetland Functions and Values: The Status of Our Understanding. Proceedings of the National Symposium on Wetlands, edited by Greeson P, Clark J, and Clark JE, 377–88. Minneapolis, MN: American Water Resources Association. [Google Scholar]
- Osterkamp WR 2008. “Annotated Definitions of Selected Geomorphic Terms and Related Terms of Hydrology, Sedimentology, Soil Science and Ecology.” USGS Open File Report 2008–1217, U.S. Department of the Interior, U.S. Geological Survey, Reston, VA. [Google Scholar]
- Phillips PJ, and Shedlock RJ. 1993. “Hydrology and Chemistry of Groundwater and Seasonal Ponds in the Atlantic Coastal Plain in Delaware, USA.” Journal of Hydrology 141 (1–4): 157–78. [Google Scholar]
- Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, and Stromburg JC. 1997. “The Natural Flow Regime.” BioScience 47 (11): 769–84. 10.2307/1313099. [DOI] [Google Scholar]
- Poff NL, Olden JD, Merritt DM, and Pepin DM. 2007. “Homogenization of Regional River Dynamics by Dams and Global Biodiversity Implications.” Proceedings of the National Academy of Sciences of the United States of America 104 (14): 5732–37. 10.1073/pnas.0609812104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porvari P, and Verta M. 2003. “Total and Methyl Mercury Concentrations and Fluxes from Small Boreal Forest Catchments in Finland.” Environmental Pollution 123 (2): 181–91. 10.1016/s0269-7491(02)00404-9. [DOI] [PubMed] [Google Scholar]
- Pyzoha JE, Callahan TJ, Sun GE, Trettin CC, and Miwa M. 2008. “A Conceptual Hydrologic Model for a Forested Carolina Bay Depressional Wetland on the Coastal Plain of South Carolina, USA.” Hydrological Processes 22 (14): 2689–98. 10.1002/hyp.6866. [DOI] [Google Scholar]
- Quinton WL, Hayashi M, and Pietroniro A. 2003. “Connectivity and Storage Functions of Channel Fens and Flat Bogs in Northern Basins.” Hydrological Processes 17: 3665–84. 10.1002/hyp.1369. [DOI] [Google Scholar]
- Racchetti E, Bartoli M, Soana E, Longhi D, Christian RR, Pinardi M, and Viaroli P. 2011. “Influence of Hydrological Connectivity of Riverine Wetlands on Nitrogen Removal Via Denitrification.” Biogeochemistry 103 (1–3): 335–54. 10.1007/s10533-010-9477-7. [DOI] [Google Scholar]
- Rains MC 2011. “Water Sources and Hydrodynamics of Closed-Basin Depressions, Cook Inlet Region, Alaska.” Wetlands 31: 377–87. 10.1007/s13157-011-0147-x. [DOI] [Google Scholar]
- Rains MC, Graham EF, Harter T, Dahlgren RA, and Williamson RJ. 2006. “The Role of Perched Aquifers in Hydrological Connectivity and Biogeochemical Processes in Vernal Pool Landscapes, Central Valley, California.” Hydrological Processes 20 (5): 1157–75. 10.1002/hyp.5937. [DOI] [Google Scholar]
- Rains MC, Leibowitz SG, Cohen MJ, Creed IF, Golden HE, Jawitz JW, Kalla P, Lane CR, Lang MW, and McLaughlin DL. 2015. “Geographically Isolated Wetlands Are Part of the Hydrological Landscape.” Hydrological Processes 30 (1): 153–60. 10.1002/hyp.10610. [DOI] [Google Scholar]
- Reddy KR, and DeLaune RD. 2008. Biogeochemistry of Wetlands: Science and Applications. Boca Raton, FL: CRC Press. [Google Scholar]
- Reddy KR, Kadlec RH, Flaig E, and Gale PM. 1999. “Phosphorus Retention in Streams and Wetlands: A Review.” Critical Reviews in Environmental Science and Technology 29 (1): 83–146. 10.1080/10643389991259182. [DOI] [Google Scholar]
- Richardson JL, Wilding LP, and Daniels RB. 1992. “Recharge and Discharge of Groundwater in Aquic Conditions Illustrated with FLOWNET Analysis.” Geoderma 53: 65–78. [Google Scholar]
- Rosenberry DO, and Winter TC. 1997. “Dynamics of Water-Table Fluctuations in an Upland between Two Prairie-Pothole Wetlands in North Dakota.” Journal of Hydrology 191: 266–89. 10.1016/S0022-1694(96)03050-8. [DOI] [Google Scholar]
- Roulet NT, and Woo MK. 1986. “Hydrology of a Wetland in the Continuous Permafrost Region.” Journal of Hydrology 89 (1–2): 73–91. [Google Scholar]
- Rovansek RJ, Hinzman LD, and Kane DL. 1996. “Hydrology of a Tundra Wetland Complex on the Alaskan Arctic Coastal Plain, USA.” Arctic and Alpine Research 28 (3): 311–17. [Google Scholar]
- Schemel LE, Sommer TR, Muller-Solger AB, and Harrell WC. 2004. “Hydrologic Variability, Water Chemistry, and Phytoplankton Biomass in a Large Floodplain of the Sacramento River, CA, USA.” Hydrobiologia 513: 129–39. 10.1023/B:hydr.0000018178.85404.1c. [DOI] [Google Scholar]
- Schiller KE, Jindal P, Basu NB, and Helmers MJ. 2012. “Impact of Artificial Subsurface Drainage on Groundwater Travel Times and Baseflow Discharge in an Agricultural Watershed, Iowa (USA).” Hydrological Processes 26: 3092–100. 10.1002/hyp.8337. [DOI] [Google Scholar]
- Schofield KA, Alexander LC, Ridley CE, Vanderhoof MK, Fritz KM, Autrey BC, DeMeester JE, Kepner WG, Lane CR, Leibowitz SG, and Pollard AI. 2018. “Biota Connect Aquatic Habitats Throughout Freshwater Ecosystem Mosaics.” Journal of the American Water Resources Association 54 (2): 372–399. 10.1111/1752-1688.12634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selvendiran P, Driscoll CT, Bushey JT, and Montesdeoca MR. 2008. “Wetland Influence on Mercury Fate and Transport in a Temperate Forested Watershed.” Environmental Pollution 154: 46–55. 10.1016/j.envpol.2007.12.005. [DOI] [PubMed] [Google Scholar]
- Serran JN, and Creed IF. 2016. “New Mapping Techniques to Estimate the Preferential Loss of Small Wetlands on Prairie Landscapes.” Hydrological Processes 30 (3): 396–409. 10.1002/hyp.10582. [DOI] [Google Scholar]
- Serran JN, Creed IF, Ameli AA, and Aldred DA. 2017. “Estimating Rates of Wetland Loss Using Power-Law Functions.” Wetlands. 10.1007/s13157-017-0960-y. [DOI] [Google Scholar]
- Shaw DA, van der Kamp G, Conly FM, Pietroniro A, and Martz L. 2012. “The Fill-Spill Hydrology of Prairie Wetland Complexes during Drought and Deluge.” Hydrological Processes 26 (20): 3147–56. 10.1002/hyp.8390. [DOI] [Google Scholar]
- Shedlock RJ, Wilcox DA, Thompson TA, and Cohen DA. 1993. “Interactions between Ground Water and Wetlands, Southern Shore of Lake Michigan, USA.” Journal of Hydrology 141: 127–55. [Google Scholar]
- Shook K, Pomeroy J, and van der Kamp G. 2015. “The Transformation of Frequency Distributions of Winter Precipitation to Spring Streamflow Probabilities in Cold Regions; Case Studies from the Canadian Prairies.” Journal of Hydrology 521: 395–409. 10.1016/j.jhydrol.2014.12.014. [DOI] [Google Scholar]
- Smith RD, Ammann A, Bartoldus C, and Brinson MM. 1995. “An Approach for Assessing Wetland Functions Using Hydrogeomorphic Classification, Reference Wetlands, and Functional Indices.” U.S. Army Corps of Engineers. [Google Scholar]
- Spence C, and Woo MK. 2003. “Hydrology of Subarctic Canadian Shield: Soil-Filled Valleys.” Journal of Hydrology 279: 151–66. 10.1016/S0022-1694(03)00175-6. [DOI] [Google Scholar]
- St. Louis VL, Rudd JWM, Kelly CA, Beaty KG, Bloom NS, and Flett RJ. 1994. “Importance of Wetlands as Sources of Methyl Mercury to Boreal Forest Ecosystems.” Canadian Journal of Fisheries and Aquatic Sciences 51 (5): 1065–76. [Google Scholar]
- Subalusky AL, Smith LL, and Fitzgerald LA. 2009a. “Ontogenetic Niche Shifts in the American Alligator Establish Functional Connectivity between Aquatic Systems.” Biological Conservation 142 (7): 1507–14. 10.1016/j.biocon.2009.02.019. [DOI] [Google Scholar]
- Subalusky AL, Smith LL, and Fitzgerald LA. 2009b. “Detection of American Alligators in Isolated, Seasonal Wetlands.” Applied Herpetology 6 (3): 199–210. . [DOI] [Google Scholar]
- Sun GW, Riekerk H, and Korhnak LV. 1995. “Shallow Groundwater Table Dynamics of Cypress Wetland Pine Upland Systems in Florida Flatwoods.” Soil and Crop Science Society of Florida Proceedings 54: 66–71. [Google Scholar]
- Thorslund J, Cohen MJ, Jawitz JW, Destouni G, Creed IF, Rains MC, Badiou P, and Jarsjö J, unpublished data. “Evidence for Hydrological Connectivity of Geographically Isolated Wetlands.” Land Degradation and Development. [Google Scholar]
- Thurman EM 1985. Organic Geochemistry of Natural Waters. Boston, MA: Martinus Nijhoff/Dr. W. Junk Publishers. [Google Scholar]
- Tiner RW 2003. “Geographically Isolated Wetlands of the United States.” Wetlands 23 (3): 494–516. 10.1672/0277-5212(2003)023[0494:GIWOTU]2.0.CO;2. [DOI] [Google Scholar]
- Tomer MD, Wilson CG, Moorman TB, Cole KJ, Heer D, and Isenhart TM. 2010. “Source-Pathway Separation of Multiple Contaminants During a Rainfall-Runoff Event in an Artificially Drained Agricultural Watershed.” Journal of Environmental Quality 39: 882–95. 10.2134/jeq2009.0289. [DOI] [PubMed] [Google Scholar]
- Tromp-van Meerveld HJ, and McDonnell JJ. 2006. “Threshold Relations in Subsurface Stormflow: 2. The Fill and Spill Hypothesis.” Water Resources Research 42 (2): W02411. 10.1029/2004wr003800. [DOI] [Google Scholar]
- Tufford DL 2011. “Shallow Water Table Response to Seasonal and Interannual Climate Variability.” Transactions of the American Society of Agricultural and Biological Engineers 54 (6): 2079–86. [Google Scholar]
- Uden DR, Hellman ML, Angeler DG, and Allen CR. 2014. “The Role of Reserves and Anthropogenic Habitats for Functional Connectivity and Resilience of Ephemeral Wetlands.” Ecological Applications 24 (7): 1569–82. 10.1890/13-1755.1. [DOI] [PubMed] [Google Scholar]
- Ullah S, and Faulkner SP. 2006. “Denitrification Potential of Different Land-Use Types in an Agricultural Watershed, Lower Mississippi Valley.” Ecological Engineering 28 (2): 131–40. 10.1016/j.ecoleng.2006.05.007. [DOI] [Google Scholar]
- Ullrich SM, Tanton TW, and Abdrashitova SA. 2001. “Mercury in the Aquatic Environment: A Review of Factors Affecting Methylation.” Critical Reviews in Environmental Science and Technology 31 (3): 241–93. 10.1080/20016491089226. [DOI] [Google Scholar]
- Urban NR, Bayley SE, and Eisenreich SJ. 1989. “Export of Dissolved Organic Carbon and Acidity from Peatlands.” Water Resources Research 25: 1619–28. [Google Scholar]
- USACE (U.S. Army Corps of Engineers). 1987. “Wetlands Delineation Manual.” Technical Report Y-87–1, U.S. Army Corps of Engineers, Waterways Experiment Station, Wetlands Research Program, Vicksburg, MS. [Google Scholar]
- USEPA (U.S. Environmental Protection Agency). 2015. “Connectivity of Streams and Wetlands to Downstream Waters: A Review and Synthesis of the Scientific Evidence.” EPA/600/R-14/475F. Washington, D.C.: USEPA Office of Research and Development. [Google Scholar]
- Vanderhoof MK, and Alexander LC. 2016. “The Role of Lake Expansion in Altering the Wetland Landscape of the Prairie Pothole Region, United States.” Wetlands 36 (Suppl. 2): 309–21. 10.1007/s13157-015-0728-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanderhoof MK, Alexander LC, and Todd JM. 2016. “Temporal and Spatial Patterns of Wetland Extent Influence Variability of Surface Water Connectivity in the Prairie Pothole Region, United States.” Landscape Ecology 31 (4): 805–24. 10.1007/s10980-015-0290-5. [DOI] [Google Scholar]
- Vanderhoof MK, Christensen JR, and Alexander LC. 2017. “Patterns and Drivers for Wetland Connections in the Prairie Pothole Region, United States.” Wetlands Ecology and Management 25 (3): 275–97. 10.1007/s11273-016-9516-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanderhoof MK, Lane CR, McManus MG, Alexander LC, and Christensen JR, in press. “Wetlands Inform How Climate Extremes Influence Surface Water Expansion and Contraction.” Hydrology and Earth Systems Science Discussion. 10.5194/hess-2017-581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Kamp G, Stolte WJ, and Clark RG. 1999. “Dying Out of Small Prairie Wetlands After Conversion of Their Catchments from Cultivation to Permanent Brome Grass.” Hydrological Sciences Journal 44 (3): 387–99. 10.1080/02626669909492234. [DOI] [Google Scholar]
- Van Meter KJ, and Basu NB. 2015. “Signatures of Human Impact: Size Distributions and Spatial Organization of Wetlands in the Prairie Pothole Landscape.” Ecological Applications 25 (2): 451–65. 10.1890/14-0662.1. [DOI] [PubMed] [Google Scholar]
- Vining KC 2002. “Simulation of Streamflow and Wetland Storage, Starkweather Coulee Subbasin, North Dakota, Water Years 1991–98.” USGS Water-Resources Investigation Report 02–4113. [Google Scholar]
- Wang X, Shang S, Qu Z, Liu T, Melesse AM, and Yang W. 2010. “Simulated Wetland Conservation-Restoration Effects on Water Quantity and Quality at Watershed Scale.” Journal of Environmental Management 91: 1511–25. 10.1016/j.jenvman.2010.02.023. [DOI] [PubMed] [Google Scholar]
- Ward JV 1989. “The Four-Dimensional Nature of Lotic Ecosystems.” Journal of the North American Benthological Society 8 (1): 2–8. [Google Scholar]
- Watmough MD, and Schmoll MJ. 2007. Environment Canada’s Prairie & Northern Region Habitat Monitoring Program, Phase II: Recent Habitat Trends in the Prairie Habitat Joint Venture. Technical Report Series No. 493, 135 pp. Edmonton, AB: Environment Canada, Canadian Wildlife Service. [Google Scholar]
- White WR, and Crisman TL. 2016. “Headwater Streams of Florida: Types, Distribution, and a Framework for Conservation.” River Research and Applications 32 (3): 452–61. 10.1002/rra.2845. [DOI] [Google Scholar]
- Whitmire S, and Hamilton S. 2008. “Rates of Anaerobic Microbial Metabolism in Wetlands of Divergent Hydrology on a Glacial Landscape.” Wetlands 28 (3): 703–14. 10.1672/06-126.1. [DOI] [Google Scholar]
- Wilcox B, Dean D, Jacob J, and Sipocz A. 2011. “Evidence of Surface Connectivity for Texas Gulf Coast Depressional Wetlands.” Wetlands 31 (3): 451–8. 10.1007/s13157-011-0163-x. [DOI] [Google Scholar]
- Winter TC 1999. “Relation of Streams, Lakes, and Wetlands to Groundwater Flow Systems.” Hydrogeology Journal 7 (1): 28–45. 10.1007/s100400050178. [DOI] [Google Scholar]
- Winter TC, and LaBaugh JW. 2003. “Hydrologic Considerations in Defining Isolated Wetlands.” Wetlands 23 (3): 532–40. 10.1672/0277-5212(2003)023[0532:HCIDIW]2.0.CO;2. [DOI] [Google Scholar]
- Winter TC, and Rosenberry DO. 1995. “The Interaction of Ground Water with Prairie Pothole Wetlands in the Cottonwood Lake Area, East-Central North Dakota, 1979–1990.” Wetlands 15 (3): 193–211. [Google Scholar]
- Winter TC, Rosenberry DO, and LaBaugh JW. 2003. “Where Does the Ground Water in Small Watersheds Come From?” Ground Water 41: 989–1000. 10.1111/j.1745-6584.2003.tb02440.x. [DOI] [Google Scholar]
- Wolf KL, Noe GB, and Ahn C. 2013. “Hydrologic Connectivity to Streams Increases Nitrogen and Phosphorus Inputs and Cycling in Soils of Created and Natural Floodplain Wetlands.” Journal of Environmental Quality 42 (4): 1245–55. 10.2134/jeq2012.0466. [DOI] [PubMed] [Google Scholar]
- Wu Q, and Lane CR. 2016. “Delineation and Quantification of Wetland Depressions in the Prairie Pothole Region of North Dakota.” Wetlands 36 (2): 215–27. 10.1007/s13157-015-0731-6. [DOI] [Google Scholar]
- Wu Q, and Lane CR. 2017. “Delineating Wetland Catchments and Modeling Hydrologic Connectivity Using LiDAR Data and Aerial Imagery.” Hydrological and Earth Systems Sciences 21: 3579–95. 10.5194/hess-21-3579-2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang W, Wang X, Liu Y, Gabor S, Boychuk L, and Badiou P. 2010. “Simulated Environmental Effects of Wetland Restoration Scenarios in a Typical Canadian Prairie Watershed.” Wetlands Ecology and Management 18 (3): 269–79. 10.1007/s11273-009-9168-0. [DOI] [Google Scholar]
- Zhang J, James RT, Ritter G, and Sharfstein B. 2009. “Lake Okeechobee Protection Program—State of the Lake and Watershed.” In South Florida Environmental Report, edited by Redfield Gand Efron S, 10.1–10.102. West Palm Beach, FL: South Florida Water Management District. [Google Scholar]