

Abstract
Emerging viral infections, including those caused by dengue virus (DENV) and Venezuelan Equine Encephalitis virus (VEEV), pose a significant global health challenge. Here, we report the preparation and screening of a series of 4-anilinoquinoline libraries targeting DENV and VEEV. This effort generated a series of lead compounds, each occupying a distinct chemical space, including 3-((6-bromoquinolin-4-yl)amino)phenol (12), 6-bromo-N-(5-fluoro-1H-indazol-6-yl)quinolin-4-amine (50) and 6-((6-bromoquinolin-4-yl)amino)isoindolin-1-one (52), with EC50 values of 0.63–0.69 µM for DENV infection. These compound libraries demonstrated very limited toxicity with CC50 values greater than 10 µM in almost all cases. Additionally, the lead compounds were screened for activity against VEEV and demonstrated activity in the low single-digit micromolar range, with 50 and 52 demonstrating EC50s of 2.3 µM and 3.6 µM, respectively. The promising results presented here highlight the potential to further refine this series in order to develop a clinical compound against DENV, VEEV, and potentially other emerging viral threats.
Keywords: dengue virus (DENV), 4-anilinoquinoline, flavivirus, alphavirus, VEEV, antivirals, emerging viruses
1. Introduction
Mosquito-borne viral infections, including those caused by the flavivirus dengue (DENV) and the alphavirus Venezuelan Equine Encephalitis virus (VEEV), represent a major public health concern [1,2,3,4]. Rapid urbanization and climate change have contributed to the expanding geographical range of DENV infections into the developed world [5] and have caused a significant increase in the number of annual infections, currently estimated at ~400 million people in over 128 endemic countries [6,7]. The majority of symptomatic DENV infected patients experience mild illness, yet approximately 5–20%, particularly those with a secondary infection with a heterologous DENV serotype, progress into a potentially life-threatening disease known as severe dengue [8,9]. Currently, there are no approved antiviral therapies for DENV infection, and the development of an effective and safe DENV vaccine has been challenged by the need to generate a balanced protective immunity against the four distinct DENV serotypes.
The alphavirus VEEV is an important causative agent of neurological disease in Central and South America [10]. While most VEEV infections in humans are mild, approximately 14% of the patients develop encephalitis, often complicated by severe neurological deficits and sometimes death [11]. Beyond mosquito bites, VEEV can be transmitted via aerosol exposure and is considered a major bioterrorism threat, underscoring the importance of developing effective countermeasures [12]. However, there are currently no approved antiviral drugs nor licensed human vaccines available for VEEV infection. In the absence of approved antiviral therapies for DENV and VEEV infections, the management of infected patients largely relies on supportive care [12,13], resulting in continued morbidity and mortality.
In the past decade, there has been a significant effort to develop small molecules for the treatment of DENV infection, yielding multiple promising candidates with a number of different scaffolds. [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33] However, none of these compounds have entered clinical trials for DENV treatment. Interestingly, many of these compounds include common kinase inhibitor scaffolds, such as the oxindole [14], isothiazolo[4,3-b]pyridine [15] and azaindoles [16]. Another kinase scaffold with reported anti-DENV activity has been the 4-anilinoquin(az)oline, present in a number of clinically approved kinase inhibitors [34,35,36,37]. This includes erlotinib, which has shown low micromolar activity against DENV, and several other, lower molecular weight analogues of the same scaffold (1–3), showing effective, low nanomolar activity against DENV (Figure 1) [14,29,30,31,32,33].
Figure 1.

A selection of previously reported quinazolines and quinolines with anti-DENV activity (the data is all for serotype 2 for consistency).
2. Results
2.1. Synthesis
Building on our previous work, to further explore the antiviral activity of the 4-anilinoquin(az)oline [14,32,33], we probed the structure activity relationships (SAR) with a series of focused libraries. We developed a series of hybrid molecules combining structural features of erlotinib and lead compound 4 to enhance the SAR in the current literature and improve the tractability of the quin(az)oline scaffold (Figure 2).
Figure 2.

Design of a hybrid series of 4-anilinoquinolines developed by combining erlotinib and 4.
We hence synthesized a series of compounds (5–56) to create the hybrid structure of erlotinib and 4. We accessed these 4-anilinoquinolines through nucleophilic aromatic displacement of 4-chloroquin(az)olines with the respective aniline (Scheme 1) [38,39,40,41,42,43,44,45,46,47,48]. We were able to access products in full range of yields, from moderate to excellent (21–94%), consistent with previous reports for literature compounds [38,39,40,41,42,43,44,45,46,47,48].
Scheme 1.

General synthetic procedure to access the 4-anilinoquinolines.
2.2. Antiviral Screening
To evaluate the compounds for broad-spectrum antiviral activity we first studied their effect on DENV2-Rluc virus infection, a wild type virus carrying a Renilla luciferase reporter gene (virus production described in Materials and Methods). We measured the effect of compound treatment on overall infection in human hepatoma (Huh7) cells 48 h following infection with DENV2 via luciferase assays and calculated the half-maximal effective concentration (EC50) relative to DMSO treated cells. In parallel, we tested the effect of these compounds on cell viability via an AlamarBlue assay in the DENV-infected Huh7 cells and measured the half-maximal cytotoxic concentration (CC50) values relative to DMSO treated cells.
In order to follow-up on the lead compound 4, we first screened a series of focused substitutions at the aniline meta-position (5–10) (Table 1) [48]. The hydroxy analogue 5 was 3-fold less potent than the methoxy 4. The nitro substitution 6 was inactive at the top concentration, while the reduced amine 7, was 7-fold weaker with respect to 4 and 2-fold weaker with respect to the direct isosteric replacement 5. The monomethyl substituted amine 8 showed no improvement, while the dimethyl substitution showed a 2-fold increase in potency against DENV with respect to 7. The methanolic substitution 10, was 2-fold weaker with respect to 5 and 8-fold with respect 4.
Table 1.
Screening results of anti-DENV activity of initial SAR around 3-methoxy aniline substitution pattern of 4.
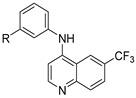
| Cmpd | R | DENV Inhibition | Cell Viability |
|---|---|---|---|
| EC50 a | CC50 b | ||
| 4 | OMe | 0.82 | >10 |
| 5 | OH | 2.7 | >10 |
| 6 | NO2 | >10 | >10 |
| 7 | NH2 | 5.9 | >10 |
| 8 | NHMe | 8.9 | >10 |
| 9 | NMe2 | 2.3 | >10 |
| 10 | CH2OH | 6.6 | >10 |
a = infectivity assay in Huh7 cells mean average n = 5; b = cytotoxicity in Huh7 cells mean average n = 5.
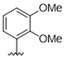
We then screened a series of matched pair meta-methoxy and meta-hydroxy analogues (11–18), changing the 6-position of the quinoline ring (Table 2) [48]. The 6-bromoquinoline methoxy analogue 11 was equipotent with the 6-triflurormethyl quinoline methoxy analogue 4. However, switching to the hydroxy 12 led to a 4-fold increase in potency against DENV (EC50 = 0.63 μM). The unsubstituted quinoline methoxy analogue 13 showed a 3-fold decrease in activity against DENV, while the hydroxy analogue 14 showed no activity at the top concentration tested. Switching to the electron donating 6-methoxyquinoline methoxy analogue 15 was 5-fold less potent with respect to 4, while the hydroxy analogue was inactive (EC50 = >10 μM). Interestingly, both the 6-methylsulfone quinoline analogues 17 and 18 were inactive.
Table 2.
Screening results of anti-DENV activity of matched pair methoxy and hydroxy aniline analogues.
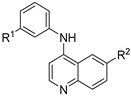
| Cmpd | R1 | R2 | DENV Inhibition |
Cell Viability |
|---|---|---|---|---|
| EC50 a | CC50 b | |||
| 4 | OMe | CF3 | 0.82 | >10 |
| 5 | OH | CF3 | 2.7 | >10 |
| 11 | OMe | Br | 0.84 | >10 |
| 12 | OH | Br | 0.63 | >10 |
| 13 | OMe | H | 2.5 | >10 |
| 14 | OH | H | >10 | >10 |
| 15 | OMe | OMe | 4.4 | >10 |
| 16 | OH | OMe | >10 | >10 |
| 17 | OMe | SO2Me | >10 | >10 |
| 18 | OH | SO2Me | >10 | >10 |
a = infectivity assay in Huh7 cells mean average n = 5; b = cytotoxicity in Huh7 cells mean average n = 5.
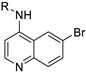
Encouraged by the result of 3-((6-bromoquinolin-4-yl)amino)phenol 12, we followed up with a series of focused analogues (19–31) fixing the 6-bromoquinoline and altering the aniline ring substituent (Table 3) [48]. The direct nitro analogue 19 and reduced amine version 20 were both inactive (EC50 = >10 μM). The substitution of the amine 20 with a methyl group to afford 21 produced a compound with a potent antiviral activity profile (EC50 = 0.24 μM), but this appeared to be in part driven by toxicity (CC50 = 5.3 μM) with a 22-fold selectivity index window. This toxicity interference appeared more likely considering 19 and 20 were inactive, as was the dimethyl analogue 22. The methanol analogue 23 was 6-fold weaker with respect to 12 and the ortho-position methanol analogue 24 a further 2-fold weaker with respect to 23. We then tested the scope of the meta-position further with several bulkier substituents (25–27). The pentafluorosulfanyl 25 was just over 2-fold less potent than 12, demonstrating that there is potentially room for expansion at this position [49]. The tert-butoxy 26 2-fold less potent than 12, with the tert-butyl 27 near equipotent (EC50 = 0.94 μM), unfortunately both showed a level of toxicity (CC50 = 9.2 μM and CC50 = 6.0 μM respectively). We then explored how torsional strain on the methoxy orientation would affect activity (28–31). The fused ring systems were overall less active against DENV, only 28 showed activity (EC50 = 1.7 μM) which was accompanied by light toxicity (CC50 = 8.6 μM).
Table 3.
Screening results of anti-DENV activity of analogues of 12.
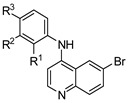
| Cmpd | R1 | R2 | R3 | DENV Inhibition | Cell Viability |
|---|---|---|---|---|---|
| EC50 a | CC50 b | ||||
| 12 | H | OH | H | 0.63 | >10 |
| 19 | H | NO2 | H | >10 | >10 |
| 20 | H | NH2 | H | >10 | >10 |
| 21 | H | NHMe | H | 0.24 | 5.3 |
| 22 | H | NMe2 | H | >10 | >10 |
| 23 | H | CH2OH | H | 3.7 | >10 |
| 24 | CH2OH | H | H | 7.1 | >10 |
| 25 | H | SF5 | H | 1.5 | >10 |
| 26 | H | OtBu | H | 2.3 | 9.2 |
| 27 | H | tBu | H | 0.94 | 6.0 |
| 28 | H | OCH2O | 1.7 | 8.6 | |
| 29 | H | CH2OCH2 | >10 | >10 | |
| 30 | OCH2O | H | >10 | >10 | |
| 31 | CH2OCH2 | H | >10 | >10 | |
a = infectivity assay in Huh7 cells mean average n = 5; b = cytotoxicity in Huh7 cells mean average n = 5.
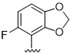
Predicting that we would identify active analogues based on the 2,3- and 3,4-connectivty, we followed up on the results of 28–31 with a series of different substitutions and fused heterocycles (32–56) (Table 4) [42]. The introduction of a fluorine to the distal ortho-position on the aniline ring of 30 produced compound 32 with good activity against DENV (IC50 = 1.9 μM). The ring opened version of 30, 33 was 4-fold weaker against DENV. The substitution of the catechol carbon linker of 30 with a gem-difluoro 34 afforded a compound with equipotency with the respective ring opened analogue 33 but was slightly more active than the parent 30. The removal of the ortho-substituted oxygen of 30 furnished 35, a compound with sub-micromolar activity against DENV (EC50 = 0.95 μM). However, the removal of the meta-substituted oxygen of 30 to afford 36 led to a 6-fold decrease in potency compared with 35. Aromatising 35 to yield 37 led to a 3-fold decrease in potency (EC50 = 2.7 μM), whereas aromatising 36 to afford 38 demonstrated a 3-fold increase in potency (EC50 = 1.9 μM). The formation of the respective isoxazole’s of 37 and 38, affording 39 and 40, produced inactive compounds at the top concentration tested (EC50 = >10 μM). Moving the nitrogen of 39 to form the oxazole 41 led to a >3-fold increase in potency (EC50 = 3.2 μM). The reversed thiazole 42, was not active (EC50 = >10 μM), but removal of the nitrogen from the thiazole ring system afforded a thiophene 43 that had equipotent activity with oxazole 41.
Table 4.
Screening results of anti-DENV activity following up on 28–31.
|
|
||||||
|---|---|---|---|---|---|---|---|
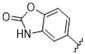
| Cmpd | R | DENV Inhibition | Cell Viability | Cmpd | R | DENV Inhibition | Cell Viability |
| EC50 a | CC50 b | EC50 a | CC50 b | ||||
| 32 |
|
1.9 | >10 | 45 |
|
5.8 | >10 |
| 33 |
|
7.8 | >10 | 46 |
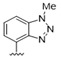
|
5.5 | >10 |
| 34 |
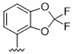
|
7.1 | >10 | 47 |
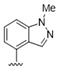
|
5.4 | >10 |
| 35 |
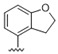
|
0.95 | >10 | 48 |
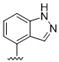
|
2.1 | >10 |
| 36 |
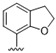
|
5.7 | >10 | 49 |
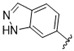
|
1.8 | >10 |
| 37 |
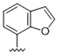
|
2.7 | >10 | 50 |
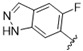
|
0.69 | >10 |
| 38 |
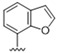
|
1.9 | >10 | 51 |
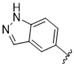
|
1.2 | >10 |
| 39 |
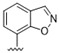
|
>10 | >10 | 52 |
|
0.64 | >10 |
| 40 |
|
>10 | >10 | 53 |
|
>10 | >10 |
| 41 |
|
3.2 | >10 | 54 |
|
>10 | >10 |
| 42 |
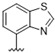
|
>10 | >10 | 55 |
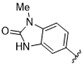
|
7.1 | >10 |
| 43 |
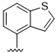
|
2.9 | >10 | 56 |
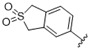
|
6.3 | >10 |
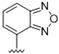
| 44 |
|
9.2 | >10 | - | - | - | - |
a = infectivity assay in Huh7 cells mean average n = 5; b = cytotoxicity in Huh7 cells mean average n = 5.
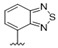
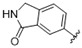
A furazan substitution 44 only showed limited activity against DENV (EC50 = 9.2 μM), whereas the introduction of a sulphur to construct a 1,2,5-thiadiazole 45 led to a 2-fold increase in potency compared with 44. The introduction of a methyl distal to the quinoline ring system via a 1-methyltriazole 46 or 1-methyl-1H-pyrazole 47 led to compounds with approximately a 5 micromolar EC50 efficacy against DENV. The removal of the methyl of 47 led to a >2-fold increase against DENV; interestingly, the reversed pyrazole (2,3 vs. 1,2) 49 was equipotent with 48. The introduction of a fluorine in the distal ortho-position of the aniline in 49 produced compound 50 and led to an almost 3-fold increase against DENV with efficacy exceeding 1 micromolar (EC50 = 0.69 μM). Moving the pyrazole from 2,3- to the 3-4-postion; 48 to 51 led to a 2-fold boost in efficacy (EC50 = 1.2 μM). An increase in diversity to produce 6-((6-bromoquinolin-4-yl)amino)isoindolin-1-one (52) led to an equipotent inhibitor compared with 49, though occupying a different chemical space. Several attempts to increase potency with 1,3-dihydro-2H-benzo[d]imidazol-2-one derivatives (53–55) and a fused cyclic sulfone (56) yielded compounds with efficacy only in the high micromolar range.
2.3. Extension Screening of Lead Compounds
The parental compound (4) and the lead compounds (12, 21, 50, 52) were then screened against VEEV (Table 5) [17]. We infected U-87 MG (human astrocytes) cells with the VEEV vaccine strain (TC-83) carrying a nanoluciferase reporter gene (virus production described in Materials and Methods) and measured the effect of compounds on viral replication via luciferase assays and cell viability via AlamarBlue assay at 18 h post-infection. The parental compound 4 was found to be inactive against VEEV at the top concentration tested. The 3-((6-bromoquinolin-4-yl)amino)phenol analogue 12 was also weakly active (EC50 = 9.9 μM). However, the N1-(6-bromoquinolin-4-yl)-N3-methylbenzene-1,3-diamine 21 showed good activity (EC50 = 1.0 μM) without the associated mild toxicity observed in Huh7 cells. Compounds 50 and 52 also both performed well with a low micromolar potency against VEEV (EC50 = 2.3 μM and 3.6 μM, respectively). Consistent with our previous reports, the antiviral effect of sunitinib was in the submicromolar range [17,36]. As an additional toxicity control, we screened these lead compounds for their effect on cellular viability in Vero (African green monkey kidney epithelial) cells [50] and found no toxicity at the top concentration tested (10 μM).
Table 5.
Extension screening on VEEV and on Vero cells for lead compounds 4, 12, 21, 50 and 52.
| Cmpd | DENV Inhibition a | VEEV Inhibition b | Vero Cells c | ||||
|---|---|---|---|---|---|---|---|
| EC50 | EC90 | CC50 | EC50 | EC90 | CC50 | CC50 | |
| 4 | 0.82 | 1.5 | >10 | >10 | >10 | >10 | >10 |
| 12 | 0.63 | 3.2 | >10 | 9.9 | >10 | >10 | >10 |
| 21 | 0.24 | 0.69 | 5.3 | 1.0 | >10 | >10 | >10 |
| 50 | 0.69 | 4.0 | >10 | 2.3 | 9.6 | >10 | >10 |
| 52 | 0.64 | 1.5 | >10 | 3.6 | >10 | >10 | >10 |
| Sunitinib | 1.1 | >10 | >10 | 4.4 | >10 | >10 | >10 |
a = infectivity assay in Huh7 cells mean average n = 5; cytotoxicity in Huh7 cells mean average n = 5; b = infectivity assay in U-87 MG cells mean average n = 5; cytotoxicity in U-87 MG cells mean average n = 5; c = cytotoxicity 48 h in Vero cells.
The lead compounds against DENV include 3-((6-bromoquinolin-4-yl)amino)phenol (12) (EC50 = 0.63 µM), 6-bromo-N-(5-fluoro-1H-indazol-6-yl)quinolin-4-amine (50) (EC50 = 0.69 µM) and 6-((6-bromoquinolin-4-yl)amino)isoindolin-1-one (52) (EC50 = 0.64 µM) (Figure 3). Each of these compounds occupy a distinct chemical space and enhances our understanding of the SAR within this series. We also identified N1-(6-bromoquinolin-4-yl)-N3-methylbenzene-1,3-diamine (21) with potent activity (EC50 = 0.24 µM), but some associated toxicity (CC50 = 5.3 µM) in the same Huh7 cell line (Figure 3). This toxicity was not observed in the Vero cell line secondary screening (CC50 = >10 µM). Compounds 21, 50 and 52 all performed well against VEEV albeit with a 4-8-fold reduction in potency relative to DENV (Figure 3). Interestingly, 4 and 12 showed weak activity toward VEEV despite a good anti-DENV activity.
Figure 3.

Compounds 21, 50 and 52 suppress both DENV2 and VEEV (TC-83) infection. Dose response of DENV2 (black) and cell viability (blue) to compounds A: 21, B: 50, C: 52 and dose response of VEEV (TC-83) (black) and cell viability (blue) to compounds D: 21, E: 50, F: 52; each measured by luciferase and AlamarBlue assays, 48 h post-infection (A–C) or 18 h post-infection (D–F). Data are plotted relative to DMSO control. Shown are representative experiments from at least two conducted, each with 4 biological replicates; shown are means ± SD.
3. Discussion
We have previously demonstrated that the 4-anilinoquinoline/quinazoline scaffold is active on both DENV and VEEV [32,33]. The current results re-enforce our interest in the 4-anilinoquin(az)oline scaffold, with the relatively low molecular weight compound 12 showing good potency (EC50 = 0.63 µM). Despite some mild toxicity (EC50 = 5.3 µM), the direct methylamine analogue 21 was even more potent (EC50 = 0.24 µM) albeit with a reduced therapeutic index. The other two lead compounds identified were 6-bromo-N-(5-fluoro-1H-indazol-6-yl)quinolin-4-amine (50) (EC50 = 0.69 µM) and 6-((6-bromoquinolin-4-yl)amino)isoindolin-1-one (52) (EC50 = 0.64 µM), both of which are isosteric replacements for the original trimethoxy of 3 and related to 3-methoxy of 4. Three of our four lead compounds (21, 50 and 52) have shown promising antiviral activity against both DENV and VEEV and offer interesting insights into the SAR within this series.
While the mechanism of the antiviral activity on both viruses is being investigated, potentially it could be targeting multiple protein targets including both cellular kinases and other proteins. Several of these compounds were originally prepared as inhibitors targeting the ATP-binding site of human kinases including cyclin-G-associated kinase (GAK), serine/threonine-protein kinase 10 (STK10) and STE20-like serine/threonine-protein kinase (SLK) [42,47,48]. While our compounds could be targeting these kinases, it is also possible that other undefined proteins may participate in the mechanism of antiviral action. Multiple examples from previous literature show that compounds with structural features impeding interaction with a kinase hinge region, have antiviral activity [35]. It is also possible that the observed phenotypes may originate from modulation of other kinases or non-ATP binding proteins [51]. Lastly, beyond targeting cellular kinases, it is possible that these compounds target a viral protein. Indeed, related quinolinones were identified as VEEV inhibitors targeting the viral nonstructural protein (nsP2) [52,53].
This set of results presents a potential step towards the identification of a novel clinical compound to combat emerging viral diseases including DENV and VEEV infections and provides a medicinal chemistry trajectory to achieve this aim. In addition, several quinoline derivatives have been shown to inhibit alpha- and beta-coronaviruses [54], demonstrating the promising potential of quinoline group as part of a broader spectrum antiviral compound.
4. Materials and Methods
4.1. Chemistry Method
General Procedure for the Synthesis of 4-Anilinoquin(az)olines
We suspended 4-chlo-quin(az)oline derivative (1.0 eq.), aniline derivative (1.1 eq.), in ethanol (10 mL) and refluxed for 18 h. The crude mixture was purified by flash chromatography using EtOAc:hexane followed by 1–5% methanol in EtOAc; After solvent removal under reduced pressure, the product was obtained as a free following solid or recrystallized from ethanol/water. Compounds 4–31 were synthesized as previous described [38,48], 32-51 were synthesized as previously described [42]. Representative supporting information provided (see Supplementary Materials).
4.2. Antiviral Screening
4.2.1. Virus Construct
DENV2 (New Guinea C strain) [55,56] Renilla reporter plasmid used for virus production (DENV2-Rluc) was a gift from Pei-Yong Shi (The University of Texas Medical Branch). The plasmid encoding infectious VEEV (TC-83) with a nanoluciferase reporter and used for virus production (VEEV-TC-83-nLuc) was a gift from Dr. William B. Klimstra (Department of Immunology, University of Pittsburgh) [57].
4.2.2. Cells
Huh7 (Apath, L.L.C, New York, NY, USA) cells were grown in Dulbecco’s Modified Eagle’s medium (DMEM) (10-013-CV: Corning, New York, NY, USA) supplemented with 10% fetal bovine serum (FBS) (Omega Scientific, Tarzana, CA, USA), nonessential amino acids, 1% L-glutamine, and 1% penicillin-streptomycin (Thermo Fisher Scientific, Waltham, MA, USA). U-87 MG cells obtained from ATCC were grown in DMEM supplemented with 10% FBS and 1% penicillin-streptomycin solution. Cells were maintained in a humidified incubator with 5% CO2 at 37 °C.
4.2.3. Virus Production
DENV2-Rluc RNA was transcribed in vitro using mMessage/mMachine (Ambion Austin, TX, USA) kits. DENV2-Rluc virus was produced by electroporating RNA into BHK-21 cells, harvesting supernatants on day 10 post-electroporation and titering via standard plaque assays on BHK-21 cells. VEEV-TC-83-nLuc RNA was transcribed in vitro from cDNA plasmid template linearized with MluI via MegaScript Sp6 kit (Invitrogen #AM1330, Thermo Fisher Scientific, Waltham, MA, USA) and electroporated into BHK-21 cells. The VEEV-TC-83-nLuc virus was harvested from the supernatant 24 h post-electroporation, clarified from cell debris and the titer determined by standard plaque assay on Vero cells.
4.2.4. Infection Assays
Huh7 cells were treated with the inhibitors or DMSO. An hour later, the cells were infected with DENV2-Rluc virus in replicates (n = 5) at a multiplicity of infection (MOI) of 0.05. The inhibitors were present for the duration of the experiment. Overall infection was measured at 48 h post-infection using a Renilla luciferase substrate. U-87 MG cells were treated with the inhibitors or DMSO. One hour later, the cells were infected with VEEV-TC-83-nLuc virus in 5 replicates at MOI of 0.01, and overall infection was measured at 18 h post-infection via a nanoluciferase assay. The inhibitors were present for the duration of the experiment. The relative light units (RLUs) were normalized to DMSO treated cells (set as 100%). GraphPad Prism nonlinear regression (curve fit) was used to generate the graphs and EC50 values.
4.2.5. Viability Assays
Viability was measured using AlamarBlue® reagent (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocol. Fluorescence was detected at 560 nm on InfiniteM1000 plate reader (Tecan, Männedorf, Switzerland). The raw fluorescence values were normalized to DMSO treated cells (set as 100%). Graphpad Prism nonlinear regression (curve fit) was used to generate the graphs and CC50 values.
Acknowledgments
This work was supported by award number W81XWH2110456 from the Department of Defense (DoD) to S.E. and C.R.M.A., Congressionally Directed Medical Research Programs (CDMRP) and HDTRA11810039 from the Defense Threat Reduction Agency (DTRA)/Fundamental Research to Counter Weapons of Mass Destruction to SE. The SGC is a registered charity (number 1097737) that receives funds from AbbVie, Bayer Pharma AG, Boehringer Ingelheim, Canada Foundation for Innovation, Eshelman Institute for Innovation, Genome Canada, Innovative Medicines Initiative (EU/EFPIA) [ULTRA-DD grant no. 115766], Janssen, Merck KGaA Darmstadt Germany, MSD, Novartis Pharma AG, Ontario Ministry of Economic Development and Innovation, Pfizer, São Paulo Research Foundation-FAPESP, Takeda, and Wellcome [106169/ZZ14/Z]. We are grateful for LC−MS/HRMS support provided by Brandie Ehrmann and Diane E. Wallace in the Mass Spectrometry Core Laboratory at the University of North Carolina at Chapel Hill. The core is supported by the National Science Foundation under grant no. CHE-1726291. The opinions, interpretations, conclusions, and recommendations are those of the authors and are not necessarily endorsed by the U.S. Army or the other funders.
Supplementary Materials
The following are available online, representative LC-HRMS, 1H and 13C NMR spectra on the lead compound compounds.
Author Contributions
S.E., C.R.M.A. conceived and designed the study; P.-T.H., S.S., C.R.M.A. performed the experiments; P.-T.H., S.S., S.E., C.R.M.A. analyzed the data; P.-T.H., S.S., S.E., C.R.M.A. edited the paper; C.R.M.A. wrote the paper and all authors approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds are available from the authors.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Paixão E.S., Teixeira M.G., Rodrigues L.C. Zika, chikungunya and dengue: The causes and threats of new and re-emerging arboviral diseases. BMJ Glob. Health. 2017;3:e000530. doi: 10.1136/bmjgh-2017-000530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Pierson T.C., Diamond M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020;5:796–812. doi: 10.1038/s41564-020-0714-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Braack L., de Almeida A.P.G., Cornel A.J., Swanepoel R., de Jager C. Mosquito-borne arboviruses of African origin: Review of key viruses and vectors. Parasit Vectors. 2018;11:29. doi: 10.1186/s13071-017-2559-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Silva N.M., Santos N.C., Martins I.C. Dengue and Zika Viruses: Epidemiological History, Potential Therapies, and Promising Vaccines. Trop. Med. Infect. Dis. 2020;5:150. doi: 10.3390/tropicalmed5040150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Campbell L.P., Luther C., Moo-Llanes D., Ramsey J.M., Danis-Lozano R., Peterson A.T. Climate change influences on global distributions of dengue and chikungunya virus vectors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015;370:20140135. doi: 10.1098/rstb.2014.0135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Murray N.E., Quam M.B., Wilder-Smith A. Epidemiology of dengue: Past, present and future prospects. Clin. Epidemiol. 2013;5:299–309. doi: 10.2147/CLEP.S34440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Guzman M.G., Gubler D.J., Izquierdo A., Martinez E., Halstead S.B. Dengue infection. Nat. Rev. Dis. Primers. 2016;2:16055. doi: 10.1038/nrdp.2016.55. [DOI] [PubMed] [Google Scholar]
- 8.Flipse J., Smit J.M. The Complexity of a Dengue Vaccine: A Review of the Human Antibody Response. PLoS Negl. Trop. Dis. 2015;9:e0003749. doi: 10.1371/journal.pntd.0003749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yang Y., Meng Y., Halloran M.E., Longini I.M., Jr. Dependency of Vaccine Efficacy on Preexposure and Age: A Closer Look at a Tetravalent Dengue Vaccine. Clin. Infect. Dis. 2018;66:178–184. doi: 10.1093/cid/cix766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sharma A., Knollmann-Ritschel B. Current Understanding of the Molecular Basis of Venezuelan Equine Encephalitis Virus Pathogenesis and Vaccine Development. Viruses. 2019;11:164. doi: 10.3390/v11020164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Guzmán-Terán C., Calderón-Rangel A., Rodriguez-Morales A., Mattar S. Venezuelan equine encephalitis virus: The problem is not over for tropical America. Ann. Clin. Microbiol Antimicrob. 2020;19:19. doi: 10.1186/s12941-020-00360-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hawley R.J., Eitzen E.M., Jr. Biological weapons—A primer for microbiologists. Annu. Rev. Microbiol. 2001;55:235–253. doi: 10.1146/annurev.micro.55.1.235. [DOI] [PubMed] [Google Scholar]
- 13.Stevens A.J., Gahan M.E., Mahalingam S., Keller P.A. The medicinal chemistry of dengue fever. J. Med. Chem. 2009;52:7911–7926. doi: 10.1021/jm900652e. [DOI] [PubMed] [Google Scholar]
- 14.Bekerman E., Neveu G., Shulla A., Brannan J., Pu S.Y., Wang S., Xiao F., Barouch-Bentov R., Bakken R.R., Mateo R., et al. Anticancer kinase inhibitors impair intracellular viral trafficking and exert broad-spectrum antiviral effects. J. Clin. Investig. 2017;127:1338. doi: 10.1172/JCI89857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pu S.Y., Wouters R., Schor S., Rozenski J., Barouch-Bentov R., Prugar L.I., O’Brien C.M., Brannan J.M., Dye J.M., Herdewijn P., et al. Optimization of Isothiazolo[4,3-b]pyridine-Based Inhibitors of Cyclin G Associated Kinase (GAK) with Broad-Spectrum Antiviral Activity. J. Med. Chem. 2018;61:6178. doi: 10.1021/acs.jmedchem.8b00613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Verdonck S., Pu S.Y., Sorrell F.J., Elkins J.M., Froeyen M., Gao L.J., Prugar L.I., Dorosky D.E., Brannan J.M., Barouch-Bentov R., et al. Synthesis and Structure-Activity Relationships of 3,5-Disubstituted-pyrrolo[2,3-b]pyridines as Inhibitors of Adaptor-Associated Kinase 1 with Antiviral Activity. J. Med. Chem. 2019;62:5810. doi: 10.1021/acs.jmedchem.9b00136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nitsche C., Steuer C., Klein C.D. Arylcyanoacrylamides as inhibitors of the Dengue and West Nile virus proteases. Bioorg. Med. Chem. 2011;19:7318. doi: 10.1016/j.bmc.2011.10.061. [DOI] [PubMed] [Google Scholar]
- 18.Yang C.C., Hu H.S., Wu R.H., Wu S.H., Lee S.J., Jiaang W.T., Chern J.H., Huang Z.S., Wu H.N., Chang C.M., et al. A novel dengue virus inhibitor, BP13944, discovered by high-throughput screening with dengue virus replicon cells selects for resistance in the viral NS2B/NS3 protease. Antimicrob. Agents Chemother. 2014;58:110. doi: 10.1128/AAC.01281-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Saudi M., Zmurko J., Kaptein S., Rozenski J., Neyts J., Van Aerschot A. Synthesis and evaluation of imidazole-4,5- and pyrazine-2,3-dicarboxamides targeting dengue and yellow fever virus. Eur. J. Med. Chem. 2014;87:529. doi: 10.1016/j.ejmech.2014.09.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Behnam M.A.M., Graf D., Bartenschlager R., Zlotos D.P., Klein C.D. Discovery of Nanomolar Dengue and West Nile Virus Protease Inhibitors Containing a 4-Benzyloxyphenylglycine Residue. J. Med. Chem. 2015;58:9354. doi: 10.1021/acs.jmedchem.5b01441. [DOI] [PubMed] [Google Scholar]
- 21.Yokokawa F., Nilar S., Noble C.G., Lim S.P., Rao R., Tania S., Wang G., Lee G., Hunziker J., Karuna R., et al. Discovery of Potent Non-Nucleoside Inhibitors of Dengue Viral RNA-Dependent RNA Polymerase from a Fragment Hit Using Structure-Based Drug Design. J. Med. Chem. 2016;59:3935. doi: 10.1021/acs.jmedchem.6b00143. [DOI] [PubMed] [Google Scholar]
- 22.Bardiot D., Koukni M., Smets W., Carlens G., McNaughton M., Kaptein S., Dallmeier K., Chaltin P., Neyts J., Marchand A. Discovery of Indole Derivatives as Novel and Potent Dengue Virus Inhibitors. J. Med. Chem. 2018;61:8390. doi: 10.1021/acs.jmedchem.8b00913. [DOI] [PubMed] [Google Scholar]
- 23.Millies B., von Hammerstein F., Gellert A., Hammerschmidt S., Barthels F., Göppel U., Immerheiser M., Elgner F., Jung N., Basic M., et al. Proline-Based Allosteric Inhibitors of Zika and Dengue Virus NS2B/NS3 Proteases. J. Med. Chem. 2019;62:11359. doi: 10.1021/acs.jmedchem.9b01697. [DOI] [PubMed] [Google Scholar]
- 24.Xu J., Xie X., Chen H., Zou J., Xue Y., Ye N., Shi P.Y., Zhou J. Design, synthesis and biological evaluation of spiropyrazolopyridone derivatives as potent dengue virus inhibitors. Bioorg. Med. Chem Lett. 2020;30:127162. doi: 10.1016/j.bmcl.2020.127162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chen W.C., Simanjuntak Y., Chu L.W., Ping Y.H., Lee Y.L., Lin Y.L., Li W.S. Benzenesulfonamide Derivatives as Calcium/Calmodulin-Dependent Protein Kinase Inhibitors and Antiviral Agents against Dengue and Zika Virus Infections. J. Med. Chem. 2020;63:1313. doi: 10.1021/acs.jmedchem.9b01779. [DOI] [PubMed] [Google Scholar]
- 26.Venkatesham A., Saudi M., Kaptein S., Neyts J., Rozenski J., Froeyen M., Van Aerschot A. Aminopurine and aminoquinazoline scaffolds for development of potential dengue virus inhibitors. Eur. J. Med. Chem. 2017;126:101. doi: 10.1016/j.ejmech.2016.10.008. [DOI] [PubMed] [Google Scholar]
- 27.Yang Y., Cao L., Gao H., Wu Y., Wang Y., Fang F., Lan T., Lou Z., Rao Y. Discovery, Optimization, and Target Identification of Novel Potent Broad-Spectrum Antiviral Inhibitors. J. Med. Chem. 2019;62:4056. doi: 10.1021/acs.jmedchem.9b00091. [DOI] [PubMed] [Google Scholar]
- 28.Opsenica I., Burnett J.C., Gussio R., Opsenica D., Todorović N., Lanteri C.A., Sciotti R.J., Gettayacamin M., Basilico N., Taramelli D., et al. A chemotype that inhibits three unrelated pathogenic targets: The botulinum neurotoxin serotype A light chain, P. falciparum malaria, and the Ebola filovirus. J. Med. Chem. 2011;54:1157. doi: 10.1021/jm100938u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chao B., Tong X.K., Tang W., Li D.W., He P.L., Garcia J.M., Zeng L.M., Gao A.H., Yang L., Li J., et al. Discovery and optimization of 2,4-diaminoquinazoline derivatives as a new class of potent dengue virus inhibitors. J. Med. Chem. 2012;55:3135. doi: 10.1021/jm2015952. [DOI] [PubMed] [Google Scholar]
- 30.Vincetti P., Caporuscio F., Kaptein S., Gioiello A., Mancino V., Suzuki Y., Yamamoto N., Crespan E., Lossani A., Maga G., et al. Discovery of Multitarget Antivirals Acting on Both the Dengue Virus NS5-NS3 Interaction and the Host Src/Fyn Kinases. J. Med. Chem. 2015;58:4964. doi: 10.1021/acs.jmedchem.5b00108. [DOI] [PubMed] [Google Scholar]
- 31.Wang Q.Y., Patel S.J., Vangrevelinghe E., Xu H.Y., Rao R., Jaber D., Schul W., Gu F., Heudi O., Ma N.L., et al. A small-molecule dengue virus entry inhibitor. Antimicrob. Agents Chemother. 2009;53:1823. doi: 10.1128/AAC.01148-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Saul S., Pu S.Y., Zuercher W.J., Einav S., Asquith C.R.M. Potent antiviral activity of novel multi-substituted 4-anilinoquin(az)olines. Bioorg. Med. Chem. Lett. 2020;30:127284. doi: 10.1016/j.bmcl.2020.127284. [DOI] [PubMed] [Google Scholar]
- 33.Saul S., Huang P.T., Einav S., Asquith C.R.M. Evaluation and identification of 4-anilinoquin(az)olines as potent inhibitors of both dengue virus (DENV) and Venezuelan equine encephalitis virus (VEEV) Bioorg. Med. Chem. Lett. 2021:128407. doi: 10.1016/j.bmcl.2021.128407. [DOI] [PubMed] [Google Scholar]
- 34.Davis M.I., Hunt J.P., Herrgard S., Ciceri P., Wodicka L.M., Pallares G., Hocker M., Treiber D.K., Zarrinkar P.P. Comprehensive analysis of kinase inhibitor selectivity. Nat. Biotechnol. 2011;29:1046–1051. doi: 10.1038/nbt.1990. [DOI] [PubMed] [Google Scholar]
- 35.Fabian M.A., Biggs W.H., Treiber D.K., Atteridge C.E., Azimioara M.D., Benedetti M.G., Carter T.A., Ciceri P., Edeen P.T., Floyd M., et al. A small molecule-kinase interaction map for clinical kinase inhibitors. Nat. Biotechnol. 2005;23:329–336. doi: 10.1038/nbt1068. [DOI] [PubMed] [Google Scholar]
- 36.Klaeger S., Heinzlmeir S., Wilhelm M., Polzer H., Vick B., Koenig P.-A., Reinecke M., Ruprecht B., Petzoldt S., Meng C., et al. The target landscape of clinical kinase drugs. Science. 2017;358:eaan4368. doi: 10.1126/science.aan4368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Attwood M.M., Fabbro D., Sokolov A.V., Knapp S., Schiöth H.B. Trends in kinase drug discovery: Targets, indications and inhibitor design. Nat. Rev. Drug Discov. 2021 doi: 10.1038/s41573-021-00252-y. [DOI] [PubMed] [Google Scholar]
- 38.Asquith C.R.M., Laitinen T., Bennett J.M., Godoi P.H., East M.P., Tizzard G.H., Graves L.M., Johnson G.L., Dornsife R.E., Wells C.I., et al. Identification and optimization of 4-anilinoquinolines as inhibitors of cyclin G associated kinase. ChemMedChem. 2018;13:48–66. doi: 10.1002/cmdc.201700663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Asquith C.R.M., Berger B.T., Wan J., Bennett J.M., Capuzzi S.J., Crona D.J., Drewry D.H., East M.P., Elkins J.M., Fedorov O., et al. SGC-GAK-1: A Chemical Probe for Cyclin G Associated Kinase (GAK) J. Med. Chem. 2019;62:2830. doi: 10.1021/acs.jmedchem.8b01213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Asquith C.R.M., Naegeli K.M., East M.P., Laitinen T., Havener T.M., Wells C.I., Johnson G.L., Drewry D.H., Zuercher W.J., Morris D.C. Design of a Cyclin G Associated Kinase (GAK)/Epidermal Growth Factor Receptor (EGFR) Inhibitor Set to Interrogate the Relationship of EGFR and GAK in Chordoma. J. Med. Chem. 2019;62:4772–4778. doi: 10.1021/acs.jmedchem.9b00350. [DOI] [PubMed] [Google Scholar]
- 41.Asquith C.R.M., Treiber D.K., Zuercher W.J. Utilizing comprehensive and mini-kinome panels to optimize the selectivity of quinoline inhibitors for cyclin G associated kinase (GAK) Bioorg. Med. Chem. Lett. 2019;29:1727. doi: 10.1016/j.bmcl.2019.05.025. [DOI] [PubMed] [Google Scholar]
- 42.Asquith C.R.M., Bennett J.M., Su L., Laitinen T., Elkins J.M., Pickett J.E., Wells C.I., Li Z., Willson T.M., Zuercher W.J. Towards the Development of an In vivo Chemical Probe for Cyclin G Associated Kinase (GAK) Molecules. 2019;24:4016. doi: 10.3390/molecules24224016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Asquith C.R.M., Fleck N., Torrice C.D., Crona D.J., Grundner C., Zuercher W.J. Anti-tubercular activity of novel 4-anilinoquinolines and 4-anilinoquinazolines. Bioorg. Med. Chem. Lett. 2019;29:2695–2699. doi: 10.1016/j.bmcl.2019.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Asquith C.R.M., Maffuid K.A., Laitinen T., Torrice C.D., Tizzard G.J., Crona D.J., Zuercher W.J. Targeting an EGFR Water Network with 4-Anilinoquin(az)oline Inhibitors for Chordoma. ChemMedChem. 2019;14:1693–1700. doi: 10.1002/cmdc.201900428. [DOI] [PubMed] [Google Scholar]
- 45.Carabajal M.A., Asquith C.R.M., Laitinen T., Tizzard G.J., Yim L., Rial A., Chabalgoity J.A., Zuercher W.J., García Véscovi E. Quinazoline-Based Antivirulence Compounds Selectively Target Salmonella PhoP/PhoQ Signal Transduction System. Antimicrob. Agents Chemother. 2019;64:e01744-19. doi: 10.1128/AAC.01744-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Asquith C.R.M., Laitinen T., Wells C.I., Tizzard G.J., Zuercher W.J. New Insights into 4-Anilinoquinazolines as Inhibitors of Cardiac Troponin I-Interacting Kinase (TNNi3K) Molecules. 2020;25:1697. doi: 10.3390/molecules25071697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Asquith C.R.M., Laitinen T., Bennett J.M., Wells C.I., Elkins J.M., Zuercher W.J., Tizzard G.J., Poso A. Design and Analysis of the 4-Anilinoquin(az)oline Kinase Inhibition Profiles of GAK/SLK/STK10 Using Quantitative Structure-Activity Relationships. ChemMedChem. 2020;15:26. doi: 10.1002/cmdc.201900521. [DOI] [PubMed] [Google Scholar]
- 48.Asquith C.R.M., Tizzard G.J., Bennett J.M., Wells C.I., Elkins J.M., Willson T.M., Poso A., Laitinen T. Targeting the Water Network in Cyclin G-Associated Kinase (GAK) with 4-Anilino-quin(az)oline Inhibitors. ChemMedChem. 2020;15:1200. doi: 10.1002/cmdc.202000150. [DOI] [PubMed] [Google Scholar]
- 49.Savoie P.R., Welch J.T. Preparation and utility of organic pentafluorosulfanyl-containing compounds. Chem. Rev. 2015;115:1130–1190. doi: 10.1021/cr500336u. [DOI] [PubMed] [Google Scholar]
- 50.Ammerman N.C., Beier-Sexton M., Azad A.F. Growth and maintenance of Vero cell lines. Curr. Protoc. Microbiol. 2008;11:A-4E. doi: 10.1002/9780471729259.mca04es11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Munoz L. Non-kinase targets of protein kinase inhibitors. Nat. Rev. Drug Discov. 2017;16:424–440. doi: 10.1038/nrd.2016.266. [DOI] [PubMed] [Google Scholar]
- 52.Haese N.N., May N.A., Taft-Benz S., Moukha-Chafiq O., Madadi N., Zhang S., Karyakarte S.D., Rodzinak K.J., Nguyen T.H., Denton M., et al. Identification of Quinolinones as Antivirals against Venezuelan Equine Encephalitis Virus. Antimicrob Agents Chemother. 2021;65:e0024421. doi: 10.1128/AAC.00244-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chung D.H., Jonsson C.B., Tower N.A., Chu Y.K., Sahin E., Golden J.E., Noah J.W., Schroeder C.E., Sotsky J.B., Sosa M.I., et al. Discovery of a novel compound with anti-venezuelan equine encephalitis virus activity that targets the nonstructural protein 2. PLoS Pathog. 2014;10:e1004213. doi: 10.1371/journal.ppat.1004213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Persoons L., Vanderlinden E., Vangeel L., Wang X., Do N.D.T., Foo S.C., Leyssen P., Neyts J., Jochmans D., Schols D., et al. Broad spectrum anti-coronavirus activity of a series of anti-malaria quinoline analogues. Antiviral Res. 2021;193:105127. doi: 10.1016/j.antiviral.2021.105127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Xie X., Gayen S., Kang C., Yuan Z., Shi P.-Y. Membrane topology and function of dengue virus NS2A protein. J. Virol. 2013;87:4609. doi: 10.1128/JVI.02424-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Zou G., Xu H.Y., Qing M., Wang Q.Y., Shi P.Y. Development and characterization of a stable luciferase dengue virus for high-throughput screening. Antiviral Res. 2011;91:11. doi: 10.1016/j.antiviral.2011.05.001. [DOI] [PubMed] [Google Scholar]
- 57.Sun C., Gardner C.L., Watson A.M., Ryman K.D., Klimstra W.B. Stable, high-level expression of reporter proteins from improved alphavirus expression vectors to track replication and dissemination during encephalitic and arthritogenic disease. J. Virol. 2014;88:2035. doi: 10.1128/JVI.02990-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data presented in this study are available in Supplementary Material.