

Abstract
Background
Lead (Pb) exposure is a global health hazard causing a wide range of adverse health outcomes. Yet, the mechanisms of Pb toxicology remain incompletely understood, especially during pregnancy. To uncover biological pathways impacted by Pb exposure, this study investigated serum metabolomic profiles during the third trimester of pregnancy that are associated with blood Pb and bone Pb.
Methods
We used data and specimens from 99 women enrolled in the Programming Research in Obesity, Growth, Environment, and Social Stressors birth cohort in Mexico City. Maternal Pb exposure was measured in whole blood samples from the third trimester of pregnancy and in the tibia and patella bones at 1 month postpartum. Third-trimester serum samples underwent metabolomic analysis; metabolites were identified based on matching to an in-house analytical standard library. A metabolome-wide association study was performed using multiple linear regression models. Class- and pathway-based enrichment analyses were also conducted.
Results
The median (interquartile range) blood Pb concentration was 2.9 (2.6) µg/dL. Median bone Pb, measured in the tibia and patella, were 2.5 (7.3) µg/g and 3.6 (9.5) µg/g, respectively. Of 215 total metabolites identified in serum, 31 were associated with blood Pb (p < 0.05). Class enrichment analysis identified significant overrepresentation of metabolites classified as fatty acids and conjugates, amino acids and peptides, and purines. Tibia and patella Pb were associated with 14 and 8 metabolites, respectively (p < 0.05). Comparing results from bone and blood Pb, glycochenodeoxycholic acid, glycocholic acid, and 1-arachidonoylglycerol were positively associated with blood Pb and tibia Pb, and 7-methylguanine was negatively associated with blood Pb and patella Pb. One metabolite, 5-aminopentanoic acid, was negatively associated with all three Pb measures.
Conclusions
This study identified serum metabolites in pregnant women associated with Pb measured in blood and bone. These findings provide insights on the metabolic profile around Pb exposure in pregnancy and information to guide mechanistic studies of toxicological effects for mothers and children.
Keywords: Pb exposure, Pregnancy, Metabolome, Bone pb, Caffeine, Fatty acids
Background
Lead (Pb) exposure is a persistent global health hazard, with no safe exposure threshold [1]. Pb is a naturally occurring non-essential element, and human industrial practices can promote Pb exposures through the contamination of dust, food, and water [2]. Once ingested, Pb often substitutes for calcium (Ca2+) in the human body and can accumulate in bone, where it is stored long-term and can re-enter the bloodstream during periods of bone remodeling [3]. Pb exposures are linked to a wide array of detrimental health outcomes [2, 4, 5], including neurotoxicity, reduced kidney function, anemia, joint weakness, hypertension, reduced immune function, and reproductive complications.
Children are particularly susceptible to the effects of Pb exposures: for a given Pb exposure, the relative exposure in children is larger by body mass than in adults, and children have higher soft tissue Pb absorption [3, 6, 7]. Further, children have a more permeable blood-brain barrier than adults, making the neurotoxic effects of Pb particularly pronounced [3]. Finally, there are multiple critical neurodevelopmental windows during childhood wherein Pb exposure may lead to life-long adverse neurological effects [8, 9]. Despite these significant health concerns, the mechanisms of Pb toxicity are not fully understood. Novel approaches to investigating Pb metabolism and toxicology may promote new avenues for discovering these mechanisms. Additionally, further examination of Pb exposure during pregnancy is particularly crucial, as exposure during this period can exert harmful effects in both mothers and children.
One powerful approach to uncover the biological impacts of Pb exposure is through metabolomics. Measuring metabolites in biological samples, such as plasma or serum, provides a snapshot of metabolic function, enabling the investigation of biological responses to exposures, including Pb. Unlike other ‘omic technologies (e.g., genomics, transcriptomics, etc.) that capture the potential for phenotypic changes, metabolomics shows the actual state of metabolic functioning. Therefore, metabolomics can reveal the metabolic links between Pb exposure and downstream health outcomes by investigating metabolites and pathways strongly associated with Pb exposure.
Metabolomics analyses of Pb exposures are relatively sparse. The few metabolomics studies of Pb have primarily been conducted in animals, adult males, and highly-exposed populations. In a study investigating the effects of Pb exposure on the gut microbiome and blood metabolome in 8-week-old female mice, Gao et al. found that Pb exposure altered multiple metabolomic pathways including nitrogen metabolism, oxidative stress, vitamin E, and bile acids [10]. A similar dosing study in 8-month-old rats found that Pb exposure reduced levels of butyryl-L-carnitine and ganglioside GD2 (d18:0/20:0) and increased levels of metabolites associated with oxidative stress pathways [11]. In an epidemiologic investigation of the urinary metabolome of highly-exposed participants residing near a Pb acid battery recycling plant, blood Pb was associated with several metabolic pathways, including porphyrin, amino acid, and chlorophyll metabolism, heme biosynthesis, and ATP-binding cassette transporters [12]. Another epidemiologic analysis in the Veterans Administration Normative Aging Study found several blood metabolites associated with blood Pb, including amino acids, lipids, and metabolites involved in oxidative stress and immune pathways [13]. Taken together, these results indicate that biologically-relevant metabolic changes occur with Pb exposure. While some findings are consistent (amino acids, oxidative stress), numerous factors, including the timing, level, and population(s) of Pb exposure, may affect metabolomic responses. Because pregnancy is a uniquely important period of the lifecourse, understanding Pb-associated metabolomic changes in pregnant women–particularly in response to low-level Pb exposures in ranges commonly observed in human populations–is critical to broadly understand the impacts of Pb exposure on human health.
In observational studies, Pb can be measured in a variety of samples–including hair, nails, urine, blood, and bone–with each matrix capturing a different window of exposure. Blood and bone, which are considered the gold standards for Pb measurement, capture short- and long-term exposure, respectively: Pb has a half-life from 1 week to 2 months in blood, depending on age and exposure history, and 5 to 20 years in bone [14]. Previous metabolomics studies primarily measured Pb in blood, which captures relatively recent Pb exposure. Metabolomic analyses in relation to bone Pb will add important information to the literature on the biological impacts of longer-term Pb exposure.
This study investigated serum metabolomic profiles during the third trimester of pregnancy to provide insights on the biological impacts of short- and long-term Pb exposures during this critically important period. We profiled metabolites and measured Pb levels in third-trimester blood samples and collected postpartum bone Pb measures for 99 women in the Programming Research in Obesity, Growth, Environment, and Social Stressors (PROGRESS) cohort based in Mexico City, Mexico.
Methods
Study subjects
The PROGRESS cohort recruited women who were pregnant and receiving prenatal care through the Mexican Social Security System (Instituto Mexicano del Seguro Social – IMSS) between July 2007 and February 2011; in total, there were 948 women with live births. Women were eligible to participate in the study if they were ≥18 years old and <20 weeks of gestation, planned to stay in Mexico City for the next 3 years, had access to a telephone, had no medical history of heart or kidney disease, did not consume alcohol daily, and did not use any steroid or anti-epilepsy medications. Procedures were approved by institutional review boards at the Harvard School of Public Health, Icahn School of Medicine at Mount Sinai, and the Mexican National Institute of Public Health. Women provided written informed consent in Spanish. For the current study, 99 women with available serum samples stored at -80oC from the third trimester of pregnancy from a fasting collection were randomly selected from the full cohort for metabolomics analysis.
Lead (Pb) measurements
All Pb measurements were previously analyzed as described [15]. In short, blood specimens in trace metal-free tubes were analyzed using an Agilent 8800 ICP Triple Quad in MS/MS mode in the trace metals laboratory at the Icahn School of Medicine at Mount Sinai. The limit of detection was <0.2 µg/dL, and the instrument precision was approximately 5% relative standard deviation. At 1 month postpartum, mother’s tibia (cortical bone) and patella (trabecular bone) Pb concentrations were measured using a K-shell X-ray fluorescence (K-XRF) instrument, a noninvasive, low-radiation-dose method of measuring bone lead content [16]. We estimated lead concentration for 30 min for each leg, and the measures were averaged by the inverse of the proportion of the measurement error corresponding to each determination. When bone lead levels are below the detection limit (2 µg/g bone mineral), the instrument can produce negative values. Bone Pb content is thought to provide an indicator of exposure over the span of decades; in particular, tibia measurements reflect longer time spans (>10 years) compared to patella (1-5 years) [17].
Metabolomics analysis
Stored serum samples were thawed on ice and vortexed, and 50-µL aliquots were transferred to a microcentrifuge tube. 150 µL of methanol containing internal standards was added, and the sample vortexed then incubated at -20oC for 30 min. Samples were centrifuged, then the supernatant was dried using a Savant SC250EXP SpeedVac concentrator at 35oC for 90 min and stored at -80oC until analysis. Before liquid chromatography high-resolution mass spectrometry (LC-HRMS) analysis, dried extracts were reconstituted in 60-µL of 80% acetonitrile or 100% methanol for analysis using zwitterionic hydrophilic interaction liquid chromatography (HILIC) or reverse phase (RP) LC connected to HRMS in positive and negative modes, respectively. The metabolite analysis was performed under the same analytical conditions as those described elsewhere [19]. In short, 2 µL sample extracts were analyzed on an UHPLC 1290 Infinity system coupled to a 6545 or 6550 iFunnel Q-TOF MS for HILIC or RP chromatography, respectively. Following the same protocol, matrix blanks (replacing the plasma with water) and a pooled QC that was prepared by combining 10 µL of each sample into a single aliquot were extracted. Samples were analyzed in a randomized order and pooled QCs injected routinely throughout the run. Metabolites were identified based upon in-house database matching considering retention time, accurate mass, and MS/MS matching (when available) with pure standards analyzed under the same conditions, providing the highest identification confidence level (Level 1 or 2) based on Metabolomics Standards Initiative criteria [20, 21]. Peaks were identified and integrated using the Personal Chemical Database Library and Profinder software (Agilent Technologies, Santa Clara, USA). There were 139 metabolites and 115 metabolites detected by HILIC and RP chromatography methods, respectively. Of these, seven metabolites were identified using both methods; only the higher intensity value was retained for analyses. Therefore, the combined dataset had 248 metabolites of interest. Metabolites were removed if they had more than 50% missing values (N = 33). Most remaining metabolites did not have any missing values (192 metabolites); for the 23 metabolites with missing values, missing values were imputed with the minimum value of the metabolite divided by the square root of 2. Imputed data were then log2-transformed to remove heteroscedasticity (unequal variability) in the metabolite intensities.
Covariates
Age and educational attainment were ascertained at enrollment. Since only one woman in the cohort reported smoking tobacco during pregnancy, environmental tobacco smoke exposure during pregnancy was characterized as the report of any tobacco smoker in the home during the second or third trimester of pregnancy. Body mass index (BMI) was determined using height (in meters [m]) and weight (in kilograms [kg]) ascertained at the third-trimester study visit. The season of the last menstrual period was defined according to weather patterns in Mexico City as dry/cold (January-February; November-December), dry/warm (March-April) and rainy (May-October). Since metabolite levels can vary by laboratory batch, models also accounted for the metabolomics batch assignment as a categorical variable.
Statistical analysis
A metabolome-wide association study approach was used to identify metabolites associated with blood and bone Pb measures [22, 23]. Using this approach, multiple linear regression models were constructed for each Pb variable (blood Pb, tibia Pb, or patella Pb) as the main predictor of each log2-transformed metabolite intensity, adjusting for all covariates listed above. For significant findings, the patterns of the associations were examined using bivariate scatterplots and locally estimated scatterplot smoothing regression. For bone Pb variables, negative instrument values were included in analyses since imputation of these values has not been reported to reduce bias [24, 25]; however, sensitivity analyses were also conducted in which negative values were imputed with simulated values between 0 and the detection limit (2 µg/g) generated from a uniform distribution.
To identify significant metabolites while accounting for the high correlation among the metabolite measurements, we implemented the “number of effective tests” adjustment approach previously used in metabolomics studies [13, 26, 27]. Briefly, we used principal components analysis to identify the number of principal components (PCs) that explain 75%, 95%, and 99% of the variance in metabolite intensities, then used these numbers to calculate adjusted p-values (a/m), where a reflects the nominal p-value threshold of 0.05, and m reflects the number of PCs that explain the defined level of variance. Herein, we calculated effective number of test (ENT) thresholds of 75% (ENT75%: 0.05/24 = 0.002083), 95% (ENT95%: 0.05/60 = 0.00083), and 99% (ENT99%: 0.05/82 = 0.000602).
For biological interpretation of the blood Pb results, we input metabolites into MetaboAnalyst [28] to perform class- and pathway-based enrichment analyses. We identified significant metabolite classes and metabolic pathways using the main-class chemical structure and the Small Molecule Pathway Database [29] pathway-based metabolite set libraries, respectively. For co-eluting metabolites, only the first listed metabolite was used as input in enrichment analyses.
Results
The study sample consisted of 99 randomly-selected pregnant women from the PROGRESS cohort with complete data on blood Pb exposure and covariates (Table 1). The median (interquartile range [IQR]) age of these women was 27.7 (6.8) years, and 63% completed a high school education or greater.
Table 1.
Descriptive characteristics for PROGRESS mothers in current study (N = 99)
| Characteristics | Level/Unit | Median (IQR) or N (%) |
|---|---|---|
| Demographics | ||
| Age at recruitment | Years | 27.7 (6.8) |
| BMI in 3rd trimester | kg/m2 | 28.6 (5.0) |
| Maternal Education | < High School | 37 (37%) |
| High School | 38 (38%) | |
| > High School | 24 (24%) | |
| Smoke exposure | Yes | 30 (30%) |
| Season of last menstrual period | Dry/cold | 29 (29%) |
| Dry/warm | 22 (22%) | |
| Rainy | 48 (48%) | |
| Lead (Pb) exposure | ||
| Blood Pb, 3rd trimester | µg/dL | 2.9 (2.6) |
| Tibia Pb, 1 month postpartuma | µg/g | 2.5 (7.3) |
| Patella Pb, 1 month postpartuma | µg/g | 3.6 (9.5) |
aN = 89
The median (IQR) Pb concentration in third-trimester blood samples was 2.9 (2.6) µg/dL (Table 1), with a range of 0.3 to 14.5 µg/dL. These levels are higher than those observed in non-pregnant women of childbearing age measured in NHANES during a similar period [30]. Current guidelines for medical management of Pb-exposed adults recommend that pregnant women avoid occupations and avocational exposures that may result in blood Pb levels >5 µg/dL [31]. In our study, nineteen mothers had blood Pb concentrations above the CDC action level of ≥5 µg/dL for pregnant women; among these women, three had Pb levels of concern (≥10 µg/dL) [32].
Bone patella and tibia Pb levels measured at 1 month postpartum were available for a large subset of the study sample (N = 89). Bone patella and tibia Pb—reflecting long-term Pb exposures from 1 to 5 years and >10 years, respectively—were 3.6 (9.5) µg/g and 2.5 (7.3) µg/g, respectively (Table 1). These bone Pb levels are lower than reported in earlier studies examining bone Pb in Mexican women in the postpartum period [33], suggesting that bone Pb might be decreasing over time in this population, but are slightly higher than those reported in pre-menopausal women in Canada [34]. The Pb levels in patella and tibia were strongly correlated (Pearson’s r = 0.51), while, as expected, bone Pb measures were less strongly correlated with blood Pb (patella and blood Pb, r = 0.34; tibia and blood Pb, r = 0.15) (Fig. 1).
Fig. 1.

Distributions and correlations of Pb in 3rd trimester blood, tibia, and patella
A total of 215 metabolites in third-trimester serum samples were included in our analyses. Of these, 31 metabolites were associated with third-trimester blood Pb at nominal significance (p < 0.05) (Table 2), and 2-hydroxybutyrate and hypoxanthine was negatively associated at ENT95% and ENT75% thresholds, respectively. Class-based enrichment analyses of these 31 metabolites identified significant overrepresentation (Holm-Bonferroni p < 0.05) of metabolites classified as fatty acids and conjugates, amino acids and peptides, and purines (noted with asterisks in Table 2).
Table 2.
Serum metabolites associated with blood Pb in the 3rd trimester
| Metabolite subclass | Compounda | Estimate (SE) | P |
|---|---|---|---|
| Fatty acids and conjugates† | Myristic acid | -0.073 (0.024) | 2.63E-03 |
| Stearic acid | -0.043 (0.014) | 3.02E-03 | |
| Adrenic acid | -0.058 (0.019) | 3.37E-03 | |
| Palmitoleic acid | -0.097 (0.032) | 3.56E-03 | |
| Oleic acid / Elaidic acid / Petroselinic acid | -0.051 (0.020) | 1.25E-02 | |
| Arachidonic acid | -0.040 (0.016) | 1.52E-02 | |
| Lauric acid | -0.058 (0.023) | 1.53E-02 | |
| Dihomo-Gamma-Linolenic acid | -0.040 (0.016) | 1.69E-02 | |
| Margaric acid / Hexadecanol | -0.046 (0.020) | 2.14E-02 | |
| Arachidic acid | -0.036 (0.016) | 2.91E-02 | |
| Palmitic acid | -0.041 (0.014) | 5.14E-03 | |
| Eicosapentaenoic acid | -0.049 (0.024) | 4.40E-02 | |
| Amino acids, peptides, and analogues† | 5-Aminopentanoic acid | -0.137 (0.053) | 1.16E-02 |
| N-Acetylalanine | -0.047 (0.019) | 1.48E-02 | |
| Asparagine | -0.035 (0.016) | 3.05E-02 | |
| N-Acetylmethionine | -0.087 (0.042) | 4.19E-02 | |
| Creatine | -0.064 (0.031) | 4.00E-02 | |
| L-Leucine | -0.039 (0.019) | 4.16E-02 | |
| Purines and purine derivatives† | Hypoxanthine | -0.047 (0.014) | 1.26E-03* |
| 7-Methylguanine | -0.015 (0.006) | 1.70E-02 | |
| Caffeine | 0.193 (0.080) | 1.84E-02 | |
| Bile acids, alcohols and derivatives | Glycocholic acid | 0.079 (0.035) | 2.73E-02 |
| Glycochenodeoxycholic acid | 0.075 (0.038) | 4.93E-02 | |
| Alpha hydroxy acids and derivatives | 2-Hydroxybutyrate | -0.055 (0.016) | 6.50E-04** |
| Dicarboxylic acids and derivatives | Succinate | -0.062 (0.024) | 1.17E-02 |
| Organosulfonic acids and derivatives | Taurine | -0.037 (0.015) | 1.26E-02 |
| Benzoic acids and derivatives | MBP (Mono-n-butyl phthalate) | -0.076 (0.036) | 3.85E-02 |
| Linoleic acids and derivatives | Alpha-linolenic acid (ALA) / Gamma-Linolenic acid (GLA) | -0.079 (0.032) | 1.50E-02 |
| Monoacylglycerols | 1-Arachidonoylglycerol | 0.066 (0.021) | 2.83E-03 |
| Other | N6-(Delta2-Isopentenyl)-Adenine | -0.049 (0.017) | 4.16E-03 |
| N-Acetylneuraminic acid | -0.087 (0.042) | 4.40E-02 |
*Significant at thresholds of ENT75% and **ENT95%;†Significantly-enriched (Holm-Bonferroni p < 0.05) subclass from enrichment analysis in MetaboAnalyst; aMetabolites were identified at MSI Level 1 confidence; compounds that co-eluted and could not be discriminated are indicated with “/” (MSI Level 2 confidence)
Additionally, pathway-based enrichment analysis identified the alpha linolenic acid and linoleic acid metabolic pathway (Holm-Bonferroni p < 0.05; Fig. 2), which contained five metabolites that were negatively associated with blood Pb. We note that linoleic acid and the n-3 fatty acid docosahexaenoic acid (DHA), the latter of which is synthesized in this pathway, were also negatively associated with blood Pb at marginal significance (linoleic acid: estimate [SE], -0.043 [0.022], p = 0.06; DHA: estimate [SE], -0.038 [0.020], p = 0.06).
Fig. 2.
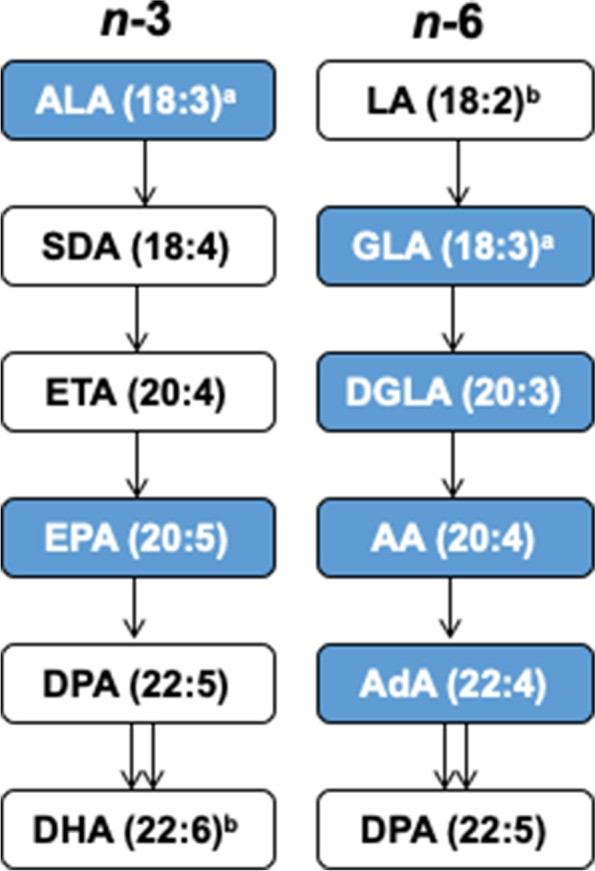
Pathway enrichment analysis of serum metabolites associated with blood Pb identifies the alpha linolenic acid and linoleic acid pathway. Metabolites negatively associated with blood Pb (p < 0.05) levels shown in blue. aALA and GLA co-eluted; bLA and DHA were negatively associated with blood Pb at marginal significance (p = 0.06). ALA: alpha linolenic acid; SDA: stearidonic acid; ETA: eicosatetraenoic acid; EPA: eicosapentaenoic acid; DPA: docosapentaenoic acid; DHA: docosahexaenoic acid; LA: linoleic acid; GLA: gamma-linolenic acid; DGLA: dihomo-gamma-linolenic acid; AA: arachidonic acid; AdA: adrenic acid
We also examined third-trimester serum metabolites associated with patella and tibia Pb, which identified 8 and 14 metabolites, respectively, at p < 0.05 (Table 3); of these, two—5-aminopentanoic acid and L-arginine—were negatively associated with both bone Pb measures. Betaine was positively associated with patella Pb at ENT99%, and 1-linoleoylglycerol was positively associated with tibia Pb at ENT75%. In sensitivity analyses, results did not materially change when negative instrument values for bone Pb were imputed with positive values below the limit of detection (data not shown).
Table 3.
Serum metabolites associated with patella and tibia Pb
| Patella Pb | Tibia Pb | |||
|---|---|---|---|---|
| Compounda | Estimate (SE) | P | Estimate (SE) | P |
| Significant in patella and tibia | ||||
| 5-Aminopentanoic acid | -0.049 (0.015) | 1.82E-03 | -0.049 (0.022) | 2.46E-02 |
| L-Arginine | -0.010 (0.005) | 4.70E-02 | -0.015 (0.007) | 2.74E-02 |
| Significant in patella | ||||
| Betaine | 0.031 (0.009) | 5.31E-04*** | 0.018 (0.014) | 0.19 |
| Biliverdin | 0.017 (0.007) | 1.97E-02 | 0.007 (0.010) | 0.45 |
| Oxoproline | 0.012 (0.005) | 2.05E-02 | 0.004 (0.007) | 0.60 |
| Pyroglutamate | 0.012 (0.005) | 2.09E-02 | 0.004 (0.007) | 0.60 |
| 7-Methylguanine | -0.004 (0.002) | 3.29E-02 | -0.003 (0.003) | 0.34 |
| POV-PC | -0.018 (0.008) | 3.42E-02 | -0.022 (0.011) | 0.05 |
| Significant in tibia | ||||
| 1-Linoleoylglycerol | 0.016 (0.012) | 0.18 | 0.049 (0.015) | 1.50E-03* |
| Glycochenodeoxycholic acid | 0.009 (0.012) | 0.44 | 0.044 (0.016) | 6.83E-03 |
| Docosahexaenoic acid (DHA) | -0.009 (0.006) | 0.16 | -0.022 (0.008) | 9.21E-03 |
| Allantoin | -0.011 (0.007) | 0.13 | -0.026 (0.010) | 9.56E-03 |
| Glycocholic acid | 0.015 (0.011) | 0.18 | 0.037 (0.015) | 1.74E-02 |
| MEP (Mono-ethyl phthalate) | 0.025 (0.027) | 0.36 | 0.085 (0.037) | 2.32E-02 |
| Oleoyl-Glycerol | -0.011 (0.010) | 0.26 | -0.030 (0.013) | 2.79E-02 |
| 1-Arachidonoylglycerol | 0.006 (0.007) | 0.34 | 0.020 (0.009) | 2.89E-02 |
| L-Isoleucine | 0.007 (0.005) | 0.17 | 0.014 (0.007) | 3.66E-02 |
| Indole-3-Carboxylic acid | 0.004 (0.006) | 0.54 | -0.017 (0.008) | 3.82E-02 |
| Erucic acid | -0.006 (0.006) | 0.32 | -0.017 (0.008) | 3.96E-02 |
| 2-Undecanone | 0.008 (0.005) | 0.07 | 0.013 (0.006) | 4.10E-02 |
*Significant at thresholds of ENT75%, **ENT95%, and ***ENT99%; a.Metabolites were identified at MSI Level 1 confidence
Finally, we compared metabolites significantly associated with bone and blood Pb. Among these, 1-arachidonoylglycerol, glycochenodeoxycholic acid, and glycocholic acid was positively associated with blood Pb and tibia Pb, and 7-methylguanine was negatively associated with blood Pb and patella Pb. Further, one metabolite, 5-aminopentanoic acid, was negatively associated with all three measures (Fig. 3).
Fig. 3.

Scatterplots of 5-aminopentanoic acid versus Pb in 3rd trimester blood, tibia, and patella
Discussion
Pb exposure is associated with many deleterious effects, but the complex downstream biological perturbations that result from Pb exposure, particularly at lower levels and during pregnancy, are not well understood. Metabolomics, which provides a global, unbiased measurement of circulating metabolites, enables discovery of metabolites and underlying mechanisms associated with exposures. However, to date, population studies on metabolomics of Pb exposures are sparse and have focused on high-exposure populations [12] or have been performed on aging adult men [13]. Our study on 99 pregnant women is the first to investigate third-trimester serum metabolomics with both blood Pb and bone Pb contents to unravel biological impacts during pregnancy of both acute and long-term Pb exposure.
Lead exposure through inhalation and ingestion results in absorption into the blood, wherein plasma levels reflect the most rapidly exchangeable fraction of Pb in the bloodstream. This fraction is assumed to be associated with the toxic effects of Pb [35]. In addition, around 95% of the total Pb burden is contained within bone, which reflects integrated or cumulative Pb exposure [36]. During rapid bone turnover, such as in pregnancy, Pb from bone may migrate into plasma, influencing blood Pb levels [37]. Indeed, we observed a greater number of metabolites associated with blood Pb (31 metabolites) than bone Pb (8 and 14 metabolites in patella and tibia, respectively) in study participants, coinciding with weak to moderate correlations between the three Pb measures (Pearson’s r = 0.15-0.51). It is likely that blood Pb, as a circulating biomarker of Pb exposure, is better able to induce transient changes in the blood metabolome compared to bone Pb. The third trimester is the period during pregnancy that contains the greatest mobilization of Pb from maternal bone and fastest fetal growth [25, 38]. Since fetuses are directly exposed to the maternal Pb fraction via umbilical cord transfer [39], higher exposure to Pb during the third trimester may coincide with adverse effects on fetal development. Thus, maternal exposure metrics of Pb are a proxy for fetal exposure, and the altered metabolomes that we observed herein may reflect both systemic biological responses (tibia and patella bone Pb) and acute responses (blood Pb) with potential to impact both maternal and fetal health.
Our results show that cumulative Pb exposures measured in bone reflected different metabolite association patterns than acute Pb exposures measured in third-trimester serum (Tables 1 and 2). However, two bile acids, glycochenodeoxycholic acid and glycocholic acid, were positively associated with both blood Pb and tibia Pb. Lead is associated with microbiome dysregulation in animal models [40, 41] and in human studies of adults [42] and children [43], and bile acids regulate the gut microbiome as well as host physiology [44]. Supporting an effect of Pb on the microbiome, though in the opposite direction of our findings, Pb exposure in mice for 13 weeks decreased bile acid concentrations measured in stool as well as altered the abundances of bacterial genera, most of which were reduced upon Pb exposure [10]. However, this relationship is also likely bidirectional, as the gut microbiome interacts with bile acids [45]. In addition, we found a significant negative association between 5-aminopentanoic acid and blood, tibia, and patella Pb. As 5-aminopentanoic acid is a lysine degradation product formed both endogenously and through bacterial catabolism of lysine, these results may further support a non-transient effect of Pb on the human microbiome [46].
We observed that acute Pb exposure (blood Pb) was associated with alterations in saturated and polyunsaturated fatty acids and metabolites in the alpha linoleic acid and linolenic acid pathway. Studies on Pb exposure and lipid alterations in humans are limited. A pilot study of pooled serum samples from occupationally exposed workers to Pb, cadmium, and arsenic found increased levels of very-low-density lipoprotein, decreased low-density lipoprotein, and increased unsaturated fatty acids associated with high heavy metal exposure. These changes in lipid fraction imply disturbance of lipid metabolism [47], though it was not possible to separate the biological effects of individual metals from this mixture. In a single study of 53 postpartum women following delivery, no correlation between fatty acid and Pb levels in maternal blood was observed; notably, that study reported concentrations and ranges of Pb were lower (1.3 [0.6] µg/dL) than in our population of mothers (2.9 [2.6] µg/dL, Table 1).
Though human studies of Pb exposure and lipid dysregulation during pregnancy are limited, several studies in rat dams and pups reported associations consistent with our current study. Pb exposure in lactating dams resulted in significantly decreased total monounsaturated fatty acids and total fatty acid concentrations; in particular, reduced levels of dihomo-gamma- linolenic acid (DGLA), linoleic acid (LA), and eicosapentaenoic acid (EPA) were observed in plasma [48]. Further, rats exposed to Pb during pregnancy and lactation had reduced DHA in milk and mammary gland tissues [49], and in pups, Pb exposure in dam’s milk, in combination with an n-3 insufficient diet, resulted in a reduction in total saturated fatty acids and total n-3 and total n-6 fatty acids in plasma [50]. Interestingly, the relationship may be bidirectional: a lower blood Pb concentration was observed in rats supplemented with a diet rich in n-3 and n-6 polyunsaturated fatty acids [51].
Acute Pb exposure was also associated with decreased 2-hydroxybutyrate, a ketone body derived from α-ketobutyrate (alpha-KB) that is produced by amino acid catabolism (threonine and methionine) and glutathione anabolism (cysteine formation pathway) and is metabolized to propionyl-CoA and carbon dioxide [52]. 2-hydroxybutyrate levels are hypothesized to be linked with lipid oxidation, oxidative stress, and glutathione synthesis [53]. Interestingly, 2-hydroxybutyrate, palmitic acid, palmitoleic acid, stearic, and lactic acid (Table 2) are all linked with gestational diabetes during the second trimester and three months postpartum [54], with 2-hydroxybuytrate indicated as an early biomarker of glucose intolerance [55, 56]. While these metabolites were higher in mothers with gestational diabetes than controls in the previous studies–the opposite direction of our findings–Pb levels may be associated with alterations in similar pathways. Indeed, Pb exposure can induce fasting hyperglycemia and glucose intolerance in rats [57]. Future mechanistic studies that investigate the role of Pb on pathways involving α-ketobutyrate, such as phenylalanine and tyrosine metabolism, branched chain amino acids metabolism, and homocysteine metabolism during pregnancy are encouraged.
Organic environmental exposures are linked with blood Pb levels in pregnant women, including caffeine [58]. Coffee, as well as tea and coca, contain high levels of Pb [59], possibly due to use of ceramic mugs containing Pb-based glaze [60] or contaminated water used in the coffee-making process and the propensity for Pb absorption into organic matter [61]. We observed that serum caffeine and 1-methyluric acid (a major metabolite of caffeine) were positively associated with blood Pb levels (the latter at p < 0.10 significance), but not bone Pb levels. The association between third-trimester blood Pb and caffeine most likely represents acute exposures since caffeine has an estimated half-life of up to 11 h during the last weeks of pregnancy [62]. Therefore, caffeine, which has been linked with both beneficial effects in adults and adverse fetal health outcomes [63], may be a particularly important Pb co-exposure to consider in future studies.
Betaine was most strongly associated with patella Pb, but also reached marginal significance with blood Pb (p = 0.07). Betaine is a vital methyl group donor in transmethylation reactions, as part of one-carbon metabolism via the methionine cycle. Since the availability of methyl groups influences methylation [64], betaine may support a mechanistic premise for the role of one-carbon metabolism in observed associations between blood Pb levels and DNA methylation in highly-exposed populations [65], maternal bone Pb levels and DNA methylation in cord blood [66], and in animal exposure models [67]. Interestingly, increased betaine levels were observed in the urine of Pb-exposed workers from a Pb-acid battery recycling site [12], suggesting that increased circulating betaine may be a biomarker of chronic or high Pb exposure levels. Betaine metabolism is also linked to the gut microbiome, specifically probiotic bacterial taxa associated with obesity prevention (Akkermansia, Bifidobacterium, and Lactobacillus) [68, 69]. The association between betaine and Pb could be due to Pb-related gut dysbiosis, potentially reducing bacteria that metabolize plant-based molecules.
Our discovery study had several limitations. First, the study included a modest sample size of 99 women in their third trimester of pregnancy from the PROGRESS cohort. While this sample captured an important pregnancy window for Pb exposure, mothers solely resided in the geographic region of Mexico City, which may limit generalizability to other populations. Second, maternal blood was sampled only once during the third trimester and may not reflect fluctuations in Pb levels or metabolite profiles. While we also used bone Pb measurements to investigate associations with cumulative Pb exposure, we cannot rule out that results were affected by sampling time. Third, this study was limited to the investigation of the main effect of Pb, even though environmental exposures to organics, nutritional compounds, and other toxic metals occur simultaneously, with potential synergistic or antagonistic effects on maternal and fetal biology. Finally, given that our study was cross-sectional, causality cannot be addressed.
Conclusions
This discovery study identified dysregulated metabolite profiles during the third trimester of pregnancy that were associated with blood Pb and bone Pb levels. Altered 2-hydroxybutyrate and metabolites in the alpha linoleic and linolenic acid pathway suggest potential mechanisms linking short-term Pb exposure to adverse maternal health, such as glucose intolerance and gestational diabetes, and a potential effect of Pb on altered gut microbiome and microbial metabolism. Further, Pb exposure levels measured in bone were associated with increased betaine, a methyl donor involved in methylation processes. Together, these results point to several possible pathways in which short-term and cumulative Pb exposure may affect maternal and fetal health. Replication studies during pregnancy that utilize larger sample numbers are needed to validate findings. Additional studies that include both untargeted and targeted metabolomics panels, as well as investigate longitudinal metabolomic changes and exposure mixtures, are encouraged.
Acknowledgements
We thank the ABC (American British Cowdray Medical Center) in Mexico for providing research facilities and all of the PROGRESS staff and study participants.
Abbreviations
- AdA
adrenic acid
- ALA
alpha-linolenic acid
- AA
arachidonic acid
- BMI
body mass index
- DGLA
dihomo-gamma-linolenic acid
- DHA
docosahexaenoic acid
- DPA
docosapentaenoic acid
- ENT
effective number of tests
- ETA
eicosatetraenoic acid
- EPA
eicosapentaenoic acid
- GLA
gamma-linolenic acid
- IQR
interquartile range
- Pb
lead
- LA
linoleic acid
- LC-HRMS
liquid chromatography high-resolution mass spectrometry
- Instituto Mexicano del Seguro Social – IMSS
Mexican Social Security System
- ‘pooled QC’
pooled quality control sample
- PCs
principal components
- PROGRESS
Programming Research in Obesity, Growth, Environment, and Social Stressors
- RP
reverse phase
- SDA
stearidonic acid
Authors’ contributions
MN and LP contributed to the conception and design of the work, data interpretation, and drafting and revision of the manuscript. SE contributed to the drafting and revision of the manuscript. AJ contributed to the data analysis and revision of the manuscript. GD and CA contributed to the sample analysis, data acquisition, and revision of the manuscript. MT-R and RW contributed to the conception and design of the work, data acquisition, and revision of the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by the National Institutes of Health (R01 ES014930, R01 ES013744, P30 ES023515, U2CES030859, T32 HD049311, R01 ES031117, R21 ES030882) and the National Institute of Public Health/Ministry of Health of Mexico.
Availability of data and materials
The data that support the findings of this study are available from the PROGRESS study team (PI: Robert O. Wright, robert.wright@mssm.edu), but restrictions apply to the availability of these data. Data are available upon reasonable request.
Declarations
Ethics approval and consent to participate
Procedures were approved by institutional review boards at the Harvard School of Public Health, Icahn School of Medicine at Mount Sinai, and the Mexican National Institute of Public Health. Women provided written informed consent in Spanish.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Betts Kellyn S. CDC Updates Guidelines for Children’s Lead Exposure. Environ Health Perspect. 2012;120(7):a268–a268. [DOI] [PMC free article] [PubMed]
- 2.Abadin H, Ashizawa A, Stevens Y-W, Llados F, Diamond G, Sage G, et al. Toxicological Profile for Lead [Internet]. Atlanta (GA): Agency for Toxic Substances and Disease Registry (US); 2007 [cited 2021 Apr 12]. (Agency for Toxic Substances and Disease Registry (ATSDR) Toxicological Profiles). Available from: http://www.ncbi.nlm.nih.gov/books/NBK158766/ [PubMed]
- 3.Lidsky TI, Schneider JS. Lead neurotoxicity in children: basic mechanisms and clinical correlates. Brain. 2003;126(1):5–19. [DOI] [PubMed]
- 4.Ramírez Ortega D, González Esquivel DF, Blanco Ayala T, Pineda B, Gómez Manzo S, Marcial Quino J, et al. Cognitive Impairment Induced by Lead Exposure during Lifespan: Mechanisms of Lead Neurotoxicity. Toxics. 2021;9(2):23. [DOI] [PMC free article] [PubMed]
- 5.Zeng X, Huo X, Xu X, Liu D, Wu W. E-waste lead exposure and children’s health in China. Sci Total Environ. 2020;734:139286. [DOI] [PubMed]
- 6.Wani AL, Ara A, Usmani JA. Lead toxicity: a review. Interdiscip Toxicol. 2015;8(2):55–64. doi: 10.1515/intox-2015-0009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tong S, von Schirnding YE, Prapamontol T. Environmental lead exposure: a public health problem of global dimensions. Bull World Health Organ. 2000;78(9):1068–77. [PMC free article] [PubMed] [Google Scholar]
- 8.Dórea JG. Environmental exposure to low-level lead (Pb) co-occurring with other neurotoxicants in early life and neurodevelopment of children. Environ Res. 2019;177:108641. doi: 10.1016/j.envres.2019.108641. [DOI] [PubMed] [Google Scholar]
- 9.Levin-Schwartz Y, Gennings C, Schnaas L, Del Carmen Hernández Chávez M, Bellinger DC, Téllez-Rojo MM, et al. Time-varying associations between prenatal metal mixtures and rapid visual processing in children. Environ Health Glob Access Sci Source. 2019;18(1):92. [DOI] [PMC free article] [PubMed]
- 10.Gao B, Chi L, Mahbub R, Bian X, Tu P, Ru H, et al. Multi-Omics Reveals that Lead Exposure Disturbs Gut Microbiome Development, Key Metabolites, and Metabolic Pathways. Chem Res Toxicol. 2017;30(4):996–1005. [DOI] [PMC free article] [PubMed]
- 11.Mani MS, Joshi MB, Shetty RR, DSouza VL, Swathi M, Kabekkodu SP, et al. Lead exposure induces metabolic reprogramming in rat models. Toxicol Lett. 2020;335:11–27. [DOI] [PubMed]
- 12.Eguchi A, Nomiyama K, Sakurai K, Kim Trang PT, Viet PH, Takahashi S, et al. Alterations in urinary metabolomic profiles due to lead exposure from a lead–acid battery recycling site. Environ Pollut. 2018;242:98–105. [DOI] [PubMed]
- 13.Kelly RS, Bayne H, Spiro A, Vokonas P, Sparrow D, Weiss ST, et al. Metabolomic signatures of lead exposure in the VA Normative Aging Study. Environ Res. 2020;190:110022. doi: 10.1016/j.envres.2020.110022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Agency for Toxic Substances and Disease Registry (ASTDR). Toxicological profile for lead [Internet]. Atlanta (GA): U.S. Department of Health and Human Services; 2020 Aug [cited 2021 Apr 12]. Available from: https://stacks.cdc.gov/view/cdc/95222
- 15.Renzetti S, Just AC, Burris HH, Oken E, Amarasiriwardena C, Svensson K, et al. The association of lead exposure during pregnancy and childhood anthropometry in the Mexican PROGRESS cohort. Environ Res. 2017 ;152:226–32. [DOI] [PMC free article] [PubMed]
- 16.Hu H, Milder FL, Burger DE. The use of K X-ray fluorescence for measuring lead burden in epidemiological studies: high and low lead burdens and measurement uncertainty. Environ Health Perspect. 1991;94:107–10. doi: 10.1289/ehp.94-1567946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hu H, Rabinowitz M, Smith D. Bone lead as a biological marker in epidemiologic studies of chronic toxicity: conceptual paradigms. Environ Health Perspect. 1998;106(1):1–8. doi: 10.1289/ehp.981061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hu J, Lesseur C, Miao Y, Manservisi F, Panzacchi S, Mandrioli D, et al. Low-dose exposure of glyphosate-based herbicides disrupt the urine metabolome and its interaction with gut microbiota. Sci Rep. 2021;11(1):3265. [DOI] [PMC free article] [PubMed]
- 19.Yu M, Dolios G, Yong-Gonzalez V, Björkqvist O, Colicino E, Halfvarson J, et al. Untargeted metabolomics profiling and hemoglobin normalization for archived newborn dried blood spots from a refrigerated biorepository. J Pharm Biomed Anal. 2020;191:113574. [DOI] [PMC free article] [PubMed]
- 20.Salek RM, Steinbeck C, Viant MR, Goodacre R, Dunn WB. The role of reporting standards for metabolite annotation and identification in metabolomic studies. GigaScience. 2013;2(1):13. [DOI] [PMC free article] [PubMed]
- 21.Sumner LW, Amberg A, Barrett D, Beale MH, Beger R, Daykin CA, et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI) Metabolomics Off J Metabolomic Soc. 2007;3(3):211–21. doi: 10.1007/s11306-007-0082-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chadeau-Hyam M, Ebbels TMD, Brown IJ, Chan Q, Stamler J, Huang CC, et al. Metabolic Profiling And The Metabolome-Wide Association Study: Significance Level For Biomarker Identification. J Proteome Res. 2010 ;9(9):4620–7. [DOI] [PMC free article] [PubMed]
- 23.Nicholson JK, Holmes E, Elliott P. The metabolome-wide association study: a new look at human disease risk factors. J Proteome Res. 2008;7(9):3637–8. [DOI] [PubMed]
- 24.Stroustrup A, Hsu H-H, Svensson K, Schnaas L, Cantoral A, Solano González M, et al. Toddler temperament and prenatal exposure to lead and maternal depression. Environ Health Glob Access Sci Source. 2016;15(1):71. [DOI] [PMC free article] [PubMed]
- 25.Téllez-Rojo MM, Hernández-Avila M, Lamadrid-Figueroa H, Smith D, Hernández-Cadena L, Mercado A, et al. Impact of Bone Lead and Bone Resorption on Plasma and Whole Blood Lead Levels during Pregnancy. Am J Epidemiol. 2004;160(7):668–78. [DOI] [PubMed]
- 26.Nyholt DR. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. Am J Hum Genet. 2004;74(4):765–9. doi: 10.1086/383251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Li M-X, Yeung JMY, Cherny SS, Sham PC. Evaluating the effective numbers of independent tests and significant p-value thresholds in commercial genotyping arrays and public imputation reference datasets. Hum Genet. 2012;131(5):747–56. doi: 10.1007/s00439-011-1118-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chong J, Soufan O, Li C, Caraus I, Li S, Bourque G, et al. MetaboAnalyst 4.0: towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018;46(W1):W486–94. [DOI] [PMC free article] [PubMed]
- 29.Jewison T, Su Y, Disfany FM, Liang Y, Knox C, Maciejewski A, et al. SMPDB 2.0: big improvements to the Small Molecule Pathway Database. Nucleic Acids Res. 2014 Jan;42(Database issue):D478-484. [DOI] [PMC free article] [PubMed]
- 30.Ettinger Adrienne S., Egan Kathryn B., Homa David M., Brown Mary Jean. Blood Lead Levels in U.S. Women of Childbearing Age, 1976–2016. Environ Health Perspect. 128(1):017012. [DOI] [PMC free article] [PubMed]
- 31.Kosnett MJ, Wedeen RP, Rothenberg SJ, Hipkins KL, Materna BL, Schwartz BS, et al. Recommendations for Medical Management of Adult Lead Exposure. Environ Health Perspect. 2007;115(3):463–71. doi: 10.1289/ehp.9784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.CDC - Adult Blood Lead Epidemiology and Surveillance (ABLES): Reference Blood Lead Levels (BLLs) for Adults in the U. S.: NIOSH Workplace Safety and Health Topic [Internet]. 2021 [cited 2021 May 21]. Available from: https://www.cdc.gov/niosh/topics/ables/ReferenceBloodLevelsforAdults.html
- 33.Liu Y, Téllez-Rojo MM, Sánchez BN, Zhang Z, Afeiche MC, Mercado-García A, et al. Early lead exposure and pubertal development in a Mexico City population. Environ Int. 2019;125:445–51. doi: 10.1016/j.envint.2019.02.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.McNeill FE, Fisher M, Chettle DR, Inskip M, Healey N, Bray R, et al. The decrease in population bone lead levels in Canada between 1993 and 2010 as assessed by in vivo XRF. Physiol Meas. 2017;39(1):015005. [DOI] [PubMed]
- 35.Schütz A, Bergdahl IA, Ekholm A, Skerfving S. Measurement by ICP-MS of lead in plasma and whole blood of lead workers and controls. Occup Environ Med. 1996;53(11):736–40. doi: 10.1136/oem.53.11.736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Barry PSI, Mossman DB. Lead concentrations in human tissues. Br J Ind Med. 1970;27(4):339–51. doi: 10.1136/oem.27.4.339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hernández-Avila M, Smith D, Meneses F, Sanin LH, Hu H. The influence of bone and blood lead on plasma lead levels in environmentally exposed adults. Environ Health Perspect. 1998106(8):473–7. [DOI] [PMC free article] [PubMed]
- 38.Ettinger AS, Lamadrid-Figueroa H, Mercado-García A, Kordas K, Wood RJ, Peterson KE, et al. Effect of calcium supplementation on bone resorption in pregnancy and the early postpartum: a randomized controlled trial in Mexican Women. Nutr J. 2014;13(1):116. doi: 10.1186/1475-2891-13-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Amaral JH, Rezende VB, Quintana SM, Gerlach RF, Barbosa F, Tanus-Santos JE. The relationship between blood and serum lead levels in peripartum women and their respective umbilical cords. Basic Clin Pharmacol Toxicol. 2010;107(6):971–5. doi: 10.1111/j.1742-7843.2010.00616.x. [DOI] [PubMed] [Google Scholar]
- 40.Xia J, Lu L, Jin C, Wang S, Zhou J, Ni Y, et al. Effects of short term lead exposure on gut microbiota and hepatic metabolism in adult zebrafish. Comp Biochem Physiol Part C Toxicol Pharmacol. 2018;209:1–8. [DOI] [PubMed]
- 41.Wu J, Wen XW, Faulk C, Boehnke K, Zhang H, Dolinoy DC, et al. Perinatal Lead Exposure Alters Gut Microbiota Composition and Results in Sex-specific Bodyweight Increases in Adult Mice. Toxicol Sci Off J Soc Toxicol. 2016;151(2):324–33. doi: 10.1093/toxsci/kfw046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Eggers S, Safdar N, Sethi AK, Suen G, Peppard PE, Kates AE, et al. Urinary lead concentration and composition of the adult gut microbiota in a cross-sectional population-based sample. Environ Int. 2019;133(Pt A):105122. [DOI] [PMC free article] [PubMed]
- 43.Sitarik AR, Arora M, Austin C, Bielak LF, Eggers S, Johnson CC, et al. Fetal and early postnatal lead exposure measured in teeth associates with infant gut microbiota. Environ Int. 2020;144:106062. [DOI] [PMC free article] [PubMed]
- 44.Ridlon JM, Kang DJ, Hylemon PB, Bajaj JS. Bile Acids and the Gut Microbiome. Curr Opin Gastroenterol. 2014;30(3):332–8. doi: 10.1097/MOG.0000000000000057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Winston JA, Theriot CM. Diversification of host bile acids by members of the gut microbiota. Gut Microbes. 2019;11(2):158–71. [DOI] [PMC free article] [PubMed]
- 46.Knorr S, Sinn M, Galetskiy D, Williams RM, Wang C, Müller N, et al. Widespread bacterial lysine degradation proceeding via glutarate and L-2-hydroxyglutarate. Nat Commun. 2018;9(1):5071. [DOI] [PMC free article] [PubMed]
- 47.Dudka I, Kossowska B, Senhadri H, Latajka R, Hajek J, Andrzejak R, et al. Metabonomic analysis of serum of workers occupationally exposed to arsenic, cadmium and lead for biomarker research: A preliminary study. Environ Int. 2014;68:71–81. [DOI] [PubMed]
- 48.Sun-Young L, Loewke J, Doherty JD, Salem N. Preferential Effect of Lead Exposure During Lactation on Non-Essential Fatty Acids in Maternal Organs. Lipids. 2005;40(7):685–93. doi: 10.1007/s11745-005-1431-z. [DOI] [PubMed] [Google Scholar]
- 49.Hossain S, Hussain J, Bhowmick S, Sarkar M, Basunia M, Al Mamun A, et al. Docosahexaenoic Acid (DHA, C22:6, ω-3) Composition of Milk and Mammary Gland Tissues of Lactating Mother Rats Is Severely Affected by Lead (Pb) Exposure. Biol Trace Elem Res. 2020;195(2):525–34. [DOI] [PubMed]
- 50.Lim S-Y, Doherty JD, Salem N Jr. Lead Exposure and (n-3) Fatty Acid Deficiency during Rat Neonatal Development Alter Liver, Plasma, and Brain Polyunsaturated Fatty Acid Composition. J Nutr. 2005;135(5):1027–33. [DOI] [PubMed]
- 51.Baranowska-Bosiacka I, Kosińska I, Jamioł D, Gutowska I, Prokopowicz A, Rębacz-Maron E, et al. Environmental Lead (Pb) Exposure Versus Fatty Acid Content in Blood and Milk of the Mother and in the Blood of Newborn Children. Biol Trace Elem Res. 2016;170:279–87. doi: 10.1007/s12011-015-0482-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Landaas S. The formation of 2-hydroxybutyric acid in experimental animals. Clin Chim Acta. 1975 Jan 6;58(1):23–32. [DOI] [PubMed]
- 53.Sekhar RV, McKay SV, Patel SG, Guthikonda AP, Reddy VT, Balasubramanyam A, et al. Glutathione Synthesis Is Diminished in Patients With Uncontrolled Diabetes and Restored by Dietary Supplementation With Cysteine and Glycine. Diabetes Care. 2011;34(1):162–7. doi: 10.2337/dc10-1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Dudzik D, Zorawski M, Skotnicki M, Zarzycki W, García A, Angulo S, et al. GC-MS based Gestational Diabetes Mellitus longitudinal study: Identification of 2-and 3-hydroxybutyrate as potential prognostic biomarkers. J Pharm Biomed Anal. 2017;144:90–8. [DOI] [PubMed]
- 55.Ferrannini E, Natali A, Camastra S, Nannipieri M, Mari A, Adam K-P, et al. Early Metabolic Markers of the Development of Dysglycemia and Type 2 Diabetes and Their Physiological Significance. Diabetes. 2013 ;62(5):1730–7. [DOI] [PMC free article] [PubMed]
- 56.Gall WE, Beebe K, Lawton KA, Adam K-P, Mitchell MW, Nakhle PJ, et al. alpha-hydroxybutyrate is an early biomarker of insulin resistance and glucose intolerance in a nondiabetic population. PloS One. 2010;5(5):e10883. [DOI] [PMC free article] [PubMed]
- 57.Tyrrell JB, Hafida S, Stemmer P, Adhami A, Leff T. Lead (Pb) exposure promotes diabetes in obese rodents. J Trace Elem Med Biol. 2017;39:221–6. [DOI] [PubMed]
- 58.Taylor CM, Golding J, Hibbeln J, Emond AM. Environmental Factors Predicting Blood Lead Levels in Pregnant Women in the UK: The ALSPAC Study. PLOS ONE. 2013;8(9):e72371. [DOI] [PMC free article] [PubMed]
- 59.Scientific Opinion on Lead in Food EFSA J. 2010;8(4):1570. [Google Scholar]
- 60.Pantic I, Tamayo-Ortiz M, Rosa-Parra A, Bautista-Arredondo L, Wright RO, Peterson KE, et al. Children’s Blood Lead Concentrations from 1988 to 2015 in Mexico City: The Contribution of Lead in Air and Traditional Lead-Glazed Ceramics. Int J Environ Res Public Health. 2018;15(10). [DOI] [PMC free article] [PubMed]
- 61.Siu Y, Mejia G, Mejia-Saavedra J, Pohlan J, Sokolov M. Heavy Metals in Wet Method Coffee Processing Wastewater in Soconusco, Chiapas, Mexico. Bull Environ Contam Toxicol. 2007;78(5):400–4. [DOI] [PubMed]
- 62.Knutti R, Rothweiler H, Schlatter C. The effect of pregnancy on the pharmacokinetics of caffeine. Arch Toxicol Suppl Arch Toxikol Suppl. 1982;5:187–92. doi: 10.1007/978-3-642-68511-8_33. [DOI] [PubMed] [Google Scholar]
- 63.Poole R, Kennedy OJ, Roderick P, Fallowfield JA, Hayes PC, Parkes J. Coffee consumption and health: umbrella review of meta-analyses of multiple health outcomes. BMJ. 2017;359:j5024. [DOI] [PMC free article] [PubMed]
- 64.Zeisel SH. Choline, Other Methyl-Donors and Epigenetics. Nutrients [Internet]. 2017 Apr 29 [cited 2021 Mar 29];9(5). Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5452175/ [DOI] [PMC free article] [PubMed]
- 65.Yohannes YB, Nakayama SMM, Yabe J, Nakata H, Toyomaki H, Kataba A, et al. Blood lead levels and aberrant DNA methylation of the ALAD and p16 gene promoters in children exposed to environmental-lead. Environ Res. 2020;188:109759. [DOI] [PubMed]
- 66.Pilsner JR, Hu H, Ettinger A, Sánchez BN, Wright RO, Cantonwine D, et al. Influence of Prenatal Lead Exposure on Genomic Methylation of Cord Blood DNA. Environ Health Perspect. 2009;117(9):1466–71. doi: 10.1289/ehp.0800497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Basha MR, Wei W, Bakheet SA, Benitez N, Siddiqi HK, Ge Y-W, et al. The fetal basis of amyloidogenesis: exposure to lead and latent overexpression of amyloid precursor protein and beta-amyloid in the aging brain. J Neurosci Off J Soc Neurosci. 2005;25(4):823–9. [DOI] [PMC free article] [PubMed]
- 68.Koistinen VM, Kärkkäinen O, Borewicz K, Zarei I, Jokkala J, Micard V, et al. Contribution of gut microbiota to metabolism of dietary glycine betaine in mice and in vitro colonic fermentation. Microbiome. 2019;7(1):103. [DOI] [PMC free article] [PubMed]
- 69.Du J, Zhang P, Luo J, Shen L, Zhang S, Gu H, et al. Dietary betaine prevents obesity through gut microbiota-drived microRNA-378a family. Gut Microbes. 2021;13(1):1–19. doi: 10.1080/19490976.2020.1854641. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data that support the findings of this study are available from the PROGRESS study team (PI: Robert O. Wright, robert.wright@mssm.edu), but restrictions apply to the availability of these data. Data are available upon reasonable request.