

Abstract
Conventions form an essential part of human social and cultural behaviour and may also be important to other animal societies. Yet, despite the wealth of evidence that has accumulated for culture in non-human animals, we know surprisingly little about non-human conventions beyond a few rare examples. We follow the literature in behavioural ecology and evolution and define conventions as systematic behaviours that solve a coordination problem in which two or more individuals need to display complementary behaviour to obtain a mutually beneficial outcome. We start by discussing the literature on conventions in non-human primates from this perspective and conclude that all the ingredients for conventions to emerge are present and therefore that they ought to be more frequently observed. We then probe the emergence of conventions by using a unique novel experimental system in which pairs of Guinea baboons (Papio papio) can voluntarily participate together in touchscreen-based cognitive testing and we show that conventions readily emerge in our experimental set-up and that they share three fundamental properties of human conventions (arbitrariness, stability and efficiency). These results question the idea that observational learning, and imitation in particular, is necessary to establish conventions; they suggest that positive reinforcement is enough.
This article is part of a discussion meeting issue ‘The emergence of collective knowledge and cumulative culture in animals, humans and machines’.
Keywords: primate social cognition, social learning, animal culture, language evolution, imitation
1. Introduction
Conventions have often been overlooked in the literature on social learning despite the fact that they form an essential part of human social and cultural behaviour [1] and may also be important to other animal societies. Only a few rare examples of non-human conventions have been documented (see [2] for a discussion of non-human conventions), despite the existence of evolutionary precursors [3]. The first part of this article discusses current evidence of conventions in non-human primates and concludes that conventions ought to be more frequently observed. The scarcity of convention has been explained by the limited social learning capacities of non-human primates, especially regarding imitation. However, in the second part of the article, we present a set of experiments showing the emergence of convention in a non-human primate using a unique novel experimental system. These experiments show that Guinea baboons (Papio papio) can readily develop conventions that share at least three properties of human conventions (arbitrariness, stability and efficiency) through simple reinforcement learning. We start by clarifying the meaning and properties of convention.
Intuitively, conventions are rule-like behaviour such as shaking hands as a greeting. Beyond this intuitive understanding however, conventions have proven difficult to define and have been much debated. Here, we will follow a ‘Lewisian’ approach [4] and define conventions as systematic behaviours that solve a coordination problem in which two or more individuals need to display complementary behaviour to obtain a mutually beneficial outcome.
A classic example of convention, used by Hume [5] and taken up by Lewis, is that of two rowers on a boat. If both rowers are synchronized, the boat will move forward smoothly, whereas if they row out of sync, it will behave erratically. In Hume's (then Lewis's) use, this example illustrates the main properties of convention: (1) rowers do not keep promises to act in concert; (2) they may express their shared interest explicitly, but not necessarily; (3) to row in rhythm, there must be a gradual installation of synchronous behaviour; (4) they may know that what is being set up between them is a convention, but not know how to describe it verbally (we start at the 90° angle and give a stroke every 2 s); and (5) they have adopted a rhythm that they keep because it works, but there are alternatives. We discuss some of these characteristics in more detail below.
Individuals can find ad hoc solutions to coordination problems when they arise, as when finding seating arrangements around the table, for instance. However, when coordination problems happen regularly and/or impose a high cost for a failure to coordinate, individuals may benefit from establishing a convention [6]. Note that compared to other types of positive interactions such as helping (one individual benefits at the expense of another) or cooperation (individuals can benefit more by being selfish), individuals mutually benefit from coordination: it is in their own best interest and in the interest of the partner(s) to coordinate. Therefore, there is no conflict of interest between individuals when solving a coordination problem or when establishing a convention. Rather, conventions are important because they help stabilize interactions by creating mutual expectations between individuals. Someone picking up his/her car in France expects other individuals to drive on the right side of the road and so adjusts his/her behaviour accordingly. These expectations need not be explicit nor mutually acknowledged for a convention to exist [7]. In fact, sometimes we are barely aware that a convention exists: you might lunch at a certain time simply because you expect other people to lunch around that time. According to Hawkins' theoretical model [8], three processes allow the emergence and maintenance of a convention: the population level (social network management and cultural transmission), the dyadic level (coordination and social interaction) and the individual level (executive functions and expectations). A convention is not necessarily created as a result of a problem encountered by the whole population at one point. If enough dyads in the population repeatedly encounter a coordination problem and each individual belongs to several dyads, a population-wide convention can emerge.
Of course, a paradigmatic example of convention in humans is language because to communicate individuals need to coordinate on word-meaning relationships. Studies have shown how humans can develop new conventions to communicate during the natural emergence of new languages such as pidgin (which occurs when individuals with no common language need to communicate, [9]) or sign language (for instance, the Nicaraguan sign language [10]) and these studies have been complemented by experiments and models showing how conventions develop during the emergence of artificial communication systems [11,12]. Conventions are traditionally distinguished from habits and norms. Habits are patterns of individual behaviour that do not (strongly) depend on other individuals, but they can be shared and cultural: taking a shower versus taking a bath for instance. In contrast, norms may be seen as solving coordination problems when the interest of individuals are not aligned. They are associated to a sense of ‘oughtness’ and punishment that conventions and habits lack, e.g. ‘crossing at a red light’ (e.g. [13]). Clearly, given the diversity of human social behaviour, there are no strict boundaries between habits, conventions and norms. What was once a habit, ‘spitting in the street’, can become a norm, ‘do not spit’ [14]. Conventions, e.g. ‘crossing at pedestrian crossings' can be felt to some like norms that ought to be followed at all times and by others as mere conventions. There are also habits, ‘eating cheese before dessert’, that, although not a coordination problem per se, are easier to deal with when everyone follows the same rules (e.g. to have a meal together). Therefore, conventions are not an ‘all or nothing’ kind of thing but a ‘more or less’ one: behaviours are more or less conventional depending on how closely they match the prototypical example of convention.
In the framework of game theory [15], a coordination problem is a problem with multiple Nash equilibriums (a strategy is a Nash equilibrium when no player can do better by changing his strategy alone; [16]) and a convention is a Nash equilibrium to such problem (see [4] for the original discussion and [2] for a more detailed discussion in the context of animal behaviour). The Nash equilibrium, i.e. the convention, is governed by initial conditions and dynamics (influenced by the way individuals choose to act and the environment). Note that conventions require multiple equilibriums because if there are no alternative options, there would be no coordination problem to solve in the first place and two individuals would necessarily converge on the unique solution. However, different equilibriums need not be equally rewarding: one may lead to higher rewards for instance (called payoff-dominant strategies) and some equilibriums may be more rewarding than others when individuals deviate from the equilibrium (risk-dominant equilibrium). Note that in the framework of game theory a convention is ‘arbitrary’ because there are multiple Nash equilibriums to a coordination problem, so a population could have ended up using another convention. It does not mean that individuals are arbitrarily choosing one behaviour over another; they may strategically choose a payoff-dominant strategy for instance. Importantly, compared to their one-shot counterpart, repeated economic games can help create conventions because with repetition multiple equilibriums often appear [17].
To summarise, when individuals regularly face coordination problems, they may develop conventions as solutions to these problems. Conventions are systematic behaviours that are mutually beneficial and provide stable expectations about other individuals. Thus conventions, like norms, help individuals stabilize their social environment and navigate their social world. It is important to note that conventions do not require individuals to do the same action (one could pull while the other pushes), they can concern as few as two individuals, and they do not necessarily lead to the optimal solution (other more profitable equilibria may exist). Conventions are usually associated with three important properties [6]:
-
1.
Arbitrariness: at least one alternative exists and would be equally acceptable if everyone coordinated on it.
-
2.
Efficiency: individuals benefit from establishing a convention.
-
3.
Stability: over multiple games behaviour converges towards an attractor.
Conventions are the natural outcome of individuals repeatedly facing coordination problems together and are therefore expected in group-living animals and especially in tightly bonded non-human primates (for instance, see [18] for a study of chimpanzees solving a coordination task using touchscreens). Recently, Stephens et al. [19] reviewed evidence of conventions in non-human animals using a game-theoretic perspective but did not discuss primates, and Kappeler et al. [3] provided an important analysis of the evolutionary behavioural, emotional and cognitive precursors of norms and conventions in non-human primates. Here, we complement this work by discussing specific cases of non-human primate behaviour in the context just discussed that is graded and grounded in a game-theoretic perspective.
2. Conventions in non-human primates?
Susan Perry and colleagues' description of social conventions in white-faced capuchin monkeys (Cebus capucinus) is one of the best description of conventions we have in non-human primates [20]. Using 13 years of data collection in their field sites in Costa Rica, Perry et al. describe the fashion-like emergence of new behaviour. During hand-sniffing for instance (first reported by [21]) two individuals are resting together and one individual holds the hand or foot of another over its own face. These behaviours are similar in many respects to human conventions such as shaking hands: they require coordination between pairs of individuals because individuals need to be together and to perform complementary behaviour, they emerge in some groups and not others, and can spread to a large proportion of individuals within the group. They can also stay stable for a long time: in one group hand-sniffing was observed over seven years before disappearing. Finally, these behaviours could have a function close to our handshakes because they may strengthen the bonds between individuals. These behaviours therefore appear to have all the properties of conventions. Similarly, the hand-clasp grooming [22–24] and social-scratch grooming of chimpanzees [25] also appear to meet the requirements of conventions: they are stable behaviour that solve coordination problems for which alternative behaviour exists. In addition, the social structure may vary between groups of the same species and some behaviours may be considered conventional under the definition proposed here. For example, van de Waal [26] showed that neighbouring groups of vervet monkeys differ in the structure of their social networks as well as how they handle conflicts. When conflicts arise within a group, alternative behaviours can be adopted to bring back stability; some groups adopt affiliative behaviours (conciliation or reconciliation) while others adopt agonistic behaviours (coalition or redirection). Another similar example is the emergence and transmission of a ‘pacific culture’ within a troop of wild olive baboons (Papio anubis) [27,28]. Due to unfortunate circumstances, a large proportion of the most aggressive males of the troop died of tuberculosis; the few remaining males were untypically affiliative and non-aggressive, creating a troop with high rates of grooming and relaxed dominance hierarchy. The low level of aggression, compared to a neighbouring group residing in the same reserve, was stable and maintained in the group for over 20 years despite the migration of new individuals. These are good examples of convention, because different behaviours can be stable solutions to coordination problems (how to respect the dominance hierarchy for instance) and individuals benefit from a stable social organisation (social stability has important fitness benefits [29–32]).
Notably, most social learning experiments (see [33] for a definition of social learning) do not result in the establishment of conventions, mostly because they do not involve coordination problems but individual solving abilities. For instance, in the classical study of Whiten et al. [34], chimpanzees can solve a puzzle box using two different techniques and they learn to do so by watching other individuals. Since only one individual solves the problem at a time, there is no coordination between them regarding the technique they use. It also emphasizes the difficulty of defining precisely what a convention is, and the fine line between social learning and convention studies.
Some social learning experiments however show suggestive evidence of conventionality. In the studies of van de Waal et al. [35] for instance, groups of vervet monkeys (Chlorocebus aethiops) learned to eat food of one colour (e.g. pink) and avoid food from another colour (e.g. blue). Once this preference had been learned, results showed that monkeys continued to prefer to eat food of only one colour even when they were exposed to edible versions of both colours. Even more surprisingly, monkeys that migrated from one group (e.g. pink) to another (e.g. blue) sometimes adopted the behaviour of members of the group against their own previous preference (e.g. they switch from pink to blue). One possible explanation for the vervet monkeys’ behaviour is that they needed to compete for food to establish their dominance within the group.1 Indeed, dominance hierarchies are examples of conventions, they are stable solutions to coordination problems (to access food and/or reproduction) and provide stable expectations regarding others' behaviour (see [36]). In a particular group there certainly are several stable solutions to organise the dominance between individuals, that organisation is therefore conventional to some extent. However, the dominance hierarchy is also typical of a species and constrained by genetic dispositions and although it could be tempting to extend the concept of conventions to behaviour that are largely genetically determined [2], here we will limit ourselves to learned conventions.
This consideration is also important to determine if non-human primate communication is conventional. Non-human primates’ vocalisations and gestures come from a largely fixed repertoire and are not conventional (e.g. [37–40]). However, note that conventions, in the form of geographical dialects, have been observed in other vocal-learning non-human species (e.g. [41,42]) and that there is some limited evidence of vocal flexibility in non-human primates [43,44]. Watson et al. [44] in particular observed the convergence of food calls during the integration of two chimpanzee groups over a three-year period.
In non-human primates, some flexible gestural communications can emerge through ‘ontogenetic ritualisation’ [45], a process through which an action at the beginning of an action sequence, such as raising an arm before play-hitting another individual, progressively becomes a signal to initiate the action sequence: raising an arm to initiate rough-and-tumble play [46]. However, ontogenetic ritualization does not systematically give rise to conventions because there is a direct link between the sequence of actions to perform and the signal that is used to initiate it, there is therefore no alternative signal through which this could be initiated [7]. Chimpanzee and bonobo gestures overlap extensively in form and meaning for instance [47]. However, in one group of mandrills (Mandrillus sphinx) Laidre [48] described the emergence of a novel eye-covering gesture probably used to signal that the individual was resting. This gesture was unique to this group and had been observed for nearly a decade in the group and never seen elsewhere where other behaviours are used to signal ‘resting’ (such as lying down or turning one's back). Therefore, this signal may be seen as conventional to some extent, even if it is iconic. Interestingly, during the emergence of sign language when signals are created, they are initially iconic, motivated by features of the situated interaction. Later, iconic signals become arbitrary symbols as they are stylised and ritualised. This happens to the point that new learners use them without awareness of the initial motivation, and, in many cases, the initial iconic motivation is undetectable [49].
Of course, the absence of conventions in non-human primates' communication contrasts sharply with human language that is often considered to be a paradigmatic example of convention since the word–meaning association is largely arbitrary [4]. Noteworthily, Tomasello [46] has proposed that linguistic and other human conventions are learned by imitation and that the lack of a capacity for imitation in non-human primates could explain both their apparent lack of conventions and their lack of language (see also [7]). Tamariz [50] goes further and argues that it is not imitation (copying goals and behaviour) but copying without paying attention to goals, that makes language uniquely human [50]. For example, children first learn to reproduce the sound of a word before they learn the conventional way to use them.
The experiments we present below come to question the role of imitation in the establishment of conventions since we observe the emergence of convention in a non-human primate without imitation or other forms of observational learning. In our experiments, two baboons are seated next to each other facing computer touchscreens and they can see each other's screen. When the experiment starts, one baboon is randomly selected and must choose between two stimuli; then the next individual is presented with the same two stimuli and must choose one too. When the same two stimuli are selected, the two baboons are rewarded. Using this coordination task, we wanted to ask the following three main questions:
-
1.
Will the baboons solve this coordination problem by developing a convention? (compared to choosing the stimuli independently or to responding by watching what the other individual selected).
-
2.
If the baboons develop a convention, is observational learning necessary or important?
-
3.
If the baboons develop a convention, to what extent are they arbitrary and depend on the group social structure?
3. Methods
(a) . Participants
Nineteen Guinea baboons (Papio papio, 13 females) from the CNRS primatology centre in Rousset-sur-Arc were tested in this study (mean age: 12.2 years; min = 3.25; max = 25.3). They belonged to a social group living in a 25 × 30 m outdoor enclosure connected to a 6 × 4 m indoor enclosure and two 8 × 4 m trailers (figure 1a). The monkeys had ad libitum access to five S-ALDMs (Social-Automated Computer Learning Devices for Monkeys; see below) two in the first trailer and three in the second.
Figure 1.

(a) Nineteen baboons living in an enclosure have ad libitum access to two trailers, each containing several S-ALDMs (social-automated computer learning devices for monkeys). The diagram shows the two bungalows and the five units. (b) Detail of the organization of a workstation, where two monkeys can work side by side, while seeing each other. (Online version in colour.)
(b) . Social-automated computer learning devices for monkeys (S-ALDMs)
The ALDMs [51,52] are operant conditioning test systems that can be used for testing non-human primates in social settings. ALDMs use an automatic radio frequency identification device (RFID) implanted in each forearm of the monkeys to detect the location and identity of specific individuals. This device makes it possible to test the animals without having to capture and isolate them (additional information of the set-up can be found in [53,54]). The ALDMs presented in Fagot et al. [54] were modified in 2018 and connected in pairs to allow an individual in one ALDM to both see its partner and its touchscreen of the neighbouring ALDM (figure 1b). We dubbed this new version of ALDMs the Social-ALDMs or S-ALDMs for short. Two individuals could therefore see each other and their responses on the screen when a transparent partition between the S-ALDM was used or they had no visual access when an opaque partition was used. All the monkeys had previously participated in studies using this S-ALDM testing system but had never performed the present experiment.
(c) . Computerised task
We used two different tasks, one when two individuals are present, the ‘dual task’, and one when an individual is alone, the ‘filler task’. These two tasks are presented below. When a monkey entered an S-ALDM, a blue screen was displayed in its monitor with a 4 s delay. If another individual was detected in the neighbouring S-ALDM during this delay, a blue screen was also displayed on the adjoining monitor, announcing the synchronization of the two machines and the start of the dual task (figure 2 and video in electronic supplementary material). Otherwise, if the neighbouring S-ALDM stayed empty for 4 s, the filler task started. The function of the filler task was mainly to keep the baboons occupied within the S-ALDM while waiting for another individual to arrive.
Figure 2.
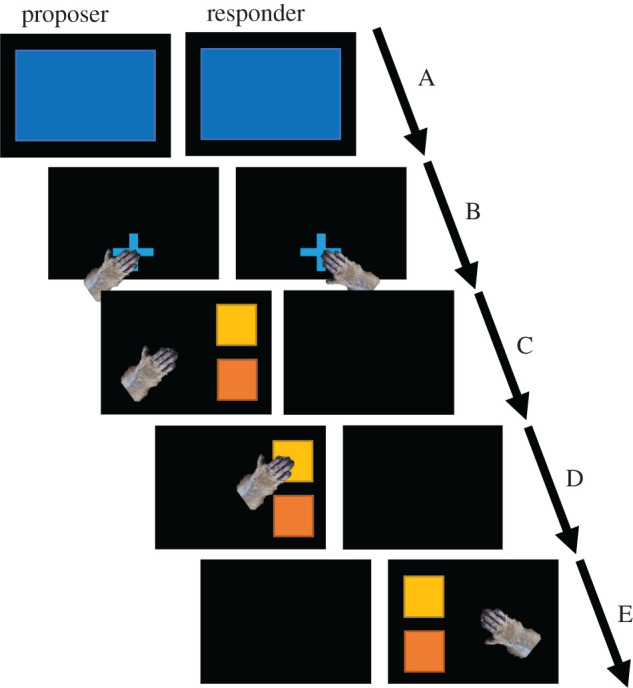
Schematic representation of the dual task on touchscreen. A to E: succession of events. (Online version in colour.)
(i) . Dual task
During the dual task (figure 2), which is the focus of this paper, the test program identified the two monkeys when they entered an S-ALDM, synchronized the two computers, and displayed a blue fixation cross at the bottom centre of both screens (figure 2B). The test started once both monkeys had pressed the fixation cross and within a delay of 4 s (otherwise the trial was aborted and re-presented). One monkey of the pair was then randomly selected. This first monkey, that we are going to call ‘proposer’, had to choose between two randomly placed visual stimuli on the screen (figure 2C). These stimuli were located on the side of the screen so that the second monkey, the ‘responder’, had the best possible view. Once the stimulus was selected by the proposer, it would flash twice (figure 2D), then both stimuli would disappear and appear on the responder's screen. The responder then had to choose one of the two stimuli according to what the neighbour had chosen (figure 2E). If both proposer and responder selected the same target, the trial was considered a success, otherwise, a miss. The success of a test was coded as a binary variable: correct response = 1 and incorrect response = 0. A success triggered the delivery of a reward to the two individuals of the pair, a miss resulted in a 3 s timeout for both. We presented experiments with sets of seven or five different stimuli, therefore 21 or 10 possible different combinations of stimulus pairs. To balance all possible combinations the trials were organized in a randomly ordered block of 42 (seven stimuli) or 40 (for five) trials.
(ii) . Filler task
If a monkey presented itself at an S-ALDM and no other monkey participated in the task in the neighbouring station (at the time of synchronization of the blue screens, figure 2A), a version of the Wisconsin Card Sorting Task (WCST, [55]) was presented. The WSCT is commonly used in executive control studies to assess cognitive flexibility and has been adapted to the ALDM system by Bonté et al. [56]. In the present study, the WCST was used as a filler task, therefore these data will not be analysed in the context of this paper.
(d) . Statistical analysis
We used two measures to understand the emergence of conventions in our experiments. The first variable of interest is the score of individuals (i.e. whether they succeeded at the task and were rewarded) because it indicates if the monkeys are solving the coordination task. To analyse the score, we consider independently the score when an individual is in the role of the proposer, from the score when in the role of the responder. This is because the roles are very different; the proposer cannot respond based on visual information, the receiver can. Note that individuals can in principle have very different scores as proposer and responder, if one individual uses visual information and not another for instance. To understand the evolution of score over time, we analyse the average score on a block of trials (40 or 42 trials, see above) performed by an individual.
To understand how the conventions emerge, we also used the Elo-score rating technique [57]. The Elo-score is used to rank chess players and to analyse dominance hierarchies in animals for instance [58]. The advantage is that it is a dynamic index: when a contest takes place, the Elo-score of the winner increases, while that of the looser decreases. In our case, we used the Elo-score package ([59]; see electronic supplementary material, and the analysis code for details) to represent the dominance between stimuli: every time a monkey chooses one stimulus over another, that stimulus ‘wins’, the other loses (the k parameter was set to 1 and the initial Elo-score was set at 0). The Elo-score technique therefore allows us to see if there is a hierarchy of stimulus preferences appearing and to track changes if they happen, which is important to assess stability.
We performed several experiments in which we manipulated the partition between individuals (either transparent or opaque) and the stimuli (all stimuli are freely available with the data and code to analyse the results). Since these experiments are based on the voluntary partition of monkeys, we could not control the number of trials done per individual or by pairs of individuals, so the number of blocks performed by individuals vary and we analyse only complete blocks. The evolution of score with the number of blocks was evaluated using a generalized linear mixed model (GLMM, [60]) with binomial error distribution and a logit link function. Models were analysed using the stan_glmer function of R's rstanarm package [61] with default weakly informative priors as recommended. Convergence was checked using the shinystan package [62] and we found no convergence issues. To consider inter-individual variability and repeated measurement, we used mixed models including a random intercept and slope (represented by the number of blocks performed) for each individual. Thus, we limit our analysis to complete blocks of trials (all the model details are presented in the electronic supplementary material). Additional analyses use non-parametric tests due to the non-independent nature of the data.
All analyses were done with the statistical software R [63].
4. Study 1: stability of convention
In this first study, the baboons had to choose between pairs of coloured stimuli of seven possible colours (see video S1 in electronic supplementary material). During the first period of the experiment, monkeys could see each other's responses through the transparent partition between them. We start by focusing on the proposer whose role is to choose which of the two colours is to be selected. We found that the proposer's score was initially high (mean score for the first block = 0.75, SEM = 0.02, min = 0.55, max = 0.95) and steadily increased at the beginning of the experiment with the number of blocks (β = 0.03, 95%CI = [0.02,0.04]; figure 3a). This increase in score was linked to the establishment of a hierarchy between colours (figure 3b), some colours (such as yellow and orange) became progressively more chosen by the proposer than others (such as light and dark blue). We found that at the end of the experiment (we used the last two blocks of trials for each proposer to get a reliable measure of score for every pair of colours) the difference in dominance rank between colours was highly correlated to the average score for that pair (Kendall, N = 21, τ = 0.71, z = 4.53, p < 0.001), for instance, the probability of success when orange and dark blue were presented together was on average 99% (a difference in Elo-score of 316) while it was 60% for light versus dark blue (an Elo-score difference of 38).
Figure 3.

Establishment of a colour convention. (a) Evolution of proposers' score with the number of blocks per individual (grey) and on average (in black, error bars represent standard error of the mean). (b) Evolution of Elo-score between colours based on proposers’ behaviour (the seven colours presented are an approximate match to the colours seen by the baboons). (c) Change in score for the responders with the change in partition, from transparent to opaque. (d) Average responder score for every combination of colour pair depending on the colour pair difference in Elo-score for three performance groups (low, medium and high performance). The fitted curve and standard errors are based on a quasi-binomial fit. (Online version in colour.)
The maintenance of the colour hierarchy can be achieved through two non-exclusive processes: the responder could watch the choice of the proposer and respond accordingly, or the responder could have learned the hierarchy and respond without watching. To tease apart these two explanations, we introduced an opaque partition between the two participants. We found that the responder's score decreased for 13/15 individuals (binomial test, p = 0.004), but that the average score remained high (figure 3c). Most of the decisions taken by the responders are therefore based on the knowledge of the hierarchy and do not require visual access. However, we noticed that all the monkeys with very high score with the transparent partition (above 85%) suffered from the introduction of the opaque partition, suggesting these individuals watched the proposer. To test this hypothesis, we divided the group of 19 individuals into three equally sized groups with low, medium and high performing individuals during the transparent condition. For each group we pooled the data and calculated the average score for every pair of colours. Since these data are non-independent (the same colour appears in multiple pairs), we used a non-parametric correlation test to evaluate the relationship between the average score of every pair of colours with the difference in Elo-score depending on the transparent and opaque conditions (figure 3d). We found that high performing monkeys used their knowledge of the hierarchy of colours when the Elo-score difference between the stimuli was high and resorted to visual checking of their social partner when the difference was small (Kendall, N = 5, τ = −0.42, z = −2.66, p = 0.008). By contrast, medium or low performing individuals seem to have only learned the dominance hierarchy of colours, and therefore have a lower score when the rank difference is small (medium: Kendall, N = 5, τ = −0.13, z = −0.85, p = 0.40; low: Kendall, N = 5, τ = −0.06, z = −0.36, p = 0.72).
The results of study 1 therefore show that when baboons had to coordinate by choosing the same colour among a pair, they quickly created a hierarchy between colours that improved their performance (the score increased). All the monkeys that participated regularly learned at least part of the hierarchy. In addition, some monkeys also watched their partner to respond appropriately when the pairs were difficult to discriminate. The hierarchical organisation of colours, although stable, does not seem to be arbitrary because the score at the beginning of the experiment was already initially high, suggesting that baboons did not choose colours arbitrarily but based on pre-existing preferences. To test the generality of our results, the arbitrariness and stability of conventions we conducted two additional sets of experiments.
5. Study 2: generalisation of the results
In experiment 2.1, we replicated the results of study 1 with a new set of five black and white images to test the generality of our conclusions (figure 4; see video S2 in electronic supplementary material). We found equivalent results to our first study: the score of the proposers increased in a similar fashion (β = 0.02, 95%CI = [0.01,0.04]) and was linked to the dominance hierarchy that emerged during the experiment (Kendall, N = 10, τ = 0.91, z = 3.67, p < 0.001). Although the deleterious effect of the opaque partition was less widespread than previously (the responder's score decreased for 11/16 individuals, binomial test, p = 0.11), we found a similar relationship between the monkeys' performance in the transparent condition and the effect of the opaque partition. The performance of high performing monkeys decreased more with the introduction of the opaque partition when the pairs of stimuli were close in Elo-scores than when they were far apart (high performance: Kendall, N = 5, τ = −0.56, z = −2.24, p = 0.025; medium: Kendall, N = 5, τ = −0.28, z = −1.16, p = 0.24; low: Kendall, N = 6, τ = 0.33, z = 1.34, p = 0.18).
Figure 4.

Establishment of conventions with black and white stimuli (a,b) and with an opaque partition (c,d). The first row shows that conventions can be obtained with new stimulus (black and white images) and the second row shows that they can also be obtained without visual contact between the participants (opaque partition). a and c, evolution of score. b and d, evolution of hierarchy. Legend is the same as in figure 3 with the only difference that colours have been arbitrarily attributed to the black and white stimuli. (Online version in colour.)
Monkeys could therefore establish a convention with different types of stimuli such as coloured squares, or black and white images. Next, in experiment 2.2, we questioned the importance of the transparent condition to establish the convention. Originally, we thought the monkeys would use the opportunity to see the action of their neighbour to choose the correct response, but the results showed that for a majority the introduction of the opaque partition had limited effect or no effect at all. We decided to question the necessity of having visual access to establish a convention by introducing five new black and white stimuli and performing the same experiment but with an opaque partition from the start. Visual inspection of figure 4 did not reveal qualitative differences between the experiment with a transparent or an opaque partition (figure 4; electronic supplementary material). We noticed that the average score was lower and the evolution of the dominance less noisy in the opaque condition; this is consistent with the fact that monkeys sometime rely on visual access, but it could also be due to differences in the stimuli we presented since we used different stimuli for the two experiments. Nonetheless, it appeared that individuals could rapidly establish a convention with or without visual access to their partner's responses.
6. Study 3: arbitrariness of convention
A distinctive feature of conventions is their arbitrariness. In experiment 3.1, we tested the possibility of establishing an arbitrary convention by individually training the same monkeys as previously to choose the five black and white stimuli of experiment 2.1 in the opposite order of dominance (we used an opaque partition so that monkeys had no visual access to their neighbours' behaviour; see video S3 and additional details in the electronic supplementary material). For instance, in experiment 2.1, we found that the ‘dog paw’ was more dominant than the ‘leaf’; we therefore individually trained the monkeys on the opposite choice by rewarding the ‘leaf’ and not the ‘paw’. This is a strong test of arbitrariness because presumably monkeys ordered the pairs according to their shared preference, therefore, if an opposite convention is established and remains stable, this shows that opposite conventions can be maintained (a less stringent test would simply try to inverse two stimuli for instance).
All individuals were trained in the same reversed order. During training, we found, as expected, that the score was initially very low (for the first two blocks of trial, mean = 0.30, s.e. = 0.02, min = 0.19, max = 0.43). However, the performance rapidly improved as the monkeys learned to choose the correct stimuli for each pair (for the last two blocks of each individual, mean score = 0.72, s.e. = 0.05, min = 0.28, max = 0.99; see also electronic supplementary material, figure S19). After the training phase we performed the same coordination experiment with the transparent partition as previously and found that the score remained high and showed no sign of decreasing (β = 0.03, 95%CI = [−0.03, 0.08]). The hierarchy was also stable (electronic supplementary material, figure S20) and in the opposite order of the one previously established (table 1). This shows that the established conventions are to some extent arbitrary.
Table 1.
Inversion of hierarchy between stimuli after training. ‘First exposition’ refers to the spontaneous Elo-scores obtained during experiment 2.1 and ‘after reversed training’ to the Elo-scores obtained after each individual had been trained on the reversed order.
| first exposition | after reversed training | |
|---|---|---|
 |
140 | −258 |
 |
63 | −93 |
 |
8 | 13 |
 |
−77 | 112 |
 |
−133 | 225 |
Finally, in experiment 3.2, we wanted to test the impact that each baboon could have on the establishment of convention based on their social relationships with others. We reasoned that well-connected individuals could either be influential and change others or, on the contrary, be more flexible and adapt to others. We selected 11 individuals that had taken part in previous experiments and used data from experiment 2.1 (generalisation of the results, transparent condition, N = 38 409) to calculate the number of times two individuals performed trials together. Using the modularity classes algorithm available in the software Gephi, we identified three clusters (figure 5). From each cluster, we selected the individual with the highest degree (Mako, Violette, Ewine) and individually trained them on five new stimuli organised in an arbitrary hierarchy. The remaining eight individuals were trained using the same procedure but in the reverse order (figure 6; see electronic supplementary material for details on training). Once trained, we tested the establishment of a new convention with a transparent partition as before.
Figure 5.
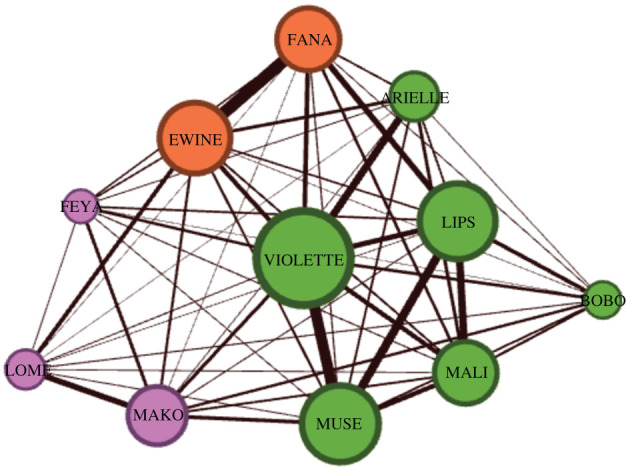
Social network of the 11 individuals tested in experiment 3.2. Individuals with the highest degree in their cluster (Mako, Violette, Ewine) were trained on the opposite convention compared to other individuals. The social network was obtained using the Force Atlas algorithm of Gephi [64]. Colours indicate different clusters, determined using modularity classes. The thickness of the links represents the weight of the links between two individuals (equivalent to their number of trials performed together). The size of the nodes represents the weighted degree of the node (i.e. the sum of the link weights). (Online version in colour.)
Figure 6.

Evolution of Elo-scores trajectories for the entire group (a) and for the first 1000 trials of two individuals (b), Violette (high degree group) and Fana (low degree group). (Online version in colour.)
Individuals rapidly learned to choose the correct stimuli during training (for the last two blocks of each individual, mean score = 0.86, s.e. = 0.03, min = 0.66, max = 0.96; see also electronic supplementary material, figure S24 for the evolution through time). During the coordination experiment, we found that the group converged on the convention of the low-degree individuals (figure 6a) because high-degree individuals changed their behaviour more than did low-degree individuals. For instance, as can be seen on figure 6b, the most dominant and least dominant stimuli were reversed for Violette (high degree) and did not change for Fana (low degree; see electronic supplementary material, figure S27 for the plots of each individual).
7. Discussion and conclusion
In this series of experiments, we showed that when monkeys were faced with a coordination problem, they could spontaneously develop efficient arbitrary stable conventions. Conventions were efficient because the score improved with the emergence of conventions and because we found a direct relationship between the score and the hierarchy among the stimuli (study 1 and 2.1). We showed that conventions were arbitrary and stable because after training to choose stimuli according to a new hierarchy, monkeys could establish and maintain a new convention based on this new hierarchy (experiment 3.1). These three properties are defining features of human conventions and our results therefore complement a small number of field studies (discussed previously) showing that non-human primates and maybe other animals can establish conventions akin to humans.
Our results also speak more generally to our understanding of conventions and how they are established. Moore [7] for instance, raises two important questions in the context of linguistic conventions: which manner of learning would allow the acquisition of convention? And what must we know in order to participate in convention? Further specifying that:
Where a coordinating tool acquires its functional properties through convention, those who learn it must be particularly attentive to the nature of the action that others perform—that is, to the means R that an observed agent employs in pursuit of her goal E. That's because in conventional coordination, unlike in the case of nut-cracking described at the outset, one couldn't (unless one was very lucky) figure out the appropriate means to a goal independently of copying the actions performed by others. ([7, p. 496]; R stands for regularity in behaviour and E for end goal)
In our study, monkeys established a new convention without any visual access to each other's behaviour (experiment 2.2), therefore showing that imitation, or more generally observational learning, is not necessary to establish conventions: in the opaque condition, baboons were able to establish a convention through positive reinforcement and knew nothing about the behaviour of their conspecifics. However, we also found that the most successful monkeys were able to copy the proposer when the task became difficult due to a small Elo-score difference between pairs of stimuli (figure 3d: when the difference in elo-score is small the effect of the opaque partition on the score of high performing monkeys is stronger). This is in agreement with the growing literature on social learning showing that individuals can learn from the observation of conspecifics [65] and with another study showing this effect with touchscreens [66].
According to Lewis [4], there are three main sources explaining the emergence of conventions: agreement, saliency and precedence. Agreement occurs when individuals can communicate and form mutual expectations (i.e. agree on a course of action), for instance when making an appointment. Conventions can also emerge when a solution is so salient that most individuals tend towards that solution (i.e. a preference system). For example, human children can use a salient solution to solve a coordination problem when they cannot communicate [67]. Finally, conventions can emerge through precedence: if a solution becomes more salient and remarkable because it was previously successfully chosen (i.e. a regularity of behaviour). In the context of our study, agreement is unlikely to explain the emergence of conventions because monkeys (i) cannot explicitly agree on which colour to choose and moreover (ii) continued to follow convention when the opaque partition prevented observation of their partner. On the other hand, monkeys can create conventions through a combination of saliency and precedence. Monkeys may all perceive certain stimuli as more salient than others (saliency) and the result of each trial also influences the stimuli future saliency (precedence).
In humans, studies have described tipping points and the role of a determined minority in changing an established convention [68,69]. We attempted to study the importance of well-connected individuals (with a high degree within their cluster), reasoning that if well-connected individuals were to change their behaviour this could change the whole group. Contrary to our expectations, we found that well-connected individuals were more likely to adapt to others rather than influence them. Given that high-degree individuals were also a minority (three versus eight), several factors could explain this result. Individuals may have been conformist and followed the majority for instance (e.g. [70]), or they may be well-connected because they are flexibly adapting to others. Additional experiments will be able to provide a more detailed understanding of the influence of network parameters and other biases by manipulating the proportion and identity of individuals that are trained on alternative conventions.
Conventions are an important aspect of human culture because like norms they stabilise cultural diversity. Surprisingly, although the evolutionary precursors of conventions and norms exist in non-human primates [3], conventions do not seem to be widespread and their contribution to non-human primate culture seems almost anecdotal. One might therefore be tempted to think that conventions require elaborate social learning mechanisms. Using a freely accessible touchscreen system in which pairs of individuals can perform experiments together, we have shown, through five experiments, that non-human primates can develop conventions using positive reinforcement learning and that although copying can be used by some individuals, it is not necessary.
Acknowledgments
We thank Julie Gullstrand and Sebastien Barniaud as well as the staff at the Rousset-sur-Arc Primate Center (CNRS-UPS846, France) for technical support.
Endnote
We thank Kate Arnold (University of St Andrews) for pointing this out to us.
Ethics
This research was carried out in accordance with European Union and French ethical standards and received approval from the French Ministère de l'Education Nationale et de la Recherche (approval no. APAFIS-2717-2015111708173794-V3).
Data accessibility
The data and analysis code used in this article are available here: https://osf.io/b68×2/.
Authors' contributions
A.F., N.C. and J.F. planned the experiment, D.P., N.C. and J.F. planned and constructed the S-ALDMs, A.F. and N.C. performed the experiment, A.F. and N.C. analysed the results, A.F., N.C. and J.F. wrote the manuscript.
Competing interests
We declare we have no competing interests.
Funding
This project received funding from the Institute for Language Communication and the Brain (ILCB) through the Agence Nationale de la Recherche grant no. ANR-16-CONV-0002.
References
- 1.O'Madagain C, Tomasello M. 2021. Shared intentionality, reason-giving and the evolution of human culture. Phil. Trans. R. Soc. B 377, 20200320. ( 10.1098/rstb.2020.0320) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Stephens DW, Heinen VK. 2018. Modeling nonhuman conventions: the behavioral ecology of arbitrary action. Behav. Ecol. 29, 598-608. ( 10.1093/beheco/ary011) [DOI] [Google Scholar]
- 3.Kappeler PM, Fichtel C, van Schaik CP. 2019. There ought to be roots: evolutionary precursors of social norms and conventions in non-human primates. In The normative animal? (eds Roughley N, Bayertz K). New York, NY: Oxford University Press. [Google Scholar]
- 4.Lewis D. 1969. Convention: A philosophical study. Cambridge, MA: Harvard University Press. [Google Scholar]
- 5.Hume D. 1888. A treatise of human nature. New York, NY: Oxford University Press. [Google Scholar]
- 6.Hawkins RXD, Goldstone RL. 2016. The formation of social conventions in real-time environments. PLoS ONE 11, e0151670. ( 10.1371/journal.pone.0151670) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Moore R. 2013. Imitation and conventional communication. Biol. Phil. 28, 481-500. ( 10.1007/s10539-012-9349-8) [DOI] [Google Scholar]
- 8.Hawkins RXD, Goodman ND, Goldstone RL. 2019. The emergence of social norms and conventions. Trends Cogn. Sci. 23, 158-169. ( 10.1016/j.tics.2018.11.003) [DOI] [PubMed] [Google Scholar]
- 9.Bickerton D. 1981. Roots of language. Ann Arbor, MI: Karoma Publishers. [Google Scholar]
- 10.Kegl J. 1994. The Nicaraguan sign language project: an overview. Signpost 7, 24-31. [Google Scholar]
- 11.Fay N, Garrod S, Roberts L, Swoboda N. 2010. The interactive evolution of human communication systems. Cogn. Sci. 34, 351-386. ( 10.1111/j.1551-6709.2009.01090.x) [DOI] [PubMed] [Google Scholar]
- 12.Galantucci B. 2005. An experimental study of the emergence of human communication systems. Cogn. Sci. 29, 737-767. ( 10.1207/s15516709cog0000_34) [DOI] [PubMed] [Google Scholar]
- 13.Bicchieri C. 2005. The grammar of society: the nature and dynamics of social norms. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 14.Nichols S. 2002. On The genealogy of norms: a case for the role of emotion in cultural evolution. Phil. Sci. 69, 234-255. ( 10.1086/341051) [DOI] [Google Scholar]
- 15.Deming W, Neumann J, Morgenstern O. 1944. Theory of games and economic behavior. J. Am. Stat. Assoc. 40, 263. ( 10.2307/2280142) [DOI] [Google Scholar]
- 16.Nash J. 1951. Non-cooperative games. Ann. Math. 54, 286-295. ( 10.2307/1969529) [DOI] [Google Scholar]
- 17.Fudenberg D, Maskin E. 2008. The folk theorem in repeated games with discounting or with incomplete information. In A long-run collaboration on long-run games (eds Fudenberg D, Levine DK), pp. 209-230. Singapore: World Scientific. [Google Scholar]
- 18.Martin CF, Biro D, Matsuzawa T. 2017. Chimpanzees spontaneously take turns in a shared serial ordering task. Sci. Rep. 7, 14307. ( 10.1038/s41598-017-14393-x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stephens DW, Heinen VK. 2018. Modeling nonhuman conventions: the behavioral ecology of arbitrary action. Behav. Ecol. 29, 598-608. ( 10.1093/beheco/ary011) [DOI] [Google Scholar]
- 20.Perry S, et al. 2003. Social conventions in wild white faced capuchin monkeys: evidence for traditions in a neotropical primate. Curr. Anthropol. 44, 241-268. ( 10.1086/345825) [DOI] [Google Scholar]
- 21.Fedigan L. 1993. Sex differences and intersexual relations in adult white-faced capuchins (Cebus capucinus). Int. J. Primatol. 14, 853-877. ( 10.1007/BF02220256) [DOI] [Google Scholar]
- 22.McGrew WC, Marchant LF, Scott SE, Tutin CEG. 2001. Intergroup differences in a social custom of wild chimpanzees: the grooming hand clasp of the Mahale Mountains. Curr. Anthropol. 42, 148-153. ( 10.1086/318441) [DOI] [Google Scholar]
- 23.van Leeuwen EJC. 2021. Temporal stability of chimpanzee social culture. Biol. Lett. 17, 20210031. ( 10.1098/rsbl.2021.0031) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.van Leeuwen EJC, Cronin KA, Haun DBM, Mundry R, Bodamer MD. 2012. Neighbouring chimpanzee communities show different preferences in social grooming behaviour. Proc. R. Soc. B 279, 4362-4367. ( 10.1098/rspb.2012.1543) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nakamura M, McGrew WC, Marchant LF, Nishida T. 2000. Social scratch: another custom in wild chimpanzees? Primates 41, 237-248. ( 10.1007/BF02557594) [DOI] [PubMed] [Google Scholar]
- 26.van de Waal E. 2018. On the neglected behavioural variation among neighbouring primate groups. Ethology 124, 845-854. ( 10.1111/eth.12815) [DOI] [Google Scholar]
- 27.Sapolsky RM. 2006. Social cultures among nonhuman primates. Curr. Anthropol. 47, 641-656. ( 10.1086/504162) [DOI] [Google Scholar]
- 28.Sapolsky RM, Share LJ. 2004. A pacific culture among wild baboons: its emergence and transmission. PLoS Biol. 2, 534-541. ( 10.1371/journal.pbio.0020106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gilby IC, Brent LJN, Wroblewski EE, Rudicell RS, Hahn BH, Goodall J, Pusey AE. 2013. Fitness benefits of coalitionary aggression in male chimpanzees. Behav. Ecol. Sociobiol. 67, 373-381. ( 10.1007/s00265-012-1457-6) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Schülke O, Bhagavatula J, Vigilant L, Ostner J. 2010. Social bonds enhance reproductive success in male macaques. Curr. Biol. 20, 2207-2210. ( 10.1016/j.cub.2010.10.058) [DOI] [PubMed] [Google Scholar]
- 31.Silk JB. 2007. Social Components of fitness in primate groups. Science 317, 1347-1351. ( 10.1126/science.1140734) [DOI] [PubMed] [Google Scholar]
- 32.Silk JB, Beehner JC, Bergman TJ, Crockford C, Engh AL, Moscovice LR, Wittig RM, Seyfarth RM, Cheney DL. 2009. The benefits of social capital: close social bonds among female baboons enhance offspring survival. Proc. R. Soc. B 276, 3099-3104. ( 10.1098/rspb.2009.0681) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Heyes CM. 1994. Social learning in animals: categories and mechanisms. Biol. Rev. 69, 207-231. ( 10.1111/j.1469-185X.1994.tb01506.x) [DOI] [PubMed] [Google Scholar]
- 34.Whiten A, Horner V, de Waal FB. 2005. Conformity to cultural norms of tool use in chimpanzees. Nature 437, 737-740. ( 10.1038/nature04047) [DOI] [PubMed] [Google Scholar]
- 35.van de Waal E, Borgeaud C, Whiten A. 2013. Potent social learning and conformity shape a wild primate's foraging decisions. Science 340, 483-485. ( 10.1126/science.1232769) [DOI] [PubMed] [Google Scholar]
- 36.Bergman TJ, Beehner JC, Cheney DL, Seyfarth RM. 2003. Hierarchical classification by rank and kinship in baboons. Science 302, 1234-1236. ( 10.1126/science.1087513) [DOI] [PubMed] [Google Scholar]
- 37.Arnold K, Zuberbühler K. 2008. Meaningful call combinations in a non-human primate. Curr. Biol. 18, R202-R203. ( 10.1016/j.cub.2008.01.040) [DOI] [PubMed] [Google Scholar]
- 38.Kemp C, Rey A, Legou T, Boë L-J, Berthommier F, Becker Y, Fagot J. 2017. Vocal repertoire of captive guinea baboons (Papio papio). Origins of human language: continuities and discontinuities with nonhuman primates (eds Boë L-J, Fagot J, Perrier P, Schwartz J-L), pp. 15–58. Berlin, Germany: Peter Lang. [Google Scholar]
- 39.Ouattara K, Lemasson A, Zuberbühler K. 2009. Campbell's monkeys use affixation to alter call meaning. PLoS ONE 4, e7808. ( 10.1371/journal.pone.0007808) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tomasello M, Zuberbühler K. 2002. Primate vocal and gestural communication. In The cognitive animal: empirical and theoretical perspectives on animal cognition (eds Bekoff M, Allen C, Burghardt G). Cambridge, MA: MIT Press. [Google Scholar]
- 41.Slater PJB. 1986. The cultural transmission of bird song. Trends Ecol. Evol. 1, 94-97. ( 10.1016/0169-5347(86)90032-7) [DOI] [PubMed] [Google Scholar]
- 42.Weilgart L, Whitehead H. 1997. Group-specific dialects and geographical variation in coda repertoire in South Pacific sperm whales. Behav. Ecol. Sociobiol. 40, 277-285. ( 10.1007/s002650050343) [DOI] [Google Scholar]
- 43.Snowdon CT, Elowson AM. 1999. Pygmy marmosets modify call structure when paired. Ethology 105, 893-908. ( 10.1046/j.1439-0310.1999.00483.x) [DOI] [Google Scholar]
- 44.Watson SK, Townsend SW, Schel AM, Wilke C, Wallace EK, Cheng L, West V, Slocombe KE. 2015. Vocal learning in the functionally referential food grunts of chimpanzees. Curr. Biol. 25, 495-499. ( 10.1016/j.cub.2014.12.032) [DOI] [PubMed] [Google Scholar]
- 45.Tomasello M, Call J. 1997. Primate cognition. Oxford, UK: Oxford University Press. [Google Scholar]
- 46.Tomasello M. 2008. Origins of human communication. New York, NY: MIT Press. [Google Scholar]
- 47.Graham KE, Hobaiter C, Ounsley J, Furuichi T, Byrne RW. 2018. Bonobo and chimpanzee gestures overlap extensively in meaning. PLoS Biol. 16, e2004825. ( 10.1371/journal.pbio.2004825) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Laidre ME. 2011. Meaningful gesture in monkeys? Investigating whether mandrills create social culture. PLoS ONE 6, e14610. (doi:10.1371%2Fjournal.pone.0014610) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Garrod S, Fay N, Lee J, Oberlander J, MacLeod T. 2007. Foundations of representation: where might graphical symbol systems come from? Cognit. Sci. 31, 961-987. ( 10.1080/03640210701703659) [DOI] [PubMed] [Google Scholar]
- 50.Tamariz M. 2019. Replication and emergence in cultural transmission. Phys. Life Rev. 30, 47-71. ( 10.1016/j.plrev.2019.04.004) [DOI] [PubMed] [Google Scholar]
- 51.Fagot J, Bonté E. 2010. Automated testing of cognitive performance in monkeys: use of a battery of computerized test systems by a troop of semi-free-ranging baboons (Papio papio). Behav. Res. Methods 42, 507-516. ( 10.3758/BRM.42.2.507) [DOI] [PubMed] [Google Scholar]
- 52.Fagot J, Paleressompoulle D. 2009. Automatic testing of cognitive performance in baboons maintained in social groups. Behav. Res. Methods 41, 396-404. ( 10.3758/BRM.41.2.396) [DOI] [PubMed] [Google Scholar]
- 53.Fagot J, Gullstrand J, Kemp C, Defilles C, Mekaouche M. 2014. Effects of freely accessible computerized test systems on the spontaneous behaviors and stress level of Guinea baboons (Papio papio). Am. J. Primatol. 76, 56-64. ( 10.1002/ajp.22193) [DOI] [PubMed] [Google Scholar]
- 54.Fagot J, Marzouki Y, Huguet P, Gullstrand J, Claidière N. 2015. Assessment of social cognition in non-human primates using a network of computerized automated learning device (ALDM) test systems. JoVE 99, e52798. ( 10.3791/52798) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Berg EA. 1948. A simple objective technique for measuring flexibility in thinking. J. Gen. Psychol. 39, 15-22. ( 10.1080/00221309.1948.9918159) [DOI] [PubMed] [Google Scholar]
- 56.Bonté E, Flemming T, Fagot J. 2011. Executive control of perceptual features and abstract relations by baboons (Papio papio). Behav. Brain Res. 222, 176-182. ( 10.1016/j.bbr.2011.03.034) [DOI] [PubMed] [Google Scholar]
- 57.Elo AE. 1978. The rating of chessplayers, past and present. New York, NY: Arco Publishing. [Google Scholar]
- 58.Strauss ED, Holekamp KE. 2019. Inferring longitudinal hierarchies: framework and methods for studying the dynamics of dominance. J. Anim. Ecol. 88, 521-536. ( 10.1111/1365-2656.12951) [DOI] [PubMed] [Google Scholar]
- 59.Sánchez-Tójar A, Schroeder J, Farine DR. 2018. A practical guide for inferring reliable dominance hierarchies and estimating their uncertainty. J. Anim. Ecol. 87, 594-608. ( 10.1111/1365-2656.12776) [DOI] [PubMed] [Google Scholar]
- 60.Baayen RH, Davidson DJ, Bates DM. 2008. Mixed-effects modeling with crossed random effects for subjects and items. J. Memory Lang. 59, 390-412. ( 10.1016/j.jml.2007.12.005) [DOI] [Google Scholar]
- 61.Goodrich B, Gabry J, Ali I, Brilleman S. 2020. rstanarm: Bayesian applied regression modeling via Stan (version 2.21.1). https://mc-stan.org/rstanarm.
- 62.Muth C, Oravecz Z, Gabry J. 2018. User-friendly Bayesian regression modeling: a tutorial with rstanarm and shinystan. Quant. Methods Psychol. 14, 99-119. ( 10.20982/tqmp.14.2.p099) [DOI] [Google Scholar]
- 63.R Core Team. 2021. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. http://www.R-project.org/. [Google Scholar]
- 64.Bastian M, Heymann S, Jacomy M. 2009. Gephi: an open source software for exploring and manipulating networks. In Paper presented at the International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA. [Google Scholar]
- 65.Whiten A. 2021. The burgeoning reach of animal culture. Science 372, eabe6514. ( 10.1126/science.abe6514) [DOI] [PubMed] [Google Scholar]
- 66.Subiaul F, Cantlon JF, Holloway RL, Terrace HS. 2004. Cognitive imitation in rhesus macaques. Science 305, 407-410. ( 10.1126/science.1099136) [DOI] [PubMed] [Google Scholar]
- 67.Grueneisen S, Wyman E, Tomasello M. 2015. Children use salience to solve coordination problems. Dev. Sci. 18, 495-501. ( 10.1111/desc.12224) [DOI] [PubMed] [Google Scholar]
- 68.Andreoni J, Nikiforakis N, Siegenthaler S. 2021. Predicting social tipping and norm change in controlled experiments. Proc. Natl Acad. Sci. USA 118, e2014893118. ( 10.1073/pnas.2014893118) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Centola D, Becker J, Brackbill D, Baronchelli A. 2018. Experimental evidence for tipping points in social convention. Science 360, 1116. ( 10.1126/science.aas8827) [DOI] [PubMed] [Google Scholar]
- 70.Haun DBM, Rekers Y, Tomasello M. 2012. Majority-biased transmission in chimpanzees and human children, but not orangutans. Curr. Biol. 22, 727-731. ( 10.1016/j.cub.2012.03.006) [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data and analysis code used in this article are available here: https://osf.io/b68×2/.