

Abstract
Food choice, in animals, has been known to change with internal nutritional state and also with variable dietary conditions. To better characterize mechanisms of diet-induced plasticity of food preference in Drosophila melanogaster, we synthesized diets with macronutrient imbalances and examined how food choice and taste sensitivity were modified in flies that fed on these diets. We found that dietary macronutrient imbalances caused compensatory behavioral shifts in both sexes to increase preference for the macronutrient that was scant in the food source, and simultaneously reduce preference for the macronutrient that was enriched. Further analysis with females revealed analogous changes in sweet taste responses in labellar neurons, with increased sensitivity on sugar-reduced diet and decreased sensitivity on sugar-enriched diet. Interestingly, we found differences in the onset of changes in taste sensitivity and behavior, which occur over 1–4 d, in response to dietary sugar reduction or enrichment. To investigate molecular mechanisms responsible for diet-induced taste modulation, we used candidate gene and transcriptome analyses. Our results indicate that signaling via Dop2R is involved in increasing cellular and behavioral sensitivity to sugar as well as in decreasing behavioral sensitivity to amino acids on dietary sugar reduction. On the other hand, cellular and behavioral sensitivity to sugar relies on dilp5 and a decrease in sugar preference following dietary sugar abundance was correlated with downregulation of dilp5. Together, our results suggest that feeding preference for sugar and amino acid can be modulated independently to facilitate food choice that accounts for prior dietary experience.
SIGNIFICANCE STATEMENT Animals adjust their feeding preferences based on prior dietary experiences. Here, we find that upon dietary macronutrient deprivation, flies undergo compensatory changes in food preference. The altered preference correlates with changes in peripheral taste sensitivity. While Dop2R mediates changes following dietary sugar reduction, downregulation of dilp5 is associated with changes caused by a sugar-enriched diet. This study contributes to a better understanding of neurophysiological plasticity of the taste system in flies, and its role in facilitating adjustment of foraging behavior based on nutritional requirements.
Keywords: drosophila, diet, dopamine, insulin, taste, plasticity
Introduction
All living animals consume food to survive and to reproduce. To maintain ideal health, animals require a balanced diet with optimum amounts of different nutrients. Macronutrients like carbohydrates and proteins are essential for growth, survival, and reproduction, but an unbalanced intake of these nutrients can be detrimental to metabolic homeostasis (Waterlow, 1974; Samuelsson et al., 2008; Malik et al., 2010; Mozaffarian et al., 2011; von Geijer and Ekelund, 2015). Low-sugar diets can lead to conditions like ketoacidosis (von Geijer and Ekelund, 2015), whereas high-sugar diets can cause metabolic disorders such as obesity, hypertension, and Type II diabetes (Malik et al., 2010; Mozaffarian et al., 2011). Similarly, protein deficiency can bring about kwashiorkor and marasmus malnutrition conditions (Waterlow, 1974), and protein excess is often associated with heart and kidney malfunction as well as adverse effects on bones (Barzel and Massey, 1998; Martin et al., 2005; Virtanen et al., 2018).
The genetic model insect, Drosophila melanogaster, also requires macronutrients such as sugars and amino acids for survival. Flies use the gustatory system to sense these nutrients and initiate feeding behaviors. A number of recent studies have shown that feeding behaviors can be altered by dietary experience (Nishimura et al., 2012; Inagaki et al., 2014; Wang et al., 2016; Steck et al., 2018; Devineni et al., 2019; May et al., 2019). Moreover, dietary macronutrient imbalance can have targeted effects on gustatory system function. Flies respond to deprivation of yeast, a natural source of proteins and amino acids, by increasing feeding preference for yeast and yeast components (Liu et al., 2017). A recent study suggests that yeast deprivation is accompanied by a concomitant decrease in sugar preference (Liu et al., 2017). Both the increase in yeast intake and decrease in sugar intake are controlled by a group of dopaminergic neurons in the brain, their effects mediated via Dop2R and Dop1R neurons, respectively (Liu et al., 2017). These previous investigations were conducted with the same amount of sugar in control and yeast-depleted diets (Inagaki et al., 2012; Marella et al., 2012; Wang et al., 2016; Liu et al., 2017), nevertheless the results align with more recent work that shows reduced taste sensitivity to sugar upon feeding on a high-sugar diet (May et al., 2019). One consideration is that in some of these studies, there were very large differences in total caloric value of experimental and control diets.
Here, we systematically investigate changes in feeding preference and taste sensitivity following dietary macronutrient imbalance using diets in which excess of one macronutrient is offset by depletion of another. We compare the effects of diets with contrasting proportions of carbohydrate and protein on feeding preference and find that dietary imbalance brings about compensatory shifts in food choice in short-term behavior assays. Evaluation of sweet taste sensitivity shows that sugar taste responses are enhanced with a sugar-reduced diet and depressed with a sugar-enriched diet. A time course analysis of diet-induced cellular and behavioral shifts in sugar sensitivity shows that the observed shifts are reversible, and restoration to “baseline” values is observed on return to the balanced diet. The increase in sugar sensitivity brought on by the sugar-reduced diet depends on Dop2R function, whereas the sugar-enriched diet-induced decrease in sugar sensitivity relies on the secretion of the insulin-like peptide Dilp5 from brain insulin-producing cells (IPCs), and its action on serotonergic neurons. We find that Dop2R function is also required for decreased amino acid preference following a low-sugar-high-protein diet, while the increase in amino acid preference following feeding with a high-sugar-low-protein diet is independent of Dilp5 function. Thus, two different neuromodulatory pathways that are triggered by different macronutrient-deprivation paradigms appear to cause antagonistic changes in peripheral taste sensitivity and feeding preference. This study contributes to our understanding of how diet-induced signaling in the brain can alter the sensitivity of peripheral chemosensory neurons.
Materials and Methods
Fly stocks
Fly stocks were maintained at 22–25°C on standard cornmeal-dextrose media. Wild-type flies were w1118 (BL5905), unless otherwise noted. elav-GAL4 (BL8765), Dop2R RNAi1 (BL26001), Dop2R RNAi2 (BL36824), Dop2R1 (BL52025), Dop2R2 (BL52157), dilp5 RNAi (BL33683), UAS-TNTG (BL28828), InR1 (BL11661), InR2 (BL9646), UAS-InR RNAi (BL51518), and Ir76b1 (BL51309) were obtained from the Bloomington Drosophila Stock Center. Gad1-GAL4, Tdc2-GAL4, Ple-GAL4, Trh-GAL4, v-Glut-GAL4, dilp2 (BL 30 881), dilp3 (BL30882), dilp5 (BL30884), npF-GAL4, and dilp2-GAL4 flies were kindly shared by Naoki Yamanaka, University of California, Riverside.
Chemicals
Sucrose (S7903), D(+) glucose (G8270), L-serine (84959), L-phenylalanine (P-5482), L-threonine (89179), and yeast extract (Y1625), were obtained from Sigma. Drosophila agar Type II (66-103) and Tegosept (20-258) were obtained from Apex Bioresearch Products. Propionic acid (UN3463) was obtained from Acros Organics, yellow cornmeal (43-375) was obtained from Quaker. For behavior experiments, tastant solutions were prepared in water. For electrophysiological recordings, tastant solutions were prepared in 30 mm tricholine citrate (Sigma, T0252).
Diets
Table 1 lists the composition of the diets used in this study, with amounts sufficient to prepare 100 vials of each diet.
Table 1.
Composition of test diets
| Balanced | Sugar-reduced/yeast-enriched | Sugar-enriched/yeast-depleted | |
|---|---|---|---|
| D(+) glucose (g) | 100 | 0 | 126.67 |
| Yeast extract (g) | 50 | 239.5 | 0 |
| Cornmeal (g) | 70 | 70 | 70 |
| Drosophila agar (g) | 6 | 6 | 6 |
| Propionic acid (ml) | 6 | 6 | 6 |
| Tegosept (ml) | 12 | 12 | 12 |
| milliQ water (autoclaved; ml) | 1025 | 1025 | 1025 |
| Calorie (100 ml) | 74.55 | 88.2 (↑ ∼18% from balanced) |
71.04 (↓ ∼5% from balanced) |
Diets were prepared by first heating the Drosophila agar with a small amount of water in a beaker or conical flask on a hot plate. The mixture was stirred continuously with a magnetic stir bar. Once the agar was dissolved, a slurry of cornmeal was added, followed by sugar, yeast extract and the remaining water, if any. The food was allowed to come to a rolling boil, taken off the heat and allowed to cool. Propionic acid and Tegosept were added when the temperature fell below 80°C. Food was dispensed into vials using a serologic pipette. Calories were calculated using Drosophila Dietary Composition Calculator (DDCC; https://www.brodericklab.com/DDCC.php#boxCalc; Lesperance and Broderick, 2020). We also used the calculator to check the diet composition, which we include in Table 2.
Table 2.
Nutrient composition and caloric content of test diets
| Balanced (g/100 ml) | Sugar-reduced/yeast-enriched (g/100 ml) | Sugar-enriched/ yeast- depleted (g/100 ml) | |
|---|---|---|---|
| Fiber | 0.97 | 0.97 | 0.97 |
| Sugar (added) | 9.76 | 0 | 12.36 |
| Protein | 0.8 | 1.91 | 0.51 |
| Fat | 0.18 | 0.29 | 0.15 |
| Carbohydrate | 15.62 | 6.18 | 18.14 |
Binary choice assays
Feeding preference assays were conducted as previously described (Wisotsky et al., 2011; Ganguly et al., 2017). Briefly, flies were starved on water-saturated Kimwipes for 24 h before the experiment. For the assays, flies were provided a choice between two different tastants prepared in 0.75% agarose, one containing blue dye (0.25 mg/ml indigocarmine, Sigma 18130) and the other containing pink dye (0.5 mg/ml sulforhodamine B, Sigma 230162), which were spotted in tight-fit Petri dishes. Flies were allowed to feed in a dark humid chamber at 25°C for 2 h and subsequently scored for color in their abdomen. Preference indices for “tastant 1” were calculated for each trial (plate) based on the following formula: (Ntastant1 + 0.5Npurple)/(Ntastant1 + Npurple + Ntastant2).
Consumption assay
Flies fed on diets as indicated for each experiment were wet-starved for 24 h before consumption experiments. Flies were then transferred to vials containing selected test diets mixed with 1% sulforhodamine B (pink dye). Vials were placed in a dark humid chamber at 25°C and flies were allowed to feed for 2 h, following which they were killed by transferring to −80°C for 30 min. Entire digestive tracts dissected from individual flies were placed in PCR tubes containing 5 µl of water. Tubes were vortexed vigorously and spun in a minicentrifuge (VWR) for 30 s. The supernatant was collected, and its absorbance was measured at 565 nm using a Nanodrop 2000c Spectrometer. Gut extract of unfed flies was obtained in the same way and used as negative control. To calculate ingested food volume from absorbance measurements, we generated a standard curve with a concentration range of sulforhodamine B.
Electrophysiology
Extracellular tip recordings were all obtained from L-type labellar sensilla as described previously (Benton and Dahanukar, 2011). Recordings were taken from age-matched mated female flies treated in parallel to the control and experimental diet regimens. Tastants were dissolved in 30 mm tricholine citrate, which served as electrolyte. Neuronal responses were calculated by counting the number of spikes in the first 500-ms period after contact.
Fluorescence imaging
Brains from 7- to 10-d-old flies were dissected in 1× PBS with 0.3% Triton X-100 (PBST) and fixed in 4% paraformaldehyde in PBST for 30 min. They were then subjected to three washes of 20 min each in PBST and mounted on slides using VECTASHIELD anti-fade mounting media (Vector Labs H-1200). GFP fluorescence was visualized using a Zeiss LSM 70 confocal microscope and images were captured in 1-µm optical sections.
Library construction, sequencing, and sequencing data analysis
For library preparation, taste tissues and brains were dissected from flies exposed to indicated control and experimental dietary regimens. Proboscis and tarsi were collected from 150 flies; whole brains were dissected from 16 flies. Tissues were mechanically crushed with disposable RNase-free plastic pestles, and total RNA was isolated using a TRIzol-based protocol. cDNA libraries were prepared from total RNA using the Illumina TruSeq RNA Sample Preparation kit (v2). Single-end sequencing (1 × 50 bp) was performed using the Hi-Seq2000 platform at the UCR Genomics Core facility. There was an average of 30.6 million reads per replicate, with an average of 81% mapped. Reads were aligned to the latest release of the D. melanogaster genome (dm6) and quantified with kallisto (version kallisto 0.43.1; Bray et al., 2016). Only libraries for which we obtained >75% alignment were used for downstream analysis. Transcript counts were summarized to gene-level using tximport package (version 1.4.0; Soneson et al., 2015). Differentially expressed gene (DEG) analysis was performed with the edgeR package (version 3.18.1; Robinson et al., 2010), using low count filtering (cpm > 0.5) and TMM normalization. All significance analyses of gene overlap were done using the GeneOverlap package in R package (version 1.14.0). GO enrichment analysis was performed using PANTHER (version 13.1; Mi et al., 2017).
PCR and qPCR analysis
For verification of mutants, genomic DNA was extracted from a single fly and used as template for PCR. For qPCR, total RNA was extracted from brain, proboscis, and tarsi using TRIzol (Invitrogen) and treated with DNase I (Invitrogen) to remove genomic DNA, according to the manufacturer's instructions. Total RNA (3 μg) was input to synthesize cDNA, using SuperScript III First-Strand Synthesis SuperMix (Invitrogen). All qPCR experiments were performed with technical replicates using SYBR Green Supermix (Bio-Rad), according to the manufacturers' instructions. Each sample was measured in triplicate, and relative expression was calculated using the 2−ΔΔCt method and normalized to the housekeeping gene rp49; t tests were performed using the CFX Maestro software (Bio-Rad).
Statistical analyses
Sample sizes for individual experiments were determined on the basis of previous literature. All statistical tests were performed using GraphPad Prism software. For experiments with multiple independent variables, we first conducted two-way ANOVA and inspected the F statistics to determine whether there was a significant effect of the independent variables on the response variable. Subsequently we conducted Sidak's or Tukey's multiple comparisons tests for post hoc analysis to generate adjusted p values for different pairwise comparisons. For pairwise comparisons with only one independent variable, Mann–Whitney U test was used. In all violin plots, the solid line indicates the median while the dotted lines represent the upper and lower quartiles, respectively. All error bars shown in the scatter plots, and column graphs represent SEM.
Data availability
Raw data are available on Mendeley Data (DOI: 10.17632/ngzwtf9tym.1).
Results
Dietary sugar-to-protein ratio sways feeding preference for sugar
The experimental strategy we adopted to investigate how changes in macronutrient balance in the diet influence taste is summarized in Figure 1A. We prepared three different diets: a “balanced” diet containing glucose and yeast extract (which contains amino acids, peptides, nucleic acids, carbohydrates including trehalose, water-soluble vitamins and trace elements) in the same proportions as in the standard cornmeal-dextrose diet, a “sugar-reduced” diet containing no added glucose and subsequently enriched in yeast extract, and a “sugar-enriched” diet containing ∼1.26× the amount of glucose and depleted of yeast extract. The composition of the diets is listed in the methods, and was designed to maintain caloric content within 20% of that of the balanced diet. Adult flies raised on standard cornmeal-dextrose media were collected at 0–2 d after eclosion, transferred to vials containing one of the three diets, and tested for cellular and/or behavioral responses to selected tastants. Male and female flies were housed together, and we assumed that all female flies would be mated at the time of testing. Sexed flies were analyzed separately, since previous studies have found sex-specific differences in feeding preference for yeast or yeast extract (Ribeiro and Dickson, 2010; Ganguly et al., 2017).
Figure 1.
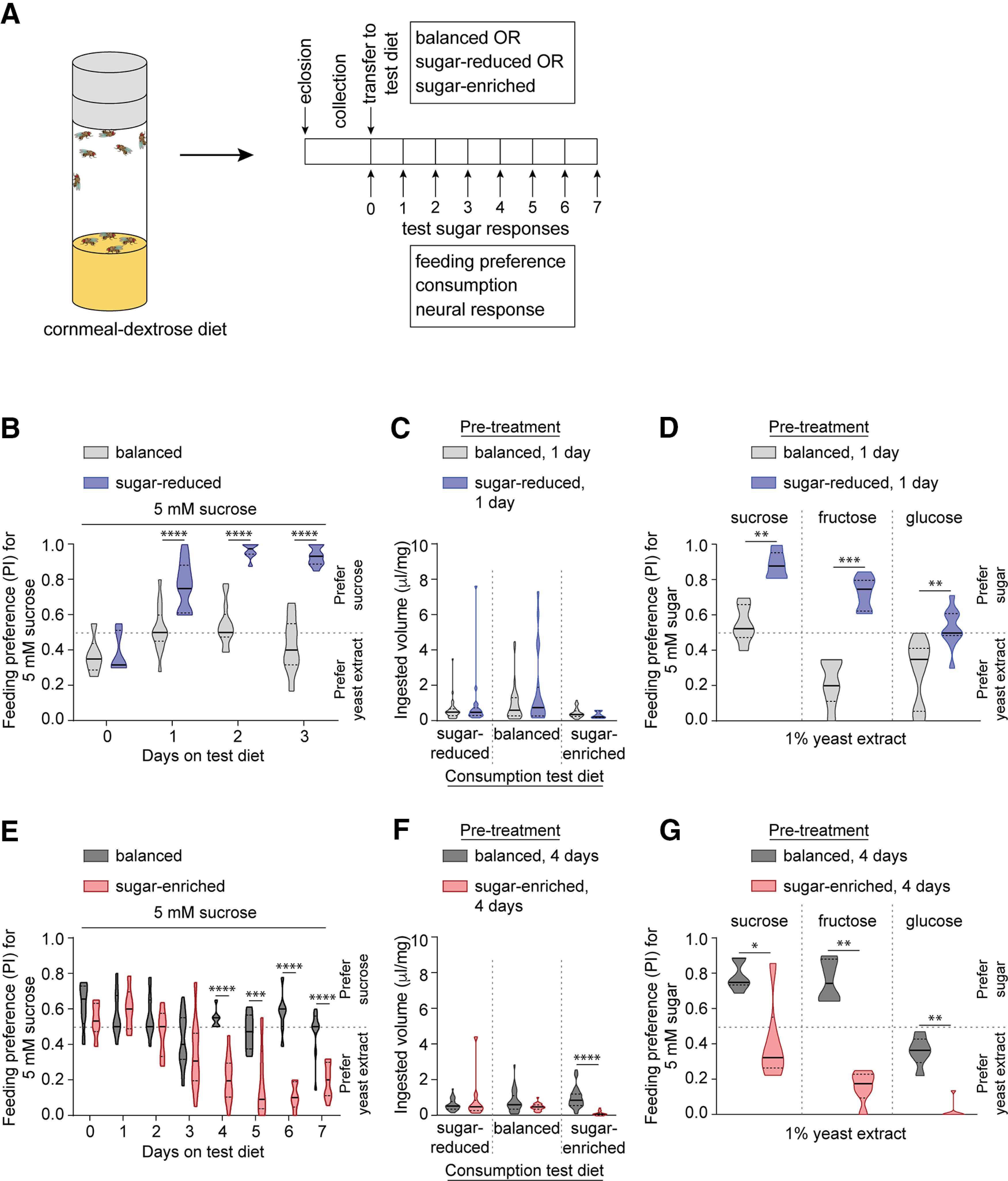
Dietary sugar-protein imbalance leads to shifts in feeding preference. A, Schematic representing the experimental strategy to evaluate diet-induced alteration of taste sensitivity and preference. B, Preference of mated females (and males, see Extended Data Fig. 1-1) fed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays; n = 6 balanced (0 d), 6 sugar-reduced (0 d), 15 balanced (1 d), 13 sugar-reduced (1 d), 13 balanced (2 d), 8 sugar-reduced (2 d), 17 balanced (3 d), 15 sugar-reduced (3 d). Feeding participation of males and females prefed on the balanced or sugar-reduced diets is shown in Extended Data Figures 1-1, 1-2. C, Ingested volume (μl) of indicated diet normalized to body weight (mg) in mated females prefed with indicated diet for 1 d and then tested for 2-h consumption of sugar-reduced (n = 35, 18), balanced (n = 35, 19), or sugar-enriched (n = 31, 10) diets. Head and body weights of females prefed on the balanced or sugar-reduced diets for 1 d are shown in Extended Data Figure 1-3. D, Preference of mated females prefed on indicated diet for 5 mm sugar (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays; n = 6 sucrose, 7 fructose, 10 glucose for both balanced and sugar-reduced. E, Preference of mated females (and males, see Extended Data Fig. 1-4) prefed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract in binary choice assays; n = 6 balanced, 6 sugar-enriched (0 d), n = 9 balanced, 6 sugar-enriched (1 d), n = 13 balanced, 6 sugar-enriched (2 d), n = 17 balanced, 10 sugar-enriched (3 d), n = 7 balanced, 16 sugar-enriched (4 d), n = 6 balanced, 6 sugar-enriched (5 d), n = 7 balanced, 6 sugar-enriched (6 d), n = 10 balanced, 11 sugar-enriched (7 d). Feeding participation of males and females prefed on the balanced or sugar-enriched diets are shown in Extended Data Figures 1-4, 1-5. F, Ingested volume (μl) of indicated diet normalized to body weight (mg) in mated females prefed with indicated diet for 4 d and then tested for 2-h consumption of sugar-reduced (n = 34, 10), balanced (n = 28, 21), or sugar-enriched (n = 31, 21) diets. Head and body weights of females prefed on the balanced or sugar-enriched diets for 4 d are shown in Extended Data Figure 1-6. G, Preference of mated females prefed on indicated diet for 5 mm sugar (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays; n = 6 sucrose, 6 fructose, 6 glucose for both balanced and sugar-enriched. Diet-induced changes in feeding preference are compared in starved and unstarved females in Extended Data Figure 1-7. For all graphs, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Data in B, C, E, F were analyzed using two-way ANOVA with Sidak's post hoc multiple comparisons test. Data in D, G were analyzed with the Mann–Whitney test.
Preference (left) and participation (right) of males prefed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays; n = 6 balanced (0 d), 6 sugar-reduced (0 d), 15 balanced (1 d), 13 sugar-reduced (1 d), 13 balanced (2 d), 8 sugar-reduced (2 d), 17 balanced (3 d), 15 sugar-reduced (3 d); *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-1, TIF file (5.9MB, tif) .
Participation of mated females fed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays; n = 6 balanced (0 d), 6 sugar-reduced (0 d), 15 balanced (1 d), 13 sugar-reduced (1 d), 13 balanced (2 d), 8 sugar-reduced (2 d), 17 balanced (3 d), 12 sugar-reduced (3 d). No significant differences were found between balanced and sugar-reduced, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-2, TIF file (2MB, tif) .
Head and body weights of mated females prefed with indicated diet for 1 d; n = 6 trials. ns = not significant, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-3, TIF file (2.1MB, tif) .
Preference (left) and participation (right) of males prefed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract in binary choice assays; n = 6 balanced, 6 sugar-enriched (0 d), n = 9 balanced, 6 sugar-enriched (1 d), n = 13 balanced, 6 sugar-enriched (2 d), n = 17 balanced, 10 sugar-enriched (3 d), n = 7 balanced, 16 sugar-enriched (4 d), n = 6 balanced, 6 sugar-enriched (5 d), n = 7 balanced, 6 sugar-enriched (6 d), n = 10 balanced, 11 sugar-enriched (7 d); *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-4, TIF file (5.7MB, tif) .
Participation of mated females fed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract in binary choice assays; n = 6 balanced, 6 sugar-enriched (0 d), n = 9 balanced, 6 sugar-enriched (1 d), n = 13 balanced, 6 sugar-enriched (2 d), n = 17 balanced, 10 sugar-enriched (3 d), n = 7 balanced, 16 sugar-enriched (4 d), n = 6 balanced, 6 sugar-enriched (5 d), n = 7 balanced, 6 sugar-enriched (6 d), n = 10 balanced, 11 sugar-enriched (7 d). No significant differences were found between balanced and sugar-enriched, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-5, TIF file (2.5MB, tif) .
Head and body weights of mated females prefed with indicated diet for 4 d; n = 7 trials. ns = not significant, ***p < 0.001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-6, TIF file (2MB, tif) .
Left, Preference of mated females prefed on balanced or sugar-reduced diet, and either wet-starved (starved) or not starved (unstarved) prior to testing in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye); n = 5. Right, Preference of mated females prefed on balanced or sugar-enriched diet, and either wet-starved (starved) or not starved (unstarved) prior to testing in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye); n = 6. For both graphs, *p < 0.05, **p < 0.01, ***p < 0.001, one-way ANOVA with Tukey's post hoc multiple comparisons test. Download Figure 1-7, TIF file (5.4MB, tif) .
We first determined whether feeding on the test diets altered preference in binary choice feeding assays. Flies were tested daily for a period of 3 to 7 d after transferring to sugar-reduced or sugar-enriched diets, respectively. Age-matched flies fed on the balanced diet were tested in parallel. Preference for sugar was assessed in feeding choice assays with 5 mm sucrose and 1% yeast extract as the two stimuli. We found that female flies that were fed on sugar-reduced food exhibited higher preference for sucrose as compared with those fed on the balanced diet. A significant increase in sugar preference was observed after just 1 d on the sugar-reduced diet, and the elevated preference remained with continued feeding on the same diet (Fig. 1B, w1118 balanced vs sugar-reduced diet for 0 d p = 0.9997, 1 d p < 0.0001, 2 d p < 0.0001, 3 d p < 0.0001, Sidak's post hoc multiple comparisons test; see also Table 3). Although the baseline sucrose preference is higher in males to begin with, a potentiation of sugar preference was observed after feeding on the sugar-reduced diet, which was significant after 3 d (Extended Data Fig. 1-1, w1118 balanced vs sugar-reduced diet for 3 d p = 0.0012, Sidak's post hoc multiple comparisons test; see also Table 3). The dietary change did not affect participation of the flies in binary choice feeding assays, except for a small drop in males at 3 d after feeding on the sugar-reduced diet (Extended Data Figs. 1-1, 1-2, w1118 balanced vs sugar-reduced diet for 3 d p < 0.0001, Sidak's post hoc multiple comparisons test; see also Table 3). To rule out the possibility that the sugar-reduced diet triggers a change in amount of food intake rather than a shift in food preference, we used short-term consumption assays to compare intake of the three test diets, presented individually, after prefeeding on either balanced or sugar-reduced diets. We found that flies fed on the sugar-reduced diet for a day consumed the same amounts of each of the three diets as those fed on the balanced diet (Fig. 1C, w1118 balanced vs sugar-reduced pretreatment, p = 0.6303 for sugar-reduced consumption diet, p = 0.0792 balanced consumption diet, p = 0.9368 for sugar-enriched consumption diet; Sidak's post hoc multiple comparisons test; see also Table 3). Neither head weight nor body weight were affected on feeding on the sugar-reduced diet (Extended Data Fig. 1-3, p = 0.0833 for head, p = 0.6883 for body, Sidak's post hoc multiple comparisons test; see also Table 3). Notably, flies subjected to the sugar-reduced diet also showed increased preference for fructose and glucose in binary choice assays with yeast extract, indicating that relative feeding preference for sugar was broadly altered with this dietary regimen (Fig. 1D, w1118 balanced vs sugar-reduced diet, p = 0.0022 sucrose, p = 0.0006 fructose, p = 0.0014 glucose; Mann–Whitney test).
Table 3.
Summary of statistical analysis
| Figure number | Independent variables | Statistical test | Result | Post hoc test | Comparison groups | Adjusted p value |
|---|---|---|---|---|---|---|
| Figure 1 | ||||||
| Figure 1 B | Diet × days | Two-way ANOVA | Diet × days F(3,85) = 19.61, p < 0.0001; Days F(3,85) = 28.6, p < 0.0001; Diet F(1,85) = 143.7, p < 0.0001 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-reduced | 0 d | 0.9997 |
| 1 d | <0.0001 | |||||
| 2 d | <0.0001 | |||||
| 3 d | <0.0001 | |||||
| Figure 1 C | Pretreatment diet × consumption test diet | Two-way ANOVA | Pretreatment diet × consumption test diet F(2,142) = 1.6, p = 0.2054; consumption diet F(2,142) = 7.406, p = 0.0009; Pretreatment diet F(1,142) = 1.936, p = 0.1662 |
Sidak's multiple comparisons test, w1118 balanced vs w1118 sugar-reduced pretreatment | Sugar-reduced consumption diet | 0.6303 |
| Balanced consumption diet | 0.0792 | |||||
| Sugar-enriched consumption diet | 0.9368 | |||||
| Figure 1 D | Diet | Mann–Whitney test | w1118 balanced vs sugar-reduced 1 d | Sucrose | 0.0022 | |
| Fructose | 0.0006 | |||||
| Glucose | 0.0014 | |||||
| Figure 1 E | Diet × days | Two-way ANOVA | Diet × days F(7,126) = 6.363, p < 0.00001; Days F(7,126) = 9.812, p < 0.0001; Diet F(1,126) = 72.14, p < 0.0001 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-enriched diet | 0 d | 0.9552 |
| 1 d | 0.9999 | |||||
| 2 d | 0.8709 | |||||
| 3 d | 0.6033 | |||||
| 4 d | <0.0001 | |||||
| 5 d | 0.0003 | |||||
| 6 d | <0.0001 | |||||
| 7 d | <0.0001 | |||||
| Figure 1 F | Pretreatment diet × consumption test diet | Two-way ANOVA | Pretreatment diet × consumption test diet F(2,139) = 9.621, p = 0.0001; Consumption diet F(2,139) = 0.6712, p = 0.5127; Pretreatment diet F(1,139) = 7.691, p = 0.0063 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-enriched pretreatment diet | Sugar-reduced consumption diet | 0.3549 |
| Balanced consumption diet | 0.135 | |||||
| Sugar-enriched consumption diet | <0.0001 | |||||
| Figure 1 G | Diet | Mann–Whitney test | w1118 balanced vs sugar-enriched diet, 4 d | Sucrose | 0.0238 | |
| Fructose | 0.0022 | |||||
| Glucose | 0.0022 | |||||
| Figure 2 | ||||||
| Figure 2 B | Diet × sucrose concentration | Two-way ANOVA | Diet × sucrose concentration F(3,124) = 5.566, p = 0.0013; Sucrose concentration F(3,124) = 108.8, p < 0.0001; Diet F(1,124) = 37.05, p < 0.0001 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-reduced, 1 d | 0 mm suc | 0.9998 |
| 10 mm suc | 0.0282 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | 0.0001 | |||||
| Figure 2 D | Diet × sucrose concentration | Two-way ANOVA | Diet × sucrose concentration F(3,156) = 3.428, p = 0.0187; Sucrose concentration F(3,156) = 153.7, p < 0.0001; Diet F(1,156) = 24.03, p < 0.0001 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-enriched, 4 d | 0 mm suc | >0.9999 |
| 10 mm suc | 0.0093 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | 0.1411 | |||||
| Figure 3 | ||||||
| Figure 3 A | Diet regimen × days | Two-way ANOVA | Diet regimen × days F(3,67) = 4.309, p = 0.0077; Days F(3,67) = 14.33, p < 0.0001; Diet regimen F(1,67) = 15.18, p = 0.0002 |
Sidak's multiple comparisons test, w1118 balanced → balanced vs sugar-reduced → balanced | 0 d | 0.9995 |
| 1 d | <0.0001 | |||||
| 2 d | 0.03 | |||||
| 3 d | 0.9645 | |||||
| Figure 3 C | Diet regimen × sucrose concentration | Two-way ANOVA | Diet regimen × sucrose concentration F(15,348) = 4.555, p < 0.0001; Diet regimen F(3,348) = 437.5, p < 0.0001; Sucrose concentration F(5,348) = 26.17, p < 0.0001 |
Tukey's multiple comparisons test, w1118 balanced 1 d vs sugar-reduced 1 d | 0 mm suc | >0.9999 |
| 10 mm suc | 0.0004 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | <0.0001 | |||||
| Tukey's multiple comparisons test, w1118 balanced 2 d vs sugar-reduced 1 d + balanced 1 d | 0 mm suc | >0.9999 | ||||
| 10 mm suc | >0.9999 | |||||
| 30 mm suc | 0.01 | |||||
| 100 mm suc | 0.0007 | |||||
| Tukey's multiple comparisons test, w1118 balanced 3 d vs sugar-reduced 1 d + balanced 2 d | 0 mm suc | >0.9999 | ||||
| 10 mm suc | 0.286 | |||||
| 30 mm suc | 0.1972 | |||||
| 100 mm suc | 0.0264 | |||||
| Figure 3 D | Diet regimen × days | Two-way ANOVA | Diet regimen × days F(7,119) = 5.144, p < 0.0001; Days F(7,119) = 8.939, p < 0.0001; Diet regimen F(1,119) = 9.152, p = 0.003 |
Sidak's multiple comparisons test, w1118 balanced → balanced vs sugar-enriched → balanced | 1 d | 0.9996 |
| 2 d | 0.7448 | |||||
| 3 d | 0.3981 | |||||
| 4 d | <0.0001 | |||||
| 5 d | 0.9312 | |||||
| 6 d | 0.8033 | |||||
| 7 d | 0.9717 | |||||
| 8 d | 0.9814 | |||||
| Figure 3 F | Diet regimen × sucrose concentration | Two-way ANOVA | Diet regimen × sucrose concentration F(21,582) = 4.349, p < 0.0001; Sucrose concentration F(3,582) = 496.5, p < 0.0001; Diet regimen F(7,582) = 20.06, p < 0.0001 |
Tukey's multiple comparisons test, w1118 balanced 4 d vs sugar-enriched 4 d | 0 mm suc | >0.9999 |
| 10 mm suc | 0.0365 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | 0.4696 | |||||
| Tukey's multiple comparisons test, w1118 balanced 5 d vs sugar-enriched 4 d + balanced 1 d | 0 mm suc | >0.9999 | ||||
| 10 mm suc | 0.0266 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | <0.0001 | |||||
| Tukey's multiple comparisons test, w1118 balanced 6 d vs sugar-enriched 4 d + balanced 2 d | 0 mm suc | >0.9999 | ||||
| 10 mm suc | 0.5622 | |||||
| 30 mm suc | >0.9999 | |||||
| 100 mm suc | 0.3759 | |||||
| Tukey's multiple comparisons test, w1118 balanced 7 d vs sugar-enriched 4 d + balanced 3 d | 0 mm suc | >0.9999 | ||||
| 10 mm suc | 0.9963 | |||||
| 30 mm suc | >0.9999 | |||||
| 100 mm suc | 0.0843 | |||||
| Figure 4 | ||||||
| Figure 4 A | Diet | Mann–Whitney test | w1118 balanced vs sugar-reduced, 1 d | 0.0043 | ||
| Figure 4 B | Diet × days | Two-way ANOVA | Diet × days F(7,85) = 10.7, p < 0.0001; Days F(7,85) = 9.944, p < 0.0001; Diet F(7,85) =78.05, p < 0.0001 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-enriched | 0 d | 0.9982 |
| 1 d | 0.99 | |||||
| 2 d | 0.9998 | |||||
| 3 d | >0.9999 | |||||
| 4 d | 0.0003 | |||||
| 5 d | <0.0001 | |||||
| 6 d | <0.0001 | |||||
| 7 d | <0.0001 | |||||
| Figure 4 C | Diet regimen × days | Two-way ANOVA | Diet regimen × days F(6,70) = 2.462, p = 0.0322; Days F(6,70) = 3.882, p = 0.0021; Diet regimen F(1,70) = 10.98, p = 0.0015 |
Sidak's multiple comparisons test, w1118 balanced → balanced vs sugar-enriched → balanced | 1 d | 0.8866 |
| 2 d | 0.9938 | |||||
| 3 d | >0.9999 | |||||
| 4 d | 0.0018 | |||||
| 5 d | 0.0283 | |||||
| 6 d | 0.9994 | |||||
| 7 d | 0.9992 | |||||
| Figure 4 D | Diet | Mann–Whitney test | balanced vs sugar-enriched, 4 d | w 1118 | 0.0017 | |
| Ir76b 1 | 0.0851 | |||||
| Figure 4 E | Diet × tastant | Two-way ANOVA | Diet × tastant F(2,63) = 5.967, p = 0.0042; Tastant F(2,63) = 110.7, p < 0.0001; Diet F(1,63) = 0.1558, p = 0.6943 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-enriched 4 d | 100 mm suc | 0.1164 |
| 3AA mix | 0.0215 | |||||
| 30 mm TCC | >0.9999 | |||||
| Figure 5 | ||||||
| Figure 5 A | Diet × genotype | Two-way ANOVA | Diet × genotype F(2,36) = 5.645, p = 0.0074; Genotype F(2,36) = 29.4, p < 0.0001; Diet F(1,36) = 7.007, p = 0.012 |
Sidak's multiple comparisons test, balanced 1 d | elav-GAL4 ctrl vs elav>Dop2R RNAi1 | 0.0719 |
| elav-GAL4 ctrl vs elav>Dop2R RNAi2 | 0.0022 | |||||
| elav>Dop2R RNAi1 vs elav>Dop2R RNAi2 | <0.0001 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-reduced, 1 d | elav-GAL4 ctrl | 0.0008 | ||||
| elav>Dop2R RNAi1 | 0.3067 | |||||
| elav>Dop2R RNAi2 | 0.7995 | |||||
| Figure 5 B | Diet × genotype | Two-way ANOVA | Diet × genotype F(2,38) = 2.084, p = 0.1384; Genotype F(2,38) = 8.539, p = 0.0009; Diet F(1,38) = 9.284, p = 0.0042 |
Sidak's multiple comparisons test, balanced, 1 d | w1118 vs Dop2R1 | 0.3804 |
| w1118 vs Dop2R2 | 0.9989 | |||||
| Dop2R1 vs Dop2R2 | 0.2273 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-reduced, 1 d | w 1118 | 0.0075 | ||||
| Dop2R 1 | 0.9423 | |||||
| Dop2R 2 | 0.3635 | |||||
| Figure 5 C | Diet × genotype | Two-way ANOVA | Diet × genotype F(2,28) = 7.765, p = 0.0021; Genotype F(2,28) = 19.98, p < 0.0001; Diet F(1,28) = 145.4, p < 0.0001 |
Sidak's multiple comparisons test, balanced, 4 d | w1118 vs Dop2R1 | 0.2904 |
| w1118 vs Dop2R2 | 0.3225 | |||||
| Dop2R1 vs Dop2R2 | 0.9999 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-enriched, 4 d | w 1118 | 0.0019 | ||||
| Dop2R 1 | <0.0001 | |||||
| Dop2R 2 | <0.0001 | |||||
| Figure 5 D | Diet × genotype | Two-way ANOVA | Diet × genotype F(2,35) = 3.487, p = 0.0416; Genotype F(2,35) = 3.903, p = 0.0295; Diet F(1,35) = 16.26, p = 0.0003 |
Sidak's multiple comparisons test, balanced, 1 d | w1118 vs Dop2R1 | 0.9987 |
| w1118 vs Dop2R2 | 0.3036 | |||||
| Dop2R1 vs Dop2R2 | 0.1933 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-reduced, 1 d | w 1118 | 0.0002 | ||||
| Dop2R 1 | 0.0993 | |||||
| Dop2R 2 | 0.3417 | |||||
| Figure 7 | ||||||
| Figure 7 A | Diet × genotype | Two-way ANOVA | Diet × genotype F(3,52) = 5.235, p = 0.0031; Genotype F(3,52) = 4.874, p = 0.0046; Diet F(1,52) = 72.05, p < 0.0001 |
Sidak's multiple comparisons test, balanced, 4 d | w1118 vs dilp2 | 0.9986 |
| w1118 vs dilp3 | 0.808 | |||||
| w1118 vs dilp5 | 0.0716 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-enriched, 4 d | w 1118 | <0.0001 | ||||
| dilp2 | <0.0001 | |||||
| dilp3 | 0.0035 | |||||
| dilp5 | 0.4748 | |||||
| Figure 7 B | Diet × genotype | Two-way ANOVA | Diet × genotype F(1,56) = 1.547, p = 0.2188; Genotype F(1,56) = 9.133, p = 0.038; Diet F(1,56) = 50.03, p < 0.0001 |
Sidak's multiple comparisons test, w1118 vs dilp5 | Balanced | 0.3738 |
| Sugar-reduced | 0.0084 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-reduced, 1 d | w 1118 | <0.0001 | ||||
| dilp5 | 0.0008 | |||||
| Figure 7 E | Diet × genotype | Two-way ANOVA | Diet × genotype F(4,95) = 4.202, p = 0.0035; Genotype F(4,95) = 19.84, p < 0.0001; Diet F(1,95) = 56.16, p < 0.0001 |
Sidak's multiple comparisons test, balanced, 4 d | dilp2-GAL4 vs UAS-dilp5 RNAi | 0.6802 |
| dilp2-GAL4 vs dilp2>dilp5 RNAi | 0.9988 | |||||
| dilp2-GAL4 vs UAS-TNTG | 0.0672 | |||||
| dilp2-GAL4 vs dilp2>TNTG | 0.8096 | |||||
| UAS-dilp5 RNAi vs dilp2>dilp5 RNAi | 0.9992 | |||||
| UAS-dilp5 RNAi vs dilp2>TNTG | 0.217 | |||||
| UAS-TNTG vs dilp2>TNTG | 0.6080 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-enriched, 4 d | dilp2-GAL4 | <0.0001 | ||||
| UAS-dilp5 RNAi | 0.0007 | |||||
| dilp2>dilp5 RNAi | 0.4489 | |||||
| UAS-TNTG | 0.0105 | |||||
| dilp2>TNTG | 0.2572 | |||||
| Figure 7 F | Diet × genotype | Two-way ANOVA | Diet × genotype F(1,26) = 7.507e-006, p = 0.9978 Genotype F(1,26) = 0.4644, p = 0.5016; Diet F(1,26) = 21.88, p < 0.0001 |
Sidak's multiple comparisons test, w1118 vs dilp5 | Balanced | 0.8703 |
| Sugar-enriched | 0.8612 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-enriched, 4 d | w 1118 | 0.0109 | ||||
| dilp5 | 0.0021 | |||||
| Figure 7 G | Diet × genotype | Two-way ANOVA | Diet × genotype F(1,26) = 24.27, p < 0.0001; Genotype F(1,26) = 0.01833, p = 0.8933; Diet F(1,26) = 21.31, p < 0.0001 |
Sidak's multiple comparisons test, w1118 vs InR1/InR2 | Balanced | 0.0045 |
| Sugar-enriched | 0.0028 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-enriched 4 d | w 1118 | <0.0001 | ||||
| InR 1 /InR 2 | 0.9753 | |||||
| Figure 7 H | Diet × genotype | Two-way ANOVA | Diet × genotype F(2,36) = 3.092, p = 0.0576; Genotype F(2,36) = 9.274, p = 0.0006; Diet F(1,36) = 10.88, p = 0.0002 |
Sidak's multiple comparisons test, balanced 4 d | UAS-InR RNAi vs elav-GAL4 | 0.0044 |
| UAS-InR RNAi vs elav-GAL4>InR RNAi | 0.8006 | |||||
| elav-GAL4 vs elav-GAL4>InR RNAi | 0.0351 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-enriched | UAS-InR RNAi | 0.0423 | ||||
| elav-GAL4 | 0.0031 | |||||
| elav-GAL4>InR RNAi | 0.999 | |||||
| Figure 7 I | Diet × genotype | Two-way ANOVA | Diet × genotype F(6,83) = 5.527, p < 0.0001; Genotype F(6,83) = 3.476, p = 0.0041; Diet F(1,83) = 179.4, p < 0.0001 |
Sidak's multiple comparisons test, balanced 4 d, genotype comparison | UAS-InR RNAi vs v-Glut>InR RNAi | 0.9987 |
| UAS-InR RNAi vs Gad1>InR RNAi | 0.9721 | |||||
| UAS-InR RNAi vs Ple>InR RNAi | 0.9993 | |||||
| UAS-InR RNAi vs Trh>InR RNAi | 0.0825 | |||||
| UAS-InR RNAi vs Tdc2>InR RNAi | 0.9527 | |||||
| UAS-InR RNAi vs npF>InR RNAi | 0.9845 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-enriched 4 d | UAS-InR RNAi | <0.0001 | ||||
| v-Glut>InR RNAi | <0.0001 | |||||
| Gad1>InR RNAi | 0.0022 | |||||
| Ple>InR RNAi | 0.0002 | |||||
| Trh>InR RNAi | 0.8235 | |||||
| Tdc2>InR RNAi | <0.0001 | |||||
| npF>InR RNAi | <0.0001 | |||||
| Extended Data Figure 1 | ||||||
| Extended Data Figure 1-1 | Diet × days (preference) | Two-way ANOVA | Diet × days F(3,85) = 1.169, p = 0.3265; Days F(3,85) = 7.171, p = 0.0002; Diet F(1,85) = 11.11, p = 0.0013 |
Sidak's multiple comparisons test, w1118 balanced vs w1118 sugar-reduced | 0 d | 0.9922 |
| 1 d | 0.4079 | |||||
| 2 d | 0.2306 | |||||
| 3 d | 0.0012 | |||||
| Extended Data Figure 1-1 | Diet × days (participation) | Two-way ANOVA | Diet × days F(3,85) = 2.911, p = 0.0391; Days F(3,85) = 6.110, p = 0.0008; Diet F(1,85) = 11.66, p = 0.0010 |
Sidak's multiple comparisons test, w1118 balanced vs w1118 sugar-reduced | 0 d | >0.9999 |
| 1 d | 0.7113 | |||||
| 2 d | 0.1281 | |||||
| 3 d | <0.0001 | |||||
| Extended Data Figure 1-2 | Diet × days (participation) | Two-way ANOVA | Diet × days F(3,85) = 0.2272, p = 0.8772; Days F(3,85) = 6.110, p = 0.0008; Diet F(1,85) = 0.2833, p = 0.0960 |
Sidak's multiple comparisons test, w1118 balanced vs w1118 sugar-reduced | 0 d | >0.9999 |
| 1 d | 0.507 | |||||
| 2 d | 0.7586 | |||||
| 3 d | 0.4953 | |||||
| Extended Data Figure 1-3 | Diet × body part (weight) | Two-way ANOVA | Diet × body part (weight) F(1,20) = 0.9544, p = 0.3403; Body part (weight) F(1,20) = 41.62, p < 0.0001; Diet F(1,20) = 4.355, p = 0.0499 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-reduced | Head | 0.0833 |
| Body | 0.6883 | |||||
| Extended Data Figure 1-4 | Diet × days (preference) | Two-way ANOVA | Diet × days F(7,126) = 4.646, p = 0.1258; Days F(7,126) = 1.656, p = 0.1258; Diet F(1,126) = 32.23, p < 0.0001 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-enriched | 0 d | 0.9999 |
| 1 d | >0.9999 | |||||
| 2 d | 0.0536 | |||||
| 3 d | >0.9999 | |||||
| 4 d | 0.1052 | |||||
| 5 d | 0.1807 | |||||
| 6 d | 0.025 | |||||
| 7 d | <0.0001 | |||||
| Extended Data Figure 1-4 | Diet × days (participation) | Two-way ANOVA | Diet × days (participation) F(7,126) = 0.8195, p = 0.5728; Days (participation) F(7,126) = 2.521, p = 0.0185; Diet F(1,126) = 1.417, p = 0.2361 |
Sidak's multiple comparisons test, w1118 balanced diet vs sugar-enriched | 0 d | 0.9979 |
| 1 d | 0.2761 | |||||
| 2 d | >0.9999 | |||||
| 3 d | >0.9999 | |||||
| 4 d | 0.9324 | |||||
| 5 d | >0.9999 | |||||
| 6 d | >0.9999 | |||||
| 7 d | 0.9876 | |||||
| Extended Data Figure 1-5 | Diet × days (participation) |
Two-way ANOVA | Diet × days F(7,116) = 0.8336, p = 0.5613; Days F(7,126) = 1.28, p = 0.2655; Diet F(1,126) = 3.735, p = 0.0555 |
Sidak's multiple comparisons test, w1118 balanced vs sugar-enriched | 0 d | 0.1439 |
| 1 d | 0.9922 | |||||
| 2 d | >0.9999 | |||||
| 3 d | 0.8270 | |||||
| 4 d | 0.9914 | |||||
| 5 d | >0.9999 | |||||
| 6 d | 0.9946 | |||||
| 7 d | >0.9999 | |||||
| Extended Data Figure 1-6 | Diet × body part | Two-way ANOVA | F(1,24) = 9.469, p = 0.0052 | Sidak's multiple comparisons test, w1118 balanced vs sugar-enriched | Head | 0.9662 |
| Body | 0.0008 | |||||
| Extended Data Figure 1-7, left | Pretreatment condition | One-way ANOVA | F(2,12) = 7.588, p = 0.0074 | Tukey's multiple comparisons test | Balanced (unstarved) vs sugar-reduced (unstarved) |
0.0394 |
| Balanced (starved) vs balanced (unstarved) |
0.6718 | |||||
| Sugar-reduced (unstarved) vs balanced (starved) | 0.0114 | |||||
| Extended Data Figure 1-7, right | Pretreatment condition | One-way ANOVA | F(2,15) = 66.59, p < 0.0001 | Tukey's multiple comparisons test | Balanced (unstarved) vs sugar-enriched (unstarved) |
<0.0001 |
| Balanced (starved) vs balanced (unstarved) |
0.3368 | |||||
| Sugar-enriched (unstarved) vs balanced (starved) | <0.0001 | |||||
| Extended Data Figure 2 | ||||||
| Extended Data Figure 2-1 | Diet pretreatment × sucrose concentration | Two-way ANOVA | Interaction F(3,88) = 4.757, p = 0.0040; Sucrose concentration F(3,88) = 109.1, p < 0.0001; Diet F(1,88) = 5.967, p = 0.0166 |
Sidak's multiple comparisons test, balanced vs sugar-reduced | 0 mm suc | >0.9999 |
| 10 mm suc | 0.0212 | |||||
| 30 mm suc | 0.0069 | |||||
| 100 mm suc | 0.5976 | |||||
| Extended Data Figure 2-2 | Diet pretreatment × sucrose concentration | Two-way ANOVA | Interaction F(3,108) = 4.706, p = 0.004; Sucrose concentration F(3,108) = 93.65, p < 0.0001; Diet F(1,108) = 31.03, p < 0.0001 |
Sidak's multiple comparisons test, balanced vs sugar-enriched | 0 mm suc | >0.9999 |
| 10 mm suc | 0.001 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | 0.1023 | |||||
| Extended Data Figure 5 | ||||||
| Extended Data Figure 5-1 | Diet | Independent samples t test | Dop2R | 0.324177 | ||
| Extended Data Figure 5-3 | Diet × genotype | Two-way ANOVA | Diet × genotype F(2,38) = 0.3505, p = 0.7066; Genotype F(2,38) = 1.861, p = 0.1694; Diet F(1,38) = 0.2966, p = 0.5892 |
Sidak's multiple comparisons test, balanced vs sugar-reduced, 1 d | w 1118 | >0.9999 |
| Dop2R 1 | >0.9999 | |||||
| Dop2R 2 | 0.6014 | |||||
| Extended Data Figure 5-4 | Pretreated diet | Log-rank test | Df = 1, χ2 = 0.5312 |
Mantel–Cox | Water vs sugar-reduced | 0.4661 |
| Extended Data Figure 5-6 | Genotype/pretreatment × sucrose concentration | Two-way ANOVA | Genotype/pretreatment × sucrose concentration F(9,373) = 6.873, p < 0.0001; Sucrose concentration F(3,373) = 449.3, p < 0.0001; Genotype/pretreatment F(3,373) = 32.83, p < 0.0001 |
Tukey's multiple comparisons test, w1118 balanced vs sugar-reduced, 1 d | 0 mm suc | 0.9969 |
| 10 mm suc | 0.0017 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | <0.0001 | |||||
| Tukey's multiple comparisons test, w1118 balanced vs Dop2R1 balanced | 0 mm suc | 0.7828 | ||||
| 10 mm suc | 0.8215 | |||||
| 30 mm suc | 0.5909 | |||||
| 100 mm suc | 0.294 | |||||
| Tukey's multiple comparisons test, Dop2R1 balanced vs Dop2R1 sugar-reduced | 0 mm suc | >0.9999 | ||||
| 10 mm suc | >0.9999 | |||||
| 30 mm suc | 0.0244 | |||||
| 100 mm suc | 0.0002 | |||||
| Extended Data Figure 5-7 | Diet × genotype | Two-way ANOVA | Diet × genotype F(2,40) = 4.420, p = 0.0184; Genotype F(2,40) = 11.24, p = 0.0001; Diet F(1,40) = 24.31, p < 0.0001 |
Sidak's multiple comparisons test, balanced 1 d | UAS-Dop2R RNAi vs Gad1-GAL4>Dop2R RNAi | 0.0428 |
| UAS-Dop2R RNAi vs Gad1-GAL4 | 0.036 | |||||
| Gad1-GAL4>Dop2R RNAi vs Gad1-GAL4 | 0.9958 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-reduced 1 d | Dop2R RNAi | 0.0303 | ||||
| Gad1-GAL4>Dop2R RNAi | 0.7004 | |||||
| Gad1-GAL4 | <0.0001 | |||||
| Extended Data Figure 5-9 | Genotype (relative expression) | Independent samples t test | Dop2R | 0.6521 | ||
| Extended Data Figure 5-10 | Diet × genotype (left) | Two-way ANOVA | Diet × genotype F(2,49) = 1.054, p = 0.3564; Genotype F(2,49) = 7.388, p = 0.0016; Diet F(1,49) = 63.32 p < 0.0001 |
Sidak's multiple comparisons test, balanced, 1 d | Gr64f-GAL4 vs UAS-D2R RNAi1 | 0.1064 |
| Gr64f-GAL4 vs Gr64f>D2R RNAi1 | 0.381 | |||||
| UAS-Dop2R RNAi1 vs Gr64f>D2R RNAi1 | 0.946 | |||||
| Sidak's multiple comparisons test, balanced vs sugar-reduced, 1 d | Gr64f-GAL4 | <0.0001 | ||||
| UAS-D2R RNAi1 | 0.0057 | |||||
| Gr64f-GAL4>D2R RNAi1 | <0.0001 | |||||
| Extended Data Figure 5-10 | Diet × genotype (right) | Two-way ANOVA | Diet × genotype F(7,97) = 3.41, p = 0.0027; Genotype F(7,97) = 4.784, p = 0.0001; Diet F(1,97) = 158.9, p < 0.0001 |
Sidak's multiple comparisons test, balanced vs sugar-reduced 1 d | Tdc2-GAL4>Dop2R RNAi | 0.0128 |
| Tdc2-GAL4 | 0.0001 | |||||
| Ple>Dop2R RNAi | 0.0026 | |||||
| Ple-GAL4 | <0.0001 | |||||
| Trh>Dop2R RNAi | 0.0002 | |||||
| Trh-GAL4 | 0.0219 | |||||
| v-Glut>Dop2R RNAi | <0.0001 | |||||
| v-Glut-GAL4 | 0.0276 | |||||
| Extended Data Figure 5-11 | Diet × sucrose concentration | Two-way ANOVA | Diet × sucrose concentration F(9,208) = 3.177, p = 0.0013; Genotype F(3,208) = 362.8, p < 0.0001; Diet F(3,208) = 13.91, p < 0.0001 |
Tukey's multiple comparisons test, UAS-D2R RNAi balanced vs sugar-reduced 1 d | 0 mm suc | >0.9999 |
| 10 mm suc | 0.0027 | |||||
| 30 mm suc | 0.0164 | |||||
| 100 mm suc | 0.0004 | |||||
| Tukey's multiple comparisons test, Gad1-GAL4>UAS-D2R RNAi balanced vs sugar-reduced 1 d | 0 mm suc | 0.9946 | ||||
| 10 mm suc | 0.0993 | |||||
| 30 mm suc | 0.2012 | |||||
| 100 mm suc | 0.1977 | |||||
| Extended Data Figure 6 | ||||||
| Extended Data Figure 6-1 | Diet (brain) | Independent samples t test | dilp3 | 0.680683 | ||
| dilp5 | 0.009507 | |||||
| Extended Data Figure 6-1 | Diet (proboscis) | Independent samples t test | dilp3 | 0.753426 | ||
| dilp5 | 0.680805 | |||||
| Extended Data Figure 7 | ||||||
| Extended Data Figure 7-2 | Diet × genotype (participation) | Two-way ANOVA | Diet × genotype F(3,52) = 0.05811, p = 0.9814; Genotype F(3,52) = 5.497, p = 0.0023; Diet F(1,52) = 0.393, p = 0.5335 |
Sidak's multiple comparisons test, balanced vs sugar-enriched 4 d | w 1118 | >0.9999 |
| dilp2 | 0.973 | |||||
| dilp3 | 0.9983 | |||||
| dilp5 | 0.9848 | |||||
| Extended Data Figure 7-3 | Diet/pretreatment × sucrose concentration | Two-way ANOVA | Diet/pretreatment × sucrose concentration F(9,379) = 7.609, p < 0.0001; Sucrose concentration F(3,379) = 428.8, p < 0.0001; Diet/pretreatment F(3,379) = 34.27, p < 0.0001 |
Tukey's multiple comparisons test, w1118 balanced vs w1118 sugar-enriched 4 d | 0 mm suc | 0.9968 |
| 10 mm suc | 0.03 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | 0.0653 | |||||
| Tukey's multiple comparisons test, dilp5 balanced vs dilp5 sugar-enriched | 0 mm suc | mm 0.9998 | ||||
| 10 mm suc | 0.0174 | |||||
| 30 mm suc | 0.9728 | |||||
| 100 mm suc | 0.0006 | |||||
| Tukey's multiple comparisons test, w1118 balanced vs dilp5 balanced | 0 mm suc | 0.9654 | ||||
| 10 mm suc | <0.0001 | |||||
| 30 mm suc | <0.0001 | |||||
| 100 mm suc | 0.0005 | |||||
| Extended Data Figure 7-4 | Genotype (brain) | Independent samples t test | dilp5 | 0.360222 | ||
| Extended Data Figure 7-5 | Genotype (brain) | Independent samples t test | InR | 0.070619 |
A summary of statistical analyses used in this study, which includes information about the statistical tests and p values.
Experiments with the sugar-enriched diet yielded complementary results, however the onset of change in preference was delayed in comparison with that elicited by the sugar-reduced diet. Females displayed a reduced feeding preference for sugar in binary choice assays with yeast extract only after 4 d on the sugar-enriched diet (Fig. 1E, w1118 balanced vs sugar-enriched diet for 4 d p < 0.0001, 5 d p = 0.0003, 6 d p < 0.0001, 7 d p < 0.0001; Sidak's post hoc multiple comparisons test; see also Table 3), and males exhibited a shift toward yeast extract preference after 6 d (Extended Data Fig. 1-4, w1118 balanced vs sugar-enriched diet for 6 d p = 0.025, 7 d p < 0.0001; Sidak's post hoc multiple comparisons test; see also Table 3). For both sexes, the altered preference persisted for the remainder of the 7-d test period, during which flies were maintained on the same diet. In no-choice consumption assays, the test flies exhibited no differences in intake of the balanced diet or of the sugar-reduced/yeast-enriched diet (Fig. 1F, w1118 balanced vs sugar-enriched pretreatment, p = 0.3549 for sugar-reduced consumption diet, p = 0.135 balanced consumption diet; Sidak's post hoc multiple comparisons test; see also Table 3), consistent with the idea that the observed shift in behavioral preference is unlikely to arise solely from changes in food intake caused by dietary protein deprivation (Steck et al., 2018). Flies fed with the sugar-enriched diet exhibited no difference in participation in binary choice feeding assays compared with those fed on balanced diet (Extended Data Figs. 1-4, 1-5; Table 3). We did, however, observe reduced intake of the sugar-enriched diet (Fig. 1F, p < 0.0001 for sugar-enriched consumption diet; Sidak's post hoc multiple comparisons test; see also Table 3), indicative of sugar-enriched diet-dependent changes in food intake over time. This was reflected in the body weight, as flies fed on the sugar-enriched diet showed a reduction in body weight, but not head weight, as compared with those fed on the balanced diet (Extended Data Fig. 1-6, w1118 balanced vs sugar-enriched diet, head weight p = 0.9662, body weight p = 0.0008; Sidak's post hoc multiple comparisons test; see also Table 3). As observed for experiments with the sugar-reduced diet, sugar preference was broadly altered with the sugar-enriched dietary treatment, with reduced preference for fructose and glucose in addition to sucrose (Fig. 1G, w1118 balanced vs sugar-enriched diet, p = 0.0238 sucrose, p < 0.0001 fructose, p = 0.0001 glucose; Mann–Whitney test). With both imbalanced diets, shifts in feeding preference were independent of starved or fed state (Extended Data Fig. 1-7; Table 3). Combined, these results suggest that relative feeding preference for sugar shifts in a manner that compensates for the amount of sugar in the diet.
Dietary sugar-protein variation alters sweet taste
One possible mechanism that would account for behavioral adjustments in feeding preference is a change in sensitivity of sweet taste neurons. Indeed, previous studies have reported that flies given very sweet diets, containing either a high concentration of sucrose or an artificial sweetener sucralose, experienced changes in neuronal sensitivity to sugar (Wang et al., 2016; May et al., 2019). We therefore measured responses of labellar taste hairs to sucrose using extracellular tip recordings. Individual hairs were tested with a range of sucrose concentrations to make comparisons of sucrose sensitivity between flies subjected to the different diet treatments. Sucrose responses from labellar L-type sensilla of flies fed on sugar-reduced diet for 1 d were significantly higher at all tested concentrations than those of age-matched flies fed on balanced diet (Fig. 2A,B, w1118 balanced vs sugar-reduced diet, p = 0.0282 for 10 mm sucrose, p < 0.0001 for 30 mm sucrose, p = 0.0001 for 100 mm sucrose; Sidak's post hoc multiple comparisons test; see also Table 3).
Figure 2.
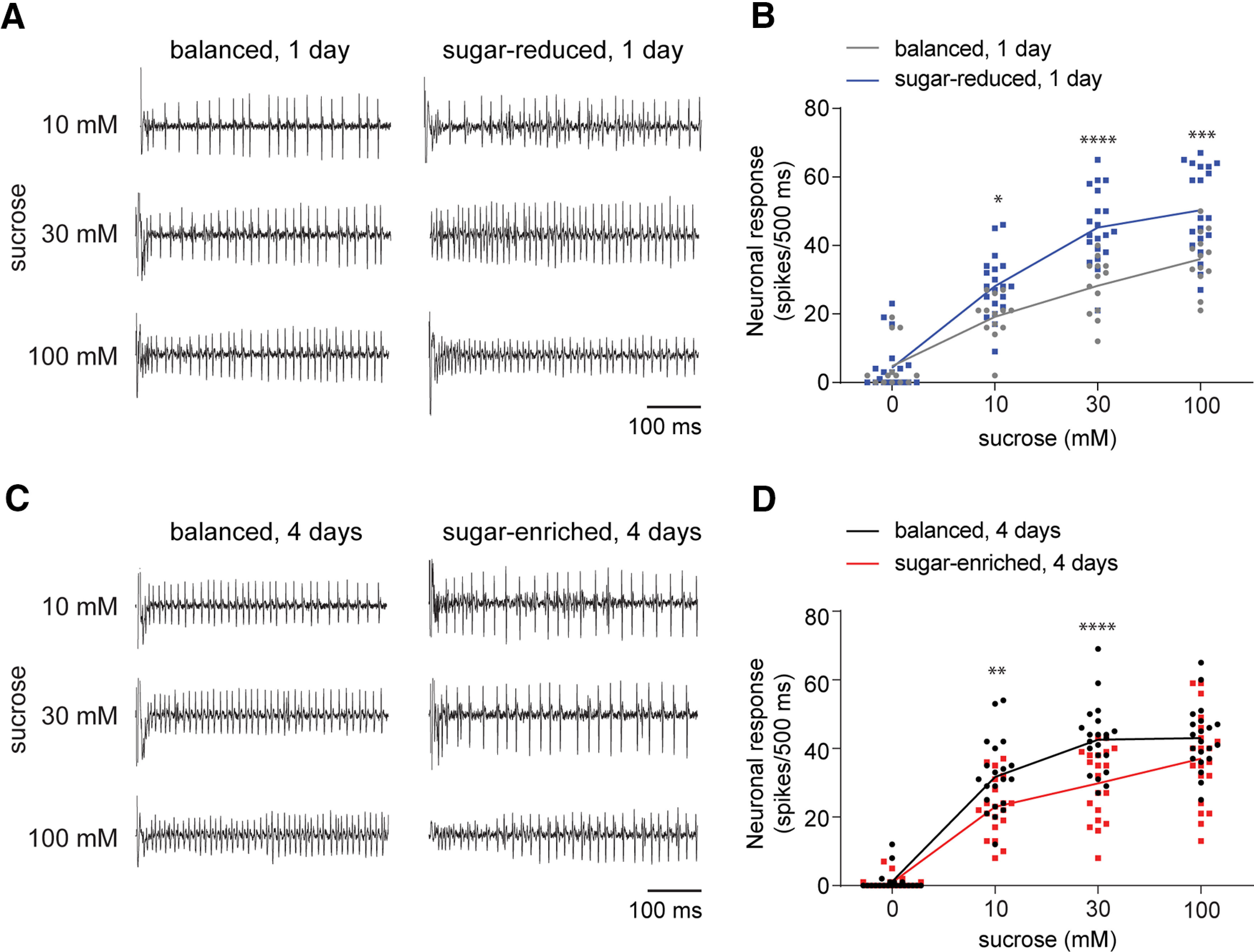
Dietary sugar imbalance alters sweet taste. A, Representative traces of the first 500-ms period of tip recordings from sensilla stimulated with the indicated concentrations of sucrose. Before testing, flies were prefed with indicated diet for 1 d. B, Neuronal responses in the first 500-ms period on stimulation with sucrose. Flies were prefed with the indicated diet for 1 d and tested without starvation (A, B) or after a 24-h period of wet starvation (Extended Data Fig. 2-1); n = 13 (balanced), n = 20 (sugar-reduced) sensilla from three to four flies. C, Representative traces of the first 500-ms period of tip recordings from sensilla stimulated with the indicated concentrations of sucrose. Before testing, flies were prefed with the indicated diet for 4 d. D, Neuronal responses in the first 500-ms period on stimulation with sucrose. Flies were prefed with the indicated diet for 4 d and tested without starvation (C, D) or after a 24-h period of wet starvation (Extended Data Fig. 2-2); n = 21 (balanced), n = 20 (sugar-enriched) sensilla from four flies. All recordings were taken from L-type labellar sensilla in mated females. Neuronal responses in B, D were compared using two-way ANOVA with Sidak's post hoc multiple comparisons test; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Representative traces (left) and mean responses (right) to sucrose in flies prefed as indicated for 1 d followed by starvation for 1 d; they were given access to water during the starvation period. Recordings were taken from L-type labellar sensilla in mated females, and action potentials in the first 500-ms period after contact were counted; n = 11 (balanced), n = 13 (sugar-reduced) sensilla from four flies each; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 2-1, TIF file (7MB, tif) .
Representative traces (left) and mean responses (right) to sucrose in flies prefed as indicated for 4 d followed by starvation for 1 d; they were given access to water during the starvation period. Recordings were taken from L-type labellar sensilla in mated females, and action potentials in the first 500-ms period after contact were counted; n = 16 (balanced), n = 13 (sugar-reduced) sensilla from four flies each; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 2-2, TIF file (7.6MB, tif) .
In complementary experiments, flies were fed on the sugar-enriched diet for 4 d and tested in parallel with age-matched control flies fed on the balanced diet. In this case, we observed a reduction in responses of labellar sweet taste neurons to sucrose, with significant differences observed between responses to 10 and 30 mm sucrose, but not to 100 mm sucrose (Fig. 2C,D, w1118 balanced vs sugar-enriched diet, p = 0.0093 for 10 mm sucrose, p < 0.0001 for 30 mm sucrose, p = 0.0.1411 for 100 mm sucrose; Sidak's post hoc multiple comparisons test; see also Table 3), indicating that response threshold but not peak response is altered. Overall, we found that taste sensitivity to sucrose was heightened by exposure to the sugar-reduced diet and dampened by the sugar-enriched diet. Since flies are starved for 24 h before binary choice assays, we verified that the changes in taste responses were associated with the specific diet rather than with starvation by recording sucrose responses from flies that were starved for 24 h after being prefed on the sugar-reduced or sugar-enriched diets (Extended Data Fig. 2-1, w1118 balanced vs sugar-reduced diet, p = 0.0212 for 10 mm sucrose, p = 0.0069 for 30 mm sucrose, p = 0.5976 for 100 mm sucrose and Extended Data Fig. 2-2, w1118 balanced vs sugar-reduced diet, p = 0.001 for 10 mm sucrose, p < 0.0001 for 30 mm sucrose, p = 0.1023 for 100 mm sucrose; Sidak's post hoc multiple comparisons test; see also Table 3). Overall, the diet-induced changes in taste sensitivity aligned with the observed shifts in behavioral preference in short-term choice assays, supporting the idea that behavioral modification in response to dietary imbalances occurs, at least in part, via alteration of taste sensitivity.
Diet-induced changes in sweet taste are persistent but reversible
Since modifications in taste sensitivity to sucrose were apparent only after a day or more following transfer of flies to test diets, we wanted to assess the extent to which the observed changes persisted on return to a balanced diet. To address this, we prefed flies on the sugar-reduced or sugar-enriched diet for 1 or 4 d, respectively, which were the time frames at which increases or decreases in sugar sensitivity were observed (Fig. 1B,E). Flies were then returned to the balanced diet and behavioral preference for sugar was tested daily with batches of flies. We found that the increase in sugar preference produced by the sugar-reduced diet persisted for at least a day following reintroduction of the balanced diet, returning by 2 d to levels observed for flies fed continuously on the balanced diet (Fig. 3A, w1118 balanced → balanced vs sugar-reduced → balanced; 1 d p < 0.0001, 2 d p = 0.03; Sidak's post hoc multiple comparisons test; see also Table 3). Consistent with these observations, we found that firing rates of labellar sweet taste neurons remained elevated at the higher concentrations tested (30 and 100 mm) for 1 d after resuming the balanced diet as compared with those obtained from control flies (Fig. 3B,C, w1118 balanced 1 d vs sugar-reduced 1 d, p = 0.0004 for 10 mm sucrose, p < 0.0001 for 30 mm sucrose, p < 0.0001 for 100 mm sucrose; w1118 balanced 2 d vs sugar-reduced 1 d + balanced 1 d, p = 0.01 for 30 mm sucrose, p = 0.0007 for 100 mm sucrose; w1118 balanced 3 d vs sugar-reduced 1 d + balanced 2 d, p = 0.0264 for 100 mm sucrose; Tukey's post hoc multiple comparisons test; see also Table 3).
Figure 3.
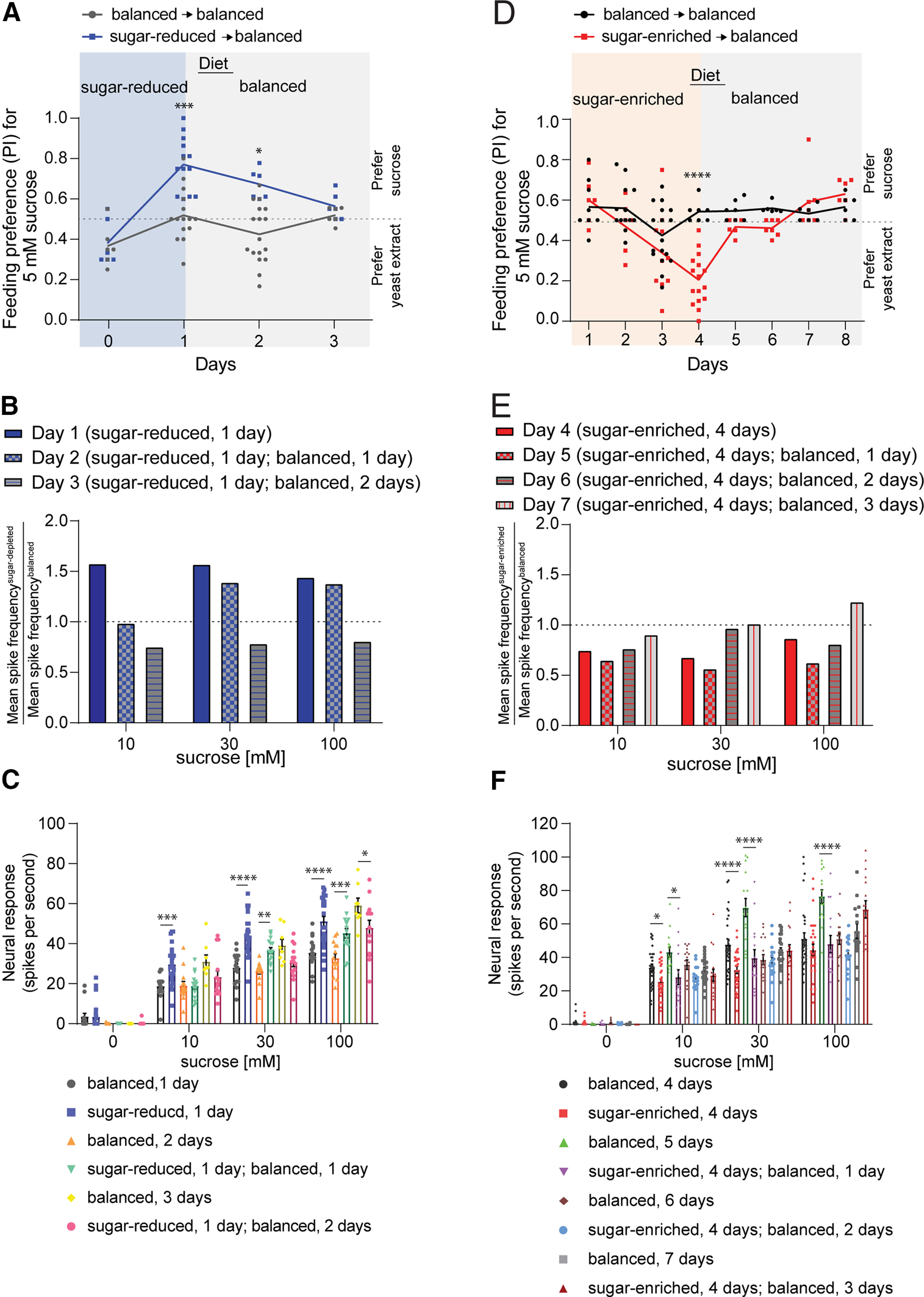
Dietary sugar-protein imbalance-induced alterations in sweet taste and behavior are reversible. A, Preference of mated females for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays. Flies were treated to indicated dietary regimens and tested daily; n = 6 balanced, 6 sugar-reduced (0 d); n = 15 balanced, 13 sugar-reduced (1 d); n = 17 balanced, 6 sugar-reduced (2 d); n = 6 balanced, 6 sugar-reduced (3 d). B, Ratio of mean sweet neuron response in flies fed on sugar-reduced diet (test) to that from flies fed on balanced diet (control). Flies were treated to different dietary regimens as in A and tested on indicated days with different concentrations of sucrose; n = 8–25 sensilla from two to five flies. The dotted line indicates a ratio of 1, i.e., equal taste sensitivity to sucrose in test and control flies. Tip recordings were taken from L-type labellar hairs in mated females. C, Mean neuronal responses used to generate the spike ratios shown in B. D, Preference of mated females for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays. Flies were treated to indicated dietary regimens and tested daily; n = 9 balanced, 6 sugar-enriched (1 d); n = 13 balanced, 6 sugar-enriched (2 d); n = 17 balanced, 10 sugar-enriched (3 d); n = 7 balanced, 16 sugar-enriched (4 d); n = 7 balanced, 6 sugar-enriched (5 d); n = 6 balanced, 7 sugar-enriched (6 d); n = 6 balanced, 7 sugar-enriched (7 d); n = 6 balanced, 6 sugar-enriched (8 d). E, Ratio of mean sweet neuron response in flies fed on sugar-enriched diet (test) to that from flies fed on balanced diet (control). Flies were treated to different dietary regimens as in C and tested on indicated days with different concentrations of sucrose; n = 15–29 sensilla from three to six flies. The dotted line indicates a ratio of 1, i.e., equal taste sensitivity to sucrose in test and control flies. F, Mean neuronal responses used to generate spike ratios shown in E. Data in A, D were analyzed using two-way ANOVA with Sidak's post hoc multiple comparisons test; data in C, F were analyzed using two-way ANOVA with Tukey's post hoc multiple comparisons test; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
In similar experiments with flies fed on the sugar-enriched diet, we found that feeding preference for sucrose bounced back quickly on resuming the balanced diet (Fig. 3D, w1118 balanced → balanced vs sugar-enriched → balanced; 4 d p < 0.0001, 5 d p = 0.9312; Sidak's post hoc multiple comparisons test; see also Table 3). However, sucrose response in labellar sweet taste neurons remained lower than that of flies fed continuously on the balanced diet, rising to comparable levels only after 2 d of recovery on the balanced diet (Fig. 3E,F, w1118 balanced 4 d vs sugar-enriched 4 d, 10 mm sucrose p = 0.0365, 30 mm sucrose p < 0.0001; w1118 balanced 5 d vs sugar-enriched 4 d + balanced 1 d, p = 0.0266 for 10 mm sucrose, p < 0.0001 for 30 mm sucrose, p < 0.0001 for 100 mm sucrose; Tukey's post hoc multiple comparisons test; see also Table 3). Together, the results suggest that dietary sugar-to-protein ratio alters sweet taste in a manner that can persist for some time after recommencement of the balanced diet. This is in agreement with a previous study, which reported that the increased preference for yeast in yeast-deprived flies disappears once the flies are reintroduced to a more complete diet (Ribeiro and Dickson, 2010; Vargas et al., 2010; Ganguly et al., 2017). Moreover, in both instances of recovery we observed a “rebound” effect of overcorrected neuronal firing rate in response to sucrose, at some part of the concentration range tested. Although analogous effects were not observed in binary choice assays, it remains possible that behavioral rebounds might be observed via other assays.
Dietary sugar-to-protein ratio alters feeding preference for amino acids
Since the proportion of protein also varies in the sugar-reduced and sugar-enriched diets we tested whether these diets affect feeding preference for amino acids. Flies were fed on sugar-reduced (protein-enriched) diet for 1 d and subsequently evaluated for feeding preference for an appetitive mixture of three amino acids (serine, phenylalanine, threonine) in binary choice assays (Ganguly et al., 2017). To promote feeding, 2 mm sucrose was added to both tastant alternatives, water and amino acid mixture, respectively. We found that flies fed on the sugar-reduced/protein-enriched diet had a lower preference for amino acids in comparison with those fed on the balanced diet (Fig. 4A, w1118 balanced vs sugar-reduced, 1 d, p = 0.0043, Mann–Whitney test), consistent with the idea that the sugar-protein imbalance affects taste sensitivity to sugar as well as to amino acids in opposing ways. The apparent aversion for the sucrose-amino acid mixture was somewhat surprising. Previous studies have found that certain amino acids, including phenylalanine, can elicit firing from S-type hairs that house bitter taste neurons (Park and Carlson, 2018), which may result in feeding avoidance when combined with a reduction in appetitive value.
Figure 4.
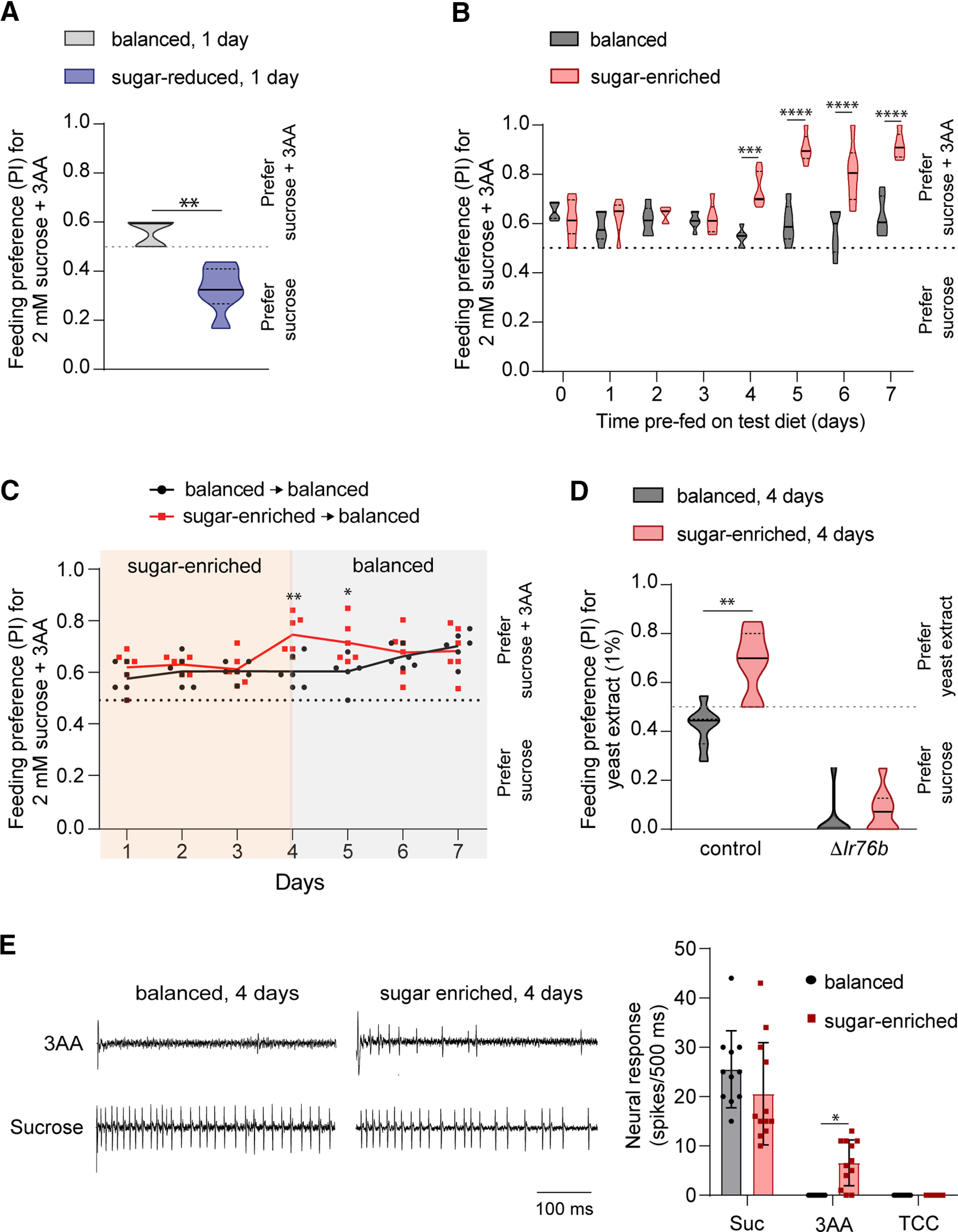
Dietary sugar-protein imbalance alters feeding preference for amino acids. A, Preference of mated females prefed on indicated diet for 1 d for 2 mm sucrose mixed with 25 mm each of serine, phenylalanine, and threonine (pink dye, sucrose + 3AA) tested against 2 mm sucrose alone (blue dye) in binary choice assays; n = 5 (balanced), n = 6 (sugar-reduced). B, Preference of mated females fed on indicated diet for 2 mm sucrose mixed with 25 mm each of serine, phenylalanine, and threonine (pink dye, sucrose + 3AA) tested against 2 mm sucrose alone (blue dye) in binary choice assays. Flies were tested daily on start of the indicated diet; n = 6 balanced (0–3 d), 10 (4 d), 6 (5–7 d); n = 6 sugar-enriched (0–3 d), 7 (4 d), 6 (5–7 d). C, Preference of mated females for 2 mM sucrose mixed with 25 mM each of serine, phenylalanine, and threonine (pink dye, sucrose + 3AA) tested against 2 mM sucrose alone (blue dye) in binary choice assays. Flies were treated to indicated dietary regimens and tested daily; n = 6 for each condition. D, Preference of mated females of control (w1118) and ΔIr76b (Ir76b1) genotypes prefed on the indicated diet for 4 d for 1% yeast extract (pink dye) tested against 5 mm sucrose (blue dye) in binary choice assays; n = 7 balanced, 7 sugar-enriched (control), n = 10 balanced, 9 sugar-enriched (ΔIr76b). E, Representative traces of the first 500-ms period of tip recordings from sensilla stimulated with a 3AA mixture of serine (100 mm), threonine (100 mm), and phenylalanine (25 mm) or 100 mm sucrose (left). Before testing, flies were prefed with the indicated diet for 4 d. Neuronal responses in the first 500-ms period on stimulation with indicated tastant or 30 mm tricholine citrate electrolyte (TCC; right). Flies were prefed with the indicated diet for 4 d; n = 11 (balanced), n = 12 (sugar-enriched) from five to six flies. For all graphs, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Statistical comparisons were made using the Mann–Whitney test (A, D) or two-way ANOVA with Sidak's post hoc multiple comparisons test (B, C, E).
Likewise, flies fed on sugar-enriched/protein-depleted diet were tested daily in choice assays and showed an enhancement of amino acid feeding preference at 4 d after starting the test diet, they preferred a mixture of amino acids and sucrose over sucrose alone as compared with flies fed on the balanced diet (Fig. 4B, w1118 balanced vs sugar-enriched, 4 d p = 0.0003, 5 d p < 0.0001, 6 d p < 0.0001, 7 d p < 0.0001; Sidak's post hoc multiple comparisons test; see also Table 3). In fact, the increased amino acid preference persisted for a day after flies were returned to a balanced diet (Fig. 4C, w1118 balanced → balanced vs sugar-enriched → balanced, 4 d p = 0.0018, 5 d p = 0.0283; Sidak's post hoc multiple comparisons test; see also Table 3). Moreover, flies fed with sugar-enriched/protein-depleted diet showed an increased preference for whole yeast extract, which was dependent on the function of Ir76b (Fig. 4D, balanced vs sugar-enriched diet w1118 p = 0.0017, Ir76b1 p = 0.0851, Mann–Whitney test), a receptor required for feeding behavior to amino acids as well as other components in yeast (Croset et al., 2016; Hussain et al., 2016; Ganguly et al., 2017; Steck et al., 2018). Taken together, the results suggest that behavioral shifts to accommodate changes in protein hunger are caused, at least in part, by altered feeding preference for amino acids.
A previous study showed that proboscis extension in response to stimulation of labellar hairs with amino acids was increased with dietary protein deprivation (Toshima and Tanimura, 2012). We therefore wished to test whether labellar taste responses to amino acids, which are ordinarily weak (Dahanukar et al., 2007; Ganguly et al., 2017; Park and Carlson, 2018), are strengthened on prefeeding with the sugar-enriched/protein-depleted diet. As reported previously, we observed no response to a mixture of three amino acids (100 mm serine, 100 mm threonine, and 25 mm phenylalanine) in L-type sensilla of flies fed on the balanced diet (Fig. 4E). A small but significant increase in the response was observed when flies were fed on the sugar-enriched/protein-depleted diet (Fig. 4E, w1118 balanced vs sugar-enriched 4 d, 100 mm sucrose p = 0.1164, 3AA mixture p = 0.0215; Sidak's post hoc multiple comparisons test; see also Table 3). Thus, the sugar-enriched/protein-depleted diet appears to have opposing effects on taste sensitivity of sucrose and amino acids that correlate with reduced feeding preference for the former and increased feeding preference for the latter.
Increased sweet taste sensitivity following dietary sugar reduction depends on Dop2R
To begin to understand the molecular mechanisms underlying diet-induced alterations in taste sensitivity, we first focused on candidate genes that are known to be involved in modulating feeding behavior responses on the basis of internal nutrient state. Many recent studies have identified such functions for the dopamine pathway (Okamoto and Nishimura, 2015; Post and Tatar, 2016), and a requirement for Dop2R has been uncovered in responses to starvation and protein hunger (Liu et al., 2017; Marella et al., 2012). Since the sugar-reduced/protein-enriched diet altered both sweet taste and amino acid preference, we sought to test whether one or both of these changes were mediated via Dop2R. With qRT-PCR analysis, the difference in Dop2R expression between flies fed on the balanced or test diet for 1 d was not statistically significant, but there was a trend toward lower Dop2R expression in the brains of flies prefed with the sugar-reduced diet (Extended Data Fig. 5-1, balanced vs sugar-reduced Dop2R expression p = 0.324177, independent sample t test). We therefore knocked down Dop2R pan-neuronally using the elav-GAL4 driver and two independent transgenic UAS-Dop2R RNAi lines (Extended Data Fig. 5-2). UAS-Dcr2 was also included in the genetic background. Flies were exposed to the sugar-reduced/protein-enriched diet as described above and tested in binary choice assays using sucrose and yeast extract as the two tastants. The baseline preference for sucrose was different in the two elav > Dop2R RNAi lines. elav> Dop2R RNAi1 showed a similar baseline sucrose preference as the GAL4 control, whereas elav>Dop2R RNAi2 exhibited an enhanced baseline sucrose preference (Table 3). The elav > Dop2R RNAi1 line had lost any diet-induced behavioral shift that was observed in control elav-GAL4 flies tested in parallel; this was also the case for elav > Dop2R RNAi2, but it is possible that the high baseline preference of these flies does not leave significant room for enhancement (Fig. 5A, balanced sugar reduced diet for 1 d, elav-GAL4 ctrl p = 0.0008, elav>Dop2R RNAi1 p = 0.3067, elav>Dop2R RNAi2 p = 0.7995; Sidak's post hoc multiple comparisons test; see also Table 3). We therefore selected Dop2R RNAi1 for additional experiments.
Figure 5.

Dop2R is required for sugar-deprivation induced rise in sweet taste and preference. A, Preference of mated females prefed on indicated diet for 1 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as follows: elav-Gal4 (n = 6 balanced, 7 sugar-reduced), elav-Gal4>UAS-Dop2R RNAi1 (elav>D2R RNAi1, n = 9 balanced, 9 sugar-reduced), and elav-Gal4>UAS-Dop2R RNAi2 (elav>D2R RNAi2, n = 6 balanced, 6 sugar-reduced). Levels of Dop2R expression were measured by reverse transcriptase-PCR in wild-type females fed on balanced or sugar-reduced diet for 1 d (see Extended Data Fig. 5-1) and in elav-Gal4>UAS-Dop2R RNAi1 flies (see Extended Data Fig. 5-2). B, Feeding preference and participation (see Extended Data Fig. 5-3) of mated females prefed on indicated diet for 1 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as follows: w1118 (control, n = 6 balanced, 6 sugar-reduced), BDSC#52025 (Dop2R1, n = 6 balanced, 6 sugar-reduced) and BDSC#52517 (Dop2R2, n = 10 balanced, 10 sugar-reduced). The observed shift in food choice differs from a starvation-induced effect (see Extended Data Figs. 5-4, 5-5). C, Preference of mated females prefed on indicated diet for 4 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were w1118 (control, n = 5 balanced, 5 sugar-enriched), BDSC#52025 (Dop2R1, n = 6 balanced, 6 sugar-enriched), and BDSC#52517 (Dop2R2, n = 6 balanced, 6 sugar-enriched). D, Preference of mated females prefed on indicated diet for 1 d and tested in binary choice assays with 2 mm sucrose mixed with 25 mm each of serine, phenylalanine, and threonine (sucrose + 3AA, pink dye) against 2 mm sucrose alone (blue dye). Genotypes were w1118 (control, n = 6 balanced, 6 sugar-reduced), BDSC#52025 (Dop2R1, n = 9 balanced, 9 sugar-reduced), and BDSC#52517 (Dop2R2, n = 6 balanced, 6 sugar-reduced). E, Representative traces obtained from Dop2R1 mutant females (left) and ratio of mean sweet neuron response (right) of flies fed on sugar-reduced diet (test) to that from flies fed on balanced diet (control) for 1 d. Mean neuronal responses are shown in Extended Data Figure 5-6. Recordings were taken from labellar L-type sensilla of five to six flies for each genotype. Genotypes tested were w1118 (control, n = 25–29) and BDSC#52025 (Dop2R1, n = 22–23). The dotted line indicates a ratio of 1, i.e., equal taste sensitivity to sucrose in test and control flies. Data in A–D were analyzed using two-way ANOVA with Sidak's post hoc multiple comparisons test, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Expression of Dop2R RNAi in Gad1-GAL4 neurons but not in Gr64f-GAL4 or other classes of neurons phenocopied the Dop2R mutant (see Extended Data Figs. 5-7, 5-8, 5-9, 5-10, 5-11).
Relative normalized expression levels of Dop2R in the brain of flies fed on indicated diet for 1 d; n = 3 biological replicates. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC. Primers for Dop2R are 5′-CTGAACTGCACCAACGAGACGC and 5′-CAGGATGTTGCCGAAGAGGGTC. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Graph shows mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 5-1, TIF file (2.4MB, tif) .
Image of agarose gel electrophoresis of RT-PCR samples using RNA derived from heads of 50–60 flies of the indicated genotypes and primers designed to amplify synaptotagmin (syt) or Dop2R sequences. Genotypes were as following: elav-GAL4; UAS-Dcr2 (elav-GAL4); UAS-Dop2R RNAi1, BDSC 26001 (UAS-Dop2R RNAi1); and elav-GAL4 > Dop2R RNAi1. Lane 1 was loaded with Generuler 1 Kb Plus (Thermo Fisher SM1331). Primers for syt were as following: 5′-TCTGGTCGTGCTTCGAGAAG and 5′-CGGATCCCTATGTCAAGGTG. Primers for Dop2R were 5′-ACTCGGTGGACTTGTCGTTC and 5′-CACCGAATTCACGCCAAGCACAAAAAT. Download Figure 5-2, TIF file (3.8MB, tif) .
Participation of mated females fed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays. w1118 (control, n = 6 balanced, 6 sugar-reduced), BDSC#52025 (Dop2R1, n = 6 balanced, 6 sugar-reduced), and BDSC#52517 (Dop2R2, n = 10 balanced, 10 sugar-reduced); *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 5-3, TIF file (2.2MB, tif) .
Starvation resistance of mated females that were given water alone or the sugar-reduced diet for 1 d and then subjected to starvation with access to water. Statistical comparison was done by the Log-rank (Mantel–Cox) test, n = 60 flies for each condition. Download Figure 5-4, TIF file (3.3MB, tif) .
Table comparing mortality of mated females that were wet-starved or fed on the sugar-reduced diet; n = 60 flies for each condition. Download Figure 5-5, TIF file (1.9MB, tif) .
Neuronal responses to various concentrations of sucrose obtained from mated females fed on the balanced or sugar-reduced diets for 1 d. Tip recordings were taken from labellar L-type sensilla. Genotypes were w1118 (control) and BDSC#52025 (Dop2R1); n = 25–29 sensilla from five to six flies (control) and n = 22–23 sensilla from five to six flies (Dop2R1); *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA with Tukey's post hoc multiple comparisons test. Download Figure 5-6, TIF file (3.7MB, tif) .
Preference of mated female flies fed on indicated diet for 1 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as following: BDSC#26001 UAS-Dop2R RNAi1 (D2R RNAi, n = 5 balanced, 5 sugar-reduced), Gad1-Gal4 > UAS-Dop2R RNAi (Gad1>D2R RNAi, n = 10 balanced, 10 sugar-reduced), and Gad1-Gal4 (n = 8 balanced, 8 sugar-reduced); *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 5-7, TIF file (2.8MB, tif) .
Confocal image of live GFP expression in Gad1-GAL4-labeled neurons in the brain. The image represents a z-projection of optical sections (1 µm) compressed in ImageJ using maximum intensity settings. Genotype: Gad1-GAL4; UAS-mCD8::GFP. The white dotted line represents the outline of the brain. Download Figure 5-8, TIF file (2.3MB, tif) .
Relative normalized expression levels of Dop2R in the brain following expression of Dop2R RNAi under the control of the Gad1-GAL4 driver. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC. Primers for Dop2R are 5′-CTGAACTGCACCAACGAGACGC and 5′-CAGGATGTTGCCGAAGAGGGTC. Graph shows mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 5-9, TIF file (2.2MB, tif) .
Preference of mated female flies expressing UAS-Dop2R RNAi in sweet taste neurons (left) or in selected subsets of neurons (right) that were prefed on indicated diet for 1 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as following: Gr64f-GAL4 (n = 14 balanced, 15 sugar-reduced), UAS-Dop2R RNAi (D2R RNAi, n = 7 balanced, 7 sugar-reduced), Gr64f-GAL4 > D2R RNAi (n = 6 balanced, 6 sugar-reduced), Tdc2 > D2R RNAi (n = 9 balanced, 10 sugar-reduced), Tdc2-GAL4 (n = 6 balanced, 6 sugar-reduced), Ple>D2R RNAi (n = 11 balanced, 11 sugar-reduced), Ple-GAL4 (n = 6 balanced, 6 sugar-reduced), TRH>D2R RNAi (n = 6 balanced, 6 sugar-reduced), TRH-GAL4 (n = 6 balanced, 6 sugar-reduced), v-Glut>D2R RNAi (n = 6 balanced, 6 sugar-reduced), v-Glut-GAL4 (n = 6, balanced, 6 sugar-reduced); *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 5-10, TIF file (6.5MB, tif) .
Neuronal responses to various concentrations of sucrose obtained from L-type labellar sensilla of mated females fed on indicated diet for 1 d. Genotypes were as following: BDSC#26001 UAS-Dop2R RNAi1, Gad1-Gal4 > UAS-Dop2R RNAi; n = 12–16 sensilla from three to four flies; *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, two-way ANOVA post-hoc Sidak's multiple comparisons test. Download Figure 5-11, TIF file (4.2MB, tif) .
To validate the observations with Dop2R RNAi, we next tested two different Dop2R mutants using the same experimental treatment and assays. Both mutants exhibited baseline sucrose preference similar to that of controls (Table 3). Consistent with the results of knock-down analysis, both failed to show increases in sucrose feeding preference on dietary sugar reduction (Fig. 5B, balanced vs sugar-reduced diet, w1118 p = 0.0075, Dop2R1 p = 0.9423, Dop2R2 p = 0.3635; Sidak's post hoc multiple comparisons test; see also Table 3). Loss of Dop2R did not affect participation of the flies in binary choice assays (Extended Data Fig. 5-3, balanced vs sugar-reduced diet, w1118 p > 0.9999, Dop2R1 p > 0.9999, Dop2R2 p = 0.6014; Sidak's post hoc multiple comparisons test; see also Table 3). However, Dop2R mutants retained the ability to adjust feeding preference in the appropriate direction on exposure to the sugar-enriched/protein-depleted diet (Fig. 5C, balanced vs sugar-enriched, w1118 p = 0.0019, Dop2R1 p < 0.0001, Dop2R2 p < 0.0001; Sidak's post hoc multiple comparisons test; see also Table 3), indicating a role for Dop2R in response to a specific dietary imbalance.
Importantly, since Dop2R has been found to be involved in response to starvation (Inagaki et al., 2012; Marella et al., 2012), we considered the possibility that flies feeding on the sugar-reduced diet were being starved. To probe this, we first determined whether flies were in fact consuming the sugar-reduced/protein-enriched food by including red dye in the diet. Flies were examined 1 d following transfer to the diet and all flies showed red dye in the abdomen (data not shown). Second, we compared starvation resistance of flies that had been subjected to 1 d of the sugar-reduced/protein-enriched diet versus water alone. No mortality was observed in the former group until ∼30–35 h of starvation. By contrast, individuals of the latter group began dying at ∼10–15 h of starvation (Extended Data Fig. 5-4, balanced vs sugar-reduced pretreatment p = 0.4661, Log-rank Mantel–Cox test; see also Table 3). Next, we compared survival rates on water (wet starvation) and the sugar-reduced/protein-enriched diet. While all the wet-starved flies succumbed in 4 d or less, the median survival time was ∼16 d for those fed on the sugar-reduced/protein-enriched diet (Extended Data Fig. 5-5). Together, these results support the idea that physiological outcomes of feeding on the sugar-reduced/protein-enriched diet are somewhat different from those of acute starvation. Thus, Dop2R-mediated changes in behavioral responses to sugar occur not only on starvation but also on exposure to a calorie-rich diet that is a poor source of sugar. Interestingly, the Dop2R mutants failed to lower preference for amino acids on feeding on the sugar-reduced/protein-enriched diet (Fig. 5D, balanced vs sugar-reduced diet, w1118 p = 0.0002, Dop2R1 p = 0.0993, Dop2R2 p = 0.3417; Sidak's post hoc multiple comparisons test; see also Table 3).
We next tested whether Dop2R is required for diet-induced alteration of sweet taste. We selected the Dop2R1 mutant for electrophysiological analysis and compared responses of labellar sweet neurons in flies fed with the balanced or sugar-reduced diet for 1 d. We found that the diet-induced enhancement of sucrose response was reduced in Dop2R mutants as compared with control flies; this effect was more pronounced for the lower concentrations that were tested (10 and 30 mm; Fig. 5E; Extended Data Fig. 5-6, w1118 balanced vs sugar-reduced, 10 mm sucrose p = 0.0017, 30 mm sucrose p < 0.0001, 100 mm sucrose p < 0.0001, Dop2R1 balanced vs Dop2R1 sugar-reduced, 10 mm sucrose p > 0.9999, 30 mm sucrose p = 0.0244, 100 mm sucrose p = 0.0002; Sidak's post hoc multiple comparisons test; see also Table 3). Overall, these results suggest that Dop2R plays a role in increasing sweet taste sensitivity on restriction of dietary sugar.
To identify the class of neurons in which Dop2R function is necessary for altering sweet taste following dietary sugar depletion, we expressed Dop2R RNAi in different subsets of neurons and compared feeding preferences of the resulting flies fed on balanced or sugar-reduced diets. Interestingly, expression of Dop2R RNAi via Gad1-GAL4, which labels GABAergic neurons, abolished the increase in sugar feeding preference produced by the sugar-reduced diet (Extended Data Fig. 5-7, balanced vs sugar-reduced diet, Gad1-GAL4>Dop2R RNAi p = 0.7004; Sidak's post hoc multiple comparisons test; see also Table 3). Both GAL4 and UAS controls, which were treated and tested in parallel, displayed diet-induced alterations in sugar preference as expected (Extended Data Fig. 5-7; Table 3). We observed Gad1-GAL4 to be widely expressed in the fly brain with particularly strong expression in the optic lobes and also in the protocerebral bridge (Extended Data Fig. 5-8). We did not find a significant reduction in Dop2R transcript levels in Gad1 > Dop2R RNAi flies using qRT-PCR analysis (Extended Data Fig. 5-9, Gad1-Gal4 vs Gad1 > Dop2R RNAi p = 0.6521, independent sample t test). One possible explanation is that there is only partial overlap between Dop2R and Gad1-GAL4 neurons, which obscures the effects of knock-down in the selected subset of cells.
Flies in which Dop2R RNAi was expressed in primary sweet taste neurons using Gr64f-GAL4 retained the ability to increase sugar preference on dietary sugar reduction (Extended Data Fig. 5-10, balanced vs sugar-reduced, Gr64f-GAL4 p < 0.0001, UAS-D2R RNAi1 p = 0.0057, Gr64f-GAL4>D2R RNAi1 p < 0.0001; Sidak's post hoc multiple comparisons test; see also Table 3). Knock-down of Dop2R in octopaminergic (Tdc2-GAL4), dopaminergic (Ple-GAL4), serotonergic (TRH-GAL4), or glutamatergic (v-Glut-GAL4) neurons also had little appreciable consequence on this phenotype (Extended Data Fig. 5-10, balanced vs sugar-reduced, Tdc2-GAL4>Dop2R RNAi p = 0.0128, Tdc2-GAL4 p = 0.0001, Ple>Dop2R RNAi p = 0.0026, Ple-GAL4 p < 0.0001, Trh>Dop2R RNAi p = 0.0002, Trh-GAL4 p = 0.0219, v-Glut>Dop2R RNAi p < 0.0001, v-Glut-GAL4 p = 0.0276; Sidak's post hoc multiple comparisons test; see also Table 3), although we cannot fairly evaluate partial contributions with these assay conditions. Consistent with the results of behavioral experiments, the differences in sweet taste neuron response to sucrose between flies fed on balanced versus sugar-reduced diet were greatly diminished or abolished in Gad1 > Dop2R RNAi flies, as compared with UAS-Dop2R RNAi controls tested in parallel (Extended Data Fig. 5-11, UAS-D2R RNAi balanced vs sugar-reduced, 10 mm sucrose p = 0.0027, 30 mm sucrose p = 0.0164, 100 mm sucrose p = 0.0004; Gad1-GAL4>UAS-D2R RNAi balanced versus sugar-reduced, 10 mm sucrose p = 0.0993, 30 mm sucrose p = 0.2012, 100 mm sucrose p = 0.1977; Sidak's post hoc multiple comparisons test; see also Table 3). Since the Gad1-GAL4 driver labels numerous neurons in the brain, the precise subsets of neurons in which Dop2R is required remain to be identified, but our results raise the possibility that GABAergic neurons in the brain are involved.
Transcriptome analysis identifies diet-induced changes in gene expression in taste tissues and the brain
To understand the molecular changes that accompany the observed physiological and behavioral shifts, we performed RNA-seq from various tissues and compared flies fed on the sugar-enriched/protein-depleted diet to flies fed on the balanced diet for 4 d. We analyzed the brain transcriptome, as well as that of tissues housing the majority of peripheral taste neurons, the proboscis and legs. In the brain, we found just over 200 genes that were differentially expressed, 120 of which were higher in flies exposed to the sugar-enriched diet and 93 that were lower (Fig. 6A, left). In the peripheral tissues, however, we found a disproportionately larger number of genes that showed reduced expression in flies given the sugar-enriched diet. In the proboscis, only 31 genes were significantly upregulated with feeding on the sugar-enriched diet, whereas 430 were downregulated (Fig. 6A, middle). Similarly, transcript levels of 145 genes increased, while those of 664 genes were significantly lower in the legs of flies fed on the sugar-enriched diet (Fig. 6A, right). Comparison of the DEGs revealed little overlap in upregulated genes between the three tissues (Fig. 6B, left). By contrast, we found significant overlap in the downregulated genes, 18 of which were decreased in all three tissues (Fig. 6B, right, C; Table 4). Among the commonly downregulated genes is Fit, which encodes the female-specific independent of transformer (FIT) peptide. Fit expression was previously shown to increase with protein intake in female flies, and it functions as a hormone to signal protein satiety (Sun et al., 2017). Consistent with these findings, we found a decrease in Fit expression in the brain, proboscis, and legs in female flies fed on sugar-enriched/yeast-depleted diet (Table 4). Interestingly, we also found that the expression of many genes encoding yolk proteins and serum proteins was reduced across all three tissues, suggesting that this dietary imbalance has profound effects on fat body gene expression in females (Table 4). The most substantial overlap, however, occurred among genes downregulated in the proboscis and legs, suggesting that similar molecular changes occur in these peripheral taste tissues (Fig. 6B,C). Gene Ontology (GO)-enrichment analysis for the genes altered in the brains of flies fed on the sugar-enriched diet revealed an enrichment for “G-protein-coupled photoreceptor activity [GO:0008020]” (Fig. 6D, top). This group includes the seven rhodopsin family genes found in Drosophila, typically expressed in the retina at high levels and markedly reduced in the brain (Senthilan et al., 2019) We found that four rhodopsin genes were significantly increased, between 2- and 3-fold, following feeding on sugar-enriched diet. In addition, two molecular function terms, “hormone activity [GO:0005179]” and “chitin binding [GO:0008061]” were enriched in genes that were downregulated in the brains of flies fed on sugar-enriched diet (Fig. 6D, bottom). Given the importance of hormones in regulating feeding behavior, we examined the expression of these hormone genes in peripheral taste tissues. We found that neuropeptide genes amn and CCAP were altered in the proboscis and leg, whereas Nplp3 was reduced in all three tissues (Fig. 6E). Similarly, two of the eight insulin-like peptide genes, dilp3 and dilp5, were downregulated in the brain and proboscis of flies fed on sugar-enriched diet; expression in the legs was also reduced for these genes, although they did not meet our false discovery rate (FDR) cutoff (Fig. 6E). Since dilp2, dilp3, and dilp5 have been reported to be expressed only in the brain (Brogiolo et al., 2001; Ikeya et al., 2002; Okamoto and Nishimura, 2015), detection of reads in other tissues, albeit at very low levels, was a little surprising. The dilp5 transcriptome findings were supported by qRT-PCR analysis, which showed a significant reduction in expression in the brains of flies fed on the sugar-enriched diet for 4 d (Extended Data Fig. 6-1). qRT-PCR also showed a trend toward lower expression of dilp3 and dilp5 in the proboscis, although the differences were not significant (Extended Data Fig. 6-1, balanced vs sugar-enriched diet 4 d, proboscis, dilp3 p = 0.753426, dilp5 p = 0.680805, independent samples t test). Overall, our results indicate profound effects of dietary sugar-enrichment on gene expression in the brain and taste organs, with changes in transcript abundance for a variety of genes. Moreover, mechanisms of diet-induced reduction in gene expression appear to have many common targets in the three tissues that we examined.
Figure 6.
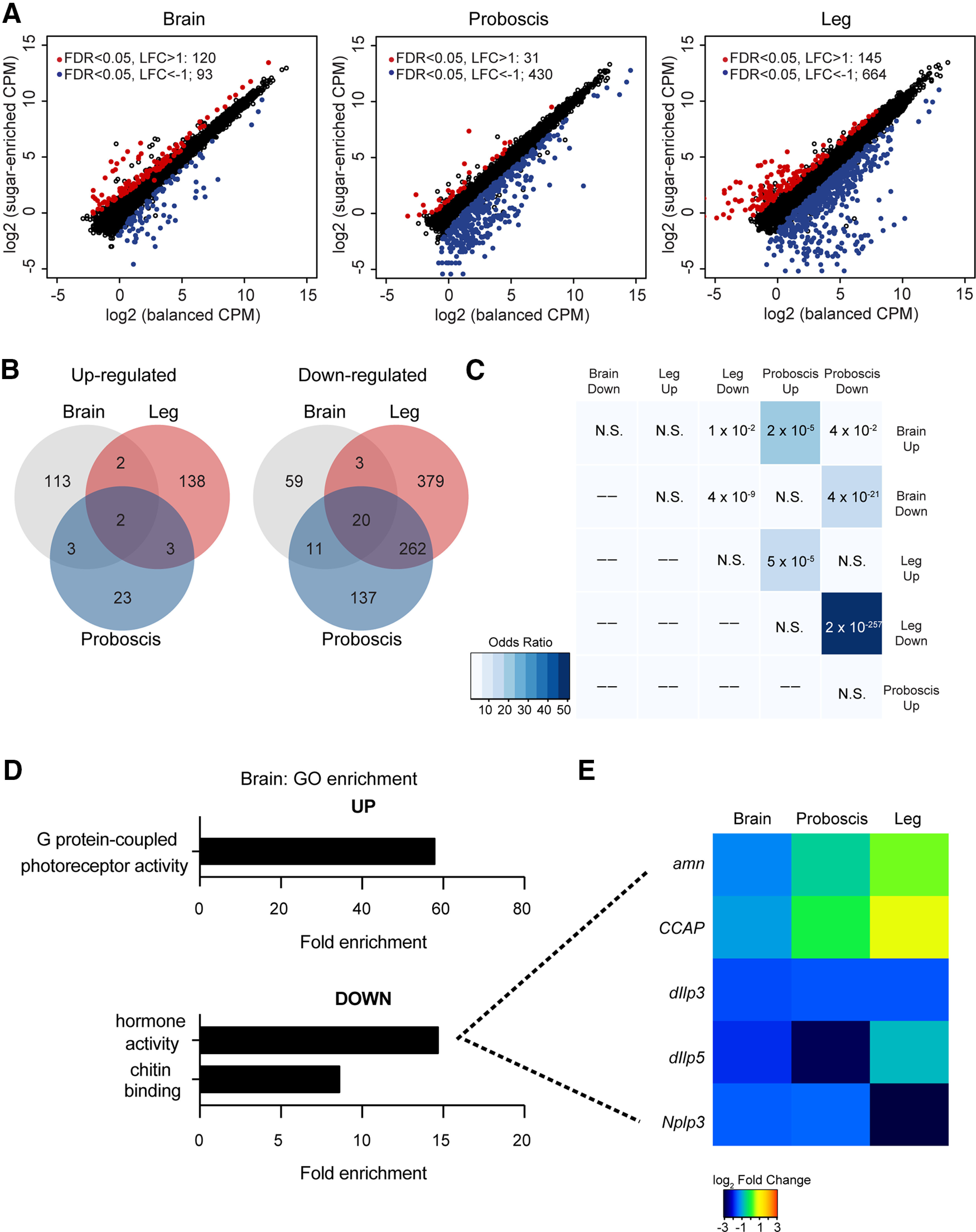
Gene expression analyses reveal a downregulation of insulin-like peptides (dILPs) on feeding on the sugar-enriched diet. A, Plots highlighting upregulated and downregulated genes in the brain, proboscis, and legs. Red and blue dots represent upregulated genes [Log2fold change (LFC) > 1, FDR < 0.05] and downregulated genes (LFC < −1, FDR < 0.05), respectively. B, Overlap of upregulated (left) and downregulated (right) genes in each of the three tissues. C, Significance of overlap of indicated gene sets (p values indicated in boxes; colors denote odds ratio from Fisher's exact test). D, Fold-enrichment for molecular function GO terms in genes upregulated and downregulated in the brain (p < 0.05). E, Heatmap depicting expression of brain-regulated genes associated with the “hormone activity” GO terms across all experiments. Each row represents the expression of one gene, normalized across samples (red = high expression, blue = low expression). All comparisons are relative to the balanced diet. Diet-induced changes in expression of dilp3 and dilp5 were tested by qRT-PCR (see Extended Data Fig. 6-1).
Table 4.
Common 18 downregulated genes in flies fed on a sugar-enriched diet
| Gene | Name | Log2fold change |
Function | ||
|---|---|---|---|---|---|
| Brain | Proboscis | Leg | |||
| Lsp2 | Larval serum protein 2 | −5.40 | −8.31 | −7.30 | Larval storage protein (LSP) which may serve as a store of amino acids for synthesis of adult proteins |
| Obp99a | Odorant-binding protein 99a | −3.87 | −5.46 | −3.40 | Present in the aqueous fluid surrounding olfactory sensory dendrites and are thought to aid in the capture and transport of hydrophobic odorants into and through this fluid |
| fit | Female-specific independent of transformer | −3.14 | −3.84 | −3.08 | - |
| Lsp1beta | Larval serum protein 1 beta | −2.90 | −4.62 | −3.65 | Larval storage protein (LSP) which may serve as a store of amino acids for synthesis of adult proteins |
| Yp2 | Yolk protein 2 | −2.62 | −3.04 | −3.10 | Vitellogenin is the major yolk protein of eggs where it is used as a food source during embryogenesis |
| Yp3 | Yolk protein 3 | −1.98 | −2.50 | −2.55 | Vitellogenin is the major yolk protein of eggs where it is used as a food source during embryogenesis |
| Cyp4g1 | Cytochrome P450- 4g1 | −1.94 | −1.34 | −1.29 | May be involved in the metabolism of insect hormones and in the breakdown of synthetic insecticides |
| Cyp12d1-p | Cyp12d1-p | −1.90 | −1.52 | −1.68 | - |
| Cp15 | Chorion protein 15 | −1.79 | −4.79 | −7.82 | Chorion membrane (egg shell) protein; plays a role in protecting the egg from the environment |
| tobi | Target of brain insulin | −1.74 | −3.46 | −2.92 | - |
| CG3699 | - | −1.57 | −1.66 | −2.93 | - |
| Nplp3 | Neuropeptide-like precursor 3 | −1.56 | −1.50 | −3.04 | - |
| CG10621 | - | −1.52 | −2.90 | −2.11 | - |
| CG3999 | - | −1.43 | −2.14 | −1.48 | - |
| dec-1 | Defective chorion 1 | −1.36 | −5.61 | −10.01 | Required for proper assembly of the eggshell |
| Vm26Aa | Vitelline membrane 26Aa | −1.36 | −4.71 | −8.72 | Major early eggshell protein |
| Vm34Ca | Vitelline membrane 34Ca | −1.35 | −4.84 | −9.19 | Major early eggshell protein |
| Yp1 | Yolk protein 1 | −1.22 | −1.78 | −1.99 | Vitellogenin is the major yolk protein of eggs where it is used as a food source during embryogenesis |
| CG4408 | - | −1.21 | −2.04 | −1.75 | - |
| CG34166 | - | −1.14 | −2.30 | −2.15 | - |
A total of 18 genes, including several that code for yolk and serum proteins, were found to decrease in the brain, proboscis, and legs of flies fed on the sugar-enriched diet. For each gene, fold change is represented as log (base2) ratios and FDR < 0.05.
Relative normalized expression levels of dilp3 and dilp5 in the brains (left) and proboscis (right) of flies fed on balanced or sugar-enriched diets for 4 d; n = 3 biological replicates. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC. Primers for dilp3 are 5′-GTCCAGGCCACCATGAAGTTGTGC and 5′-CTTTCCAGCAGGGAACGGTCTTCG. Primers for dilp5 are 5′-TGTTCGCCAAACGAGGCACCTTGG and 5′-CACGATTTGCGGCAACAGGAGTCG. Graphs show mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 6-1, TIF file (4.9MB, tif) .
Dilp5 is required in IPCs for diet-induced dampening of sweet taste
We focused on dilp5, whose transcripts were downregulated in the brain after flies were transferred to the sugar-enriched diet (Fig. 6E; Extended Data Fig. 6-1, balanced vs sugar-enriched diet 4 d, brain, dilp3 p = 0.680683, dilp5 p = 0.009507, independent samples t test). These findings are consistent with previous studies that report changes in dilp5 expression with altered protein or amino acid concentration in food (Manière et al., 2016). Incidentally Dilp2, Dilp3, and Dilp5 are secreted from the same group of cells, namely the IPCs in the brain. Moreover, a recent study has shown that IPCs can directly sense the amino acid leucine, and secrete Dilp2 and Dilp5 in response (Manière et al., 2016).
Based on our transcriptome analysis, we hypothesized that mutant flies lacking dilp5 that are fed on the balanced diet would behave as if they were fed on the sugar-enriched diet and exhibit reduced feeding preference for sugar as compared with control flies. Indeed, this is what we observed for dilp5 mutants, but not for dilp2 or dilp3 mutants (Fig. 7A; Extended Data Fig. 7-1, balanced vs sugar-enriched diet for 4 d w1118 p < 0.0001, dilp2 p < 0.0001, dilp3 p = 0.0035, dilp5 p = 0.4748; Sidak's post hoc multiple comparisons test; see also Table 3). We note that there were some apparent differences in baseline sucrose preference between the different genotypes, and in particular, dilp5 mutants had a lower mean preference for sucrose as compared with control flies or dilp2 and dilp3 mutants, whose behavior was identical to control flies. However, the differences in baseline preference across the genotypes were not statistically significant (Table 3). Moreover, dilp5 mutants participated to the same extent as controls in binary choice assays (Extended Data Fig. 7-2, participation in balanced vs sugar-enriched, w1118 p > 0.9999, dilp2 p = 0.973, dilp3 p = 0.9983, dilp5 p = 0.9848; Sidak's post hoc multiple comparisons test; see also Table 3), but were not able to significantly reduce sucrose feeding preference in response to the sugar-enriched diet; dilp2 and dilp3 mutants, however, retained the ability to do so (Fig. 7A). Interestingly, we found no differences in behavioral preference for sucrose between control and dilp5 mutants following exposure to the sugar-reduced diet, indicating a requirement for dilp5 only in adjusting behavioral responses to dietary sugar enrichment but not to sugar depletion (Fig. 7B, balanced vs sugar-reduced, w1118 p < 0.0001, dilp5 = 0.0008; Sidak's post hoc multiple comparisons test; see also Table 3). Consistent with these results, dilp5 mutants did not exhibit a diminished neuronal response to sugar after feeding on the sugar-enriched diet, as compared with wild-type flies that were co-tested (Fig. 7C,D; Extended Data Fig. 7-3, w1118 balanced vs w1118 sugar-enriched 4 d, 10 mm sucrose p = 0.03, 30 mm sucrose p < 0.0001, 100 mm sucrose p = 0.0653; dilp5 balanced vs dilp5 sugar-enriched, 10 mm sucrose p = 0.0174, 30 mm sucrose p = 0.9728, 100 mm sucrose p = 0.0006; Sidak's post hoc multiple comparisons test; see also Table 3). On the contrary, at least for some concentrations there was a slight increase in neuronal response to sucrose in the dilp5 mutants (Fig. 7D; Extended Data Fig. 7-3; Table 3). Interestingly, baseline levels of sweet neuron response and sugar preference of dilp5 mutants fed on balanced diet were lower than that of control flies (Fig. 7A,D; Extended Data Fig. 7-3; Table 3), suggesting that an optimum titer of dilp5 may be required to maintain sugar preference in flies.
Figure 7.

dilp5 is required for dietary sugar-enrichment-induced decline in sweet taste and preference. A, Preference of mated females prefed on indicated diet for 4 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were w1118 (control, n = 8 balanced, 8 sugar-enriched), dilp2 (n = 7 balanced, 8 sugar-enriched), dilp3 (n = 7 balanced, 6 sugar-enriched), and dilp5 (n = 7 balanced, 9 sugar-enriched). dilp3 and dilp5 mutants were verified by genomic PCR (see Extended Data Fig. 7-1). Feeding participation of females prefed on balanced or sugar-enriched diet is shown in Extended Data Figure 7-2. B, Preference of mated females prefed on indicated diet for 1 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were w1118 (control, n = 19 balanced, 17 sugar-reduced), dilp5 (n = 12 balanced, 12 sugar-reduced). C, Representative traces obtained from dilp5 mutant females fed on indicated diet for 4 d. D, Ratio of mean sweet neuron response of flies fed on sugar-enriched diet (test) to that from flies fed on balanced diet (control) for 4 d. Mean neuronal responses are shown in Extended Data Figure 7-3. Recordings were taken from labellar L-type sensilla in mated females. Genotypes were as follows: w1118 (control) and dilp5, n = 19–30 sensilla from four to six flies. The dotted line indicates a ratio of 1, i.e., equal taste sensitivity to sucrose in test and control flies. E, Preference of mated females prefed on indicated diet for 4 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as follows: dilp2-Gal4 (n = 9 balanced, 10 sugar-enriched), UAS-dilp5 RNAi (n = 6 balanced, 6 sugar-enriched), dilp2>dilp5 RNAi (n = 10 balanced, 12 sugar-enriched), UAS-TNTG (n = 9 balanced, 9 sugar-enriched), and dilp2>TNTG (n = 17 balanced, 17 sugar-enriched). Expression of dilp5 in dilp2>dilp5 RNAi flies was tested by qRT-PCR (see Extended Data Fig. 7-4). F, Preference of mated female flies prefed on indicated diet for 4 d and tested in binary choice assays with 2 mm sucrose mixed with 25 mm each of serine, phenylalanine, and threonine (sucrose + 3AA, pink dye) against 2 mm sucrose alone (blue dye). Genotypes were as follows: w1118 (control, n = 6 balanced, 6 sugar-enriched), dilp5 (8 balanced, 10 sugar-enriched). G, Preference of mated females prefed on indicated diet for 4 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as follows: w1118 (control, n = 9 balanced, 9 sugar-enriched), InR1 (BL11661)/InR2 (BL9646; n = 6 balanced, 6 sugar-enriched). H, Preference of mated females prefed on indicated diet for 4 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as follows: UAS-InR RNAi (n = 7 balanced, 6 sugar-enriched), elav-Gal4 (n = 9 balanced, 8 sugar-enriched), elav-Gal4 > UAS-InR RNAi (n = 6 balanced, 6 sugar-enriched). I, Preference of mated females prefed on indicated diet for 4 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as follows: UAS-InR RNAi (control n = 6 balanced, 6 sugar-enriched), v-Glut>UAS-InR RNAi (n = 7 balanced, 7 sugar-enriched), Gad1>UAS-InR RNAi (n = 7 balanced, 7 sugar-enriched), Ple>UAS-InR RNAi (n = 6 balanced, 7 sugar-enriched), Trh>UAS-InR RNAi (n = 8 balanced, 8 sugar-enriched), Tdc2>UAS-InR RNAi (n = 7 balanced, 7 sugar-enriched), npF>UAS-InR RNAi (n = 7 balanced, 7 sugar-enriched). Expression of InR in Trh-GAL4>UAS-InR RNAi flies was measured by qRT-PCR (see Extended Data Fig. 7-5). For all except C, D, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns - not significant, two-way ANOVA with Sidak's post hoc multiple comparisons test.
Image of agarose gel electrophoresis of PCR samples using genomic DNA templates from w1118 and dilp5 (left) or dilp3 (right) mutant flies. Lane 1 was loaded with Generuler 1 Kb Plus (Thermo Fisher SM1331). Primers for synaptotagmin (syt) were as in Extended Data Figure 5-2. Primers for dilp5 were as following: 5′-CGCCGAATGCTCGACAGTGA and 5′-CTCCGTGATCCCAGTTCTCCT. Primers for dilp3 were as following: 5′-GACTTCAGGCAACACTCGTC and 5′-ATCCTTATGATCGGCGGTGTC. Arrows mark positions of expected amplicons. Download Figure 7-1, TIF file (4.7MB, tif) .
Participation of mated females prefed on indicated diet for 4 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were w1118 (control, n = 8 balanced, 8 sugar-enriched), dilp2 (n = 7 balanced, 8 sugar-enriched), dilp3 (n = 7 balanced, 6 sugar-enriched), and dilp5 (n = 7 balanced, 9 sugar-enriched); *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 7-2, TIF file (2.1MB, tif) .
Neuronal responses to various concentrations of sucrose obtained from mated females fed on balanced or sugar-enriched diet for 4 d. Tip recordings were taken from labellar L-type sensilla. Genotypes were w1118 (control) and dilp5; n = 29–30 sensilla from five to six flies (control) and n = 19–21 sensilla from five to six flies (dilp5); *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA with Tukey's post hoc multiple comparisons test. Download Figure 7-3, TIF file (3.4MB, tif) .
We next expressed dilp5 RNAi specifically in the IPCs under the control of dilp2-GAL4. We chose dilp2-GAL4 since among the dilps, dilp2 had the most stable expression between balanced and test diet treatments. Although not statistically significant, we found a decrease in the mean relative expression level of dilp5 in the dilp2 > dilp5 RNAi flies (Extended Data Fig. 7-4, dilp5 expression in brain p = 0.360222, independent samples t test). Interestingly, the dilp2 > dilp5 RNAi flies phenocopied the dilp5 mutant, in terms of the failure to decrease sugar preference following feeding on the sugar-enriched diet (Fig. 7E, balanced vs sugar-enriched diet, dilp2-GAL4 p < 0.0001, UAS-dilp5 RNAi = 0.0007, dilp2>dilp5 RNAi= 0.4489, UAS-TNTG p = 0.0105, dilp2>TNTG p = 0.2572; Sidak's post hoc multiple comparisons test; see also Table 3). Inhibition of vesicle secretion in dilp2-GAL4 neurons by expressing tetanus toxin (TNTG) had the same effect (Fig. 7E), consistent with the expectation that secretion of Dilps from the IPCs is important for their functional roles. We next tested whether the increase in amino acid preference following exposure to the sugar-enriched/protein-depleted diet was dependent on dilp5. We observed similar increases in feeding preference for the three-amino acid mixture in dilp5 mutants and control flies (Fig. 7F, balanced vs sugar-enriched, w1118 p = 0.0109, dilp5 p = 0.0021; Sidak's post hoc multiple comparisons test; see also Table 3), suggesting that Dilp5 may be specifically required for altering behavioral responses to sugar, but not to amino acids, following dietary macronutrient imbalance.
Relative normalized expression levels of dilp5 in the brain following expression of dilp5 RNAi in dilp2-GAL4 neurons; n = 3 biological replicates. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC. Primers for dilp5 are 5′-TGTTCGCCAAACGAGGCACCTTGG and 5′-CACGATTTGCGGCAACAGGAGTCG. Graph shows mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 7-4, TIF file (2.1MB, tif) .
dilp5 acts via InR in serotonergic neurons to control sugar preference
Since InR is the only known receptor for Dilp5 we hypothesized that loss of InR would phenocopy the dilp5 mutant. We examined feeding preference of trans-heterozygous flies of two different InR mutant alleles, namely InR05545 (InR1) and InRE19 (InR2), such flies have been reported to have ∼90% reduction in InR transcript expression (Tatar et al., 2001). The baseline sucrose preference of the InR mutant flies was significantly lower than that of the control flies (Table 3), somewhat reminiscent of the trend observed for dilp5 mutants (Fig. 7A,F). As predicted, InR mutants failed to register a decrease in sugar preference after feeding on the sugar-enriched diet, whereas control flies that were tested in parallel did so (Fig. 7G, balanced vs sugar enriched, w1118 p < 0.0001, InR1/InR2 = 0.9753; Sidak's post hoc multiple comparisons test; see also Table 3). Pan-neuronal expression of InR RNAi was able to recapitulate the defect in sugar-enriched diet-induced alteration of sugar preference (Fig. 7H, balanced vs sugar-enriched, elav-GAL4>InR RNAi p = 0.999; Sidak's post hoc multiple comparisons test; see also Table 3). Like wild-type flies, both control lines were able to reduce sugar preference after feeding on the sugar-enriched diet (Fig. 7H, UAS-InR RNAi p = 0.0423, elav-GAL4 p = 0.0031; Sidak's post hoc multiple comparisons test; see also Table 3).
To identify the class of neurons in which InR activity is required for modulating sugar preference, we expressed InR RNAi in different subsets of neurons and examined feeding preference of the resulting flies after feeding on balanced or sugar-enriched diets. Similar to dilp5 and InR mutants, flies in which InR RNAi was expressed in Trh-GAL4 (Extended Data Fig. 7-5, brain InR p = 0.070619, independent samples t test) serotonergic neurons exhibited a lower baseline sugar preference when prefed on the balanced diet, and sugar preference was found at that same level regardless of whether the flies were given a balanced or sugar-enriched diet (Fig. 7I, balanced vs sugar-enriched, Trh>InR RNAi p = 0.8235; Sidak's post hoc multiple comparisons test; see also Table 3). UAS control flies, which were treated and tested in parallel, displayed a diet-induced shift in sugar preference as expected (Fig. 7I, balanced vs sugar-enriched, UAS-InR RNAi p < 0.0001, Sidak's post hoc multiple comparisons test; see also Table 3). As did flies in which InR RNAi was expressed in glutamatergic (v-Glut-GAL4), GABAergic (Gad1-GAL4), dopaminergic (Ple-GAL4), octopaminergic (Tdc2-GAL4), or npF (npF-GAL4) neurons (Fig. 7I, balanced vs sugar-enriched, v-Glut>InR RNAi p < 0.0001, Gad1>InR RNAi p = 0.0022, Ple>InR RNAi p = 0.0002, Tdc2>InR RNAi p < 0.0001, npF>InR RNAi p < 0.0001; Sidak's post hoc multiple comparisons test; see also Table 3). A previous study found that serotoninergic (Trh-GAL4) neurons and the serotonin receptor 5HT2A mediate increased protein feeding on protein starvation by altering relative value of protein in higher order brain neurons (Ro et al., 2016). Serotonergic neurons have also been shown to regulate IPCs via 5-HT(1A) receptors present on the IPCs (Luo et al., 2012). Combining our observations with these previous findings suggests the presence of a feedback loop between serotonergic neurons and IPCs to regulate protein feeding in flies.
Relative normalized expression levels of InR in the brain following expression of InR RNAi in Trh-GAL4 neurons; n = 3 biological replicates. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC′. Primers for InR are 5′-TGAGCATGTGGAGCACATCAAGATG and 5′-CGTAGGAGATTTTCTCGTTTGGCTG. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Graphs show mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 7-5, TIF file (2MB, tif) .
Discussion
Here, we compare cellular and behavioral responses to sugar in flies exposed to diets that vary in the amount of carbohydrate and protein. Our results support a model in which contrasting sugar-protein imbalances in the diet can activate independent signaling pathways that act either on sweet taste or amino acid taste or both to bring about compensatory changes in feeding behavior. Response to dietary carbohydrate deprivation is more rapid than that to protein deprivation, likely reflecting differences in energetic and physiological needs fulfilled by the two macronutrients. Notably, dietary imbalance results in behavioral shifts in food choice, but does not appear to have significant effects on consumption in the short term. Our results suggest that flies can regulate food choice to mitigate nutrient deficit or excess, and this occurs, at least in part, via regulation of taste sensitivity. Thus, primary taste neurons may be fairly plastic and can undergo changes to suit the physiological requirements of an organism.
A number of studies have investigated behavioral shifts caused by dietary macronutrient imbalance. For example, insect predators pretreated with protein-poor/lipid-rich or protein-rich/lipid-poor diets adjusted their food selection or consumption to compensate for lack of the specific macronutrient in the pretreatment diet (Mayntz et al., 2005). In locusts, compensatory changes in behavior have been linked to changes in sensitivity of peripheral taste neurons, i.e., increased sugar responsiveness in protein-fed animals and increased amino acid-responsiveness in carbohydrate-fed animals (Abisgold and Simpson, 1988; Simpson and Simpson, 1992). Similar studies in Drosophila show that dietary protein or amino acid deprivation increased feeding preference for protein or amino acids (Ribeiro and Dickson, 2010; Vargas et al., 2010; Toshima and Tanimura, 2012; Corrales-Carvajal et al., 2016; Leitão-Gonçalves et al., 2017; Steck et al., 2018). One study also reported an increase in proboscis extension responses to amino acids following amino acid deprivation for 6 d (Toshima and Tanimura, 2012), and recent calcium imaging experiments found that pharyngeal and peg taste neurons become more responsive to yeast on amino acid deprivation (Steck et al., 2018). More recently, longer term exposure to high-sugar diet has been shown to increase consumption of the diet, which is also dependent on diet-induced depression of sweet taste (May et al., 2019). Thus, for generalist foragers, nutrient diversity and availability could influence changes in peripheral neuron sensitivity, food choice and consumption in various ways.
We find a subset of genes that are differentially expressed on prolonged feeding on a sugar-enriched/protein-depleted diet in the brain and in the peripheral taste tissues of the female fly: the proboscis and legs. Although only subsets of genes in the Drosophila genome are altered with this dietary change in each tissue, we find a larger fraction of downregulated genes in the proboscis and legs. Surprisingly, we find no change in the expression of peripheral taste receptors (Grs and Irs) in the various taste tissues. Among the downregulated genes, however, we find Drosophila insulin-like peptide genes (dilps) that were altered in all three tissues. This includes dilp5, which our work shows to be necessary for compensatory responses to dietary sugar imbalance.
Comparing genes that changed in abundance in our study with those reported in other nutrient imbalance studies, we find only modest overlap. However, one study examined changes in gene expression with a whole-fly microarray performed using flies raised on a high-sugar, high-calorie diet (Liu et al., 2017). In the other, transcriptomes were analyzed from proboscis tissue of flies fed on a high-sugar (and high-calorie) diet for 7 d (May et al., 2019). The disproportionately large number of genes that we found downregulated in the proboscis was far more pronounced to the reports in this study, which found the number of genes reduced in expression only slightly higher than those increased in expression (May et al., 2019). The different results could be attributed to many factors, including differences in diet exposure time (days vs weeks) or in calorie intake. Nevertheless, we find 42 genes that overlap with the differentially regulated set identified by May et al. (2019), which merit further attention. Our finding that the recovery of baseline taste responses on return to the balanced diet occurs rapidly suggests also that mechanisms exist to take immediate advantage of nutrient-balanced food sources. An expanded time-course transcriptome analysis would reveal the extent to which such mechanisms depend on transcriptional regulation, and how they relate to changes in neurophysiological and behavioral responses.
An important role for dopamine signaling has emerged from studies investigating the effects of starvation on sugar feeding behavior. Sweet-sensing neurons were found to respond to dopamine via the DopEcR receptor, undergoing presynaptic potentiation consistent with a lower behavioral acceptance threshold for sucrose, particularly in the first 6 h after starvation (Inagaki et al., 2012). Another study found that starvation-induced increase in behavioral sensitivity to sucrose is mediated via the Dop2R receptor, although the neuronal populations in which it is required were not identified (Marella et al., 2012). The same study identified the dopaminergic TH-VUM neuron as a signaling source, and as expected its activity was observed to be modulated by satiety state (Marella et al., 2012). Interestingly, Dop2R was recently implicated in increased yeast preference on dietary protein deprivation (Liu et al., 2017). The study describes that protein deprivation caused an increase in yeast preference as well as a reduction in sugar preference, and a role for Dop2R was found for the former but not the latter. While we also observed that flies given a sugar-enriched/yeast-depleted diet compensated with opposite effects on sugar and amino acid preference, we found that Dop2R was dispensable for these shifts. One possible explanation for this is that Dop2R signaling is involved in protein hunger when the overall energetic requirements are not met. Our observation that Dop2R is required for modulation of both sweet and amino acid preference in response to the sugar-reduced/yeast extract-enriched diet lends support to this idea. Whether dopaminergic DA-WED neurons, which have been suggested to have antagonistic effects on sugar and amino acid feeding, are involved remains to be seen. Moreover, the identity of specific Dop2R neuronal substrates, and determination of whether the same or different sets of neurons regulates sugar and amino acid choice, will be interesting to pursue.
Insulin and insulin-like peptides have long been known to be regulated by nutrient status. Many studies have examined nutrient-dependent expression of dilps (Ikeya et al., 2002; Géminard et al., 2009; Sun et al., 2017; Lin et al., 2018) and subsequent regulation of nutrient uptake (Sun et al., 2017; Semaniuk et al., 2018). Various studies have shown that in both larvae and adult flies an increase or decrease in dietary yeast upregulates or downregulates dilp5, respectively (Géminard et al., 2009; Okamoto and Nishimura, 2015; Post and Tatar, 2016; Lin et al., 2018). Leucine intake has also been found to induce production and release of Dilp2 and Dilp5 (Manière et al., 2016). Additionally, dilp5 expression is controlled by dietary carbohydrates; a high-sucrose diet upregulates dilp5 mRNA in larvae (Pasco and Léopold, 2012), whereas high dietary fructose but not glucose downregulates dilp2 and dilp5 in the brain, leading to increases in food consumption and obesity (Rovenko et al., 2015). However, a specific effect on nutrient choice rather than intake has not been described previously. Our studies suggest that dilp5 is involved in preserving sweet taste sensitivity under standard dietary conditions, since loss of dilp5 phenocopies the effects of a sugar-enriched diet. Ultimately, the question of how sweet neuron sensitivity is altered remains unanswered. A straightforward possibility is that levels or activity of sweet taste receptors are modulated. However, at least at the transcriptional level we found no evidence for differences in expression of members of the sweet Gr clade in flies prefed the various diets, although one previous study found upregulation of Gr64a in starved flies (Nishimura et al., 2012). An alternative possibility is that dietary nutrient imbalance alters taste sensitivity by regulation of neuronal excitability. A recent study has shown that sensitization of sweet taste neurons on chronic exposure to unsweetened diet is controlled by dopaminergic pathways acting via Dop1R in sweet taste neurons and downstream cAMP/CREB pathways, eventually converging on PGC1α, which is necessary and sufficient for sweet neuron sensitization (Wang et al., 2020). Since sweet taste neurons have been found to respond to many other classes of tastants (Masek and Keene, 2013; Ganguly et al., 2017; Tauber et al., 2017; Jaeger et al., 2018), it would be interesting to determine whether dietary variation has precise effects on taste reception, or whether it broadly affects responses to all tastants that are detected by selected neurons.
Footnotes
This work was supported in part by the National Science Foundation Grant IOS–1149667 and the University of California-Riverside Agricultural Experimental Station and National Institute of Food and Agriculture–United States Department of Agriculture Hatch Project Grant 1011543. We thank B. Jablonska for technical help and members of the Dahanukar, Ray, and Yamanaka laboratories for helpful discussions. Stocks were obtained from the Bloomington Drosophila Stock Center (National Institutes of Health Grant P40OD018537) and N. Yamanaka.
The authors declare no competing financial interests.
References
- Abisgold JD, Simpson SJ (1988) The effect of dietary protein levels and haemolymph composition on the sensitivity of the maxillary palp chemoreceptors of locusts. J Exp Biol 135:215–229. 10.1242/jeb.135.1.215 [DOI] [Google Scholar]
- Barzel US, Massey LK (1998) Excess dietary protein can adversely affect bone. J Nutr 128:1051–1053. 10.1093/jn/128.6.1051 [DOI] [PubMed] [Google Scholar]
- Benton R, Dahanukar A (2011) Electrophysiological recording from Drosophila taste sensilla. Cold Spring Harb Protoc 2011:839–850. 10.1101/pdb.prot5631 [DOI] [PubMed] [Google Scholar]
- Bray NL, Pimentel H, Melsted P, Pachter L (2016) Erratum: near-optimal probabilistic RNA-seq quantification. Nat Biotechnol 34:888. 10.1038/nbt0816-888d [DOI] [PubMed] [Google Scholar]
- Brogiolo W, Stocker H, Ikeya T, Rintelen F, Fernandez R, Hafen E (2001) An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr Biol 11:213–221. 10.1016/s0960-9822(01)00068-9 [DOI] [PubMed] [Google Scholar]
- Corrales-Carvajal VM, Faisal AA, Ribeiro C (2016) Internal states drive nutrient homeostasis by modulating exploration-exploitation trade-off. Elife 5:e19920. 10.7554/eLife.19920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croset V, Schleyer M, Arguello JR, Gerber B, Benton R (2016) A molecular and neuronal basis for amino acid sensing in the Drosophila larva. Sci Rep 6:34871. 10.1038/srep34871 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahanukar A, Lei YT, Kwon JY, Carlson JR (2007) Two Gr genes underlie sugar reception in Drosophila. Neuron 56:503–516. 10.1016/j.neuron.2007.10.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devineni AV, Sun B, Zhukovskaya A, Axel R (2019) Acetic acid activates distinct taste pathways in to elicit opposing, state-dependent feeding responses. Elife 8:e47677. 10.7554/eLife.47677 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ganguly A, Pang L, Duong VK, Lee A, Schoniger H, Varady E, Dahanukar A (2017) A molecular and cellular context-dependent role for Ir76b in detection of amino acid taste. Cell Rep 18:737–750. 10.1016/j.celrep.2016.12.071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Géminard C, Rulifson EJ, Léopold P (2009) Remote control of insulin secretion by fat cells in Drosophila. Cell Metab 10:199–207. 10.1016/j.cmet.2009.08.002 [DOI] [PubMed] [Google Scholar]
- Hussain A, Zhang M, Üçpunar HK, Svensson T, Quillery E, Gompel N, Ignell R, Grunwald Kadow IC (2016) Ionotropic chemosensory receptors mediate the taste and smell of polyamines. PLoS Biol 14:e1002454. 10.1371/journal.pbio.1002454 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikeya T, Galic M, Belawat P, Nairz K, Hafen E (2002) Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Curr Biol 12:1293–1300. 10.1016/s0960-9822(02)01043-6 [DOI] [PubMed] [Google Scholar]
- Inagaki HK, Ben-Tabou de-Leon S, Wong AM, Jagadish S, Ishimoto H, Barnea G, Kitamoto T, Axel R, Anderson DJ (2012) Visualizing neuromodulation in vivo: TANGO-mapping of dopamine signaling reveals appetite control of sugar sensing. Cell 148:583–595. 10.1016/j.cell.2011.12.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inagaki HK, Panse KM, Anderson DJ (2014) Independent, reciprocal neuromodulatory control of sweet and bitter taste sensitivity during starvation in Drosophila. Neuron 84:806–820. 10.1016/j.neuron.2014.09.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaeger AH, Stanley M, Weiss ZF, Musso PY, Chan RC, Zhang H, Feldman-Kiss D, Gordon MD (2018) A complex peripheral code for salt taste. Elife 7:e37167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leitão-Gonçalves R, Carvalho-Santos Z, Francisco AP, Fioreze GT, Anjos M, Baltazar C, Elias AP, Itskov PM, Piper MDW, Ribeiro C (2017) Commensal bacteria and essential amino acids control food choice behavior and reproduction. PLoS Biol 15:e2000862. 10.1371/journal.pbio.2000862 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lesperance DNA, Broderick NA (2020) Meta-analysis of diets used in Drosophila microbiome research and introduction of the Drosophila Dietary Composition Calculator (DDCC). G3 (Bethesda) 10:2207–2211. 10.1534/g3.120.401235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin WS, Yeh SR, Fan SZ, Chen LY, Yen JH, Fu TF, Wu MS, Wang PY (2018) Insulin signaling in female Drosophila links diet and sexual attractiveness. FASEB J 32:3870–3877. 10.1096/fj.201800067R [DOI] [PubMed] [Google Scholar]
- Liu Q, Tabuchi M, Liu S, Kodama L, Horiuchi W, Daniels J, Chiu L, Baldoni D, Wu MN (2017) Branch-specific plasticity of a bifunctional dopamine circuit encodes protein hunger. Science 356:534–539. 10.1126/science.aal3245 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo J, Becnel J, Nichols CD, Nässel DR (2012) Insulin-producing cells in the brain of adult Drosophila are regulated by the serotonin 5-HT1A receptor. Cell Mol Life Sci 69:471–484. 10.1007/s00018-011-0789-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malik VS, Popkin BM, Bray G,-P, Despres J, Willett WC, Hu FB (2010) Sugar-sweetened beverages and risk of metabolic syndrome and type 2 diabetes: a meta-analysis. Diabetes Care 33:2477–2483. 10.2337/dc10-1079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manière G, Ziegler AB, Geillon F, Featherstone DE, Grosjean Y (2016) Direct sensing of nutrients via a LAT1-like transporter in Drosophila insulin-producing cells. Cell Rep 17:137–148. 10.1016/j.celrep.2016.08.093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marella S, Mann K, Scott K (2012) Dopaminergic modulation of sucrose acceptance behavior in Drosophila. Neuron 73:941–950. 10.1016/j.neuron.2011.12.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin W, Armstrong L, Rodriguez N (2005) Dietary protein intake and renal function. Nutr Metab (Lond) 2:25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masek P, Keene AC (2013) Drosophila fatty acid taste signals through the PLC pathway in sugar-sensing neurons. PLoS Genet 9:e1003710. 10.1371/journal.pgen.1003710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- May CE, Vaziri A, Lin YQ, Grushko O, Khabiri M, Wang Q-P, Holme KJ, Pletcher SD, Freddolino PL, Neely GG, Dus M (2019) High dietary sugar reshapes sweet taste to promote feeding behavior in Drosophila melanogaster. Cell Rep 27:1675–1685.e7. 10.1016/j.celrep.2019.04.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayntz D, Raubenheimer D, Salomon M, Toft S, Simpson SJ (2005) Nutrient-specific foraging in invertebrate predators. Science 307:111–113. 10.1126/science.1105493 [DOI] [PubMed] [Google Scholar]
- Mi H, Huang X, Muruganujan A, Tang H, Mills C, Kang D, Thomas PD (2017) PANTHER version 11: expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res 45:D183–D189. 10.1093/nar/gkw1138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mozaffarian D, Hao T, Rimm EB, Willett WC, Hu FB (2011) Changes in diet and lifestyle and long-term weight gain in women and men. N Engl J Med 364:2392–2404. 10.1056/NEJMoa1014296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura A, Ishida Y, Takahashi A, Okamoto H, Sakabe M, Itoh M, Takano-Shimizu T, Ozaki M (2012) Starvation-induced elevation of taste responsiveness and expression of a sugar taste receptor gene in Drosophila melanogaster. J Neurogenet 26:206–215. 10.3109/01677063.2012.694931 [DOI] [PubMed] [Google Scholar]
- Okamoto N, Nishimura T (2015) Signaling from glia and cholinergic neurons controls nutrient-dependent production of an Insulin-like peptide for Drosophila body growth. Dev Cell 35:295–310. 10.1016/j.devcel.2015.10.003 [DOI] [PubMed] [Google Scholar]
- Park J, Carlson JR (2018) Physiological responses of the Drosophila labellum to amino acids. J Neurogenet 32:27–36. 10.1080/01677063.2017.1406934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasco MY, Léopold P (2012) High sugar-induced insulin resistance in Drosophila relies on the lipocalin Neural Lazarillo. PLoS One 7:e36583. 10.1371/journal.pone.0036583 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Post S, Tatar M (2016) Nutritional geometric profiles of Insulin/IGF expression in Drosophila melanogaster. PLoS One 11:e0155628. 10.1371/journal.pone.0155628 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ribeiro C, Dickson BJ (2010) Sex peptide receptor and neuronal TOR/S6K signaling modulate nutrient balancing in Drosophila. Curr Biol 20:1000–1005. 10.1016/j.cub.2010.03.061 [DOI] [PubMed] [Google Scholar]
- Ro J, Pak G, Malec PA, Lyu Y, Allison DB, Kennedy RT, Pletcher SD (2016) Serotonin signaling mediates protein valuation and aging. Elife 5:e16843. 10.7554/eLife.16843 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson MD, McCarthy DJ, Smyth GK (2010) edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26:139–140. 10.1093/bioinformatics/btp616 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rovenko BM, Kubrak OI, Gospodaryov DV, Perkhulyn NV, Yurkevych IS, Sanz A, Lushchak OV, Lushchak VI (2015) High sucrose consumption promotes obesity whereas its low consumption induces oxidative stress in Drosophila melanogaster. J Insect Physiol 79:42–54. 10.1016/j.jinsphys.2015.05.007 [DOI] [PubMed] [Google Scholar]
- Samuelsson AM, Matthews PA, Argenton M, Christie MR, McConnell JM, Eugene HJ, Piersma AH, Ozanne SE, Twinn DF, Remacle C, Rowlerson A, Poston L, Taylor PD (2008) Diet-induced obesity in female mice leads to offspring hyperphagia, adiposity, hypertension, and Insulin resistance. Hypertension 51:383–392. 10.1161/HYPERTENSIONAHA.107.101477 [DOI] [PubMed] [Google Scholar]
- Semaniuk UV, Gospodaryov DV, Feden'ko KM, Yurkevych IS, Vaiserman AM, Storey KB, Simpson SJ, Lushchak O (2018) Insulin-like peptides regulate feeding preference and metabolism in Drosophila. Front Physiol 9:1083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Senthilan PR, Grebler R, Reinhard N, Rieger D, Helfrich-Förster C (2019) Role of rhodopsins as circadian photoreceptors in Drosophila melanogaster. Biology (Basel) 8:6. 10.3390/biology8010006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson SJ, Simpson CL (1992) Mechanisms controlling modulation by haemolymph amino acids of gustatory responsiveness in the locust. J Exp Biol 168:269–287. 10.1242/jeb.168.1.269 [DOI] [Google Scholar]
- Soneson C, Love MI, Robinson MD (2015) Differential analyses for RNA-seq: transcript-level estimates improve gene-level inferences. F1000Res 4:1521. 10.12688/f1000research.7563.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steck K, Walker SJ, Itskov PM, Baltazar C, Moreira J-M, Ribeiro C (2018) Internal amino acid state modulates yeast taste neurons to support protein homeostasis in Drosophila. Elife 7:e31625. 10.7554/eLife.31625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun J, Liu C, Bai X, Li X, Li J, Zhang Z, Zhang Y, Guo J, Li Y (2017) Drosophila FIT is a protein-specific satiety hormone essential for feeding control. Nat Commun 8:14161. 10.1038/ncomms14161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatar M, Kopelman A, Epstein D, Tu MP, Yin CM, Garofalo RS (2001) A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 292:107–110. 10.1126/science.1057987 [DOI] [PubMed] [Google Scholar]
- Tauber JM, Brown EB, Li Y, Yurgel ME, Masek P, Keene AC (2017) A subset of sweet-sensing neurons identified by IR56d are necessary and sufficient for fatty acid taste. PLoS Genet 13:e1007059. 10.1371/journal.pgen.1007059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toshima N, Tanimura T (2012) Taste preference for amino acids is dependent on internal nutritional state in Drosophila melanogaster. J Exp Biol 215:2827–2832. 10.1242/jeb.069146 [DOI] [PubMed] [Google Scholar]
- Vargas MA, Luo N, Yamaguchi A, Kapahi P (2010) A Role for S6 kinase and serotonin in postmating dietary switch and balance of nutrients in D. melanogaster. Curr Biol 20:1006–1011. 10.1016/j.cub.2010.04.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Virtanen HEK, Voutilainen S, Koskinen TT, Mursu J, Tuomainen T-P, Virtanen JK (2018) Intake of different dietary proteins and risk of heart failure in men: the Kuopio Ischaemic Heart Disease risk factor study. Circ Heart Fail 11:e004531. 10.1161/CIRCHEARTFAILURE.117.004531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Geijer L, Ekelund M (2015) Ketoacidosis associated with low-carbohydrate diet in a non-diabetic lactating woman: a case report. J Med Case Rep 9:224. 10.1186/s13256-015-0709-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang QP, Lin YQ, Zhang L, Wilson YA, Oyston LJ, Cotterell J, Qi Y, Khuong TM, Bakhshi N, Planchenault Y, Browman DT, Lau MT, Cole TA, Wong ACN, Simpson SJ, Cole AR, Penninger JM, Herzog H, Gregory Neely G (2016) Sucralose promotes food intake through NPY and a neuronal fasting response. Cell Metab 24:75–90. 10.1016/j.cmet.2016.06.010 [DOI] [PubMed] [Google Scholar]
- Wang QP, Lin YQ, Lai ML, Su Z, Oyston LJ, Clark T, Park SJ, Khuong TM, Lau MT, Shenton V, Shi YC, James DE, Ja WW, Herzog H, Simpson SJ, Neely GG (2020) PGC1α controls sucrose taste sensitization in Drosophila. Cell Rep 31:107480. 10.1016/j.celrep.2020.03.044 [DOI] [PubMed] [Google Scholar]
- Waterlow JC (1974) Letter: evolution of kwashiorkor and marasmus. Lancet 2:712. 10.1016/s0140-6736(74)93283-8 [DOI] [PubMed] [Google Scholar]
- Wisotsky Z, Medina A, Freeman E, Dahanukar A (2011) Evolutionary differences in food preference rely on Gr64e, a receptor for glycerol. Nat Neurosci 14:1534–1541. 10.1038/nn.2944 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Preference (left) and participation (right) of males prefed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays; n = 6 balanced (0 d), 6 sugar-reduced (0 d), 15 balanced (1 d), 13 sugar-reduced (1 d), 13 balanced (2 d), 8 sugar-reduced (2 d), 17 balanced (3 d), 15 sugar-reduced (3 d); *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-1, TIF file (5.9MB, tif) .
Participation of mated females fed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays; n = 6 balanced (0 d), 6 sugar-reduced (0 d), 15 balanced (1 d), 13 sugar-reduced (1 d), 13 balanced (2 d), 8 sugar-reduced (2 d), 17 balanced (3 d), 12 sugar-reduced (3 d). No significant differences were found between balanced and sugar-reduced, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-2, TIF file (2MB, tif) .
Head and body weights of mated females prefed with indicated diet for 1 d; n = 6 trials. ns = not significant, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-3, TIF file (2.1MB, tif) .
Preference (left) and participation (right) of males prefed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract in binary choice assays; n = 6 balanced, 6 sugar-enriched (0 d), n = 9 balanced, 6 sugar-enriched (1 d), n = 13 balanced, 6 sugar-enriched (2 d), n = 17 balanced, 10 sugar-enriched (3 d), n = 7 balanced, 16 sugar-enriched (4 d), n = 6 balanced, 6 sugar-enriched (5 d), n = 7 balanced, 6 sugar-enriched (6 d), n = 10 balanced, 11 sugar-enriched (7 d); *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-4, TIF file (5.7MB, tif) .
Participation of mated females fed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract in binary choice assays; n = 6 balanced, 6 sugar-enriched (0 d), n = 9 balanced, 6 sugar-enriched (1 d), n = 13 balanced, 6 sugar-enriched (2 d), n = 17 balanced, 10 sugar-enriched (3 d), n = 7 balanced, 16 sugar-enriched (4 d), n = 6 balanced, 6 sugar-enriched (5 d), n = 7 balanced, 6 sugar-enriched (6 d), n = 10 balanced, 11 sugar-enriched (7 d). No significant differences were found between balanced and sugar-enriched, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-5, TIF file (2.5MB, tif) .
Head and body weights of mated females prefed with indicated diet for 4 d; n = 7 trials. ns = not significant, ***p < 0.001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 1-6, TIF file (2MB, tif) .
Left, Preference of mated females prefed on balanced or sugar-reduced diet, and either wet-starved (starved) or not starved (unstarved) prior to testing in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye); n = 5. Right, Preference of mated females prefed on balanced or sugar-enriched diet, and either wet-starved (starved) or not starved (unstarved) prior to testing in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye); n = 6. For both graphs, *p < 0.05, **p < 0.01, ***p < 0.001, one-way ANOVA with Tukey's post hoc multiple comparisons test. Download Figure 1-7, TIF file (5.4MB, tif) .
Representative traces (left) and mean responses (right) to sucrose in flies prefed as indicated for 1 d followed by starvation for 1 d; they were given access to water during the starvation period. Recordings were taken from L-type labellar sensilla in mated females, and action potentials in the first 500-ms period after contact were counted; n = 11 (balanced), n = 13 (sugar-reduced) sensilla from four flies each; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 2-1, TIF file (7MB, tif) .
Representative traces (left) and mean responses (right) to sucrose in flies prefed as indicated for 4 d followed by starvation for 1 d; they were given access to water during the starvation period. Recordings were taken from L-type labellar sensilla in mated females, and action potentials in the first 500-ms period after contact were counted; n = 16 (balanced), n = 13 (sugar-reduced) sensilla from four flies each; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 2-2, TIF file (7.6MB, tif) .
Relative normalized expression levels of Dop2R in the brain of flies fed on indicated diet for 1 d; n = 3 biological replicates. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC. Primers for Dop2R are 5′-CTGAACTGCACCAACGAGACGC and 5′-CAGGATGTTGCCGAAGAGGGTC. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Graph shows mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 5-1, TIF file (2.4MB, tif) .
Image of agarose gel electrophoresis of RT-PCR samples using RNA derived from heads of 50–60 flies of the indicated genotypes and primers designed to amplify synaptotagmin (syt) or Dop2R sequences. Genotypes were as following: elav-GAL4; UAS-Dcr2 (elav-GAL4); UAS-Dop2R RNAi1, BDSC 26001 (UAS-Dop2R RNAi1); and elav-GAL4 > Dop2R RNAi1. Lane 1 was loaded with Generuler 1 Kb Plus (Thermo Fisher SM1331). Primers for syt were as following: 5′-TCTGGTCGTGCTTCGAGAAG and 5′-CGGATCCCTATGTCAAGGTG. Primers for Dop2R were 5′-ACTCGGTGGACTTGTCGTTC and 5′-CACCGAATTCACGCCAAGCACAAAAAT. Download Figure 5-2, TIF file (3.8MB, tif) .
Participation of mated females fed on indicated diet for 5 mm sucrose (blue dye) tested against 1% yeast extract (pink dye) in binary choice assays. w1118 (control, n = 6 balanced, 6 sugar-reduced), BDSC#52025 (Dop2R1, n = 6 balanced, 6 sugar-reduced), and BDSC#52517 (Dop2R2, n = 10 balanced, 10 sugar-reduced); *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 5-3, TIF file (2.2MB, tif) .
Starvation resistance of mated females that were given water alone or the sugar-reduced diet for 1 d and then subjected to starvation with access to water. Statistical comparison was done by the Log-rank (Mantel–Cox) test, n = 60 flies for each condition. Download Figure 5-4, TIF file (3.3MB, tif) .
Table comparing mortality of mated females that were wet-starved or fed on the sugar-reduced diet; n = 60 flies for each condition. Download Figure 5-5, TIF file (1.9MB, tif) .
Neuronal responses to various concentrations of sucrose obtained from mated females fed on the balanced or sugar-reduced diets for 1 d. Tip recordings were taken from labellar L-type sensilla. Genotypes were w1118 (control) and BDSC#52025 (Dop2R1); n = 25–29 sensilla from five to six flies (control) and n = 22–23 sensilla from five to six flies (Dop2R1); *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA with Tukey's post hoc multiple comparisons test. Download Figure 5-6, TIF file (3.7MB, tif) .
Preference of mated female flies fed on indicated diet for 1 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as following: BDSC#26001 UAS-Dop2R RNAi1 (D2R RNAi, n = 5 balanced, 5 sugar-reduced), Gad1-Gal4 > UAS-Dop2R RNAi (Gad1>D2R RNAi, n = 10 balanced, 10 sugar-reduced), and Gad1-Gal4 (n = 8 balanced, 8 sugar-reduced); *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 5-7, TIF file (2.8MB, tif) .
Confocal image of live GFP expression in Gad1-GAL4-labeled neurons in the brain. The image represents a z-projection of optical sections (1 µm) compressed in ImageJ using maximum intensity settings. Genotype: Gad1-GAL4; UAS-mCD8::GFP. The white dotted line represents the outline of the brain. Download Figure 5-8, TIF file (2.3MB, tif) .
Relative normalized expression levels of Dop2R in the brain following expression of Dop2R RNAi under the control of the Gad1-GAL4 driver. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC. Primers for Dop2R are 5′-CTGAACTGCACCAACGAGACGC and 5′-CAGGATGTTGCCGAAGAGGGTC. Graph shows mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 5-9, TIF file (2.2MB, tif) .
Preference of mated female flies expressing UAS-Dop2R RNAi in sweet taste neurons (left) or in selected subsets of neurons (right) that were prefed on indicated diet for 1 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were as following: Gr64f-GAL4 (n = 14 balanced, 15 sugar-reduced), UAS-Dop2R RNAi (D2R RNAi, n = 7 balanced, 7 sugar-reduced), Gr64f-GAL4 > D2R RNAi (n = 6 balanced, 6 sugar-reduced), Tdc2 > D2R RNAi (n = 9 balanced, 10 sugar-reduced), Tdc2-GAL4 (n = 6 balanced, 6 sugar-reduced), Ple>D2R RNAi (n = 11 balanced, 11 sugar-reduced), Ple-GAL4 (n = 6 balanced, 6 sugar-reduced), TRH>D2R RNAi (n = 6 balanced, 6 sugar-reduced), TRH-GAL4 (n = 6 balanced, 6 sugar-reduced), v-Glut>D2R RNAi (n = 6 balanced, 6 sugar-reduced), v-Glut-GAL4 (n = 6, balanced, 6 sugar-reduced); *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 5-10, TIF file (6.5MB, tif) .
Neuronal responses to various concentrations of sucrose obtained from L-type labellar sensilla of mated females fed on indicated diet for 1 d. Genotypes were as following: BDSC#26001 UAS-Dop2R RNAi1, Gad1-Gal4 > UAS-Dop2R RNAi; n = 12–16 sensilla from three to four flies; *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, two-way ANOVA post-hoc Sidak's multiple comparisons test. Download Figure 5-11, TIF file (4.2MB, tif) .
Relative normalized expression levels of dilp3 and dilp5 in the brains (left) and proboscis (right) of flies fed on balanced or sugar-enriched diets for 4 d; n = 3 biological replicates. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC. Primers for dilp3 are 5′-GTCCAGGCCACCATGAAGTTGTGC and 5′-CTTTCCAGCAGGGAACGGTCTTCG. Primers for dilp5 are 5′-TGTTCGCCAAACGAGGCACCTTGG and 5′-CACGATTTGCGGCAACAGGAGTCG. Graphs show mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 6-1, TIF file (4.9MB, tif) .
Image of agarose gel electrophoresis of PCR samples using genomic DNA templates from w1118 and dilp5 (left) or dilp3 (right) mutant flies. Lane 1 was loaded with Generuler 1 Kb Plus (Thermo Fisher SM1331). Primers for synaptotagmin (syt) were as in Extended Data Figure 5-2. Primers for dilp5 were as following: 5′-CGCCGAATGCTCGACAGTGA and 5′-CTCCGTGATCCCAGTTCTCCT. Primers for dilp3 were as following: 5′-GACTTCAGGCAACACTCGTC and 5′-ATCCTTATGATCGGCGGTGTC. Arrows mark positions of expected amplicons. Download Figure 7-1, TIF file (4.7MB, tif) .
Participation of mated females prefed on indicated diet for 4 d and tested in binary choice assays with 5 mm sucrose (blue dye) against 1% yeast extract (pink dye). Genotypes were w1118 (control, n = 8 balanced, 8 sugar-enriched), dilp2 (n = 7 balanced, 8 sugar-enriched), dilp3 (n = 7 balanced, 6 sugar-enriched), and dilp5 (n = 7 balanced, 9 sugar-enriched); *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA with Sidak's post hoc multiple comparisons test. Download Figure 7-2, TIF file (2.1MB, tif) .
Neuronal responses to various concentrations of sucrose obtained from mated females fed on balanced or sugar-enriched diet for 4 d. Tip recordings were taken from labellar L-type sensilla. Genotypes were w1118 (control) and dilp5; n = 29–30 sensilla from five to six flies (control) and n = 19–21 sensilla from five to six flies (dilp5); *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA with Tukey's post hoc multiple comparisons test. Download Figure 7-3, TIF file (3.4MB, tif) .
Relative normalized expression levels of dilp5 in the brain following expression of dilp5 RNAi in dilp2-GAL4 neurons; n = 3 biological replicates. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC. Primers for dilp5 are 5′-TGTTCGCCAAACGAGGCACCTTGG and 5′-CACGATTTGCGGCAACAGGAGTCG. Graph shows mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 7-4, TIF file (2.1MB, tif) .
Relative normalized expression levels of InR in the brain following expression of InR RNAi in Trh-GAL4 neurons; n = 3 biological replicates. Primers for rp49 are 5′-CAGTCGGATCGATATGCTAAGCTG and 5′-TAACCGATGTTGGGCATCAGATAC′. Primers for InR are 5′-TGAGCATGTGGAGCACATCAAGATG and 5′-CGTAGGAGATTTTCTCGTTTGGCTG. Expression levels were calculated using the 2−ΔΔCt method and normalized to rp49. Graphs show mean ± SEM from three independent experiments; *p < 0.05, independent samples t test. Download Figure 7-5, TIF file (2MB, tif) .
Data Availability Statement
Raw data are available on Mendeley Data (DOI: 10.17632/ngzwtf9tym.1).