

Abstract
Matrix metalloproteinases 9 (MMP9) are enzymes involved in regulating neuroplasticity in the hippocampus. This, combined with evidence for disrupted hippocampal structure and function in schizophrenia, has prompted our current investigation into the relationship between MMP9 and hippocampal volumes in schizophrenia. 34 healthy individuals (mean age = 32.50, male = 21, female = 13) and 30 subjects with schizophrenia (mean age = 33.07, male = 19, female = 11) underwent a blood draw and T1-weighted magnetic resonance imaging. The hippocampus was automatically segmented utilizing FreeSurfer. MMP9 plasma levels were measured with ELISA. ANCOVAs were conducted to compare MMP9 plasma levels (corrected for age and sex) and hippocampal volumes between groups (corrected for age, sex, total intracranial volume). Spearman correlations were utilized to investigate the relationship between symptoms, medication, duration of illness, number of episodes, and MMP9 plasma levels in patients. Last, we explored the correlation between MMP9 levels and hippocampal volumes in patients and healthy individuals separately. Patients displayed higher MMP9 plasma levels than healthy individuals (F(1, 60) = 21.19, p < 0.0001). MMP9 levels correlated with negative symptoms in patients (R = 0.39, p = 0.035), but not with medication, duration of illness, or the number of episodes. Further, patients had smaller left (F(1,59) = 9.12, p = 0.0040) and right (F(1,59) = 6.49, p = 0.013) hippocampal volumes. Finally, left (R = −0.39, p = 0.034) and right (R = −0.37, p = 0.046) hippocampal volumes correlated negatively with MMP9 plasma levels in patients. We observe higher MMP9 plasma levels in SCZ, associated with lower hippocampal volumes, suggesting involvement of MMP9 in the pathology of SCZ. Future studies are needed to investigate how MMP9 influences the pathology of SCZ over the lifespan, whether the observed associations are specific for schizophrenia, and if a therapeutic modulation of MMP9 promotes neuroprotective effects in SCZ.
Subject terms: Schizophrenia, Biomarkers
Introduction
Neuroplasticity refers to the brain’s ability to reorganize neuronal pathways in response to internal and external stimuli and comprises several levels, including micro-and macro connectivity, activity-dependent brain function, and synaptic transmission [1–3]. While previous studies have characterized altered neuroplasticity as one of the core pathologies of schizophrenia [4, 5], recent research tries to identify which factors might underly these abnormalities. Interestingly, several lines of evidence highlighted the extracellular matrix’s role in modulating neuronal pathways in response to stimuli. Specifically, matrix metalloproteinases (MMPs), a large family of extracellularly acting zinc-dependent proteases [6], have been implicated in the dysregulation of forming and eliminating synapses in schizophrenia.
Matrix metalloproteinase 9 (MMP9) is the most prevalent MMP in the central nervous system [7]. It is expressed in neural and glial cells in multiple brain regions, including the hippocampus [8, 9]. It is released under the influence of glutamate and can bind to various substrates, including beta-amyloid, precursors of growth factors, chemokines, tissue inhibitors, and adhesion molecules [10]. At the physiological level, it controls hippocampal [11–16] and dendritic development and maintains the extracellular environment of the postsynaptic part of excitatory synapses (by modifying both NMDA and AMPA receptors) [9, 17], which are vital for synaptic plasticity and long-term potentiation [18, 19]. Long-term potentiation is a physiological readout of neuroplasticity that a) reflects the functional ability for memory acquisition and storage and b) is often disturbed in schizophrenia [4, 20]. Interestingly, increased MMP9 levels are associated with structural (e.g., abnormally long and thin dendritic spines [7]) and functional (e.g., impaired long-term potentiation [14, 21, 22]) synaptic plasticity abnormalities.
MMP9 has repeatedly been implicated in the pathophysiology of schizophrenia [23]. Several studies have demonstrated MMP9 upregulation in the blood of patients with schizophrenia when compared to healthy individuals [24, 25]. Upregulation of mRNA expression of MMP9 was reported in peripheral blood mononuclear cells in treatment-naive schizophrenia patients, with a marked downregulation following treatment [26]. Preliminary evidence also suggests that minocycline, an antibiotic and MMP9 inhibitor, ameliorates symptom severity in patients with schizophrenia - further pointing towards the potential role of MMP9 as a treatment target [27–29].
While this accumulating evidence suggests a possible relationship between abnormal MMP9 regulation and brain structural and functional pathology in schizophrenia, no studies have investigated this link in humans. In this preliminary study, we propose to combine and analyze the relationship between imaging markers of the hippocampus structure, the blood marker MMP9, and clinical variables (including symptom severity, medication, duration of illness, number of episodes). Given the crucial role of the hippocampus for schizophrenia [30–33], the role of MMP9 in regulating hippocampus structure and function [8, 9, 12–15], and its role in the regulation of dendritic and synaptic plasticity, we postulate that a) patients with schizophrenia will present with higher plasma levels of MMP9 than healthy individuals and b) MMP9 abnormalities will be associated with smaller hippocampal volumes in patients.
Materials and methods
Participants
The present study includes data from 34 healthy individuals and 30 patients with schizophrenia. Patients were recruited in the Department of Psychiatry at University Hospital Brno, Brno, Czech Republic, following hospital admission for an acute psychotic episode. Patients were diagnosed utilizing the Structured clinical interview for DSM5 - Research version (SCID-RV) criteria [34]. The Positive and Negative Symptoms Scales (PANSS) [35] were administered to determine symptom severity. We recruited healthy individuals via advertising within the local community. Exclusion criteria for healthy individuals and patients were a history of neurological injury, brain disorder, substance abuse, and an inability to undergo MRI. Additional exclusion criteria for healthy individuals were any history of psychiatric illness themselves, first or second-degree relatives (assessed with Mini-International Neuropsychiatric Interview [36]).
All participants provided written informed consent for participation, and the Institutional Ethical Committee of the University Hospital Brno approved the study.
Data collection
MMP9
Venous blood from people with schizophrenia was collected approximately three weeks after hospitalization for an acute psychotic episode, on the day of MRI scanning. At this time, patients have been intensively treated with antipsychotics, and positive symptoms started to subside. The blood was collected at 7 am in a fasting state to account for possible circadian fluctuations. Blood was collected into tubes with EDTA and immediately stored on ice (0 °C). Plasma was separated by centrifugation within 30 min in a refrigerated centrifuge, and aliquots were immediately stored at −75 °C until the time of analysis.
Plasma MMP-9 values were obtained using the Quantikine sandwich ELISA kit (#DMP900, R&D Systems, Minneapolis, MN). The Quantikine immunoassay control group 130 (#QC130) was used as a quantitative control for the determination of MMP9. The MMP9 immunoassay recognizes both the pro-MMP9 (92 kDa) and the active MMP9 (82 kDa), but not the 65 kDa form. Before the analysis, plasma samples were diluted 33 times according to the manufacturer’s specifications for the assay and measured in duplicate. The detection limit was 0.156 ng/mL, and the intra-assay coefficient of variation was <2%. The optical densities were measured using the Spectramax 340PC microplate reader (Molecular Devices, San Jose, CA).
Image acquisition
3T Siemens Magnetom Prisma (Erlangen, Germany) with 64 channel Head-Neck coil was utilized to collect the structural MRI data. A whole brain, high-resolution three-dimensional T1-weighted magnetization prepared rapid gradient echo (MPRAGE) sequence was used to collect 240 sagittal slices (field of view = 224 × 224 mm2, 1 mm3 isotropic voxel, TR = 2.3 s, TE = 2.33 ms, flip angle = 8).
Image processing
Structural T1-weighted images underwent visual quality control (for artifacts). Next, images were axis-aligned and centered. Consecutively, brain masks were automatically generated using 3D Slicer (software version 4.5; www.slicer.org) and manually edited to guarantee anatomical precision. FreeSurfer 5 was run on the structural imaging data, and the output was checked for quality.
Statistical analyses
All statistical analyses were performed with SPSS version 26 and GraphPad Prism 8.
MMP9
Group comparison
To test for group differences in MMP9, we conducted an ANCOVA with MMP9 plasma level as the dependent variable, group as the independent variable (healthy individuals versus patients with schizophrenia), and age and sex as covariates. Please note that (a) MMP9 plasma values were not normally distributed in healthy individuals (healthy: D(34) = 0.20, p < 0.001; patients: D(30) = 0.11, p < 0.20) and (b) the variance of MMP9 plasma levels was different between groups (F = 8.51, p < 0.005). Thus, we repeated the group comparisons for MMP9 levels with a non-parametric Mann–Whitney U test.
Association between MMP9 and clinical measures
Post hoc Spearman’s correlation analyses between positive/ negative symptom severity, chlorpromazine equivalent dosage (CPZ), duration of illness, number of episodes, and MMP9 plasma levels were carried out for patients. We chose Spearman’s rank order because not all variables were normally distributed. We utilized Fisher’s Exact Score to test if the correlation coefficients significantly differed from zero.
Hippocampus
Group comparison
To test for group differences between the patients and the HC groups in the hippocampus, we used two ANCOVAs with left or right hippocampal volume as the independent variable, group (healthy individuals versus patients with schizophrenia) as the dependent variable, and age, sex, and total intracranial volume as covariates.
Association between MMP9 and hippocampus
For patients with schizophrenia and healthy individuals separately, we applied Spearman’s correlation analyses to correlate the left and right hippocampus volume with MMP9 plasma levels. Fisher’s Exact Score was utilized to test whether the correlation coefficient significantly differed from zero.
Results
Demographics
Patients with schizophrenia and healthy individuals did not show group differences in critical demographic variables (see Table 1).
Table 1.
Demographics.
| Healthy individuals (N = 34) | Individuals with schizophrenia (N = 30) | Test-statistic | |
|---|---|---|---|
| Age in years (mean ± std) | 32.50 ± 9.10 | 33.07 ± 9.48 | T = −0.24, df = 62, p < 0.81 |
| Sex | Male = 21, Female = 13 | Male = 19, Female = 11 | X2 = 0.017, df = 1, p < 0.90 |
| Education in years (mean ± std) | 14.91 ± 2.71 | 13.93 ± 2.95 | T = 1.38, df = 62, p < 0.17 |
| Duration of illness in years (mean ± std) | 7.96 ± 6.45 | ||
| Number of episodes | 3.37 ± 2.39 | ||
| Chlorpromazine equivalent dosage in mg | 740.77 ± 385.08 | ||
| PANSS Negative Symptoms | 15.60 ± 6.31 | ||
| PANSS Positive Symptoms | 11.80 ± 4.77 |
Table 1 shows that healthy individuals and patients were matched for age, sex, and education years and displays clinical characteristics for patients.
std standard deviation, PANSS Positive and Negative Symptom Scale [74].
MMP9
Group comparison
ANCOVA with MMP9 plasma levels as the dependent variable showed a significant effect of group (F(1,60) = 21.19, p < 0.0001, Cohen’s d = 1.09). The average MMP 9 plasma levels of patients with schizophrenia were almost twice as high as those of healthy individuals (89.76% increase, patients = 119.72 ± 62.65 ng/ml; healthy individuals = 63.09 ± 37.89 ng/ml) (Fig. 1). In addition to a significant group effect, we also observed a significant sex effect F(1,60) = 8.46, p < 0.005). Age did not have a significant effect (F(1,60) = 0.006, p < 0.94). To follow up on the significant sex effect, we conducted a sex x group interaction analysis, which did not yield a significant result (F(1,60) = 2.85, p < .097). Please note that we still observed significant group differences when repeating the analyses with a non-parametric Mann–Whitney U test (U = 188.00, p < 0.0001).
Fig. 1. Group comparison of MMP9 values.
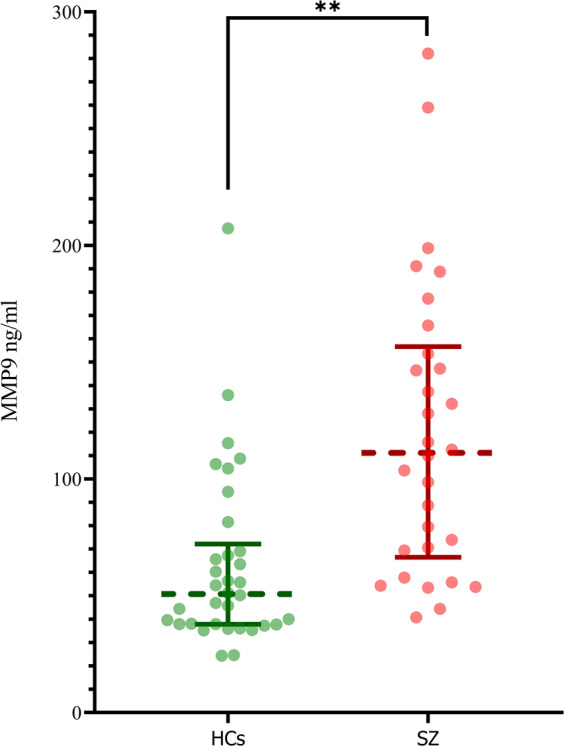
Group comparisons for matrix metalloproteinase 9 (MMP9) plasma values (in ng/ml) for healthy individuals (HC) and patients with schizophrenia (SCZ). The dotted lines display the median, the whiskers the interquartile range. Single dots represent individual values. An ANCOVA (corrected for age and sex) demonstrates that patients with SCZ display significantly higher MMP9 plasma values than HC (F(1,60) = 21.19, p < .0001, Cohen’s d = 1.09). This result was confirmed by a non-parametric Mann–Whitney U test (U = 188.00, p < 0.0001).
Association between MMP9 and clinical measures
To assess whether MMP9 levels were associated with clinical measures, we performed correlations in patients with schizophrenia. Positive symptom severity did not significantly correlate with MMP9 plasma levels (R = −0.23, p = 0.23). However, there was a statistically significant correlation between MMP9 plasma levels and negative symptom severity (R = 0.39, p = 0.035). MMP9 plasma levels were not correlated with CPZ (R = −0.015, p = 0.94), duration of illness (R = −0.014, p < 0.94), or number of episodes (R = 0.036, p < 0.85).
Hippocampus
ANCOVA with the left hippocampal volume as dependent variable (F(1,59) = 9.12, p = 0.004, Cohen’s d = 0.44) and the ANCOVA with the right hippocampal volume as dependent variable (F(1,59) = 6.49, p = 0.013, Cohen’s d = 0.33) both demonstrated a significant group effect. Patients with schizophrenia had smaller left (4.35% decrease, healthy individuals = 3.91 cm3 ± 0.37 cm3, patients = 3.74 cm3 ± 0.40 mm3) and right (3.50% decrease, healthy individuals = 4.00 cm3 ± 0.38 cm3, patients = 3.86 mm3 ± 0.46 cm3) hippocampal volumes than healthy individuals. For associations between hippocampal volumes and clinical variables please see Supplementary Table 1.
Association between MMP9 and hippocampus
For healthy individuals we did not observe an association between MMP9 plasma levels and either left (R = −0.030, p = 0.97) or right (R = 0.003, p = 0.99) hippocampal volumes. However, in patients MMP9 plasma levels were negatively correlated with left (R = −0.39, p = 0.034) and right (R = −0.37, p = 0.046) hippocampal volumes (Fig. 2).
Fig. 2. Correlation of MMP9 levels and hippocampal volume.

Correlation between left and right hippocampal volume (in cm3) and matrix metalloproteinase 9 MMP9 plasma levels (in ng/ml) in patients with schizophrenia. In patients MMP9 plasma levels were negatively correlated with both left (R = −0.39, p = 0.034) and right (R = −0.37, p = 0.046) hippocampal volumes.
Discussion
The present exploratory study characterizes the association between MMP9 and the hippocampus in schizophrenia. We observed lower hippocampal volumes and higher MMP9 plasma levels in patients. Most interestingly, in patients, hippocampal volumes and MMP9 plasma levels were negatively correlated.
Group comparisons
MMP9
Our finding of elevated MMP9 plasma levels is in line with previous studies demonstrating higher peripheral activity of MMP9 [37] and its increased blood concentration [24, 25] in patients with schizophrenia. It has been proposed that abnormal MMP9 levels might be a useful peripheral biomarker of schizophrenia [24], potentially discriminating patients from healthy individuals [38].
Indeed, given MMP9 distribution and function, upregulation of this protein might explain several processes that are disturbed throughout the lifespan in schizophrenia. MMP9 is an essential regulator of neuroplasticity. Both up- and downregulation of MMP9 can lead to impaired neuroplasticity [19], possibly in part by regulating the development and integrity of perineuronal nets. Moreover, a recent animal study suggests that MMP9 upregulation might be a core mechanism that translates oxidative stress into neuroinflammatory dysregulation and cytotoxic responses [39]. In that paper, the authors propose a “vicious cycle” between oxidative stress and neuroinflammation, especially during development, controlled and modulated by MMP9 expression, leading to long-term detrimental effects on PNNs and PVI neurons.
While our study, correlational in nature, is not designed to test for consequences of MMP9 upregulation, we see an association between MMP9 plasma levels and negative symptoms in patients with schizophrenia. Previous research suggested that negative symptoms may indicate neuronal damage [40] and that predominantly negative symptoms are associated with neurodevelopmental abnormalities [41]. Additionally, our finding is in line with recently published studies reporting that MMP9 mRNA expression is more pronounced in deficit schizophrenia [38, 42]. Last, it has previously been demonstrated that MMP9 influences hippocampal and prefrontal N-methyl-D-aspartate receptor (NMDA) activity [38, 42]. We speculate that this modulation of NMDA activity might mediate the association between MMP9 and negative symptoms in schizophrenia.
Hippocampal volume
Given the crucial role of the hippocampus for memory, emotion processing, and higher-level cognition [43], which are all affected in schizophrenia, it is one of the most studied brain structures in psychosis. Evidence from postmortem [32, 44, 45] and imaging research [43, 46, 47] implies structural and functional abnormalities of the hippocampus and the hippocampal circuits in schizophrenia. Our finding of overall hippocampal atrophy is, thereby, in line with a recent meta-analysis that showed that the hippocampus is the most affected subcortical brain region in SCZ [33].
Association between MMP9 and brain structure
We observed a negative association between MMP9 plasma levels and hippocampal volume. We have focused our analyses on the hippocampus, given the (1) vital role of the hippocampus for the schizophrenia pathophysiology and (2) known importance of MMP9 for hippocampal development and function [11]. MMP9 regulates dendritic and synaptic morphology during development in the hippocampus [9, 14, 48–50] and is expressed in the adult human hippocampus [9]. Additionally, animal studies have demonstrated the role of MMP9 expression in the hippocampus for memory performance [18, 19] and its upregulation in reaction to stress, drugs, or fear [51–53]. Our study supports these previous findings by linking MMP9 plasma levels with macrostructural volumetric measurements in humans. However, more extensive studies are needed to examine whether the observed associations are specific for schizophrenia, how they develop over the trajectory of the disorder, and if therapeutic modifications of MMP9 plasma levels and activity can influence the pathology of SCZ [54]. Last, given the small sample size and exploratory nature of the present study, we did not explore if the association between MMP9 and hippocampal volume might be specific to subregions of the hippocampus. However, some animal studies imply that MMP9 is upregulated explicitly in CA1 and not in the whole hippocampus [19, 52] and experiments in ischemic insult mice models reveal that MMP9 inhibitors [55, 56] have spatially specific neuroprotective effects in the striatum and CA1/CA2 subfields of the hippocampus.
Limitations and future directions
We acknowledge several limitations of the present study. First, this is a preliminary study testing a novel approach of linking MMP9 plasma levels and brain pathologies. Given its novelty and relatively small sample size, we only assessed one brain region. However, there is evidence suggesting a broader role of MMP9 for brain structure and SCZ pathology. Specifically, MMP9 might also play a pivot role in modulating the central inflammatory response [7, 53, 57]. Future studies are needed to examine how MMP9 and brain pathologies’ association develops with age [58] and over the SCZ trajectory. Additionally, we acknowledge that while the reported correlations are medium strong, the statistical tests were not corrected for multiple comparisons, and the significance would not survive a Bonferroni multiple-comparison correction (likely due to the small sample size). While we demonstrated that MMP9 is not associated with antipsychotic medication, more extensive studies should examine additional factors, such as sex [59, 60], nicotine [61, 62], alcohol [63], and metabolic dysregulations [64]. Furthermore, while we do not see an association between MMP9 levels and hippocampal volumes in healthy individuals, this does not mean that our finding is specific to SCZ. While some previous studies have suggested that MMP9 might be an excellent marker to discriminate patients with SCZ from healthy individuals [25], trans-diagnostic studies are missing. Importantly there is preliminary evidence that MMP9 might also be involved in the pathology of mood disorders [65, 66], autism [67], multiple sclerosis [68, 69], and stroke [70, 71]. Last, while most studies propose that peripheral MMP9 levels can directly be associated with MMP9 levels in the brain [72], this assumption needs further validation.
Conclusion
We provide evidence for the association between MMP9 and hippocampal volume in patients with schizophrenia. Additionally, we show abnormal MMP9 plasma levels and hippocampal volumes in patients with SCZ, hinting towards the role of MMP9 in the pathology of SCZ. For example, MMP9 inhibitors such as minocycline can regulate MMP9 and promote neuroplasticity, reverse neurodegeneration and improve negative symptoms in SCZ [73]. More extensive studies are needed to replicate our findings and investigate the association between MMP9 and brain pathology along the SCZ spectrum.
Funding and disclosure
This work was supported by the National Institutes of Health (NIH) (grant numbers, R01MH102377 [to MK], K24MH110807 [to MK], R01MH108574 [to OP], P41EB015902 and R01MH074794 [to CFW]. R01 MH112748 [to MK, NM]). We acknowledge the core facility MAFIL supported by the Czech-BioImaging large RI project (LM2018129 funded by MEYS CR) for their support with obtaining scientific data presented in this paper. The authors declare no competing interests.
Supplementary information
Acknowledgements
This work is part of the Ph.D. thesis of Carina Heller.
Author contributions
JS-H Ph.D. MD: Substantial contributions to the design of the work, analysis, and interpretation of data; drafting the work; final approval of the version to be published; agreement to be accountable for all aspects of the work. MS MD: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. NM Ph.D. MD: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. JR Ph.D.: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. K-IKC Ph.D.: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. ER: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. MV Ph.D.: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. OSS MD: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. CH: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. OP Ph.D.: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. FAS Ph.D.: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. C-FW Ph.D.: Substantial contributions to the analysis of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. JL Ph.D. MD: Substantial contributions to the acquisition of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. LU Ph.D. MD: Substantial contributions to the acquisition of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. JT Ph.D. MD: Substantial contributions to the acquisition of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. LV Ph.D.: Substantial contributions to the acquisition of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. PKI: Substantial contributions to the acquisition of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. MJ: Substantial contributions to the acquisition of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. TWW Ph.D. MD: Substantial contributions to the design of the work, analysis, and interpretation of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. TK Ph.D. MD: Substantial contributions to the acquisition of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. ZK Ph.D.: Substantial contributions to the design of the work, analysis, and interpretation of data; revising the draft; final approval of the version to be published; agreement to be accountable for all aspects of the work. MK Ph.D. MD: Substantial contributions to the design of the work, analysis, and interpretation of data; drafting the work; final approval of the version to be published; agreement to be accountable for all aspects of the work
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
The online version contains supplementary material available at 10.1038/s41386-021-00997-5.
References
- 1.De Picker LJ, Morrens M, Chance SA, Boche D. Microglia and brain plasticity in acute psychosis and schizophrenia illness course: a meta-review. Front Psychiatry. 2017;8:238. doi: 10.3389/fpsyt.2017.00238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Oberman L, Pascual-Leone A. Changes in plasticity across the lifespan: cause of disease and target for intervention. Prog Brain Res. 2013;207:91–120. doi: 10.1016/B978-0-444-63327-9.00016-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bernardinelli Y, Nikonenko I, Muller D. Structural plasticity: mechanisms and contribution to developmental psychiatric disorders. Front Neuroanat. 2014;8:123. doi: 10.3389/fnana.2014.00123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.McCullumsmith RE, Clinton SM, Meador-Woodruff JH. Schizophrenia as a disorder of neuroplasticity. Int Rev Neurobiol. 2004;59:19–45. doi: 10.1016/S0074-7742(04)59002-5. [DOI] [PubMed] [Google Scholar]
- 5.Crabtree GW, Gogos JA. Synaptic plasticity, neural circuits, and the emerging role of altered short-term information processing in schizophrenia. Front Synaptic Neurosci. 2014;6:28. doi: 10.3389/fnsyn.2014.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rybakowski JK. Matrix metalloproteinase-9 (MMP9)-a mediating enzyme in cardiovascular disease, cancer, and neuropsychiatric disorders. Cardiovasc Psychiatry Neurol. 2009;2009:904836. doi: 10.1155/2009/904836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Vafadari B, Salamian A, Kaczmarek L. MMP-9 in translation: from molecule to brain physiology, pathology, and therapy. J Neurochem. 2016;139:91–114. doi: 10.1111/jnc.13415. [DOI] [PubMed] [Google Scholar]
- 8.Sathyanesan M, Girgenti MJ, Banasr M, Stone K, Bruce C, Guilchicek E, et al. A molecular characterization of the choroid plexus and stress-induced gene regulation. Transl Psychiatry. 2012;2:e139. doi: 10.1038/tp.2012.64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dziembowska M, Wlodarczyk J. MMP9: a novel function in synaptic plasticity. Int J Biochem cell Biol. 2012;44:709–13. doi: 10.1016/j.biocel.2012.01.023. [DOI] [PubMed] [Google Scholar]
- 10.Bronisz E, Kurkowska-Jastrzębska I. Matrix metalloproteinase 9 in epilepsy: the role of neuroinflammation in seizure development. Mediators Inflamm. 2016;2016:7369020. doi: 10.1155/2016/7369020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Murase S, McKay RD. Matrix metalloproteinase-9 regulates survival of neurons in newborn hippocampus. J Biol Chem. 2012;287:12184–94. doi: 10.1074/jbc.M111.297671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sidhu H, Dansie LE, Hickmott PW, Ethell DW, Ethell IM. Genetic removal of matrix metalloproteinase 9 rescues the symptoms of fragile X syndrome in a mouse model. J Neurosci. 2014;34:9867–79. doi: 10.1523/JNEUROSCI.1162-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Aujla PK, Huntley GW. Early postnatal expression and localization of matrix metalloproteinases-2 and -9 during establishment of rat hippocampal synaptic circuitry. J Comp Neurol. 2014;522:1249–63. doi: 10.1002/cne.23468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fragkouli A, Papatheodoropoulos C, Georgopoulos S, Stamatakis A, Stylianopoulou F, Tsilibary EC, et al. Enhanced neuronal plasticity and elevated endogenous sAPPalpha levels in mice over-expressing MMP9. J Neurochem. 2012;121:239–51. doi: 10.1111/j.1471-4159.2011.07637.x. [DOI] [PubMed] [Google Scholar]
- 15.Gkogkas CG, Khoutorsky A, Cao R, Jafarnejad SM, Prager-Khoutorsky M, Giannakas N, et al. Pharmacogenetic inhibition of eIF4E-dependent Mmp9 mRNA translation reverses fragile X syndrome-like phenotypes. Cell Rep. 2014;9:1742–55. doi: 10.1016/j.celrep.2014.10.064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wojcik-Stanaszek L, Gregor A, Zalewska T. Regulation of neurogenesis by extracellular matrix and integrins. Acta Neurobiol. Exp. 2011;71:103–12. doi: 10.55782/ane-2011-1827. [DOI] [PubMed] [Google Scholar]
- 17.Tsilibary E, Tzinia A, Radenovic L, Stamenkovic V, Lebitko T, Mucha M, et al. Neural ECM proteases in learning and synaptic plasticity. Prog Brain Res. 2014;214:135–57. doi: 10.1016/B978-0-444-63486-3.00006-2. [DOI] [PubMed] [Google Scholar]
- 18.Wiera G, Nowak D, van Hove I, Dziegiel P, Moons L, Mozrzymas JW. Mechanisms of NMDA receptor- and voltage-gated L-type calcium channel-dependent hippocampal LTP critically rely on proteolysis that is mediated by distinct metalloproteinases. J Neurosci. 2017;37:1240–56. doi: 10.1523/JNEUROSCI.2170-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wiera G, Wozniak G, Bajor M, Kaczmarek L, Mozrzymas JW. Maintenance of long-term potentiation in hippocampal mossy fiber-CA3 pathway requires fine-tuned MMP-9 proteolytic activity. Hippocampus. 2013;23:529–43. doi: 10.1002/hipo.22112. [DOI] [PubMed] [Google Scholar]
- 20.Salavati B, Rajji TK, Price R, Sun Y, Graff-Guerrero A, Daskalakis ZJ. Imaging-based neurochemistry in schizophrenia: a systematic review and implications for dysfunctional long-term potentiation. Schizophr Bull. 2015;41:44–56. doi: 10.1093/schbul/sbu132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Meighan SE, Meighan PC, Choudhury P, Davis CJ, Olson ML, Zornes PA, et al. Effects of extracellular matrix-degrading proteases matrix metalloproteinases 3 and 9 on spatial learning and synaptic plasticity. J Neurochem. 2006;96:1227–41. doi: 10.1111/j.1471-4159.2005.03565.x. [DOI] [PubMed] [Google Scholar]
- 22.Wiera G, Szczot M, Wojtowicz T, Lebida K, Koza P, Mozrzymas JW. Impact of matrix metalloproteinase-9 overexpression on synaptic excitatory transmission and its plasticity in rat CA3-CA1 hippocampal pathway. J Physiol Pharmacol. 2015;66:309–15. [PubMed] [Google Scholar]
- 23.Bitanihirwe BKY, Woo TW. A conceptualized model linking matrix metalloproteinase-9 to schizophrenia pathogenesis. Schizophr Res. 2020;218:28–35. doi: 10.1016/j.schres.2019.12.015. [DOI] [PubMed] [Google Scholar]
- 24.Yamamori H, Hashimoto R, Ishima T, Kishi F, Yasuda Y, Ohi K, et al. Plasma levels of mature brain-derived neurotrophic factor (BDNF) and matrix metalloproteinase-9 (MMP-9) in treatment-resistant schizophrenia treated with clozapine. Neurosci Lett. 2013;556:37–41. doi: 10.1016/j.neulet.2013.09.059. [DOI] [PubMed] [Google Scholar]
- 25.Ali FT, Abd El-Azeem EM, Hamed MA, Ali MAM, Abd Al-Kader NM, Hassan EA. Redox dysregulation, immuno-inflammatory alterations and genetic variants of BDNF and MMP-9 in schizophrenia: Pathophysiological and phenotypic implications. Schizophr Res. 2017;188:98–109. doi: 10.1016/j.schres.2017.01.016. [DOI] [PubMed] [Google Scholar]
- 26.Kumarasinghe N, Beveridge NJ, Gardiner E, Scott RJ, Yasawardene S, Perera A, et al. Gene expression profiling in treatment-naive schizophrenia patients identifies abnormalities in biological pathways involving AKT1 that are corrected by antipsychotic medication. Int J Neuropsychopharmacol. 2013;16:1483–503. doi: 10.1017/S1461145713000035. [DOI] [PubMed] [Google Scholar]
- 27.Solmi M, Veronese N, Thapa N, Facchini S, Stubbs B, Fornaro M, et al. Systematic review and meta-analysis of the efficacy and safety of minocycline in schizophrenia. CNS Spectr. 2017;22:415–26. doi: 10.1017/S1092852916000638. [DOI] [PubMed] [Google Scholar]
- 28.Chaudhry IB, Hallak J, Husain N, Minhas F, Stirling J, Richardson P, et al. Minocycline benefits negative symptoms in early schizophrenia: a randomised double-blind placebo-controlled clinical trial in patients on standard treatment. J Psychopharmacol. 2012;26:1185–93. doi: 10.1177/0269881112444941. [DOI] [PubMed] [Google Scholar]
- 29.Levkovitz Y, Mendlovich S, Riwkes S, Braw Y, Levkovitch-Verbin H, Gal G, et al. A double-blind, randomized study of minocycline for the treatment of negative and cognitive symptoms in early-phase schizophrenia. J Clin Psychiatry. 2010;71:138–49. doi: 10.4088/JCP.08m04666yel. [DOI] [PubMed] [Google Scholar]
- 30.Walker MA, Highley JR, Esiri MM, McDonald B, Roberts HC, Evans SP, et al. Estimated neuronal populations and volumes of the hippocampus and its subfields in schizophrenia. Am J Psychiatry. 2002;159:821–8. doi: 10.1176/appi.ajp.159.5.821. [DOI] [PubMed] [Google Scholar]
- 31.Highley JR, Walker MA, McDonald B, Crow TJ, Esiri MM. Size of hippocampal pyramidal neurons in schizophrenia. Br J Psychiatry. 2003;183:414–7. doi: 10.1192/bjp.183.5.414. [DOI] [PubMed] [Google Scholar]
- 32.Zaidel DW, Esiri MM, Harrison PJ. Size, shape, and orientation of neurons in the left and right hippocampus: investigation of normal asymmetries and alterations in schizophrenia. Am J Psychiatry. 1997;154:812–8. doi: 10.1176/ajp.154.6.812. [DOI] [PubMed] [Google Scholar]
- 33.Haukvik UK, Tamnes CK, Soderman E, Agartz I. Neuroimaging hippocampal subfields in schizophrenia and bipolar disorder: a systematic review and meta-analysis. J Psychiatr Res. 2018;104:217–26. doi: 10.1016/j.jpsychires.2018.08.012. [DOI] [PubMed] [Google Scholar]
- 34.First MB. Structured Clinical Interview for the DSM (SCID). The Encyclopedia of Clinical Psychology. Hoboken, NJ, USA: John Wiley & Sons; 2015. p. 1-6.
- 35.Kay SR, Fiszbein A, Opler LA. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull. 1987;13:261–76. doi: 10.1093/schbul/13.2.261. [DOI] [PubMed] [Google Scholar]
- 36.Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, et al. The Mini-International Neuropsychiatric Interview (M.I.N.I.): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry. 1998;59:34–57. [PubMed] [Google Scholar]
- 37.Chang SH, Chiang SY, Chiu CC, Tsai CC, Tsai HH, Huang CY, et al. Expression of anti-cardiolipin antibodies and inflammatory associated factors in patients with schizophrenia. Psychiatry Res. 2011;187:341–6. doi: 10.1016/j.psychres.2010.04.049. [DOI] [PubMed] [Google Scholar]
- 38.Gao J, Yi H, Tang X, Feng X, Yu M, Sha W, et al. DNA methylation and gene expression of matrix metalloproteinase 9 gene in deficit and non-deficit schizophrenia. Front Genet. 2018;9:646. doi: 10.3389/fgene.2018.00646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dwir D, Giangreco B, Xin L, Tenenbaum L, Cabungcal JH, Steullet P, et al. MMP9/RAGE pathway overactivation mediates redox dysregulation and neuroinflammation, leading to inhibitory/excitatory imbalance: a reverse translation study in schizophrenia patients. Mol Psychiatry. 2020;25:2889–904. [DOI] [PMC free article] [PubMed]
- 40.Mitelman SA, Newmark RE, Torosjan Y, Chu KW, Brickman AM, Haznedar MM, et al. White matter fractional anisotropy and outcome in schizophrenia. Schizophr Res. 2006;87:138–59. doi: 10.1016/j.schres.2006.06.016. [DOI] [PubMed] [Google Scholar]
- 41.Bucci P, Mucci A, Piegari G, Nobile M, Pini S, Rossi A, et al. Characterization of premorbid functioning during childhood in patients with deficit vs. non-deficit schizophrenia and in their healthy siblings. Schizophr Res. 2016;174:172–76. doi: 10.1016/j.schres.2016.01.032. [DOI] [PubMed] [Google Scholar]
- 42.Gao J, Tang X, Kang J, Xie C, Yu M, Sha W, et al. Correlation between neurocognitive impairment and DNA methylation of MMP-9 gene in patients with deficit schizophrenia. Schizophr Res. 2019;204:455–57. doi: 10.1016/j.schres.2018.10.005. [DOI] [PubMed] [Google Scholar]
- 43.Baglivo V, Cao B, Mwangi B, Bellani M, Perlini C, Lasalvia A, et al. Hippocampal subfield volumes in patients with first-episode psychosis. Schizophr Bull. 2018;44:552–59. doi: 10.1093/schbul/sbx108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Law AJ, Weickert CS, Hyde TM, Kleinman JE, Harrison PJ. Reduced spinophilin but not microtubule-associated protein 2 expression in the hippocampal formation in schizophrenia and mood disorders: molecular evidence for a pathology of dendritic spines. Am J Psychiatry. 2004;161:1848–55. doi: 10.1176/ajp.161.10.1848. [DOI] [PubMed] [Google Scholar]
- 45.Barbeau D, Liang JJ, Robitalille Y, Quirion R, Srivastava LK. Decreased expression of the embryonic form of the neural cell adhesion molecule in schizophrenic brains. Proc Natl Acad Sci USA. 1995;92:2785–9. doi: 10.1073/pnas.92.7.2785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Francis AN, Seidman LJ, Tandon N, Shenton ME, Thermenos HW, Mesholam-Gately RI, et al. Reduced subicular subdivisions of the hippocampal formation and verbal declarative memory impairments in young relatives at risk for schizophrenia. Schizophr Res. 2013;151:154–7. doi: 10.1016/j.schres.2013.10.002. [DOI] [PubMed] [Google Scholar]
- 47.Haukvik UK, Westlye LT, Morch-Johnsen L, Jorgensen KN, Lange EH, Dale AM, et al. In vivo hippocampal subfield volumes in schizophrenia and bipolar disorder. Biol Psychiatry. 2015;77:581–8. doi: 10.1016/j.biopsych.2014.06.020. [DOI] [PubMed] [Google Scholar]
- 48.Niculescu D, Michaelsen-Preusse K, Guner U, van Dorland R, Wierenga CJ, Lohmann C. A BDNF-mediated push-pull plasticity mechanism for synaptic clustering. Cell Rep. 2018;24:2063–74. doi: 10.1016/j.celrep.2018.07.073. [DOI] [PubMed] [Google Scholar]
- 49.Jasinska M, Milek J, Cymerman IA, Leski S, Kaczmarek L, Dziembowska M. miR-132 regulates dendritic spine structure by direct targeting of matrix metalloproteinase 9 mRNA. Mol Neurobiol. 2016;53:4701–12. doi: 10.1007/s12035-015-9383-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kondratiuk I, Leski S, Urbanska M, Biecek P, Devijver H, Lechat B, et al. GSK-3beta and MMP-9 cooperate in the control of dendritic spine morphology. Mol Neurobiol. 2017;54:200–11. doi: 10.1007/s12035-015-9625-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ganguly K, Rejmak E, Mikosz M, Nikolaev E, Knapska E, Kaczmarek L. Matrix metalloproteinase (MMP) 9 transcription in mouse brain induced by fear learning. J Biol Chem. 2013;288:20978–91. doi: 10.1074/jbc.M113.457903. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 52.van der Kooij MA, Fantin M, Rejmak E, Grosse J, Zanoletti O, Fournier C, et al. Role for MMP-9 in stress-induced downregulation of nectin-3 in hippocampal CA1 and associated behavioural alterations. Nat Commun. 2014;5:4995. doi: 10.1038/ncomms5995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Beroun A, Mitra S, Michaluk P, Pijet B, Stefaniuk M, Kaczmarek L. MMPs in learning and memory and neuropsychiatric disorders. Cell Mol life Sci. 2019;76:3207–28. doi: 10.1007/s00018-019-03180-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lepeta K, Kaczmarek L. Matrix metalloproteinase-9 as a novel player in synaptic plasticity and schizophrenia. Schizophr Bull. 2015;41:1003–9. doi: 10.1093/schbul/sbv036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cho KO, Kim SK, Cho YJ, Sung KW, Kim SY. Regional differences in the neuroprotective effect of minocycline in a mouse model of global forebrain ischemia. Life Sci. 2007;80:2030–5. doi: 10.1016/j.lfs.2007.03.005. [DOI] [PubMed] [Google Scholar]
- 56.Lee JH, Lee SR. The effect of Baicalein on hippocampal neuronal damage and metalloproteinase activity following transient global cerebral ischaemia. Phytother Res. 2012;26:1614–9. doi: 10.1002/ptr.4644. [DOI] [PubMed] [Google Scholar]
- 57.Strzelecki D, Kaluzynska O, Szyburska J, Wysokinski A. MMP-9 serum levels in schizophrenic patients during treatment augmentation with sarcosine (Results of the PULSAR Study). Int J Mol Sci. 2016;17:1075. [DOI] [PMC free article] [PubMed]
- 58.Romero JR, Vasan RS, Beiser AS, Au R, Benjamin EJ, DeCarli C, et al. Association of matrix metalloproteinases with MRI indices of brain ischemia and aging. Neurobiol Aging. 2010;31:2128–35. doi: 10.1016/j.neurobiolaging.2008.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Rai AL, Jeswar U. Immunohistochemical colocalization of estrogen receptor-alpha and GABA in adult female rat hippocampus. Ann Neurosci. 2012;19:112–5. doi: 10.5214/ans.0972.7531.190305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Corvino V, Di Maria V, Marchese E, Lattanzi W, Biamonte F, Michetti F, et al. Estrogen administration modulates hippocampal GABAergic subpopulations in the hippocampus of trimethyltin-treated rats. Front Cell Neurosci. 2015;9:433. doi: 10.3389/fncel.2015.00433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Niitsu T, Ishima T, Yoshida T, Hashimoto T, Matsuzawa D, Shirayama Y, et al. A positive correlation between serum levels of mature brain-derived neurotrophic factor and negative symptoms in schizophrenia. Psychiatry Res. 2014;215:268–73. doi: 10.1016/j.psychres.2013.12.009. [DOI] [PubMed] [Google Scholar]
- 62.Shu H, Zheng GQ, Wang X, Sun Y, Liu Y, Weaver JM, et al. Activation of matrix metalloproteinase in dorsal hippocampus drives improvement in spatial working memory after intra-VTA nicotine infusion in rats. J Neurochem. 2015;135:357–67. doi: 10.1111/jnc.13283. [DOI] [PubMed] [Google Scholar]
- 63.Yin L, Li F, Li J, Yang X, Xie X, Xue L, et al. Chronic intermittent ethanol exposure induces upregulation of matrix metalloproteinase-9 in the rat medial prefrontal cortex and hippocampus. Neurochem Res. 2019;44:1593–601. doi: 10.1007/s11064-019-02783-8. [DOI] [PubMed] [Google Scholar]
- 64.Arabska J, Margulska A, Strzelecki D, Wysokinski A. Does metabolic status affect serum levels of BDNF and MMP-9 in patients with schizophrenia? Nord J Psychiatry. 2019:73:515–21. [DOI] [PubMed]
- 65.Domenici E, Wille DR, Tozzi F, Prokopenko I, Miller S, McKeown A, et al. Plasma protein biomarkers for depression and schizophrenia by multi analyte profiling of case-control collections. PLoS One. 2010;5:e9166. doi: 10.1371/journal.pone.0009166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Rybakowski JK, Remlinger-Molenda A, Czech-Kucharska A, Wojcicka M, Michalak M, Losy J. Increased serum matrix metalloproteinase-9 (MMP-9) levels in young patients during bipolar depression. J Affect Disord. 2013;146:286–9. doi: 10.1016/j.jad.2012.07.019. [DOI] [PubMed] [Google Scholar]
- 67.Fiorentino M, Sapone A, Senger S, Camhi SS, Kadzielski SM, Buie TM, et al. Blood-brain barrier and intestinal epithelial barrier alterations in autism spectrum disorders. Mol Autism. 2016;7:49. doi: 10.1186/s13229-016-0110-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Waubant E, Goodkin DE, Gee L, Bacchetti P, Sloan R, Stewart T, et al. Serum MMP-9 and TIMP-1 levels are related to MRI activity in relapsing multiple sclerosis. Neurology. 1999;53:1397–401. doi: 10.1212/wnl.53.7.1397. [DOI] [PubMed] [Google Scholar]
- 69.Avolio C, Filippi M, Tortorella C, Rocca MA, Ruggieri M, Agosta F, et al. Serum MMP-9/TIMP-1 and MMP-2/TIMP-2 ratios in multiple sclerosis: relationships with different magnetic resonance imaging measures of disease activity during IFN-beta-1a treatment. Mult Scler. 2005;11:441–6. doi: 10.1191/1352458505ms1193oa. [DOI] [PubMed] [Google Scholar]
- 70.Montaner J, Rovira A, Molina CA, Arenillas JF, Ribo M, Chacon P, et al. Plasmatic level of neuroinflammatory markers predict the extent of diffusion-weighted image lesions in hyperacute stroke. J Cereb Blood Flow Metab. 2003;23:1403–7. doi: 10.1097/01.WCB.0000100044.07481.97. [DOI] [PubMed] [Google Scholar]
- 71.Barr TL, Latour LL, Lee KY, Schaewe TJ, Luby M, Chang GS, et al. Blood-brain barrier disruption in humans is independently associated with increased matrix metalloproteinase-9. Stroke. 2010;41:e123–8. doi: 10.1161/STROKEAHA.109.570515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Fainardi E, Castellazzi M, Bellini T, Manfrinato MC, Baldi E, Casetta I, et al. Cerebrospinal fluid and serum levels and intrathecal production of active matrix metalloproteinase-9 (MMP-9) as markers of disease activity in patients with multiple sclerosis. Mult Scler. 2006;12:294–301. doi: 10.1191/135248506ms1274oa. [DOI] [PubMed] [Google Scholar]
- 73.Alaiyed S, Bozzelli PL, Caccavano A, Wu JY, Conant K. Venlafaxine stimulates PNN proteolysis and MMP-9-dependent enhancement of gamma power; relevance to antidepressant efficacy. J Neurochem. 2019;148:810–21. doi: 10.1111/jnc.14671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kay SR, Fizbin A, Lindenmayer JP, Opler LA. Positive and negative syndromes in schizophrenia as a function of chronicity. Acta Psychiatr Scand. 1986;74:507–18. doi: 10.1111/j.1600-0447.1986.tb06276.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.