

Abstract
Organoids are three-dimensional structures self-organized from human pluripotent stem cells or primary tissue, potentially serving as a traceable and manipulatable platform to facilitate our understanding of organogenesis. Despite the ongoing advancement in generating organoids of diverse systems, biological applications of in vitro generated organoids remain major challenge in part due to a substantial lack of intricate complexity. The studies of development and regeneration enumerate the essential roles of highly diversified non-epithelial populations such as mesenchyme and endothelium in directing fate specification, morphogenesis and maturation. Furthermore, organoids with physiological and homeostatic functions require direct and indirect inter-organ crosstalk that is seen in organogenesis. We herein review the evolving organoid technology at cell, tissue, organ, and system level with a main emphasis on endoderm-derivatives.
Graphical Abstract
INTRODUCTION
Organogenesis is a robust and complex developmental process of organs, conserved across species. The process has long been studied using embryos of zebrafish, xenopus, chicks and rodents, whereas tension exists around the use of human embryos and fetuses as a research tool in development due to the ongoing ethical debates. Alternatively, the discovery of human pluripotent stem cells (hPSCs), including both embryonic stem cells [1] and induced pluripotent stem cells [2] paved a way for developmental biology research, enabling the study of human organogenesis in vitro.
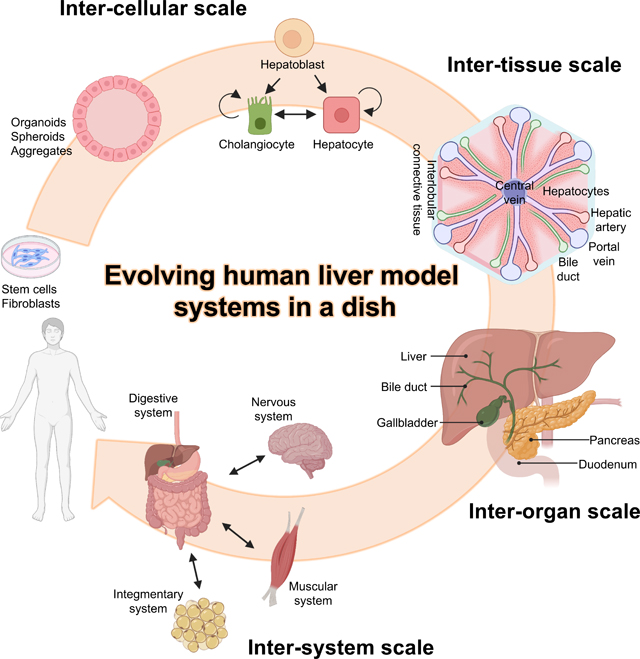
Organoids are three-dimensional (3D) structures self-organized through cell-cell and cell-matrix interactions either from stem cells or adult tissue. In comparison to two-dimensional (2D) monolayer cultures that result in altered gene and protein expressions [3], organoids recapitulate some level of the tissue structure and function [4,5]. Over the past decade, generation of organoids have been tackled mainly by controlling the signaling pathways with small molecules or recombinant proteins, utilizing the molecular principles governing organogenesis. However, rapid advancement in genomics and transcriptomics at single cell level elucidated numerous inter-cellular and inter-tissue interactions through the direct or indirect activation of essential signaling pathways mediating specification, proliferation and maturation. Organoids are further challenged by a substantial lack of additional complexity, for example, the incorporation of the excretory pathways and exocrine/endocrine regulations (Fig1). In the past few years, exploration of organ-specific mesenchyme and neighboring organ communications are providing new possibilities to the field. Here we discuss frontiers of in vitro organogenesis, focusing on the non-cell autonomous reactions and the hierarchical sub-structures of the human body at multiple dimensions (Fig2).
Fig1. Cell-Tissue-Organ-System of liver and equivalent organoid models.

Schematic of the hierarchical sub-structures relating to the liver and its equivalent organoid models. Liver consists cells derived from all three germ layers, namely endoderm derived cholangiocytes and hepatocytes, mesoderm derived Kuppfer cells and stellate cells, and ectoderm derived hepatic neurons. These cells produce epithelial and connective tissues to form the liver which belongs to the digestive system.
Fig2. Organoids at multiple levels and field of use.

Examples of inter-cellular organoids (e.g., hepatic organoids and hepato-biliary organoids), intertissue organoids (e.g., liver organoids with connective tissues), inter-organ organoids (e.g., hepatobiliary-pancreatic organoid) and inter-system organoids (e.g., heart forming organoids with liver domain) and its field of use are shown.
1. Inter-cellular scale organogenesis in a dish.
Endoderm-derived primordial gut is the key source of epithelial cells in respiratory, endocrine and digestive systems including the hepato-biliary-pancreatic (HBP) organs. The two other germ layers, ectoderm and mesoderm also take a part in directing endoderm organogenesis. Epithelial organoids are one prominent example consisting a layer of organ-specific epithelial cells that possess apical-basal polarity induced by the extra-cellular matrix protein[6]. These epithelial organoids can be generated by self-organization of hPSC differentiated cells or from adult stem cells from extracted adult tissues (e.g., liver [7] pancreas [8], intestine[9] and bile duct [10,11]). Because of inherent simplicity, protocols tend to be robust and reproducible, thus encompassing diverse biomedical applications including disease modeling and precision screening [12].
In the liver, hepatocytes and cholangiocytes are the two main epithelial components both differentiated from hepatoblasts. The differentiation of hepatoblasts into cholangiocytes begins at around E13 of mouse development, initially forming a double epithelial layer called the ductal plate around the portal vein. The intrahepatic bile duct (IHBD) is formed from the focal dilations of these epithelial layers, surrounded by the portal mesenchyme. Hepatocytes are epithelialized around E17 in mice aligned by the liver sinusoidal endothelial cells (LSEC) on the basal side and by bile canaliculi on the apical side. Recent advances in inter-cellular scale hepatic organoids are hepatobiliary organoids that contain a mixture of hepatocytes and cholangiocytes [13,14], enabling long-term survival [13] and the formation of bile canaliculi-like network to model cholestatic diseases [14] (Fig 1). Hepatocytes are divided by localization and function into three different zones, zone1 (periportal) to zone3 (pericentral) controlled by WNT signaling from neighboring LSECs. Additional findings relating to the regulation and zonal characteristics within the liver have been reported by incorporating single cell RNA-sequencing [15] and in vivo models [16,17]. Attempts to produce zonal hepatocytes in vitro are using an oxygen gradient in a microscale device [18,19] or genetically engineered cells with inducible WNT signaling [20]. Shinozawa et al. have recently established liver organoids containing cholangiocytes and both pericentral and periportal hepatocytes [12].Thus, inter-cellular scale organoid harbors polarized epithelial cell types serving as a powerful tool to investigate epithelial health and disease due to its relative simplicity (Fig 2, bottom).
3. Inter-tissue scale organogenesis in a dish.
An inter-tissue scale organoid contains multiple tissue types such includes epithelial, connective, hematopoietic, neural and muscular tissues, which function together as a unit to component for organogenesis (Fig2). For instance, the role oforgan-specific endothelial cells has long been discussed in promoting liver[21] and pancreatic organogenesis [22], further characterized by advanced analysis like scRNAseq [23], genome-wide expression, and CpG methylation profiling [24].
The epithelial-mesenchymal interaction is an progressively studied context in development and regeneration [25,26] (Fig3). Underneath epithelium, mesenchyme is motile, lacking tight intracellular adhesions and maintaining surrounding extracellular matrices [26]. Although functional and structural divergence across organs have been an ongoing topic of investigation, recent studies have identified spatial and genetic differences with an organ-specific regional identity [27,28]. Han et al. identified the emergence of organ-specific mesenchyme as early as E8.5 to E9.5 in mouse embryos by utilizing single-cell RNA-sequencing (scRNAseq). There are 17 definable mesenchymal sub-clusters at E9.5 just in the foregut region, including transcription factor signatures enriched explicitly in a neighboring organ-specific manner [27] (Fig4a). Mesenchyme provides paracrine signals for differentiation and maturation like Neuregulin 1 (NRG1) in intestine, a protein that induced cellular diversity validated in enteroid cultures [28] and Wnt for trachea specification [29]. Additional mesenchymal roles involves morphogenetic induction, for example, by promoting medial constriction between the trachea and esophagus [30] (Fig 4b).
Fig3. Increasing studies relating to mesenchyme.

PubMed search counts using the query (epithelial OR epithelium), (mesenchymal OR mesenchyme), (endothelial OR endothelium) demonstrates an accelerated increase in mesenchyme related papers in the past two decades.
Fig4. Organ-specific heterogeneity of mesenchyme and the epithelial-mesenchymal interaction at the early stage of organogenesis.

(a) Summarized schematic of the identification of organ-specific mesenchyme at an early stage of development and the signaling pathways involved for the differentiation by Han et al. [27]. Examples of epithelial-mesenchymal interactions involved in early organogenesis, where (b) the splanchnic mesenchyme induces medial constriction of the foregut to initiate the diversification of the Sox2+ esophageal and Nkx2-1+ tracheal epithelium and (c) the hepatoblasts invaginate and delaminate into the septum transversum mesenchyme for accelerated proliferation. Abbreviations, Single cell RNA sequencing (scRNA-seq), embryonic day (E), Lateral plate mesoderm (LPM), Foregut (FG), Bone morphogenetic protein (B), Fibroblast growth factor (F), Hedgehog (H), Notch (N), Retinoic acid (R), Wnt (W), Septum transversum mesenchyme (STM).
Mesenchyme and endothelial cells (ECs) are also inevitable components in the early development of the liver, when the invaginated hepatoblasts delaminate into the adjacent septum transversum mesenchyme (STM) induced by the endothelial cells aligned between them around E9.5 (Fig 4c). To recapitulate the intricate niche for the development and maturation of the liver, Takebe et al. have generated organoids including mesenchymal stem cells and human umbilical vein endothelial cells (HUVECs) in addition to hepatocytes [31] later produced solely from hPSCs [32]. Since STM is also known to be the progenitors of hepatic stellate cells [33], a differentiation protocol with mesenchymal co-differentiation included hepatic stellate-like cells to recapitulate the inflammatory and fibrotic responses in steatohepatitis models [34]. (Fig 1). Organ-specific ECs in the liver, namely, liver sinusoidal endothelial cells (LSECs) are characterized with the presence of fenestrae. Gata4 was identified as one of the transcriptomic determinants for LSECs [35,36], which resulted in liver hypoplasia in Gata4 knock-out LSECs [36], indicating that organ-specific characterization of the ECs to LSECs contribute to liver development. Multiple STM [27,32,37,38] and LSEC [38–41] differentiation protocols have been established using hPSCs recently, facilitating maturation and better survival of cells in organoids .
The neuronal innervation in the liver is another field with increasing attention. It has been shown that the intrahepatic bile ducts (IHBD) guides the intrahepatic nerve network development [42], which are dominantly sympathetic nerves [43]. They contribute to regeneration [42], regulates the phagocytic activity in resident macrophages [44], and are damaged by metabolic stress [43]. It was also shown that the hepatic vagal sensory afferent nerves indirectly sense the gut microenvironment to maintain the peripheral regulatory T cells [45]. Interestingly, an inter-tissue scale organoid by Guye et al. included co-differentiating neuronal niche within their liver organoid [46]. Although the neuronal tissue addition remains largely unexplored in the liver, further investigation of regulatory signals will be required to self-organize innervated liver organoids.
4. Inter-organ scale organogenesis in a dish
Anatomically related organs are essential to make up a tract (e.g., digestive, respiratory, urinary, musculoskeletal). The inter-organ connections are critical components in a system that refine the function (e.g., hormone regulations between the hypothalamus and pituitary) and prolong the survival by incorporating the excretory tract of toxic substances (e.g., ureter that excretes urine from the kidney). Thus far, inter-organ scale organoid has just recently began explored to attain such interconnections. Suga et al. were the forerunners in establishing inter-organ scale organoids generating an organoid with both hypothalamus and pituitary-like structures [47], allowing in vitro modeling of hypothalamic regulation on congenital pituitary hypoplasia [48]. Attempts to produce kidney and ureter were studied by combining two different progenitors, the ureteric bud and the mesenchymal nephron progenitors, resulting in architectures resembling nephrons and interconnected ureteric epithelium. [49–51]. Koike et al. established an inter-organ scale organoid of HBP domains that possess inter-connections to duodenum-like structures (Fig2) [52]. In this model, the HBP domains emerged spontaneously without extrinsic factors from the boundary of the hPSC derived foregut-midgut spheroids resulting in four organ domains to co-develop in parallel. The specification of HBP was induced by endogenous activation of the retinoic acid (RA) pathway, promoted by the co-developing mesenchyme [52].
Future biomedical application includes the modeling of diseases affecting inter-organ connection that warrants mechanistic and therapeutic investigation. For example, Hes family bHLH transcription factor 1 (HES1) is a known transcription factor that regulate pancreato-biliary segregation, and the Hes1-knockout mice result in conversion of the biliary system to pancreatic cells [53]. This phenotype was recapitulated in the HES1 knockout hPSC derived HBP organoids with less ductal tissue and more pancreatic structures. Thus, inter-organ scale organoids serve as a platform to emulate abnormal organogenesis amenable for genetic and pharmacological manipulation, otherwise inaccessible.
5. Inter-system scale organogenesis in a dish
Organ systems present in our body involve the circulatory, respiratory, musculoskeletal, digestive, integumentary, endocrine, reproductive, and nervous systems. Inter-system scale organoids incorporate orchestrations occurring across multiple systems. Examples of inter-system scale interactions include; Gut-Liver-Brain-axis, an effect on liver and brain via the increased permeability of the gut [54], thyroid hormone regulation of metabolic homeostasis in the liver [55], growth hormone induced insulin-like growth factor synthesis in the liver that regulate the insulin resistance [56], and kernicterus, a neurological disorder induced by hyperbilirubinemia [57]. In vitro cell, tissue and organoid based modeling efforts are limited at this point but begun explored. Studies are emerging with the use of biomedical devices such as organs/body-on-chip, thereby connecting preformed tissue constructs for the study of pathophysiology. However, given that each organ systems are established earlier in development, predominantly 1st trimester, future studies will be needed to understand the essential and non-essential roles of such system-scale interactions in realizing targeted organogenesis in a dish.
Interestingly, human heart forming organoids (HFOs) by Drakhlis et al. modeling the early development of the heart, contained endocardial-like cells surrounded by septum transversum mesenchyme-like structures, and co-differentiating anterior and posterior foregut endoderm tissues [58]. The posterior foregut endoderm population expressed albumin and alfa-fetoprotein (AFP) suggesting its differentiation to hepatoblasts. This model illuminates how the cardiovascular and digestive systems develops synchronously (Fig 2). Additionally, recently established blastoids, a blastocyst-like structure derived from hPSCs [59] or gastruloids, 3D multicellular aggregates that differentiate to form derivatives of the three germ layers organized spatiotemporally [60] might be a starting tool to infer essential inter-system communications, resulting in the methods to induce selective organogenesis in a dish.
Conclusion and perspectives
Organogenesis involves multiple inter-cellular, -tissue, -organ, and -system interactions to generate the robust and complex function of each organ. Although the technologies and knowledge of organoid generation are rapidly evolving, recapitulating the organ in vitro with maturity and robustness is an ongoing challenge. The scale of the organoid should be selected depending on the study since inter-cellular scale organoids are generally simpler with high throughput potential, whereas inter-system scale organoids are more complex with lower throughput capabilities. Organoids can be utilized to study the development in a dish both for discovery and validation and additional discoveries in understanding the developmental process will open opportunities to improve the organoid function.
ACKNOWLEDGEMENTS
We would like to express sincere gratitude to the other Takebe lab members, Wells-Zorn lab members, PSCF lab members for their support and excellent technical assistance. Illustrations were created using Biorender.com. This work was supported by Cincinnati Children’s Research Foundation grant, NIH Director’s New Innovator Award (DP2 DK128799-01) and CREST (20gm1210012h0001) grant from Japan Agency for Medical Research and Development (AMED) to TT. This work was also supported by an NIH grant UH3 DK119982, Cincinnati Center for Autoimmune Liver Disease Fellowship Award, PHS Grant P30 DK078392 (Integrative Morphology Core and Pluripotent Stem Cell and Organoid Core) of the Digestive Disease Research Core Center in Cincinnati, the Falk Catalyst Research Awards Program, JST Moonshot R&D Grant Number JPMJMS2033 and JPMJMS2022, Takeda Science Foundation award, Mitsubishi Foundation award and AMED JP19fk0210037, JP19bm0704025, JP19fk0210060, JP19bm0404045, and JSPS JP18H02800, 19K22416. TT is a New York Stem Cell Foundation –Robertson Investigator.
Footnotes
Conflict of Interest
These authors declare no conflict of interest associated with the current manuscript.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM: Embryonic Stem Cell Lines Derived from Human Blastocysts. Science 1998, 282:1145–1147. [DOI] [PubMed] [Google Scholar]
- 2.Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S: Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131:861–872. [DOI] [PubMed] [Google Scholar]
- 3.Jensen C, Teng Y: Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front Mol Biosci 2020, 7:33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Takebe T, Wells JM: Organoids by design. Science 2019, 364:956–959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Marsee A, Roos FJM, Verstegen MMA, Marsee A, Roos F, Verstegen M, Clevers H, Vallier L, Takebe T, Huch M, et al. : Building consensus on definition and nomenclature of hepatic, pancreatic, and biliary organoids. Cell Stem Cell 2021, 28:816–832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Co JY, Margalef-Català M, Li X, Mah AT, Kuo CJ, Monack DM, Amieva MR: Controlling Epithelial Polarity: A Human Enteroid Model for Host-Pathogen Interactions.Cell Rep 2019, 26:2509–2520.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hu H, Gehart H, Artegiani B, LÖpez-Iglesias C, Dekkers F, Basak O, van Es J, Chuva de Sousa Lopes SM, Begthel H, Korving J, et al.: Long-Term Expansion of Functional Mouse and Human Hepatocytes as 3D Organoids. Cell 2018, 175:1591–1606.e19. [DOI] [PubMed] [Google Scholar]
- 8.Georgakopoulos N, Prior N, Angres B, Mastrogiovanni G, Cagan A, Harrison D, Hindley CJ, Arnes-Benito R, Liau SS, Curd A, et al. : Long-term expansion, genomic stability and in vivo safety of adult human pancreas organoids. BMC Dev Biol 2020, 20:1–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sugimoto S, Kobayashi E, Fujii M, Ohta Y, Arai K, Matano M, Ishikawa K, Miyamoto K, Toshimitsu K, Takahashi S, et al. : An organoid-based organ-repurposing approach to treat short bowel syndrome.Nature 2021, 592:99–104. [DOI] [PubMed] [Google Scholar]
- 10.Sampaziotis F, Muraro D, Tysoe OC, Sawiak S, Beach TE, Godfrey EM, Upponi SS, Brevini T, Wesley BT, Garcia-Bernardo J, et al. : Cholangiocyte organoids can repair bile ducts after transplantation in the human liver.Science 2021, 371:839–846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Aloia L, McKie MA, Vernaz G, Cordero-Espinoza L, Aleksieva N, van den Ameele J, Antonica F, Font-Cunill B, Raven A, Aiese Cigliano R, et al. : Epigenetic remodelling licences adult cholangiocytes for organoid formation and liver regeneration.Nat Cell Biol 2019, 21:1321–1333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Shinozawa T, Kimura M, Cai Y, Saiki N, Yoneyama Y, Ouchi R, Koike H, Maezawa M, Zhang R-R, Dunn A, et al. : High-Fidelity Drug-Induced Liver Injury Screen Using Human Pluripotent Stem Cell-Derived Organoids.Gastroenterology 2021, 160:831–846.e10. •• Liver organoids derived from hPSCs, containing diverse zonal hepatocytic populations that are polarized with bile canaliculi-like structures enabling unidirectional bile acid transport to study bile transport function.
- 13.Akbari S, Sevinç GG, Ersoy N, Basak O, Kaplan K, Sevinç K, Ozel E, Sengun B, Enustun E, Ozcimen B, et al. : Robust, Long-Term Culture of Endoderm-Derived Hepatic Organoids for Disease Modeling. Stem Cell Reports 2019, 13:627–641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Ramli MN Bin, Lim YS, Koe CT, Demircioglu D, Tng W, Gonzales KAU, Tan CP, Szczerbinska I, Liang H, Soe EL, et al. : Human Pluripotent Stem Cell-Derived Organoids as Models of Liver Disease. Gastroenterology 2020, 159:1471–1486.e12. •• Generation of organoids containing both hepatocytes and cholangiocytes with functional bile canaliculi system enabling to study drug induced cholestasis and ductular reactions in nonalcoholic fatty liver disease.
- 15.Aizarani N, Saviano A, Sagar, Mailly L, Durand S, Herman JS, Pessaux P, Baumert TF, Grün D : A human liver cell atlas reveals heterogeneity and epithelial progenitors. Nature 2019, 572:199–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. He L, Pu W, Liu X, Zhang Z, Han M, Li Y, Huang X, Han X, Li Y, Liu K, et al. : Proliferation tracing reveals regional hepatocyte generation in liver homeostasis and repair.Science 2021, 371. • Incorporating dual recombinases for lineage tracing purposes, the authors developed proliferation tracer (ProTracer) system, which revealed that hepatocyte proliferation were mainly in zone 2 during liver homeostasis.
- 17.Wei Y, Wang YG, Jia Y, Li L, Yoon J, Zhang S, Wang Z, Zhang Y, Zhu M, Sharma T, et al. : Liver homeostasis is maintained by midlobular zone 2 hepatocytes.Science 2021, 371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Matsumoto S, Safitri AR, Danoy M, Maekawa T, Kinoshita H, Shinohara M, Sakai Y, Fujii T, Leclerc E: Investigation of the hepatic respiration and liver zonation on rat hepatocytes using an integrated oxygen biosensor in a microscale device. Biotechnol Prog 2019, 35:1–13. [DOI] [PubMed] [Google Scholar]
- 19.Tonon F, Giobbe GG, Zambon A, Luni C, Gagliano O, Floreani A, Grassi G, Elvassore N: In vitro metabolic zonation through oxygen gradient on a chip. Sci Rep 2019, 9:1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wahlicht T, Vièyres G, Bruns SA, Meumann N, Büning H, Hauser H, Schmitz I, Pietschmann T, Wirth D: Controlled Functional Zonation of Hepatocytes in Vitro by Engineering of Wnt Signaling. ACS Synth Biol 2020, 9:1638–1649. [DOI] [PubMed] [Google Scholar]
- 21.Matsumoto K, Yoshitomi H, Rossant J, Zaret KS: Liver Organogenesis Promoted by Endothelial Cells Prior to Vascular Function. Science 2001, 294:559 LP –563. [DOI] [PubMed] [Google Scholar]
- 22.Lammert E, Cleaver O, Melton D: Induction of Pancreatic Differentiation by Signals from Blood Vessels. Science 2001, 294:564 LP –567. [DOI] [PubMed] [Google Scholar]
- 23. Paik DT, Tian L, Williams IM, Rhee S, Zhang H, Liu C, Mishra R, Wu SM, Red-Horse K, Wu JC: Single-Cell RNA Sequencing Unveils Unique Transcriptomic Signatures of Organ-Specific Endothelial Cells. Circulation 2020, 142:1848–1862. •• Incorportating scRNAseq to profile tissue specific mouse endothelial cells. The authors identified novel markers of ECs specific to organs, and the signaling network to maintain their organ-specific characteristic.
- 24.Jamil MA, Singer H, Al-Rifai R, Nüsgen N, Rath M, Strauss S, Andreou I, Oldenburg J, El-Maarri O: Molecular analysis of fetal and adult primary human liver sinusoidal endothelial cells: A comparison to other endothelial cells. Int J Mol Sci 2020, 21:1–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ferretti E, Hadjantonakis AK: Mesoderm specification and diversification: from single cells to emergent tissues. Curr Opin Cell Biol 2019, 61:110–116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Carmona R, Ariza L, Cano E, Jiménez-Navarro M, Muñoz-Chápuli R: Mesothelial-mesenchymal transitions in embryogenesis. Semin Cell Dev Biol 2019, 92:37–44. [DOI] [PubMed] [Google Scholar]
- 27. Han L, Chaturvedi P, Kishimoto K, Koike H, Nasr T, Iwasawa K, Giesbrecht K, Witcher PC, Eicher A, Haines L, et al. : Single cell transcriptomics identifies a signaling network coordinating endoderm and mesoderm diversification during foregut organogenesis. Nat Commun 2020, 11:4158. •• Application of scRNAseq to profile the early foregut region of mouse embryos from E8.5 to E9.5. The authors identified a diversity of splanchnic mesoderm populations with neighboring organ specific characteristics and the signaling network of epithelial-mesenchymal interactions coordinating organogenesis
- 28.Holloway EM, Czerwinski M, Tsai YH, Wu JH, Wu A, Childs CJ, Walton KD, Sweet CW, Yu Q, Glass I, et al. : Mapping Development of the Human Intestinal Niche at Single-Cell Resolution. Cell Stem Cell 2021, 28:568–580.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kishimoto K, Furukawa KT, Luz-Madrigal A, Yamaoka A, Matsuoka C, Habu M, Alev C, Zorn AM, Morimoto M: Bidirectional Wnt signaling between endoderm and mesoderm confers tracheal identity in mouse and human cells. Nat Commun 2020, 11:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nasr T, Mancini P, Rankin SA, Edwards NA, Agricola ZN, Kenny AP, Kinney JL, Daniels K, Vardanyan J, Han L, et al. : Endosome-Mediated Epithelial Remodeling Downstream of Hedgehog-Gli Is Required for Tracheoesophageal Separation. Dev Cell 2019, 51:665–674.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Takebe T, Sekine K, Enomura M, Koike H, Kimura M, Ogaeri T, Zhang RR, Ueno Y, Zheng YW, Koike N, et al. : Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499:481–484. [DOI] [PubMed] [Google Scholar]
- 32.Takebe T, Sekine K, Kimura M, Yoshizawa E, Ayano S, Koido M, Funayama S, Nakanishi N, Hisai T, Kobayashi T, et al. : Massive and Reproducible Production of Liver Buds Entirely from Human Pluripotent Stem Cells.Cell Rep 2017, 21:2661–2670. [DOI] [PubMed] [Google Scholar]
- 33.Asahina K, Zhou B, Pu WT, Tsukamoto H: Septum transversum-derived mesothelium gives rise to hepatic stellate cells and perivascular mesenchymal cells in developing mouse liver.Hepatology 2011, 53:983–995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ouchi R, Togo S, Kimura M, Shinozawa T, Koido M, Koike H, Thompson W, Karns RA, Mayhew CN, McGrath PS, et al. : Modeling Steatohepatitis in Humans with Pluripotent Stem CellDerived Organoids. Cell Metab 2019, 30:374–384.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.De Haan W, Øie C, Benkheil M, Dheedene W, Vinckier S, Coppiello G, Aranguren XL, Beerens M, Jaekers J, Topal B, et al. : Unraveling the transcriptional determinants of liver sinusoidal endothelial cell specialization. Am J Physiol - Gastrointest Liver Physiol 2020, 318:G803–G815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Géraud C, Koch PS, Zierow J, Klapproth K, Busch K, Olsavszky V, Leibing T, Demory A, Ulbrich F, Diett M, et al. : GATA4-dependent organ-specific endothelial differentiation controls liver development and embryonic hematopoiesis. J Clin Invest 2017, 127:1099–1114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Miyoshi M, Kakinuma S, Kamiya A, Tsunoda T, Tsuchiya J, Sato A, Kaneko S, Nitta S, Kawai-Kitahata F, Murakawa M, et al. : LIM homeobox 2 promotes interaction between human iPS-derived hepatic progenitors and iPS-derived hepatic stellate-like cells. Sci Rep 2019, 9:1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Koui Y, Kido T, Ito T, Oyama H, Chen S-W, Katou Y, Shirahige K, Miyajima A: An In Vitro Human Liver Model by iPSC-Derived Parenchymal and Non-parenchymal Cells.Stem cell reports 2017, 9:490–498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ardalani H, Sengupta S, Harms V, Vickerman V, Thomson JA, Murphy WL: 3-D culture and endothelial cells improve maturity of human pluripotent stem cell-derived hepatocytes. Acta Biomater 2019, 95:371–381. [DOI] [PubMed] [Google Scholar]
- 40.Gage BK, Liu JC, Innes BT, MacParland SA, McGilvray ID, Bader GD, Keller GM: Generation of Functional Liver Sinusoidal Endothelial Cells from Human Pluripotent Stem-Cell-Derived Venous Angioblasts.Cell Stem Cell 2020, 27:254–269.e9. [DOI] [PubMed] [Google Scholar]
- 41.De Smedt J, van Os EA, Talon I, Ghosh S, Toprakhisar B, Furtado Madeiro Da Costa R, Zaunz S, Vazquez MA, Boon R, Baatsen P, et al. : PU.1 drives specification of pluripotent stem cell-derived endothelial cells to LSEC-like cells. Cell Death Dis 2021, 12:84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Tanimizu N, Ichinohe N, Mitaka T: Intrahepatic bile ducts guide establishment of the intrahepatic nerve network in developing and regenerating mouse liver.Development 2018, 145. [DOI] [PubMed] [Google Scholar]
- 43. Liu K, Yang L, Wang G, Liu J, Zhao X, Wang Y, Li J, Yang J: Metabolic stress drives sympathetic neuropathy within the liver.Cell Metab 2021, 33:666–675.e4. • Identification of a sympathetic neuronal innervation and network formation in the liver. The first study to reveal the hepatic sympathetic neuropathy under metabolic stress.
- 44.Fonseca RC, Bassi GS, Brito CC, Rosa LB, David BA, Araújo AM, Nóbrega N, Diniz AB, Jesus ICG, Barcelos LS, et al. : Vagus nerve regulates the phagocytic and secretory activity of resident macrophages in the liver. Brain Behav Immun 2019, 81:444–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Teratani T, Mikami Y, Nakamoto N, Suzuki T, Harada Y, Okabayashi K, Hagihara Y, Taniki N, Kohno K, Shibata S, et al. : The liver–brain–gut neural arc maintains the Treg cell niche in the gut. Nature 2020, 585:591–596. [DOI] [PubMed] [Google Scholar]
- 46.Guye P, Ebrahimkhani MR, Kipniss N, Velazquez JJ, Schoenfeld E, Kiani S, Griffith LG, Weiss R: Genetically engineering self-organization of human pluripotent stem cells into a liver bud-like tissue using Gata6.Nat Commun 2016, 7:10243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Suga H, Kadoshima T, Minaguchi M, Ohgushi M, Soen M, Nakano T, Takata N, Wataya T, Muguruma K, Miyoshi H, et al. : Self-formation of functional adenohypophysis in three-dimensional culture.Nature 2011, 480:57–62. [DOI] [PubMed] [Google Scholar]
- 48.Matsumoto R, Suga H, Aoi T, Bando H, Fukuoka H, Iguchi G, Narumi S, Hasegawa T, Muguruma K, Ogawa W, et al. : Congenital pituitary hypoplasia model demonstrates hypothalamic OTX2 regulation of pituitary progenitor cells.J Clin Invest 2020, 130:641–654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Tsujimoto H, Kasahara T, Sueta S ichi, Araoka T, Sakamoto S, Okada C, Mae S ichi, Nakajima T, Okamoto N, Taura D, et al. : A Modular Differentiation System Maps Multiple Human Kidney Lineages from Pluripotent Stem Cells. Cell Rep 2020, 31:107476. [DOI] [PubMed] [Google Scholar]
- 50.Taguchi A, Nishinakamura R: Higher-Order Kidney Organogenesis from Pluripotent Stem Cells.Cell Stem Cell 2017, 21:730–746.e6. [DOI] [PubMed] [Google Scholar]
- 51.Howden SE, Wilson SB, Groenewegen E, Starks L, Forbes TA, Tan KS, Vanslambrouck JM, Holloway EM, Chen YH, Jain S, et al. : Plasticity of distal nephron epithelia from human kidney organoids enables the induction of ureteric tip and stalk. Cell Stem Cell 2021, 28:671–684.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Koike H, Iwasawa K, Ouchi R, Maezawa M, Giesbrecht K, Saiki N, Ferguson A, Kimura M, Thompson WL, Wells JM, et al. : Modelling human hepato-biliary-pancreatic organogenesis from the foregut–midgut boundary. Nature 2019, 574:112–116. •• Using the boundary interactions between the anterior and the posterior gut spheroids derived from hPSCs, the authors show the spontaneous emergence of hepato-biliary-pancreatic anlage at the foregut-midgut boundary in the absence of extrinsic factors. This is the first study to generate inter-connected hepato-biliary-pancreatic organoid.
- 53.Sumazaki R, Shiojiri N, Isoyama S, Masu M, Keino-Masu K, Osawa M, Nakauchi H, Kageyama R, Matsui A: Conversion of biliary system to pancreatic tissue in Hes1-deficient mice.Nat Genet 2004, 36:83–87. [DOI] [PubMed] [Google Scholar]
- 54.Shukla PK, Meena AS, Dalal K, Canelas C, Samak G, Pierre JF, Rao R: Chronic stress and corticosterone exacerbate alcohol-induced tissue injury in the gut-liver-brain axis.Sci Rep 2021, 11:826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ritter MJ, Amano I, Hollenberg AN: Thyroid Hormone Signaling and the Liver.Hepatology 2020, 72:742–752. [DOI] [PubMed] [Google Scholar]
- 56.Riedel EO, Hinrichs A, Kemter E, Dahlhoff M, Backman M, Rathkolb B, Prehn C, Adamski J, Renner S, Blutke A, et al. : Functional changes of the liver in the absence of growth hormone (GH) action - Proteomic and metabolomic insights from a GH receptor deficient pig model.Mol Metab 2020, 36:100978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Miura S, Tsuchie K, Fukushima M, Arauchi R, Tsumori T, Otsuki K, Hayashida M, Hashioka S, Wake R, Miyaoka T, et al. : Normalizing hyperactivity of the Gunn rat with bilirubin-induced neurological disorders via ketanserin.Pediatr Res 2021, doi: 10.1038/s41390-021-01446-1. [DOI] [PubMed] [Google Scholar]
- 58. Drakhlis L, Biswanath S, Farr C-MM, Lupanow V, Teske J, Ritzenhoff K, Franke A, Manstein F, Bolesani E, Kempf H, et al. : Human heart-forming organoids recapitulate early heart and foregut development.Nat Biotechnol 2021, doi: 10.1038/s41587-021-00815-9. •• Heart forming organoids composed of not only myocardium surrounded by septum transversum mesenchyme, but also both anterior and posterior foregut endoderm tissues recapitulating the development of the early native heart anlagen and its interaction with the foregut endoderm.
- 59. Yu L, Wei Y, Duan J, Schmitz DA, Sakurai M, Wang L, Wang K, Zhao S, Hon GC, Wu J: Blastocyst-like structures generated from human pluripotent stem cells.Nature 2021, 591:620–626. • Generation of blastoids derived from naïve hPSC in 3D cultures, recapitulating the human blastocyst by morphology, size and cell number as well as the composition of different cell lineages.
- 60. Moris N, Anlas K, van den Brink SC, Alemany A, Schröder J, Ghimire S, Balayo T, van Oudenaarden A, Martinez Arias A: An in vitro model of early anteroposterior organization during human development. Nature 2020, 582:410–415. • Generation of gastruloids derived from hESCs in 3D cultures that possess the potential to differentiate into three germ layers in a spatiotemporal manner, recapitulating the human gastrulation.