

SUMMARY
The organismal body axes that are formed during embryogenesis are intimately linked to intrinsic asymmetries established at the cellular scale in oocytes 1. However, the mechanisms that generate cellular asymmetries within the oocyte, and then transduce that polarity to organismal scale body axes are poorly understood outside of select model organisms. Here, we report an axis-defining event in meiotic oocytes of the sea star Patiria miniata. Dishevelled is a cytoplasmic Wnt pathway effector required for axis development in diverse species 2–4, but the mechanisms governing its function and distribution remain poorly defined. Using time-lapse imaging, we find that Dishevelled localizes uniformly to puncta throughout the cell cortex in Prophase I-arrested oocytes, but becomes enriched at the vegetal pole following meiotic resumption through a dissolution-reassembly mechanism. This process is driven by an initial disassembly phase of Dvl puncta, followed by selective reformation of Dvl assemblies at the vegetal pole. Rather than being driven by Wnt signaling, this localization behavior is coupled to meiotic cell cycle progression and influenced by Lamp1+ endosome association and Frizzled receptors pre-localized within the oocyte cortex. Our results reveal a cell cycle-linked mechanism by which maternal cellular polarity is transduced to the embryo through spatially-regulated Dishevelled dynamics.
eTOC blurb
The primary body axis in diverse species is prefigured within the egg. Using sea star oocytes, Swartz et al. find that the Wnt pathway protein Dishevelled (Dvl) localizes to the vegetal pole to direct body axis specification. Dvl re-localization occurs in meiosis through coupled dissolution and reformation of dynamic molecular assemblies.
Graphical Abstract
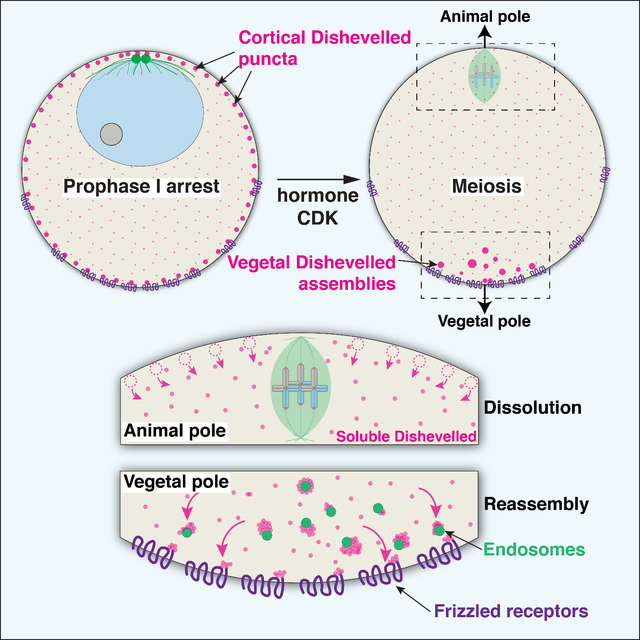
RESULTS AND DISCUSSION
Dishevelled is required for specification of the primary body axis in sea star embryos
Developmental determinants of the primary body axis are established maternally within the oocytes of diverse animal species 5. In many organisms, including echinoderms such as sea stars, the cytologically-defined Animal-Vegetal axis of the oocyte correlates with and prefigures the Anterior-Posterior axis of the embryo (Figure1A) 5–7. The Wnt/beta-catenin pathway is an important determinant of the primary body axis, and provides a link between oocyte asymmetries and embryonic polarity 8–10. However, the mechanisms by which the Wnt pathway becomes asymmetrically activated within subsets of embryonic cells to direct axis development are poorly defined. Dishevelled (Dvl) is an essential cytoplasmic transducer of Wnt signaling, and is localized asymmetrically in diverse organisms, including sea urchin eggs and embryos and in Xenopus oocytes following fertilization 11–14. We sought to define the mechanisms that govern maternal Dvl localization and function, which have been challenging to study in these other systems.
Figure 1. Dishevelled is required for posterior identity and undergoes relocalization in meiosis.

(A) Schematic of sea star meiosis and development. The Animal-Vegetal axis in oocytes, defined by the site of the meiotic divisions, correlates with the Anterior-Posterior axis in the embryo. (B) Depletion of Dvl by morpholino injection disrupts primary invagination at the posterior pole (25 hours control n=35, Dvl knockdown n=26 embryos), and the formation of endomesodermal structures in subsequent embryogenesis (72 hours control n=45, Dvl knockdown n=50). Scale bars = 50 μm. (C) Expression of posteriorly expressed cWnt pathway genes is downregulated with Dvl knockdown as measured by qPCR. The anteriorly expressed factor Six3 is unaffected (mean and standard deviation of 2 biological replicates each performed in technical triplicate). (D) 4D partial z-stacks of live oocyte expressing Dvl-GFP and EMTB-mCherry. Upper images are of Dvl-GFP channel alone in inverted grayscale. In Prophase I arrest (t=00:00), Dvl localizes to uniformly distributed puncta at the cortex. In addition to the centrosomal microtubule array at the animal pole, EMTB-mCherry revealed a band of microtubule density at the vegetal pole. Following hormonal stimulation, Dvl puncta dissolve (t=16:40), and then reform at the vegetal pole (t=50:00). Scale bars = 50 μm. (E) Kymograph depiction of the oocyte in (D), defined by the white dashed lines in the lower left image. A transverse kymograph of cytoplasmic Dvl, along with circumferential kymographs for cortical Dvl and microtubules are provided. Regions of interest to which the kymographs correspond are outlined in (D).
To define the function of Dvl in body axis specification, we used the sea star Patiria miniata, which provides a powerful system to study oogenesis, meiosis, and embryogenesis 15. We found that Dvl depletion using morpholino injection disrupted gastrulation and the expression of known Wnt-beta catenin target genes in the posterior embryo (Figure 1B,C) 16, consistent with an essential role for Dvl in primary axis specification and prior dominant negative studies in sea urchins17. We confirmed Dvl depletion by Western blotting of 24-hour embryos using antibodies raised against P. miniata Dvl (Figure S1A). Following 72 hours of development, wild-type embryos form a differentiated tripartite gut and mesodermally-derived coelomic pouch structures. However, Dvl knockdown embryos failed to form normal endomesodermal structures, indicative of a failure of endomesoderm specification (Figure 1B,C). Anterior-Posterior patterning is initiated by beta-catenin nuclearization, endomesoderm specification, and the secretion of factors that restrict anterior fate 16,18,19. These phenotypes are therefore consistent with Dvl acting to specify endomesodermal fates at the posterior pole in the sea star embryo.
Dishevelled localization becomes polarized during meiotic cell cycle progression
Given the essential role of Dvl in axis determination, we next sought to define its localization during the oocyte-to-embryo transition by live imaging (Figure 1D, Video S1). In Prophase I-arrested oocytes, Dvl-GFP localized to puncta uniformly throughout the cell cortex, as well as diffusely to the cytoplasm (Figure 1D,E). However, following hormonal stimulation, just prior to germinal vesicle breakdown (GVBD), these Dvl-GFP puncta were lost (Figure 1D,E, 16:40). Subsequently, Dvl-GFP assemblies reappeared in the vegetal subcortical cytoplasm as the first meiotic spindle formed. These structures increased in prevalence as the first meiotic division completed (Video S1). Following the completion of meiosis, the first mitotic spindle is oriented perpendicularly with the cleavage plane parallel to the animal-vegetal axis (see also 20). Due to the localization of Dvl to the vegetal pole, this cleavage pattern distributes the vegetal Dvl domain equally to both blastomeres where it remains asymmetrically localized (Figure S1C).
To confirm the behavior observed for ectopically-expressed Dvl-GFP, we additionally visualized endogenous Dvl mRNA and protein. Using fluorescent in situ hybridization, we detected Dvl mRNA uniformly throughout the oocyte and embryo (Figure S1B). By immunofluorescence, endogenous Dvl protein localized throughout Prophase I-arrested oocytes, but formed assemblies that were enriched at the vegetal pole in meiosis (Figure S1C), consistent with our observations of Dvl-GFP. Immunostaining of embryos in which Dvl was knocked down indicated that the antibody selectively recognized Dvl protein (Figure S1D). In summary, Dvl localization becomes polarized in a process coupled temporally to meiotic cell cycle progression, which could serve as a critical event in primary body axis specification during embryogenesis.
Dvl localization to the vegetal pole is correlated temporally with cytological changes during meiotic progression. We therefore considered whether key cell cycle regulators influence Dvl localization. Sea star meiosis occurs largely in the absence of major changes in protein abundance 21. Consistent with this general protein stability, translational inhibition with emetine did not affect Dvl vegetal pole relocalization (Figure S2A). In contrast, inhibiting cyclin-dependent kinase (Cdk) activity with flavopiridol to disrupt meiotic progression prevented vegetal Dvl localization (Figure S2A). Cdk and other meiotic kinases are opposed by the protein phosphatases PP1 and PP2A. PP1/PP2A inhibition is sufficient to induce partial meiotic resumption, even in the absence of hormonal stimulation 21. PP1 and PP2A inhibition using Calyculin A in arrested oocytes was sufficient to induce the dissolution of uniformly distributed Dvl puncta (Figure S2B). However, it did not result in vegetal Dvl localization. Collectively, we propose that the Dvl dissolution and reassembly phases are separable events that require distinct post-translational mechanisms. Dvl reassembly may require the activity of a kinase downstream of hormonal stimulation, possibly including Cdk itself.
Polarized dissolution and reassembly coupled to meiotic progression drives Dvl localization
We next sought to define the mechanisms that underlie Dvl localization to the vegetal pole, including whether this requires cytoplasmic flow or active transport along cytoskeletal elements. We did not observe substantial changes in the bulk cytoplasmic Dvl-GFP concentration along the AV axis when viewed as kymograph over meiotic time (Figure 1E). Furthermore, we did not observe Dvl puncta moving substantial distances across the oocyte, arguing against the presence of directional transport. In addition, pharmacological disruption of microtubule or actin networks did not prevent Dvl vegetal enrichment following hormonal stimulation (Figure S2 C,D), suggesting that Dvl distribution occurs through alternative mechanisms.
Given the lack of visible transport of Dvl puncta across the oocyte, we instead hypothesized that Dvl enrichment to the vegetal pole is a regionalized process. To analyze this behavior, we specifically imaged the vegetal pole at higher magnification and performed particle tracking analyses. This unbiased analysis revealed two discrete populations of Dvl granules based on proximity to the cell surface (Figure 2A, Video S2) - one in the cytoplasm, and another in close association (<3.25 μm) with the cell surface (Video S2). We therefore tracked these two populations over time separately to assess their differential behaviors (Video S2, middle and bottom panels). Mean squared displacement (MSD) measurements indicated that the vegetal cortex granules are largely immobile and achieve lower velocities, whereas the cytoplasmic granules traverse greater distances (Figure 2B, S2D). However, on a population level, the movements of the cytoplasmic granules were not directionally biased, based on the mean velocities of all particle trajectories across angles relative to the Animal-Vegetal axis (Figure 2C). We therefore conclude that polarized Dvl localization is not a result of directional transport or flow.
Figure 2. Dishevelled localization by polarized dissolution and condensation.

(A) Time lapse vegetal pole images of oocyte expressing Dvl-GFP and EMTB-mCherry at high magnification (scale bar = 10 μm). Appearance of rows of Dvl-GFP puncta are due to z-step size during rapid acquisition followed by maximal projection. Top images are of the Dvl-GFP channel alone in inverted grayscale. (B) Mean squared displacement for Dvl assemblies over time, indicating cortex assemblies are less mobile. (C) Mean velocity of cytoplasmic assemblies across angles relative to the Animal-Vegetal axis reveals no directionally biased movement. (D) Area quantification of Dvl assemblies from maximally projected 3D time lapse shown in (A). The images were segmented to separately measure cortex-associated or cytoplasmic assemblies. Area measurements reveal a period of dissolution and condensation for both cytoplasmic and cortex Dvl assemblies. (B,D) Error bars represents the standard error of puncta over time, the sample size of which is provided in Figure S2E. (E) Magnified views of single Dvl puncta during the dissolution or condensation period. Top row images are 1.5 μm wide, and lower row images are 3.0 μm wide.
Dvl self-assembles into higher order structures via its N-terminal DIX domain and central DEP domain 22. We therefore hypothesized that biased assembly and disassembly from a uniformly-available pool of monomers could favor Dvl puncta assembly at the vegetal pole, but disfavor Dvl assembly in the animal region of the oocyte. To test this, we measured the size of Dvl assemblies over time, using the area of maximally projected time lapse videos as a proxy. In Prophase I-arrested oocytes, the cortex-associated granules were larger and more abundant than the cytoplasmic granules. However, following hormonal stimulation, the size of cortex-associated Dvl puncta decreased rapidly, displaying behavior characteristic of dissolution (Figure 2D,E, S2C). As meiosis progressed, both the cortex and cytoplasmic granules began to reform, reaching a mean area of approximately 0.5 μm2 (Figure 2D,E). Given this strikingly dynamic behavior in response to hormonal signaling, we conclude that Dvl localization is a result of polarized dissolution and reassembly. This process occurs through biochemical signals downstream of meiotic resumption, and is spatially regulated to asymmetrically favor Dvl self-association at the vegetal pole.
Dvl granules are dynamic molecular assemblies
The rapid dissolution and reformation behavior suggests that Dvl incorporates into dynamic molecular assemblies. Indeed, by time lapse imaging, we identified fusion events amongst the mobile cytoplasmic Dvl granules suggesting that these assemblies engage in higher-order molecular interactions (Figure 3A, Video S3). To test the nature of Dvl binding within the larger assemblies, we conducted fluorescent recovery after photobleaching (FRAP) on the cortical Dvl puncta in Prophase I-arrested oocytes, and the immobile puncta at the vegetal cortex after Dvl re-localization (Figure 3B,C). We found that Dvl exchanged readily between the puncta and cytoplasmic pool with a halftime for recovery of ~2.5 minutes for both cortical and vegetal cortex-localized granules. However, although Prophase I assemblies recovered fully (mobile fraction = 1.05+/−0.32 S.D.), the vegetal cortically-localized granules displayed lower recovery (mobile fraction = 0.66+/−0.19 S.D.) (Figure 3C). These differences may reflect different binding modes for Dvl in these different contexts, such as a distinct vegetal receptor for Dvl in meiosis from that in Prophase I-arrested oocytes. Although these data do not allow us to determine conclusively the physical nature of the phase transition state that Dvl undergoes, based on the dynamic properties measured by FRAP and the ability of assemblies to fuse, we propose that Dvl forms dynamic biomolecular assemblies, which may form by phase-separation or using structured protein-protein interactions.
Figure 3. Dishevelled forms dynamic molecular assemblies and associates with cytoplasmic organelles.

(A) Time lapse video stills of a cytoplasmic Dvl assembly (in area schematized to the right) undergoing fusion. Arrowheads indicate the two separate assemblies. (B) FRAP analysis of uniform cortex puncta in Prophase I, or vegetal cortical domain (VCD) puncta in meiosis. Stills from example FRAP experiments are provided. (C) FRAP quantification of Prophase I or VCD granules (Pro I n=6 oocytes, mean of 3 puncta each, VCD n=4 oocytes, mean of 3 puncta each). (D) Vegetal pole image of a live oocyte expressing Lamp1–3xGFP and Dvl-HaloTag showing cytoplasmic Dvl assemblies are associated with endosomes. (E) Fraction of Dvl assemblies that co-localize with Lamp1-positive endosomes in Prophase I, Meiosis I, and Meiosis II. Black circle is the mean value over N=3 oocytes while blue circle is the number from individual oocyte. Error bars represent the standard deviation over 10 minute time lapse videos. (F) Zoom views of insets indicated in (D). Images are 4.88 μm wide. (G) Still images of individual Dvl-endosome complexes tracked over time. Images are 16.93 μm wide.
Dvl assemblies associate with Lamp1-positive endosomes
The molecular interactions and cellular states driving Dvl self-association into punctate structures are not well understood 22. The distinct Dvl populations within the oocyte display differing properties, which could reflect different binding partners or differing microenvironments. The movement and geometry of cytoplasmic Dvl assemblies suggest that they are influenced by interactions with cellular structures (Figure 2A, 3A, Video S2). To test this, we visualized endomembrane compartments, including putative lysosomes as labeled with Lamp1–3xGFP. We observed a striking association between Dvl-GFP granules and the surface of Lamp1-positive endosomes (Figure 3D,F, Video S4). Dvl-associated endosomes were also Rab7 positive, suggesting a subclass of late endosomes or lysosome-related organelles (Figure S3A,B). Immunofluorescence further revealed that endogenous Dvl protein assembled into granules that co-localized with Lamp1-positive endosomes (Figure S3C,D). This unexpected interaction between Dvl granules and Lamp1-positive endosomes could facilitate nucleation of Dvl assemblies, or serve as anchors to retain them at the oocyte’s vegetal pole. Based on image segmentation, the relative proportion of Dvl in granules associated with endosomes was low in Prophase I and peaked in meiosis I (Figure 3E), consistent with cell cycle regulation. Furthermore, the association with Dvl assemblies was maintained stably even as the endosomes moved (Figure 3G, Video S4). We hypothesize that Dvl assembly on the surface of these endosomes could provide a reservoir of Dvl protein that stably maintains the vegetal Dvl domain as the cortex is remodeled across the cell cycle. Endocytosis has been found to play an important role in Wnt signaling downstream of Frizzled receptors 23, and may also be necessary here for Dvl localization in oocytes.
We next evaluated which Dvl protein regions mediate interaction with endosomes. Prior work identified residues within the DIX domain that bind actin and lipids in vitro, and are necessary for Dvl self-association 4,24,25. These residues were later found to be important for Dvl localization and function in sea urchin embryos 12,17. We found that mutations in the putative actin and lipid binding domains in sea star Dvl (K47A, and K57A K58A, respectively) were deficient in their ability to localize to uniform puncta in Prophase I-arrested oocytes (Figure S3E). These substitutions may interfere with the ability of Dvl to oligomerize via its DIX domain, or to associate with a specific binding partner in Prophase I 25. However, in contrast with prior work in sea urchins, both mutants were capable of localizing to the vegetal pole following hormonal stimulation, as well as endosome association (Figure S3F)17. This distinction could represent a species difference, or differential localization requirements in meiosis versus embryogenesis.
Pre-localized cues are required for vegetal Dvl assembly
The polarization of Dvl localization occurs in single oocytes denuded of associated somatic cells in open seawater. Because this would preclude Wnt signaling from an adjacent cell, we reasoned that Dvl must be autonomously regulated within the oocyte. We first hypothesized that a gradient of diffusible factors influences Dvl behavior, analogous to MEX-5 regulation of P-granules in C. elegans, or the CyclinB-Cdk1 activity gradient in sea star oocytes 26,27. In this model, a repressive signal from the animal pole could prevent Dvl condensation except at the point farthest from its source. Importantly, such a gradient system would be sensitive to changes in oocyte geometry. Therefore, we used microfabricated oval-shaped PDMS chambers to alter oocyte shape by shortening the Animal-Vegetal axis (Figure S4A)28,29. If proximity to the animal pole represses Dvl granule formation, this shape change would cause Dvl to condense at the corners of the oval - the points farthest from the animal pole. Instead, we found that Dvl-GFP localized to the original vegetal pole within the microtubule ring (Figure S4B). This suggests that a cortical cue, rather than an activity gradient, regulates the reassembly of Dvl at the vegetal pole.
We next tested whether the vegetal cortex and adjacent cytoplasm contain pre-established signals that enable Dvl condensation. Using microdissection, we removed the vegetal portion of the oocyte during Prophase I arrest, prior to Dvl polarization (Figure 4A). As a control, we removed an equivalent amount of the lateral side of the oocyte cortex and cytoplasm. In vegetally-cut oocytes, Dvl-GFP assemblies failed to reform at the vegetal pole during meiosis (Figure 4A,B). Immunofluorescence of vegetally-cut oocytes for endogenous Dvl yielded similar results (Figure S4C). Reciprocally, isolated vegetal oocyte fragments displayed autonomous Dvl-GFP assembly in response to hormonal stimulation, suggesting that the vegetal region is capable of directing Dvl reassembly in the absence of other cellular structures or signals (Figure 4B). Vegetally ablated oocytes matured and fertilized normally, but produced embryos that failed to gastrulate, consistent with a primary axis defect (Figure S4D, see also 30). Thus, a cue established at the vegetal pole prior to meiotic resumption is essential for the subsequent local Dvl reassembly, which in turn is necessary for gastrulation during embryogenesis.
Figure 4. Localization mechanisms driving Dishevelled behavior.

(A) Live oocytes expressing Dvl-GFP and EMTB-mCherry from which the vegetal pole was removed by microdissection as described by schematic to the right. As a control, the side of the oocyte was removed. Upper images are Dvl-GFP alone in inverted grayscale. (B) Dvl-GFP Pixel intensity quantification of vegetal regions of control or vegetally-cut animal fragments, and isolated vegetal fragments (control n=11, animal fragment n=12, vegetal fragment n=9 oocytes, ****p<0.0001 Mann Whitney test). (C) Expression of GFP-tagged Frizzled 1 in Prophase I arrested and metaphase I oocytes, displayed as partial z-stacks. Frizzled 1 localization is enriched towards the vegetal cortex. Left panels are magnified views of insets. (D) Immunofluorescence images of endogenous Dvl in colocalization with Fzd1-GFP. Upper images are magnified views of inset indicated below. Scale bars = 50 μm. (E) Immunofluorescence images of endogenous Dvl in 2-cell embryos following control injection, Fzd1 morpholino, or morpholino combined with Fzd1-GFP overexpression. Scale bars = 50 μm. (F) Relative pixel intensity quantification of vegetal cortex Dvl signal (control n=24, Fzd1 knockdown n=25, Fzd1 overexpression n=26 oocytes, ****p < 0.0001Mann-Whitney Test).
See also Figure S4.
Maternally-expressed Frizzled 1 is required for Dvl localization
Based on our microdissection experiments, we reasoned that a pre-localized membrane-associated factor drives vegetal Dvl reassembly. Dvl can be recruited to the membrane through interactions with Frizzled transmembrane receptors, which detect Wnt signals. Analysis of a sea star ovary transcriptome revealed the expression of Frizzled 1 (Fzd1) 31. By fluorescent in situ analysis, we found that Fzd1 mRNA is present in small primordial oocytes and large, full-grown oocytes (Figure S4E). Strikingly, ectopically-expressed Fzd1-GFP localized to punctate structures that were enriched towards the vegetal pole in prophase I-arrested oocytes (Figure 4C, S4F). Fzd1 also localized to larger cytoplasmic compartments, possibly corresponding to the Golgi apparatus, in a gradient that was strongest at the vegetal pole, suggesting polarized trafficking of Fzd1 to the vegetal membrane. Consistent with a role in Dvl localization, we observed substantial colocalization between endogenous Dvl assemblies and Fzd1-GFP puncta at the vegetal pole (Figure 4D).
We next tested whether Fzd1 is required for Dvl localization by maternal knockdown. In contrast to controls, Fzd1-depleted embryos displayed significantly reduced punctate Dvl localization to the vegetal cortex (Figure 4D,E). Dvl localization was rescued and further increased by expressing knockdown-insensitive Fzd1-GFP (Figure 4D,E). Collectively, these results indicate that Fzd1 is localized maternally to the vegetal pole, and is essential for Dvl localization to the vegetal cortex. However, although Fzd1-GFP is vegetally enriched, it is present throughout the oocyte, suggesting other mechanisms are additionally required to restrict Dvl to the vegetal pole.
A conserved paradigm for primary body axis specification in animals
A fundamental problem in development is the generation of asymmetries along body axes to pattern a multi-functionalized body plan. Comparative studies point to asymmetric activation of the Wnt pathway as a deeply conserved determinant of the primary body axis in both cnidarians and bilaterian animals 8,10. Here, we identified a symmetry-determining event involving the Wnt pathway effector Dvl. In contrast to other polarization events influenced by sperm entry, such as in C. elegans, the sea star body axis is specified entirely by maternal control. We find that Dvl undergoes a cycle of polarized dissolution and reassembly to distribute asymmetrically to the vegetal pole. This redistribution is essential for primary axis specification and is regulated cell-autonomously during meiotic progression, and by cues including the receptor Fzd1, that are prelocalized to the vegetal pole.
Dvl has been noted to form large “puncta” or “granules” in several animal models, as well as by overexpression experiments in mammalian cell culture, but the physiological relevance and mechanisms by which these granules form has been debated 32,33. Here, we define a paradigm by which Dvl granules form at endogenous levels. An important future direction will be to determine the biophysical mechanisms underlying this dissolution-reassembly process. The ability of Dvl to self-associate through multivalent interactions to form dynamic assemblies could suggest a phase separation-based mechanism 22,34. The switch-like response of cortical Dvl dissolution dynamics in response to hormonal reception (<5 mins, Figure 1E, 2C) could result from a rapidly propagating signaling wave that lowers the condensation point of Dvl puncta across the cortex. In contrast, the gradual reassembly process (>30 mins, Figure 1E, 2C) suggests a nucleation-and-growth process by condensation that is seeded by endosomes and Fzd1 in a diffusion-limited manner. Alternatively, Dvl granules could instead form through canonical, structured interactions.
Prior work has identified potential associations between Dvl and endomembrane compartments, including putative vesicle-related organelles in Xenopus oocytes, although the physiological significance of these interactions remains unclear 2,4,13,24. Similarly, interactions with endoplasmic reticulum regulate the biogenesis and fission of membraneless organelles including p-bodies and stress granules 35, suggesting that endomembrane associations may broadly regulate protein assemblies such as Dvl granules. In sea star meiosis, we identified stable associations between Dvl and endosomes (Figure 3D–G), which could provide a local reservoir of Dvl protein to support a stable posterior-specifying signaling domain through the meiotic and embryonic mitotic divisions. Putative lipid-binding residues within the Dvl DIX domain are dispensable for this association, and instead suggest that it may occur through a specific binding partner, such as Frizzled receptors internalized by endocytosis 4,12,17,23. Given the broad diversity of organisms in which Dvl granule formation has been observed, Dvl and its capacity for spatially regulated self-assembly may occupy a critical node in the evolution of the Metazoan body plan.
STAR METHODS
Resource Availability
Lead contact
Requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, Iain Cheeseman (icheese@wi.mit.edu).
Materials availability
Plasmids generated in this study have been deposited with Addgene. All unique/stable reagents generated in this study are available from the Lead Contact with a completed Materials Transfer Agreement.
Data and code availability
Original Western blot images and image quantification values have been deposited with Mendeley Data and are publicly available as of the date of publication (DOI listed in the Key Resources Table). All other data, including microscopy data, are available from the Lead Contact on request.
This paper does not report original code.
Any additional information required to reanalyze the data reported in this paper is available from the Lead Contact upon request.
KEY RESOURCES TABLE.
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| Mouse anti-tubulin (DM1a) unconjugated | Sigma-Aldrich | Cat# T9026; RRID: AB_477593 |
| Rabbit anti-starfish Dvl (1:2000 IF, 1:5000 IB) | This study | N/A |
| Anti-digoxigenin-peroxidase antibody | Roche | Cat# #11207733 |
| GFP Booster ATTO488 antibody | ChromoTek | Cat# gba488–100; RRID: AB_2631386 |
| Chemicals, peptides, and recombinant proteins | ||
| Janelia Fluor 646 HaloTag Ligand | Promega | Cat# GA1120 |
| 1-Methyladenine | Acros Organics | Cat# AC201310250 |
| Calyculin A | Santa Cruz Biotechnology | Cat# SC-24000A |
| Emetine | Sigma Aldrich | Cat# E2375–500MG |
| Flavopiridol (L86–8275) | Selleckchem | Cat# S1230 |
| Trimethoprim | Santa Cruz Biotechnology | Cat# sc-203302 |
| Sulfamethoxazole | Santa Cruz Biotechnology | Cat# sc-208405 |
| digoxigenin-11-UTP | Sigma Aldrich | Cat# 11277073910 |
| Critical commercial assays | ||
| mMessage mMachine Sp6 Transcription Kit | Thermo Fisher | Cat# AM1340 |
| Poly(A) Tailing Kit | Thermo Fisher | Cat# AM1350 |
| TSA Plus Kit | Perkin-Elmer | Cat# NEL752001KT |
| Deposited Data | ||
| Raw Western blot images and microscopy quantification values | Mendeley Data | http://dx.doi.org/10.17632/z42m3kkvt4.1 |
| Experimental models: Organisms/strains | ||
| Patiria miniata | South-Coast Bio LLC | N/A |
| Oligonucleotides | ||
| Standard Control Morpholino 5′–CCTCTTACCTCAGTTACAATTTATA–3′ |
Gene-Tools | N/A |
| Dishevelled Morpholino 5′—ATGATTTTTGTCTCTTCCATGTTGG—3′ |
Gene-Tools | N/A |
| Fzd1 Morpholino 5′—GCTTGACTGAGGATGAAATTTAG—3′ |
Gene-Tools | N/A |
| Ubiquitin qPCR primers F: 5′—TTCGGTGAAAGCCAAGATTC—3′ R: 5′—CCCACCTCTCATGGCTAGAA—3′ |
16 | N/A |
| Wnt8 qPCR primers F: 5′—GGGCAGAAACCCAGAACGAC—3′ R: 5′—TACAGCTCCGTGCTTCCCAC—3′ |
16 | N/A |
| Bra qPCR primers F: 5′—CGAGATGATCGTCACCAAGA—3′ R: 5′—GGGGTGAAGTCCAGAAGGAT—3′ |
16 | N/A |
| Blimp1 qPCR primers F: 5′—CCAAACACGACGAAGCACTA—3′ R: 5′—GGCGTTGAGGACAGGTTTAG—3′ |
16 | N/A |
| Ets1/2 qPCR primers F: 5′—ATGCCTGTCTACGCGAGAGT—3′ R: 5′—TCCAGGCTGAACTCCTTGAT—3′ |
16 | N/A |
| FoxA qPCR primers F: 5′—TGGCGCATATAATCCTCACC—3′ R: 5′—GTCTCTCAGTGGCGCAAGAT—3′ |
16 | N/A |
| Six3 qPCR primers F: 5′—GTGAGACGTTGGAGGAGAGC—3′ R: 5′—TGTGGTTTTCCAGGATGTGA—3′ |
16 | N/A |
| Recombinant DNA | ||
| Dvl-GFP, pCS2+8 | This study | pZS217 |
| Dvl-HaloTag, pCS2+8 | This study | pZS181 |
| DvlK47A –GFP, pCS2+8 | This study | pZS220 |
| DvlK57A,K58A –GFP, pCS2+8 | This study | pZS221 |
| Lamp1–3xGFP, pCS2+ | This study | pZS222 |
| Lamp1-mCherry, pCS2+8 | This study | pSZ212 |
| mCherry-Rab7, pCS2+8 | This study | pZS218 |
| Frizzled 1 – GFP, pCS2+8 | This study | pZS202 |
| GST-Dvl, pGEX | This study | pZS230 |
| EMTB-mCherry | 39 | Addgene 26742 |
| Software and Algorithms | ||
| Fiji version 2.1.0/1.53c | 44 | RRID:SCR_002285 |
| Matlab version 2016b | MathWorks | RRID:SCR_001622 |
| Prism version 8.3.0 (328) | GraphPad | RRID:SCR_002798 |
Experimental Model and Subject Details
Sea stars (Patiria miniata) were wild-caught by South Coast Bio Marine (http://scbiomarine.com/), kept in artificial seawater aquariums at 15° C, and fed ad libitum with frozen shrimp. As they were wild collected animals, genetic backgrounds are mixed and ages are undefined. Individuals were randomly selected for isolation of oocytes or sperm. Males and females were housed in separate aquaria to avoid spawning. Intact ovary and testis fragments were surgically extracted from small incisions on the oral side of the animal using forceps and kept in filtered seawater containing 10 μg/ml trimethoprim and 50- μg/ml sulfamethoxazole at 15° C 36. The ovary fragments were transferred to fresh seawater and antibiotics every 2 days, and oocytes that were spontaneously released from the ovary were discarded. Intact ovary fragments were cultured this way for up to 1 week until oocytes were to be microinjected for experimentation.
Method Details
Oocyte Culture
Isolated oocytes were cultured for a maximum of 24 hours in artificial seawater with 10 μg/ml trimethoprim and 50 μg/ml sulfamethoxazole. To induce meiotic resumption, 1-methyladenine (Acros Organics) was added to the culture at a final concentration of 10 μM. For fertilization, extracted sperm was added to the culture at a 1:1,000,000 dilution. Latrunculin b was used at a final concentration of 10 μM, and nocodazole was used at 3.3 μM. For knockdown of maternal Fzd1 (e.g. Figure 4D,E), oocytes were injected with 500 μM morpholino and cultured for 2 days in sulfamethoxazole/trimethoprim-containing seawater (50 μg/ml sulfamethoxazole, 10 μg/ml trimethoprim).
Construct and antibody generation
Patiria miniata gene homologs were identified using genomics tools at echinobase.org 37. Fluorescent protein fusion constructs for mRNA in vitro transcription were cloned into vectors derived from pCS2+8 using standard restriction enzyme-based methods 38. Dvl protein corresponding to the DIX domain (residues 1–90) was expressed as an n-terminal GST fusion in the pGEX-6P vector in BL21 bacteria. Expression was induced with 0.1 mM IPTG and the cells were grown overnight at 18° C. GST-DIX was then purified from bacterial lysate using Glutathione-agarose (Sigma), eluted with reduced L-glutathione (Sigma). Using this immunogen, antibodies were then generated in rabbits (Covance). For affinity purification, serum was pre-depleted against GST, and then purified with GST-DIX bound to HiTrap NHS-activated columns (GE Healthcare). EMTB-mCherry was a gift from William Bement (Addgene plasmid # 26742; http://n2t.net/addgene:26742; RRID:Addgene_26742) 39.
Oocyte Microinjection and Manipulation
For expression of constructs in oocytes, plasmids were linearized with NotI to yield linear template DNA. mRNA was transcribed in vitro using mMessage mMachine SP6 and the polyadenylation kit (Life Technologies), then precipitated using lithium chloride solution. Prophase I arrested oocytes were injected horizontally in Kiehart chambers with 10–20 picoliters of mRNA solution in nuclease free water as described 40. Dvl-GFP wildtype and mutant constructs were injected at 800 ng/μl. After microinjection, oocytes were cultured 18–24 hours to allow time for the constructs to translate before 1-methyladenine stimulation. Custom synthesized morpholinos, or the Gene Tools standard control, were injected at 1000 μM for Dvl and 500 μM for Fzd1 (Gene Tools). For microsurgery experiments (Figure 4A, S4C,D), oocytes were cut by hand using a pulled capillary tube as a knife and a mouth pipette to hold in place. For shape mold experiments (Figure S4A,B), chambers were fabricated by casting PDMS onto patterned silicon wafers as described previously 41. The chamber shapes were designed with a height of 80 μm and surface area of around 27000 μm2, to match typical volumes of the oocytes. The patterned silicon wafer was manufactured using photolithography (Microfactory SAS, France). The silicon wafer was silanized with Trichlorosilane (Sigma 448931). PDMS was made by mixing Dow SYLGARD™ 184 Silicone Elastomer Clear solution at a 10:1 base-to-curing agent ratio. After mixing thoroughly, the elastomer was poured over the silicon master mold, degassed in a vacuum chamber and cured at 60 °C in an oven for one hour.
Immunofluorescence, In Situ Hybridization, and Imaging
For endogenous Dvl immunofluorescence (Figure 4D, S1C,D), oocytes were fixed at various stages in a microtubule stabilization buffer (1% paraformaldehyde, 0.1% Triton X-100, 100 mM HEPES, pH 7.0, 50 mM EGTA, 10 mM MgSO4, 400 mM dextrose) for 15 minutes at room temperature 42. The oocytes were then transferred to a solution of 80% methanol, 20% DMSO, and incubated on ice for at least 1 hour and up to overnight. Oocytes were then washed 3 times in PBS with 0.1% Triton X-100 (PBSTx), transferred to sodium citrate buffer (10 mM tri-sodium citrate dihydrate, 0.05% Tween 20, pH 6.0) and heated at 55° C for 1 hour for antigen retrieval. Oocytes were then washed 3 times with PBSTx and blocked for 15 minutes in AbDil (3% BSA, 1 X TBS, 0.1% triton X-100, 0.1% Na Azide). Primary antibodies diluted in AbDil were then applied and the oocytes were incubated overnight at room temperature. Anti-alpha tubulin (DM1α, Sigma) and Dishevelled (this study) antibodies were used at 1:5,000. DNA was stained with Hoechst. GFP fusion construct expressing oocytes were instead fixed with 2% PFA in Millonig’s Buffered Fixative (0.2 M NaH2PO4*H2O, 0.136 M NaCl, pH 7.0) overnight at room temperature, permeabilized by three washes with PBSTx, and blocked with AbDil as described above. GFP booster (Chromotek) was used at 1:1000 to improve the signal from GFP fusion constructs (e.g. Figure S2A–D, 4C,D). The oocytes were compressed under coverslips in ProLong Gold Antifade Mountant (ThermoFisher).
Fluorescent whole mount in situ hybridization (FISH) was performed as previously described 43. Embryos were collected and left in fixation solution (4% paraformaldehyde, 32.5% sea water, 32.5 mM MOPS pH 7, 162.5 mM NaCl) overnight at 4°C, then washed in MOPS and stored in 70% ethanol. After rehydration, embryos were hybridized with the Frzd1 probe (0.1 ng/μl) in hybridization buffer (70% formamide, 100 mM MOPS pH 7, 500 mM NaCl, 0.1%Tween 20, 1 mg/ml BSA.) at 60°C for a week. Signal was developed with the fluorophore-conjugated tyramide kit (Perkin Elmer, Cat. #:NEL752001KT; RRID:AB_2572409). After post hybridization washes in maleic acid buffer, embryos were blocked in the blocking buffer provided by the kit and left overnight with anti-digoxigenin-peroxidase antibody ( Roche cat #11207733 ) diluted 1:2000. Signal was detected by staining for 30 min in 1:400 cy3 in amplification diluent (provided in the kit).
Fixed oocytes were imaged with a Nikon Ti2 microscope with a CoolSnap HQ2 CCD camera and a 100× 1.40 NA Olympus U-PlanApo objective (Figure S2D), or a Zeiss 710 confocal (Figure S1–4). Live confocal imaging was performed with a Yokogawa W1 spinning disk microscope (Figure 2–4, S3A,E,F) or a Zeiss 710 (Figure S4B). Images were processed with Fiji, and scaled equivalently across conditions unless otherwise specified 44. For images collected with the Zeiss 710 in Figure 4C,D, S1D, S3C,D, S4F, the ROF Denoise plugin in Fiji was used with theta value 5.
RNA extraction and quantitative real-time PCR (qPCR)
RNA from 100 embryos was isolated with the RNeasy Micro kit (Qiagen, Cat#:74004). RNA was quantified with a nanodrop and the whole pool of embryos was used to make cDNA. cDNA synthesis was performed using Maxima kit (Life Technologies, Cat#:K1641) and random hexamer primers were used to prime synthesis of first strand cDNA. qPCR was performed using ABI900 real time instrument with Maxima SYBR master mix (Life Technologies, Cat#:FERK0222). qPCR volume was 25 μl per well containing 1 μl of cDNA diluted 1:10 (1 embryo equivalent), 2.5 pMoles of each primer (forward and reverse), 12.5 μl of SybrGreen Mix and water. The integrity of the amplicons was determine by melt curves. ddCt values were calculated between experiment and control embryos and converted to fold differences using ubiquitin as a reference (FD = 2 ΔΔCt).Experiments were run in two biological replicates, and each biological replicate was run on the qPCR machine with three technical replicates.
Quantification and Statistical Analysis
Statistical analyses were performed using Prism (8.3.0 GraphPad Software). Details of statistical tests and sample sizes are provided in the figure legends. Unless stated otherwise in the figure legends, images comparing the same signal across conditions are scaled equivalently. In all cases in which pixel intensity is quantified, unscaled images with appropriate background subtraction were used, as outlined below.
Kymograph depictions of Dvl localization dynamics (Figure 1E) were generated in MATLAB. The space time kymograph of cytoplasmic Dvl IC(y, t) was obtained by extracting the fluorescence intensity IC(y) along the animal-vegetal (AV) axis for all time frame t. Here, we used y to denote the distance along AV axis. We first performed a Gaussian filtering step (with standard deviation of 2 pixels) to smooth the fluorescence image. We then computed the average of the intensity values 10 pixels to the left and to the right of the AV axis to obtain IC(y). To construct the full kymograph IC(y, t), the intensity IC(y) at each time frame t is aligned such that the animal and vegetal poles correspond to the same position and resampled at the appropriate distance y. The space time kymograph of cortical fluorescence IR(s, t) is computed by first extracting the boundary of the oocyte and then extracting the fluorescence intensity IR(s) along the boundary for all time frames t. Here, we used s to parameterize the arclength of the oocyte boundary. For each time frame t, we performed a Gaussian filtering step (with a standard deviation of 1.2 pixels) before applying a thresholding step to make a binary image. The oocyte boundary is obtained by using the bwboundaries function in MATLAB on the binary image. The intensity IR(s) is obtained by first identifying a local window of size 8-by-8 pixels centered at and taking the mean intensity of the pixels in the top 50 percentile intensity within the local window. To construct the full kymograph IR(s, t), the intensity IR(s) at each time frame t is aligned such that the AP corresponds to the same arclength position and resampled at the appropriate arclength s.
To segment the cortical and cytoplasmic Dvl puncta at a single time point (Figure 2B–D), each z slice of the confocal z-stack was first demarcated into a cytoplasmic and a cortical region. By applying a thresholding to identify the boundary of the oocyte, the region around 30 pixels (3.25 μm) extending from the cell boundary into the cytoplasm is defined as the cortical region, while the rest of the oocyte is considered the cytoplasmic region. After performing this step on all z-slices, a maximum intensity projection is done separately on the cortical and cytoplasmic region. To segment the Dvl puncta in the respective region, a 2D Gaussian blur is first performed before a thresholding step. Once the binary image is obtained, the MATLAB function regionprops is used to calculate the area and centroid of each puncta. The mean squared displacement with respect to initial position is calculated as , where the average is done over all available puncta. To generate the plot of mean velocity as a function of angle, the instantaneous velocity along the puncta trajectory is first calculated as , where dt is the frame duration. All recorded instantaneous velocities were then binned according to the angle they make with the x-axis.
To quantify the fraction of Dvl co-localizing with Lamp1+ endosomes (Figure 3E, S3F), the segmentation of Dvl puncta and Lamp puncta were first performed separately. Briefly, a 2D Gaussian blur was performed before the appropriate thresholding step was applied to obtain a binary image for each channel. The fraction of Dvl on endosome fDvl is calculated as , where is the total area of all Dvl puncta that overlap with Lamp puncta, and 𝐴𝐷𝑣𝑙 is the total area of all Dvl puncta. The final reported fDvl is the temporal mean averaged over the appropriate cell cycle duration.
Quantification of cortex-associated Dvl signal was performed using Fiji 44. To measure the pixel intensity of Dvl localized to the membrane in Figure 4F, a single z-lice encompassing the membrane was selected. A 50-pixel circular region of interest was defined within the vegetal Dvl domain, and a background area was defined in an in-focus region of the oocyte membrane but outside of the Dvl domain. These regions were measured using RawIntDen parameter, and the background was subtracted from the Dvl measurement. These background subtracted values were then normalized to the mean of the control oocytes. For quantification of Dvl mutants (Figure S3E,F) background subtraction was performed using a rolling ball radius of 1 pixel and the sliding paraboloid function. Gaussian Blur was then applied with radius of 1 pixel. Next, Auto Threshold was performed using the Triangle method. To identify puncta, the Watershed function followed by Analyze Particles was performed to create a selection of ROIs that were then applied to the original image, and then the intensities were determined with Measure and the RawIntDen parameter. The total number of puncta are reported relative to image area in square microns, along with the mean of RawIntDen across puncta per individual oocyte.
Supplementary Material
Video S1. Dvl localization during meiotic resumption, related to Figure 1. Timelapse video of the oocyte depicted in Figure 1D,E, with Dvl-GFP (magenta) and the microtubule probe EMTB-mCherry (green).
Video S2. Dvl dissolution and condensation, related to Figure 2. (Top panel) Timelapse video of the oocyte vegetal pole at higher magnification, corresponding to the oocyte depicted in Figure 2A, with Dvl-GFP (magenta) and the microtubule probe EMTB-mCherry (green). (Middle and bottom panels) Image segmentation analysis to track particles within the cytoplasm (middle) or within 3.25 μm of the cortex (bottom).
Video S3. Fusion of cytoplasmic Dvl assemblies, related to Figure 3. Example fusion event between two Dvl assemblies in the vegetal pole cytoplasm.
Video S4. Association of Dvl assemblies and Lamp1+ endosomes, related to Figure 3. Association of Dvl assemblies (magenta) associated with moving Lamp1+ endosomes (green) over time.
Highlights.
The primary embryonic body axis is maternally specified in sea star oocytes.
Posterior fate specification requires the Wnt pathway effector Dishevelled.
Dishevelled localization polarizes by cell cycle-linked dissolution and reassembly.
Maternal cues and endosome association drive Dishevelled localization.
Acknowledgements
We thank the Marine Biological Laboratory and the students, faculty, and staff of the Embryology Course, where the pilot experiments for this work began. We thank Gene-Tools for donating the PmDvl morpholino to the course. We thank Hannah Rosenblatt for suggesting an examination of lysosomes. We thank Peter Lenart, Brad Shuster, Peter Reddien, and members of the Cheeseman laboratory for helpful discussions. We thank Veronica Hinman and her laboratory for sharing in situ probes. This research was supported by 1K99HD099315 to S.Z.S., R35GM126930 to I.M.C., and a grant to I.M.C. from the Gordon and Betty Moore Foundation. The Embryology course is supported by R25HD094666.
Footnotes
Declaration of Interests
I.M.C. is a member of the Editorial Advisory Board for Current Biology.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.St Johnston D, and Ahringer J (2010). Cell polarity in eggs and epithelia: parallels and diversity. Cell 141, 757–774. 10.1016/j.cell.2010.05.011. [DOI] [PubMed] [Google Scholar]
- 2.Schwarz-Romond T, Merrifield C, Nichols BJ, and Bienz M (2005). The Wnt signalling effector Dishevelled forms dynamic protein assemblies rather than stable associations with cytoplasmic vesicles. J. Cell Sci. 118, 5269–5277. 10.1242/jcs.02646. [DOI] [PubMed] [Google Scholar]
- 3.Schaefer KN, and Peifer M (2019). Wnt/Beta-Catenin Signaling Regulation and a Role for Biomolecular Condensates. Dev. Cell 48, 429–444. 10.1016/j.devcel.2019.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Capelluto DG, Kutateladze TG, Habas R, Finkielstein CV, He X, and Overduin M (2002). The DIX domain targets dishevelled to actin stress fibres and vesicular membranes. Nature 419, 726–729. 10.1038/nature01056. [DOI] [PubMed] [Google Scholar]
- 5.Wikramanayake AH (2013). Heads or Tails? Karl Ernst von Baer, Robert Remak, and characterization of the primary axis in animal eggs. Mol. Reprod. Dev. 80, i. 10.1002/mrd.22156. [DOI] [PubMed] [Google Scholar]
- 6.Martindale MQ (2005). The evolution of metazoan axial properties. Nat. Rev. Genet. 6, 917–927. 10.1038/nrg1725. [DOI] [PubMed] [Google Scholar]
- 7.Kominami T (1983). Establishment of embryonic axes in larvae of the starfish, Asterina pectinifera. J. Embryol. Exp. Morphol. 75, 87–100. [PubMed] [Google Scholar]
- 8.Martindale MQ, and Hejnol A (2009). A developmental perspective: changes in the position of the blastopore during bilaterian evolution. Dev. Cell 17, 162–174. 10.1016/j.devcel.2009.07.024. [DOI] [PubMed] [Google Scholar]
- 9.Wikramanayake AH, Hong M, Lee PN, Pang K, Byrum CA, Bince JM, Xu R, and Martindale MQ (2003). An ancient role for nuclear beta-catenin in the evolution of axial polarity and germ layer segregation. Nature 426, 446–450. 10.1038/nature02113. [DOI] [PubMed] [Google Scholar]
- 10.Petersen CP, and Reddien PW (2009). Wnt signaling and the polarity of the primary body axis. Cell 139, 1056–1068. 10.1016/j.cell.2009.11.035. [DOI] [PubMed] [Google Scholar]
- 11.Peng CJ, and Wikramanayake AH (2013). Differential regulation of disheveled in a novel vegetal cortical domain in sea urchin eggs and embryos: implications for the localized activation of canonical Wnt signaling. PLoS One 8, e80693. 10.1371/journal.pone.0080693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Leonard JD, and Ettensohn CA (2007). Analysis of dishevelled localization and function in the early sea urchin embryo. Dev. Biol. 306, 50–65. 10.1016/j.ydbio.2007.02.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Miller JR, Rowning BA, Larabell CA, Yang-Snyder JA, Bates RL, and Moon RT (1999). Establishment of the dorsal-ventral axis in Xenopus embryos coincides with the dorsal enrichment of dishevelled that is dependent on cortical rotation. J. Cell Biol. 146, 427–437. 10.1083/jcb.146.2.427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lee PN, Kumburegama S, Marlow HQ, Martindale MQ, and Wikramanayake AH (2007). Asymmetric developmental potential along the animal-vegetal axis in the anthozoan cnidarian, Nematostella vectensis, is mediated by Dishevelled. Dev. Biol. 310, 169–186. 10.1016/j.ydbio.2007.05.040. [DOI] [PubMed] [Google Scholar]
- 15.Kishimoto T (2018). MPF-based meiotic cell cycle control: Half a century of lessons from starfish oocytes. Proc. Jpn. Acad. Ser. B. Phys. Biol. Sci. 94, 180–203. 10.2183/pjab.94.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McCauley BS, Akyar E, Saad HR, and Hinman VF (2015). Dose-dependent nuclear beta-catenin response segregates endomesoderm along the sea star primary axis. Development 142, 207–217. 10.1242/dev.113043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Weitzel HE, Illies MR, Byrum CA, Xu R, Wikramanayake AH, and Ettensohn CA (2004). Differential stability of beta-catenin along the animal-vegetal axis of the sea urchin embryo mediated by dishevelled. Development 131, 2947–2956. 10.1242/dev.01152. [DOI] [PubMed] [Google Scholar]
- 18.Wikramanayake AH, Huang L, and Klein WH (1998). beta-Catenin is essential for patterning the maternally specified animal-vegetal axis in the sea urchin embryo. Proc. Natl. Acad. Sci. USA. 95, 9343–9348. 10.1073/pnas.95.16.9343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Range RC, Angerer RC, and Angerer LM (2013). Integration of canonical and noncanonical Wnt signaling pathways patterns the neuroectoderm along the anterior-posterior axis of sea urchin embryos. PLoS Biol. 11, e1001467. 10.1371/journal.pbio.1001467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kitajima A, and Hamaguchi Y (2005). Determination of first cleavage plane: the relationships between the orientation of the mitotic apparatus for first cleavage and the position of meiotic division-related structures in starfish eggs. Dev. Biol. 280, 48–58. 10.1016/j.ydbio.2004.12.033. [DOI] [PubMed] [Google Scholar]
- 21.Swartz SZ, Nguyen HT, McEwan BC, Adamo ME, Cheeseman IM, and Kettenbach AN (2021). Selective dephosphorylation by PP2A-B55 directs the meiosis I - meiosis II transition in oocytes. Elife 10. 10.7554/eLife.70588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bienz M (2020). Head-to-Tail Polymerization in the Assembly of Biomolecular Condensates. Cell 182, 799–811. 10.1016/j.cell.2020.07.037. [DOI] [PubMed] [Google Scholar]
- 23.Brunt L, and Scholpp S (2018). The function of endocytosis in Wnt signaling. Cell. Mol. Life Sci. 75, 785–795. 10.1007/s00018-017-2654-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Capelluto DG, and Overduin M (2005). Secondary structure, 1H, 13C and 15N resonance assignments and molecular interactions of the dishevelled DIX domain. J. Biochem. Mol. Biol. 38, 243–247. 10.5483/bmbrep.2005.38.2.243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Schwarz-Romond T, Fiedler M, Shibata N, Butler PJ, Kikuchi A, Higuchi Y, and Bienz M (2007). The DIX domain of Dishevelled confers Wnt signaling by dynamic polymerization. Nat. Struct. Mol. Biol. 14, 484–492. 10.1038/nsmb1247. [DOI] [PubMed] [Google Scholar]
- 26.Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, Julicher F, and Hyman AA (2009). Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 324, 1729–1732. 10.1126/science.1172046. [DOI] [PubMed] [Google Scholar]
- 27.Bischof J, Brand CA, Somogyi K, Majer I, Thome S, Mori M, Schwarz US, and Lenart P (2017). A cdk1 gradient guides surface contraction waves in oocytes. Nat. Commun. 8, 849. 10.1038/s41467-017-00979-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Minc N, Burgess D, and Chang F (2011). Influence of cell geometry on division-plane positioning. Cell 144, 414–426. 10.1016/j.cell.2011.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wigbers MC, Tan TH, Brauns F, Liu J, Swartz SZ, Frey E, and Fakhri N (2021). A hierarchy of protein patterns robustly decodes cell shape information. Nat. Phys, 1–7. [Google Scholar]
- 30.Kuraishi R, and Osanai K (1994). Contribution of Maternal Factors and Cellular Interaction to Determination of Archenteron in the Starfish Embryo. Development 120, 2619–2628. [Google Scholar]
- 31.Reich A, Dunn C, Akasaka K, and Wessel G (2015). Phylogenomic analyses of Echinodermata support the sister groups of Asterozoa and Echinozoa. PLoS One 10, e0119627. 10.1371/journal.pone.0119627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ma W, Chen M, Kang H, Steinhart Z, Angers S, He X, and Kirschner MW (2020). Single-molecule dynamics of Dishevelled at the plasma membrane and Wnt pathway activation. Proc. Natl. Acad. Sci. USA. 10.1073/pnas.1910547117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kan W, Enos MD, Korkmazhan E, Muennich S, Chen DH, Gammons MV, Vasishtha M, Bienz M, Dunn AR, Skiniotis G, and Weis WI (2020). Limited Dishevelled/Axin oligomerization determines efficiency of Wnt/beta-catenin signal transduction. Elife 9. 10.7554/eLife.55015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gammons MV, Renko M, Johnson CM, Rutherford TJ, and Bienz M (2016). Wnt Signalosome Assembly by DEP Domain Swapping of Dishevelled. Mol. Cell 64, 92–104. 10.1016/j.molcel.2016.08.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lee JE, Cathey PI, Wu H, Parker R, and Voeltz GK (2020). Endoplasmic reticulum contact sites regulate the dynamics of membraneless organelles. Science 367. 10.1126/science.aay7108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Swartz SZ, McKay LS, Su K-C, Bury L, Padeganeh A, Maddox PS, Knouse KA, and Cheeseman IM (2019). Quiescent Cells Actively Replenish CENP-A Nucleosomes to Maintain Centromere Identity and Proliferative Potential. Dev. Cell 51, 35–48.e37. 10.1016/j.devcel.2019.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cary GA, Cameron RA, and Hinman VF (2018). EchinoBase: Tools for Echinoderm Genome Analyses. Methods Mol. Biol. 1757, 349–369. 10.1007/978-1-4939-7737-6_12. [DOI] [PubMed] [Google Scholar]
- 38.Gokirmak T, Campanale JP, Shipp LE, Moy GW, Tao H, and Hamdoun A (2012). Localization and substrate selectivity of sea urchin multidrug (MDR) efflux transporters. J. Biol. Chem. 287, 43876–43883. 10.1074/jbc.M112.424879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Miller AL, and Bement WM (2009). Regulation of cytokinesis by Rho GTPase flux. Nat. Cell Biol. 11, 71–77. 10.1038/ncb1814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Swartz SZ, McKay LS, Su K-C, Bury L, Padeganeh A, Maddox PS, Knouse KA, and Cheeseman IM (2019). Quiescent Cells Actively Replenish CENP-A Nucleosomes to Maintain Centromere Identity and Proliferative Potential. Developmental cell 51, 35–48.e37. 10.1016/j.devcel.2019.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wigbers MC, Tan TH, Brauns F, Liu J, Swartz SZ, Frey E, and Fakhri N (2021). A hierarchy of protein patterns robustly decodes cell shape information. Nature Physics, 1–7. [Google Scholar]
- 42.von Dassow G, Verbrugghe KJC, Miller AL, Sider JR, and Bement WM (2009). Action at a distance during cytokinesis. J. Cell Biol. 187, 831–845. 10.1083/jcb.200907090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Perillo M, Paganos P, Spurrell M, Arnone MI, and Wessel GM (2021). Methodology for Whole Mount and Fluorescent RNA In Situ Hybridization in Echinoderms: Single, Double, and Beyond. Methods. Mol. Biol. 2219, 195–216. 10.1007/978-1-0716-0974-3_12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, et al. (2012). Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682. 10.1038/Nmeth.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Video S1. Dvl localization during meiotic resumption, related to Figure 1. Timelapse video of the oocyte depicted in Figure 1D,E, with Dvl-GFP (magenta) and the microtubule probe EMTB-mCherry (green).
Video S2. Dvl dissolution and condensation, related to Figure 2. (Top panel) Timelapse video of the oocyte vegetal pole at higher magnification, corresponding to the oocyte depicted in Figure 2A, with Dvl-GFP (magenta) and the microtubule probe EMTB-mCherry (green). (Middle and bottom panels) Image segmentation analysis to track particles within the cytoplasm (middle) or within 3.25 μm of the cortex (bottom).
Video S3. Fusion of cytoplasmic Dvl assemblies, related to Figure 3. Example fusion event between two Dvl assemblies in the vegetal pole cytoplasm.
Video S4. Association of Dvl assemblies and Lamp1+ endosomes, related to Figure 3. Association of Dvl assemblies (magenta) associated with moving Lamp1+ endosomes (green) over time.
Data Availability Statement
Original Western blot images and image quantification values have been deposited with Mendeley Data and are publicly available as of the date of publication (DOI listed in the Key Resources Table). All other data, including microscopy data, are available from the Lead Contact on request.
This paper does not report original code.
Any additional information required to reanalyze the data reported in this paper is available from the Lead Contact upon request.
KEY RESOURCES TABLE.
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| Mouse anti-tubulin (DM1a) unconjugated | Sigma-Aldrich | Cat# T9026; RRID: AB_477593 |
| Rabbit anti-starfish Dvl (1:2000 IF, 1:5000 IB) | This study | N/A |
| Anti-digoxigenin-peroxidase antibody | Roche | Cat# #11207733 |
| GFP Booster ATTO488 antibody | ChromoTek | Cat# gba488–100; RRID: AB_2631386 |
| Chemicals, peptides, and recombinant proteins | ||
| Janelia Fluor 646 HaloTag Ligand | Promega | Cat# GA1120 |
| 1-Methyladenine | Acros Organics | Cat# AC201310250 |
| Calyculin A | Santa Cruz Biotechnology | Cat# SC-24000A |
| Emetine | Sigma Aldrich | Cat# E2375–500MG |
| Flavopiridol (L86–8275) | Selleckchem | Cat# S1230 |
| Trimethoprim | Santa Cruz Biotechnology | Cat# sc-203302 |
| Sulfamethoxazole | Santa Cruz Biotechnology | Cat# sc-208405 |
| digoxigenin-11-UTP | Sigma Aldrich | Cat# 11277073910 |
| Critical commercial assays | ||
| mMessage mMachine Sp6 Transcription Kit | Thermo Fisher | Cat# AM1340 |
| Poly(A) Tailing Kit | Thermo Fisher | Cat# AM1350 |
| TSA Plus Kit | Perkin-Elmer | Cat# NEL752001KT |
| Deposited Data | ||
| Raw Western blot images and microscopy quantification values | Mendeley Data | http://dx.doi.org/10.17632/z42m3kkvt4.1 |
| Experimental models: Organisms/strains | ||
| Patiria miniata | South-Coast Bio LLC | N/A |
| Oligonucleotides | ||
| Standard Control Morpholino 5′–CCTCTTACCTCAGTTACAATTTATA–3′ |
Gene-Tools | N/A |
| Dishevelled Morpholino 5′—ATGATTTTTGTCTCTTCCATGTTGG—3′ |
Gene-Tools | N/A |
| Fzd1 Morpholino 5′—GCTTGACTGAGGATGAAATTTAG—3′ |
Gene-Tools | N/A |
| Ubiquitin qPCR primers F: 5′—TTCGGTGAAAGCCAAGATTC—3′ R: 5′—CCCACCTCTCATGGCTAGAA—3′ |
16 | N/A |
| Wnt8 qPCR primers F: 5′—GGGCAGAAACCCAGAACGAC—3′ R: 5′—TACAGCTCCGTGCTTCCCAC—3′ |
16 | N/A |
| Bra qPCR primers F: 5′—CGAGATGATCGTCACCAAGA—3′ R: 5′—GGGGTGAAGTCCAGAAGGAT—3′ |
16 | N/A |
| Blimp1 qPCR primers F: 5′—CCAAACACGACGAAGCACTA—3′ R: 5′—GGCGTTGAGGACAGGTTTAG—3′ |
16 | N/A |
| Ets1/2 qPCR primers F: 5′—ATGCCTGTCTACGCGAGAGT—3′ R: 5′—TCCAGGCTGAACTCCTTGAT—3′ |
16 | N/A |
| FoxA qPCR primers F: 5′—TGGCGCATATAATCCTCACC—3′ R: 5′—GTCTCTCAGTGGCGCAAGAT—3′ |
16 | N/A |
| Six3 qPCR primers F: 5′—GTGAGACGTTGGAGGAGAGC—3′ R: 5′—TGTGGTTTTCCAGGATGTGA—3′ |
16 | N/A |
| Recombinant DNA | ||
| Dvl-GFP, pCS2+8 | This study | pZS217 |
| Dvl-HaloTag, pCS2+8 | This study | pZS181 |
| DvlK47A –GFP, pCS2+8 | This study | pZS220 |
| DvlK57A,K58A –GFP, pCS2+8 | This study | pZS221 |
| Lamp1–3xGFP, pCS2+ | This study | pZS222 |
| Lamp1-mCherry, pCS2+8 | This study | pSZ212 |
| mCherry-Rab7, pCS2+8 | This study | pZS218 |
| Frizzled 1 – GFP, pCS2+8 | This study | pZS202 |
| GST-Dvl, pGEX | This study | pZS230 |
| EMTB-mCherry | 39 | Addgene 26742 |
| Software and Algorithms | ||
| Fiji version 2.1.0/1.53c | 44 | RRID:SCR_002285 |
| Matlab version 2016b | MathWorks | RRID:SCR_001622 |
| Prism version 8.3.0 (328) | GraphPad | RRID:SCR_002798 |