

Abstract
Evolution of dihydrofolate reductase (DHFR) has been studied using the enzyme from Escherichia coli DHFR (ecDHFR) as a model, but less studies have used the enzyme from Homo sapiens DHFR (hsDHFR). Each enzyme maintains a short and narrow distribution of hydride donor-acceptor distances (DAD) at the tunneling ready state (TRS). Evolution of the enzyme was previously studied in ecDHFR where three key sites were identified as important to the catalyzed reaction. The corresponding sites in hsDHFR are F28, 62-PEKN, and 26-PPLR. Each of these sites was studied here through the creation of mutant variants of the enzyme and measurements of the temperature dependence of the intrinsic kinetic isotope effects (KIEs) on the reaction. F28 is mutated first to M (F28M) and then to the L of the bacterial enzyme (F28L). The KIEs of the F28M variant are larger and more temperature-dependent than wild-type (WT), suggesting a broader and longer average DAD at the TRS. To more fully mimic ecDHFR, we also study a triple mutant of the human enzyme (F32L-PP26N-PEKN62G). Remarkably, the intrinsic KIEs, while larger in magnitude, are temperature-independent like the WT enzymes. We also construct deletion mutations of hsDHFR removing both the 62-PEKN and 26-PPLR sequences. The results mirror those described previously for insertion mutants of ecDHFR. Taken together, these results suggest a balancing act during DHFR evolution between achieving an optimal TRS for hydride transfer and preventing product inhibition arising from the different intercellular pools of NADPH and NADP+ in prokaryotic and eukaryotic cells.
Introduction
The extraordinary rate accelerations achieved by enzymes have been studied extensively, but a comprehensive understanding of the physical principles governing enzyme catalysis has yet to be achieved. In particular, considerable debate exists over the possible roles of fast (fs to ps) protein motions in enzymatic reactions.3−16 Several computational, bioinformatic, and experimental studies have suggested a role for such motions in the hydride transfer reaction catalyzed by dihydrofolate reductase (DHFR; for reviews, see refs (3, 17)). The enzyme catalyzes the stereospecific transfer of the pro-R hydride from reduced nicotinamide adenine diphosphate (NADPH) to the C6 position of the dihydropterin ring of 7,8-dihydrofolate (DHF) to generate 5,6,7,8-tetrahydrofolate (THF). The reaction catalyzed by DHFR is essential in maintaining the intercellular pool of THF, which is required for the anabolism of purine nucleotides and some amino acids, thus making the enzyme an important drug target.
Experimental evidence regarding the role of active-site conformational sampling in the DHFR reaction has largely come from measurements of the temperature dependence of the intrinsic kinetic isotope effects (KIEs) on hydride transfer for the wild-type (WT)18 and mutant19−24 forms of the enzyme from Escherichia coli DHFR (ecDHFR). Theoretical studies have demonstrated that the magnitude and distribution of the hydride donor–acceptor distance (DAD) are the dominant factors in determining the temperature dependence of the KIEs.5,7−9 These studies support an interpretation in which the protein scaffold reorganizes the active site to achieve a DAD conducive for hydride tunneling.7−9 In ecDHFR, as in most WT enzymes with their physiological substrate, this reorganization results in a short and narrow distribution of DADs so that the effects of thermal fluctuations in the optimal tunneling ready state (TRS) are the same for all hydrogen isotopes. As a result, the intrinsic KIEs are temperature-independent.18 Disruption of the optimized TRS through site-directed mutagenesis, for example, results in DADs that are longer and more broadly distributed. Thus, temperature-dependent KIEs are observed for many mutants of the enzyme because at lower temperatures there is insufficient wavefunction overlap for effective tunneling of the heavier isotopes at the average DAD.
Previous studies have linked the functional motions of DHFR to the evolution of the enzyme.1,19,25,26 Phylogenetic analysis of DHFR sequences in organisms ranging from bacteria to Homo sapiens DHFR (hsDHFR) identified three key sites that were predicted to influence the motions of the enzyme (Figure 1) and were denoted as phylogenetically coherent events (PCEs).1 The first PCE was a four-amino-acid insert (62-PEKN in hsDHFR) that appeared ∼797 million years ago and is conserved in higher organisms. A second PCE occurred ∼325 million years ago in the M20 loop of the enzyme and consists of a polyproline sequence (26-PPLR in hsDHFR).1 A third PCE is at the end of the M20 loop and corresponds to F32 in the human enzyme. The corresponding residue in the extensively studied E. coli enzyme is a leucine that first evolved to a methionine before becoming a phenylalanine ∼499 million years ago.
Figure 1.
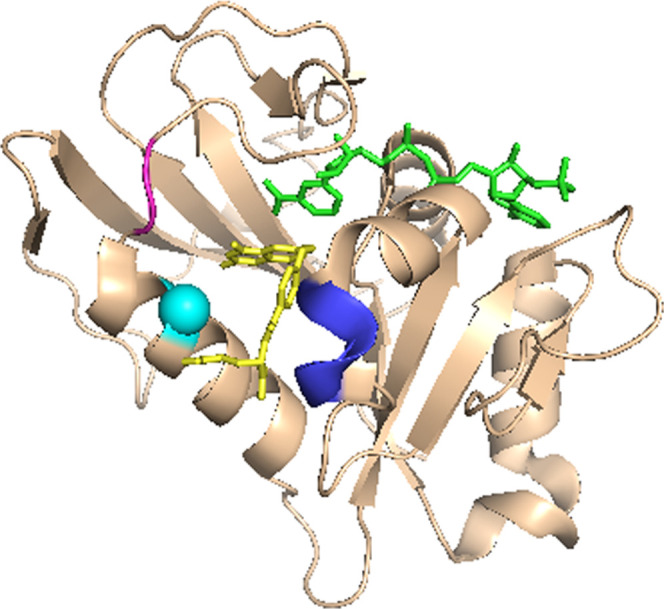
hsDHFR (PDB ID 4M6K) bound to NADP+ and folate. The ligands are shown as green (NADP+) and yellow (folate) sticks; hsDHFR residues corresponding to the PCEs described in1 are shown in cyan (F32), magenta (26-PPLR), and blue (62-PEKN).
In their study, Benkovic et al. created and characterized several ecDHFR mutants to test their ideas about DHFR evolution and the PCEs that they identified and developed a predictive model of the forces that drove these events.1 A notable difference between the cellular environments of prokaryotic and eukaryotic cells is the intracellular pool of NADP+ and NADPH.27 Since the catalytic turnover of both prokaryotic and eukaryotic DHFRs is limited by product release,28,29 there was evolutionary pressure to respond to changing cellular environments while maintaining an efficient rate of enzyme-catalyzed hydride transfer. The PCE that introduced the polyproline sequence restricts the motion of the M20 loop that undergoes dramatic conformational changes during the turnover of ecDHFR.30 The restricted motion prevents significant product inhibition by NADP+ but also dramatically reduces both the efficiency of hydride transfer and the overall turnover rate of the enzyme.1,19 Thus, it appears that other PCEs compensate for the impaired hydride transfer resulting from the polyproline insertion in ecDHFR.
NMR relaxation measurements have shown that insertion of N23PP alone into ecDHFR restricts the ms dynamics of the enzyme.30 In addition, quantum-mechanics/molecular-mechanic (QM/MM) simulations demonstrated that the magnitudes of the conformational changes between the ground state and the transition state of the reaction of N23PP are increased relative to ecDHFR, which is indicative of greater motion of many residues across the enzyme to achieve the short DAD required for hydride transfer.1,31 This increase in the dynamics correlates with a decrease in the single turnover rates1 of the mutant and an increased temperature dependence of the KIEs19 compared to WT ecDHFR. Since there are no natural DHFRs containing only the 23PP insertion, a “humanized” form of ecDHFR was created through insertions of both the polyproline sequence and PEKN (N23PP-G51PEKN ecDHFR). QM/MM simulations of this variant found that the dynamics were restored to those found in WT ecDHFR.1 In addition, both the single turnover rates and the temperature dependence of the KIEs were identical to WT ecDHFR.1,19 Since the “rescuing” PEKN insertion occurred first in the evolution of the enzyme, it appears that the protein motions in DHFR were preserved through evolution and likely play a critical role in enzyme catalysis.
The network of coupled residues that have been extensively studied in the E. coli enzyme20,21,23 has also been shown to contribute to hydride transfer in hsDHFR.25 In the bacterial enzyme, residues M42 and G121, which are distal to the active site, function synergistically to afford an efficient hydride transfer. Sequence alignments show that the equivalent residues in hsDHFR are M53 and S145, respectively. Applying a similar approach of site-directed mutagenesis and investigation of the temperature dependence of the intrinsic KIEs on hydride transfer catalyzed by the variants of the human enzyme showed a similar synergistic effect of these residues. It thus appears that the network of coupled residues in DHFR catalysis is also conserved in evolution. This led us to hypothesize that the PCEs act synergistically to conserve the properties of the enzyme at the TRS, especially the short and narrow distribution of DADs, during evolution.
The current study tests similar effects of PCEs as the aforementioned studies have done in ecDHFR, except we now focus on the effects of backward mutations from hsDHFR. Specifically, we examine the effects of these PCEs on the temperature dependence of the KIE in several site-directed mutants of the human enzyme in which we mutate residues of the human enzyme back to a sequence like that found in the ecDHFR. First, F32 in hsDHFR is sequentially mutated back toward the bacterial enzyme by creating an F32M variant to mimic the form that first appeared ∼797 million years ago and then the F32L variant to mimic ecDHFR. We also study deletion mutants of the PCE insertions identified by Benkovic et al., 26PP and 62-PEKN. Finally, we combine all of these to create a “bacterialized” version of the human enzyme. Each of the residues studied here is highlighted in the X-ray structure of the enzyme with NADP+ and folate bound (Figure 1), and the variants are summarized in Table 1, which also notes which PCE each variant is designed to assess. These studies test the evolutionary importance of 32F, 26PP, and 62-PEKN in maintaining a short and narrow distribution of DADs for effective hydride tunneling. If the effects of these residues on the catalyzed hydride transfer are preserved through evolution, it supports the general idea that evolution will retain optimal TRS properties for the catalyzed H-transfer reactions even when the chemistry is not rate limiting.
Table 1. Summary of hsDHFR Variants Characterized.
| hsDHFR variants | PCEa | divergence (mya)b | region of the enzyme |
|---|---|---|---|
| Δ62-PEKN | 62-PEKN | ∼797 | distal to M20 loop |
| F32M, F32L | F32 | ∼499 | at base of M20 loop |
| Δ26-PPLR-Δ62-PEKN | 26-PPLR | ∼325 | within M20 loop |
| F32L-PP26N-PEKN62G | all PCE |
Phylogenetically coherent events identified in ref (32).
Time of divergence in millions of years (mya).
Materials and Methods
Materials
All chemicals are reagent grade and purchased from Sigma-Aldrich (St. Louis, MO), unless otherwise mentioned. [Carbonyl-14C]-nicotinamide was from Moravek, while [3H]-sodium borohydride was from Perkin-Elmer. Glucose dehydrogenase from Bacillus megaterium is from MP Biomedical. [Carbonyl-14C]-NADPH, 4R-[carbonyl -14C, 4-2H]-NADPH, 4R-3H-NADPH, and DHF were synthesized and stored according to previously published protocols.33−39
Preparation of Mutant Variants
All mutant variants were constructed using the Agilent Stratagene QuickChange site-directed mutagenesis kit using the manufacturer’s instructions. The F32L, F32M, PP26N, and the Δ62-PEKN deletion mutants were derived from the hsDHFR WT plasmid. We constructed the Δ26-PPLR-Δ62-PEKN and the F32L-PP26N-PEKN62G mutants through sequential reactions using different mutant plasmids. For the double-deletion mutant (Δ26-PPLR-Δ62-PEKN), we performed site-directed mutagenesis using the Δ62-PEKN plasmid derived from the WT. Similarly, we constructed the triple mutant (F32L-PP26N-PEKN62G) by replacing 26PP with N using the PP26N primers and then using the resulting plasmid for a subsequent round of mutagenesis in which G replaces 62-PEKN. All mutagenic primers used to generate the hsDHFR variants were from Integrated DNA Technologies (Coralville, IA) and their sequences are listed in Table S1. The PCR products were transformed into E. coli DH5α cells. We extracted plasmids from overnight cultures to confirm the sequences by automated DNA sequencing at the University of Iowa DNA Core Facility. All hsDHFR variants were expressed and purified following published protocols described previously.2
Competitive Intrinsic KIE Measurements
We measured intrinsic KIEs as described previously for human and ecDHFR.2,18 In summary, reacting the protium- or tritium-labeled NADPH at the 4R position with unlabeled DHF at the desired temperature at pH 9.0 in 50 mM MTEN buffer (50 mM MES, 25 mM Tris, 25 mM ethanolamine, and 100 mM sodium chloride) yielded a measure of the H/T KIE. The carbonyl carbon on the nicotinamide ring of the cofactor was labeled with 14C to serve as a tracer for the conversion of protiated NADPH to product (NADP+). An excess of methotrexate quenched the reactions at different time intervals, and we stored the reaction mixture on dry ice. Oxygen bubbled into the quenched reaction mixture ensured that THF was completely oxidized before high-performance liquid chromatography (HPLC) analysis. D/T experiments followed the same protocol but with deuterium at the 4R position. The depletion of tritium in the substrate as a function of fractional conversion yielded the observed KIE on the second-order rate constant kcat/KM. Intrinsic KIEs were then extracted from the observed H/T and D/T KIEs using the Northrop equation (eq 1)40
| 1 |
In eq 1, T(V/K)Hobs and T(V/K)Dobs are the observed H/T and D/T KIEs, respectively, and kH/kT is the intrinsic H/T KIE. The isotope effects on the activation parameters for the intrinsic KIEs are calculated by a nonlinear fit of the data to the Arrhenius equation for intrinsic KIEs (eq 2)
| 2 |
where kl and kh are the rate constants for light and heavy isotopes, respectively, Al/Ah is the isotope effect on the Arrhenius pre-exponential factor, ΔEa is the difference in energy of activation between the two isotopologues, R is the gas constant, and T is the absolute temperature.
Steady-State Kinetics
The steady-state kinetic parameters of the deletion mutants were determined in 50 mM MTEN buffer at pH 7.65 and 25 °C for comparison to the WT enzyme.2 We were unable to measure the single turnover rate of hsDHFR due to a high rate constant even at 4 °C leading to the reaction occurring within the dead time of the stopped flow instrument (∼2 ms). Thus, the effects of the deletion mutants of hsDHFR residues were limited to the steady-state turnover of the enzyme. In short, initial rates were determined with ∼50 nM of the enzyme by following the decrease in absorbance of NADPH at 340 nm, which occurred when DHF and NADPH convert to THF and NADP+. The kinetic parameters with DHF were determined using the 0.1–1.0 μM substrate and those with NADPH were determined using 0.3–1.5 μM nicotinamide. For each set of experiments, the second substrate was kept constant at a saturating concentration of 100 μM. Plots of the initial rate as a function of the substrate concentration were fit to the Michaelis–Menten equation for one substrate to determine the kinetic parameters.
Results and Discussion
The first PCE investigated here is F32, which was originally an L (L28 in ecDHFR). This PCE occurred after the PEKN insertion but before the development of PPLR. The transition away from the L residue in ecDHFR occurs essentially simultaneously with the insertion of PEKX that ultimately becomes PEKN in hsDHFR. It is reasonable to speculate that mutation of L to F32 occurred as a way of restricting the motion of the M20 loop to regulate the binding of NADPH and release of NADP+ before the more substantial PPLR insertion developed. To test this idea, we use site-directed mutagenesis of hsDHFR and measure the intrinsic KIEs on hydride transfer. Following the evolutionary process backward, we first mutate F32 in hsDHFR to M to mimic the transitional form of DHFR and then to L as in ecDHFR.
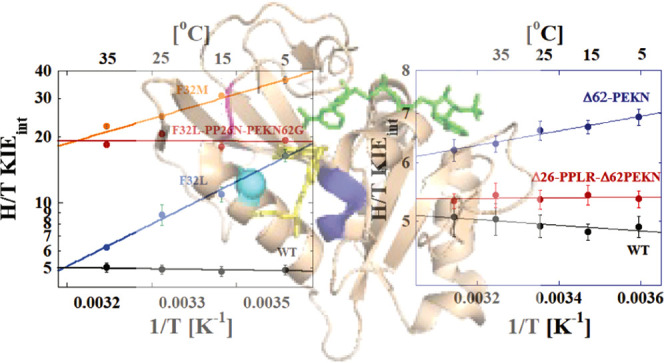
Figure 2 shows the temperature dependence of the intrinsic KIEs on hydride transfer for the F32 variants of hsDHFR. Replacing F32 with M to resemble a transitional DHFR has a large effect on both the magnitude and temperature dependence of the intrinsic KIE for the enzymatic reaction. The size of the KIE is higher than any hsDHFR measured previously. Importantly, the temperature dependence of the KIE (ΔEa(T-H)) is substantially larger than for the WT enzyme with a value of 2.8 ± 0.2 kcal/mol. Fitting the data to the phenomenological activated tunneling model41 yields an average DAD at the TRS of 3.20 ± 0.01 Å (Table 2). Note that this distance is the average DAD, not the DAD at which tunneling occurs. Tunneling can only occur at shorter DADs, which are thermally accessible in the distribution but rare. The significant temperature dependence of the KIE reflects the thermal sampling of the DAD distribution within the active site that is required to reach a DAD that is suitable for hydride tunneling. For F32L, the temperature dependence is even larger, with a ΔEa(T-H) value of 6.5 ± 0.3 kcal/mol. This corresponds to an average DAD at the TRS of 3.48 ± 0.02 Å.
Figure 2.

Arrhenius plots of the H/T KIEs for each hsDHFR PCE at pH 9.0. The data points are the average of at least 5 independent measurements with their standard deviations and the lines are nonlinear fits of all measured KIEs to eq 2. The lines are for WT hsDHFR (black),2 F32L (blue), F3M (orange), and F32L-PP26N-PEKN62G (red).
Table 2. Comparative Isotope Effects for hsDHFR Mutants at pH 9.0.
| hsDHFR variant | AH/AT | ΔEa(T-H) kcal/mol | DADavga (Å) |
|---|---|---|---|
| wild-type hsDHFR | 6.6 ± 0.8 | –0.1 ± 0.1 | 3.05 ± 0.01 |
| F32M | 0.2 ± 0.1 | 2.8 ± 0.2 | 3.20 ± 0.01 |
| F32L | 0.1 ± 0.1 | 6.5 ± 0.3 | 3.48 ± 0.02 |
| F32L-PP26N-PEKN62G | 19.3 ± 6.7 | –0.01 ± 0.2 | 3.10 ± 0.01 |
| Δ62-PEKN | 3.1 ± 0.3 | 0.5 ± 0.1 | 3.10 ± 0.01 |
| Δ26-PPLR-Δ62-PEKN | 5.2 ± 0.5 | 0.1 ± 0.1 | 3.05 ± 0.01 |
From a fit of the data to the model described in ref (41).
Two additional PCEs occurred along with evolution to F32. To fully mimic ecDHFR using the human enzyme as a starting point, we created a triple mutant to account for all three PCEs identified by Benkovic et al.1 This triple mutant replaces F32 with L and removes both the rigidifying PP mutation through the replacement of 26PP with N and the flexibility inducing PEKN insertion by replacing 62-PEKN with G. We identify this triple mutant as F32L-PP26N-PEKN62G. Remarkably, the triple mutant is not only still active but also, based on the temperature-independent KIE values in Figure 2, able to achieve a narrow distribution of DADs at the TRS similar to both hsDHFR and ecDHFR. The ΔEa(T-H) value is −0.01 ± 0.05 kcal/mol and an average DAD of 3.09 ± 0.01 Å, which is strikingly similar to the value for WT hsDHFR. Notably, however, the average KIE is ∼2–3 times larger than that for ecDHFR, which reflects the fact that while the distribution of DADs is narrow and close to that for the more evolved enzyme, it is not fully optimized at the shortest possible DAD that would be expected for a more evolved enzyme.
After identifying the PCEs of DHFR evolution, Benkovic et al. explored these in the bacterial enzyme through insertion of both PEKN and PP into the ecDHFR.1 Insertion of PP alone was found to dramatically impair both the steady-state and single turnover rates of the enzyme. These insertions were also later shown to affect the temperature dependence of the intrinsic KIEs on the reaction.19 The effect of the PP insertion was mitigated by also inserting PEKN in a manner similar to what we find for the triple mutant, the F32L-PP26N-PEKN62G hsDHFR variant described above. To test the effects of the 62-PEKN and the 26-PPLR PCEs in hsDHFR, we construct single and double-deletion mutants of these sites and measure the temperature dependence of the intrinsic KIEs. If our current hypotheses described above for the role of these insertions are correct, we would expect that these deletion mutants would behave in a fashion similar to the ecDHFR insertion variants. In addition, since the single turnover rate of WT hsDHFR cannot be measured due to a rate constant that is too fast for the dead time of the apparatus,2 we measure the steady-state turnover of the deletion mutants with both DHF and NADPH as substrates.
The first deletion variant we consider removes the first PCE in the hsDHFR evolution (62-PEKN; blue in Figure 3) but leaves the PPLR sequence intact. Deletion of 62-PEKN results in KIEs that are both larger and more temperature-dependent compared to the WT hsDHFR enzyme. A fit of the data for the Δ62-PEKN mutant to the Arrhenius equation yields a ΔEa(T-H) of 0.5 ± 0.1 kcal/mol (Table 1). Within the context of the activated tunneling model,41 this result suggests that the mutant enzyme is unable to achieve the short and narrow distribution of DADs required for efficient hydride tunneling with an average DAD of 3.10 Å. This enzyme retains the polyproline sequence that restricts M20 loop movement to control ligand flux but lacks the compensatory effects that the PEKN insertion provides. The deletion of PEKN alone also significantly reduces the overall turnover rate and the catalytic efficiency of the enzyme with both NADPH and DHF as substrates (Table 3). Similar effects were observed for the N23PP mutant of ecDHFR1 where restricting the M20 loop alone in the enzyme resulted in a disrupted TRS that manifests in impaired steady-state kinetic parameters1 and increased temperature dependence of the intrinsic KIE.19
Figure 3.

Arrhenius plots of the H/T KIEs for hsDHFR and the deletion mutant variants at pH 9.0. The data points are the average of at least 5 independent measurements with their standard deviations and the lines are nonlinear fits of all measured KIEs to eq 2. The lines are for WT hsDHFR (black),2 Δ62-PEKN (blue), and Δ26-PPLR-Δ62-PEKN (red).
Table 3. Steady-State Kinetic Parameters of Wild-Type and Mutant hsDHFR at pH 7.65 and 25 °Ca,b.
| wild-typec | Δ26-PPLR-Δ62-PEKN | Δ62-PEKN | |
|---|---|---|---|
| kcatDHF, (s–1) | 14.03 ± 0.02 | 11.28 ± 0.26 | 1.72 ± 0.04 |
| KmDHF, (μM) | 0.29 ± 0.05 | 0.40 ± 0.04 | 0.59 ± 0.06 |
| (kcat/Km)DHF × 107 (M–1 s–1) | 4.84 ± 0.83 | 2.82 ± 0.29 | 0.29 ± 0.03 |
| kcatNADPH, (s–1) | 14.10 ± 0.01 | 11.23 ± 0.13 | 1.73 ± 0.04 |
| KmNADPH, (μM) | 0.36 ± 0.05 | 0.54 ± 0.03 | 0.77 ± 0.08 |
| (kcat/Km)NADPH × 107 (M–1 s–1) | 3.92 ± 0.54 | 2.08 ± 0.12 | 0.22 ± 0.02 |
kcatDHF and KmDHF are the turnover number and Michaelis constant with DHF as the substrate, measured in the presence of 100 μM NADPH and varying concentrations of DHF.
kcatNADPH and KmNADPH are the turnover number and Michaelis constant with NADPH as the substrate, measured in the presence of 100 μM DHF and varying concentrations of NADPH.
From ref (2).
The PCE that introduced flexibility in DHFR occurred incredibly early in the evolution of the enzyme (PEKN). This flexibility likely allowed for the subsequent rigidification that the polyproline sequence produces to modulate the binding and release of NADPH and NADP+. Consistent with the results for the F32L-PP26N-PEKN62G described above, the double-deletion variant that removes both 62-PEKN and 26-PPLR from hsDHFR restores the ability of the enzyme to achieve an optimal TRS for hydride transfer as is evident from the temperature-independent KIE (red line in Figure 3; ΔEa(T-H) = 0.1 ± 0.1 kcal/mol in Table 1). As shown in Table 3, deletion of 26-PPLR and 62-PEKN largely restores the overall rate of catalytic turnover as well as the catalytic efficiencies with both NADPH and DHF as substrates. Finally, fitting the data for the Δ26-PPLR-Δ62-PEKN variant to the phenomenological activated tunneling model41 gives an average DAD at the TRS that is identical to that of the WT (3.05 Å; Table 1).
The results of these variants show that the changes caused by the deletion of the PEKN and PPLR PCEs follow precisely the expected trend based on the previous studies of the insertions of these sequences in ecDHFR. That result is significant because it corroborates the model suggested by the previous studies regarding the effects of these sequences on the substrate binding and the catalyzed reaction. Although that outcome may seem obvious and expected based on the prior studies, it is nevertheless impressive because the effects of evolutionary changes are not always so consistent with predictions.
Our results provide strong support for the model that Benkovic et al.1 put forth to understand the physical effects of the PCEs that occur between ecDHFR and hsDHFR. The changing ligand concentrations in the intracellular environments of prokaryotic and eukaryotic cells placed evolutionary pressure on DHFR whose activity is essential for cellular function. Large-scale conformational changes of the M20 loop in ecDHFR allow the enzyme to achieve the short and narrow distribution of DADs at the TRS for efficient hydride transfer, but it also makes the enzyme susceptible to inhibition by NADP+.1 Such inhibition reduces both the turnover rate and catalytic efficiency of the enzyme, which would be catastrophic to cellular metabolism in the presence of higher concentrations of NADP+. This sequence suggests that the enzyme may have evolved initially to increase the flexibility of the entire protein through insertion of the PEKN sequence. The subsequent PP insertion and mutations at L28 could then restrict the motions of the M20 loop while maintaining enough flexibility in the active site to achieve the short and narrow distribution of DADs required for efficient hydride transfer and avoiding product inhibition. The evolutionary studies of DHFR in this and previous work suggest that a delicate balance is required for efficient catalysis in different cellular environments.
Conclusions
We use site-directed mutagenesis and measurements of intrinsic KIEs to study the PCEs identified by Benkovic et al.1 in the human DHFR enzyme. In each case, we prepare mutant variants that remove the PCEs from the hsDHFR sequence to bacterialize the enzyme. We find that single mutations or deletions of some of the key PCEs perturb the distribution of DADs at the TRS leading to longer average DADs with a wider distribution that results in thermal sampling giving rise to a temperature-dependent KIE for the catalyzed hydride transfer. Preparing either the triple mutant (F32L-PP26N-PEKN62G) or the double-deletion variant (Δ26-PPLR-Δ62-PEKN), however, effectively rescues the catalytic properties of the enzyme resulting in a temperature-independent KIE suggesting a narrow DAD distribution centered at an average that is only slightly larger than that for the WT hsDHFR, Thus, the bacterialized hsDHFR has properties that are very similar to those for WT ecDHFR. These results are consistent with prior studies of the humanized ecDHFR.19 In that study, the PP insertion into ecDHFR altered the DAD distribution leading to a temperature-dependent KIE. In contrast, the double insertion mutant with both the PP and PEKN sequences added to ecDHFR exhibited a temperature-independent KIE indicative of recovery of a DAD distribution that was similar to that for the WT ecDHFR. Thus, our results support the physical model for these PCEs that was first hypothesized by Benkovic et al., and they are consistent with prior studies of ecDHFR variants that mimic the same evolutionary PCEs.
Acknowledgments
This work was supported by NIH (R01GM65368), NSF (CHE-1149023) to A.K., NSF CHE-1707598 to C.M.C. and a Departmental Research Grant from The Robert A. Welch Foundation AC-0006 to K.F.
Glossary
Abbreviations
- (DHFR)
dihydrofolate reductase
- (NADPH)
reduced nicotinamide adenine diphosphate
- (DHF)
7,8-dihydrofolate
- (THF)
5,6,7,8-tetrahydrofolate
- (KIEs)
kinetic isotope effects
- (WT)
wild-type
- (ecDHFR)
Escherichia coli dihydrofolate reductase
- (hsDHFR)
human dihydrofolate reductase
- (DAD)
donor–acceptor distance
- (TRS)
tunneling ready state
- (PCEs)
phylogenetically coherent events
Supporting Information Available
The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/acs.biochem.1c00558.
Tables showing the primers used in this study and the observed and intrinsic H/T and D/T KIEs for each variant studied are provided (PDF)
Accession Codes
Author Contributions
A.K. and C.M.C. led and oversaw the studies and J.L., P.S., and K.F. conducted the experiments. The manuscript was written through contributions of all authors who have given approval to the final version of the manuscript.
The authors declare no competing financial interest.
Supplementary Material
References
- Liu C. T.; Hanoian P.; French J. B.; Pringle T. H.; Hammes-Schiffer S.; Benkovic S. J. Functional significance of evolving protein sequence in dihydrofolate reductase from bacteria to humans. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 10159–10164. 10.1073/pnas.1307130110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis K.; Sapienza P. J.; Lee A. L.; Kohen A. The Effect of Protein Mass Modulation on Human Dihydrofolate Reductase. Biochemistry 2016, 55, 1100–1106. 10.1021/acs.biochem.5b00945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis K.; Kohen A. Protein motions and the activation of the CH bond catalyzed by dihydrofolate reductase. Curr. Opin. Chem. Biol. 2014, 21, 19–24. 10.1016/j.cbpa.2014.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glowacki D. R.; Harvey J. N.; Mulholland A. J. Taking Ockham’s razor to enzyme dynamics and catalysis. Nat. Chem. 2012, 4, 169–176. 10.1038/nchem.1244. [DOI] [PubMed] [Google Scholar]
- Hay S.; Scrutton N. S. Good vibrations in enzyme-catalysed reactions. Nat. Chem. 2012, 4, 161–168. 10.1038/nchem.1223. [DOI] [PubMed] [Google Scholar]
- Kamerlin S. C.; Warshel A. At the dawn of the 21st century: Is dynamics the missing link for understanding enzyme catalysis?. Proteins 2010, 78, 1339–1375. 10.1002/prot.22654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klinman J. P.; Kohen A. Hydrogen tunneling links protein dynamics to enzyme catalysis. Annu. Rev. Biochem. 2013, 82, 471–496. 10.1146/annurev-biochem-051710-133623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klinman J. P.; Offenbacher A. R. Understanding Biological Hydrogen Transfer Through the Lens of Temperature Dependent Kinetic Isotope Effects. Acc. Chem. Res. 2018, 51, 1966–1974. 10.1021/acs.accounts.8b00226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagel Z. D.; Klinman J. P. Update 1 of: Tunneling and dynamics in enzymatic hydride transfer. Chem. Rev. 2010, 110, PR41–67. 10.1021/cr1001035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schramm V. L. Enzymatic transition states, transition-state analogs, dynamics, thermodynamics, and lifetimes. Annu. Rev. Biochem. 2011, 80, 703–732. 10.1146/annurev-biochem-061809-100742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz S. D. Protein dynamics and the enzymatic reaction coordinate. Top. Curr. Chem. 2013, 337, 189–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruiz-Pernía J. J.; Behiry E.; Luk L. Y. P.; Loveridge E. J.; Tunon I.; Moliner V.; Allemann R. K. Minimization of dynamic effects in the evolution of dihydrofolate reductase. Chem. Sci. 2016, 7, 3248–3255. 10.1039/C5SC04209G. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luk L. Y. P.; Loveridge E. J.; Allemann R. K. Protein motions and dynamic effects in enzyme catalysis. Phys. Chem. Chem. Phys. 2015, 17, 30817–30827. 10.1039/C5CP00794A. [DOI] [PubMed] [Google Scholar]
- Scott A. F.; Luk L. Y. P.; Tunon I.; Moliner V.; Allemann R. K. Heavy Enzymes and the Rational Redesign of Protein Catalysts. ChemBioChem 2019, 20, 2807–2812. 10.1002/cbic.201900134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy S.; Schopf P.; Warshel A. Origin of the Non-Arrhenius Behavior of the Rates of Enzymatic Reactions. J. Phys. Chem. B 2017, 121, 6520–6526. 10.1021/acs.jpcb.7b03698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warshel A.; Bora R. P. Perspective: Defining and quantifying the role of dynamics in enzyme catalysis. J. Chem. Phys. 2016, 144, 180901 10.1063/1.4947037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohen A. Dihydrofolate reductase as a model for studies of enzyme dynamics and catalysis [version 1; peer review: 2 approved]. F1000Res. 2015, 4, 1464 10.12688/f1000research.6968.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sikorski R. S.; Wang L.; Markham K. A.; Rajagopalan P. T.; Benkovic S. J.; Kohen A. Tunneling and coupled motion in the Escherichia coli dihydrofolate reductase catalysis. J. Am. Chem. Soc. 2004, 126, 4778–4779. 10.1021/ja031683w. [DOI] [PubMed] [Google Scholar]
- Francis K.; Stojkovic V.; Kohen A. Preservation of protein dynamics in dihydrofolate reductase evolution. J. Biol. Chem. 2013, 288, 35961–35968. 10.1074/jbc.M113.507632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh P.; Francis K.; Kohen A. Network of remote and local protein dynamics in dihydrofolate reductase catalysis. ACS Catal. 2015, 5, 3067–3073. 10.1021/acscatal.5b00331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh P.; Sen A.; Francis K.; Kohen A. Extension and limits of the network of coupled motions correlated to hydride transfer in dihydrofolate reductase. J. Am. Chem. Soc. 2014, 136, 2575–2582. 10.1021/ja411998h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stojković V.; Perissinotti L. L.; Willmer D.; Benkovic S. J.; Kohen A. Effects of the donor-acceptor distance and dynamics on hydride tunneling in the dihydrofolate reductase catalyzed reaction. J. Am. Chem. Soc. 2012, 134, 1738–1745. 10.1021/ja209425w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L.; Goodey N. M.; Benkovic S. J.; Kohen A. Coordinated effects of distal mutations on environmentally coupled tunneling in dihydrofolate reductase. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 15753–15758. 10.1073/pnas.0606976103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C. T.; Francis K.; Layfield J. P.; Huang X.; Hammes-Schiffer S.; Kohen A.; Benkovic S. J. Escherichia coli dihydrofolate reductase catalyzed proton and hydride transfers: temporal order and the roles of Asp27 and Tyr100. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 18231–18236. 10.1073/pnas.1415940111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J.; Fortunato G.; Lin J.; Agarwal P. K.; Kohen A.; Singh P.; Cheatum C. M. Evolution Conserves the Network of Coupled Residues in Dihydrofolate Reductase. Biochemistry 2019, 58, 3861–3868. 10.1021/acs.biochem.9b00460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh P.; Vandemeulebroucke A.; Li J.; Schulenburg C.; Fortunato G.; Kohen A.; Hilvert D.; Cheatum C. M. Evolution of the Chemical Step in Enzyme Catalysis. ACS Catal. 2021, 11, 6726–6732. 10.1021/acscatal.1c00442. [DOI] [Google Scholar]
- Weikl T. R.; Boehr D. D. Conformational selection and induced changes along the catalytic cycle of Escherichia coli dihydrofolate reductase. Proteins 2012, 80, 2369–2383. 10.1002/prot.24123. [DOI] [PubMed] [Google Scholar]
- Appleman J. R.; Beard W. A.; Delcamp T. J.; Prendergast N. J.; Freisheim J. H.; Blakley R. L. Unusual transient- and steady-state kinetic behavior is predicted by the kinetic scheme operational for recombinant human dihydrofolate reductase. J. Biol. Chem. 1990, 265, 2740–2748. 10.1016/S0021-9258(19)39864-3. [DOI] [PubMed] [Google Scholar]
- Fierke C. A.; Johnson K. A.; Benkovic S. J. Construction and evaluation of the kinetic scheme associated with dihydrofolate reductase from Escherichia coli. Biochemistry 1987, 26, 4085–4092. 10.1021/bi00387a052. [DOI] [PubMed] [Google Scholar]
- Bhabha G.; Lee J.; Ekiert D. C.; Gam J.; Wilson I. A.; Dyson H. J.; Benkovic S. J.; Wright P. E. A dynamic knockout reveals that conformational fluctuations influence the chemical step of enzyme catalysis. Science 2011, 332, 234–238. 10.1126/science.1198542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luk L. Y.; Javier Ruiz-Pernia J.; Dawson W. M.; Roca M.; Loveridge E. J.; Glowacki D. R.; Harvey J. N.; Mulholland A. J.; Tunon I.; Moliner V.; Allemann R. K. Unraveling the role of protein dynamics in dihydrofolate reductase catalysis. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 16344–16349. 10.1073/pnas.1312437110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C. T.; Hanoian P.; French J. B.; Pringle T. H.; Hammes-Schiffer S.; Benkovic S. J. Functional significance of evolving protein sequence in dihydrofolate reductase from bacteria to humans. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 10159–10164. 10.1073/pnas.1307130110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agrawal N.; Kohen A. Microscale synthesis of 2-tritiated isopropanol and 4R-tritiated reduced nicotinamide adenine dinucleotide phosphate. Anal. Biochem. 2003, 322, 179–184. 10.1016/j.ab.2003.08.012. [DOI] [PubMed] [Google Scholar]
- Blakley R. L. Crystalline dihydropterinoylglutamic acid. Nature 1960, 188, 231–232. 10.1038/188231a0. [DOI] [Google Scholar]
- Markham K. A.; Sikorski R. S.; Kohen A. Synthesis and utility of 14C-labeled nicotinamide cofactors. Anal. Biochem. 2004, 325, 62–67. 10.1016/j.ab.2003.10.027. [DOI] [PubMed] [Google Scholar]
- Markham K. A.; Sikorski R. S.; Kohen A. Purification, analysis, and preservation of reduced nicotinamide adenine dinucleotide 2′-phosphate. Anal. Biochem. 2003, 322, 26–32. 10.1016/j.ab.2003.07.010. [DOI] [PubMed] [Google Scholar]
- Sen A.; Stojkovic V.; Kohen A. Synthesis of radiolabeled nicotinamide cofactors from labeled pyridines: versatile probes for enzyme kinetics. Anal. Biochem. 2012, 430, 123–129. 10.1016/j.ab.2012.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sen A.; Yahashiri A.; Kohen A. Triple isotopic labeling and kinetic isotope effects: exposing H-transfer steps in enzymatic systems. Biochemistry 2011, 50, 6462–6468. 10.1021/bi2003873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh P.; Guo Q.; Kohen A. Chemoenzymatic Synthesis of Ubiquitous Biological Redox Cofactors. Synlett 2017, 28, 1151–1159. 10.1055/s-0036-1588768. [DOI] [Google Scholar]
- Northrop D. B.Enzyme Mechanism from Isotope Effects; CRC Press: Boca Raton, FL, 1991; pp 181–202. [Google Scholar]
- Roston D.; Cheatum C. M.; Kohen A. Hydrogen donor-acceptor fluctuations from kinetic isotope effects: a phenomenological model. Biochemistry 2012, 51, 6860–6870. 10.1021/bi300613e. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.