

Abstract
This study aims at evaluating the ameliorative role of Terminalia catappa aqueous leaf extract (TCA) on hyperglycaemia-induced oxidative stress and inflammation in a high-fat, low dose streptozotocin-induced type 2 diabetic rat model. Experimental rats were treated orally with 400 and 800 mg/kg bw TCA daily for four weeks. Antioxidant enzyme activities, plasma glucose concentration, protein concentration, oxidative stress, and inflammation biomarkers were assayed using standard methods. Hepatic relative expressions of tumour necrosis factor-alpha (TNF-α), interleukin-six (IL-6), and nuclear factor-erythroid 2 related factor 2 (Nrf-2) were also assessed. Molecular docking and prediction of major TCA phytoconstituents' biological activity related to T2DM-induced oxidative stress were evaluated in silico. Induction of diabetes significantly (p < 0.05) reduced superoxide dismutase, glutathione-S-transferase, and peroxidase activities. Glutathione and protein stores were significantly (p < 0.05) depleted, while glucose, MDA, interleukin-six (IL-6), and tumour necrosis factor-α (TNF-α) concentrations were significantly (p < 0.05) increased. A significant (p < 0.05) upregulation of hepatic TNF-α and IL-6 expression and downregulation (p < 0.05) of Nrf-2 expression were observed during diabetes onset. TCA treatment significantly (p < 0.05) modulated systemic diabetic-induced oxidative stress and inflammation, mRNA expression dysregulation, and dysregulated macromolecule metabolism. However, only 800 mg/kg TCA treatment significantly (p < 0.05) downregulated hepatic TNF-α expression. 9-Oxabicyclo[3.3.1]nonane-2,6-diol and 1,2,3-Benzenetriol bound comparably to glibenclamide in Nrf-2, IL-6, and TNF-α binding pockets. They were predicted to be GST A and M substrate, JAK2 expression, ribulose-phosphate 3-epimerase, NADPH peroxidase, and glucose oxidase inhibitors. These results suggest that TCA ameliorates hyperglycaemia-induced oxidative stress and inflammation by activating Nrf-2 gene.
1. Introduction
Diabetes mellitus (DM) is a metabolic and endocrine disease of chronic nature which arises from a lack of insulin secretion, action, or both. It is characterised majorly by chronic hyperglycaemia, disturbance in the intermediary metabolism of major macromolecules (carbohydrate, protein, and lipid), and vascular complications arising from organ/system dysfunction [1, 2]. According to the International Diabetes Foundation (IDF), as of 2019, over 450 million adults were living with diabetes worldwide. This number is expected to increase by 2045 to about 700 million, with most of the diabetes risk and burden being type 2 diabetes mellitus (T2DM) [3]. There is a paradigm shift in Africa, in which the major focus of the continent's health system has moved from infectious diseases to noncommunicable diseases. This recent trend is due to increased urbanisation and lifestyle changes [4–6]. Irrespective of the paradigm shift, the use of medicinal plants in disease management has not changed. Terminalia catappa is one of such plants found in Nigeria with numerous pharmacological properties [7]. The leaf is a rich source of antioxidant and anti-inflammatory principles, notably flavonoids, phenols, and terpenes [1, 8].
The progression of many chronic diseases has been hinged on inflammation and oxidative stress interactions. Different studies have shown that, just like other noncommunicable diseases, oxidative stress plays a significant role in T2DM pathogenesis, progression, and complications [9–12]. Oxidative stress arises whenever the cellular redox balance is disrupted, leading to loss of biomolecular integrity and membrane damage. It promotes diabetes onset and aggravates this disorder and its associated complications by disrupting insulin release and action [13–15]. An increase in free radicals generates further oxidative stress by inducing mitochondria uncoupling, depleting the antioxidant defence system, and eliciting chronic inflammation by activating stress-activated kinases. Stimulation of proinflammatory cytokine and chemokine gene expression with subsequent increase in the circulation of these proinflammatory mediators results from stress kinase activation [16]. Dysfunction in glucose metabolism and chronic hyperglycaemia utilise oxidative stress and inflammatory mechanisms in the pathogenesis of diabetes-related complications [17]. Hyperglycaemia stimulates intra- and extracellular free radical generation inducing oxidative stress, concomitantly stimulating the generation of proinflammatory mediators [18]. It leads to the formation of toll-like receptors and inflammasome-complexes, which are key inducers for inflammation through nuclear factor-kappa B (NF-κB) activation and oxidative stress progression [19]. Irregular epigenetic modifications and activation of interleukin-6 (IL-6), tumour necrosis factor-α (TNF-α), and NF-κB genes with concomitant downregulation of superoxide dismutase 2 (SOD2), nuclear factor-erythroid 2 related factor 2 (Nrf-2), paraoxonase-1 (PON 1), and other antioxidant-related genes are also common features [20]. Damage of macromolecules is the downstream consequence of hyperglycaemic-induced oxidative stress, with the degree of damage associated with the duration of exposure [21].
Numerous pieces of information show that hyperglycaemia-induced oxidative stress is a major player in the pathogenesis and progression of T2DM and its related complications; hence, tackling it cannot be overemphasised [11, 17, 22–25]. With the increasing prevalence and incidence of T2DM in Nigeria and sub-Saharan Africa [2, 26], oxidative stress has become a focal point of interest in diabetic research [11]. Despite the reported antioxidant activities of oral antioxidant agents such as vitamins, their usefulness in preventing diabetic complications is elusive as human studies have shown that they do not decrease oxidative stress [27–29]. This observation has been attributed to their null effect on blood glucose levels [30]. Therefore, controlling oxidative stress and inflammation while maintaining glycaemic control is imperative in managing T2DM and preventing complications associated with diabetes. Medicinal plants and natural antioxidants with hypoglycaemic activity are hypothesised as a possible solution due to a plethora of phytochemicals and nonnutritional compounds [31]. Despite the reports on the antioxidant property of T. catappa in diabetic and nondiabetic animal models [32–35], information on the molecular mode of antioxidant and anti-inflammatory activity of T. catappa in T2DM animal model is lacking. This study evaluates the role of Terminalia catappa aqueous leaf extract in ameliorating hyperglycaemia-induced oxidative stress and inflammation in high-fat diet (HFD)/low-dose streptozotocin-induced diabetic rats. The extract's role on the expression of some genes relevant to redox status and inflammation was also evaluated.
2. Materials and Methods
2.1. Chemicals and Reagents
One-step RT-PCR kit (TransGen EasyScript®) was purchased from TransGen Biotech Co., Ltd. (Beijing, China). Glucose diagnostic kit was purchased from Randox Diagnostics (Ireland, UK). Streptozotocin (STZ), tumour necrosis factor-alpha (TNF-α), and interleukin-6 (IL-6) ELISA kits were products of Solarbio Science and Technology (Beijing, China). All other chemicals, unless stated otherwise, were purchased from Sigma-Aldrich, Germany.
2.2. Collection, Identification, and Preparation of Plants
Fresh mature leaves of T. catappa (TC) were acquired from Covenant University, Ota, Nigeria, and authenticated (FHI 112775) in Forest Research Institute of Nigeria (FRIN), Ibadan, Nigeria. The leaves were then air-dried and aqueous crude extract (TCA) was prepared as outlined by Iheagwam, Okeke [36]. Briefly, TC leaves were washed, air-dried under shade, and macerated in distilled water and the resulting filtrate was concentrated to dryness in a rotary evaporator to yield TCA.
2.3. Experimental Animals and Husbandry
Six-to-eight-week-old Male Wistar rats (n = 30) weighing 200 ± 20 g were used for this study. The animals were acclimatised for two weeks before the experiment, maintained under standard conditions (12 hrs light/dark cycle, room temperature (23 ± 2°C), humidity (50 ± 5%)), and provided food and water ad libitum. The experimental protocol was approved by Covenant University Health, Research and Ethics Committee (CHREC/031/2018), following institution guidelines for animal care.
2.4. Experimental Design
Rats (n = 30) were randomly divided into five [5] groups of 6 rats: Group I, normal rats treated with vehicle (distilled water) alone; Group II, diabetic rats treated with vehicle alone; Group III, diabetic rats treated with glibenclamide (10 mg/kg bw); Group IV, diabetic rats treated with 400 mg/kg bw of TCA; and Group V, diabetic rats treated with 800 mg/kg bw of TCA. The experimental period and dosage were based on a similar design utilised in the safety evaluation study of TCA [36, 37].
2.5. Diabetes Induction
T2DM was induced using a high-fat diet (HFD) that was self-formulated as shown in Table 1 and low-dose STZ (30 mg/kg bw) according to the method of Stalin et al. [38] with slight modification. Briefly, rats were fed with HFD to induce insulin resistance for eight weeks. Thereafter, DM was induced by intraperitoneal injection of STZ (30 mg/kg bw in citrate buffer [0.1 M, pH 4.5]) at 1 mL/kg bw. Five days after STZ injection, the tail vein of the rats was pricked and fasting blood glucose (FBG) was analysed. Rats with FBG > 200 mg/dL were considered diabetic and included in the study in which TCA treatment started on the 6th day after STZ induction. Administration of TCA, glibenclamide, and vehicle was done daily according to the experimental design by gastric intubation for 28 days. Fasting blood glucose levels and animal body weight were also recorded during the experiment. Rats were fasted for over 12 hours and thereafter anaesthetised using xylazine/ketamine (5 : 50 g/g) after the experimental period.
Table 1.
Normal and high-fat diet chow formulation.
| Feed composition | Weight (g/kg) | |
|---|---|---|
| Normal diet | High-fat diet | |
| Maize | 450 | 50 |
| Beef tallow | 0 | 400 |
| Groundnut cake | 100 | 100 |
| Flour binder | 100 | 100 |
| Soybean meal | 50 | 50 |
| Full fat soya | 50 | 50 |
| Palm kernel cake | 50 | 50 |
| Fish meal | 100 | 100 |
| Wheat offal | 50 | 50 |
| Bone | 10 | 10 |
| Premix∗ | 30 | 30 |
| Methionine | 6 | 6 |
| Lysine | 4 | 4 |
∗ Contained (per kg): vitamin A (4 000 000 IU), vitamin D3 (800 000 IU), vitamin E (8 000 IU), vitamin K3 (0.9 g), thiamine (0.7 g), riboflavine (2 g), pyridoxine (1.2 g), vitamin B12 (0.006 g), nicotinic acid (11 g), pantothenic acid (3 g), folic acid (3 g), biotin (0.02 g), choline (120 g), CuSO4·5H2O (2 g), CoCl2·6H2O (0.008), NaCl (2 g), FeSO4·7H2O (8 g), KI (0.48 g), MnSO4·7H2O (32 g), CaSO4 (14 g), and ZnSO4 (20 g).
2.6. Sample Collection and Preparation
Blood was collected from the heart through cardiac puncture, placed in heparin, and separated to obtain plasma and erythrocytes [39]. Liver and kidney were excised, prepared, and stored as previously reported [36, 40].
2.7. Oxidative Stress and Biochemical Assays
Assessment of oxidative stress, inflammation, and other biochemical parameters was carried out in the plasma, erythrocytes, kidney, and liver. The activity of superoxide dismutase (SOD) was determined using the pyrogallol method of Marklund and Marklund [41], while the formation of 1-chloro-2,4-dinitrobenzene and glutathione conjugate was used to analyse glutathione-S-transferase (GST) activity according to the method of Habig et al. [42]. The concentration of reduced glutathione (GSH) was estimated using the method of Sedlak and Lindsay [43], while peroxidase (Px) activity was assayed following the method of Chance and Maehly [44]. The concentration of thiobarbituric reactive substances (MDA) was tested to determine lipid peroxidation as described by Buege and Aust [45]. Tumour necrosis factor-α (TNF-α) and interleukin-6 (IL-6) were analysed using Solarbio ELISA kits, while plasma blood glucose was assayed using Randox kit according to the manufacturers' instructions. Protein concentration was assayed according to the method of Lowry et al. [46].
2.8. RNA Extraction and Expression Analysis
Total hepatic RNA extraction using Trizol® reagent (TransGen Biotech Co., China) and reverse transcriptase-polymerase chain reaction (RT-PCR) analysis for TNF-α, IL-6, and nuclear factor-erythroid 2 related factor 2 (Nrf-2) were determined according to the methodology of Stalin et al. [38] with slight modification. RT-PCR was performed using TransScript® II One-Step RT-PCR kit (TransGen Biotech Co., China) and run in a C1000 thermal cycler (Bio-Rad, CA, USA) following set parameters:
Initial denaturation at 95°C for 5 min;
Denaturation at 95°C for 30 sec;
Annealing at 51–53°C for 30 sec as indicated in Table 2 for each primer;
Extension at 72°C for 30 sec;
Final extension at 72°C for 7 min. (×40 cycles).
Table 2.
Gene-specific primer sequence.
| Gene | Sequence (5′-3′) | Annealing temperature (°C) |
|---|---|---|
| IL-6 | 5′-ATTGTATGAACAGCGATGATGCAC-3′ (F) | 51 |
| 5′-CCAGGTAGAAACGGAACTCCAGA-3′ (R) | ||
| Nrf-2 | 5′-GGGCAAAAGCTCTCCATATTCC-3′ (F) | 52 |
| 5′-GAGCGGCAACTTTATTCTTCCC-3′ (R) | ||
| TNF-α | 5′-ACGGCATGGATCTCAAAGAC-3′ (F) | 53 |
| 5′-CGGACTCCGCAAAGTCTAAG-3′ (R) | ||
| GAPDH | 5′-CTGACATGCCGCCTGAAAC-3′ (F) | 51 |
| 5′-CCAGCATCAAAGGTGGAAGAA-3′ (R) |
The gene-specific primers in Table 2 were used for first-strand cDNA synthesis, while GAPDH served as a reference. Ethidium bromide-stained agarose gel (1.5%) (Sigma Aldrich, Germany) was used to run the PCR products and viewed under UV light (UVP BioDoc-It™ Imaging System, Upland, CA, USA). Each experiment was repeated three times.
2.9. In Silico Analysis of TCA Major Phytochemicals
2.9.1. Protein Modelling, Ligand Modelling, and Molecular Docking
In a previous report [1], 9-Oxabicyclo[3.3.1]nonane-2,6-diol and 1,2,3-Benzenetriol were identified as the major phytochemicals in TCA, accounting for 11.02% and 9.63%, respectively, of the total phytochemical constituents. Their structures were downloaded from PubChem and energy was minimised as reported by Iheagwam et al. [1]. The 3D crystal structures of human TNF-α and IL-6 were obtained from the RCSB protein data bank with PDB codes 5MU8 and 4O9H, respectively. There was no crystal structure of human Nrf-2 present in the RCSB protein data bank. The amino acid sequence of Nrf-2 (AAB32188.1) was obtained from NCBI for homology modelling using SWISS-MODEL. The sequence was queried using BLASTp and a suitable template was selected based on the target sequence, identity, and query coverage correlation. The modelled Nrf-2 was validated using the Ramachandran plot [47].
2.9.2. Molecular Docking
Molecular docking was carried out according to the methodology of Iheagwam et al. [47]. AutoDock 4.2 was used to add Gasteiger charges and assign nonpolar hydrogen to ligands, selected diabetic drugs (metformin and glibenclamide), and protein targets (Nrf-2, TNF-α, and IL-6). AutoDock Vina was used to run the docking simulation of the prepared ligands and selected diabetic drugs in the active sites of the prepared proteins with the set grid map for each target at 0.375 Å, as shown in Table S1.
2.9.3. Computational Activity and ADMET Prediction
The biological activity profile of 9-Oxabicyclo[3.3.1]nonane-2,6-diol and 1,2,3-Benzenetriol related to T2DM-induced oxidative stress was predicted using the prediction of activity spectra for substances- (PASS-) based approach [48]. The predicted activity of the compounds is estimated based on their structural formula identifying probable activity (Pa) and inactivity (Pi), which ranges from 0.000 to 1.000. However, only activities in which Pa > Pi and Pa > 0.7 were considered probable. The pharmacokinetics and toxicity properties of the compounds were predicted using SwissADME [49] and vNN-ADMET [50], respectively.
2.10. Data Analysis
Generated data were analysed using IBM Statistical Package for the Social Sciences v.23 (IBM Inc., New York, USA) and expressed as mean ± SEM of six biological replicates except otherwise stated. Statistical significance (p < 0.05) was determined using one-way analysis of variance (ANOVA) supplemented with Duncan multiple range test for post hoc analyses.
3. Results
3.1. T. Catappa Effect on HFD/STZ-Induced Weight Loss and Hyperglycaemia
In Figure 1, diabetes onset led to a significant (p < 0.05) weight loss of 35.2% in HFD/STZ-induced diabetic rats compared to 19.7% weight gain in normal rats at the end of the experiment. Administration of TCA significantly (p < 0.05) reduced the weight loss, with 0.5% weight gain recorded for 800 TCA administered group compared with glibenclamide (weight loss of 6.5%) and diabetic groups. Unlike animal weight, after diabetes induction, the fasting blood sugar was significantly (p < 0.05) raised in the diabetic rats compared with normal rats. Diabetic rats treated with TCA exhibited a dose-dependent decrease (p < 0.05) in fasting blood sugar after 14 and 28 days compared to untreated diabetic rats (Figure 2).
Figure 1.
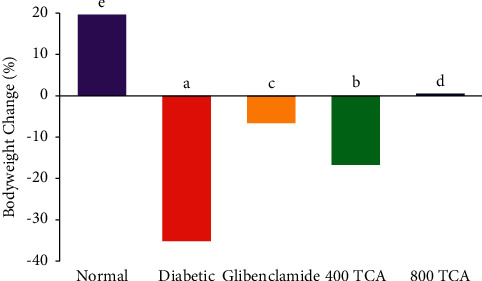
Effect of T. catappa aqueous extract treatment on bodyweight changes in HFD/STZ-induced diabetic rats. Bars represent proportion. Bars with different superscripts are significantly different at p < 0.05.
Figure 2.
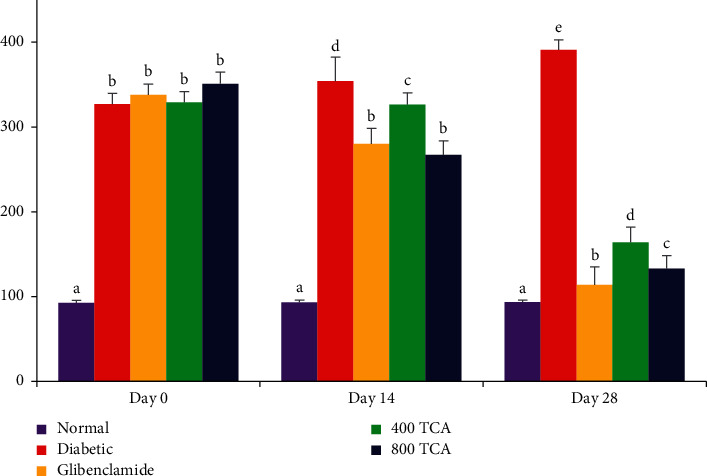
Effect of T. catappa aqueous extract treatment on fasting blood glucose in HFD/STZ-induced diabetic rats after 14 and 28 days of treatment. Bars represent mean ± SEM (n = 6). Bars with different superscripts are significantly different, while those with the same superscripts are not significantly different at p < 0.05.
3.2. T. Catappa Ameliorates HFD/STZ-Induced Systemic Oxidative Stress
The data for TCA treatment on enzymatic antioxidant activities are illustrated in Figures 3–5. In HFD/STZ-induced diabetic rats, there was a significant (p < 0.05) reduction in plasma, hepatic, and erythrocyte SOD activities by 47.1, 27.9, and 18.0% compared with normal rats. The plasma and hepatic SOD activities in diabetic rats treated with TCA were restored significantly (p < 0.05) to normal in a dose-dependent manner with a 110.9 and 54.8% respective increase recorded at the highest dose compared with the diabetic group. Nonetheless, the opposite was the case as TCA administration could not improve (p > 0.05) erythrocyte SOD activity at both doses compared with the diabetic group. There was no significant (p > 0.05) difference in renal SOD activity after diabetic insult and TCA treatment (Figure 3). Induction of diabetes did not significantly (p > 0.05) alter the activities of Px in the liver, kidney, and erythrocytes. However, the activity of plasma Px was significantly (p < 0.05) reduced by 87.2% in diabetic rats compared with normal rats. This observation was significantly (p < 0.05) improved upon by 333.3 and 486.7% in a dose-dependent manner after 400 and 800 mg/kg bw TCA treatment, respectively, compared with the diabetic group (Figure 4). In Figure 5, diabetes induction led to a significant (p < 0.05) reduction in plasma and hepatic GST activities by 77.7 and 80.9%, respectively, compared with normal rats. After 4-week TCA administration, plasma and hepatic GST activities were significantly (p < 0.05) increased in diabetic rats compared with the diabetic group. The observation was dose-dependent only in plasma GST activity as 76.2 and 206% increases were the respective ratios in 400 and 800 mg/kg bw TCA treatment. Induction of diabetes did not significantly (p > 0.05) alter the activities of GST in the kidney and erythrocytes (Figure 5).
Figure 3.
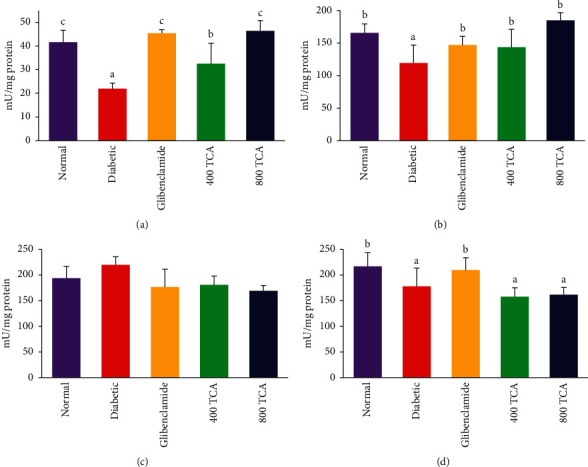
Effect of T. catappa aqueous extract treatment on (a) plasma, (b) hepatic, (c) renal, and (d) erythrocyte superoxide dismutase (SOD) activity in HFD/STZ-induced diabetic rats. Bars represent mean ± SEM (n = 6). Bars with different superscripts are significantly different, while those without superscripts are not significantly different at p < 0.05.
Figure 4.
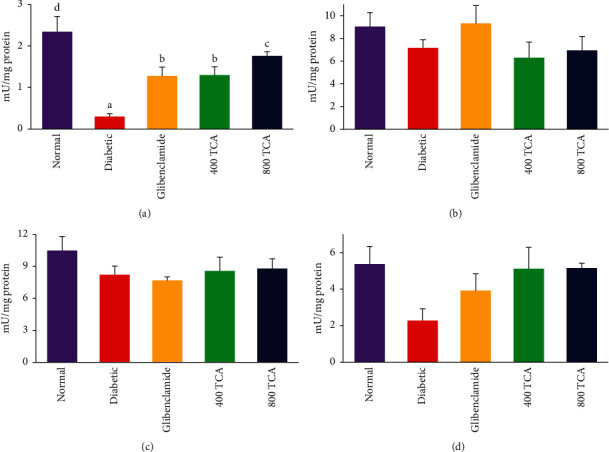
Effect of T. catappa aqueous extract treatment on (a) plasma, (b) hepatic, (c) renal, and (d) erythrocyte peroxidase (Px) activity in HFD/STZ-induced diabetic rats. Bars represent mean ± SEM (n = 6). Bars with different superscripts are significantly different, while those without superscripts are not significantly different at p < 0.05.
Figure 5.
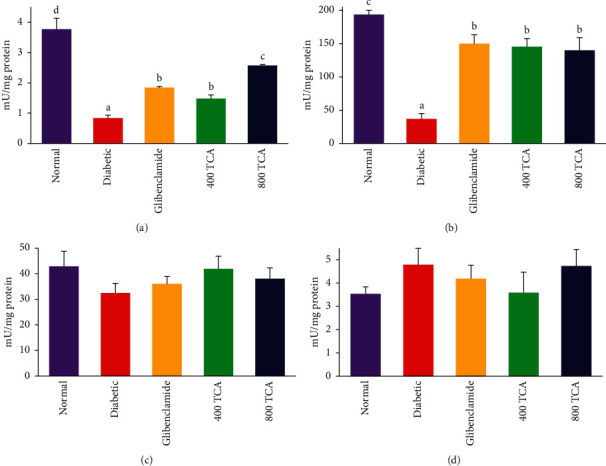
Effect of T. catappa aqueous extract treatment on (a) plasma, (b) hepatic, (c) renal, and (d) erythrocyte glutathione-S-transferase (GST) activity in HFD/STZ-induced diabetic rats. Bars represent mean ± SEM (n = 6). Bars with different superscripts are significantly different, while those without superscripts are not significantly different at p < 0.05.
The data for TCA treatment on nonenzymatic antioxidant concentrations are illustrated in Figures 6 and 7. Administration of TCA at 400 and 800 mg/kg bw to diabetic rats significantly (p < 0.05) improved the plasma (95.4 and 97.4%, respectively), hepatic (97 and 127.1%, respectively), renal (254.1 and 308.6%, respectively), and erythrocytes (53.4 and 90.4%, respectively) GSH concentrations to near-normal levels when compared with the diabetic group. Plasma, hepatic, renal, and erythrocytes GSH concentrations were initially depleted significantly (p < 0.05) by 58.3, 76.3, 83.7, and 55.9%, respectively, after induction of diabetes when compared with normal rats (Figure 6). After diabetes induction, plasma, hepatic, renal, and erythrocyte MDA concentrations were significantly (p < 0.05) increased by 70.9, 784.6, 502.7, and 665.2%, respectively, compared with the normal rats. Upon 400 and 800 mg/kg bw TCA treatment, the MDA concentration of the diabetic rats was reduced significantly (p < 0.05). On the other hand, only renal and erythrocyte MDA levels of the rats were reduced to the normal level, while the reduction was dose-dependent in the kidney (57.5 and 84.4%, respectively) and liver (63.7 and 81.2%, respectively) (Figure 7).
Figure 6.
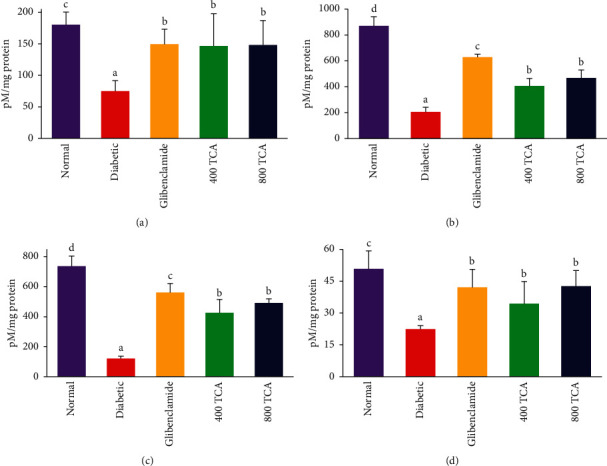
Effect of T. catappa aqueous extract treatment on (a) plasma, (b) hepatic, (c) renal, and (d) erythrocyte reduced glutathione (GSH) concentration in HFD/STZ-induced diabetic rats. Bars represent mean ± SEM (n = 6). Bars with different superscripts are significantly different at p < 0.05.
Figure 7.
Effect of T. catappa aqueous extract treatment on (a) plasma, (b) hepatic, (c) renal, and (d) erythrocyte lipid peroxidation (MDA) concentrations in HFD/STZ-induced diabetic rats. Bars represent mean ± SEM (n = 6). Bars with different superscripts are significantly different at p < 0.05.
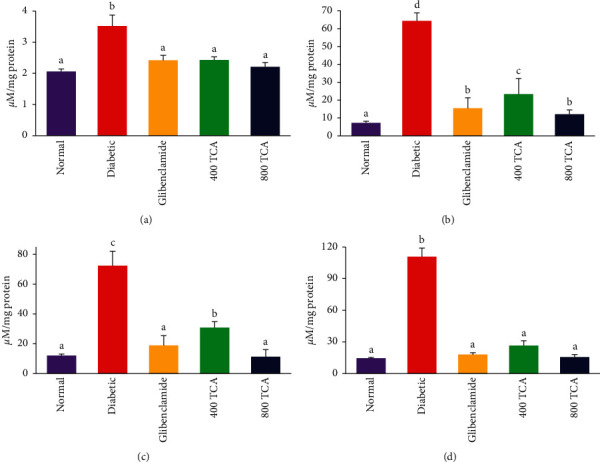
3.3. T. Catappa Suppresses HFD/STZ-Induced Inflammation, Glucose, and Protein Dysregulation
Diabetic onset significantly (p < 0.05) elevated plasma glucose by 153.5% compared with normal rats. TCA 400 and 800 mg/kg bw treatment significantly reduced (p < 0.05) the elevated glucose concentration by 38.3 and 50.9%, respectively, in a dose-dependent manner when compared with the diabetic group. The reduction was not as significant as that observed for the drug group (55.4%). Induction of diabetes resulted in a significant (p < 0.05) increase in plasma and hepatic IL-6 (645.3 and 104.5%, respectively) and TNF-α (69.4 and 479.8%, respectively) concentrations when compared with normal rats. Nonetheless, there was no significant (p > 0.05) difference in renal IL-6 and TNF-α concentrations across all groups. Oral 400 and 800 mg/kg bw TCA treatment significantly (p < 0.05) decreased IL-6 and TNF-α concentrations in the liver by 53.4 and 69.1%, respectively, while 71.9 and 32.6% decreases were the respective ratios in the plasma when compared with diabetic rats. The inverse was the case for plasma and organ protein levels as they were significantly (p < 0.05) depleted after induction of diabetes compared with normal rats. However, upon treatment with TCA, these parameters were significantly (p < 0.05) improved compared to the untreated diabetic rats (Table 3).
Table 3.
Effect of T. catappa aqueous extract treatment on other biochemical parameters in HFD/STZ-induced diabetic rats.
| Normal | Diabetic | Glibenclamide | 400 TCA | 800 TCA | |
|---|---|---|---|---|---|
| Plasma glucose (mg/dL) | 101.42 ± 14.10a | 257.06 ± 15.04d | 114.62 ± 13.13a | 158.51 ± 11.42c | 126.32 ± 14.51b |
|
| |||||
| TNF-α (pg/mL) | |||||
| Plasma | 9.37 ± 1.56a | 15.87 ± 1.58d | 11.27 ± 1.60c | 12.66 ± 3.70c | 10.69 ± 1.11b |
| Liver | 3.37 ± 0.65a | 19.54 ± 1.40c | 11.83 ± 22.10b | 10.21 ± 16.86b | 6.03 ± 10.98a |
| Kidney | 9.92 ± 1.36 | 9.39 ± 2.67 | 9.91 ± 1.88 | 10.33 ± 1.01 | 8.47 ± 1.73 |
|
| |||||
| IL-6 (pg/mL) | |||||
| Plasma | 28.38 ± 6.77a | 211.52 ± 27.00e | 94.22 ± 8.06c | 147.05 ± 14.97d | 59.39 ± 10.28b |
| Liver | 7.75 ± 0.88a | 15.85 ± 1.12e | 13.99 ± 4.62d | 9.36 ± 1.78c | 7.39 ± 1.41b |
| Kidney | 5.36 ± 0.32 | 7.74 ± 0.85 | 6.45 ± 1.87 | 6.35 ± 0.36 | 5.91 ± 1.49 |
|
| |||||
| Total protein (mg/mL) | |||||
| Plasma | 539.60 ± 14.09c | 463.49 ± 2.07a | 501.81 ± 13.75b | 509.76 ± 13.59b | 532.23 ± 13.16c |
| Liver | 154.33 ± 13.49c | 109.35 ± 3.18a | 138.45 ± 4.22b | 132.31 ± 5.04b | 137.11 ± 4.99b |
| Kidney | 124.99 ± 8.12d | 99.55 ± 1.49a | 120.39 ± 0.91c | 109.80 ± 1.90b | 125.41 ± 0.93d |
| Erythrocyte | 133.60 ± 6.38c | 95.08 ± 2.85a | 127.62 ± 10.91b | 123.33 ± 4.24b | 135.71 ± 3.05c |
Data are represented as mean ± SEM (n = 6). Values with different superscripts across a row are significantly different, while those without superscripts are not significantly different at p < 0.05.
3.4. T. Catappa Modulates HFD/STZ-Induced Dysregulation of Inflammatory mRNA Expression
Induction of diabetes significantly (p < 0.05) reduced the expression of hepatic Nrf-2 mRNA in rats by 62.5% compared with the normal group. Treatment of diabetic rats with 400 and 800 mg/kg bw TCA significantly (p < 0.05) improved Nrf-2 expression by 533.3 and 633.3%, respectively, in the experimental group compared with the diabetic animals. The reverse was the case for hepatic IL-6 and TNF-α, as diabetes onset significantly (p < 0.05) increased the expression of these genes by 228.0 and 95.1%, respectively. Upon 400 and 800 mg/kg bw TCA treatment, IL-6 mRNA expression was significantly (p < 0.05) reduced by 77.6 and 65.4%, while only the highest treatment dosage (800 mg/kg) was able to reduce TNF-α mRNA expression by 21.0% when compared with the diabetic group (Figure 8).
Figure 8.
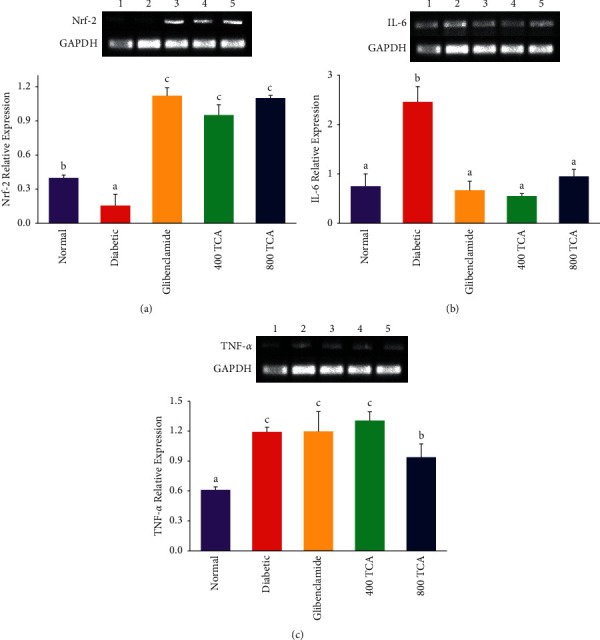
Effect of TCA on (a) Nrf-2, (b) IL-6, and (c) TNF-α mRNA expression in the liver of HFD/STZ-induced diabetic rats. Bars represent mean ± SEM (n = 3). Bars with different superscripts are significantly different at p < 0.05.
3.5. Molecular Docking Study of Major T. Catappa Aqueous Leaf Extract Phytoconstituents
The selected template for the modelled Nrf-2 structure was 2LZ1 based on sequence identity (100%), sequence similarity (0.6), global model quality estimate (GMQE = 0.05), QMEANDisCo score (0.75 ± 0.11), oligomeric state (monomer), and experimental comparison plot superiority over other templates. In the modelled Nrf-2 structure, 89.2, 10.6, and 1.4% of the amino acid residues were in the most favoured, additional allowed, and disallowed region, respectively (Figure S1).
In Table 4, 9-Oxabicyclo[3.3.1]nonane-2,6-diol (−4.5, −4.7, and −5.0 kcal/mol, respectively) and 1,2,3-Benzenetriol (−4.3, −5.3, and −5.6 kcal/mol, respectively) exhibited better binding capacity than metformin (−4.2, −4.6, and −4.3 kcal/mol, respectively) in the pocket of Nrf-2, IL-6, and TNF-α. However, glibenclamide (−7.2, −7.6, and −6.4 kcal/mol, respectively) exhibited the best binding affinity for the three protein targets. For the binding interactions, all ligands were stabilised by hydrogen bonds, pi bonds, and Van der Waals interactions with amino acid residues in the binding site. Unlike other ligands, metformin did not form a pi bond in the binding pocket of the targets but rather distinct interactions such as salt-bridge interaction and attractive bond with Nrf-2 (Glu442) and IL-6 (Glu42), respectively. Pro453 was the only amino acid residue in the binding pocket involved in the stabilising of all ligands in the binding pocket of Nrf-2, while numerous residues were observed for IL-6 (Leu39, Glu42, Thr43, Leu101, Arg104, Ala112, Thr163, and Leu167) and TNF-α (Val17, Ala18, Pro20, Phe144, Ser147, Gly148, Gln149, and Val150) (Table 4). The binding conformations of all ligands in the binding pockets of Nrf-2, IL-6, and TNF-α are shown in Figures S2–S4, respectively.
Table 4.
Molecular docking of hypoglycaemic agents and major TCA phytoconstituents in the binding sites of Nrf-2, IL-6, and TNF-α proteins.
| Compound | BA∗ | H-Bond | Π-Bond | VdW Interactions | Others |
|---|---|---|---|---|---|
| Nrf2 | |||||
| 9-Oxabicyclo[3.3.1]nonane-2,6-diol | −4.5 | Arg501 | Lys500, Arg501, Ala495 | Pro453, Gln496, Asn497, Arg499 | Cys498a |
| 1,2,3-Benzenetriol | −4.3 | His437 | Val454 | Glu435, Leu438, Thr439, Glu442, Pro453, Glu455 | Arg433a |
| Glibenclamide | −7.2 | Asn505, Arg501b | Lys500, Leu503, Pro453, Val454 | Glu435, Leu438, Glu442, Ala445, Lys446, Ile450, Phe452, Glu455, Val507 | — |
| Metformin | −4.2 | Glu435, His437 | — | Arg433, Leu438, Thr439, Pro453, Val454, Glu455, Arg501, Leu503, Asn505 | Glu442c |
|
| |||||
| IL-6 | |||||
| 9-Oxabicyclo[3.3.1]nonane-2,6-diol | −4.7 | Glu99, Asn144 | Val96, Pro141 | Lue92, Glu95, Pro139, Leu148 | — |
| 1,2,3-Benzenetriol | −5.3 | Leu39, Ser108 | Leu39, Leu101, Ala112 | Glu42, Thr43, Arg104, Thr163, Leu167 | — |
| Glibenclamide | −7.6 | Thr43, Arg104, Gln156 | Leu39, Tyr100, Leu101, Ala112 | Glu42, Lys46, Asn103, Ser107, Gln152, Gln159, Asp160, Thr163, Leu167 | — |
| Metformin | −4.6 | Leu39, Glu42b, Thr43b, Arg104 | — | Arg40, Leu101, Ser107, Ser108, Ala112, Thr163, Leu167 | Glu42d |
|
| |||||
| TNF-α | |||||
| 9-Oxabicyclo[3.3.1]nonane-2,6-diol | −5.0 | Ala145, Glu146, Ser147b | Val17, Ala18, Pro20, Val150 | Asp143, Phe144, Gly148, Gln149 | — |
| 1,2,3-Benzenetriol | −5.6 | Ala18, Gly148 | Ala18, Pro20, Val150 | Val17, Phe144, Ala145, Ser147, Gln149, Tyr151 | Val150a, Glu146a |
| Glibenclamide | −6.4 | Glu146, Gly148, Gln149, Val150 | Val17, Pro20 | Ala18, Leu29, Arg32, Ala33, Asn34, Phe144, Ala145, Ser147 | — |
| Metformin | −4.3 | Ala18, Asn34, Ser147b, Gly148 | — | Val17, Pro20, Leu29, Phe144, Gln149, Val150 | — |
∗ kcal/mol, aunfavourable bonds, bcarbon-hydrogen bonds, csalt-bridge interaction, dattractive bond, BA: binding affinity, H: conventional hydrogen, VdW: Van der Waals.
3.6. Predicted Bioactivity and ADMET Properties of Major T. Catappa Aqueous Leaf Extract Phytoconstituents
For the predicted bioactivity, 9-Oxabicyclo[3.3.1]nonane-2,6-diol and 1,2,3-Benzenetriol were predicted to be GST A and M substrates, JAK2 expression, ribulose-phosphate 3-epimerase, NADPH peroxidase, and glucose oxidase inhibitors. HIF1A expression and bisphosphoglycerate phosphatase inhibition were peculiar to 9-Oxabicyclo[3.3.1]nonane-2,6-diol and 1,2,3-Benzenetriol, respectively (Table 5). From Table 6, 9-Oxabicyclo[3.3.1]nonane-2,6-diol and 1,2,3-Benzenetriol were predicted to be very soluble, easily absorbed in the human intestine, bioavailable, easily synthesized, and stable in the hepatic microsome. They were also not predicted to be P-glycoprotein substrate and inhibitors, violators of Lipinski, Veber and Egan rule of drug-likeness, and cytochrome P450 isoforms inhibitor. These ligands were predicted to be unable to cause cardiotoxicity, drug-induced liver injury, cytotoxicity, mitochondrial toxicity, and mutagenicity. However, 1,2,3-Benzenetriol was predicted to easily permeate the blood-brain barrier, inhibit CYP3A4, and cause mitochondrial toxicity. 384 and 90 mg/day were the predicted maximum recommended therapeutic doses for 9-Oxabicyclo[3.3.1]nonane-2,6-diol and 1,2,3-Benzenetriol, respectively (Table 6).
Table 5.
PASS-predicted T2DM-induced oxidative stress activity of major T. catappa aqueous leaf extract phytoconstituents.
| Pa | Pi | Activity |
|---|---|---|
| 9-Oxabicyclo[3.3.1]nonane-2,6-diol | ||
| 0.881 | 0.003 | Ribulose-phosphate 3-epimerase inhibitor |
| 0.855 | 0.008 | HIF1A expression inhibitor |
| 0.809 | 0.008 | JAK2 expression inhibitor |
| 0.802 | 0.009 | GST A substrate |
| 0.798 | 0.012 | NADPH peroxidase inhibitor |
| 0.768 | 0.016 | Glucose oxidase inhibitor |
|
| ||
| 1,2,3-Benzenetriol | ||
| 0.907 | 0.003 | NADPH peroxidase inhibitor |
| 0.898 | 0.003 | JAK2 expression inhibitor |
| 0.892 | 0.004 | Glucose oxidase inhibitor |
| 0.858 | 0.004 | Ribulose-phosphate 3-epimerase inhibitor |
| 0.854 | 0.004 | Bisphosphoglycerate phosphatase inhibitor |
| 0.787 | 0.011 | GST A substrate |
| 0.711 | 0.004 | GST M substrate |
Table 6.
Predicted ADMET properties of major T. catappa aqueous leaf extract phytoconstituents and standard diabetic drugs.
| Absorption and distribution | Drug-likeness | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Solubility class | HIA | BBB | Pgp-S | Pgp-I | LV | VV | EV | B | SA | MRTD∗ | |
| 9-Oxabicyclo[3.3.1]nonane-2,6-diol | Very | High | No | No | No | 0 | 0 | 0 | 0.55 | 4.31 | 384 |
| 1,2,3-Benzenetriol | Very | High | Yes | No | No | 0 | 0 | 0 | 0.55 | 1 | 90 |
| Glibenclamide | Poor | Low | No | No | Yes | 1 | 1 | 0 | 0.55 | 3.42 | 183 |
| Metformin | High | High | No | No | No | 0 | 0 | 0 | 0.55 | 3.02 | 3000 |
|
| |||||||||||
| Metabolism | Toxicity | ||||||||||
| HLM | Cytochrome P450 inhibitor | hERG | DILI | HepG2 | MMP | Ames test | |||||
| 1A2 | 2C19 | 2C9 | 2D6 | 3A4 | |||||||
| 9-Oxabicyclo[3.3.1]nonane-2,6-diol | Yes | No | No | No | No | No | No | No | No | No | No |
| 1,2,3-Benzenetriol | Yes | No | No | No | No | Yes | No | No | No | Yes | No |
| Glibenclamide | No | No | Yes | Yes | No | Yes | No | Yes | No | No | No |
| Metformin | Yes | No | No | No | No | No | No | No | No | No | No |
∗ mg/day, HIA: human intestinal absorption, BBB: blood-brain barrier permeation, Pgp-S: P-glycoprotein substrate, Pgp-I: P-glycoprotein inhibitor, LV: Lipinski violation, VV: Veber violation, EV: Egan violation, B: bioavailability, SA: synthetic accessibility, MRTD: maximum recommended therapeutic dose, HLM: human liver microsomal stability, hERG: human ether-à-go-go-related gene, DILI: drug-induced liver injury, HepG2: HepG2 cytotoxicity, MMP: mitochondrial membrane potential.
4. Discussion
The significant weight loss observed in the diabetic rats may be due to poor glycaemic control. This major anthropometric observation shifts the energy demand to fat and protein stores. Various studies have reported similar findings on the weight reduction in diabetic rats buttressing this study [51–53]. The modulation of weight loss by TCA may be ascribed to the improvement of glycaemic control. The onset of hyperglycaemia in the untreated diabetic rats could result from poor utilisation of circulating glucose as an energy source by GLUT-4-dependent tissues [54]. The hypoglycaemic activity of TCA in diabetic rats, especially at the highest dose, might indicate improvement in the utilisation of glucose by peripheral tissues, further corroborating the modulatory effect of TCA on HFD/STZ-induced weight loss.
In diabetes progression, there is a de novo generation of reactive species, which promotes oxidative stress [55]. In conditions where chronic hyperglycaemia occurs, these generated reactive species further suppress the antioxidant defence systems in various tissues, exacerbating oxidative stress to maintain T2DM progression and complication development [56]. Enzymatic (SOD, Px, and GST) and nonenzymatic antioxidants (GSH and vitamins E and C) are endogenous biomolecules that scavenge free radicals preventing oxidative stress onslaught in cells. SOD scavenges superoxide radicals via enzymatic conversion to hydrogen peroxide, which GPx and catalase then catalyse to yield water and oxygen. The oxidative stress markers were changed in the rats as a result of the onset of diabetes. GSH concentration, SOD, GST, and Px activities were reduced with a concomitant increase in MDA concentration, reflecting impaired antioxidant defence with a subsequent increase in oxidative stress [57, 58]. However, TCA treatment improved the impaired antioxidant system while reducing oxidative stress (Figures 1–5). Previous studies corroborated these findings on the antioxidant effect of plant extracts and isolated natural compounds on HFD/STZ-induced diabetic rats [59, 60]. The increased hepatic GST activity could be ascribed to the phytochemicals present in TCA. Their biological activity prediction as GST substrates further lents credence to the findings. The reduction in the activities of the antioxidant enzymes in the diabetic group could be attributed to an increase in free radicals, glycosylation of enzymes, and loss of cofactors [61]. The deleterious effect of diabetes on SOD activity and lipid peroxidation impacted the erythrocytes more than other tissues. The observed high susceptibility of the erythrocyte to oxidative stress may be due to molecular oxygen, ferrous ions, and polyunsaturated fatty acids presence [16]. In addition, due to the absence of a nucleus and other cell organelles, there is limited repair capacity on oxidative stress-related damage [62]. The absence of these organelles could be the reason why TCA was unable to increase the reduced activity of SOD in the erythrocyte. The inhibition of ATP production by glycation products may have been responsible for the depletion of the erythrocyte GSH level [63]. These findings were similar to a previous report in alloxan-induced diabetic rats [64]. The inability of diabetic onset to affect renal enzymatic antioxidants might be due to the length of experimental duration, which was not sufficient to cause this complication, with nephropathy reported to occur gradually [65].
The observed improvement of the impaired antioxidant system of the organs and plasma might also be ascribed to the upregulation of Nrf-2 expression and proinflammatory gene downregulation (Figure 5). Nrf-2 is a transcription factor that induces phase II detoxifying and antioxidant enzymes, playing an important role in the antioxidant response, glucose metabolism, and prevention of DM onset [66–68]. It has also been reported to interact with Kelch-like ECH-associated protein 1 (Keap1) to protect organs/tissues of diabetics against diabetes-induced organopathy [69]. The observed increase in the concentration of these inflammatory mediators (IL-6 and TNF-α) after diabetes induction in rats was suppressed by TCA treatment, suggesting the anti-inflammatory effect of TCA in diabetic rats (Table 3). Proinflammatory cytokines are associated with diabetes progression through subclinical chronic inflammation, an independent risk factor for T2DM development [59]. Impaired hepatocyte insulin signalling and inhibition of insulin secretion from the β-cells of the pancreas after glucose stimulation is associated with elevated IL-6 [59]. During excessive ROS generation, activated NFκB activates the release of other proinflammatory cytokines, instigating further ROS generation and establishing oxidative stress in the process [70, 71]. These inflammatory cytokines can interfere with insulin receptors on tissues and their signalling pathways, leading to complications such as insulin resistance [72]. Insulin resistance is directly associated with an increase in inflammatory biomarker concentration. These inflammatory mediators alter β-cell function through direct action or other secondary pathways to maintain T2DM onset [16]. Upregulation of proinflammatory cytokines mRNA would increase the level of tissue and systemic circulating levels of these cytokines. They are well documented to be associated with dysregulating downstream processes in the insulin signalling pathway [73, 74]. TCA's downregulation of these inflammatory genes may have resulted in reversing the dysregulation of insulin signalling, concomitantly reducing hyperglycaemia and its resultant oxidative and inflammatory effects. This probable mechanism has been postulated by a previous study [75]. In addition, the synthesis of proteins could be realised as one of the downstream effects of insulin signalling dysregulation, repair, and reversal. It is noteworthy that systemic protein levels are affected due to a defect in glucose metabolism as protein store impairment is a consequence of diabetes [76]. Proper utilisation of plasma glucose as a source of energy in TCA treatment groups may have resulted in suppressing protein store dysregulation, making it available for the synthesis of antioxidant enzymes and glutathione.
The binding conformations of the ligands in Nrf-2, IL-6, and TNF-α active sites suggest that TCA phytoconstituents and glibenclamide interacted with these proteins in their active site. Thus, TCA may probably ameliorate oxidative stress and inflammation via a similar mechanism as glibenclamide [38]. Also, the allosteric binding of 9-Oxabicyclo[3.3.1]nonane-2,6-diol in IL-6 active pocket may hint at TCA's probable additional anti-inflammatory mechanism. A study on the network pharmacology of andrographolide reported similar allosteric binding and interactive amino acid residues in the binding pocket of IL-6 [77]. A study on the possible activation of Nrf-2 by vitamin E and curcumin by Mishra et al. [78] reported a different binding conformation to our study. This observation could result from the size of their ligands, hence the preference for the binding pocket located at the c-terminal. The better binding affinity exhibited by glibenclamide over metformin on proteins associated with hyperglycaemia-induced oxidative stress and inflammation buttresses the choice of hypoglycaemic agent in this study.
The predicted inhibition of Janus kinase-2 (JAK2) expression, hypoxia-inducible factor-1α (HIF1A) expression, and NADPH peroxidase would suggest possible mechanisms through which TCA might alleviate glucose-induced toxicity (Table 4). Oxidative stress is known to be closely associated with activation or overexpression of NADPH peroxidase, while JAK2 expression enhances reactive species and proinflammatory cytokine generation [79]. Inhibition of JAK2 and HIF1A expression can partially attenuate hyperglycaemia-elicited oxidative stress [80, 81]. Glucose oxidase catalyses glucose conversion to gluconolactone and subsequently to glucuronic acid and H2O2 [82]. Ribulose-phosphate 3-epimerase and bisphosphoglycerate phosphatase are enzymes found in the pentose phosphate pathway and the glycolytic pathway. During diabetes onset, these enzymes catalyse reactions that give rise to increased glyceraldehyde-3-phosphate, which is subsequently converted to methylglyoxal inducing free radical generation, proinflammation, and oxidative stress [83]. Predicted inhibition of these enzymes by TCA phytochemicals may be potential mechanisms of tackling oxidative stress in the diabetic state. Pa was >0.7, suggesting that the pharmacological action experimental validation would probably be similar to the predicted results. The predicted mitochondria toxicity of 1,2,3-Benzenetriol might be due to its ability to generate H2O2. This toxic effect is usually counteracted by reduction of therapeutic dose as predicted and use of other natural antioxidants [84]. Irrespectively, the predicted ADMET properties and obedience of 9-Oxabicyclo[3.3.1]nonane-2,6-diol and 1,2,3-Benzenetriol to the drug-likeness rules would suggest these compounds as potential leads in controlling oxidative stress and inflammation while maintaining glycaemic control.
5. Conclusion
In conclusion, the aqueous extract of T. catappa (TCA) leaves possesses significant antioxidant and anti-inflammatory effects, which may be ascribed to the upregulation of the Nrf-2 gene and downregulation of inflammatory cytokine genes. These pharmacological activities are reported for the first time and may help slow down and overcome DM-related complications associated with hyperglycaemia-induced oxidative stress and inflammation. The inability to investigate the effect of TCA on glucose metabolism and Nrf-2 signalling pathway elements was a limitation in this study which requires further research. Further studies can be carried out to isolate these antioxidant principles from T. catappa leaves and experimentally validate their in silico prediction results as possible antioxidant mechanisms of TCA in T2DM.
Acknowledgments
The authors acknowledge Olawumi Toyin Iheagwam for proofreading the manuscript. Covenant University Centre for Research, Innovation and Discovery (CUCRID) is duly acknowledged.
Data Availability
All the data are included in the manuscript and supplementary text.
Conflicts of Interest
The authors declare that they have no actual or potential competing financial interest.
Authors' Contributions
FNI, OOO, and SNC conceptualized the study. FNI and GEB developed the methodology and performed formal analysis and investigation. FNI wrote the original draft. All the authors reviewed, edited, and approved the manuscript.
Supplementary Materials
Table S1: grid map specifications for molecular docking. Figure S1: representation of (a) modelled Nrf-2 structure and (b) corresponding Ramachandran plot. Figure S2: 2D representation of (a) 9-Oxabicyclo[3.3.1]nonane-2,6-diol, (b) 1,2,3-Benzenetriol, (c) glibenclamide, and (d) metformin interactions in the binding pocket of Nrf-2. Green, light blue, pink, orange, violet, and broken red lines represent conventional hydrogen, carbon-hydrogen, pi-alkyl, salt bridge, pi-sigma, and unfavourable bonds, respectively. Light green circles represent Van der Waals interactions. Figure S3: 2D representation of (a) 9-Oxabicyclo[3.3.1]nonane-2,6-diol, (b) 1,2,3-Benzenetriol, (c) glibenclamide, and (d) metformin interactions in the binding pocket of IL-6. Green, light blue, pink, orange, and violet broken lines represent conventional hydrogen, carbon-hydrogen, pi-alkyl, attractive, and pi-sigma bonds, respectively. Light green circles represent Van der Waals interactions. Figure S4: 2D representation of (a) 9-Oxabicyclo[3.3.1]nonane-2,6-diol, (b) 1,2,3-Benzenetriol, (c) glibenclamide, and (d) metformin interactions in the binding pocket of TNF-α. Green, light blue, pink, violet, and broken red lines represent conventional hydrogen, carbon-hydrogen, pi-alkyl, pi-sigma, and unfavourable bonds, respectively. Light green circles represent Van der Waals interactions.
References
- 1.Iheagwam F. N., Israel E. N., Kayode K. O., De Campos O. C., Ogunlana O. O., Chinedu S. N. GC-MS analysis and inhibitory evaluation of Terminalia catappa leaf extracts on major enzymes linked to diabetes. Evidence-Based Complementary and Alternative Medicine . 2019;2019 doi: 10.1155/2019/6316231.6316231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yesufu J. O., Oluwasile O. D., Oluranti O. I., Fasanmade A. A., Soladoye A. O. Cardiopulmonary health indices and diabetes risk scores in undergraduate students of a private university in Nigeria. Beni-Suef University Journal of Basic and Applied Sciences . 2020;9(1):1–7. doi: 10.1186/s43088-019-0032-x. [DOI] [Google Scholar]
- 3.International Diabetes Federation. IDF Diabetes Atlas . 9th. Brussels, Belgium: IDF; 2019. [PubMed] [Google Scholar]
- 4.Gassasse Z., Smith D., Finer S., Gallo V. Association between urbanisation and type 2 diabetes: an ecological study. BMJ Global Health . 2017;2(4) doi: 10.1136/bmjgh-2017-000473.e000473 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Koch M. Urbanisation, inequality, and non-communicable disease risk. The Lancet Diabetes and Endocrinology, . 2017;5(5):p. 313. doi: 10.1016/S2213-8587(17)30116-X. [DOI] [PubMed] [Google Scholar]
- 6.Shi Y., Hu F. B. The global implications of diabetes and cancer. The Lancet . 2014;383(9933):1947–1948. doi: 10.1016/s0140-6736(14)60886-2. [DOI] [PubMed] [Google Scholar]
- 7.Das G., Kim D.-Y., Fan C., et al. Plants of the genus Terminalia: an insight on its biological potentials, pre-clinical and clinical studies. Frontiers in Pharmacology . 2020;11 doi: 10.3389/fphar.2020.561248.561248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Divya N., Vijaya Anand A. Phytochemical investigation and in vitro anti-diabetic activity of Terminalia catappa leaves. International Journal of Phytopharmacy . 2014;4:132–134. [Google Scholar]
- 9.Alipour M., Salehi I., Ghadiri Soufi F. Effect of exercise on diabetes-induced oxidative stress in the rat hippocampus. Iranian Red Crescent Medical Journal . 2012;14(8):222–228. [PMC free article] [PubMed] [Google Scholar]
- 10.Brownlee M. The pathobiology of diabetic complications: a unifying mechanism. Diabetes . 2005;54(6):1615–1625. doi: 10.2337/diabetes.54.6.1615. [DOI] [PubMed] [Google Scholar]
- 11.Pitocco D., Tesauro M., Alessandro R., Ghirlanda G., Cardillo C. Oxidative stress in diabetes: implications for vascular and other complications. International Journal of Molecular Sciences . 2013;14(11):21525–21550. doi: 10.3390/ijms141121525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Prattichizzo F., De Nigris V., Mancuso E., et al. Short-term sustained hyperglycaemia fosters an archetypal senescence-associated secretory phenotype in endothelial cells and macrophages. Redox Biology . 2018;15:170–181. doi: 10.1016/j.redox.2017.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ferrannini E., Baldi S., Frascerra S., et al. Shift to fatty substrate utilization in response to sodium-glucose cotransporter 2 inhibition in subjects without diabetes and patients with type 2 diabetes. Diabetes . 2016;65(5):1190–1195. doi: 10.2337/db15-1356. [DOI] [PubMed] [Google Scholar]
- 14.Rorsman P., Braun M. Regulation of insulin secretion in human pancreatic islets. Annual Review of Physiology . 2013;75(1):155–179. doi: 10.1146/annurev-physiol-030212-183754. [DOI] [PubMed] [Google Scholar]
- 15.Speakman J. R., Selman C. The free-radical damage theory: accumulating evidence against a simple link of oxidative stress to ageing and lifespan. BioEssays . 2011;33(4):255–259. doi: 10.1002/bies.201000132. [DOI] [PubMed] [Google Scholar]
- 16.Oguntibeju O. O. Type 2 diabetes mellitus, oxidative stress and inflammation: examining the links. International Journal of Physiology, Pathophysiology and Pharmacology . 2019;11(3):45–63. [PMC free article] [PubMed] [Google Scholar]
- 17.Butkowski E. G., Jelinek H. F. Hyperglycaemia, oxidative stress and inflammatory markers. Redox Report . 2017;22(6):257–264. doi: 10.1080/13510002.2016.1215643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dayre A., Pouvreau C., Butkowski E. G., de Jong B., Jelinek H. F. Diabesity increases inflammation and oxidative stress. International Journal of Pharmaceutical Sciences and Developmental Research . 2016;2(1):12–18. doi: 10.17352/ijpsdr.000006. [DOI] [Google Scholar]
- 19.Luc K., Schramm-Luc A., Guzik T. J., Mikolajczyk T. P. Oxidative stress and inflammatory markers in prediabetes and diabetes. Journal of Physiology and Pharmacology . 2019;70(6):809–824. doi: 10.26402/jpp.2019.6.01. [DOI] [PubMed] [Google Scholar]
- 20.Kang Q., Yang C. Oxidative stress and diabetic retinopathy: molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biology . 2020;37 doi: 10.1016/j.redox.2020.101799.101799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rehman K., Akash M. S. H. Mechanism of generation of oxidative stress and pathophysiology of type 2 diabetes mellitus: how are they interlinked? Journal of Cellular Biochemistry . 2017;118(11):3577–3585. doi: 10.1002/jcb.26097. [DOI] [PubMed] [Google Scholar]
- 22.Charlton A., Garzarella J., Jandeleit-Dahm K. A., Jha J. C. Oxidative stress and inflammation in renal and cardiovascular complications of diabetes. Biology . 2021;10(1):p. 18. doi: 10.3390/biology10010018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ortega Á. L. Oxidative stress in diabetic retinopathy. Antioxidants . 2021;10(1):p. 50. doi: 10.3390/antiox10010050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pouvreau C., Dayre A., Butkowski E., De Jong B., Jelinek H. F. Inflammation and oxidative stress markers in diabetes and hypertension. Journal of Inflammation Research . 2018;11:61–68. doi: 10.2147/jir.s148911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tiwari B. K., Pandey K. B., Abidi A. B., Rizvi S. I. Markers of oxidative stress during diabetes mellitus. Journal of Biomarkers . 2013;2013 doi: 10.1155/2013/378790.378790 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Uloko A. E., Musa B. M., Ramalan M. A., et al. Prevalence and risk factors for diabetes mellitus in Nigeria: a systematic review and meta-analysis. Diabetes Therapy . 2018;9(3):1307–1316. doi: 10.1007/s13300-018-0441-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ceriello A., Testa R. Antioxidant anti-inflammatory treatment in type 2 diabetes. Diabetes Care . 2009;32(2):S232–S236. doi: 10.2337/dc09-s316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Choi S. W., Ho C. K. Antioxidant properties of drugs used in Type 2 diabetes management: could they contribute to, confound or conceal effects of antioxidant therapy? Redox Report . 2018;23(1):1–24. doi: 10.1080/13510002.2017.1324381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Golbidi S., Alireza Ebadi S., Laher I. Antioxidants in the treatment of diabetes. Current Diabetes Reviews . 2011;7(2):106–125. doi: 10.2174/157339911794940729. [DOI] [PubMed] [Google Scholar]
- 30.Rajendiran D., Packirisamy S., Gunasekaran K. A review on role of antioxidants in diabetes. Asian Journal of Pharmaceutical and Clinical Research . 2018;11(2):48–53. doi: 10.22159/ajpcr.2018.v11i2.23241. [DOI] [Google Scholar]
- 31.Jebur A. B., Mokhamer M. H., El-Demerdash F. M. A review on oxidative stress and role of antioxidants in diabetes mellitus. Austin Endocrinology and Diabetes Case Reports . 2016;1(1):1006–1011. [Google Scholar]
- 32.Annegowda H. V., Mordi M. N., Ramanathan S., Mansor S. M. Analgesic and antioxidant properties of ethanolic extract of Terminalia catappa L. leaves. International Journal of Pharmacology . 2010;6(6):910–915. doi: 10.3923/ijp.2010.910.915. [DOI] [Google Scholar]
- 33.Ben E. E., Asuquo A. E., Owu D. U. Comparative effect of aspirin, meloxicam and Terminalia catappa leaf extract on serum levels of some inflammatory markers in alloxan induced diabetic rats. Asian Journal of Research in Biochemistry . 2019;4(1):1–10. doi: 10.9734/ajrb/2019/v4i130058. [DOI] [Google Scholar]
- 34.Ben E. E., Asuquo A. E., Owu D. U. Possible amelioration of oxidative stress damage via cyclo-oxygenase pathway by aqueous extract of Terminalia catappa leaves in alloxan induced diabetic rats. GSC Biological and Pharmaceutical Sciences . 2021;16(2):38–48. doi: 10.30574/gscbps.2021.16.2.0215. [DOI] [Google Scholar]
- 35.Dada F. A., Oyeleye S. I., Adefegha S. A., Babatola L. J., Adebayo A. Evaluation of different almond (Terminalia catappa) extracts against oxidative stress induced by cyclosporine in brain and liver of rats. Journal of Complementary and Integrative Medicine . 2021 doi: 10.1515/jcim-2020-0193. [DOI] [PubMed] [Google Scholar]
- 36.Iheagwam F. N., Okeke C. O., DeCampos O. C., Okere D. U., Ogunlana O. O., Chinedu S. N. Safety evaluation of Terminalia catappaLinn (Combretaceae) aqueous leaf extract: sub-acute cardio-toxicopathological studies in albino Wistar rats. Journal of Physics: Conference Series . 2019;1299(1) doi: 10.1088/1742-6596/1299/1/012109.012109 [DOI] [Google Scholar]
- 37.Iheagwam F. N., Okeke C. O., De Campos O. C., Adegboye B. E., Ogunlana O. O., Chinedu S. N. Toxicopathological, proinflammatory and stress response evaluation of Terminalia catappa extract in male Wistar rats. Toxicology Reports . 2021;8:1769–1776. doi: 10.1016/j.toxrep.2021.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Stalin A., Irudayaraj S. S., Gandhi G. R., Balakrishna K., Ignacimuthu S., Al-dhabi N. A. Hypoglycemic activity of 6-bromoembelin and vilangin in high-fat diet fed-streptozotocin-induced type 2 diabetic rats and molecular docking studies. Life Sciences . 2016;153:100–117. doi: 10.1016/j.lfs.2016.04.016. [DOI] [PubMed] [Google Scholar]
- 39.Rotimi O. A., Rotimi S. O., Oluwafemi F., Ademuyiwa O., Balogun E. A. Oxidative stress in extrahepatic tissues of rats co-exposed to aflatoxin B1 and low protein diet. Toxicological Research . 2018;34(3):211–220. doi: 10.5487/tr.2018.34.3.211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rotimi S. O., Bankole G. E., Adelani I. B., Rotimi O. A. Hesperidin prevents lipopolysaccharide-induced endotoxicity in rats. Immunopharmacology and Immunotoxicology . 2016;38(5):364–371. doi: 10.1080/08923973.2016.1214142. [DOI] [PubMed] [Google Scholar]
- 41.Marklund S., Marklund G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. European Journal of Biochemistry . 1974;47(3):469–474. doi: 10.1111/j.1432-1033.1974.tb03714.x. [DOI] [PubMed] [Google Scholar]
- 42.Habig W. H., Pabst M. J., Jakoby W. B. Glutathione S-transferases. Journal of Biological Chemistry . 1974;249(22):7130–7139. doi: 10.1016/s0021-9258(19)42083-8. [DOI] [PubMed] [Google Scholar]
- 43.Sedlak J., Lindsay R. H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Analytical Biochemistry . 1968;25:1192–1205. doi: 10.1016/0003-2697(68)90092-4. [DOI] [PubMed] [Google Scholar]
- 44.Chance B., Maehly A. C. Assay of catalase and peroxidase. Methods in Enzymology . 1955;2:773–775. [Google Scholar]
- 45.Buege J. A., Aust S. D. [30] Microsomal lipid peroxidation. Methods in Enzymology . 1978;52:302–310. doi: 10.1016/s0076-6879(78)52032-6. [DOI] [PubMed] [Google Scholar]
- 46.Lowry O., Rosebrough N., Farr A. L., Randall R. Protein measurement with the folin phenol reagent. Journal of Biological Chemistry . 1951;193(1):265–275. doi: 10.1016/s0021-9258(19)52451-6. [DOI] [PubMed] [Google Scholar]
- 47.Iheagwam F. N., Ogunlana O. O., Chinedu S. N. Model optimization and in silico analysis of potential dipeptidyl peptidase iv antagonists from GC-MS identified compounds in Nauclea latifolia leaf extracts. International Journal of Molecular Sciences . 2019;20(23):p. 5913. doi: 10.3390/ijms20235913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Filimonov D. A., Lagunin A. A., Gloriozova T. A., et al. Prediction of the biological activity spectra of organic compounds using the PASS online web resource. Chemistry of Heterocyclic Compounds . 2014;50(3):444–457. doi: 10.1007/s10593-014-1496-1. [DOI] [Google Scholar]
- 49.Daina A., Michielin O., Zoete V. SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Scientific Reports . 2017;7(1) doi: 10.1038/srep42717.42717 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Schyman P., Liu R., Desai V., Wallqvist A. vNN web server for ADMET predictions. Frontiers in Pharmacology . 2017;8:p. 889. doi: 10.3389/fphar.2017.00889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Saeedan A. S., Soliman G. A., Abdel-Rahman R. F., et al. Possible synergistic antidiabetic effects of quantified artemisia judaica extract and glyburide in streptozotocin-induced diabetic rats via restoration of PPAR-α mRNA expression. Biology . 2021;10(8):p. 796. doi: 10.3390/biology10080796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Reuben Okoduwa S., Umar I., James D., Inuwa H. Validation of the antidiabetic effects of Vernonia amygdalina delile leaf fractions in fortified diet-fed streptozotocin-treated rat model of type-2 diabetes. Journal of Diabetology . 2017;8(3):74–85. doi: 10.4103/jod.jod_19_17. [DOI] [Google Scholar]
- 53.Kumar V., Sachan R., Rahman M., Sharma K., Al-Abbasi F. A., Anwar F. Prunus amygdalus extract exert antidiabetic effect via inhibition of DPP-IV: in-silico and in-vivo approaches. Journal of Biomolecular Structure and Dynamics . 2021;39(11):4160–4174. doi: 10.1080/07391102.2020.1775124. [DOI] [PubMed] [Google Scholar]
- 54.Sakuludomkan W., Yeewa R., Subhawa S., Khanaree C., Bonness A. I., Chewonarin T. Effects of fermented houttuynia cordata thunb. On diabetic rats induced by a high-fat diet with streptozotocin and on insulin resistance in 3T3-L1 adipocytes. Journal of Nutrition and Metabolism . 2021;2021 doi: 10.1155/2021/6936025.6936025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Asmat U., Abad K., Ismail K. Diabetes mellitus and oxidative stress-A concise review. Saudi Pharmaceutical Journal . 2016;24(5):547–553. doi: 10.1016/j.jsps.2015.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ighodaro O. M. Molecular pathways associated with oxidative stress in diabetes mellitus. Biomedicine and Pharmacotherapy . 2018;108:656–662. doi: 10.1016/j.biopha.2018.09.058. [DOI] [PubMed] [Google Scholar]
- 57.Giacco F., Brownlee M. Oxidative stress and diabetic complications. Circulation Research . 2010;107(9):1058–1070. doi: 10.1161/circresaha.110.223545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Bilanda D. C., Bidingha R. à. G., Djomeni Dzeufiet P. D., et al. Antihypertensive and antidiabetic activities of Erythrina senegalensis DC (Fabaceae) stem bark aqueous extract on diabetic hypertensive rats. Journal of Ethnopharmacology . 2020;246 doi: 10.1016/j.jep.2019.112200.112200 [DOI] [PubMed] [Google Scholar]
- 59.Al-Trad B., Alkhateeb H., Alsmadi W., Al-Zoubi M. Eugenol ameliorates insulin resistance, oxidative stress and inflammation in high fat-diet/streptozotocin-induced diabetic rat. Life Sciences . 2019;216:183–188. doi: 10.1016/j.lfs.2018.11.034. [DOI] [PubMed] [Google Scholar]
- 60.Ogar I., Egbung G. E., Nna V. U., Atangwho I. J., Itam E. H. Hyptis verticillata attenuates dyslipidaemia, oxidative stress and hepato-renal damage in streptozotocin-induced diabetic rats. Life Sciences . 2019;219:283–293. doi: 10.1016/j.lfs.2019.01.027. [DOI] [PubMed] [Google Scholar]
- 61.Seven A., Güzel S., Seymen O., et al. Effects of vitamin E supplementation on oxidative stress in streptozotocin induced diabetic rats: investigation of liver and plasma. Yonsei Medical Journal . 2004;45(4):703–710. doi: 10.3349/ymj.2004.45.4.703. [DOI] [PubMed] [Google Scholar]
- 62.Maurya P. K., Kumar P., Chandra P. Biomarkers of oxidative stress in erythrocytes as a function of human age. World Journal of Methodology . 2015;5(4):216–222. doi: 10.5662/wjm.v5.i4.216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Carelli-Alinovi C., Misiti F. Erythrocytes as potential link between diabetes and alzheimer’s disease. Frontiers in Aging Neuroscience . 2017;9:p. 276. doi: 10.3389/fnagi.2017.00276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hamden K., Carreau S., Jamoussi K., et al. 1α,25 dihydroxyvitamin D3: therapeutic and preventive effects against oxidative stress, hepatic, pancreatic and renal injury in alloxan-induced diabetes in rats. Journal of Nutritional Science & Vitaminology . 2009;55(3):215–222. doi: 10.3177/jnsv.55.215. [DOI] [PubMed] [Google Scholar]
- 65.Sagoo M. K., Gnudi L. Diabetic nephropathy: is there a role for oxidative stress? Free Radical Biology and Medicine . 2018;116:50–63. doi: 10.1016/j.freeradbiomed.2017.12.040. [DOI] [PubMed] [Google Scholar]
- 66.Wan Hasan W. N., Kwak M.-K., Makpol S., Ngah W. Z. W., Yusof Y. A. M. Piper betle induces phase I & II genes through Nrf2/ARE signaling pathway in mouse embryonic fibroblasts derived from wild type and Nrf2 knockout cells. BMC Complementary and Alternative Medicine . 2014;14(1):p. 72. doi: 10.1186/1472-6882-14-72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Uruno A., Furusawa Y., Yagishita Y., et al. The Keap1-Nrf2 system prevents onset of diabetes mellitus. Molecular and Cellular Biology . 2013;33(15):2996–3010. doi: 10.1128/mcb.00225-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Uruno A., Yagishita Y., Yamamoto M. The Keap1-Nrf2 system and diabetes mellitus. Archives of Biochemistry and Biophysics . 2015;566:76–84. doi: 10.1016/j.abb.2014.12.012. [DOI] [PubMed] [Google Scholar]
- 69.Ding X., Jian T., Wu Y., et al. Ellagic acid ameliorates oxidative stress and insulin resistance in high glucose-treated HepG2 cells via miR-223/keap1-Nrf2 pathway. Biomedicine and Pharmacotherapy . 2019;110:85–94. doi: 10.1016/j.biopha.2018.11.018. [DOI] [PubMed] [Google Scholar]
- 70.Ni Z., Guo L., Liu F., Olatunji O. J., Yin M. Allium tuberosum alleviates diabetic nephropathy by supressing hyperglycemia-induced oxidative stress and inflammation in high fat diet/streptozotocin treated rats. Biomedicine and Pharmacotherapy . 2019;112 doi: 10.1016/j.biopha.2019.108678.108678 [DOI] [PubMed] [Google Scholar]
- 71.Donath M. Y., Dalmas É., Sauter N. S., Böni-Schnetzler M. Inflammation in obesity and diabetes: islet dysfunction and therapeutic opportunity. Cell Metabolism . 2013;17(6):860–872. doi: 10.1016/j.cmet.2013.05.001. [DOI] [PubMed] [Google Scholar]
- 72.Bastard J. P., Maachi M., Lagathu C., et al. Recent advances in the relationship between obesity, inflammation, and insulin resistance. European Cytokine Network . 2006;17(1):4–12. [PubMed] [Google Scholar]
- 73.Weigert C., Hennige A. M., Lehmann R., et al. Direct cross-talk of interleukin-6 and insulin signal transduction via insulin receptor substrate-1 in skeletal muscle cells. Journal of Biological Chemistry . 2006;281(11):7060–7067. doi: 10.1074/jbc.m509782200. [DOI] [PubMed] [Google Scholar]
- 74.Sosale B., Chandrashekara S., Aravind S. R., Renuka P., Anupama K. R. Influence of cytokine status on insulin resistance and circulating endothelial progenitor cells in type 2 diabetes mellitus. Cytokine . 2017;99:179–185. doi: 10.1016/j.cyto.2017.09.012. [DOI] [PubMed] [Google Scholar]
- 75.Fan X., Zhang C., Niu S., et al. Ginsenoside Rg1 attenuates hepatic insulin resistance induced by high-fat and high-sugar by inhibiting inflammation. European Journal of Pharmacology . 2019;854:247–255. doi: 10.1016/j.ejphar.2019.04.027. [DOI] [PubMed] [Google Scholar]
- 76.Nandini H. S., Naik P. R. Antidiabetic, antihyperlipidemic and antioxidant effect of vincamine, in streptozotocin-induced diabetic rats. European Journal of Pharmacology . 2019;843:233–239. doi: 10.1016/j.ejphar.2018.11.034. [DOI] [PubMed] [Google Scholar]
- 77.Li L., Li S.-h., Jiang J.-p., Liu C., Ji L.-l. Investigating pharmacological mechanisms of andrographolide on non-alcoholic steatohepatitis (NASH): a bioinformatics approach of network pharmacology. Chinese Herbal Medicines . 2021;13(3):342–350. doi: 10.1016/j.chmed.2021.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Mishra P., Paital B., Jena S., et al. Possible activation of NRF2 by Vitamin E/Curcumin against altered thyroid hormone induced oxidative stress via NFĸB/AKT/mTOR/KEAP1 signalling in rat heart. Scientific Reports . 2019;9(1):7408–7416. doi: 10.1038/s41598-019-43320-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Lee S. R., An E. J., Kim J., Bae Y. S. Function of NADPH oxidases in diabetic nephropathy and development of Nox inhibitors. Biomolecules & Therapeutics . 2020;28(1):25–33. doi: 10.4062/biomolther.2019.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Song J., Tang Z., Li H., et al. Role of JAK2 in the pathogenesis of diabetic erectile dysfunction and an intervention with Berberine. The Journal of Sexual Medicine . 2019;16(11):1708–1720. doi: 10.1016/j.jsxm.2019.08.014. [DOI] [PubMed] [Google Scholar]
- 81.Wang L., Fan J., Yan C.-y., Ling R., Yun J. Activation of hypoxia-inducible factor-1α by prolonged in vivo hyperinsulinemia treatment potentiates cancerous progression in estrogen receptor-positive breast cancer cells. Biochemical and Biophysical Research Communications . 2017;491(2):545–551. doi: 10.1016/j.bbrc.2017.03.128. [DOI] [PubMed] [Google Scholar]
- 82.Wang X., Gu C., He W., et al. Glucose oxidase induces insulin resistance via influencing multiple targets in vitro and in vivo: the central role of oxidative stress. Biochimie . 2012;94(8):1705–1717. doi: 10.1016/j.biochi.2012.03.024. [DOI] [PubMed] [Google Scholar]
- 83.Ge T., Yang J., Zhou S., Wang Y., Li Y., Tong X. The role of the pentose phosphate pathway in diabetes and cancer. Frontiers in Endocrinology . 2020;11:p. 365. doi: 10.3389/fendo.2020.00365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Upadhyay G., Gupta S. P., Prakash O., Singh M. P. Pyrogallol-mediated toxicity and natural antioxidants: triumphs and pitfalls of preclinical findings and their translational limitations. Chemico-Biological Interactions . 2010;183(3):333–340. doi: 10.1016/j.cbi.2009.11.028. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1: grid map specifications for molecular docking. Figure S1: representation of (a) modelled Nrf-2 structure and (b) corresponding Ramachandran plot. Figure S2: 2D representation of (a) 9-Oxabicyclo[3.3.1]nonane-2,6-diol, (b) 1,2,3-Benzenetriol, (c) glibenclamide, and (d) metformin interactions in the binding pocket of Nrf-2. Green, light blue, pink, orange, violet, and broken red lines represent conventional hydrogen, carbon-hydrogen, pi-alkyl, salt bridge, pi-sigma, and unfavourable bonds, respectively. Light green circles represent Van der Waals interactions. Figure S3: 2D representation of (a) 9-Oxabicyclo[3.3.1]nonane-2,6-diol, (b) 1,2,3-Benzenetriol, (c) glibenclamide, and (d) metformin interactions in the binding pocket of IL-6. Green, light blue, pink, orange, and violet broken lines represent conventional hydrogen, carbon-hydrogen, pi-alkyl, attractive, and pi-sigma bonds, respectively. Light green circles represent Van der Waals interactions. Figure S4: 2D representation of (a) 9-Oxabicyclo[3.3.1]nonane-2,6-diol, (b) 1,2,3-Benzenetriol, (c) glibenclamide, and (d) metformin interactions in the binding pocket of TNF-α. Green, light blue, pink, violet, and broken red lines represent conventional hydrogen, carbon-hydrogen, pi-alkyl, pi-sigma, and unfavourable bonds, respectively. Light green circles represent Van der Waals interactions.
Data Availability Statement
All the data are included in the manuscript and supplementary text.