

ABSTRACT
Research on the gut–brain axis has accelerated substantially over the course of the last years. Many reviews have outlined the important implications of understanding the relation of the gut microbiota with human brain function and behavior. One substantial drawback in integrating gut microbiome and brain data is the lack of integrative multivariate approaches that enable capturing variance in both modalities simultaneously. To address this issue, we applied a linked independent component analysis (LICA) to microbiota and brain connectivity data.We analyzed data from 58 healthy females (mean age = 21.5 years). Magnetic Resonance Imaging data were acquired using resting state functional imaging data. The assessment of gut microbial composition from feces was based on sequencing of the V4 16S rRNA gene region. We used the LICA model to simultaneously factorize the subjects’ large-scale brain networks and microbiome relative abundance data into 10 independent components of spatial and abundance variation.LICA decomposition resulted in four components with non-marginal contribution of the microbiota data. The default mode network featured strongly in three components, whereas the two-lateralized fronto-parietal attention networks contributed to one component. The executive-control (with the default mode) network was associated to another component. We found that the abundance of Prevotella genus was associated with the strength of expression of all networks, whereas Bifidobacterium was associated with the default mode and frontoparietal-attention networks.We provide the first exploratory evidence for multivariate associative patterns between the gut microbiota and brain network connectivity in healthy humans considering the complexity of both systems.
KEYWORDS: Linked ICA, gut microbiota, brain connectivity networks, fMRI, resting state, Bifidobacterium, Prevotella
Introduction
The gut–brain axis (GBA) is a bidirectional biochemical signaling that takes place between the gastrointestinal tract (GI tract) and the central nervous system (CNS).1 The microbiota–GBA is used to describe the complex effects of the commensal gut bacteria (the microbiota) in the interplay between the gut and the brain. Recently, many studies have outlined the important implications of understanding the relation of the gut microbiota with human brain function and behavior. Several intermediary pathways have been proposed, specifically, bi-directional interactions between microbiota and the brain are plausible via modulation of vagal nerve activity, via neuromodulators or their precursors such as serotonin or tryptophan, via the Hypothalamic–Pituitary–Adrenal System (HPA-axis) and via interactions with the immune system.1–4
In recent years, researchers aimed at elucidating these interactions, highlighting putative pathways, hormonal or immunological agents, and targeting the activity and interaction of certain bacterial strains.3 However, these studies have not taken into account the complexity and, especially, the full multivariate nature of both the brain and the gut microbiome.
One of these complex traits of the brain is the intrinsic connectivity between different brain regions. So far, studies assessing the relation between gut microbiome composition and intrinsic brain connectivity – with resting state fMRI – are rare, limited in rigor, and inconclusive.5 A recent study tested the effects of four weeks multi-strain probiotics supplementation.6 The authors report mild probiotics-induced changes in resting state connectivity of some of the 10 networks tested. The strongest modulation was found in differences between the placebo (n = 15) and probiotics (n = 15) group, with the latter showing a relatively stronger increase in connectivity of the salience network to superior frontal brain regions. In another placebo-controlled trial of probiotics (n = 20 per group), Bifidobacterium longum influenced resting neural oscillations measured with magnetoencephalography (MEG), which correlated with enhanced vitality and reduced mental fatigue during a social stress induction task. Modulations of theta and alpha band oscillations by probiotics were localized in the frontal and cingulate cortex and supramarginal gyrus.7 However, these results (in relatively small samples) have not been related to probiotics-induced effects on gut microbiota composition.
A few studies did assess the relation between gut microbiome composition and intrinsic brain connectivity. One resting state fMRI study (n = 30), which included a subgroup of smokers, focused on the association of gut microbiota composition with insula connectivity and found its connection to several brain regions, such as occipital and lingual gyrus, frontal pole and cerebellar regions, to be associated with microbiota diversity and structure.8 Other exploratory region-of-interest (ROI) analyses did not reveal significant associations. Another resting-state fMRI study (n = 28 vs 19) demonstrated that in end-stage renal disease, the integrity of the default mode network (DMN) was decreased along with alterations in the gut–microbiota composition.9 A recent study (n = 157) focused on certain enteroptypes and diversity measures in the gut and their association to specific large-scale brain networks.10 The authors found an association between gut microbial diversity measures with network connectivity of executive control, default mode and sensorimotor control, as well as within executive control network. Enterotypes were linked to executive control network. Another very recent study investigated MRI measures of cortical thickness, regional homogeneity (ReHo) and fractional amplitude of low-frequency fluctuation (fALFF) in relation to relative abundance in a schizophrenia group comparison.11 The ‘functional’ measures (ReHo & fALFF) used in this study should be interpreted with caution, as they are potentially strongly biased by non-neuronal sources (cp.12). They found differences in relative abundance of Ruminococcus and Roseburia genera and associated differences in brain measures, which might be associated with neuropathology of schizophrenia.
Most importantly, previous studies on healthy individuals have exclusively performed bivariate associations (albeit controlled for multiple comparisons or partial effects) between one gut–microbiome composition measure and one brain connectivity measure (i.e. within one network or between two brain networks). Taken together, these results are difficult to integrate and comprehend, as studies focus on one aspect of the modalities, such as connectivity from one particular ROI or with different types of interventions. Furthermore, investigation of the gut–brain axis in targeted patient groups9–11,13 is of great merit, especially for the relevance of the gut microbiota in the context of a particular disease, but as long as the health functionality of the gut–brain axis is not sufficiently understood on a macroscopic level, assessing the consequences of certain pathologies is difficult. We here argue for a multivariate integration of healthy adult individuals as a starting point.
Indeed, in research, one approach to understand complex systems is to try to elucidate the function of all its components sequentially and then to integrate interactions between a limited number of components. The opposite approach of investigation is to aim at integration at a macroscopic level. In this approach as many components as possible are sampled and patterns are investigated by dimensionality reduction. This has been attempted for the gut–brain axis very often narratively, in multiple reviews. Yet, no empirical attempt has been made so far to try to integrate the functions of the brain and the gut microbiome at a macroscopic level to determine associations between variance in macroscopic components of the two systems. In a similar approach, using data from the Human Connectome, researchers linked several lifestyle, demographic and psychometric measures in a positive–negative mode of brain connectivity.14
Here, we aim to assess the relation between these two complex, multivariate modalities, focusing on canonically established brain networks in resting state that represent major modes of brain functioning in an unperturbed fashion, i.e. in healthy individuals.15 We asked if the inter-individual variability in the abundance of gut microbiome genera was linked to variability in brain functional connectivity in canonical brain networks, when taking into account the full complexity of both. In doing this, we hope to (1) validate past research that has demonstrated associations between brain connectivity and gut microbiota, be it via probiotica intervention or by focusing on select networks/regions and bacterial strains, (2) provide a targeted explorative ‘map’ of potential candidate links between sets of associated gut microbiota and brain connectivity that have this far not be explored and (3) provide a similar guidance to studies investigating clinical perturbations in brain connectivity – gut microbiota associations as well as an elegant way to investigate complex intervention-induced changes in the gut–brain axis.
One substantial methodological challenge is the multivariate and simultaneous integration of gut microbiome and brain data that enables capturing variance in both modalities simultaneously. To address this issue, we applied a linked independent component analysis (LICA)16,17 to microbiota and brain connectivity data (Figure 1). LICA enables data reduction in several modalities simultaneously and thereby can demonstrate joint inter-individual variation patterns in different modalities. We chose to investigate four very well characterized and often replicated brain networks.15 In the previous work, we used this selection to investigate the impact of fasting on functional connectivity at rest.18 We limited our study to a set of four networks of interest (the lateralized fronto-parietal (left/right) attention networks, FPN; the executive control network, ECN; and the default mode network, DMN) due to their importance in the neuroimaging field, their comparatively clear and cognitive functional profile and their importance in mental disease or previous microbiome research.14,15,18–20 This pre-selection enables us to better conceptually understand the explorative associations between gut and these brain networks, limiting the number but not the type of interactions.
Figure 1.
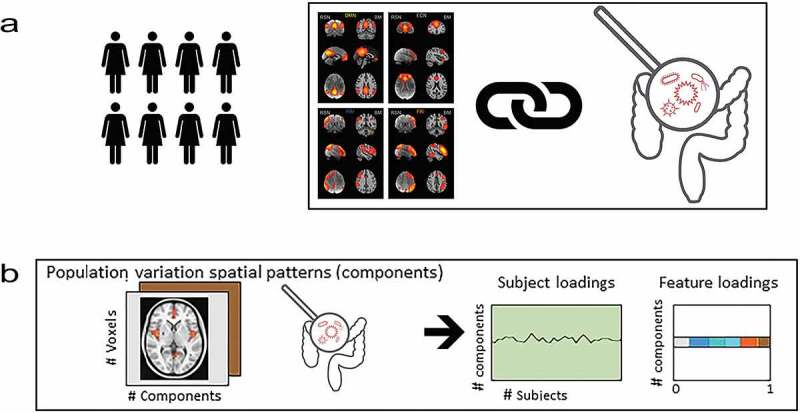
We linked functional brain connectivity in four well-established brain networks with relative abundance of human gut bacteria (microbiota). Panel A describes the Linked ICA that decomposed, simultaneously, the variability in functional connectivity of the four networks and the relative abundance of bacterial taxa (genera). This resulted in 10 components for which we have individual subject loadings as well as the loadings of each input feature depicted in panel B. The loadings represent voxel-wise association to the component in functional connectivity per network and genera-wise association to the component in the gut–microbiome
Results
For 58 subjects, the spatial template maps of right and left frontoparietal-attention networks (FPN), executive control (ECN), default mode network (DMN)15 were projected onto the subjects resting state fMRI time-courses to create network maps per subject. The ECN largely consists of middle frontal and superior frontal gyri, paracingulate cortex and dorsal posterior parietal cortex.15,21 The ECN has been demonstrated to overlap spatially with brain activity observed in cognitive control tasks, emotion tasks and response inhibition.15 The DMN is the large-scale brain network that was identified first and it is probably the most often studied of all so-called resting-state networks. DMN modulations have been implicated in a broad range of disorders.22–25 The most prominent feature of the DMN is its task-negative nature; the areas of the DMN deactivate when an individual is engaged in most tasks.26–28 It has been associated to a broad range of cognitive processes such as self-referenced thought and self-monitoring,26 passive, broad attention,23,29 auto-biographical memory retrieval,30,31 imparting meaning to the current sensory input depending on prior experiences,32 mind-wandering and future thinking32,33 as well as homeostatic functions.26,27,34,35 As the name of these networks suggest, the fronto-parietal-attention networks encompass fronto-parietal brain regions, which are commonly and reliably associated with brain activity in attention tasks15 and are modulated with varying degree of attention demand.36,37
Gut microbiome composition was based on sequencing of the V4 region of the 16S rRNA gene on the Illumina HiSeq platform. We used the LICA model to simultaneously factorize the subjects’ brain networks and gut microbiome relative abundance into 10 independent components.16,17
Joint decomposition of brain networks and microbiome relative abundance
From the 10 components, six showed a non-marginal (proportion >0.2) contribution on both the gut microbiota relative abundance and the brain connectivity patterns (Component 0, 1, 3, 4, 6 and 7ö Figure 2). From these six components, the first extracted component (component 0) was explained by a single subject; therefore, this component was disregarded for further analyses. Additionally, sanity checks on brain connectivity showed that, for component 4, equal values for all voxels in the brain data. This renders the interpretation of this component hardly possible and could potentially be related to residual noise being picked up and explained. This component was therefore also discarded.
Figure 2.
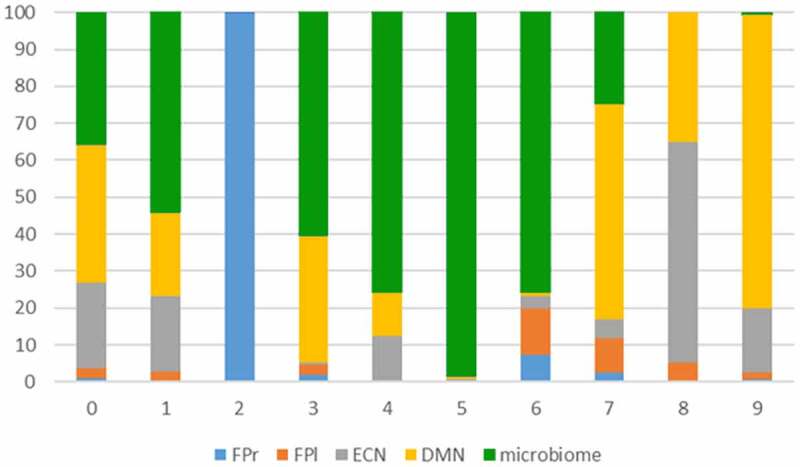
Decomposition of brain connectivity and microbiome. The plot shows the percentage of contribution per input modality. FPr and FPl are right and left fronto-parietal networks, DMN is default mode and ECN is executive control network
We subsequently investigated the association between brain connectivity and gut microbiota relative abundance in the four remaining components. We characterized each component by the contribution of the different modalities (proportion >0.2 for brain or microbiome). For each component, we plotted the brain-network and their voxel-wise loading and listed the bacterial genera that were non-marginally associated with the component. For brain connectivity data, the z-maps from the LICA were thresholded at a z > 3 for display purposes (see Neurovault: https://neurovault.org/collections/TRVFBPAB/ for z maps of the brain data four components). The z-scores reflect how strongly a voxel covaries in connectivity with the respective input network. For microbiome data, high loadings reflect a robust covariation in relative abundance of a particular genus in that particular component. Similarly, we thresholded the microbiome loading at z > 2.3 as they were more sparse compared to brain loadings (see Figure 3 for a visualization of these results).
Figure 3.
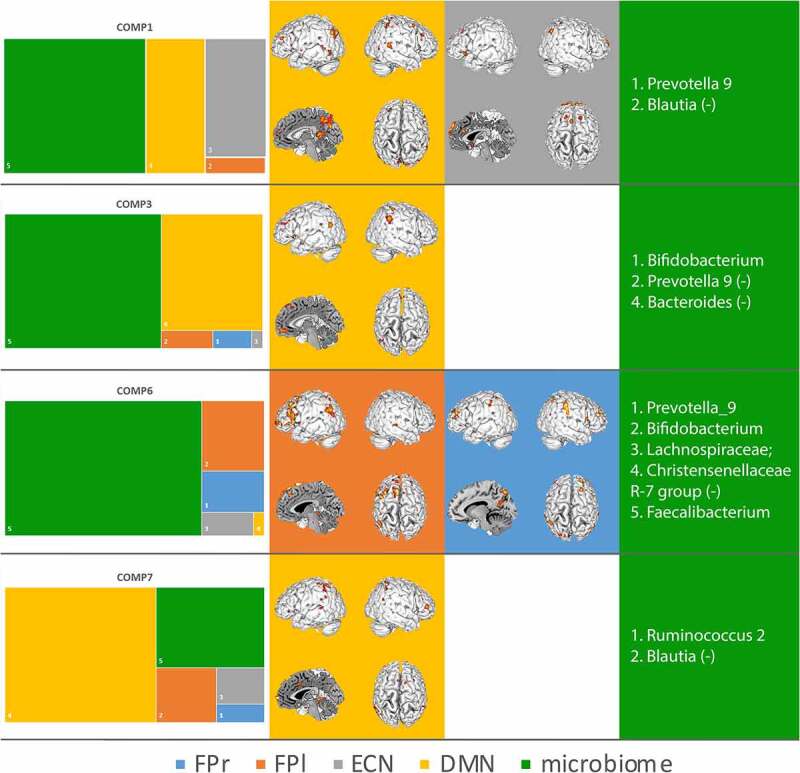
Summary results of the contribution of each modality are shown in the first column (left). Second and third columns display the spatial project of the brain modalities, e.g. which voxels covary most strongly with covariation in other modalities (brain networks and microbiota abundance). Fourth column (right) displays the genera that show a covariance in abundance that is linked to covariance in the brain networks. The colors align with the modalities of the LICA (the four brain networks and the gut microbiota). For display purposes genera loading were cut at z > 2.3 (for more details see the Method section)
The microbiota accounted for the majority of the variability that could be explained by component 1, 3 and 6. The DMN explained the majority of variability in component 7, yet the microbiota is contributing by more than .2. We will discuss components 1, 3, 6 and 7 in chronological order. On the brain connectivity side, LICA enables us to say which voxels show variation in functional connectivity to the respective network between subjects, which could be understood as an inter-individual variation in the strength of network connectivity. For the microbiota, LICA gives an indication of between-subject variation in the relative abundance of the gut microbiota genera that load on that component.
The first component (#1) has strong contributions from variability in the DMN and the ECN and the microbiota. The ECN in component 1 varied between subjects in core hubs of the ECN, such as the dorsal paracingulate cortex, middle frontal, and superior frontal gyri. This covariance in core hubs of the ECN can be interpreted as this component explaining the functional connectivity strength, or the strength of the expression of the ECN in the subjects, and this strength of expression being related to variation in the relative abundance of gut microbiota. Similarly, for the DMN, component 1 picked up on variability in the posterior core hub of the DMN (the posterior cingulate and retrosplenial cortex; 13, 16). Prevotella_9 was more abundant and Blautia was less abundant with increasing between-subject functional connectivity of these hubs of the two networks. We therefore in the discussion call it ECN-DMN-Prevotella+-Blautia− component.
The next component (#2) had a contribution of over 50% from variability in the microbiota abundance and the DMN. Bifidobacterium was more abundant and Prevotella_9 and Bacteroides were less abundant with increasing functional connectivity in the anterior core hubs of the DMN. We therefore in the discussion call it the DMN-Bifido+-Prevotella− component
Component #6 had the strongest contribution of all components from microbiota of around 75%, yet also explained variability in the two-lateralized fronto-parietal attention networks. The topology of the loading of these networks on this component overlaps with their common, canonical spatial profile in lateral frontal and parietal brain areas. Thus, again this component is associated with the between-subjects variation of the strength of expression of the lateralized attention networks. Prevotella_9, Bifidobacterium, genera belonging to Lachnospiracaceae family, and Faecalibacterium were more abundant and Christensenellacea_R-7_group was less abundant with stronger expression of the attention networks. We therefore in the discussion call it the FPN-Bifido-Prevotella component.
Component #7 was associated with variability in the DMN again and to roughly 25% of the microbiota. The spatial pattern of between-subject variation could be interpreted as elevated connectivity of the DMN to parts of the so-called salience network (such as dorsal anterior cingulate cortex (dorsal ACC) and ventrolateral prefrontal cortex (VLPFC)), which has in the previous literature been associated with effects of elevated stress on DMN resting state connectivity.19,20 Ruminocuccus_2 was more abundant and Blautia was less abundant in individuals that showed this elevated connectivity pattern of DMN to dorsal ACC and VPLFC. We therefore in the discussion call it the DMN-Ruminocuccus+-Blautia− component.
Discussion
In this study, we provide the first evidence for multivariate associative patterns between the gut microbiota and brain network connectivity in healthy humans. We used a novel multivariate modality integration technique to explain inter-individual differences in brain connectivity in four canonical networks and the gut microbiota. We see our exploratory results as a map that could show high potential to guide future research on the relation of gut–brain interactions in a hypothesis-generating manner. We have linked ECN connectivity to an abundance of Prevotella_9 and Blautia; DMN connectivity to Prevotella_9, Blautia, Ruminococcus_2, Bifidobacterium, and Bacteroides; fronto-parietal attention network connectivity to Prevotella_9, Bifidobacterium, Faecalibacterium, Christensenellacea_R-7_group, and certain genera belonging to Lachnospiracaceae. DMN connectivity that has been linked to stress is associated with Ruminocuccus_2 and Blautia. The spatial associations in the components to core hubs of the respective networks can be seen as a conceptual validation of our approach.15,20,38 Furthermore, we observe the between-subject variation in functional connectivity in core hubs of the respective networks in three of the four components as a link between an individual’s connectivity strength and the relative abundance of certain microbiota. These findings can be taken as an indication that certain microbial genera are associated with the normal expression of all four canonical resting state networks and their natural variation between healthy subjects.
On the side of the bacterial genera that were associated with brain network connectivity, we found that inter-individual variation in abundance of the Bifidobacterium genus was prominently contributing to two of our four identified components. Variation in abundance of Bifidobacteria were associated with increased connectivity of the medial prefrontal cortex of the DMN and parietal regions in the ECN-DMN-Prevotella+-Blautia− and in the FPN-Bifido-Prevotella with modulated connectivity of the core hubs of the fronto-parietal attention network. The Bifidobacterium genus is probably one of the most noticeable targets in current gut–brain axis research.2,39,40 This strong focus is potentially related to a landmark study, which showed that germ-free mice have altered HPA-axis function, and this altered HPA activity was reversed by colonization with a Bifidobaterium.1 Bifidobacteria are one of the most important and abundant genera during development and have been associated with decreased levels of inflammation in human development.41 Bifidobacterium longum, a strain commonly used in probiotic products, influenced resting neural activity that correlated with enhanced vitality and reduced mental fatigue during a social stress induction task.7 The medial prefrontal cortex and the DMN have been related to autobiographic and episodic memory or prior knowledge structures,42 which fits to findings of the link between increased Bifidobacteria after interventions and elevated verbal episodic memory.6 Furthermore, Bagga and colleagues also found altered functional connectivity of the DMN after probiotic use (including B. longum).6 Component 3 might therefor partially reflect episodic memory-related modulations in Bifidobacteria and DMN connectivity. A probiotics trial with Bidfidobacterium longum using electrophysiological resting state brain recordings found evidence for an association of increased frontal midline mobility and improved memory after probiotics consumption compared to placebo.43 The authors related the brain recordings to attention-related brain activity. Moreover, the fact that out of >50 genera that featured in our analysis, Bifidobacteria featured in two of the four components both underscores their putative influence in the gut–brain interaction and the validity of our integrative approach. In summary, we found evidence for a relation of Bifidobacteria abundance to attention- and potentially memory-related brain network activity at rest.
For the DMN-Ruminocuccus+-Blautia− component, the spatial patterns of association were similar to results showing an alerted state of the DMN after social stress induction.19 In data using a similar paradigm, Bifidobacterium longum modulated activity in similar regions that were influenced by social stress and also in the hippocampus, a region that is part of the DMN.7 This pattern particularly varied with abundance in Blautia and Ruminococcus 2. Although Bifidobacterium did not covary with this component, the Bifidobacterium intake might have indirectly affected DMN connectivity in stress, potentially via modulation of the abundance of Ruminococcus 2 and Blautia. Indeed, the preexisting levels of Blautia and Ruminococcae correlated with the metabolic outcomes of a Bifidobacterium-targeting prebiotic intervention in obese patients.44 Furthermore, Blautia has been found to be the only genus to be enriched in depression-model rats45 and both Blautia and Ruminococcae correlated with stress-related depression-like behavior in mice.46 In summary, variations in Blautia and Ruminococcus 2 abundance might relate to stress-induced modulation of DMN connectivity.
The association of all brain networks in three different components with Prevotella_9 is interesting as this genus has previously been involved in psychiatric disorders, cognition, and brain connectivity changes. For example, in autism spectrum disorder (ASD), which is characterized by atypical brain network organization (including DMN and ECN, as in component 1),47 a higher relative abundance of Prevotella (and Bifidobacterium) has been linked with a beneficial effect of Microbiota Transfer Therapy.48 Accordingly, lower relative abundance of Prevotella has been associated with psychiatric disorders like ADHD in children,49 Parkinson’s disease,50 and ASD.51 Furthermore, the gut–brain axis may play a role in the disturbed executive functioning of ASD (for a review, see13). Our findings of a positive correlation of Prevotella with DMN and ECN functioning and also fronto-parietal attention network modulations support these results. Prevotella dominated enterotype has been demonstrated to be associated with an elevated functional intra-network connectivity of the ECN.10 Furthermore, this connectivity mediated reaction times in an executive control task, further supporting the functional profile of the ECN-DMN-Prevotella+-Blautia− and FPN-Bifido-Prevotella to executive control and attention. Our study may add more facets to this picture by the division between attention related to Bifidobacterium and Prevotella covariance patterns and executive control with DMN association and lower abundance of Blautia. Previous work showed a link between gut microbiota and resting-state functional connectivity, as assessed here.9,52 Interestingly, in one study assessing bivariate relationships, Prevotella and Bacteroides were associated with insular connectivity.8 The insula has not only been discussed as part of the salience network, but also as an important component of the general task positive network.53,54 In our case, both of the Prevotella and Bacteroides genera were negatively associated with DMN in component 3. As the DMN is thought to be anti-correlated with the task positive network, our finding is in line with previous results.53 Prevotella seem to be associated to healthy modulation in brain connectivity related to attention, cognitive control, episodic memory and a range of other psychological functions.
Our design and approach have limitations in the interpretation of the results. First, these findings are necessarily limited to more common genera. We capped our analysis at genera that are at least detectable in 30% of our subjects. Genera with lower occurrence rates in individuals might have unequally strong leverage on the LICA. As a consequence of this methodological choice, we cannot exclude an overestimation of the loadings for the more common taxa (given the sample size) and we cannot assess the rare genera and their association with brain network connectivity.
Second, the selection of brain networks was motivated by their role in cognition specifically to high-level cognitive constructs such as attention and cognitive control and their relevance in the literature. While we perceive this selection as well motivated and we have demonstrated their sensitivity,18 it is a subjective pre-selection. We might not cover other cognitive processes and associated brain networks equally well. Nevertheless, we chose a pre-selected networks approach for several methodological reasons: (1) we wanted to use representations of brain network connectivity that have been previously well characterized and understood, both conceptually (i.e. in their psychological function) and biologically (i.e. large-scale brain networks of temporally synchronized BOLD activation). (2) A problem with using a correlation matrix as input would be that single voxels, or sets of potentially scattered voxels might link to gut microbiota relative abundance. Cognitive neuroscience is far from an understanding of those single voxel or scattered correlation patterns and understanding those would require reverse inference, which is a tremendous problem in the field55 2011). (3) While the whole brain connectivity matrix represents neuronal correlations over time, it will – despite rigorous denoising – retain spurious or noise correlations. Thus, our approach can be viewed as an additional safe-guard against spurious correlations. (4) We focus on a selected set of important networks as a certain constraint to limit the number of possible interactions/contributions, but we do not constrain the type of interactions/contributions in any way. In this way, this method is still clearly data driven.
Third, we investigated a very homogenous, healthy, and young group of only female participants. Although this naturally limits the generalizability of the results, we believe that our data still serves an orientating purpose and is therefore valuable. In replication attempts, this homogeneity and special characteristic of our sample should be considered. We would like to reiterate that we see a strong need to replicate the current results in larger and more diverse samples.
Conclusion
In summary, we provided the first evidence for multivariate associative patterns between large-scale brain network functional connectivity of four very well-established brain networks and the relative abundance of gut microbiota in a sample of healthy female individuals. This link provides a map for future research, involving the full complexity of both measures into account. For example, interventions targeting improvement in attention (for example in neurodevelopmental disorders) could investigate the influence on the bacterial genera associated to the attention networks. Moreover, it can provide a roadmap to investigate how the effect of probiotic intervention trials can modulate brain networks (and associated cognitive functions) in relation to the changes in certain genera of the gut microbiome. Furthermore, future research might investigate the mechanistic nature of our multivariate associative patterns and aim to assess the generalizability to other healthy samples as well as their potential disruption in the diseased brain.
Material and methods
Sample
We analyzed pre-intervention data from a probiotics intervention study on 64 healthy female participants (mean age = 21.5 (0.45) years).56 In total, 58 of the 64 participants were included in the analyses. Six participants were excluded from the final analyses, due to high depression scores (N = 1), missing feces samples (N = 2), and movement exceeding 4 mm between acquisitions (n = 3). For more detailed characteristic of the samples and exclusion criteria as well as the ethical declaration, please see the Material and Methods section of Papalini and colleagues.56 Briefly, participants with relevant medical history of e.g. psychiatric and/or gastrointestinal disorder were excluded. Also, use of antibiotics and diet like e.g. vegan diet were part of the exclusion criteria.
fMRI data acquisition
Participants were screened for compatibility with magnetic resonance imaging (MRI). MRI data were acquired using a 3 T MAGNETOM Prisma system, equipped with a 32-channel head coil. After three short task-related fMRI scans (see Papalini et al.), 9 min of resting state fMRI was acquired. 3D echo planar imaging (EPI) scans using a T2*weighted gradient echo multi-echo sequence (Poser, Versluis et al. 2006) were acquired (voxel size 3.5 × 3.5 × 3 mm isotropic, TR = 2070 ms, TE = 9 ms; 19.25 ms; 29.5 ms; 39.75 ms, FoV = 224 mm). The slab positioning and rotation (average angle of 14 degrees to AC axis) optimally covered both prefrontal and deep brain regions. Subjects were instructed to lie still with their eyes open and refrain from directed thought. A whole-brain high-resolution T1-weighted anatomical scan was acquired using a MPRAGE sequence (voxel size 1.0 × 1.0 × 1.0 isotropic, TR = 2300 ms, TE = 3.03 ms, 192 slices).
MRI data preprocessing: FSL (FMRIB, University of Oxford, UK; www.fmrib.ox.ac.uk/fsl;57 was used for pre-processing, data-denoising, and generation of subject-specific network maps. Pre-processing steps included three-dimensional movement correction, and spatial smoothing using a 5 mm full-width at half maximum (FWHM) Gaussian kernel to reduce inter-subject variability and a high-pass filter (> 0.007 Hz) was applied. All pre-processing steps, except temporal filtering, were conducted before AROMA data denoising.58,59 Briefly, ICA-AROMA is designed to identify motion-related artifacts by matching single subject ICA components to four robust and standardized features. The data is denoised by linear regression of ICA components identified as noise by AROMA and subsequently the high pass filter was applied. Prior to all group analyses, data were normalized to MNI space and re-sampled to 2 mm3 resolution using FMRIB’s Nonlinear Image Registration Tool (FNIRT).
Generation of subject-specific functional connectivity maps
Dual (spatial and temporal) regression was used to generate subject-specific spatial maps of well-studied, canonical large-scale brain networks15 from the individuals’ data. These canonical large-scale brain networks consist of 10 resting state networks for which the template can be freely downloaded (https://www.fmrib.ox.ac.uk/datasets/brainmap+rsns/). We downloaded the 10 well-matched resting state networks for use as templates in this study. The z-maps of these networks are temporally concatenated in one 4D file and used as input for the dual regression. These maps were used in a linear model fit against the individual fMRI data, resulting in the subject-specific temporal dynamics. Subsequently, these time-course matrices are employed in a linear model fit against the subject’s fMRI data set to estimate subject-specific spatial maps. From these subject-wise expressions of the 10 networks, we selected four networks of interest (the left and right lateralized fronto-parietal attention networks, FPN; the executive control network, ECN; and the default mode network, DMN), due to their importance in the neuroimaging field, their comparatively clear functional profile and their importance in mental disease or previous microbiome research.14,15,18–20 The different spatial maps for all participants are combined into a single 4D file per target network. In this way, we generated four files for the four respective networks of interest that contain one spatial z-map per subject that indicates for each voxel the connectivity strength of the respective network in that individual. These four network files were used as inputs to the LICA.
Gut microbiome analysis
Fecal samples were collected by using OMNIgene•GUT kit (DNAGenotek, Ottawa, CA) within 24 hours after the MRI scan.60 Collected fecal samples were transported to the laboratory and aliquoted into 1.5 mL Eppendorf tubes and stored at −80 ℃ for microbiome analysis. DNA was isolated from the fecal pellets using the Maxwell® 16 Instrument (Promega, Leiden, The Netherlands) as described previously.61 Briefly, in the 2-step PCR protocol the 16S rRNA gene V4 variable region was targeted by using 515 F (GTGYCAGCMGCCGCGGTAA) and 806 R (GGACTACNVGGGTWTCTAAT) primers, and unique barcodes were used to identify each sample. Sequencing was performed on the Illumina HiSeq PE300 platform by GATC Biotech AG (Konstanz, Germany). The sequences were processed using NG-Tax62 analysis pipeline as described previously.63 NG-Tax identified the taxonomy of the samples based on 16S sequences using three core elements: (i) barcode-primer filtering, (ii) operational taxonomic unit (OTU) picking, in which unique sequences with the relative abundance above 0.1% were clustered into OTUs based on a sequence similarity ≥98.5%, and (iii) taxonomic assignment using the SILVA reference database (version 128).64 This resulted in an Operational Taxonomical Unit (OTU) table containing 844 OTUs. We applied a prevalence-filtering at the genus level, selecting genera present in at least 30% of the samples. After this step, the OTU-table containing 644 OTUs was used for the downstream analyses. The gut microbiome composition tables at the phylum and genus taxonomic levels were provided by the ‘phyloseq’ package available in R.65 In sum, the composition table is the result of the normalization step where the read counts where transformed into relative abundance.
Linked analyses
We used the LICA model17 to simultaneous factorize the functional network maps (of ECN, FPNs and DMN) and the microbiome data of 58 subjects into independent sources (or components) of variation. In the brain networks, spatial variation was explained; while in the microbiome data, variation in relative abundance of bacterial genera was explained. In brief, LICA is an extension of Bayesian ICA66 to multiple input sets, where all individual ICA factorizations are linked through a shared common mixing matrix that reflect the subject-wise contribution to each component (Figure 1).
This operation is represented in Figure 1. Factorization provides a set of spatial maps (one per feature modality and component), a vector of feature loadings that reflects the degree to which the component ’represents’ the different modalities, and a vector that reflects the contribution of the individual subject to a given component. All mathematical derivations involved in the LICA factorization can be found in the original paper describing the original algorithm.17 Further details and code implementing each feature extraction procedure as well as the LICA factorization are publicly available at.67 Given the sample size, we forced a 10 components solution. We disregarded components estimated with marginal (proportion <0.2) contribution of the microbiome or brain networks, respectively.
Acknowledgments
We would like to thank Silvia Papalini, Franziska Michels and Joost Wegman for their support in data analysis and acquisition. We are grateful to our participants and to the excellent support staff and infrastructure at the Donders Center for Cognitive Neuroimaging in Nijmegen.
Funding Statement
The study was supported by the Dutch Ministry of Economic Affairs under the TKI Life Science and Health, project LSHM15034. CB and AL have received funding from the Innovative Medicines Initiative 2 Joint Undertaking under grant agreement No 777394 for the project AIMS-2-TRIALS. This Joint Undertaking receives support from the European Union’s Horizon 2020 research and innovation program and EFPIA and AUTISM SPEAKS, Autistica, SFARI. This work was further supported by the European Union Horizon2020 program CANDY (Grant Agreement No. 847818). EA received funding from the European Research Council (ERC_StG2019_852189). JST and AAV have received support from the European Union’s Horizon 2020 research and innovation program under Grant Agreement No. 728018 (Eat2beNICE);Autism Speaks European Federation of Pharmaceutical Industries and Associations Simons Foundation Autism Research Initiative
Disclosure statement
No potential conflict of interest was reported by the author(s).
Authors’ contributions
NK and EA conceived the study, EA and AAV conceived the project and acquired funding, JST conducted the microbiome analyses, AL and CB conceived the integratory analyses and conducted these, NK conducted processing and analysis of the fMRI data and the LICA results, AL, EA, AAV supported and supervised LICA results integration, NK and JST wrote the first draft of the manuscript, all authors contributed and agreed to the final version of the manuscript.
Ethics approval and consent to participate
The study was conducted following the Declaration of Helsinki with human subjects and the complete procedure was approved by the local Ethics Committee (CMO Arnhem-Nijmegen, NL55406.091.15) and registered at the Dutch trial register (protocol number: NTR5845). Written informed consent was obtained from each participant.
Data availability statement
All data, documentation and code will be shared via the DondersSharingCollection (https://doi.org/10.34973/3j3h-ts61).
References
- 1.Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu X-N, Kubo C, Koga Y.. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol (Lond). 2004;558:263–14. doi: 10.1113/jphysiol.2004.063388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cryan JF, Dinan TG.. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012;13:701–712. doi: 10.1038/nrn3346. [DOI] [PubMed] [Google Scholar]
- 3.Cryan JF, O’Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, Boehme M, Codagnone MG, Cussotto S, Fulling C, Golubeva AV, et al. The microbiota-gut-brain axis. Physiol Rev. 2019;99:1877–2013. doi: 10.1152/physrev.00018.2018. [DOI] [PubMed] [Google Scholar]
- 4.Fülling C, Dinan TG, Cryan JF. Gut microbe to brain signaling: what happens in vagus. Neuron. 2019;101:998–1002. doi: 10.1016/j.neuron.2019.02.008. [DOI] [PubMed] [Google Scholar]
- 5.Liu P, Peng G, Zhang N, Wang B, Luo B. Crosstalk between the gut microbiota and the brain: an update on neuroimaging findings. Front Neurol. 2019;10:883. doi: 10.3389/fneur.2019.00883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bagga D, Aigner CS, Reichert JL, Cecchetto C, Fischmeister F, Holzer P, Moissl-Eichinger C, Schöpf V. Influence of 4-week multi-strain probiotic administration on resting-state functional connectivity in healthy volunteers. Eur J Nutr. 2019;58:1821–1827. doi: 10.1007/s00394-018-1732-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang H, Braun C, Murphy EF, Enck P. Bifidobacterium longum 1714TM strain modulates brain activity of healthy volunteers during social stress. Am J Gastroenterol. 2019;114:1152–1162. doi: 10.14309/ajg.0000000000000203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Curtis K, Stewart CJ, Robinson M, Molfese DL, Gosnell SN, Kosten TR, Petrosino JF, De La Garza R, Salas R. Insular resting state functional connectivity is associated with gut microbiota diversity. Eur J Neurosci. 2019;50:2446–2452. doi: 10.1111/ejn.14305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wang YF, Zheng LJ, Liu Y, Ye YB, Luo S, Lu GM, Gong D, Zhang LJ. The gut microbiota-inflammation-brain axis in end-stage renal disease: perspectives from default mode network. Theranostics. 2019;9:8171–8181. doi: 10.7150/thno.35387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cai H, Wang C, Qian Y, Zhang S, Zhang C, Zhao W, Zhang T, Zhang B, Chen J, Liu S, et al. Large-scale functional network connectivity mediate the associations of gut microbiota with sleep quality and executive functions. Hum Brain Mapp. 2021;42:3088–3101. doi: 10.1002/hbm.25419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li S, Song J, Ke P, Kong L, Lei B, Zhou J, Huang Y, Li H, Li G, Chen J, et al. The gut microbiome is associated with brain structure and function in schizophrenia. Sci Rep. 2021;11:9743. doi: 10.1038/s41598-021-89166-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bijsterbosch J, and Beckmann C. An introduction to resting state FMRI functional connectivity. UK: Oxford University Press; 2017. [Google Scholar]
- 13.Roman P, Rueda-Ruzafa L, Cardona D, Cortes-Rodríguez A. Gut–brain axis in the executive function of austism spectrum disorder. Behav Pharmacol. 2018;29:654–663. doi: 10.1097/FBP.0000000000000428. [DOI] [PubMed] [Google Scholar]
- 14.Smith S, Nichols T, Vidaurre D, Winkler A, Behrens T, Glasser M, Ugurbil K, Barch D, Van Essen D, Miller K. A positive-negative mode of population covariation links brain connectivity, demographics and behavior. Nat Neurosci. 2015;18:1565–1567. doi: 10.1038/nn.4125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Smith SM, Fox PT, Miller KL, Glahn DC, Fox PM, Mackay CE, Filippini N, Watkins KE, Toro R, Laird AR, et al. Correspondence of the brain’s functional architecture during activation and rest. PNAS. 2009;106:13040–13045. doi: 10.1073/pnas.0905267106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Llera A, Wolfers T, Mulders P, Beckmann CF. Inter-individual differences in human brain structure and morphology link to variation in demographics and behavior. eLife. 2019;8:e44443. doi: 10.7554/eLife.44443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Groves AR, Beckmann CF, Smith SM, Woolrich MW. Linked independent component analysis for multimodal data fusion. NeuroImage. 2011;54:2198–2217. doi: 10.1016/j.neuroimage.2010.09.073. [DOI] [PubMed] [Google Scholar]
- 18.Orfanos S, Toygar T, Berthold-Losleben M, Chechko N, Durst A, Laoutidis Z, Vocke S, Weidenfeld C, Schneider F, and Karges W, et al. Investigating the impact of overnight fasting on intrinsic functional connectivity: a double-blind fMRI study. Brain Imaging Behav. 2018;12(4) :1150–1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Clemens B, Wagels L, Bauchmüller M, Bergs R, Habel U, Kohn N. Alerted default mode: functional connectivity changes in the aftermath of social stress. Sci Rep. 2017;7:1–9. doi: 10.1038/srep40180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hermans EJ, Henckens MJAG, Joëls M, Fernández G. Dynamic adaptation of large-scale brain networks in response to acute stressors. Trends Neurosci. 2014;37:304–314. doi: 10.1016/j.tins.2014.03.006. [DOI] [PubMed] [Google Scholar]
- 21.Kohn N, Hermans EJ, Fernández G. Cognitive benefit and cost of acute stress is differentially modulated by individual brain state. Soc Cogn Affect Neurosci. 2017;12:1179–1187. doi: 10.1093/scan/nsx043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Raichle ME. The brain’s default mode network. Annu Rev Neurosci. 2015;38:433–447. doi: 10.1146/annurev-neuro-071013-014030. [DOI] [PubMed] [Google Scholar]
- 23.Buckner RL, Andrews-Hanna JR, Schacter DL. The brain’s default network: anatomy, function, and relevance to disease. Ann N Y Acad Sci. 2008;1124:1–38. doi: 10.1196/annals.1440.011. [DOI] [PubMed] [Google Scholar]
- 24.Andrews-Hanna JR, Smallwood J, Spreng RN. The default network and self-generated thought: component processes, dynamic control, and clinical relevance. Ann N Y Acad Sci. 2014;1316:29–52. doi: 10.1111/nyas.12360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Broyd SJ, Demanuele C, Debener S, Helps SK, James CJ, Sonuga-Barke EJS. Default-mode brain dysfunction in mental disorders: a systematic review. Neurosci Biobehav Rev. 2009;33:279–296. doi: 10.1016/j.neubiorev.2008.09.002. [DOI] [PubMed] [Google Scholar]
- 26.Gusnard DA, Akbudak E, Shulman GL, Raichle ME. Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function. Proc Natl Acad Sci U S A. 2001;98:4259–4264. doi: 10.1073/pnas.071043098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci U S A. 2001;98:676–682. doi: 10.1073/pnas.98.2.676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Shulman G, Fiez J, and Corbetta M. Common blood flow changes across visual tasks: II. Decreases in cerebral cortex. J Cogn . 1997;9(5):648–663. [DOI] [PubMed] [Google Scholar]
- 29.Raichle ME, Snyder AZ. A default mode of brain function: a brief history of an evolving idea. NeuroImage. 2007;37:1083–1090. doi: 10.1016/j.neuroimage.2007.02.041. [DOI] [PubMed] [Google Scholar]
- 30.Mevel K, Landeau B, Fouquet M, La Joie R, Villain N, Mézenge F, Perrotin A, Eustache F, Desgranges B, Chételat G. Age effect on the default mode network, inner thoughts, and cognitive abilities. Neurobiol Aging. 2013;34:1292–1301. doi: 10.1016/j.neurobiolaging.2012.08.018. [DOI] [PubMed] [Google Scholar]
- 31.Sestieri C, Corbetta M, Romani GL, Shulman GL. Episodic memory retrieval, parietal cortex, and the default mode network: functional and topographic analyses. J Neurosci. 2011;31:4407–4420. doi: 10.1523/JNEUROSCI.3335-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bar M. The proactive brain: using analogies and associations to generate predictions. Trends Cogn Sci. 2007;11:280–289. doi: 10.1016/j.tics.2007.05.005. [DOI] [PubMed] [Google Scholar]
- 33.Mason MF, Norton MI, Van Horn JD, Wegner DM, Grafton ST, Macrae CN. Wandering Minds: the Default Network and Stimulus-Independent Thought. Science. 2007;315:315. doi: 10.1126/science.315.5810.315a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Raichle ME. The restless brain: how intrinsic activity organizes brain function. Philosoph Trans Royal Soc B. 2015;370:20140172–20140172. doi: 10.1098/rstb.2014.0172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kohn N, Toygar T, Weidenfeld C, Berthold-Losleben M, Chechko N, Orfanos S, Vocke S, Durst A, Laoutidis ZG, Karges W, et al. In a sweet mood? Effects of experimental modulation of blood glucose levels on mood-induction during fMRI. NeuroImage. 2015;113:246–256. doi: 10.1016/j.neuroimage.2015.03.024. [DOI] [PubMed] [Google Scholar]
- 36.Petersen SE, Posner MI. The attention system of the human brain: 20 years after. Annu Rev Neurosci. 2012;35:73–89. doi: 10.1146/annurev-neuro-062111-150525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Andrews-Hanna JR, Reidler JS, Sepulcre J, Poulin R, Buckner RL. Functional-anatomic fractionation of the brain’s default network. Neuron. 2010;65:550–562. doi: 10.1016/j.neuron.2010.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Menon V. Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cogn Sci. 2011;15:483–506. doi: 10.1016/j.tics.2011.08.003. [DOI] [PubMed] [Google Scholar]
- 39.Foster JA, Rinaman L, Cryan JF. Stress & the gut-brain axis: regulation by the microbiome. Neurobio Stress. 2017;7:124–136. doi: 10.1016/j.ynstr.2017.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sandhu KV, Sherwin E, Schellekens H, Stanton C, Dinan TG, Cryan JF. Feeding the microbiota-gut-brain axis: diet, microbiome, and neuropsychiatry. Trans Res. 2017;179:223–244. doi: 10.1016/j.trsl.2016.10.002. [DOI] [PubMed] [Google Scholar]
- 41.de Weerth C. Do bacteria shape our development? Crosstalk between intestinal microbiota and HPA axis. Neurosci Biobehav Rev. 2017;83:458–471. doi: 10.1016/j.neubiorev.2017.09.016. [DOI] [PubMed] [Google Scholar]
- 42.Müller NCJ, Dresler M, Janzen G, Beckmann CF, Fernández G, Kohn N. Medial prefrontal decoupling from the default mode network benefits memory. NeuroImage. 2020;210:116543. doi: 10.1016/j.neuroimage.2020.116543. [DOI] [PubMed] [Google Scholar]
- 43.Allen AP, Hutch W, Borre YE, Kennedy PJ, Temko A, Boylan G, Murphy E, Cryan JF, Dinan TG, Clarke G. Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl Psychiatry. 2016;6:e939–e939. doi: 10.1038/tp.2016.191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Rodriguez J, Hiel S, Neyrinck AM, Roy TL, Pötgens SA, Leyrolle Q, Pachikian BD, Gianfrancesco MA, Cani PD, Paquot N, et al. Discovery of the gut microbial signature driving the efficacy of prebiotic intervention in obese patients. Gut [Internet]. 2020;69(11):1975–1987. [accessed 2020 Aug 10]. https://gut.bmj.com/content/early/2020/02/10/gutjnl-2019-319726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Jianguo L, Xueyang J, Cui W, Changxin W, Xuemei Q. Altered gut metabolome contributes to depression-like behaviors in rats exposed to chronic unpredictable mild stress. Transl Psychiatry. 2019;9:1–14. doi: 10.1038/s41398-018-0355-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tian T, Xu B, Qin Y, Fan L, Chen J, Zheng P, Gong X, Wang H, Bai M, Pu J, et al. Clostridium butyricum miyairi 588 has preventive effects on chronic social defeat stress-induced depressive-like behaviour and modulates microglial activation in mice. Biochem Biophys Res Commun. 2019;516:430–436. doi: 10.1016/j.bbrc.2019.06.053. [DOI] [PubMed] [Google Scholar]
- 47.Abbott AE, Nair A, Keown CL, Datko M, Jahedi A, Fishman I, Müller R-A. Patterns of Atypical Functional Connectivity and Behavioral Links in Autism Differ Between Default, Salience, and Executive Networks. Cereb Cortex. 2016;26:4034–4045. doi: 10.1093/cercor/bhv191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kang D-W, Adams JB, Coleman DM, Pollard EL, Maldonado J, McDonough-Means S, Caporaso JG, Krajmalnik-Brown R. Long-term benefit of Microbiota Transfer Therapy on autism symptoms and gut microbiota. Sci Rep. 2019;9:1–9. doi: 10.1038/s41598-018-37186-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Prehn-Kristensen A, Zimmermann A, Tittmann L, Lieb W, Schreiber S, Baving L, Fischer A. Reduced microbiome alpha diversity in young patients with ADHD. PLoS One [Internet]. 2018;13:e0200728. [accessed 2020 Apr 20]. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6042771/. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gerhardt S, Mohajeri MH. Changes of colonic bacterial composition in Parkinson’s disease and other neurodegenerative diseases. Nutrients [Internet] 2018;10:708. [accessed 2020 Apr 20]. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6024871/. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ho LKH, Tong VJW, Syn N, Nagarajan N, Tham EH, Tay SK, Shorey S, Tambyah PA, Law ECN. Gut microbiota changes in children with autism spectrum disorder: a systematic review. Gut Pathog. 2020;12:6. doi: 10.1186/s13099-020-0346-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Tengeler AC, Dam SA, Wiesmann M, Naaijen J, van Bodegom M, Belzer C, Dederen PJ, Verweij V, Franke B, Kozicz T, et al. Gut microbiota from persons with attention-deficit/hyperactivity disorder affects the brain in mice. Microbiome. 2020;8:44. doi: 10.1186/s40168-020-00816-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Di X, Biswal BB. Modulatory interactions between the default mode network and task positive networks in resting-state. PeerJ [Internet]. 2014;2:e367. [accessed 2020 Apr 20]. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4017816/. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, Raichle ME. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc Natl Acad Sci U S A. 2005;102:9673–9678. doi: 10.1073/pnas.0504136102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Poldrack RA. Inferring mental states from neuroimaging data: from reverse inference to large-scale decoding. Neuron. 2011;72:692–697. doi: 10.1016/j.neuron.2011.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Papalini S, Michels F, Kohn N, Wegman J, van Hemert S, Roelofs K, Arias-Vasquez A, Aarts E. Stress matters: randomized controlled trial on the effect of probiotics on neurocognition. Neurobio Stress. 2019;10:100141. doi: 10.1016/j.ynstr.2018.100141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jenkinson M, Beckmann CF, Behrens TEJ, Woolrich MW, Smith SM. FSL. NeuroImage. 2012;62:782–790. doi: 10.1016/j.neuroimage.2011.09.015. [DOI] [PubMed] [Google Scholar]
- 58.Pruim RHR, Mennes M, van Rooij D, Llera A, Buitelaar JK, Beckmann CF. ICA-AROMA: a robust ICA-based strategy for removing motion artifacts from fMRI data. Neuroimage. 2015;112:267–277. doi: 10.1016/j.neuroimage.2015.02.064. [DOI] [PubMed] [Google Scholar]
- 59.Pruim RHR, Mennes M, Buitelaar JK, Beckmann CF. Evaluation of ICA-AROMA and alternative strategies for motion artifact removal in resting state fMRI. Neuroimage. 2015;112:278–287. doi: 10.1016/j.neuroimage.2015.02.063. [DOI] [PubMed] [Google Scholar]
- 60.Szopinska JW, Gresse R, van der Marel S, Boekhorst J, Lukovac S, van Swam I, Franke B, Timmerman H, Belzer C, Arias Vasquez A. Reliability of a partici pant-friendly fecal collec tion method for micr obiome analyses: a step towards large sample size investigation. BMC Micr obiol. 2018;18:110. doi: 10.1186/s12866-018-1249-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Fernández-Calleja JMS, Konstanti P, Swarts HJM, Bou wman LMS, Garcia-Camp ayo V, Bill ecke N, Oosting A, Smidt H, Keijer J, van Schot horst EM. Non-invasive continuous real-time in vivo analysis of microbial hydr ogen produ ction shows adaptation to ferm entable carbo hydrates in mice. Sci Rep. 2018;8:1–16. doi: 10.1038/s41598-018-33619-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ramiro-Garcia J, Hermes GDA, Giatsis C, Sipkema D, Zoetendal EG, Schaap PJ, Smidt H. NG-Tax, a highly accurate and validated pipeline for analysis of 16S rRNA amplicons from complex biomes. F1000Res. 2018;5:1791. doi: 10.12688/f1000research.9227.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Szopinska-Tokov J, Dam S, Naaijen J, Konstanti P, Rommelse N, Belzer C, Buitelaar J, Franke B, Aarts E, Arias Vasquez A. Investigating the gut microbiota composition of individuals with attention-deficit/hyperactivity disorder and association with symptoms. Microorganisms. 2020;8:406. doi: 10.3390/microorganisms8030406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner FO. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2013;41:D590–6. doi: 10.1093/nar/gks1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.McMurdie PJ, Holmes S. Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLOS ONE. 2013;8:e61217. doi: 10.1371/journal.pone.0061217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Choudrey R. Variational methods for bayesian independent component analysis. [DOI] [PubMed]
- 67.Llera A. Linked ICA in HCP500 [Internet]. GitHub2019. Accessed 20 Apr 2021. https://github.com/allera/Llera_elife_2019_1
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data, documentation and code will be shared via the DondersSharingCollection (https://doi.org/10.34973/3j3h-ts61).