

Abstract
Traditionally used for bulk biochemical assays, Xenopus laevis egg extracts have emerged as a powerful imaging-based tool for studying cytoplasmic phenomena, such as cytokinesis, mitotic spindle formation and assembly of the nucleus. Building upon early methods that imaged fixed extracts sampled at sparse time points, recent approaches image live extracts using time-lapse microscopy, revealing more dynamical features with enhanced temporal resolution. These methods usually require sophisticated surface treatments of the imaging vessel. Here we introduce an alternative method for live imaging of egg extracts that require no chemical surface treatment. It is simple to implement and utilizes mass-produced laboratory consumables for imaging. We describe a system that can be used for both wide-field and confocal microscopy. It is designed for imaging extracts in a 2-dimensional (2D) field, but can be easily extended to imaging in 3D. It is well-suited for studying spatial pattern formation within the cytoplasm. With representative data, we demonstrate the typical dynamic organization of microtubules, nuclei and mitochondria in interphase extracts prepared using this method. These image data can provide quantitative information on cytoplasmic dynamics and spatial organization.
Introduction
The cytoplasm constitutes the main volume of a cell and has a distinct organization. The ingredients of the eukaryotic cytoplasm can self-assemble into a wide range of spatial structures, such as microtubule asters and the Golgi apparatus, which in turn are dynamically arranged and turned over depending on the cell’s identity and physiological state. Understanding the spatial organization of the cytoplasm and its link to cellular functions is thus important for understanding how the cell works. Xenopus laevis egg extracts have traditionally been used for bulk biochemical assays1,2,3,4,5,6,7,8, but recent work establishes them as a powerful live imaging system for mechanistic studies of cytoplasmic structures and their cellular functions9,10,11,12,13,14,15,16,17,18. These undiluted extracts preserve many structures and functions of the cytoplasm, while allowing direct manipulations of cytoplasmic contents not achievable in conventional cellbased models19,20. This makes them ideal for characterizing cytoplasmic phenomena and dissecting their mechanistic underpinnings.
Existing methods for imaging extracts require chemical surface modifications, or fabrication of microfluidic devices. One coverslip-based method requires polyethylene glycol (PEG) passivation of glass coverslips21. A microemulsion-based method requires vapor deposition of trichloro(1H,1H,2H,2H-perfluorooctyl)silane on glass surfaces22,23. Microfluidic-based systems allow precise control of the volume, geometry and composition of extract droplets, but require specialized microfabrication facilities11,12,24.
Here we introduce an alternative method for imaging egg extracts that is easy to implement and utilizes readily available, low-cost materials. This includes preparation of an imaging chamber with a slide and a coverslip coated with fluorinated ethylene propylene (FEP) tape. The chamber can be used for imaging extracts with a variety of microscopy systems, including stereoscopes and upright and inverted microscopes. This method requires no chemical treatment of surfaces while achieving similar optical clarity obtained with existing glass-based methods discussed above. It is designed to image a layer of extracts with a uniform thickness across a 2D field, and can be easily extended to image a 3D volume of extracts. It is well suited for time-lapse imaging of collective cytoplasmic behavior over a large field of view.
We have used interphase-arrested egg extracts to demonstrate our imaging method. The extract preparation follows the protocol of Deming and Kornbluth19. Briefly, eggs naturally arrested in metaphase of meiosis II are crushed by a low speed spin. This spin releases the cytoplasm from meiotic arrest and allows the extract to proceed into interphase. Normally, cytochalasin B is added prior to the crushing spin to inhibit F-actin formation. However, it can be omitted if F-actin is desired. Cycloheximide is also added prior to the crushing spin to prevent the interphase extract from entering the next mitosis. The extracts are subsequently placed in the aforementioned imaging chambers and placed on a microscope. Finally, images are recorded over time at defined intervals by a camera connected to the microscope, producing time-lapse image series that capture the dynamical behavior of the extract in a 2D field.
Protocol
All methods described here have been approved by the Institutional Animal Care and Use Committee (IACUC) of Stanford University.
1. Preparation of slides and coverslips
Apply a layer of fluorinated ethylene propylene (FEP) adhesive tape to a glass slide with a roller applicator. Cut off excessive tape over the edges with a clean razor blade. Prepare FEP tape-coated coverslips in the same way (Figure 1A).
-
Apply a double-sided sticky imaging spacer to the FEP tape-coated side of the slide. Leave the protective liner on the top unpeeled (Figure 1A).
NOTE: The slides and coverslips should be prepared before the experiment. They can be used immediately, or stored in boxes to prevent dust accumulation on surfaces. The well in the imaging spacer is 120 μm deep and has a diameter of 9 mm.
Figure 1: Schematics and photos related to the experimental procedure.

(A) Schematic diagram for preparing FEP tape-coated glass coverslips and slides. (B) Xenopus laevis eggs deposited in egg laying buffer, with examples of poor-quality eggs indicated by arrows. Yellow arrows, examples of eggs that look like white puffy balls. Red arrows, examples of eggs that appear in a string. (C) Examples of Xenopus laevis eggs with normal appearance. (D) Examples of poor-quality eggs with irregular or mottled pigment. (E) A deteriorating egg with a growing white region. (F) An egg that shows darkened and contracted pigmented area, possibly due to parthenogenetic activation. (G) The layers formed by ruptured Xenopus laevis eggs after the 12,000 × g centrifugation in step 2.11. (H) Schematics for preparing extract imaging chamber in step 2.15. (I) A photo of a prepared imaging chamber with an egg extract inside. (C) and (D) share the same scale bar at the bottom of (D). (E) and (F) share the same scale bar at the bottom of (F). The scale bars in (B) (C) (D) (E) (F) and (I) are approximate.
2. Preparation and live imaging of interphase-arrested egg extracts
NOTE: The following protocol is adapted from Deming and Kornbluth19, Murray20, and Smythe and Newport25 with modifications. All steps should be performed at room temperature unless otherwise noted.
Three to ten days before egg collection, inject mature female Xenopus laevis frogs subcutaneously into the dorsal lymph sac with 100 IU of pregnant mare serum gonadotropin (PMSG).
Sixteen to eighteen hours prior to planned egg collection, inject the frogs from step 2.1 with 500 IU of human chorionic gonadotropin (hCG). Leave frogs at 18 °C in egg laying buffer (100 mM NaCl, 2 mM KCl, 1 mM MgSO4·7H2O, 2.5 mM CaCl2·2H2O, 0.5 mM HEPES, 0.1 mM EDTA, prepare as a 20x stock solution at pH 7.4 and dilute with clean frog tank water to 1x before use) until egg collection.
-
On the day of the experiment, collect eggs in a large glass Petri dish and assess egg quality. Discard any eggs that look like white puffy balls or appear in a string (Figure 1B). Examine the eggs under a stereoscope, keep the eggs with normal appearance (Figure 1C), and discard those with irregular or mottled pigment (Figure 1D).
NOTE: This protocol works with eggs collected from a single frog, which typically lays 25 mL of eggs by 16 hours after hCG injection. Usually, a total of 3 to 6 frogs are induced by hCG, and the frog with the highest egg quality is chosen for the extract preparation experiment.
Transfer eggs to a 400 mL glass beaker, and remove as much egg laying buffer as possible by decanting.
Incubate the eggs in 100 mL of freshly prepared dejellying solution (2% w/v L-cysteine in water, adjust to pH 8.0 with NaOH) and gently swirl them periodically. After about 3 minutes, pour off the solution, and add 100 mL of fresh dejellying solution. Continue the incubation until the eggs are tightly packed (no space between eggs), but avoid leaving eggs in the dejellying solution for more than a total of 5 minutes.
Remove as much of the dejellying solution as possible by decanting, and wash the eggs in 0.25x MMR buffer (25 mM NaCl, 0.5 mM KCl, 0.25 mM MgCl2, 0.5 mM CaCl2, 0.025 mM EDTA, 1.25 mM HEPES, prepared as a 10x stock solution, adjusted to pH 7.8 with NaOH, and diluted in Milli-Q water before use) by adding the buffer, swirling the eggs, and then pouring off the buffer. Repeat a few times until a total of 1 L of the buffer is used for the washes.
-
Wash the eggs a few times with a total of 400 mL of egg lysis buffer (250 mM sucrose, 10 mM HEPES, 50 mM KCl, 2.5 mM MgCl2, 1 mM DTT, made fresh and adjusted to pH 7.7 with KOH). Remove eggs with abnormal appearance using a Pasteur pipette between the washes.
NOTE: Eggs with abnormal appearance refer to those that look like white puffy balls (Figure 1B), have mottled pigmentation (Figure 1D), are deteriorating with a growing white region (Figure 1E), or show darkened and contracted pigmented area in the animal hemisphere (Figure 1F).
Using a transfer pipette with its tip cut wide open, transfer the eggs to a 17 mL round-bottom centrifuge tube containing 1 mL of egg lysis buffer. Spin the tube in a clinical centrifuge at 400 × g for 15 seconds to pack the eggs.
-
Remove as much of the egg lysis buffer as possible from the top of packed eggs using a Pasteur pipette.
NOTE: It is important to remove as much buffer from the packed eggs as possible, in order to minimize the dilution of the egg extract. Sometimes it is necessary to remove some loose eggs along with the residual buffer to accomplish this.
-
Determine the approximate volume of the packed eggs, and then add 5 μg/mL aprotinin, 5 μg/mL leupeptin, 5 μg/mL cytochalasin B, and 50 μg/mL cycloheximide directly on top of the packed eggs.
NOTE: Aprotinin and leupeptin are protease inhibitors. Cytochalasin B inhibits actin polymerization, preventing the extract from contracting and gelling26. Cycloheximide inhibits protein synthesis, thereby keeping the extract in the interphase of the cell cycle.
-
Crush the eggs by centrifuging the tube at 12,000 × g, 4 °C, for 15 minutes, in a swinging bucket rotor.
NOTE: At the end of the centrifugation, the eggs should have ruptured and the lysate separated into three main layers: a yellow lipid layer on top, the cytoplasmic extract (also called crude extract) in the middle, and a dark dense layer containing the pigment granules at the bottom (Figure 1G).
-
Attach an 18-gauge needle to a syringe. With the needle tip bevel facing up, puncture the tube from the side at the bottom of the cytoplasmic layer, and recover the extract by drawing slowly.
NOTE: Draw the cytoplasmic extract slowly to avoid the inclusion of contaminating content from the yellow lipid layer.
Transfer the recovered cytoplasmic extract to a new microcentrifuge tube and hold it on ice. Use the extract within 1 hour.
-
When ready to image, supplement the extract with desired reagents and fluorescence imaging probes.
NOTE: Fluorescence imaging probes label specific cytoplasmic structures so that they can be visualized by a fluorescence microscope.
Remove the top protective liner from the imaging spacer on the slide prepared in step 1.2, and deposit approximately 7 μL of extract at the center of the well. Immediately apply the FEP tape-coated coverslip with the FEP side facing the extract to seal the well. Quickly proceed to imaging (Figure 1H,I).
-
Set the slide on an inverted or upright microscope with a motorized stage and a digital camera. Image the extracts at desired spatial positions and time intervals in both bright-field and fluorescence channels.
NOTE: Typically, a 5x objective is used for imaging. The motorized stage enables automated image acquisition at multiple defined spatial positions. Bright-field microscopy visualizes cytoplasmic structures with different degrees of transparency. Fluorescence microscopy visualizes the cytoplasmic structures specifically labeled by the fluorescence imaging probes added in step 2.14. The camera records time-lapse images of these structures, thereby capturing the dynamics of cytoplasmic organization.
Representative Results
Xenopus laevis egg extracts can be used to study the self-organization of the cytoplasm during interphase. Figure 2A shows results from a successful experiment.We supplemented interphase-arrested extracts with demembranated Xenopus laevis sperm nuclei19 at a concentration of 27 nuclei/μL and 0.38 μM purified GST-GFP-NLS27,28,29,30 (fusion protein consisting of glutathione-S-transferase, green fluorescent protein, and a nuclear localization sequence) to allow reconstitution and visualization of interphase nuclei. We also added 1 μM fluorescently labeled tubulin to visualize microtubules, and 500 nM MitoTracker Red CMXRos to visualize mitochondria. Moments after the extract was placed in the imaging chamber, the cytoplasm appeared disorganized. In the course of the next 30 minutes at room temperature, the cytoplasm started to self-organize into cell-like compartments. Microtubule asters grew from centrosomes introduced with the sperm nuclei, and formed microtubule-depleted border zones when met with microtubules from neighboring asters. GST-GFP-NLS protein translocated into the round interphase nuclei self-assembled from the added demembranated sperm nuclei. Areas depleted of light scattering cytoplasmic components were visible in both bright-field and mitochondria channels (Figure 2A, 20 min and 35 min). Mitochondria also became depleted from the borders established by microtubules, and became enriched in isolated compartments that aligned with microtubule compartments. By 60 min at room temperature, a spatial pattern consisting of cell-like compartments should be well-established, with microtubules forming a hollow wreath-like structure, and mitochondria clearly partitioned into each compartment (Figure 2A, 53 min).
Figure 2: Interphase-arrested egg extracts self-organize into cell-like compartments.
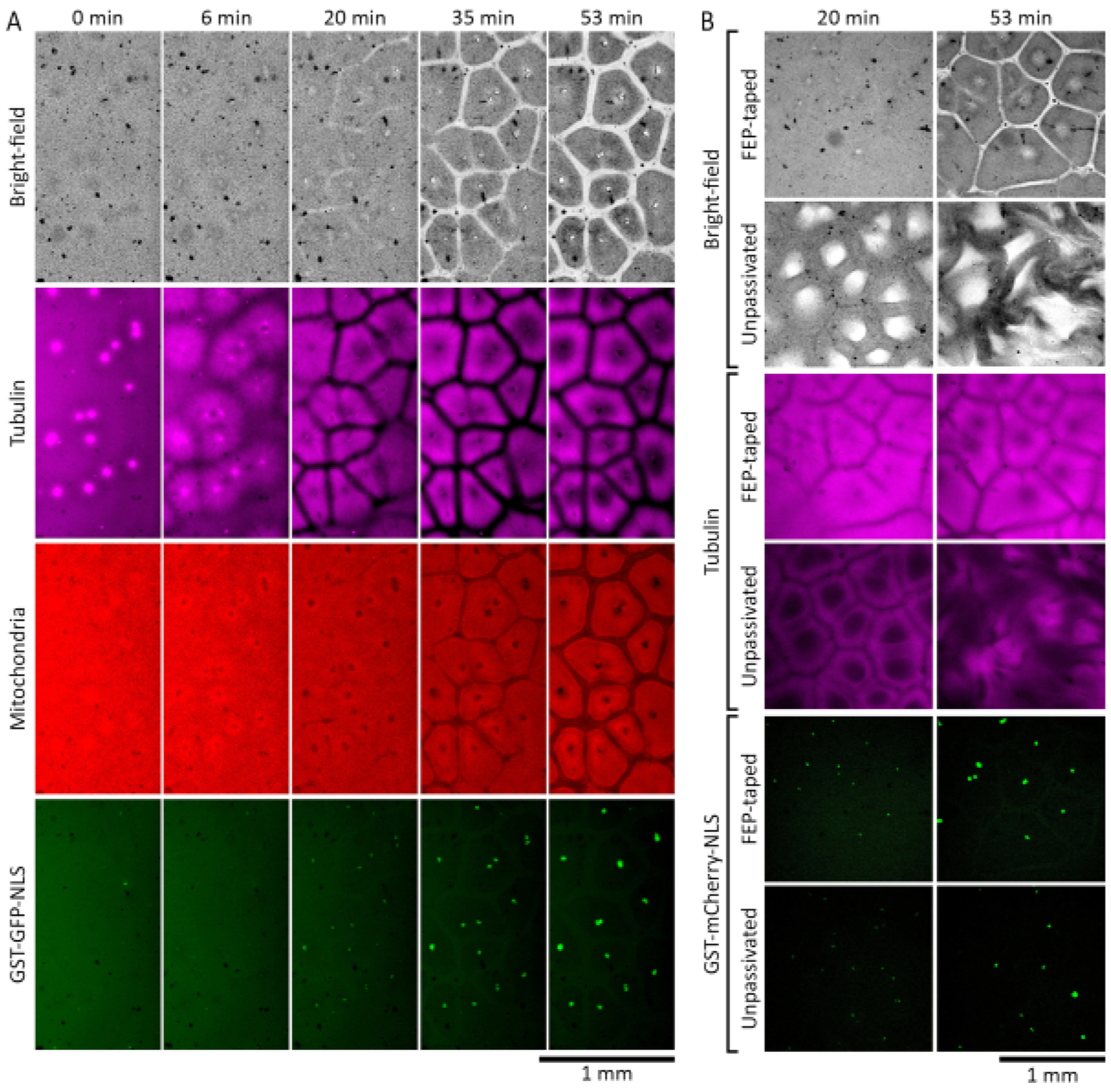
(A)Time-lapse montage of self-organized pattern formation in a thin layer (120 μm) of interphase-arrested Xenopus laevis egg extract. The extract was supplemented with 27 nuclei/μL of demembranated Xenopus laevis sperm nuclei to allow reconstitution of the interphase nuclei. Microtubules were visualized by HiLyte 647-labeled tubulin (shown in magenta), mitochondria by MitoTrackerRed CMXRos (shown in red), and nuclei by GST-GFP-NLS (shown in green). (B) Self-organized pattern formation in interphase-arrested Xenopus laevis egg extracts placed in chambers with and without FEP-tape covered glass surfaces. The extracts were supplemented with 27 nuclei/μL of demembranated Xenopus laevis sperm nuclei to allow reconstitution of the interphase nuclei. Microtubules were visualized by HiLyte 488-labeled tubulin (shown in magenta) and nuclei by GST-mCherry-NLS (shown in green).
Figure 2B compares extract performance in imaging chambers with and without FEP tapes on glass. We supplemented interphase-arrested extracts with demembranated Xenopus laevis sperm nuclei19 at a concentration of 27 nuclei/μL and 0.35 μM GST-mCherry-NLS27,28,29,30 (fusion protein consisting of glutathione-S-transferase, mCherry fluorescent protein, and a nuclear localization sequence) to allow reconstitution and visualization of interphase nuclei. We also added 1 μM fluorescently labeled tubulin to visualize microtubules. Differences in dynamics became obvious by approximately 20 min at room temperature. In the chamber made with FEP-taped glass, the extract self-organized into normal cell-like patterns (Figure 2B, images in rows 1 and 3). However, in the chamber where glass surfaces were not covered by the FEP tape (unpassivated), the extract showed abnormal bright-field and microtubule patterns that became increasingly disrupted over time (Figure 2B, images in rows 2 and 4). No significant differences were observed in nuclear import of the GST-mCherry-NLS protein (Figure 2B, images in rows 5 and 6).
Discussion
Xenopus laevis egg extracts have emerged as a powerful model system for imaging-based studies of various subcellular structures10,14,15,16,17,18,21,31,32,33,34,35,36 and cytoplasmic organization on a whole cell scale9. Here we have described a live imaging method suitable for visualizing dynamic cytoplasmic organization in 2D. The effectiveness of the method is demonstrated by representative results.
Several steps are critical to the success of the method. Egg quality is important for extracts, so steps 2.3 and 2.7 are critical. In our experience, the animal hemisphere of high-quality eggs has uniform pigmentation, a distinct white dot at the center (indicative of germinal vesicle breakdown, GVBD), and a clear border with the vegetal hemisphere (Figure 1C). The extracts made from high-quality eggs have better performance under our imaging conditions. If more than 5% of the eggs have mottled pigment (Figure 1D), have darkened and contracted pigmented area (Figure 1F), look like white puffy balls (Figure 1B), appear in a string (Figure 1B), or show signs of deterioration after the washes in steps 2.6 and 2.7 (Figure 1E), then the whole batch of eggs should be discarded. The extracts retain remarkable biological activity because they are essentially undiluted cytoplasm. Therefore, the step that aims to minimize dilution of the extract (step 2.9) is important for the success of the experiment. For imaging, extracts are handled in very small volumes, and will evaporate quickly if in prolonged contact with air. This will negatively affect their activity. Therefore, in step 2.15, after the extract is deposited, it must be sealed with the FEP tape-coated side of the coverslip as soon as possible. Sealing can be visually confirmed by the texture change at the contact site between the adhesive on the spacer and the coverslip. The spacer should be able to create a complete seal if applied properly. However, if an additional seal is desired, mineral oil can be dispensed between the overhang of the coverslip and the glass slide. The oil can form an additional seal around the spacer by capillary action (Figure 3). Passivation of glass surfaces can reduce non-specific adsorption of molecules, and it is important for interphase microtubule asters in extracts21,37. The application of FEP tape over a glass surface described here appears to offer similar benefits, as suggested by the assembly of normal interphase microtubule asters (Figure 2, 6 min). Therefore, step 1.1 is also critical.
Figure 3: Optional secondary seal for the imaging chamber.
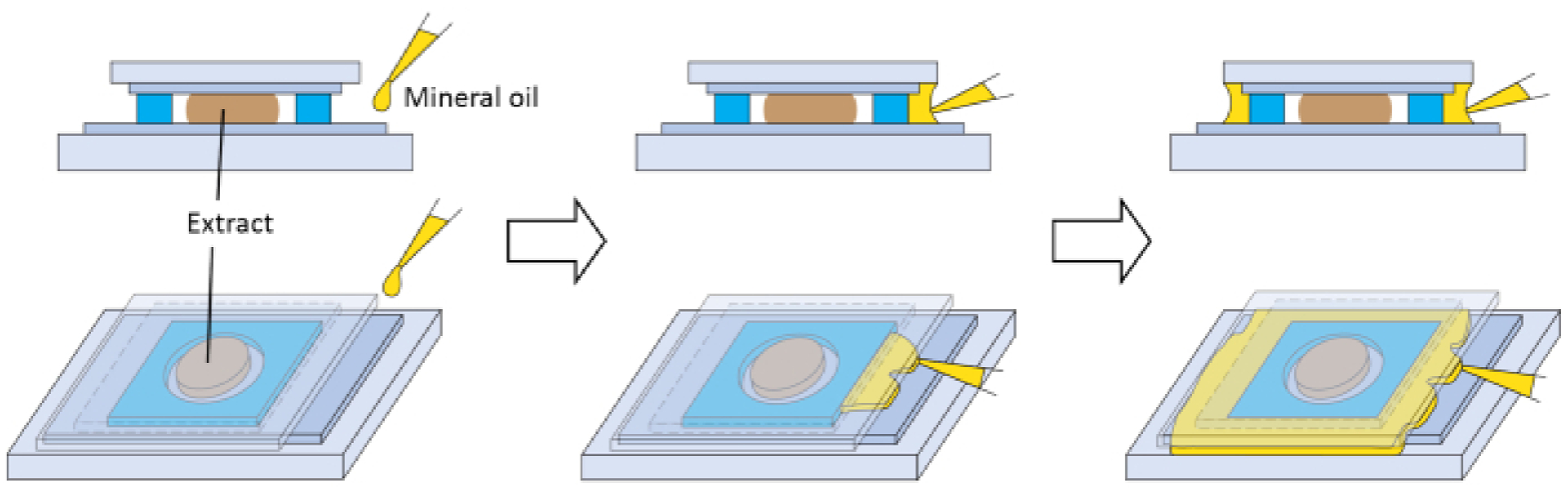
Schematics for preparing an optional secondary seal with mineral oil to prevent the extract from prolonged contact with air.
We demonstrated the application of an imaging method using interphase-arrested egg extracts following the protocol of Deming and Kornbluth19. By default, the protocol supplements the extracts with the actin polymerization inhibitor cytochalasin B to prevent gelation-contraction in the extracts after extended incubation at room temperature26. To allow actin polymerization and observe actin dynamics, one can leave out the cytochalasin B in step 2.10 of the protocol9. A modification we have made to the Deming and Kornbluth protocol is that we do not supplement extracts with an energy mix to regenerate ATP19. This is because in our hands, in extracts supplemented with this ATP-regenerating mix19 and sperm nuclei, microtubules occasionally form a crosslinked lattice that interferes with cytoplasmic pattern formation. Therefore, the protocol does not include the step that adds the energy mix to the extracts19.
The interphase extract protocol relies on crushing eggs in EGTA-free buffer to release them from meiotic arrest (CSF-arrest)37. The extracts subsequently progress into interphase, and are kept in interphase by cycloheximide. There are other established methods for preparing interphase extracts. Some methods first prepare extracts that maintain meiotic arrest by crushing eggs in lysis buffer with EGTA38, and then release the arrest by adding calcium, driving the extract into interphase21,37,39. The extracts can be subsequently held in interphase by addition of protein synthesis inhibitors such as cycloheximide37,39. Other methods parthenogenetically activate eggs with calcium ionophore (A23187) or electric shock to release them from meiotic arrest, before crushing the eggs in the absence of EGTA20,28 (reviewed in Field et al.37). These extracts can enter interphase but typically will not stay there as they are able to undergo multiple cell cycles. Likewise, well-established methods optimized for preparing extracts with intact actin cytoskeleton have been developed10,37,39. The imaging method may be suitable for these types of extracts, but we have not tested it with them.
For the purpose of imaging the internal organization of the cytoplasm, the imaging system presented here are relatively easy to set up, requiring only applying an FEP tape to glass surfaces. It allows assembly of cytoskeletal structures in extracts9 reported in more sophisticated imaging systems where glass surfaces are passivated with poly-L-lysine-g-polyethylene glycol (PLL-g-PEG) treatment or coated with supported lipid bilayers10,21. The method allows the extract layer to form with a defined thickness (determined by the depth of the spacer, which is 120 μm for the system shown in Figure 1A, 1H, 1I, and Figure 2). We can adjust the thickness by stacking additional spacers. We have stacked up to 6 such spacers (720 μm thick) and the compartments formed normally. This flexibility enables future applications such as imaging of the extracts in 3D using confocal or light-sheet microscopy.
Acknowledgments
We thank J. Kamenz, Y. Chen, and W. Y. C. Huang for comments on the manuscript. This work was supported by grants from the National Institutes of Health (R01 GM110564, P50 GM107615, and R35 GM131792) awarded to James E. Ferrell, Jr.
Footnotes
A complete version of this article that includes the video component is available at http://dx.doi.org/10.3791/61923.
Disclosures
The authors have nothing to disclose.
References
- 1.Murray AW, Kirschner MW Cyclin synthesis drives the early embryonic cell cycle. Nature. 339 (6222), 275–280 (1989). [DOI] [PubMed] [Google Scholar]
- 2.Dunphy WG, Brizuela L, Beach D, Newport J The Xenopus cdc2 protein is a component of MPF, a cytoplasmic regulator of mitosis. Cell. 54 (3), 423–431 (1988). [DOI] [PubMed] [Google Scholar]
- 3.Minshull J, Golsteyn R, Hill CS, Hunt T The A- and B-type cyclin associated cdc2 kinases in Xenopus turn on and off at different times in the cell cycle. EMBO J. 9 (9), 2865–2875 (1990). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wu JQ et al. PP1-mediated dephosphorylation of phosphoproteins at mitotic exit is controlled by inhibitor-1 and PP1 phosphorylation. Nature Cell Biology. 11 (5), 644–651 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pomerening JR, Sontag ED, Ferrell JE Jr. Building a cell cycle oscillator: hysteresis and bistability in the activation of Cdc2. Nature Cell Biology. 5 (4), 346–351 (2003). [DOI] [PubMed] [Google Scholar]
- 6.Mochida S, Maslen SL, Skehel M, Hunt T Greatwall phosphorylates an inhibitor of protein phosphatase 2A that is essential for mitosis. Science. 330 (6011), 1670–1673 (2010). [DOI] [PubMed] [Google Scholar]
- 7.Blow JJ, Laskey RA Initiation of DNA replication in nuclei and purified DNA by a cell-free extract of Xenopus eggs. Cell. 47 (4), 577–587 (1986). [DOI] [PubMed] [Google Scholar]
- 8.Lohka MJ, Masui Y Formation in vitro of sperm pronuclei and mitotic chromosomes induced by amphibian ooplasmic components. Science. 220 (4598), 719–721 (1983). [DOI] [PubMed] [Google Scholar]
- 9.Cheng X, Ferrell JE Jr. Spontaneous emergence of cell-like organization in Xenopus egg extracts. Science. 366 (6465), 631–637 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nguyen PA et al. Spatial organization of cytokinesis signaling reconstituted in a cell-free system. Science. 346 (6206), 244–247 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Good MC, Vahey MD, Skandarajah A, Fletcher DA, Heald R Cytoplasmic volume modulates spindle size during embryogenesis. Science. 342 (6160), 856–860 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hazel J et al. Changes in cytoplasmic volume are sufficient to drive spindle scaling. Science. 342 (6160), 853–856 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Brownlee C, Heald R Importin alpha Partitioning to the Plasma Membrane Regulates Intracellular Scaling. Cell. 176 (4), 805–815 e808 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nguyen PA, Field CM, Mitchison TJ Prc1E and Kif4A control microtubule organization within and between large Xenopus egg asters. Molecular Biology of the Cell. 29 (3), 304–316 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Desai A, Maddox PS, Mitchison TJ, Salmon ED Anaphase A chromosome movement and poleward spindle microtubule flux occur At similar rates in Xenopus extract spindles.Journal of Cell Biology. 141 (3), 703–713 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mitchison TJ et al. Roles of polymerization dynamics, opposed motors, and a tensile element in governing the length of Xenopus extract meiotic spindles. Molecular Biology of the Cell. 16 (6), 3064–3076 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mitchison TJ et al. Bipolarization and poleward flux correlate during Xenopus extract spindle assembly. Molecular Biology of the Cell. 15 (12), 5603–5615 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Murray AW, Desai AB, Salmon ED Real time observation of anaphase in vitro. Proceedings of the National Academy of Sciences of the United States of America. 93 (22), 12327–12332 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Deming P, Kornbluth S Study of apoptosis in vitro using the Xenopus egg extract reconstitution system. Methods in Molecular Biology. 322 379–393 (2006). [DOI] [PubMed] [Google Scholar]
- 20.Murray AW Cell cycle extracts. Methods in Cell Biology. 36 581–605 (1991). [PubMed] [Google Scholar]
- 21.Field CM, Mitchison TJ Assembly of Spindles and Asters in Xenopus Egg Extracts. Cold Spring Harbor Protocols. 2018 (6) (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Guan Y et al. A robust and tunable mitotic oscillator in artificial cells. Elife. 7 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Guan Y, Wang S, Jin M, Xu H, Yang Q Reconstitution of Cell-cycle Oscillations in Microemulsions of Cell-free Xenopus Egg Extracts. Journal of Visualized Experiments. 10.3791/58240 (139) (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Oakey J, Gatlin JC Microfluidic Encapsulation of Demembranated Sperm Nuclei in Xenopus Egg Extracts. Cold Spring Harbor Protocols. 2018 (8) (2018). [DOI] [PubMed] [Google Scholar]
- 25.Smythe C, Newport JW Systems for the study of nuclear assembly, DNA replication, and nuclear breakdown in Xenopus laevis egg extracts. Methods in Cell Biology. 35 449–468 (1991). [DOI] [PubMed] [Google Scholar]
- 26.Field CM et al. Actin behavior in bulk cytoplasm is cell cycle regulated in early vertebrate embryos. Journal of Cell Science. 124 (Pt 12), 2086–2095 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chang JB, Ferrell JE Jr. Mitotic trigger waves and the spatial coordination of the Xenopus cell cycle. Nature. 500 (7464), 603–607 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chang JB, Ferrell JE Jr. Robustly Cycling Xenopus laevis Cell-Free Extracts in Teflon Chambers. Cold Spring Harbor Protocols. 2018 (8) (2018). [DOI] [PubMed] [Google Scholar]
- 29.Chatterjee S, Javier M, Stochaj U In vivo analysis of nuclear protein traffic in mammalian cells. Experimental Cell Research. 236 (1), 346–350 (1997). [DOI] [PubMed] [Google Scholar]
- 30.Mochida S, Hunt T Calcineurin is required to release Xenopus egg extracts from meiotic M phase. Nature. 449 (7160), 336–340 (2007). [DOI] [PubMed] [Google Scholar]
- 31.Heald R et al. Self-organization of microtubules into bipolar spindles around artificial chromosomes in Xenopus egg extracts. Nature. 382 (6590), 420–425 (1996). [DOI] [PubMed] [Google Scholar]
- 32.Helmke KJ, Heald R TPX2 levels modulate meiotic spindle size and architecture in Xenopus egg extracts. Journal of Cell Biology. 206 (3), 385–393 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sawin KE, Mitchison TJ Mitotic spindle assembly by two different pathways in vitro. Journal of Cell Biology. 112 (5), 925–940 (1991). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Belmont LD, Hyman AA, Sawin KE, Mitchison TJ Real-time visualization of cell cycle-dependent changes in microtubule dynamics in cytoplasmic extracts. Cell. 62 (3), 579–589 (1990). [DOI] [PubMed] [Google Scholar]
- 35.Krauss SW, Lee G, Chasis JA, Mohandas N, Heald R Two protein 4.1 domains essential for mitotic spindle and aster microtubule dynamics and organization in vitro. Journal of Biological Chemistry. 279 (26), 27591–27598 (2004). [DOI] [PubMed] [Google Scholar]
- 36.Wang S, Romano FB, Field CM, Mitchison TJ, Rapoport TA Multiple mechanisms determine ER network morphology during the cell cycle in Xenopus egg extracts. Journal of Cell Biology. 203 (5), 801–814 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Field CM, Nguyen PA, Ishihara K, Groen AC, Mitchison TJ Xenopus egg cytoplasm with intact actin. Methods in Enzymology. 540 399–415 (2014). [DOI] [PubMed] [Google Scholar]
- 38.Good MC, Heald R Preparation of Cellular Extracts from Xenopus Eggs and Embryos. Cold Spring Harbor Protocols. 2018 (6) (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Field CM, Pelletier JF, Mitchison TJ Xenopus extract approaches to studying microtubule organization and signaling in cytokinesis. Methods in Cell Biology. 137 395–435 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]