

Abstract
It has recently been reported that Parkinson’s disease (PD) is preceded and accompanied by daytime sleep attacks, nocturnal insomnia, REM sleep behaviour disorder, hallucinations and depression, symptoms which are frequently as troublesome as the motor symptoms of PD. All these symptoms are present in narcolepsy, which is linked to a selective loss of hypocretin (Hcrt) neurons. In this study, the Hcrt system was examined to determine if Hcrt cells are damaged in PD. The hypothalamus of 11 PD (mean age 79±4) and 5 normal (mean age 77±3) brains was examined. Sections were immunostained for Hcrt-1, melanin concentrating hormone (MCH) and alpha synuclein and glial fibrillary acidic protein (GFAP). The substantia nigra of 10 PD brains and 7 normal brains were used for a study of neuromelanin pigmented cell loss. The severity of PD was assessed using the Hoehn and Yahr scale and the level of neuropathology was assessed using the Braak staging criteria. Cell number, distribution and size were determined with stereologic techniques on a one in eight series.
We found an increasing loss of hypocretin cells with disease progression. Similarly, there was an increased loss of MCH cells with disease severity. Hcrt and MCH cells were lost throughout the anterior to posterior extent of their hypothalamic distributions. The percentage loss of Hcrt cells was minimal in stage I (23%) and was maximal in stage V (62%). Similarly, the percentage loss of MCH cells was lowest in stage I (12%) and was highest in stage V (74%). There was a significant increase (P=0.0006, t=4.25, df=15) in the size of neuromelanin containing cells in PD patients, but no difference in the size of surviving Hcrt (P=0.18, t=1.39, df=14) and MCH (P=0.28, t=1.39, df=14) cells relative to controls.
In summary, we found that PD is characterized by a massive loss of Hcrt neurons. Thus, the loss of Hcrt cells may be a cause of the narcolepsy-like symptoms of PD and may be ameliorated by treatments aimed at reversing the Hcrt deficit. We also saw a substantial loss of hypothalamic MCH neurons. The losses of Hcrt and MCH neurons are significantly correlated with the clinical stage of PD, not disease duration, whereas the loss of neuromelanin cells is significantly correlated only with disease duration. The significant correlations that we found between the loss of Hcrt and MCH neurons and the clinical stage of PD, in contrast to the lack of a relationship of similar strength between loss of neuromelanin containing cells and the clinical symptoms of PD, suggests a previously unappreciated relationship between hypothalamic dysfunction and the time course of the overall clinical picture of PD.
Keywords: Parkinson, narcolepsy, sleep, hypocretin, orexin, melanin concentrating hormone
Introduction
Sleep disturbances with a prevalence that ranges from 74% to 98% (Parkkinen et al., 2005; Mochizuki et al., 2006) are major problems in Parkinson’s disease (PD), often more disturbing than its motor symptoms. Most PD patients have daytime sleep attacks that resemble narcoleptic sleep attacks and that may be increased with the use of dopaminergic agonists, but that also occur independently of these agents (Frucht et al., 1999; Arnulf et al., 2000, 2002; Frucht, 2002; Arnulf, 2006; Rye, 2006; Savitt et al., 2006). Many PD patients have REM sleep at sleep onset (Arnulf et al., 2002; Onofrj et al., 2003). REM sleep behaviour disorder is common in PD (Schenck and Mahowald, 1992; Gagnon et al., 2002) as are hallucinations, some of which have been found to be linked to REM sleep phenomena (Arnulf et al., 2000; Benbir et al., 2006). Recent work has shown that the sleepiness complaints of PD typically precede the motor symptoms and intensify as the disease progresses (Abbott et al., 2005; Dhawan et al., 2006). All of the above symptoms are also characteristic of narcolepsy, suggesting that these symptoms of narcolepsy and PD may have a common cause.
Other symptoms that are common, but not universal, in narcolepsy are also found in PD. Eighty percent of PD patients experience sleep fragmentation resulting from frequent and prolonged awakenings (Askenasy, 2001). This may be exacerbated by the movement disorders of PD but does not appear to be entirely the result of this symptom (Stocchi et al., 1998; Priano et al., 2003; Barone et al., 2004; Grandas and Iranzo, 2004; Arnulf, 2006). The incidence of major depression is markedly elevated in PD. Other chronic diseases are not accompanied by a similar incidence of depression (Frosh, 2006). Disrupted nighttime sleep and depression are also common in narcolepsy (Aldrich, 1998; Siegel, 1999). One element of narcolepsy that appears to be absent in PD is cataplexy.
Human narcolepsy is caused by a loss of hypocretin (Hcrt) neurons (Peyron et al., 2000; Thannickal et al., 2000a; Thannickal et al., 2000b, 2003). Measurement of Hcrt in the CSF of PD patients has produced inconsistent results. Some studies have reported abnormally low levels, whereas others have reported values in the normal range (Mignot et al., 2002; Overeem et al., 2002; Drouot et al., 2003; Yasui et al., 2006). We have reported that Hcrt levels rise by as much as 100% when dogs or cats play, as compared to levels in quiet waking (Kiyashchenko et al., 2002; Wu et al., 2002). These findings and other similar findings suggest that any reduction in Hcrt level in PD may be secondary to the reduced movement caused by PD, rather than resulting from primary pathology of the Hcrt system. It has been speculated that the loss of dopamine neurons may be responsible for the sleepiness symptoms of PD (Dzirasa et al., 2006), but this does not appear to explain the early onset of these symptoms nor their striking similarity to those of narcolepsy. Only by examining the Hcrt system directly can we determine if Hcrt cells are damaged in PD.
Materials and methods
The hypothalamus of 11 PD (mean age 79±4) and 5 normal (mean age 77±3) brains was examined (Table 1). Details of the sleep quality of the PD patients and controls were not available, although other reports cited above demonstrate that a high percentage of PD patients have sleep abnormalities. Brains were fixed in 10% buffered formalin containing 0.1M phosphate buffer (pH=7.4). The hypothalamus was cut into 40 μm sections. Sections were immunostained for hypocretin (Hcrt-1), melanin concentrating hormone (MCH), alpha synuclein and glial fibrillary acidic protein (GFAP). The substantia nigra of 10 PD brains and 7 normal brains were used for the study of neuromelanin pigmented cell loss. The severity of Parkinson’s disease was assessed using the Hoehn and Yahr scale (Hoehn and Yahr, 2001). The level of neuropathology was assessed using the Braak staging criteria (Braak et al., 2003). Cell number, distribution and size were determined with stereology techniques on a one in eight series. All values are reported as mean and SEM. Comparisons were made using the t-test.
Table 1.
Clinical data of Parkinson’s and control subjects, and characteristics of Hcrt and MCH cells
| Subjects | Age | Sex | No of Hcrt cell | Hcrt cell size area (μm2) | No of MCH cells | MCH cell size area (μm2) | Clinical diagnosis | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Controls | |||||||||||
| C–A | 61 | M | 65833 | 408.48 | 131616 | 409.15 | Pneumonia, testicular tumor | ||||
| C–B | 78 | M | 68932 | 312.41 | 123332 | 407.95 | Non neuronal | ||||
| C–C | 81 | F | 83330 | 355.67 | 138425 | 388.65 | Cerebrovascular | ||||
| C–D | 85 | F | 79219 | 309.71 | 130840 | 346.69 | Cancer — adenocarcinoma | ||||
| C–E | 82 | M | 80632 | 379.42 | 137325 | 436.34 | Cerebrovascular | ||||
| C–F | 73 | F | only substantia nigra | Breast cancer | |||||||
| C–G | 80 | M | only substantia nigra | Cancer – renal | |||||||
| Parkinson’s | Clin. Stage (Hoehn and Yahr, 2001) | Path. Stage (Braak et al., 2003) | Duration (years) | Medications | |||||||
| PD-A | 85 | F | 58124 | 340.07 | 120250 | 338.30 | PD | I | 3 | 20 | n/a- |
| PD-B | 68 | M | 59728 | 347.44 | 98924 | 336.85 | PD, dysphagia, dementia | II | 4 | 23 | Sinemet, bromocriptine, dopamine. |
| PD-C | 77 | M | 55505 | 318.68 | 110555 | 392.45 | PD, Alzheimer’s | II | 3 | 18.5 | bromocriptine, Sinemet, Parlodel. |
| PD-D | 70 | F | 53374 | 347.39 | 79582 | 440.46 | PD | III | 3 | 4 | Sinemet |
| PD-E | 81 | M | 44266 | 306.61 | 69732 | 373.01 | PD | III | 4 | 13 | Sinemet, Parlodel |
| PD-F | 62 | M | 46800 | 304.49 | 71466 | 335.61 | PD | IV | 3 | 5 | n/a |
| PD-G | 90 | M | 45600 | 360.45 | 77600 | 413.50 | PD, Alzheimer’s | IV | 4 | 5 | Sinemet |
| PD-H | 97 | F | 39642 | 297.49 | 62329 | 330.46 | PD, basilar vasculature | IV | 4 | 12 | Sinemet |
| PD-I | 62 | M | 29716 | 335.98 | 78571 | 341.44 | PD | IV | 4 | 9 | Sinemet, Permax |
| PD-J | 103 | F | 25866 | 342.41 | 32400 | 401.08 | PD | V | 6 | 21 | n/a |
| PD-K | 67 | M | 31742 | 345.47 | 40176 | 415.48 | PD, strokes | V | 5 | 19 | Sinemet, Eldepryl, Requip, Mirapex, Provigil |
| PD-L | 86 | F | only substantia nigra | PD | II | n/a | 11 | Sinemet, Permax, Eldepryl | |||
Note: Clin.=clinical, Path.=pathological.
Hcrt, MCH and alpha synuclein immunohistochemistry
The sections were treated with 0.5% sodium borohydride in PBS for 30 min and washed with PBS, and then incubated for 30 min in 0.5% H2O2 for blocking of endogenous peroxidase activity. For antigen retrieval, sections were heated for 30 min at 80°C in a water bath with 10mM sodium citrate (pH 8.5) solution. The sections were cooled to room temperature in sodium citrate and washed with PBS. Water bath heating produces less tissue damage and more uniform antigen retrieval than other heating techniques (Jiao et al., 1999). After thorough washing with PBS the sections were placed for 2 h in 1.5% normal goat serum in PBS and incubated for 72 h at 4°C with a 1 : 2000 dilution of Hcrt-1 (Orexin-A, Calbiochem, San Diego, CA). Sections were then incubated in a secondary antibody (biotinylated goat anti-rabbit IgG; Vector Laboratories, Burlingame, CA) followed by avidin–biotin peroxidase (ABC Elite Kit; Vector laboratories), for 2 h each at room temperature. The tissue-bound peroxidase was visualized by a diaminobenzidine reaction (Vector laboratories). Adjacent series of sections were immunostained for MCH (1 : 20 000, polyclonal rabbit anti-melanin concentrating hormone, Phoenix Pharmaceuticals, Inc., Belmont, CA). Pretreatment and staining was carried out as described for Hcrt staining. Another series of one in twenty-four sections were used for α-synuclein staining (1 : 10000, mouse anti-alpha synuclein monoclonal antibody, Chemicon International, Temecula, CA). Sections were then incubated in a secondary antibody (biotinylated goat anti-mouse IgG; Vector Laboratories) followed by avidin–biotin peroxidase (ABC Elite Kit; Vector laboratories), for 2 h each at room temperature. The tissue-bound peroxidase was visualized by a diaminobenzidine reaction (Vector Laboratories).
Double immunolabelling
After antigen retrieval treatment, sections immunochemically stained for orexin and α-synuclein were incubated with a mixture of primary antibodies for orexin-A (1 : 2000) and α-synuclein (1 : 10 000) for 72 h at 4°C. After being rinsed, sections were sequentially incubated in biotinylated goat anti-mouse IgG (Vector Laboratories) for α-synuclein or biotinylated goat anti-rabbit IgG (Vector Laboratories) for Orexin A and followed by avidin–biotin peroxidase (ABC Elite Kit; Vector Laboratories) for 2 h at room temperature. The final product of α-synuclein was visualized with nickel-DAB solution (Vector Laboratories). The color of α-synuclein immunohistochemical products was black. The hypocretin immunohistochemical products were visualized with DAB, which had a yellow–brown colour.
GFAP immunohistochemistry
For GFAP staining, sections were immunostained with a 1:2000 dilution of primary polyclonal rabbit anti- GFAP antibody (DAKO, Carpinteria, CA). Antigen retrieval was not required for GFAP staining. After a hydrogen peroxide treatment and blocking serum, the sections were immunostained with GFAP antibody followed by biotinylated goat anti-rabbit secondary antibody, and an avidin–biotin–HRP complex (Vectastain ABC kit, Vector Laboratories). Incubation times were 24 h (at 4°C) for the primary antibody, 30 min (at room temperature) for the secondary antibody, and 1 h (at room temperature) for the avidin–biotin–HRP complex. Sections were treated with the DAB reaction (Vector Laboratories).
Immunohistochemistry in substantia nigra
Substantia nigra of 10 PD brains and 7 neurologically normal brains were used (Table 1). The substantia nigra were cut into 40-μm thick coronal sections. Haematoxylin and eosin (FD Neurotechnologies Inc, Baltimore, MD) staining were used for the identification of neuromelanin pigmented cells. A one in twenty-four series of sections was stained for GFAP and alpha synuclein immunohistochemistry, with the same procedure used for the hypothalamic sections.
Control sections from each brain were processed without the primary antibody and did not show staining. Brain regions and nuclei were identified using the ‘Atlas of the Human Brain’ (Mai et al., 2004). Digital image acquisition was carried out with a Micro Fire camera (Optronics, Goleta, CA) and imported to the Corel Draw program. Contrast and brightness were corrected.
Quantitative analysis
Hcrt and MCH cell number and distribution were determined with stereological techniques on a one in eight series of sections through the complete hypothalamus. We used a Nikon E600 microscope with three axis motorized stage, video camera, Neurolucida interface and Stereoinvestigator software (MicroBrightfield Corp., Colchester, Vermont). To find out whether alpha synuclein was colocalized with either Hcrt or MCH cells, we used Neurolucida mapping of the double immunolabelled sections.
The density of GFAP cells in the thalamus and posterior hypothalamus was calculated as the number of cells per unit area (mm2). After delineating the nucleus, we used 250 × 250 μm as the counting frame size for random sampling with stereological procedures. All values of each nucleus were calculated for each subject. These were pooled to give means and SEM for each region and each group.
To calculate the percentage loss of neuromelanin pigmented cells in the substantia nigra, we used Neurolucida mapping of each section stained with haematoxylin and eosin. The numbers of neuromelanin pigmented cells of PD brains were compared with matching sections of normals and the percentage loss was calculated.
The ‘nucleator probe’ in the Stereology program was used to estimate the mean cross-sectional area of the Hcrt, MCH and neuromelanin pigmented cells. Neurons with a clear nucleus were chosen for analysis. The nucleator probe was used with the optical fractionator and stereology procedures for systematic random sampling to identify cells (Gundersen, 1988). In the sampling results, the volume estimate associated with each cell was displayed, along with the average volume for the group of cells measured. A total of 606 Hcrt cells from normal (n=5) and 702 cells from PD (n=11) were measured. For MCH a total of 1032 (n=5) from normal and 1109 (n=11) from PD were measured. In the case neuromelanin pigmented cells, 1986 cells from normal (n=7) and 1518 cells from PD (n=10) were measured.
Results
Hcrt and MCH cell loss
We found an increasing loss of hypocretin cells with disease progression (Figs 1 and 2) as measured by the Hoehn and Yahr rating scale (Hoehn and Yahr, 2001). Similarly, there was higher loss of MCH cells with disease severity (Figs 2 and 3). Hcrt and MCH cells were lost throughout the A–P extent of their hypothalamic distributions (Fig. 2C). The percentage loss of Hcrt cells was minimal in stage I (23%) and was maximal in stage V (62%). Similarly, the percentage loss of MCH cells was lowest in stage I (12%) and was highest in stage V (74%). There was a significant increase (P=0.0006, t=4.25, df=15) in the size of neuromelanin containing cells in PD patients as has been reported (Cabello et al., 2002), but no difference in the size of surviving Hcrt (P=0.18, t=1.39, df=14) and MCH (P=0.28, t=1.39, df=14) cells relative to control (Fig. 2B).
Fig. 1.
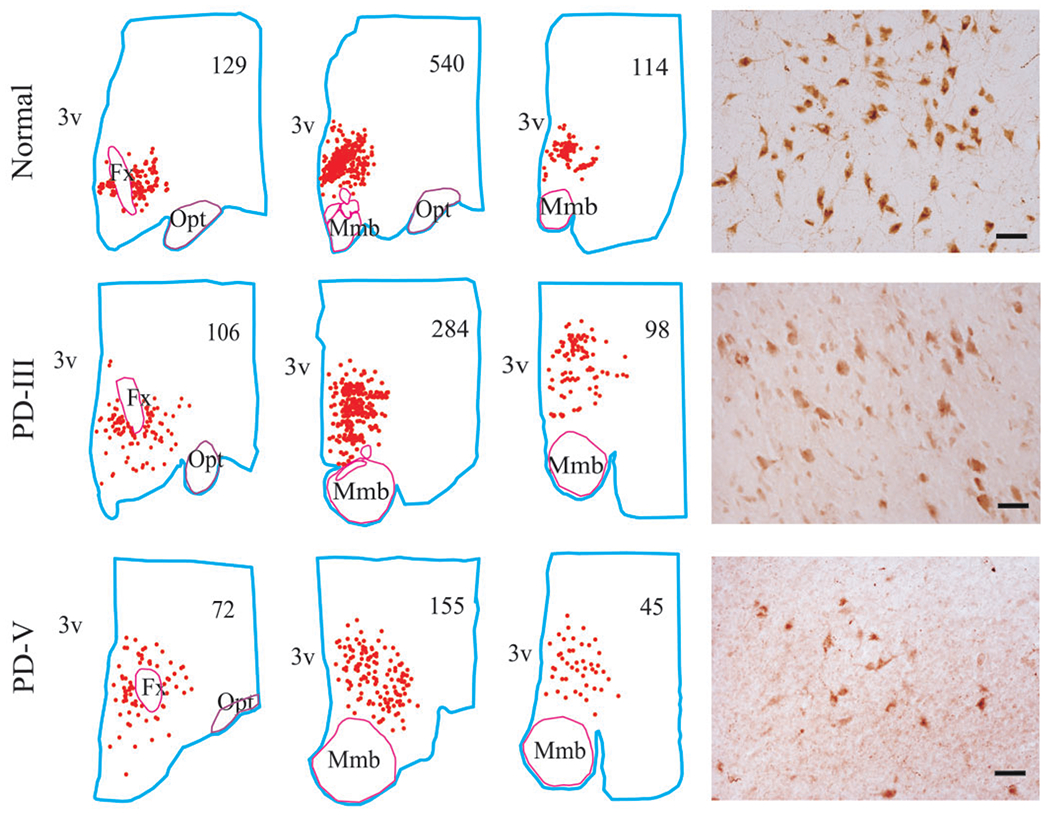
Distribution of Hcrt cells in normal and across PD stages. The clinical stages of PD are based on Hoehn and Yahr criteria. The cell distribution and count from a section of anterior, middle and posterior part of the hypothalamus were mapped from a normal, stage III and stage V of PD brains. The cell counts are listed for each section. The number of Hcrt cells is decreased with severity of the disease. 3v—third ventricle, Fx—fornix, Mmb—mammillary body, Opt—optic tract. Scale bars—50 μm.
Fig. 2.
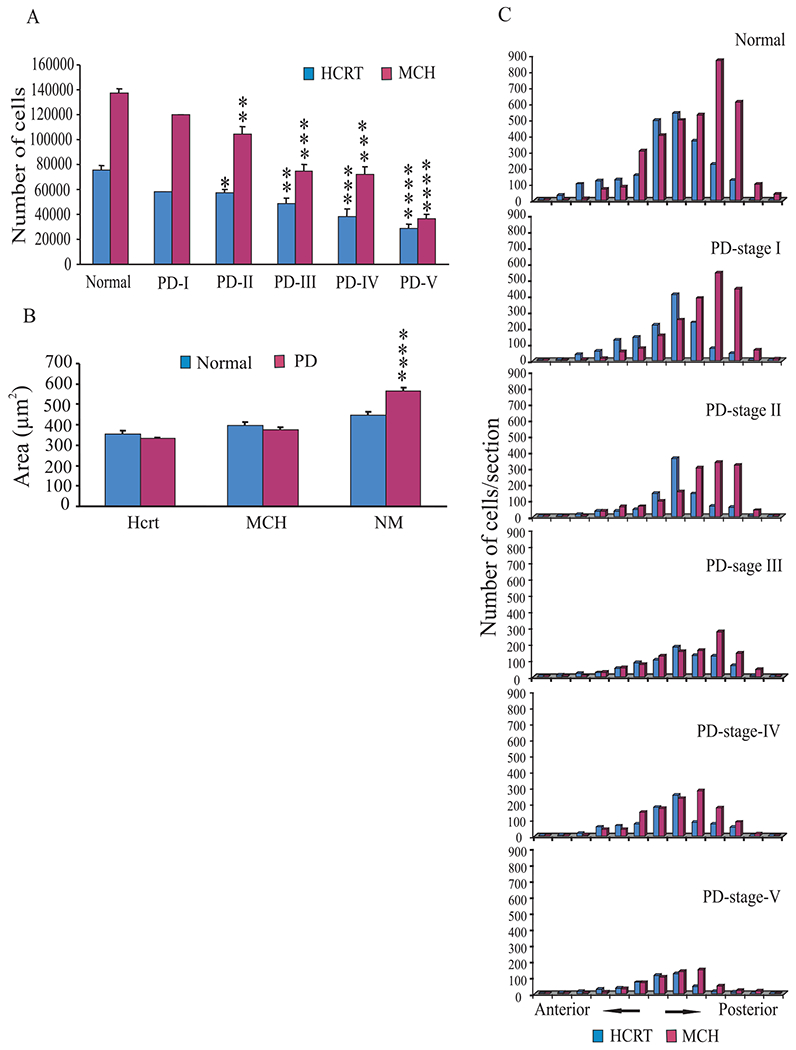
Hcrt and MCH pathology in different stages of PD. (A) The total number of Hcrt and MCH cells in normal and PD-I, PD-II, PD-III, PD-IV and PD-V. The values are compared to cell numbers in the normal brains. (B) The size of the Hcrt, MCH and neuromelanin pigmented cells estimated by nucleator method. Hcrt and MCH cells in PD did not differ in size from those in normal brains. Neuromelanin pigmented cells showed hypertrophy (27%) compared with normal cells. (C) Hcrt and MCH cells were mapped in individual sections from anterior to posterior hypothalamus with 1200 μm section interval. One brain from a normal and one from each stage (Hoehn and Yahr, I–V) of PD were used for Neurolucida mapping. There was a generalized loss of Hcrt and MCH cells with severity of the disease. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001, Student’s t-test.
Fig. 3.
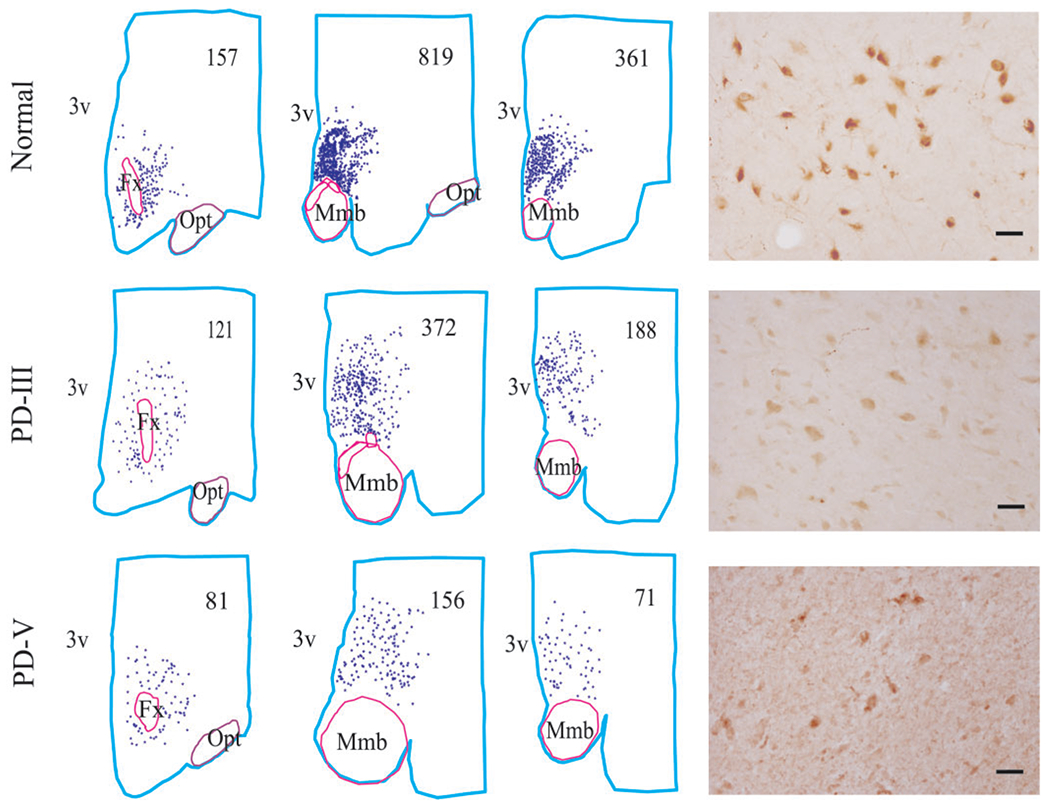
Distribution of MCH cells in normal and Parkinson stages. Cell counts are listed in each section. The number of MCH cell was decreased with severity of the disease. The abbreviations are same as in Fig. 1. Scale bars—50 μm.
Distribution of alpha synuclein, gliosis and neuromelanin pigmented cell loss
Alpha synuclein immunostaining showed a pattern of Lewy body formation in different stages of PD (Fig. 4A). We did not see Lewy bodies in surviving Hcrt (Fig. 4B and D) or MCH cells (Fig. 4C and E), but they were present in surviving neuromelanin containing cells of the substantia nigra (Fig. 4F). We hypothesize that these cells either die by a different mechanism than neuromelanin cells or that they die more rapidly, leaving few in an intermediate state to be observed. There was 50–75% loss of neuromelanin pigmented cells in the substantia nigra (Fig. 5A) compared to control. In the hypothalamus, we saw increasing levels of GFAP with disease progression in PD (Fig. 5B and C).
Fig. 4.
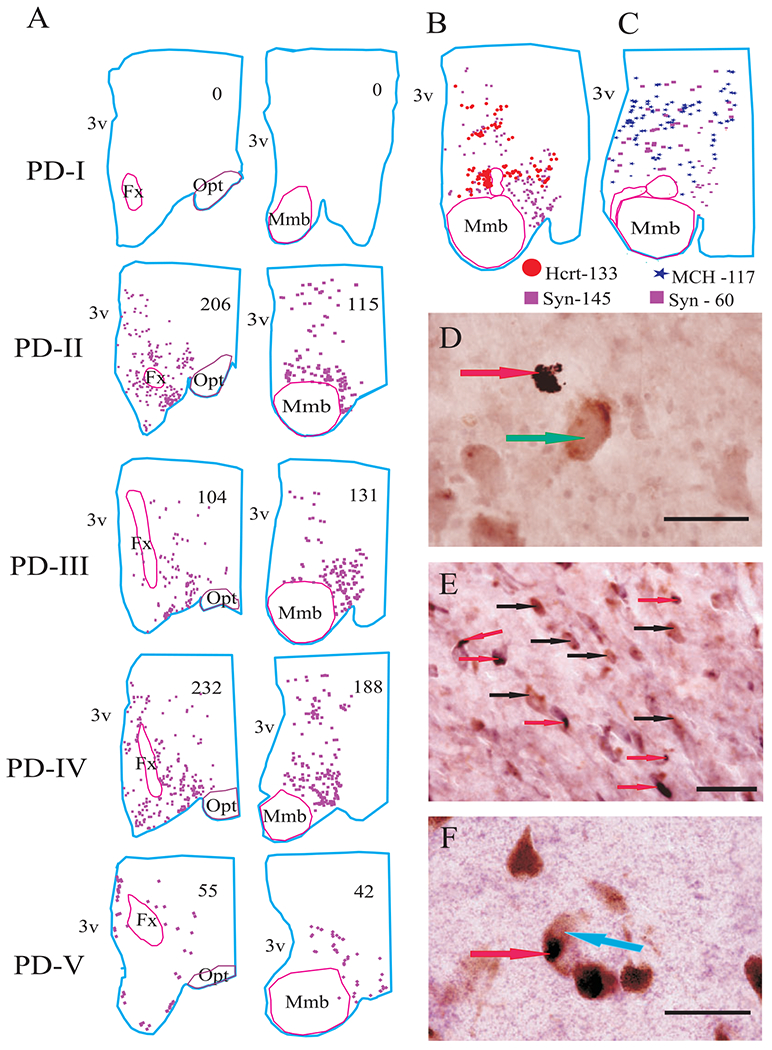
Distribution of alpha synuclein in the hypothalamus in different stages of PD. (A) Neurolucida mapping of alpha synuclein in PD stages with single immunostaining. (B) Mapping of Hcrt and alpha synuclein in double-labelled section. (C) Mapping of MCH and alpha synuclein in double-labelled section. Alpha synuclein was not colocalized with Hcrt and MCH cells (D and E), but it was colocalized with neuromelanin pigmented cells in substantia nigra (F). Arrows: red—alpha synuclein, green—Hcrt cell, black—MCH cells and blue—neuromelanin pigmented cell. Scale bars—50 μm.
Fig. 5.
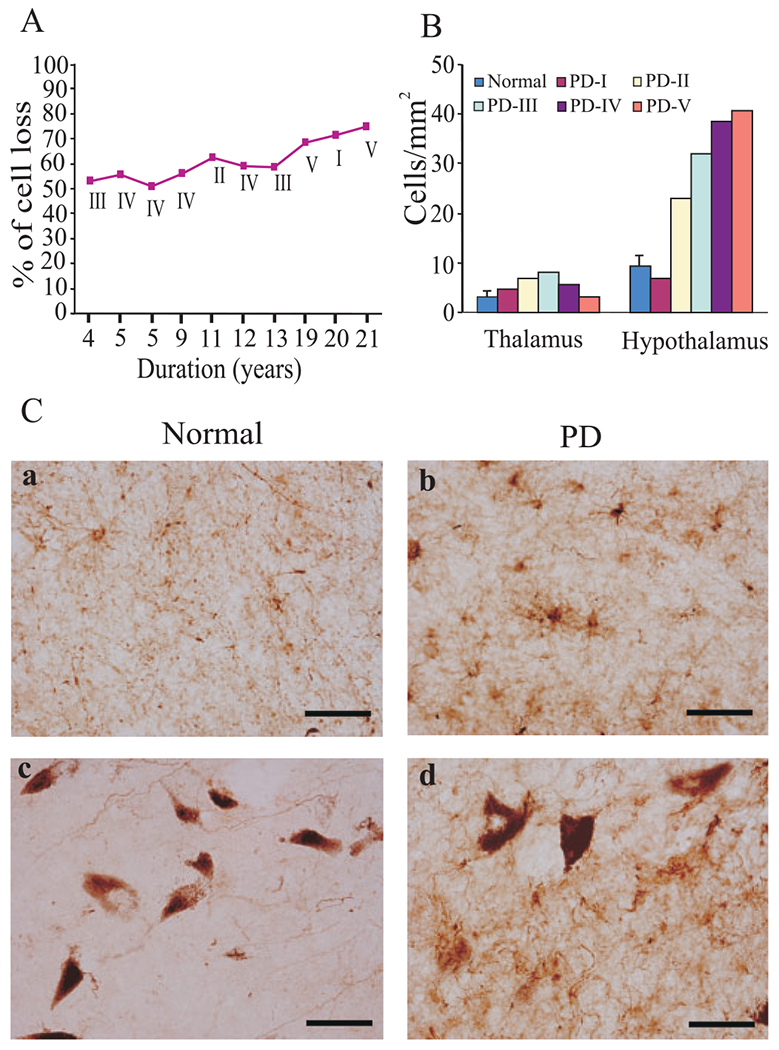
Gliosis and neuromelanin pigmented cell loss in PD. (A) The percentage loss of neuromelanin pigmented cell loss in the substantia nigra was correlated with duration of the disease. (B) The number of glial fibrillary acidic protein-labelled astrocytes (GFAP) in the thalamus and posterior hypothalamus. (C) GFAP in the hypothalamus of normal (a) and PD (b). GFAP density in the substantia nigra of normal (c) and PD (d) brain. The number of GFAP-labelled astrocytes were increased with severity of the disease. Scale bars—50 μm.
Clinicopathological correlations
We used the pathological variables (number of Hcrt, MCH and neuromelanin pigmented cells) and the clinical variables (severity and duration of disease) for the correlation study (Table 2). We found an increasing loss of hypocretin cells with disease progression as measured by the Hoehn and Yahr rating scale (Hoehn and Yahr, 2001). Similarly, MCH cell loss was correlated with disease stage but not with disease duration. In contrast, the loss of neuromelanin pigmented cells was not correlated with disease stage but was with disease duration, extending the conclusions of a recent study which showed that alpha synuclein pathology in neuromelanin cells does not correlate well with PD symptoms (Parkkinen et al., 2005). The Braak stages were correlated with percentage loss of neuromelanin pigmented cells, MCH, Hcrt and the Hoehn and Yahr staging (Table 3).
Table 2.
Correlation analysis of Hcrt, MCH and Neuromelanin pigmented cell loss in PD with clinical stages (Hoehn and Yahr), duration and % cell loss
| Correlations | r | P |
|---|---|---|
| % loss of cells versus PD stages | ||
| Hcrt cells and PD stages | 0.87 | 0.0005 |
| MCH cells and PD stages | 0.96 | 0.0001 |
| NM cells and PD stages | 0.03 | 0.94 |
| % loss of cells versus PD duration | ||
| Hcrt cells and PD duration | 0.35 | 0.28 |
| MCH cells and PD duration | 0.04 | 0.90 |
| NM cells and PD duration | 0.92 | 0.002 |
| % loss of cells versus% loss of cells | ||
| Hcrt cells and NM cells | 0.45 | 0.60 |
| MCH cells and NM cells | 0.25 | 0.58 |
| Hcrt cells and MCH cells | 0.83 | 0.001 |
Note: r=correlation; P=significance; NM=neuromelanin pigmented cells.
Table 3.
Correlation analysis of pathological stages (Braak et al.) in PD with Hcrt, MCH and Neuromelanin pigmented cell loss and duration
| Correlations | r | P |
|---|---|---|
| % loss of Hcrt cells and pathological stages | 0.56 | 0.06 |
| % loss of MCH cells and pathological stages | 0.78 | 0.004 |
| % of NM cells and pathological stages | 0.86 | 0.001 |
| Duration and pathological stages | 0.53 | 0.09 |
| Duration and clinical stages | 0.09 | 0.78 |
| Pathological stages and clinical stages | 0.71 | 0.01 |
Note: r=correlation; P=significance; NM=neuromelanin pigmented cells.
Discussion
The early loss of Hcrt cells may be related to the early appearance of narcolepsy-like signs in PD patients. This loss is occurring prior to the onset of drug treatment in many PD patients. The loss of Hcrt cells may also explain the orthostatic hypotension reported in PD (Hoehn and Yahr, 2001) which parallels the low BP seen in Hcrt null mutant mice (Kayaba et al., 2003) and the abnormal regulation of body temperature that has been reported in both PD (Elliott et al., 1974) and Hcrt null mutant mice (Mochizuki et al., 2006).
The sleepiness experienced by PD patients may not be solely attributable to the loss of Hcrt neurons. It may be at least partially due to the other neurodegenerative changes in PD, including the loss of dopamine, norepinephrine and serotonin neurons (Braak et al., 2003, 2004), all of which have alerting properties (Siegel, 1990; Wisor et al., 2001; Aston-Jones and Cohen, 2005; Siegel, 2005). The role of the loss of MCH cells reported here in the sleepiness of PD is unclear. In contrast to the maximal activity of Hcrt cells in waking (Lee et al., 2005; Mileykovskiy et al., 2005), MCH cells appear to be maximally active in sleep and are reciprocally connected with Hcrt neurons (Verret et al., 2003; Alam et al., 2005; Modirrousta et al., 2005; Torterolo et al., 2006). The loss of MCH neurons in PD may therefore alter the expression of symptoms produced by loss of Hcrt neurons, which are selectively lost in narcolepsy.
If the loss of Hcrt cells is responsible for the symptoms common to both disorders, PD’s narcoleptic like symptoms may respond to the same treatments found effective in narcolepsy. Especially promising would be treatment with hypocretin or hypocretin analogs (Stocchi et al., 1998; John et al., 2000, 2003; Siegel and Boehmer, 2006). The significant correlations that we find between the loss of Hcrt and MCH neurons and the clinical stage of PD, in contrast to the lack of a relationship of similar strength between loss of neuromelanin containing cells and the clinical symptoms of PD, suggests a previously unappreciated relationship between hypothalamic dysfunction and the time course of the overall clinical picture of PD (Langston and Forno, 1978; Kremer and Bots, 1993). The demonstrated relation between Hcrt release and mood (Kiyashchenko et al., 2002; Wu et al., 2002; Siegel, 2004; Mileykovskiy et al., 2005; Siegel and Boehmer, 2006) encourages the investigation of therapies targeted at reversing Hcrt dysfunction to treat depression in PD.
Acknowledgements
Tissue specimens were obtained from the Human Brain and Spinal Fluid Resource Center, West Los Angeles Healthcare Center, Los Angeles, CA. Supported by NS14610, HL41370, MH64109, NS42566 (YYL) and the Medical Research Service of the Department of Veterans Affairs. Funding to pay the Open Access publication charges for this article was provided by The National Institutes of Health.
Abbreviations:
- GFAP
glial fibrillary acidic protein
- Hcrt
hypocretin
- MCH
melanin concentrating hormone
- PD
Parkinson’s disease
Footnotes
An abstract of this work was submitted to the Society for Neuroscience annual meeting on May 15, 2006 and presented at the Society for Neuroscience meeting in Atlanta on October 15, 2006.
References
- Abbott RD, Ross GW, White LR, Tanner CM, Masaki KH, Nelson JS, et al. Excessive daytime sleepiness and subsequent development of Parkinson disease. Neurology 2005; 65: 1442–6. [DOI] [PubMed] [Google Scholar]
- Alam MN, Kumar S, Bashir T, Suntsova N, Methippara MN, Szymusiak R, et al. GABA-mediated control of hypocretin- but not melanin-concentrating hormone-immunoreactive neurones during sleep in rats. J Physiol 2005; 563: 569–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aldrich MS. Diagnostic aspects of narcolepsy. Neurology 1998; 50: S2–7. [DOI] [PubMed] [Google Scholar]
- Arnulf I. Sleep and wakefulness disturbances in Parkinson’s disease. J Neural Transm Suppl 2006; 70: 357–60. [DOI] [PubMed] [Google Scholar]
- Arnulf I, Bonnet AM, Damier P, Bejjani BP, Seilhean D, Derenne JP, Agid Y, et al. Hallucinations, REM sleep, and Parkinson’s disease: a medical hypothesis. Neurology 2000; 55: 281–8. [DOI] [PubMed] [Google Scholar]
- Arnulf I, Konofal E, Merino-Andreu M, Houeto JL, Mesnage V, Welter ML, et al. Parkinson’s disease and sleepiness: an integral part of PD. Neurology 2002; 58: 1019–24. [DOI] [PubMed] [Google Scholar]
- Askenasy JJ. Approaching disturbed sleep in late Parkinson’s Disease: first step toward a proposal for a revised UPDRS. Parkinsonism Relat Disord 2001; 8: 123–31. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G, Cohen JD. Adaptive gain and the role of the locus coeruleus-norepinephrine system in optimal performance. J Comp Neurol 2005; 493: 99–110. [DOI] [PubMed] [Google Scholar]
- Barone P, Amboni M, Vitale C, Bonavita V. Treatment of nocturnal disturbances and excessive daytime sleepiness in Parkinson’s disease. Neurology 2004; 63: S35–8. [DOI] [PubMed] [Google Scholar]
- Benbir G, Ozekmekci S, Cinar M, Beskardes F, Apaydin H, Erginoz E. Features associated with the development of hallucinations in Parkinson’s disease. Acta Neurol Scand 2006; 114: 239–43. [DOI] [PubMed] [Google Scholar]
- Braak H, Del TK, Rub U, De Vos RA, Jansen Steur EN, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging 2003; 24: 197–211. [DOI] [PubMed] [Google Scholar]
- Braak H, Ghebremedhin E, Rub U, Bratzke H, Del TK. Stages in the development of Parkinson’s disease-related pathology. Cell Tissue Res 2004; 318: 121–34. [DOI] [PubMed] [Google Scholar]
- Cabello CR, Thune JJ, Pakkenberg H, Pakkenber B. Ageing of substantianigra in humans: cell loss may be compensated by hypertrophy. Neurobiol Appl Neurobiol 2002; 28: 283–91. [DOI] [PubMed] [Google Scholar]
- Dhawan V, Healy DG, Pal S, Chaudhuri KR. Sleep-related problems of Parkinson’s disease. Age Ageing 2006; 35: 220–8. [DOI] [PubMed] [Google Scholar]
- Drouot X, Moutereau S, Nguyen JP, Lefaucheur JP, Creange A, Remy P, et al. Low levels of ventricular CSF orexin/hypocretin in advanced PD. Neurology 2003; 61: 540–3. [DOI] [PubMed] [Google Scholar]
- Dzirasa K, Ribeiro S, Costa R, Santos LM, Lin SC, Grosmark A, et al. Dopaminergic control of sleep-wake states. J Neurosci 2006; 26: 10577–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott K, Cote LJ, Frewin DB. Vascular responses in the hands of Parkinson’s disease patients. Neurology 1974; 24: 857–62. [DOI] [PubMed] [Google Scholar]
- Frosh WA. Psychiatric Issues in Parkinson’s disease: a practical guide. Am J Psychiatry 2006; 163: 1456–7. [Google Scholar]
- Frucht S Sudden-onset sleep in Parkinson disease. JAMA 2002; 287: 2076–77. [DOI] [PubMed] [Google Scholar]
- Frucht S, Rogers JD, Greene PE, Gordon MF, Fahn S. Falling asleep at the wheel: motor vehicle mishaps in persons taking pramipexole and ropinirole. Neurology 1999; 52: 1908–10. [DOI] [PubMed] [Google Scholar]
- Gagnon JF, Bedard MA, Fantini ML, Petit D, Panisset M, Rompre S, et al. REM sleep behavior disorder and REM sleep without atonia in Parkinson’s disease. Neurology 2002; 59: 585–9. [DOI] [PubMed] [Google Scholar]
- Grandas F, Iranzo A. Nocturnal problems occurring in Parkinson’s disease. Neurology 2004; 63: S8–11. [DOI] [PubMed] [Google Scholar]
- Gunderson HJ. The nucleator. J Microscopy 1988; 151: 3–21. [DOI] [PubMed] [Google Scholar]
- Hoehn MM, Yahr MD. Parkinsonism: onset, progression, and mortality. 1967. Neurology 2001; 57: S11–26. [PubMed] [Google Scholar]
- Jiao Y, Sun Z, Lee T, Fusco FR, Kimble TD, Meade CA, et al. A simple and sensitive antigen retrieval method for free floating and slide mounted tissue sections. J Neurosci Meth 1999; 93: 149–62. [DOI] [PubMed] [Google Scholar]
- John J, Wu M-F, Kodama T, Siegel JM. Intravenously administered hypocretin-1 alters brain amino acid release: an in vivo microdialysis study in rats. J Physiol (Lond) 2003; 548.2: 557–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- John J, Wu MF, Siegel JM. Systemic administration of hypocretin-1 reduces cataplexy and normalizes sleep and waking durations in narcoleptic dogs. Sleep Res Online 2000; 3: 23–8. [PMC free article] [PubMed] [Google Scholar]
- Kayaba Y, Nakamura A, Kasuya Y, Ohuchi T, Yanagisawa M, Komuro I, et al. Attenuated defense response and low basal blood pressure in orexin knockout mice. Am J Physiol Regul Integr Comp Physiol 2003; 285: R581–93. [DOI] [PubMed] [Google Scholar]
- Kiyashchenko LI, Mileykovskiy BY, Maidment N, Lam HA, Wu MF, John J, et al. Release of hypocretin (orexin) during waking and sleep states. J Neurosci 2002; 22: 5282–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kremer HP, Bots GT. Lewy bodies in the lateral hypothalamus: do they imply neuronal loss? Mov Disord 1993; 8: 315–20. [DOI] [PubMed] [Google Scholar]
- Langston JW, Forno LS. The hypothalamus in Parkinson disease. Ann Neurol 1978; 3: 129–33. [DOI] [PubMed] [Google Scholar]
- Lee MG, Hassani OK, Jones BE. Discharge of identified orexin/hypocretin neurons across the sleep-waking cycle. J Neurosci 2005; 25: 6716–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mai JK, Assheur J, Paxinos G. Atlas of the human brain. Elsevier Academic Press; 2004. [Google Scholar]
- Mignot E, Lammers GJ, Ripley B, Okun M, Nevsimalova S, Overeem S, et al. The role of cerebrospinal fluid hypocretin measurement in the diagnosis of narcolepsy and other hypersomnias. Arch Neurol 2002; 59: 1553–62. [DOI] [PubMed] [Google Scholar]
- Mileykovskiy BY, Kiyashchenko LI, Siegel JM. Behavioral correlates of activity in identified hypocretin/orexin neurons. Neuron 2005; 46: 787–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mochizuki T, Klerman EB, Sakurai T, Scammell TE. Elevated body temperature during sleep in orexin knockout mice. Am J Physiol Regul Integr Comp Physiol 2006; 291: R533–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Modirrousta M, Mainville L, Jones BE. Orexin and MCH neurons express c-Fos differently after sleep deprivation vs. recovery and bear different adrenergic receptors. Eur J Neurosci 2005; 21: 2807–16. [DOI] [PubMed] [Google Scholar]
- Onofrj M, Luciano AL, Iacono D, Thomas A, Stocchi F, Papola F, et al. HLA typing does not predict REM sleep behaviour disorder and hallucinations in Parkinson’s disease. Mov Disord 2003; 18: 337–40. [DOI] [PubMed] [Google Scholar]
- Overeem S, van Hilten JJ, Ripley B, Mignot E, Nishino S, Lammers GJ. Normal hypocretin-1 levels in Parkinson’s disease patients with excessive daytime sleepiness. Neurology 2002; 58: 498–9. [DOI] [PubMed] [Google Scholar]
- Parkkinen L, Kauppinen T, Pirttila T, Autere JM, Alafuzoff I. Alpha-synuclein pathology does not predict extrapyramidal symptoms or dementia. Ann Neurol 2005; 57: 82–91. [DOI] [PubMed] [Google Scholar]
- Peyron C, Faraco J, Rogers W, Ripley B, Overeem S, Charnay Y, et al. A mutation in a case of early onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains. Nat Med 2000; 6: 991–7. [DOI] [PubMed] [Google Scholar]
- Priano L, Albani G, Brioschi A, Guastamacchia G, Calderoni S, Lopiano L, et al. Nocturnal anomalous movement reduction and sleep microstructure analysis in parkinsonian patients during 1-night transdermal apomorphine treatment. Neurol Sci 2003; 24: 207–8. [DOI] [PubMed] [Google Scholar]
- Rye DB. Excessive daytime sleepiness and unintended sleep in Parkinson’s disease. Curr Neurol Neurosci Rep 2006; 6: 169–76. [DOI] [PubMed] [Google Scholar]
- Savitt JM, Dawson VL, Dawson TM. Diagnosis and treatment of Parkinson disease: molecules to medicine. J Clin Invest 2006; 116: 1744–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schenck CH, Mahowald MW. Motor dyscontrol in narcolepsy: rapid-eye-movement (REM) sleep without atonia and REM sleep behavior disorder. Ann Neurol 1992; 32: 3–10. [DOI] [PubMed] [Google Scholar]
- Siegel JM. Mechanisms of sleep control. J Clin Neurophysiol 1990; 7: 49–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siegel JM. Narcolepsy: a key role for hypocretins (orexins). Cell 1999; 98: 409–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siegel JM. Hypocretin (orexin): role in normal behavior and neuropathology. Annu Rev Psychol 2004; 55: 125–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siegel JM. REM sleep. In: Kryger MH, Roth T, Dement WC, editors. Principles and practice of sleep medicine. Philadelphia: Elsevier Saunders; 2005. p. 120–35. [Google Scholar]
- Siegel JM, Boehmer LN. Narcolepsy and the hypocretin system-where motion meets emotion. Nat Clin Pract Neurol 2006; 2: 548–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stocchi F, Barbato L, Nordera G, Berardelli A, Ruggieri S. Sleep disorders in Parkinson’s disease. J Neurol 1998; 245 (Suppl 1): S15–8. [DOI] [PubMed] [Google Scholar]
- Thannickal TC, Moore RY, Aldrich M, Albin R, Cornford M, Siegel JM. Human narcolepsy is linked to reduced number, size and synaptic bouton density in hypocretin-2 labeled neurons. Abstr Soc Neurosci 2000a; 26: 2061. [Google Scholar]
- Thannickal TC, Moore RY, Nienhuis R, Ramanathan L, Gulyani S, Aldrich M, et al. Reduced number of hypocretin neurons in human narcolepsy. Neuron 2000b; 27: 469–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thannickal TC, Siegel JM, Moore RY. Pattern of hypocretin (orexin) soma and axon loss, and gliosis, in human narcolepsy. Brain Pathol 2003; 13: 340–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torterolo P, Sampogna S, Morales FR, Chase MH. MCH-containing neurons in the hypothalamus of the cat: searching for a role in the control of sleep and wakefulness. Brain Res 2006; 1119: 101–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verret L, Goutagny R, Fort P, Cagnon L, Salvert D, Leger L, et al. A role of melanin-concentrating hormone producing neurons in the central regulation of paradoxical sleep. BMC Neurosci 2003; 4: 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wisor JP, Nishino S, Sora I, Uhl GH, Mignot E, Edgar DM. Dopaminergic role in stimulant-induced wakefulness. J Neurosci 2001; 21: 1787–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu MF, John J, Maidment N, Lam HA, Siegel JM. Hypocretin release in normal and narcoleptic dogs after food and sleep deprivation, eating, and movement. Am J Physiol Regul Integr Comp Physiol 2002; 283: R1079–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasui K, Inoue Y, Kanbayashi T, Nomura T, Kusumi M, Nakashima K. CSF orexin levels of Parkinson’s disease, dementia with Lewy bodies, progressive supranuclear palsy and corticobasal degeneration. J Neurol Sci 2006; 250: 120–3. [DOI] [PubMed] [Google Scholar]