

Abstract
Ischemic and traumatic insults to the central nervous system account for most serious acute and fatal brain injuries and are usually characterized by primary and secondary damage. Secondary damage presents the greatest challenge for medical staff; however, there are currently few effective therapeutic targets for secondary damage. Homer proteins are postsynaptic scaffolding proteins that have been implicated in ischemic and traumatic insults to the central nervous system. Homer signaling can exert either positive or negative effects during such insults, depending on the specific subtype of Homer protein. Homer 1b/c couples with other proteins to form postsynaptic densities, which form the basis of synaptic transmission, while Homer1a expression can be induced by harmful external factors. Homer 1c is used as a unique biomarker to reveal alterations in synaptic connectivity before and during the early stages of apoptosis in retinal ganglion cells, mediated or affected by extracellular or intracellular signaling or cytoskeletal processes. This review summarizes the structural features, related signaling pathways, and diverse roles of Homer proteins in physiological and pathological processes. Upregulating Homer1a or downregulating Homer1b/c may play a neuroprotective role in secondary brain injuries. Homer also plays an important role in the formation of photoreceptor synapses. These findings confirm the neuroprotective effects of Homer, and support the future design of therapeutic drug targets or gene therapies for ischemic and traumatic brain injuries and retinal disorders based on Homer proteins.
Keywords: brain injury, calcium signaling, cerebral ischemia, dendritic spine, glutamate receptor, Homer scaffolding protein, neuron, neuroprotection, retinal ganglion cell, review, traumatic brain injury
Introduction
An investigation into the global burden of diseases identified neurological injuries and diseases, such as acute brain injury and neurodegenerative diseases, as significant threats to humans (Wang, 2017). The main acute brain injuries are ischemic and traumatic brain injuries, both of which are associated with high morbidity and mortality. The pathology of acute brain injury progresses from primary to secondary damage. It is usually difficult to prevent the primary brain injury, which is largely determined by external forces or other primary disease, and the roles of medical staff and researchers are thus to mitigate secondary brain damage, including brain edema, ischemic injuries, epilepsy, and cognitive disorders (Baratz et al., 2015; Hausott and Klimaschewski, 2019; Ferrara et al., 2020; Rauen et al., 2020). However, the unclear pathophysiological mechanisms underlying secondary brain damage and the poor efficacy of neuroprotective factors mean that there are currently no reliable and effective treatments. It is therefore necessary to elucidate the pathophysiological mechanisms responsible for these fatal conditions.
Homer proteins may play important roles in the development of secondary brain damage after acute brain injury. The Homer protein family includes three main members (Homer1, Homer2, Homer3), each encoded by several transcript variants due to alternative mRNA splicing. Homer proteins are divided into short (Homer1a (Vesl-1s), 2c and d, and 3c) and long Homers (Homer1b–d, 2a and b, and 3a and b), according to the length of the amino acid chains (Reibring et al., 2020). The most extensively researched short Homer, Homer1a, was first discovered and identified as an immediate-early gene (IEG) normally induced by neuronal activation (Brakeman et al., 1997; Kato et al., 1997). In contrast, long Homers form vital components of postsynaptic densities (PSDs) and are constitutively expressed at the postsynaptic zones of neurons, where they form protein complexes mediating downstream intracellular signaling pathways. Accumulating evidence suggests that regulation of Homer expression, especially Homer1, could exert a biological role in many central nervous system pathologies, and may thus act as a potential novel therapeutic target. This review summarizes recent advances in our understanding of the roles of Homer proteins in ischemic and traumatic brain injuries and retinal disorders.
Search Strategy
We searched the PubMed database for papers describing Homer proteins published from 1946 to 2021 using the sequential queries shown in Figure 1. We screened the results of each step by abstract and title and excluded non-Homer-related experiments and reviews. We then reviewed the roles of Homer proteins in physiological and pathological processes based on the selected publications.
Figure 1.
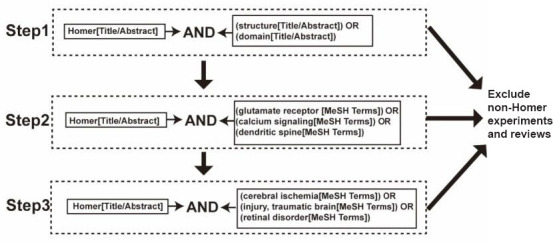
Flowchart for literature retrieval.
Structural Characteristics of Homer Proteins
All Homer proteins share a common amino-terminal domain, called the enabled/vasodilator-stimulated phosphoprotein homology 1 (EVH1) domain (Buonaguro et al., 2020). Long Homers also have a carboxy-terminal coiled-coil (CC) domain and leucine zipper (Lzip) motif (Hayashi, 2019), which are lacking in short Homers (Wang et al., 2020).
The EVH1 domain can bind to the proline-rich sequence PPXXF in various proteins, such as metabotropic glutamate receptors (mGluRs), inositol trisphosphate receptors (IP3R), transient receptor potential canonical (TRPC) channels, drebrin, and dynamin3 (Hassani and Kreienkamp, 2018; Orgovan et al., 2019; Chen et al., 2020; Luo et al., 2020; Ahumada-Castro et al., 2021). In addition, the CC domain mainly mediates self-assembly or multimerization among long Homers (Figure 2) to form dimers or polymers, and the EVH1 domains of these resulting multimers then interact with other target proteins with P motifs. The presence of long Homers at the postsynaptic area thus results in the creation of diverse protein complexes, which in turn facilitate intracellular signal transduction. In contrast, short Homers lack the carboxy-terminal domains required to form multimers, but tend to compete by interacting with the target proteins of long Homers via their EVH1 domains. For instance, the first-discovered Homer protein, Homer1a, is a short-type Homer that disrupts the clustering of postsynaptic molecules mediated by long Homers and plays a dominant-negative role in regulating related signaling pathways (Table 1). Moreover, Lzip motifs are also involved in the multimerization of Homer1c, mGluR1, and N-methyl-D-aspartate (NMDA) receptors (NMDARs) (Osmankovic et al., 2018).
Figure 2.
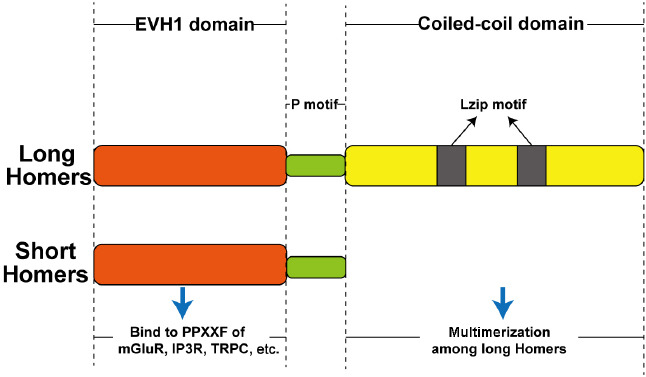
Primary structures of Homer proteins.
All Homer proteins share a common enabled/vasodilator-stimulated phosphoprotein homology 1 (EVH1) domain at their amino terminal, as well as a carboxy-terminal coiled-coil (CC) domain and leucine zipper (Lzip) motif. The EVH1 domain binds the proline-rich sequence PPXXF in various proteins, such as metabotropic glutamate receptors (mGluR), inositol trisphosphate receptors (IP3R), and transient receptor potential canonical (TRPC) channels. The CC domain mainly mediates self-assembly or multimerization among long Homers.
Table 1.
Homer protein family structural characteristics
| Homers | Homer subtypes | Domain composition | Main function |
|---|---|---|---|
| Long homers | Homer1b–d, Homer2a/b, Homer3a/b | Amino terminal: enabled/vasodilator-stimulated phosphoprotein (Ena/VASP) homology 1 (EVH1) domain carboxy-terminal: coiled-coil (CC) domain |
Form diverse protein complexes; Facilitate intracellular signaling transduction |
| Short homers | Homer1a, Homer2c/d, Homer3c | Amino-terminal: enabled/vasodilator-stimulated phosphoprotein (Ena/VASP) homology 1 (EVH1) domain | Disrupt diverse protein complexes; Regulate intracellular signaling transduction |
Homer Signaling in Nerve Cells
Protein function is determined by its structure. Homer-related signaling depends largely on upstream and downstream signaling by target proteins that bind to Homer proteins. We classified Homer-associated signaling into three categories: glutamate receptor signaling, calcium signaling, and dendritic spine morphogenesis signaling. These three types of signaling are inter-related (Figure 3), and are closely involved in secondary brain damage after ischemic and traumatic insults. For example, glutamate receptor signaling contributes to the formation of brain edema after cerebral ischemia (Shi et al., 2017; Sladojevic et al., 2020); calcium signaling plays an important role in TBI-induced inflammation (Wofford et al., 2019); and dendritic spine morphogenesis is a key factor in neurite outgrowth and synapse recovery following ischemic and traumatic insults (Yang et al., 2020; Zhao et al., 2021). Homer proteins can thus have major impacts on these types of damage via regulation of the associated signaling pathways.
Figure 3.
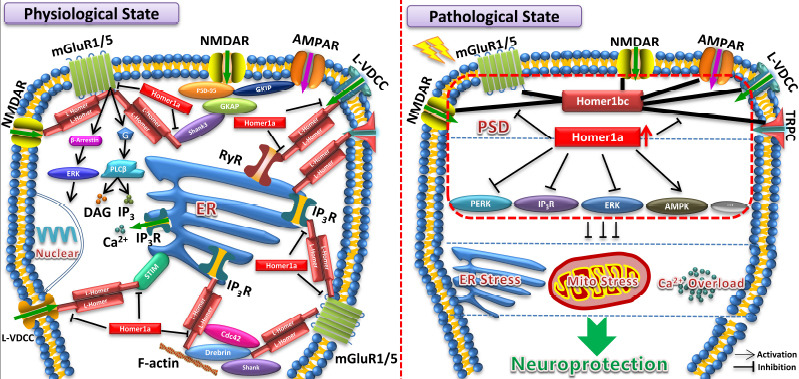
Homer signaling underlying physiological and pathological states.
In the physiological state, long Homers mediate the formation of various protein complexes with N-methyl-D-aspartate receptors (NMDARs), metabotropic glutamate receptors 1/5 (mGluR1/5), α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPAR), L-voltage-dependent calcium channels (L-VDCC), transient receptor potential canonical (TRPC) channels, Shank3, ryanodine receptors (RyR), inositol trisphosphate receptors (IP3R), stromal interaction molecule (STIM), and drebrin. Conversely, Homer1a inhibits this process and regulates glutamate receptor, calcium, and dendritic spine morphogenesis signaling. In pathological states, induced Homer1a facilitates cell survival through disrupting Homer1b/c-mediated protein complexes at the postsynaptic density (PSD), inhibiting protein kinase-like ER kinase (PERK), IP3R, and extracellular signal-regulated kinase (ERK) pathways, and activating the AMP-activated protein kinase (AMPK) pathway. Homer1a also exerts neuroprotection by mitigating endoplasmic reticulum stress (ERS), mitochondrial stress, and Ca2+ overload. DAG: diacylglycerol; GKAP: guanylate kinase-associated protein; GKIP: cGMP-dependent protein kinase interacting protein.
Role of Homer in glutamate receptor signaling
The excitatory neurotransmitter glutamate exerts a crucial role in determining neuronal activity. After release from the presynaptic membrane, glutamate binds to glutamate receptors at the postsynaptic membrane. Glutamate receptors include ionotropic glutamate receptors (iGluRs) and mGluRs. Of these iGluRs, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, kainate receptors, and NMDARs are responsible for the induction of action potentials. As G protein-coupled receptors, mGluRs can be divided into three subgroups (І, II and III) and eight subtypes (mGluR1-mGluR8) on the basis of intracellular signaling, of which group I mGluRs, including mGluR1 and mGluR5, are the most extensively studied (Petzold et al., 2021). Long Homer proteins act as important scaffold proteins through binding glutamate receptors and other proteins. First, as constituent molecules of PSDs, Homer proteins bind proline-rich motifs in group I mGluRs, IP3Rs, Shank, and NMDARs and couple these proteins in a signaling complex (O’Neill et al., 2018; Schaffler et al., 2019; Petzold et al., 2021; Docampo and Huang, 2021). Shank can also function as a scaffold protein linking NMDARs and PSD-95 (Jin et al., 2021). Homer proteins may thus mediate the linkage of mGluRs and NMDARs via direct binding of NMDARs or indirect binding of Shank. Moreover, the Homer-dependent mGluR-NMDAR link has been shown to have a bearing on synaptic potentiation (Sylantyev et al., 2013). Homer proteins may play an essential role in bridging glutamate transmission and intracellular signaling transduction, and downregulation of Homer1b/c was previously shown to decrease extracellular signal-regulated kinase (ERK) phosphorylation mediated by mGluR5, suggesting that Homer1b/c forms a central signaling pathway linking mGluR5 to ERK (May et al., 2021).
In contrast to long Homer proteins, short Homer proteins, particularly Homer1a, play a key role in regulating synaptic glutamate receptor signaling. Homer1a, which lacks a CC domain, functions as a dominant-negative factor via competitively binding to receptors with long Homers. For example, upregulating endogenous Homer1a by applying brain-derived neurotrophic factor or a proteasome inhibitor resulted in the reduction of clusters of Homer1c and F-actin (Zuo et al., 2020). Moreover, Homer1a modulated mGluR–NMDAR communication by disrupting the Homer-Shank3 interaction (Zhu et al., 2020). Second, Homer1a constitutively activated group I mGluRs including mGluR1a and mGluR5, independent of an agonist (Martin et al., 2019), and affected homeostatic scaling by regulating group I mGluR activation and contributing to neuronal excitability and neural plasticity (Witharana et al., 2019). Homer1a also affected long-term potentiation in hippocampal CA1 pyramidal neurons by decreasing AMPA/NMDA current ratios, which was dependent on uncoupling the binding between Homer1b/c and the AMPA receptor subunit GluA2 (Rozov et al., 2012). As indicated above, activity-dependent Homer1a is essential for excitatory synaptic transmission via regulation of glutamate receptor signaling.
Role of Homer in calcium signaling
Homer proteins play an essential role in regulating calcium homeostasis by binding calcium signaling proteins, such as mGluRs, TRPC channels, IP3Rs, ryanodine receptors (RyRs), and some L-type calcium channels.
Homers may contribute to intracellular calcium signaling through glutamate receptor signaling. Group I mGluRs activated phospholipase C (PLC) β, leading to activation of a diacylglycerol-mediated protein kinase C pathway and calcium signaling, including the opening of membrane calcium channels and the release of calcium from intracellular calcium stores (Mahato et al., 2018). Homer proteins can bind to mGluRs, resulting in calcium signaling changes; however, the effects of Homer proteins on mGluR-mediated calcium signaling depend on the cell type. For example, long Homers cross-linked group I mGluRs and the intracellular calcium pool depending on IP3Rs in rat superior cervical ganglion sympathetic neurons, while Homer1a disrupted this coupling and caused calcium release from intracellular stores (Sato et al., 2020). In addition, overexpression of Homer1b caused the translocation of IP3Rs accompanied by increased endoplasmic reticulum (ER) calcium ATPase, calreticulin, and calbindin (Lee et al., 2021). In contrast, upregulating Homer1a reduced group I mGluR-induced intracellular calcium responses in rat autaptic hippocampal cultures (Shan et al., 2018). The reasons why Homer proteins have different impacts on mGluR-mediated calcium signaling in different cells remain unclear, but may be partly explained by the different composite forms of mGluRs and Homers in distinct neurons, as well as the different activities of Homer1a. In addition to mGluR, Homer proteins may affect calcium signaling through other pathways. Long Homers cross-linked TRPC1 and IP3R to form TRPC1–Homer–IP3R complexes, and Homer1a activated calcium influx via blocking this linkage and promoting the opening of TRPC1 channels (Sun et al., 2020). In addition, Homer 1a increased the expression of voltage-dependent calcium channels and enhanced spike-induced calcium increase depending on IP3Rs (Colecraft, 2020). Homer1b/c also reduced the sensitivity of the cell to membrane depolarization-induced calcium elevations by facilitating the interaction between RyR2 and Cav1.2 calcium channels, whereas Homer1a increased the sensitivity by disrupting this interaction (Perni and Beam, 2021). These results suggest that Homer1a acts as an inducer of calcium signaling, while long Homers seem to stabilize the process. Recent evidence suggests that Homer proteins also play an important role in store operated calcium entry (SOCE), mediated by STIM and Orai proteins: Homer1b/c mediated the interactions between stromal interaction molecule 1 (STIM1) and Cav1.2 channels and between Orai1 and TRPC channels, and downregulation of Homer1b/c partially inhibited SOCE (Dionisio et al., 2015; Jia et al., 2017). Disruption of the TRPC–Homer–IP3R complex allowed STIM1 to open TRPC channels (Yuan et al., 2012). Furthermore, we found that Homer1a was also involved in the regulation of SOCE, and overexpression of Homer1a suppressed SOCE by disrupting the STIM1–Oria1 association (Rao et al., 2016).
Role of Homer in dendritic spine morphogenesis
The dendritic spine forms the basis of the synapse, which receives stimulation and transmits impulses into the cell body. Dendritic spine morphogenesis is associated with the formation of learning and memory. In addition to being distributed at the PSD, some Homer proteins are concentrated in the dendritic spine and play an important role in dendritic spine morphogenesis, with different Homer proteins exerting distinctive functions during the process.
Drebrin functions as a crucial molecule in dendritic spine morphogenesis. Long Homers interact with drebrin at the dendritic spine through their EVH1 domains, resulting in the formation of a tetramer (Hayashi, 2019), which facilitates the actin-bundling activity of drebrin resulting in filopodia formation and elongation in neurons (Li et al., 2019). In addition to drebrin, the small GTPase Cdc42 also boosts dendritic spine morphogenesis, and Homer2 contributed to spine morphogenesis by coupling Shank, drebrin, and activated Cdc42 (Pichaud et al., 2019). Based on these two aspects, long Homers can enhance dendritic spine morphogenesis by mediating the interaction with drebrin and Cdc42.
In contrast, Homer1a may play a negative-dominant role in this process. Homer1a reduced the density and size of dendritic spines in cultured hippocampal neurons (Witharana et al., 2019), and although the exact mechanisms remain to be elucidated, this may be partially attributed to the inhibition of long Homers mediated by Homer1a. Moreover, the complementary effects of long Homers and Homer1a may work as a “molecular switch” to regulate dendritic spine morphogenesis.
Homer Signaling in Ischemic and Traumatic Brain Injury
Homer signaling not only plays an important role in physiological processes, but is also involved in various pathological conditions. Recent studies have indicated that Homer signaling is closely associated with the pathologies and development of acute brain injury, especially ischemic and traumatic brain injuries. Moreover, upregulating or downregulating the expression of Homer1 may have beneficial or detrimental effects, respectively, in secondary brain injury after primary insults from stroke or injuries. Here, we focus on the roles of Homer1 and other related intracellular signaling pathways in these processes.
The process of acute brain injury often consists of primary and secondary brain injuries. Because the primary brain injury is uncontrollable, medical staff are concerned with attenuating secondary brain injury and protecting the brain. It is therefore important to understand the mechanism of secondary brain damage and identify clues for the development of ideal treatment modalities. There are many common pathophysiological changes underlying secondary brain damage, such as apoptosis, oxidative stress, mitochondrial dysfunction, ER stress (ERS), calcium overload, and inflammation. Numerous studies have implicated Homer1 in these processes and have shown that regulation of Homer1 expression can affect the outcome of secondary brain damage. Importantly, upregulating Homer1a or downregulating Homer1b/c can exert neuroprotective roles in the process of secondary brain injury, although the mechanisms differ among experimental models.
Role of Homer in ischemic brain damage
Homer1a expression was shown to increase after ischemic brain injury and may be involved in secondary brain injury. Su et al. (2014) investigated the effect of ischemic brain injury on the expression of Homer1a using quantitative reverse transcriptase-polymerase chain reaction (RT-PCR) in a middle cerebral artery occlusion model, and showed that Homer1a mRNA levels peaked at 1 hour, and high levels were maintained up to 24 hours after cerebral reperfusion in rat brain cortex. Wei et al. (2019) also explored the expression pattern of Homer1a protein in cultured mouse cortical neurons using western blot assay after oxygen and glucose deprivation/reperfusion (OGD/R), and revealed that Homer1a expression increased significantly at 6, 12, 24, and 48 hours after reperfusion. Homer1a expression thus increased after ischemic brain injury at both the transcriptional and translation levels.
The role of Homer1a expression following ischemic brain injury remains unclear; however, accumulating evidence suggests that overexpression of Homer1a confers a neuroprotective function. To clarify the role of Homer1a in ischemic brain injury, Wei et al. (2019) investigated the effects of Homer1a via detecting lactate dehydrogenase, reactive oxygen species production, mitochondrial membrane potential, and ATP levels in neurons in an OGD/R model. Homer1a protected against neuronal ischemia/reperfusion damage via inhibiting ERS to conserve mitochondrial function. This suggests that the mechanism by which Homer1a reduces ERS to preserve mitochondrial function is dependent on the protein kinase-like ER kinase (PERK) pathway (Wei et al., 2019). However, the exact molecular mechanism by which Homer1a affects the PERK pathway and its effect on the ultimate outcome of neurons remains unclear. In addition to its role in neurons, Paquet et al. (2013) reported that a cell permeable Tat-tagged peptide that disrupted mGluR-Homer1b binding reduced OGD-induced apoptosis of primary mouse cortical astrocytes, while Homer1a had the same effect on binding as the peptide. These findings suggest that Homer1a may act as an endogenous neuroprotector. In addition to in vitro and in vivo researches, Zhu et al. (2016) also investigated expression levels of Homer proteins in peripheral blood leukocytes using RT-PCR in a population-based study, and showed that mRNA levels of Homer1 and Homer2, but not Homer3, were positively correlated with the incidence of large-artery atherosclerosis strokes. Zhu et al. (2016) reported that Homer proteins were involved in the pathological process of ischemic brain injury, suggesting a diagnostic potential of Homers in ischemic cerebral diseases.
Importantly, evidence for the neuroprotective effect of Homer1a in ischemic brain damage has mainly come from experiments using primary, embryonic neuronal cultures, and the differences between embryonic and mature neurons indicate the need for further studies to elucidate the role of Homer1a in mature neurons, and to determine the roles of other Homer proteins.
Role of Homer in TBI
Recent studies have placed great emphasis on studying the relationship between Homer1 and TBI. In 2005, Huang et al. (2005) scratched primary rat neurons using a punch device consisting of 28 joined stainless steel blades to simulate traumatic injury in vitro. This traumatic stimulation increased Homer1a expression levels detected by both RT-PCR and western blot, but there was no significant change in Homer1b/c expression. They also found that the increase in Homer1a was positively correlated with increased lactate dehydrogenase, further suggesting that Homer1a is involved in traumatic neuronal injury. Luo et al. (2014) subsequently used a sterile plastic pipette tip to scrape primary mouse cortical neurons to induce traumatic injury in vitro, and showed that traumatic insult upregulated Homer1a at both the transcriptional and translational levels. Furthermore, the results of western blot and immunostaining showed that Homer1a expression was upregulated in the cortex around the lesion in an in vivo weight-drop TBI model (Luo et al., 2014). These findings indicated that Homer1a increased dynamically whereas Homer1b/c stayed steady after TBI.
Accumulating evidence has verified that upregulating Homer 1a or downregulating Homer1b/c has neuroprotective effects. Huang et al. (2012) reported that downregulating Homer1b/c using small interfering RNA protected primary rat cortical neurons against traumatic injury, partially as a result of decreased mGluR1a transfer and calcium influx. Fei et al. (2014) confirmed that downregulation of Homer1b/c attenuated the intracellular calcium concentration and mGluR1a expression and improved neuronal survival after TBI. Importantly however, both these were in vitro studies, and further in vivo studies are required to confirm the neuroprotective role of Homer1b/c downregulation. Luo et al. (2014) accordingly found that overexpressing Homer1a via lentiviral transduction decreased cytotoxicity and cell death in neurons undergoing traumatic injury whereas inhibiting Homer1a exacerbated cell damage, and overexpression of Homer1a by infusing lentiviruses into ipsilateral cortex reduced TBI-induced brain edema and neurological deficits in vivo. These results confirmed that Homer1a acts as an endogenous neuroprotector against TBI.
The neuroprotective molecular mechanisms involved in downregulating Homer1b/c and upregulating Homer1a appear to be closely associated with events such as alleviating calcium overload, glutamate excitotoxity, and oxidative stress. On one hand, neuroprotection conferred by downregulating Homer1b/c is highly dependent on the modulation of ER-associated signaling pathways. In 2012, Chen et al. (2012) found that knockdown of Homer1b/c attenuated glutamate-induced intracellular calcium overload by modulating the group I mGluR–IP3R–ER axis in primary rat cortical neurons. They also reported that downregulating Homer1b/c repressed ERS, including PERK, C/EBP homologous protein, and caspase-12 pathways, and preserved mitochondrial function (Chen et al., 2012). Moreover, Guo et al. (2016) reported that downregulation of Homer1b/c attenuated tert-butyl hydroperoxide-induced oxidative stress by inhibiting IP3R/RyR-mediated calcium release and ERS activation in brain endothelial cells.
On the other hand, Homer1a may exert a neuroprotective role through diverse signaling pathways. Luo et al. (2014) showed that Homer 1a protected against traumatic neuronal injury through disruption of mGluR1-ERK signaling by inhibiting the release of intracellular calcium. In addition, Wang et al. (2015) reported that NMDA-induced neuronal injury was more severe in Homer1a-homozygous knockout mice compared with wild-type mice, suggesting that Homer1a may attenuate NMDA excitotoxity by uncoupling NR2B-PSD95-neuronal nitric oxide synthase complexes and inhibiting the membrane distribution of NMDARs, in both in vivo and in vitro studies. Moreover, Luo et al. (2012a) reported that Homer1a prevented hydrogen peroxide-induced oxidative damage by inhibiting the accumulation of reactive oxygen species and mitochondrial apoptosis, and these protective effects were dependent on the regulation of intracellular calcium homeostasis. Wu et al. (2018) reported that Homer1a attenuated hydrogen peroxide-induced oxidative injury by upregulating autophagy by facilitating the phosphorylation of AMP-activated protein kinase.
Important Role of Homer in Retinal Disorders
Role of Homer in structural and functional formation of retina
The retina has evolved to manage different kinds of visual information including brightness, darkness, contrast, color, and motion, by developing a complex network of neurons releasing different neurotransmitters and expressing numerous receptor proteins that mediate their actions (Chakraborty and Pardue, 2015). The ribbon synapses of photoreceptor and bipolar cells involve glutamatergic signals transmitted at a single presynaptic site to several postsynaptic elements, heterogeneously expressing different types of iGluRs and mGluRs. Homer1 has been shown to act as a central organizer at different PSDs in glutamatergic retinal synapses, to bridge different types of glutamate receptors and link them to the cytoskeleton and downstream signaling pathways (Chokshi et al., 2019a, b).
Homer may play an important role in photoreceptor synaptogenesis. Wahlin et al. (2008) investigated changes in photoreceptor presynaptic components during chick embryo retinal development and early post-hatching life using RT-PCR, dissociated retinal cells, laser-capture microdissection, immunocytochemistry, and confocal microscopy. No mRNAs for synaptic molecules such as Homer1 and Homer2 were detected before the onset of photoreceptor synaptogenesis on embryonic day 13, but they became readily detectable thereafter, even before the appearance of visual pigments such as red and green opsins. By using the laser-capture microdissection approach, Homer1 was observed in both the outer nuclear layer and inner retina in samples of retinas on embryonic day 21, while Homer2 was only found in the inner retina. This study showed a very precise timeline of Homer expression during photoreceptor synaptogenesis in the chick retina (Wahlin et al., 2008). Brandstatter et al. (2004) further reported that Homer1 worked as a central organizer at different PSDs in glutamatergic retinal synapses. Using immunocytochemistry and light and electron microscopy, they examined the cellular, synaptic, and postnatal developmental expression of ProSAP1/Shank2 at synapses in the rat retina, and showed that ProSAP1/Shank2 was present postsynaptically at the glutamatergic ribbon synapses of photoreceptor and bipolar cells. However, double-labeling experiments revealed a high rate of co-localization of ProSAP1/Shank2 with Homer1 (Brandstatter et al., 2004). Little is known about the role of Homer1 in the retina, except for the study by Kaja et al. (2003), and Brandstatter et al. (2004) were thus the first to determine the distribution and synaptic localization of Homer1 in the rat retina. In addition, they further showed that ProSAP1 and Homer1 were present postsynaptically in dendrites of ON and OFF bipolar cells and in the processes of horizontal cells of rod and cone photoreceptor ribbon synapses. ProSAP1 and Homer1, mGluR1, mGluR5, and IP3 receptors have also been detected in ON bipolar cell dendrites, where Homer1 might physically and functionally connect with mGluR1 (Li et al., 2020), mGluR5, and IP3Rs to activate their downstream effectors and release intracellular calcium. The high rate of co-localization between ProSAP1 and Homer1 at bipolar cell ribbon synapses in the inner plexiform layer (IPL) thus suggests roles for ProSAP1 and Homer1 as central organizers at different PSDs in glutamatergic retinal synapses (Brandstatter et al., 2004). Homer may also contribute to the functional interplay of pituitary adenylate cyclase activating polypeptide and glutamate, which are co-stored in the rat retinal hypothalamic tract and are involved in photic entrainment of the circadian pacemaker (Prosser et al., 1989). Using quantitative in situ hybridization histochemistry, Nielsen et al. (2001) found that light stimulation of rats early and late at night induced Homer1 gene expression in the suprachiasmatic nucleus (SCN) at time points where light induced phase-delay or phase-advance, respectively (Nielsen et al., 2001). Homer1 mRNA levels in the SCN also displayed modest diurnal variation in a rat brain-slice model, similar to that in vivo, suggesting that the Homer1 gene could be clock-controlled. Interestingly, Homer1 mRNA levels tended to be high at mid-subjective day, coinciding with the peak of neuronal activity of the SCN (Prosser et al., 1989) and expression of the important clock gene, period1 (Per1) (Masis-Vargas et al., 2020). The mitogen-activated protein kinase (MAPK)/ERK kinase-ERK cascade was shown to play a role in glutamate-stimulated induction of Homer1a mRNA in cerebellar granule cells (Denkena et al., 2020). Given that light has been shown to activate ERK/MAPK signaling in the SCN, attenuated by glutamate receptor antagonists (Huang et al., 2020), this signaling mechanism may be responsible for the activity-dependent Homer1a mRNA induction in the SCN (Nielsen et al., 2002).
Role of Homer in retinal ischemic disorders
Homer proteins have been shown to link neurotransmitter receptors, plasma membrane ion channels, intracellular calcium channels, and the cytoskeleton, as a postsynaptic clustering molecule (Rybchyn et al., 2019; Zhu et al., 2019; Bridi et al., 2020; Fjell et al., 2020; Yoon et al., 2021). The Homer protein, Homer1c, was thus used as a unique biomarker to reveal alterations in synaptic connectivity preceding apoptosis and during the early stages of apoptosis in retinal ganglion cells (RGCs), mediated or affected by extracellular signaling, intracellular signaling, or cytoskeletal processes. Kaja et al. (2003) quantitatively analyzed the effects of estrogen administration on synaptic connections of neurons in the ganglion cell layer (GCL) of the retina in a model of mild retinal ischemia, affecting only a small percentage of neurons in the GCL, including RGCs. This model resembled pathophysiological processes with slow progression or mild onset characteristics (glaucoma, retinal ischemia during diabetic retinopathy, sickle cell retinopathy, carotid artery stenosis) (Abdalla et al., 2019; Zhang et al., 2021). Interestingly, there was a significantly higher percentage reduction in Homer1c-positive synapses in the IPL compared with terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL)-positive apoptotic neurons in the GCL, and estrogen prevented both ischemia-induced loss of Homer1c-immunoreactive synapses in the IPL and apoptosis of cells in the GCL. This indicated that Homer1c-immunoreactive synapses can function as an early indicator of neurodegeneration, and the protective effects of estrogen on neurons in the GCL, especially Homer1c-positive contacts of RGCs to cone bipolar cells in the IPL, are highly specific and reproducible. This may support the development of novel treatments for neurodegenerative diseases and acute neurotoxic insults to RGCs (Kaja et al., 2003). In an in vitro study, Wang et al. (2017) also used Homer1c as a biomarker of PSD, and showed that elevated hydrostatic pressure in retinal neuron cultures had no effect on the expression of presynaptic or postsynaptic proteins like Homer1c, while the addition of recombinant thrombospondin 2 protein to retinal neuron cultures upregulated the level of presynaptic proteins.
Kaja et al. (2014) further studied mild retinal ischemia by measuring molecular determinants in the retina responsible for elevated susceptibility of RGCs to degeneration during glaucoma. They measured and assessed the effects of Homer expression in the retina of DBA/2J mice using behavioral analyses, quantitative gene expression, quantitative immunoblotting, immunohistochemistry, and microfluorimetry. They showed that Homer proteins were differentially expressed in aged and glaucomatous DBA/2J retinas, at both the transcriptional and translational levels. Immunoreactivity for the long Homer1c isoform, but not the IEG product Homer1a, was increased in the synaptic layers of the retina, and increased Homer1c protein levels were correlated with increased disease severity and decreased visual performance. IP3Rs and RyRs in the membranes of the ER and mGluRs at the plasma membrane were identified as binding partners of Homer proteins (Serwach and Gruszczynska-Biegala, 2020), and in addition to providing a critical clustering role, Homer isoforms also differentially altered the biophysical properties of RyRs (Del, 2013). It was speculated that increased Homer1c expression would result in increased calcium release, and an increased Homer1c:1a ratio in the aged DBA/2J retina would favor synaptic coupling that was likely to contribute to overall hyperexcitability and calcium toxicity (Umanskaya et al., 2014), whereas a lower ratio, as observed in young, non-glaucomatous DBA/2J mice, would promote homeostasis. Furthermore, the effect of elevated Homer1c levels was potentially exacerbated by increased release of calcium from intracellular stores following cellular oxidative stress (Lencesova and Krizanova, 2012; Chrysostomou et al., 2013; Guo et al., 2016), as occurs during glaucoma and related neurodegenerative pathologies (Droge and Kinscherf, 2008), and may be amplified through polycystin-2 intracellular calcium release channels present on the ER in the mouse retina (Kaja et al., 2014). Synaptic clustering of Homer1c was strongly correlated with functional markers of the severity of glaucoma in DBA/2J mice (Umanskaya et al., 2014). Homer1c represents a novel, potential drug target for the future development of anti-glaucoma therapies aimed at reducing hyperexcitability and aberrant neuronal calcium signaling in the glaucomatous retina (Kaja et al., 2014).
Homer1a and Homer1b/c have been shown to play important roles in the development of TBI via the regulation of group I mGluRs (Luo et al., 2011, 2014; Chen et al., 2012; Fei et al., 2014), and Homer1 proteins were also shown to be involved in the regulation of neuronal injury in models of inflammation (Luo et al., 2012b) and oxidative stress (Luo et al., 2012a). Fei et al. (2014) investigated changes in the expression and effects of Homer1a in RGCs both in vitro and in vivo after ischemia/reperfusion (I/R) injury using western blot and TUNEL assays, gene interference and overexpression, and gene knockout procedures. Levels of Homer1a and phosphorylated ERK (p-ERK) increased in RGCs and retinas after I/R injury. Upregulation of Homer1a in RGCs after I/R injury decreased p-ERK and mitigated RGC apoptosis, while downregulation of Homer1a increased p-ERK and augmented RGC apoptosis. Inhibition of p-ERK reduced RGC apoptosis and increased the expression of Homer1a after I/R injury. Finally, there were significantly fewer dendrites and RGCs in the retinas of Homer1a-knockout mice following I/R injury compared with Homer1a-wild-type mice. These results suggested that Homer1a may contribute to RGC survival after I/R injury by interacting with the ERK pathway (Fei et al., 2015).
Limitations
Importantly, this review had several limitations. First, the current review mainly focused on Homer1b/c and Homer1a, and an integrated understanding of other Homer proteins is also needed. Second, the role of Homer proteins in hemorrhagic strokes has rarely been discussed. This review therefore did not consider hemorrhagic stroke, and the roles of Homer proteins in hemorrhagic brain injuries thus deserves further study. Third, we mainly reviewed the effects of Homer proteins in acute brain injuries, and their roles in psychiatric disorders and neurodegenerative diseases were not reviewed. Fourth, Homer1b/c and Homer1a conditional knockout mice have rarely been used in research, and further studies are therefore needed to review the role of Homer in specific brain regions and nerve cells in vivo in the context of acute brain injury.
Conclusion
Ischemic and traumatic brain injury and retinal disorders are relatively common and can lead to serious social and economic burdens. These conditions are usually characterized by primary and secondary damage, and it is essential to inhibit secondary damage to improve neuronal survival; however, there is currently a lack of reliable and effective molecular targets for preventing secondary damage, particularly in relation to endogenous neuroprotective factors. Accumulating evidence suggests that Homer proteins play crucial roles ischemic and traumatic brain injuries and retinal lesions (Table 2). This review thus summarized the structural features, important related signaling pathways, and diverse roles of Homer proteins in both physiological and pathological processes. Specifically, we addressed the neuroprotective effects of downregulating Homer1b/c and upregulating Homer1a. As a key scaffolding protein, Homer1b/c couples with other proteins to form PSDs, which act as the basis of synaptic transmission, suggesting that extrinsic modulation of Homer1b/c expression might have some adverse effects. Interestingly, as an IEG, Homer1a expression is induced by detrimental external stimuli. Given the increasing therapeutic use of adeno-associated viruses in clinical practice, it is necessary to verify the effects of modulating Homer expression using adeno-associated virus transduction.
Table 2.
Selected studies of Homer proteins
| References | Types | Models | Signaling transduction | Indications (disease or significance) | |
|---|---|---|---|---|---|
| Prosser et al., 1989 | Homer1 | In vivo and in vitro | Rat circadian model | ERK/MAPK signaling | Generates circadian rhytms |
| Kaja et al., 2003 | Homer1c | In vivo | Rat mild retinal ischemia by MCAO | Glutamatergic retinal synapses | Glaucoma, retinal ischemia |
| Brandstatter et al., 2004 | Homer1a | In vitro | Rat retinal cells | Glutamatergic retinal synapses | Homer as important transmitter |
| Wahlin et al., 2008 | Homer1, 2 | In vitro | Chick retinal cells | Photoreceptor synaptogenesis | – |
| Chen et al., 2012 | Homer1bc | In vitro | Glutamate excitotoxity on primary rat cortical neurons | I mglur-IP3R-ER axis | TBI |
| Luo et al., 2014 | Homer 1a | In vivo and in vitro | Scratching mouse cortical neurons and weight-drop TBI model | Mglur1-ERK signaling | TBI |
| Umanskaya et al., 2014 | Homer 1c | In vivo | DBA/2J mice, a preclinical genetic glaucoma model | Increased Ca2+ release | Glaucoma |
| Fei et al., 2015 | Homer 1a | In vitro and in vivo | Retinal I/R | ERK pathway | Glaucoma, traumatic optic neuropathy, anterior ischemic |
| Wang et al., 2015 | Homer 1a | In vivo and in vitro | NMDA-induced injury | Nr2b-PSD95-NNOS | TBI |
| Guo et al., 2016 | Homer 1bc | In vitro | T-BHP-induced oxidative injury on brain endothelial cells | IP3R/RyR mediated Ca2+ release | Cerebral ischemia, TBI |
| Wu et al., 2018 | Homer 1a | In vitro | Hydrogen peroxide-induced oxidative injury | AMPK pathway | Cerebral ischemia, TBI |
| Wei et al., 2019 | Homer1a | In vitro | OGD/R on mouse cortical neurons | PERK pathway | Cerebral ischemia |
ER: Endoplasmic reticulum; ERK: extracellular signal-regulated kinase; IP3R: inositol trisphosphate receptor; MCAO: middle cerebral artery occlusion; mGluR: metabotropic glutamate receptor; NMDA: N-methyl-D-aspartate; OGD/R: oxygen and glucose deprivation/reperfusion; PERK: protein kinase-like ER kinase; RyR: ryanodine receptor; TBI: traumatic brain injury.
In summary, the roles of Homer proteins in ischemic and traumatic brain injuries and retinal disorders are interesting and complex. The current review indicated that both downregulating Homer1b/c and upregulating Homer1a may offer promising therapeutic approaches. However, further studies are needed to investigate many aspects of the neuroprotective effects of Homer1 and to clarify its potential role as a therapeutic target for clinical translational research.
Additional file: Open peer review reports 1 (87.2KB, pdf) and 2 (94.2KB, pdf) .
Footnotes
Conflicts of interest: The authors declare that there are no conflicts of interest associated with this manuscript.
Financial support: This work was supported by the National Natural Science Foundation of China, Nos. 81600738 (to FF), 81771239 (to ZF), 81801300 (to NS).
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewers: Janne Koskimäki, University of Turku, Finland; Peng Li, VCU Massey Cancer Center, USA.
Funding: This work was supported by the National Natural Science Foundation of China, Nos. 81600738 (to FF), 81771239 (to ZF), 81801300 (to NS).
P-Reviewers: Koskimäki J, Li P; C-Editor: Zhao M; S-Editors: Wang J, Li CH; L-Editors: Furness S, Song LP; T-Editor: Jia Y.
References
- 1.Abdalla EM, Mura M, Al DH, Schellini S, Malik R, Kozak I, Schatz P. Sickle cell retinopathy. A focused review. Graefes Arch Clin Exp Ophthalmol. 2019;257:1353–1364. doi: 10.1007/s00417-019-04294-2. [DOI] [PubMed] [Google Scholar]
- 2.Ahumada-Castro U, Bustos G, Silva-Pavez E, Puebla-Huerta A, Lovy A, Cardenas C. In the right place at the right time: regulation of cell metabolism by IP3R-mediated inter-organelle Ca(2+) fluxes. Front Cell Dev Biol. 2021;9:629522. doi: 10.3389/fcell.2021.629522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Baratz R, Tweedie D, Wang JY, Rubovitch V, Luo W, Hoffer BJ, Greig NH, Pick CG. Transiently lowering tumor necrosis factor-alpha synthesis ameliorates neuronal cell loss and cognitive impairments induced by minimal traumatic brain injury in mice. J Neuroinflammation. 2015;12:45. doi: 10.1186/s12974-015-0237-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Brakeman PR, Lanahan AA, O’Brien R, Roche K, Barnes CA, Huganir RL, Worley PF. Homer: a protein that selectively binds metabotropic glutamate receptors. Nature. 1997;386:284–288. doi: 10.1038/386284a0. [DOI] [PubMed] [Google Scholar]
- 5.Brandstatter JH, Dick O, Boeckers TM. The postsynaptic scaffold proteins ProSAP1/Shank2 and Homer1 are associated with glutamate receptor complexes at rat retinal synapses. J Comp Neurol. 2004;475:551–563. doi: 10.1002/cne.20194. [DOI] [PubMed] [Google Scholar]
- 6.Bridi M, Schoch H, Florian C, Poplawski SG, Banerjee A, Hawk JD, Porcari GS, Lejards C, Hahn CG, Giese KP, Havekes R, Spruston N, Abel T. Transcriptional corepressor SIN3A regulates hippocampal synaptic plasticity via Homer1/mGluR5 signaling. JCI Insight. 2020;5:e92385. doi: 10.1172/jci.insight.92385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Buonaguro EF, Morley-Fletcher S, Avagliano C, Vellucci L, Iasevoli F, Bouwalerh H, Van Camp G, Nicoletti F, Maccari S, de Bartolomeis A. Glutamatergic postsynaptic density in early life stress programming: topographic gene expression of mGlu5 receptors and Homer proteins. Prog Neuropsychopharmacol Biol Psychiatry. 2020;96:109725. doi: 10.1016/j.pnpbp.2019.109725. [DOI] [PubMed] [Google Scholar]
- 8.Chakraborty R, Pardue MT. Molecular and biochemical aspects of the retina on refraction. Prog Mol Biol Transl Sci. 2015;134:249–267. doi: 10.1016/bs.pmbts.2015.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chen T, Fei F, Jiang XF, Zhang L, Qu Y, Huo K, Fei Z. Down-regulation of Homer1b/c attenuates glutamate-mediated excitotoxicity through endoplasmic reticulum and mitochondria pathways in rat cortical neurons. Free Radic Biol Med. 2012;52:208–217. doi: 10.1016/j.freeradbiomed.2011.10.451. [DOI] [PubMed] [Google Scholar]
- 10.Chen X, Sooch G, Demaree IS, White FA, Obukhov AG. Transient receptor potential canonical (TRPC) channels: then and now. Cells. 2020;9:1983. doi: 10.3390/cells9091983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chokshi V, Druciak B, Worley PF, Lee HK. Homer1a is required for establishment of contralateral bias and maintenance of ocular dominance in mouse visual cortex. J Neurosci. 2019a;39:3897–3905. doi: 10.1523/JNEUROSCI.3188-18.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chokshi V, Gao M, Grier BD, Owens A, Wang H, Worley PF, Lee HK. Input-specific metaplasticity in the visual cortex requires homer1a-mediated mGluR5 signaling. Neuron. 2019b;104:736–748. doi: 10.1016/j.neuron.2019.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chrysostomou V, Rezania F, Trounce IA, Crowston JG. Oxidative stress and mitochondrial dysfunction in glaucoma. Curr Opin Pharmacol. 2013;13:12–15. doi: 10.1016/j.coph.2012.09.008. [DOI] [PubMed] [Google Scholar]
- 14.Colecraft HM. Designer genetically encoded voltage-dependent calcium channel inhibitors inspired by RGK GTPases. J Physiol. 2020;598:1683–1693. doi: 10.1113/JP276544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Denkena J, Zaisser A, Merz B, Klinger B, Kuhl D, Bluthgen N, Hermey G. Neuronal activity regulates alternative exon usage. Mol Brain. 2020;13:148. doi: 10.1186/s13041-020-00685-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dionisio N, Smani T, Woodard GE, Castellano A, Salido GM, Rosado JA. Homer proteins mediate the interaction between STIM1 and Cav1.2 channels. Biochim Biophys Acta. 2015;1853:1145–1153. doi: 10.1016/j.bbamcr.2015.02.014. [DOI] [PubMed] [Google Scholar]
- 17.Docampo R, Huang G. The IP3 receptor and Ca2+ signaling in trypanosomes. Biochim Biophys Acta Mol Cell Res. 2021;1868:118947. doi: 10.1016/j.bbamcr.2021.118947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Droge W, Kinscherf R. Aberrant insulin receptor signaling and amino acid homeostasis as a major cause of oxidative stress in aging. Antioxid Redox Signal. 2008;10:661–678. doi: 10.1089/ars.2007.1953. [DOI] [PubMed] [Google Scholar]
- 19.Fei F, Li J, Rao W, Liu W, Chen X, Su N, Wang Y, Fei Z. Upregulation of homer1a promoted retinal ganglion cell survival after retinal ischemia and reperfusion via interacting with Erk pathway. Cell Mol Neurobiol. 2015;35:1039–1048. doi: 10.1007/s10571-015-0198-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fei F, Rao W, Zhang L, Chen BG, Li J, Fei Z, Chen Z. Downregulation of Homer1b/c improves neuronal survival after traumatic neuronal injury. Neuroscience. 2014;267:187–194. doi: 10.1016/j.neuroscience.2014.02.037. [DOI] [PubMed] [Google Scholar]
- 21.Ferrara F, Zeisig V, Pietsch S, Rutten R, Dreyer AY, Pieper L, Schatzl AK, McLeod DD, Barthel H, Boltze J, Schrodl W, Nitzsche B. Hypothesis and theory: a pathophysiological concept of stroke-induced acute phase response and increased intestinal permeability leading to secondary brain damage. Front Neurosci. 2020;14:272. doi: 10.3389/fnins.2020.00272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fjell AM, Sederevicius D, Sneve MH, de Lange AG, Brathen AC, Idland AV, Watne LO, Wang Y, Reinbold C, Dobricic V, Kilpert F, Blennow K, Zetterbergj H, Hong S, Bertram L, Walhovd KB. Self-reported sleep problems related to amyloid deposition in cortical regions with high HOMER1 gene expression. Cereb Cortex. 2020;30:2144–2156. doi: 10.1093/cercor/bhz228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Guo ZY, Zhang YH, Xie GQ, Liu CX, Zhou R, Shi W. Down-regulation of Homer1 attenuates t-BHP-induced oxidative stress through regulating calcium homeostasis and ER stress in brain endothelial cells. Biochem Biophys Res Commun. 2016;477:970–976. doi: 10.1016/j.bbrc.2016.07.012. [DOI] [PubMed] [Google Scholar]
- 24.Hassani NF, Kreienkamp HJ. Functional relevance of missense mutations affecting the N-terminal part of shank3 found in autistic Patients. Front Mol Neurosci. 2018;11:268. doi: 10.3389/fnmol.2018.00268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hausott B, Klimaschewski L. Sprouty2-a novel therapeutic target in the nervous system? Mol Neurobiol. 2019;56:3897–3903. doi: 10.1007/s12035-018-1338-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hayashi Y. Drebrin-homer interaction at an atomic scale. Structure. 2019;27:3–5. doi: 10.1016/j.str.2018.12.008. [DOI] [PubMed] [Google Scholar]
- 27.Huang TL, Wen YT, Ho YC, Wang JK, Lin KH, Tsai RK. Algae oil treatment protects retinal ganglion cells (RGCs) via ERK signaling pathway in experimental optic nerve ischemia. Mar Drugs. 2020;18:83. doi: 10.3390/md18020083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Huang W, Liu X, Fei Z, Zhang Y, Yang J. Down-regulation of Homer1b/c expression protects cultured neurons after traumatic injury. Neural Regen Res. 2012;7:2176–2181. doi: 10.3969/j.issn.1673-5374.2012.028.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Huang WD, Fei Z, Zhang X. Traumatic injury induced homer-1a gene expression in cultured cortical neurons of rat. Neurosci Lett. 2005;389:46–50. doi: 10.1016/j.neulet.2005.07.014. [DOI] [PubMed] [Google Scholar]
- 30.Jia S, Rodriguez M, Williams AG, Yuan JP. Homer binds to Orai1 and TRPC channels in the neointima and regulates vascular smooth muscle cell migration and proliferation. Sci Rep. 2017;7:5075. doi: 10.1038/s41598-017-04747-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jin C, Lee Y, Kang H, Jeong K, Park J, Zhang Y, Kang HR, Ma R, Seong H, Kim Y, Jung H, Kim JY, Kim YK, Han K. Increased ribosomal protein levels and protein synthesis in the striatal synaptosome of Shank3-overexpressing transgenic mice. Mol Brain. 2021;14:39. doi: 10.1186/s13041-021-00756-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kaja S, Naumchuk Y, Grillo SL, Borden PK, Koulen P. Differential up-regulation of Vesl-1/Homer 1 protein isoforms associated with decline in visual performance in a preclinical glaucoma model. Vision Res. 2014;94:16–23. doi: 10.1016/j.visres.2013.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kaja S, Yang SH, Wei J, Fujitani K, Liu R, Brun-Zinkernagel AM, Simpkins JW, Inokuchi K, Koulen P. Estrogen protects the inner retina from apoptosis and ischemia-induced loss of Vesl-1L/Homer 1c immunoreactive synaptic connections. Invest Ophthalmol Vis Sci. 2003;44:3155–3162. doi: 10.1167/iovs.02-1204. [DOI] [PubMed] [Google Scholar]
- 34.Kato A, Ozawa F, Saitoh Y, Hirai K, Inokuchi K. vesl, a gene encoding VASP/Ena family related protein, is upregulated during seizure, long-term potentiation and synaptogenesis. FEBS Lett. 1997;412:183–189. doi: 10.1016/s0014-5793(97)00775-8. [DOI] [PubMed] [Google Scholar]
- 35.Lee J, Rosales JL, Byun HG, Lee KY. D, L-Methadone causes leukemic cell apoptosis via an OPRM1-triggered increase in IP3R-mediated ER Ca(2+) release and decrease in Ca(2+) efflux, elevating [Ca(2+)]i. Sci Rep. 2021;11:1009. doi: 10.1038/s41598-020-80520-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lencesova L, Krizanova O. IP3 receptors, stress and apoptosis. Gen Physiol Biophys. 2012;31:119–130. doi: 10.4149/gpb_2012_014. [DOI] [PubMed] [Google Scholar]
- 37.Li R, Wang Y, Chen P, Meng J, Zhang H. G-protein coupled estrogen receptor activation protects the viability of hyperoxia-treated primary murine retinal microglia by reducing ER stress. Aging (Albany NY) 2020;12:17367–17379. doi: 10.18632/aging.103733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Li Z, Liu H, Li J, Yang Q, Feng Z, Li Y, Yang H, Yu C, Wan J, Liu W, Zhang M. Homer tetramer promotes actin bundling activity of drebrin. Structure. 2019;27:27–38. doi: 10.1016/j.str.2018.10.011. [DOI] [PubMed] [Google Scholar]
- 39.Luo L, Huang M, Zhang Y, Wang W, Ma X, Shi H, Worley PF, Kim DK, Fedorovich SV, Jiang W, Xu T. Disabling phosphorylation at the homer ligand of the metabotropic glutamate receptor 5 alleviates complete Freund's adjuvant-induced inflammatory pain. Neuropharmacology. 2020;170:108046. doi: 10.1016/j.neuropharm.2020.108046. [DOI] [PubMed] [Google Scholar]
- 40.Luo P, Fei F, Zhang L, Qu Y, Fei Z. The role of glutamate receptors in traumatic brain injury: implications for postsynaptic density in pathophysiology. Brain Res Bull. 2011;85:313–320. doi: 10.1016/j.brainresbull.2011.05.004. [DOI] [PubMed] [Google Scholar]
- 41.Luo P, Chen T, Zhao Y, Xu H, Huo K, Zhao M, Yang Y, Fei Z. Protective effect of Homer 1a against hydrogen peroxide-induced oxidative stress in PC12 cells. Free Radic Res. 2012a;46:766–776. doi: 10.3109/10715762.2012.678340. [DOI] [PubMed] [Google Scholar]
- 42.Luo P, Chen T, Zhao Y, Zhang L, Yang Y, Liu W, Li S, Rao W, Dai S, Yang J, Fei Z. Postsynaptic scaffold protein Homer 1a protects against traumatic brain injury via regulating group I metabotropic glutamate receptors. Cell Death Dis. 2014;5:e1174. doi: 10.1038/cddis.2014.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Luo P, Zhao Y, Li D, Chen T, Li S, Chao X, Liu W, Zhang L, Qu Y, Jiang X, Lu G, Poon W, Fei Z. Protective effect of Homer 1a on tumor necrosis factor-alpha with cycloheximide-induced apoptosis is mediated by mitogen-activated protein kinase pathways. Apoptosis. 2012b;17:975–988. doi: 10.1007/s10495-012-0736-z. [DOI] [PubMed] [Google Scholar]
- 44.Mahato PK, Ramsakha N, Ojha P, Gulia R, Sharma R, Bhattacharyya S. Group I metabotropic glutamate receptors (mGluRs): ins and outs. Adv Exp Med Biol. 2018;1112:163–175. doi: 10.1007/978-981-13-3065-0_12. [DOI] [PubMed] [Google Scholar]
- 45.Martin SC, Monroe SK, Diering GH. Homer1a and mGluR1/5 signaling in homeostatic sleep drive and output. Yale J Biol Med. 2019;92:93–101. [PMC free article] [PubMed] [Google Scholar]
- 46.Masis-Vargas A, Ritsema W, Mendoza J, Kalsbeek A. Metabolic effects of light at night are time- and wavelength-dependent in Rats. Obesity (Silver Spring) 2020;28(Suppl 1):S114–S125. doi: 10.1002/oby.22874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.May V, Johnson GC, Hammack SE, Braas KM, Parsons RL. PAC1 receptor internalization and endosomal MEK/ERK activation is essential for pacap-mediated neuronal excitability. J Mol Neurosci. 2021 doi: 10.1007/s12031-021-01821-x. DOI: 10.1007/s12031-021-01821-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Menard C, Quirion R. Successful cognitive aging in rats: a role for mGluR5 glutamate receptors, homer 1 proteins and downstream signaling pathways. PLoS One. 2012;7:e28666. doi: 10.1371/journal.pone.0028666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Nielsen HS, Hannibal J, Knudsen SM, Fahrenkrug J. Pituitary adenylate cyclase-activating polypeptide induces period1 and period2 gene expression in the rat suprachiasmatic nucleus during late night. Neuroscience. 2001;103:433–441. doi: 10.1016/s0306-4522(00)00563-7. [DOI] [PubMed] [Google Scholar]
- 50.Nielsen HS, Georg B, Hannibal J, Fahrenkrug J. Homer-1 mRNA in the rat suprachiasmatic nucleus is regulated differentially by the retinohypothalamic tract transmitters pituitary adenylate cyclase activating polypeptide and glutamate at time points where light phase-shifts the endogenous rhythm. Brain Res Mol Brain Res. 2002;105:79–85. doi: 10.1016/s0169-328x(02)00395-9. [DOI] [PubMed] [Google Scholar]
- 51.O’Neill N, McLaughlin C, Komiyama N, Sylantyev S. Biphasic modulation of NMDA receptor function by metabotropic glutamate receptors. J Neurosci. 2018;38:9840–9855. doi: 10.1523/JNEUROSCI.1000-18.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Orgovan Z, Ferenczy GG, Keseru GM. Fragment-based approaches for allosteric metabotropic glutamate receptor (mGluR) modulators. Curr Top Med Chem. 2019;19:1768–1781. doi: 10.2174/1568026619666190808150039. [DOI] [PubMed] [Google Scholar]
- 53.Osborne NN, Del OS. Maintenance of retinal ganglion cell mitochondrial functions as a neuroprotective strategy in glaucoma. Curr Opin Pharmacol. 2013;13:16–22. doi: 10.1016/j.coph.2012.09.002. [DOI] [PubMed] [Google Scholar]
- 54.Osmankovic D, Doric S, Pojskic N, Lukic BL. New approach to detect coiled coil and leucine zipper motifs in protein sequences. J Comput Biol. 2018;25:1278–1283. doi: 10.1089/cmb.2018.0104. [DOI] [PubMed] [Google Scholar]
- 55.Paquet M, Ribeiro FM, Guadagno J, Esseltine JL, Ferguson SS, Cregan SP. Role of metabotropic glutamate receptor 5 signaling and homer in oxygen glucose deprivation-mediated astrocyte apoptosis. Mol Brain. 2013;6:9. doi: 10.1186/1756-6606-6-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Perni S, Beam K. Neuronal junctophilins recruit specific CaV and RyR isoforms to ER-PM junctions and functionally alter CaV2.1 and CaV2.2. Elife. 2021;10:e64249. doi: 10.7554/eLife.64249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Petzold J, Szumlinski KK, London ED. Targeting mGlu5 for methamphetamine use disorder. Pharmacol Ther. 2021;224:107831. doi: 10.1016/j.pharmthera.2021.107831. [DOI] [PubMed] [Google Scholar]
- 58.Pichaud F, Walther RF, Nunes DAF. Regulation of Cdc42 and its effectors in epithelial morphogenesis. J Cell Sci. 2019;132:jcs217869. doi: 10.1242/jcs.217869. [DOI] [PubMed] [Google Scholar]
- 59.Prosser RA, McArthur AJ, Gillette MU. cGMP induces phase shifts of a mammalian circadian pacemaker at night, in antiphase to cAMP effects. Proc Natl Acad Sci U S A. 1989;86:6812–6815. doi: 10.1073/pnas.86.17.6812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Rao W, Peng C, Zhang L, Su N, Wang K, Hui H, Dai SH, Yang YF, Luo P, Fei Z. Homer1a attenuates glutamate-induced oxidative injury in HT-22 cells through regulation of store-operated calcium entry. Sci Rep. 2016;6:33975. doi: 10.1038/srep33975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Rauen K, Pop V, Trabold R, Badaut J, Plesnila N. Vasopressin v1a receptors regulate cerebral aquaporin 1 after traumatic brain injury. J Neurotrauma. 2020;37:665–674. doi: 10.1089/neu.2019.6653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Reibring CG, Hallberg K, Linde A, Gritli-Linde A. Distinct and overlapping expression patterns of the homer family of scaffolding proteins and their encoding genes in developing murine cephalic tissues. Int J Mol Sci. 2020;21:1264. doi: 10.3390/ijms21041264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rozov A, Zivkovic AR, Schwarz MK. Homer1 gene products orchestrate Ca(2+)-permeable AMPA receptor distribution and LTP expression. Front Synaptic Neurosci. 2012;4:4. doi: 10.3389/fnsyn.2012.00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Rybchyn MS, Islam KS, Brennan-Speranza TC, Cheng Z, Brennan SC, Chang W, Mason RS, Conigrave AD. Homer1 mediates CaSR-dependent activation of mTOR complex 2 and initiates a novel pathway for AKT-dependent beta-catenin stabilization in osteoblasts. J Biol Chem. 2019;294:16337–16350. doi: 10.1074/jbc.RA118.006587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Sato S, Bunney BG, Vawter MP, Bunney WE, Sassone-Corsi P. Homer1a undergoes bimodal transcriptional regulation by CREB and the circadian clock. Neuroscience. 2020;434:161–170. doi: 10.1016/j.neuroscience.2020.03.031. [DOI] [PubMed] [Google Scholar]
- 66.Schaffler MD, Middleton LJ, Abdus-Saboor I. Mechanisms of tactile sensory phenotypes in autism: current understanding and future directions for research. Curr Psychiatry Rep. 2019;21:134. doi: 10.1007/s11920-019-1122-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Serwach K, Gruszczynska-Biegala J. Target molecules of STIM proteins in the central nervous system. Front Mol Neurosci. 2020;13:617422. doi: 10.3389/fnmol.2020.617422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Shan W, Nagai T, Tanaka M, Itoh N, Furukawa-Hibi Y, Nabeshima T, Sokabe M, Yamada K. Neuronal PAS domain protein 4 (Npas4) controls neuronal homeostasis in pentylenetetrazole-induced epilepsy through the induction of Homer1a. J Neurochem. 2018;145:19–33. doi: 10.1111/jnc.14274. [DOI] [PubMed] [Google Scholar]
- 69.Shi Z, Zhang W, Lu Y, Lu Y, Xu L, Fang Q, Wu M, Jia M, Wang Y, Dong L, Yan X, Yang S, Yuan F. Aquaporin 4-mediated glutamate-induced astrocyte swelling is partially mediated through metabotropic glutamate receptor 5 activation. Front Cell Neurosci. 2017;11:116. doi: 10.3389/fncel.2017.00116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Sladojevic N, Yu B, Liao JK. Regulator of G-Protein signaling 5 maintains brain endothelial cell function in focal cerebral ischemia. J Am Heart Assoc. 2020;9:e17533. doi: 10.1161/JAHA.120.017533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Su JJ, Pan H, Zhou HG, Tang YP, Dong Q, Liu JR. Acid-sensing ion channels activation and hypoxia upregulate Homer1a expression. Cns Neurosci Ther. 2014;20:264–274. doi: 10.1111/cns.12206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sun ZC, Ma SB, Chu WG, Jia D, Luo C. Canonical transient receptor potential (TRPC) channels in nociception and pathological pain. Neural Plast. 2020;2020:3764193. doi: 10.1155/2020/3764193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Sylantyev S, Savtchenko LP, Ermolyuk Y, Michaluk P, Rusakov DA. Spike-driven glutamate electrodiffusion triggers synaptic potentiation via a homer-dependent mGluR-NMDAR link. Neuron. 2013;77:528–541. doi: 10.1016/j.neuron.2012.11.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Umanskaya A, Santulli G, Xie W, Andersson DC, Reiken SR, Marks AR. Genetically enhancing mitochondrial antioxidant activity improves muscle function in aging. Proc Natl Acad Sci U S A. 2014;111:15250–15255. doi: 10.1073/pnas.1412754111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Wahlin KJ, Moreira EF, Huang H, Yu N, Adler R. Molecular dynamics of photoreceptor synapse formation in the developing chick retina. J Comp Neurol. 2008;506:822–837. doi: 10.1002/cne.21582. [DOI] [PubMed] [Google Scholar]
- 76.Wang H. Global, regional, and national under-5 mortality, adult mortality, age-specific mortality, and life expectancy, 1970-2016: a systematic analysis for the global burden of disease study 2016. Lancet. 2017;390:1084–1150. doi: 10.1016/S0140-6736(17)31833-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wang S, Hu T, Wang Z, Li N, Zhou L, Liao L, Wang M, Liao L, Wang H, Zeng L, Fan C, Zhou H, Xiong K, Huang J, Chen D. Macroglia-derived thrombospondin 2 regulates alterations of presynaptic proteins of retinal neurons following elevated hydrostatic pressure. PLoS One. 2017;12:e185388. doi: 10.1371/journal.pone.0185388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wang Y, Rao W, Zhang C, Zhang C, Liu MD, Han F, Yao LB, Han H, Luo P, Su N, Fei Z. Scaffolding protein Homer1a protects against NMDA-induced neuronal injury. Cell Death Dis. 2015;6:e1843. doi: 10.1038/cddis.2015.216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Wang Y, Zhao M, Shang L, Zhang Y, Huang C, He Z, Luo M, Wu B, Song P, Wang M, Duan F. Homer1a protects against neuronal injury via PI3K/AKT/mTOR signaling pathway. Int J Neurosci. 2020;130:621–630. doi: 10.1080/00207454.2019.1702535. [DOI] [PubMed] [Google Scholar]
- 80.Wei J, Wu X, Luo P, Yue K, Yu Y, Pu J, Zhang L, Dai S, Han D, Fei Z. Homer1a attenuates endoplasmic reticulum stress-induced mitochondrial stress after ischemic reperfusion injury by inhibiting the PERK pathway. Front Cell Neurosci. 2019;13:101. doi: 10.3389/fncel.2019.00101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Witharana W, Clark BJ, Trivedi V, Mesina L, McNaughton BL. Immediate-early gene Homer1a intranuclear transcription focus intensity as a measure of relative neural activation. Hippocampus. 2019;29:481–490. doi: 10.1002/hipo.23036. [DOI] [PubMed] [Google Scholar]
- 82.Wofford KL, Loane DJ, Cullen DK. Acute drivers of neuroinflammation in traumatic brain injury. Neural Regen Res. 2019;14:1481–1489. doi: 10.4103/1673-5374.255958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Wu X, Luo P, Rao W, Dai S, Zhang L, Ma W, Pu J, Yu Y, Wang J, Fei Z. Homer1a attenuates hydrogen peroxide-induced oxidative damage in HT-22 cells through AMPK-dependent autophagy. Front Neurosci. 2018;12:51. doi: 10.3389/fnins.2018.00051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Yang K, Zhou Y, Zhou L, Yan F, Guan L, Liu H, Liu W. Synaptic plasticity after focal cerebral ischemia was attenuated by gap26 but enhanced by GAP-134. Front Neurol. 2020;11:888. doi: 10.3389/fneur.2020.00888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Yoon S, Piguel NH, Khalatyan N, Dionisio LE, Savas JN, Penzes P. Homer1 promotes dendritic spine growth through ankyrin-G and its loss reshapes the synaptic proteome. Mol Psychiatry. 2021 doi: 10.1038/s41380-020-00991-1. doi: 10.1038/s41380-020-00991-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Yuan JP, Lee KP, Hong JH, Muallem S. The closing and opening of TRPC channels by Homer1 and STIM1. Acta Physiol (Oxf) 2012;204:238–247. doi: 10.1111/j.1748-1716.2011.02319.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Zhang J, Lai ZP, Chen P, Ying Y, Zhuang J, Yu KM. Glycogen synthase kinase-3beta inhibitor SB216763 promotes DNA repair in ischemic retinal neurons. Neural Regen Res. 2021;16:394–400. doi: 10.4103/1673-5374.290913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Zhao C, Deng Y, He Y, Huang X, Wang C, Li W. Decreased level of exosomal mir-5121 released from microglia suppresses neurite outgrowth and synapse recovery of neurons following traumatic brain injury? Neurotherapeutics. 2021 doi: 10.1007/s13311-020-00999-z. doi: 10.1007/s13311-020-00999-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Zhu J, Hafycz J, Keenan BT, Guo X, Pack A, Naidoo N. Acute sleep loss upregulates the synaptic scaffolding protein, homer1a, in non-canonical sleep/wake brain regions, claustrum, piriform and cingulate cortices. Front Neurosci. 2020;14:188. doi: 10.3389/fnins.2020.00188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Zhu L, Zhu J, Guo SW. Homer1/mGluR1-mediated ER stress contributes to lysophosphatidic acid-induced neurotoxicity in cortical neurons. Neurochem Int. 2019;129:104515. doi: 10.1016/j.neuint.2019.104515. [DOI] [PubMed] [Google Scholar]
- 91.Zhu M, Zuo J, Shen J, Jing W, Luo P, Li N, Wen X, Wang C, Yu M, Liang C, Tu J. Diagnostic potential of differentially expressed homer1 and homer2 in ischemic stroke. Cell Physiol Biochem. 2016;39:2353–2363. doi: 10.1159/000447927. [DOI] [PubMed] [Google Scholar]
- 92.Zuo C, Cao H, Ding F, Zhao J, Huang Y, Li G, Huang S, Jiang H, Jiang Y, Wang F. Neuroprotective efficacy of different levels of high-frequency repetitive transcranial magnetic stimulation in mice with CUMS-induced depression: involvement of the p11/BDNF/Homer1a signaling pathway. J Psychiatr Res. 2020;125:152–163. doi: 10.1016/j.jpsychires.2020.03.018. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.