

Abstract
Candidiasis, caused by the opportunistic yeast Candida albicans, is the most common fungal infection today. Resistance of C. albicans to current antifungal drugs has emerged over the past decade leading to the need for novel antifungal agents. Our aim was to select new antifungal compounds by library-screening methods and to assess their antifungal effects against C. albicans. After screening 90 potential antifungal compounds from JUNIA, a chemical library, two compounds, 1-(4-chlorophenyl)-4-((4-chlorophenyl)amino)-3,6-dimethylpyridin-2(1H)-one (PYR) and (Z)-N-(2-(4,6-dimethoxy-1,3,5-triazin-2-yl)vinyl)-4-methoxyaniline (TRI), were identified as having potential antifungal activity. Treatment with PYR and TRI resulted in a significant reduction of C. albicans bioluminescence as well as the number of fungal colonies, indicating rapid fungicidal activity. These two compounds were also effective against clinically isolated fluconazole- or caspofungin-resistant C. albicans strains. PYR and TRI had an inhibitory effect on Candida biofilm formation and reduced the thickness of the mannan cell wall. In a Caenorhabditis elegans infection model, PYR and TRI decreased the mortality of nematodes infected with C. albicans and enhanced the expression of antimicrobial genes that promote C. albicans elimination. Overall, PYR and TRI showed antifungal properties against C. albicans by exerting fungicidal activities and enhancing the antimicrobial gene expression of Caenorhabditis elegans.
Keywords: Candida albicans, antifungal agents, biofilm formation, C. elegans infection model
1. Introduction
Opportunistic fungi have become an increasingly important cause of nosocomial bloodstream infections, with high rates of morbidity and mortality in intensive care units [1,2]. These fungal infections are particularly problematic for immunocompromised patients, as well as patients with solid-organ malignancies or those recovering from abdominal surgery [1,2]. Candida albicans is an opportunistic yeast that colonizes the oropharyngeal, esophageal, and gastrointestinal mucosa in most healthy humans. Overgrowth of C. albicans in these niches can result in mucosal infections and life-threatening systemic disease, making C. albicans an important opportunistic pathogen. C. albicans is the most commonly identified Candida species in clinical settings and is one of the leading causes of hospital-acquired infections. Antifungal resistance is an emerging problem worldwide and this complicates the selection of appropriate antifungal therapy [3,4]. A subset of Candida strains are now resistant to first-line antifungal agents such as fluconazole and echinocandins [4]. This increasing prevalence of strains that are resistant to current antifungal drugs correlates with high azole and echinocandin use in clinical wards hosting patients at high risk of invasive candidiasis.
C. albicans is capable of adhering to catheters and other medical devices contributing to the formation of biofilms, which are heterogeneous structures containing hyphae, pseudohyphae, and yeast cells. These develop at the interface between an aqueous medium and a solid [5]. Biofilms are difficult for antifungals to penetrate and represent a potential source of candidemia relapses, leading to high rates of morbidity and mortality. Biofilms are therefore an important virulence factor and closely correlate with invasive fungal infections [6].
The nematode Caenorhabditis elegans has been widely used as a model for the study of C. albicans pathogenesis and the host immune system [7]. The C. elegans model enables the screening of different compounds for antifungal activity. The C. elegans immune response also plays a critical role during host–microbe interactions because many fungal and bacterial pathogens that cause illness in humans also cause tissue damage in C. elegans [8]. Antimicrobial peptides are the key components of innate immunity in C. elegans, including the carbohydrate-binding C-type lectin encoding clec gene repertoire, lysozymes, and caenacins [9,10]. In addition, a p38 mitogen activated kinase (MAPK) pathway, involving MAPK PMK-1, is among the different signaling pathways involved in modulating the innate immune response in C. elegans [10,11].
Recently, 2,3-dihydroxy-4-methoxybenzaldehyde (DHMB) was identified as a compound with potential antifungal activity against fluconazole- and caspofungin-resistant C. albicans [12]. DHMB also reduced the clinical and histological scores for inflammation in DSS-induced colitis in mice and promoted the elimination of C. albicans from the gut [12].
In the present study, we screened 90 small aromatic compounds from the JUNIA chemical library for their possible antifungal activity against C. albicans. Two compounds were identified as novel small molecules with antifungal activity against Candida viability, Candida biofilms, and cell wall fitness. The antifungal effects of these two compounds on C. albicans virulence and the host immune response were studied in a C. elegans infection model.
2. Results
2.1. Screening of the Chemical Library
Ninety compounds from the chemical library were screened for their antifungal and antibacterial activity. (Z)-N-(2-(4,6-dimethoxy-1,3,5-triazin-2-yl)vinyl)-4-methoxyaniline (TRI) and 1-(4-chlorophenyl)-4-((4-chlorophenyl)amino)-3,6-dimethylpyridin-2(1H)-one (PYR) were found to have antifungal activity against C. albicans (Figure 1). These two compounds shared some similar features, including a 6-membered N-heterocycle, a free -NH- link, and a para-substituted aromatic moiety (see Figure 1).
Figure 1.

Active compounds TRI and PYR identified from initial screening; structure of compound C and reported active compounds similar to TRI and PYR.
Some derivatives of TRI and PYR were also screened against C. albicans to determine the importance of each moiety in antifungal activity. In a first experiment, modification of the amine moiety was performed, while keeping the 4,6-dimethoxy-1,3,5-triazine and ethylene linkers constant (Table 1). Suppression of the para-methoxy substituent on the phenyl ring (TRI1) led to a decrease in activity, measured at 32 µg/mL and confirmed at 0.8 mmol/L (Table 1, entry 2). Polymethoxylated substitution or addition of a dioxolane ring did not improve the activity against C. albicans, whatever the position of the substituent on the aromatic ring (Table 1, entries 3–8). Introducing a 3-hydroxy-4-methoxyphenyl unit, similar to DHMB, an antifungal previously described by our group, did not result in any improvement in activity (Table 1, entry 6) [12]. Moreover, replacing the methoxy groups with chloro substituents did not increase the antifungal potency (Table 1, entry 9). Introducing diethylamine in compound TRI9 instead of p-anisidine of TRI slightly decreased the activity, which could be related to the importance of the aromatic ring or the H-bonding capacity of the -NH- link (Table 1, entry 10). To confirm the role of the -NH- link, compound C, in which the p-methoxyphenyl group was conserved but the amino-vinyl linker was modified, was also screened (Figure 1). Compound C was 6-fold less active than TRI against C. albicans. This highlights the importance of a -NH- linker for activity.
Table 1.
Chemical modifications of TRI and PYR and their impact on antifungal activity.
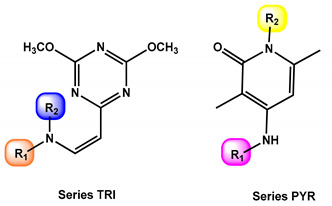
| Entry | CPD | R1 | R2 | %Inhibition at 32 µg/mLa |
%Inhibition at 0.8 mmol/L (µg/mL) b |
|---|---|---|---|---|---|
| 1 | TRI | 4-CH3O-C6H4- | H | 32.1 | 98 (230) |
| 2 | TRI1 | C6H5- | H | 3.9 | 72 (206) |
| 3 | TRI2 | 2,5-(CH3O)2-C6H4- | H | 1.7 | 77 (255) |
| 4 | TRI3 | 2,4-(CH3O)2-C6H4- | H | 0.5 | ND |
| 5 | TRI4 | 3,4,5-(CH3O)3-C6H4- | H | 1.9 | ND |
| 6 | TRI5 | 3-(OH),4-(CH3O)-C6H4- | H | 0.8 | ND |
| 7 | TRI6 |
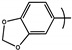
|
H | 1.4 | ND |
| 8 | TRI7 | 3,5-(CH3O)2-C6H4- | H | 3.9 | ND |
| 9 | TRI8 | 3,5-(Cl)2-C6H4- | H | 0.5 | ND |
| 10 | TRI9 | CH3-CH2- | CH3-CH2- | 3.8 | 84 (190) |
| 11 | PYR | 4-Cl-C6H4- | 4-Cl-C6H4- | 14.0 | 98 (287) |
| 12 | PYR1 | 4-CH3-C6H4- | 4-CH3-C6H4- | 8.1 | 88 (255) |
| 13 | PYR2 | C6H5- | C6H5- | 3.3 | ND |
| 14 | PYR3 | 4-CH3O-C6H4- | 4-CH3O-C6H4- | 4.2 | ND |
| 15 | PYR4 | 3,4-(CH3O)2-C6H4- | 3,4-(CH3O)2-C6H4- | 2.5 | ND |
| 16 | PYR5 |
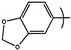
|

|
3.4 | ND |
| 17 | PYR6 | 3,4-(CH3O)2-C6H4- |
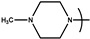
|
3.0 | ND |
| 18 | PYR7 | 3,4-(CH3O)2-C6H4- |
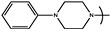
|
3.6 | ND |
Biological activity was evaluated at a concentration of a 32 µg/mL or b 0.8 mmol/L corresponding to 230, 206, 255, 190, 287, and 255 µg/mL for TRI, TRI1, TR2, TRI9, PYR, and PYR1, respectively, on C. albicans strain ATCC90028 grown for 3 days on yeast extract–peptone dextrose (YPD) agar at 30 °C. A yeast suspension of 1 × 106 –5 × 106 cfu/mL (determined by OD550) was prepared from five colonies. The suspension was diluted and added to each well of the compound-containing plates to give a final fungal cell concentration of 2.5 × 103 cfu/mL and a total volume of 50 µL. All plates were covered and incubated at 35 °C for 24 h without shaking. Inhibition of C. albicans growth was determined by measuring the absorbance at 530 nm (OD530). Fluconazole was used as a positive control. CPD: compound. ND: not determined.
Derivatives of PYR compounds were also evaluated, with chemical modifications on the aromatic moieties (Table 1, series B). In this experiment, it is notable that the para-chloroaniline moiety had the highest activity against C. albicans. Introducing inductive or mesomeric electro-donating substituents such as methyl- or methoxy- on the aromatic rings led to a decrease in biological activity (Table 1, entries 12–16). Moreover, replacing one aromatic ring by a piperazine part as in compounds PYR7 and PYR8 did not improve the activity (Table 1, entries 17–18). While in series A, the p-methoxyaniline was the most effective ring for antifungal activity, it appeared that in series B, p-chloroaniline was the best choice. Gathering these data, compounds TRI and PYR were selected for further biological studies.
2.2. Effect of PYR and TRI on C. albicans Viability
To determine the antifungal effect of PYR and TRI on C. albicans viability, C. albicans cells were challenged with PYR or TRI and viability was assessed by culture-plate assay (Figure 2). PYR and TRI at their 1× MIC (12.5 µg/mL and 50 µg/mL, respectively) reduced C. albicans viability and this antifungal activity was more pronounced than with caspofungin at 2× MIC (Figure 2). Additionally, the viability of C. albicans cells was monitored in real time using a bioluminescent C. albicans strain challenged with either caspofungin, fluconazole, PYR, or TRI (at 0, 30, 60, and 120 min). PYR and TRI at their MICs (12.5 µg/mL and 50 µg/mL, respectively) led to a significant reduction in bioluminescence of C. albicans when compared to that of C. albicans unchallenged with antifungal compounds (Figure 3). This reduction was observed rapidly after C. albicans treatment with PYR and TRI indicating the fungicidal effect of these compounds against C. albicans. We then analyzed whether derivatives of PYR (PYR1 and PYR2) and TRI (TRI1) had antifungal activity against C. albicans (Figure 3). In contrast to TRI1, the derivatives PYR1 and PYR2 had antifungal activity against C. albicans. Of note, the derivatives PYR1 and PYR2 had less antifungal activity against C. albicans when compared to their original compound, PYR (Figure 3). These data confirmed that the substitution of the aromatic moiety is highly important in the activity. For PYR derivatives, p-chloro substituent is superior to the p-methoxy one whereas the p-methoxy group is needed for activity in the TRI series of compounds.
Figure 2.

Viability of C. albicans by culture assay in the presence of caspofungin, fluconazole, PYR, or TRI. C. albicans SC5314 cells were challenged with caspofungin (Caspo), fluconazole (Fluco), PYR, or TRI at their (A) 1× MIC, or (B) 2× MIC. C. albicans (control) corresponds to C. albicans alone without any antifungal treatment. Error bars indicate standard deviations.
Figure 3.

Impact of PYR and TRI and their derivatives on C. albicans viability. Bioluminescent C. albicans strain was treated with PYR, TRI, and their derivatives (PYR-1, PYR-2, and TRI-1) at their 1x MIC and monitored at (A) 0, (B) 30, (C) 60, and (D) 120 min. CTL corresponds to PBS and substrate. CTL+ represents C. albicans challenged with PBS. Caspo corresponds to C. albicans cells challenged with caspofungin (1× MIC). Fluco represents C. albicans cells challenged with fluconazole (1× MIC). Error bars indicate standard deviations.
To evaluate whether PYR and TRI had antifungal activity against drug-resistant C. albicans clinical isolates, we selected five C. albicans clinical isolates resistant to either fluconazole or caspofungin (Table 2 and Table 3). PYR and TRI significantly decreased the viability of drug-resistant C. albicans clinical isolates at 1× MIC and 2× MIC in terms of viable colony counts using fungal culture media (Table 3). We next assessed the effect of PYR and TRI on the C. albicans cell wall (Figure 4). Quantification of the fluorescence intensity of C. albicans cells labeled with concanavalin A from 30 different images extracted from confocal microscopy using ImageJ showed that PYR and TIR reduced the thickness of the mannan part of the C. albicans cell wall while the labeling of C. albicans with wheat germ agglutinin (WGA) did not show any changes in the chitin part of the cell wall (Figure 4).
Table 2.
Antifungal activity of PYR and TRI versus caspofungin and fluconazole.
| Strain | Description | MIC Caspofungin µg/mL |
MIC Fluconazole µg/mL |
MIC PYR µg/mL |
MIC TRI µg/mL |
Strain Ref. |
|---|---|---|---|---|---|---|
| C. albicans SC5314 | Wild-type | 0.03 | 0.5 | 12.5 | 50 | [13] |
| Bioluminescent C. albicans |
C. albicans strain CAI4 (ura3::imm434/ura3::imm434) |
0.03 | 0.5 | 12.5 | 50 | [12] |
| C. albicans 17292c3367 | Venous catheter, caspofungin-resistant | 8 | 0.5 | 12.5 | 50 | This study |
| C. albicans 15343c3523 | Blood, caspofungin-resistant | 2 | 0.5 | 12.5 | 50 | This study |
| C. albicans 15351c6859 | Venous catheter, caspofungin-resistant | 4 | 1 | 12.5 | 50 | This study |
| C. albicans 14402c5521 | Abdominal lesions, fluconazole-resistant | 0.125 | 256 | 12.5 | 50 | This study |
Table 3.
Effect of PYR and TRI on the viability of drug-resistant C. albicans clinical isolates.
| Strain | Viability of C. albicans at 2× MIC (µg/mL) | Viability of C. albicans at 4x MIC (µg/mL) | ||
|---|---|---|---|---|
| TRI vs. Caspo (%) | TRI vs. Fluco (%) | PYR vs. Caspo (%) | PYR vs. Fluco (%) | |
| Caspofungin-resistant C. albicans 15343c3523 |
14.81 ± 7.8 vs. 83.56 ± 32.05 * | 24.96 ± 2.23 vs. 33.47 ± 18.6 * | ||
| Caspofungin-resistant C. albicans 15351c6859 |
13.49 ± 17.04 vs. 194.03 ± 8.014 * | 22.08 ± 13.95 vs. 231.89 ± 50.58 * | ||
| Caspofungin-resistant C. albicans 17292c3367 |
29.54 ± 17.8 vs. 130.76 ± 6.101 * | 10.79 ± 8.04 vs. 125.11 ± 62.04 * | ||
| Fluconazole-resistant C. albicans 14402c5521 |
11.30 ± 13.2 vs. 33.68 ± 26.2 * | 10.95 ± 9.49 vs. 29.12 ± 20.03 * | ||
* p < 0.05.
Figure 4.

Modulation of chitin and α-mannan in the C. albicans cell wall after antifungal challenge. (A) Confocal microscopy of C. albicans SC5314 strain challenged with PBS (CTL: a and b), caspofungin (Caspo: c and d), fluconazole (Fluco: e and f), PYR (g and h), or TRI (i and j) at their 1× MIC for 2 h. α-mannans were labelled with concanavalin A-rhodamine (red) and chitin was labelled with WGA-IFTC (green). Nuclei were labelled with DAPI. Scale bars represent 5 µm for (a, c, e, g, and j) and 1 µm for (b, d, f, h, and j). Magnification 100× with 1.4 numerical aperture. (B) Thickness of C. albicans cell wall α-mannan (µm). Error bars indicate standard deviations.
To determine whether PYR and TRI had an impact on biofilm formation, which is involved in resistance to antifungal agents by acting as a shield that delays or prevents drug diffusion to host cells, C. albicans biofilms were challenged with the two new antifungal compounds (Figure 5). The cell density of C. albicans challenged with caspofungin, fluconazole, PYR, or TRI at their MICs (0.03 µg/mL, 0.5 µg/mL, 12.5 µg/mL, and 50 µg/mL, respectively) was clearly lower when compared to that of untreated C. albicans. In this assay, the measurement of C. albicans cell-wall thickness was difficult from microscope images since the biofilm matrix was a dense and highly compact structure in all conditions (C. albicans alone or C. albicans challenged with antifungal compounds).
Figure 5.

Effect of PYR and TRI on C. albicans biofilm formation. C. albicans SC5314 cells were allowed to form a biofilm for 24 h. C. albicans biofilms were treated with caspofungin (Caspo), fluconazole (Fluco), PYR, or TRI at their 1× MIC for 24 h. CTL represents C. albicans alone without any antifungal treatment. Error bars indicate standard deviations.
2.3. PYR and TRI Reduced the Virulence of C. albicans in the C. elegans Infection Model
The effects of PYR and TRI on C. albicans virulence in vivo were investigated using a C. elegans nematode infection model. Nematodes infected with C. albicans were treated with either PYR or TRI (Figure 6). The survival of nematodes was followed daily by microscopic observation. C. albicans infection was observed to cause 85% mortality of C. elegans at day 4. PYR treatment of C. elegans infected with C. albicans at 12.5 µg/mL increased nematode survival to about 53% at day 4 while TRI treatment at 50 µg/mL protected the nematodes to about 30% survival indicating that PYR and TRI are active agents in prolonging nematode survival against C. albicans infection (Figure 6).
Figure 6.

Effects of PYR and TRI on C. albicans virulence in C. elegans. Nematodes infected with C. albicans SC5314 strain were examined for survival daily for 4 days and the percentage nematode survival was determined on day 4. Nematodes were considered dead when they did not respond to being touched with a platinum-wire pick. Ca + PYR represents C. elegans infected with C. albicans and treated with PYR. Ca + TRI represents C. elegans infected with C. albicans and treated with TRI. Data are presented as mean ± SD of four independent measurements.
To explore the effect of these two compounds on the immune response of C. elegans, we studied the expression of lys-1, lys-7, and cnc-4 involved in the antimicrobial response and pmk-1 (p38 MAPK signaling pathway), which concerns the immune reaction. In comparison with Escherichia coli OP50 control conditions, C. albicans increased the expression of lys-1, lys-7, cnc-4, and pmk-1 (Figure 7). PYR treatment of C. elegans infected with C. albicans enhanced the expression of lys-7 and cnc-4 while no significant change in pmk-1 expression was observed. TRI treatment of C. elegans infected with C. albicans induced the up-regulation of lys-1, lys-7, and cnc-4 while a reduction of pmk-1 expression was noted indicating that these antimicrobial genes, including the p38 MAPK pathway, have a significant role in C. elegans during C. albicans infection. Thus, PYR or TRI treatment enhanced the expression of antimicrobial genes that promote the elimination of C. albicans (Figure 7).
Figure 7.

Relative expression of antimicrobial peptides (lys-1, lys-7, and cnc-4) and the immune gene (pmk-1) in C. elegans infected with C. albicans. L4 nematodes were infected with C. albicans SC5314 lawns for 6 h and then treated with PYR or TRI at their 1x MIC for 12 h. E. coli represents C. elegans fed with E. coli. Ca represents C. elegans infected with C. albicans. Ca + PYR corresponds to C. elegans infected with C. albicans and treated with PYR. Ca + TRI represents C. elegans infected with C. albicans and treated with PYR. Data are presented as mean ± SD of four independent measurements.
3. Discussion
The development of new antifungal drugs remains a major challenge to overcoming the spread of drug-resistant clinically-relevant fungal pathogens, as strains are increasingly becoming less sensitive to conventional antifungal compounds [14,15].
In the present study, we selected two compounds, TRI and PYR, belonging to the triazine and pyridine chemical families, and showing antifungal activity against C. albicans and improved efficacy against fluconazole- or caspofungin-resistant clinical Candida strains.
Triazine is an important heterocyclic nucleus, which is a major component in some natural products such as reurhycin, toxoflavin, and fervenulin [16]. The chemistry of 1,2,4-triazines and their derivatives has attracted considerable attention owing to their broad spectrum of biological activity, including antifungal, anti-cancer, antiviral, cyclin-dependent kinase inhibitory, and anti-inflammatory activities [17,18,19,20,21]. Some 1,3,5-triazine derivatives, such as altretamine or almitrine have been approved by the FDA as antineoplastic agents or as respiratory stimulants, but none as antimicrobial agents. Moreover, very few reports have been published on the antimicrobial activity of such derivatives. In 1989, Reich et al. reported pyrido[3,4-e]-1,2,4-triazines and related heterocycles as having antifungal activity, with moderate activity against C. albicans, with MIC values ranging from 8 to 16 µg/mL for the lead compounds [22]. As shown in Figure 1, the reported compounds are bis-heterocycles and differ chemically from TRI.
Pyridin-2(1H)-compounds are naturally occurring products and display a variety of biological properties, including antimicrobial activity. Oxysporidinone was isolated after the fermentation of Fusarium oxysporum and has demonstrated antifungal activity against Aspergillus niger, Botrytis cinerea, Alternaria alternata, and Venturia inequalis, with MIC values of 10, 1, 50, and 10 µg/mL, respectively [23]. Its parent compound, funicolosin, is also a natural broad-spectrum antibiotic [24]. The related compound, ciclopirox, is a synthetic approved antifungal agent used as a topical treatment for superficial mycoses, particularly against Tinea versicolor [25]. As observed for TRI series, the compounds from the PYR series are chemically different from the reported oxysporidone, funicolosin, or ciclopirox parent molecules. Although these three reported antibiotics bear cyclic aliphatic moieties, PYR derivatives are composed of a pyridinone moiety decorated with two distinct aromatic rings.
Treatment with PYR and TRI resulted in a rapid reduction of C. albicans viability indicating that these compounds have fungicidal activity against C. albicans. Additionally, they reduced the viability of clinically isolated fluconazole- or caspofungin-resistant Candida strains. C. albicans cells in biofilms exhibit phenotypic traits, such as increased resistance to antifungal drugs and protection from host defenses that are dramatically different from their planktonic counterparts [5]. In the present study, we showed that challenge of C. albicans biofilms with either TRI or PYR contributed to a significant reduction in biofilm formation. Derivatives of PYR had low antifungal activity against C. albicans when compared to the original compound, while the derivative TRI1 did not have any antifungal activity at all.
The cell wall of C. albicans is the first point of contact between the fungus and the innate immune system [2]. The cell wall is composed of an outer layer enriched in mannan and mannosylated glycoproteins and an inner layer enriched in β-glucan and chitin [2,26]. We observed that PYR or TRI reduced the thickness of the mannan part of the C. albicans cell wall indicating that these two compounds can modulate the fungal cell wall and affect C. albicans virulence. However, no changes in chitin levels were observed in the fungal cells. These observations are consistent with our previous experimental studies, which showed that mannans were involved in Candida virulence and a deficiency in cell wall mannan contributed to a reduction in mouse mortality and intestinal inflammation induced by DSS [27].
In the C. elegans model, both TRI and PYR reduced the mortality of a high percentage of nematodes infected with C. albicans supporting the antifungal properties of these compounds against C. albicans infection [28]. C. elegans relies on its innate immune system to defend itself against fungal and bacterial pathogens by producing different antimicrobial proteins [29]. Caenacins and lysozyme, which are expressed in C. elegans, play a direct role against pathogens that infect worms via the intestinal lumen or cuticle [9,30,31]. Mallo et al. reported that overexpression of the lysozyme gene, lys-1, augmented the protection of C. elegans against Serratia marcescens infection [9]. In the current study, we showed that treatment with the two compounds enhanced the expression of antimicrobial genes (lys-1, lys-7, and cnc-4) in nematodes infected with C. albicans, indicating that PYR and TIR treatments not only had beneficial antifungal effects during infection but were also capable of improving the innate immune response of nematodes against C. albicans infection.
4. Materials and Methods
4.1. C. albicans Growth Conditions
C. albicans strain SC5314 used in this study was maintained at 4 °C in yeast peptone dextrose broth (YPD; 1% yeast extract, 2% dextrose, 2% peptone). To prepare the C. albicans suspension, fungal cells were cultured in Sabouraud dextrose broth (Sigma-Aldrich, St. Quentin Fallavier, France) for 24 h at 37 °C in a rotary shaker [32]. The fungal culture obtained was then centrifuged at 2500 rpm for 5 min and washed twice in PBS (phosphate-buffered saline). The clinical isolates were cultured in Sabouraud agar for 24–48 h. To identify these clinical isolates on the plates, the colonies were mixed with 1.5 μL of matrix solution (α-cyano-4-hydroxycinnamic acid; Bruker Daltonics, Bremen, Germany) dissolved in 50% acetonitrile, 47.5% water, and 2.5% trifluoroacetic acid, and analyzed by MALDI-TOF MS (Microflex-Bruker Daltonic, Bremen, Germany) [33,34].
4.2. Chemical Synthesis of PYR and TRI
Compound PYR was synthesized as described previously by Boisse et al. [35]. The reaction was performed between an amine (4-chloroaniline for Cpd PYR) and an allenic precursor (Figure 8). This methodology led to new 1-aryl-3,6-dimethyl-4-aminoaryl-2-pyridones 1 in good yields. These syntheses can be performed in two distinct steps, allowing for the possibility of introducing different substituents in the positions 1 and 4. Using this strategy, all compounds from the PYR series (1) were obtained (Figure 8). All the procedures and chemical characterizations of PYR compounds are detailed in the publication described by Boisse et al. [35].
Figure 8.

General pathway of PYR synthesis.
The synthesis of TRI compounds has not yet been described in the literature. The full synthetic study will be reported in due course in a global article of organic chemistry. Briefly, TRI compounds were obtained by reaction of arylamine with 2-ethinyl-4,6-dimethoxy-1,3,5-triazine in dichloromethane under magnetic stirring at room temperature for 2 days (Figure 9).
Figure 9.

General pathway for the synthesis of TRI compounds.
4.3. Antifungal Compounds
PYR and TRI were designed, synthesized, and provided by JUNIA (a graduate school of general engineering-Hautes Etudes d’Ingénieur, Lille, France). Several small aliquots were prepared for each compound and stored in the freezer at −20 °C. These fresh aliquots were adjusted to the appropriate dilutions in PBS for each experiment. PYR and TRI were used at their minimum inhibitory concentrations (MICs; 12.5 µg/mL and 50 µg/mL, respectively), diluted in PBS during the various in vitro and in vivo experiments. Commercially available caspofungin (Merck, Semoy, France) and fluconazole (Fresenius, Sèvres, France) were used as positive controls. The negative control was PBS.
4.4. Fungal Viability Assays
MICs were measured using the broth microdilution method and Clinical Laboratory Standards Institute (CLSI) procedures [36]. The screening was carried out by CO-ADD (Community for Antimicrobial Drug Discovery) and the University of Queensland (Santa Lucia, Australia). After the culture of C. albicans ATCC 90028 for 3 days on YPD agar at 30 °C, a C. albicans suspension of 1 × 106 to 5 × 106 CFU/mL was prepared [12]. This suspension was next diluted and added to each well of the PYR- or TRI-compound-containing plates giving a final cell density of fungi of 2.5 × 103 CFU/mL in a total volume of 50 μL. All plates were covered and incubated at 35 °C for 36 h without shaking. After the addition of resazurin (0.001% final concentration) and incubation at 35 °C for 2 h, the growth inhibition of C. albicans ATCC 90028 was assessed by measuring absorbance at 630 nm. The absorbance was assessed using a Biotek Multiflo Synergy HTX plate-reader. The percentage growth inhibition was assessed for each well, using the negative control (media only) and positive control (fungi without inhibitor) on the same plate. The MIC was determined as the lowest concentration at which fungal growth was completely inhibited (Table 1) [12], defined by inhibition of ≥80% for C. albicans (total inhibition in the case of PYR at 12.5 µg/mL and TRI at 50 µg/mL). These two compounds showed no cytotoxicity at fungicidal concentrations on different human lung, breast, renal, prostate, or colon cell lines [37]. A bioluminescent C. albicans strain was used for the viability assays [38]. This bioluminescent strain was suspended in PBS at a volume of 106 cells/well (96-well black plates, Chimney well). PYR, TRI, caspofungin, or fluconazole were then added at their final MIC (Table 1). The coelenterazine substrate was then added to the wells at a concentration of 2 μM [12]. Bioluminescence kinetics were then measured (at 0, 30, 60, and 120 min) and analyzed with a FLUOstar Omega Fluorometer (BMG Labtech, Champigny sur Marne, France). The positive control consisted of C. albicans challenged with PBS only. The viability of C. albicans was also evaluated by culture assay in the presence of PYR, TRI, fluconazole, and caspofungin. C. albicans strain SC5314 was suspended in PBS at a concentration of 105 C. albicans cells and antifungal compounds were then added. PYR, TRI, fluconazole, and caspofungin were used at their MIC (12.5 µg/mL, 50 µg/mL, 0.5 µg/mL, and 0.03 µg/mL, respectively). Serial dilutions ranging from 10−1 to 10−4 were performed on the samples suspended in PBS and 100 μL of each dilution was spread onto Sabouraud agar at 37 °C.
4.5. Effect of PYR and TRI on C. albicans Biofilm Formation
C. albicans at a concentration 107 colony-forming units (cfu) in 200 µL of RPMI was added to each well of a 96-well polystyrene plate (Greiner Bio-one) and incubated for 24 h. PYR and TRI were then added to the plate at their 1× MIC (12.5 µg/mL and 50 µg/mL, respectively) for 24 h. Caspofungin or fluconazole were also used at their MICs as positive controls. After three washes with PBS to remove non-adherent cells, 110 μL of crystal violet (0.4%; Fluka) was added to each well. After three washes with PBS, crystal violet staining was decolorized by adding 200 µL of ethanol to the plate. The absorbance of the decolorization solution that reflected the numbers of viable cells was measured at 550 nm using a spectrophotometer (FLUOstar; BMG Labtech, Champigny sur Marne, France). The results are presented as the mean of six replicates from two independent experiments.
4.6. C. elegans Survival Assay and RT-PCR Quantification of Antimicrobial Genes of C. elegans
C. albicans SC5314 strain was inoculated in 2 mL of Sabouraud broth and incubated at 36 °C for 24 h. Candida lawns were prepared by spreading 10 µL of the C. albicans culture onto plates containing solid BHI medium with amikacin (45 µg/mL). These plates were then incubated at 36 °C for 24 h. N2 wild-type C. elegans was grown on nematode growth medium seeded with E. coli OP50 as a food source at room temperature for 72 h, until they reached the L4 stage. Approximately 100 worms were selected and washed with M9 buffer containing 90 µg/mL amikacin to eliminate E. coli. These worms were then picked onto the Candida lawn and allowed to feed for 6 h. The nematodes were washed three times with M9 buffer to remove C. albicans cells from their cuticles. Approximately 70–80 nematodes were then picked to wells in a 6-well microtiter dish that contained 2 mL of liquid 80% M9 buffer, 20% BHI, 10 µg/mL cholesterol in ethanol, and 90 µg/mL amikacin. TRI and PYR at their MICs (12.5 µg/mL and 50 µg/mL, respectively) were added to each well. The plates were incubated at room temperature for 12 h. The worms were examined for survival daily for 4 days. Nematodes were considered dead when they did not respond to being touched by a platinum-wire pick.
For the RT-PCR assay, total RNA was isolated from N2 after 12 h of antifungal treatment using a NucleoSpin RNA® kit (Macherey-Nagel, Hoerdt, France). Nematode RNA was quantified by spectrophotometry (Nanodrop; Nyxor Biotech, Paris, France). cDNA synthesis was carried out using a High Capacity DNA Reverse Transcription (RT) kit, with Master Mix (Applied Biosystems, CA, USA). To amplify the cDNA, Fast SYBR green (Applied Biosystems) was employed in a one-step system (Applied Biosystems) [39]. SYBR green dye intensity was assessed using one-step software. All results were normalized to the reference gene, act-2.
4.7. Statistical Analysis
The Mann–Whitney U test was used to analyze the differences between the groups and the results were considered to be statistically significant when the p value was as follows: p < 0.05; p < 0.01; p < 0.001. All statistical analyses were carried out using GraphPad Prism version 6 (GraphPad, La Jolla, CA, USA).
5. Conclusions
In conclusion, two series of triazine (TRI) and pyridinone (PYR) derivatives were selected for their antifungal potential. These series share some similarities such as a 6-membered N-heterocycle, a free -NH- link and a para-substituted aromatic moiety. Whereas in series TRI, the p-methoxyaniline was the most effective ring for antifungal activity, it appeared that in series PYR, p-chloroaniline was the best choice. TRI and PYR were found to have fungicidal activity against C. albicans which was more pronounced than their analogues, and both reduced the viability of clinically isolated fluconazole- or caspofungin-resistant Candida strains. PYR and TRI decreased C. albicans biofilm formation and decreased the thickness of the mannan part of the cell wall. In a C. elegans nematode infection model, PYR and TRI decreased the mortality of nematodes infected with C. albicans and enhanced the expression of antimicrobial genes that promote the elimination of C. albicans. Overall, this study provides evidence that PYR and TRI are able to reduce the viability of drug-resistant C. albicans clinical isolates and offer promising therapeutic applications against biofilm-associated C. albicans infections. The identification of these two compounds PYR and TRI will enable us to widen our study to in silico approaches and to predict the possible binding sites of these two compounds in due course.
Acknowledgments
The authors thank Thomas Boulin (Université Claude Bernard Lyon 1) for providing the nematodes N2 and E. coli OP50. The authors thank Antonino Bongiovanni, Clovis Bortolus, and Nadine François for their excellent technical assistance. The authors would like to thank the Community for Antimicrobial Drug Discovery (CO-ADD), funded by the Welcome Trust (UK), and the University of Queensland (Australia) for performing the antifungal screening of compounds on Candida albicans strain ATCC 90028.
Author Contributions
L.M., M.B., R.C., B.D., A.G. and S.J. performed the experiments. L.M., R.C., B.D., B.S. and S.J. analyzed the data. L.M., R.C., B.D., B.S. and S.J. interpreted the results of the experiments. M.B. and A.G. contributed to the reagents/materials/analysis tools. S.J. designed the experiments. S.J. drafted the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially funded by the Agence Nationale de la Recherche (ANR) in the setting of project “InnateFun”, promotional reference ANR-16-IFEC-0003-05, in the “Infect-ERA” program.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Poulain D., Sendid B., Standaert-Vitse A., Fradin C., Jouault T., Jawhara S., Colombel J.F. Yeasts: Neglected pathogens. Dig. Dis. 2009;27:104–110. doi: 10.1159/000268129. [DOI] [PubMed] [Google Scholar]
- 2.Jawhara S. How Fungal Glycans Modulate Platelet Activation via Toll-Like Receptors Contributing to the Escape of Candida albicans from the Immune Response. Antibiotics. 2020;9:385. doi: 10.3390/antibiotics9070385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Billamboz M., Fatima Z., Hameed S., Jawhara S. Promising Drug Candidates and New Strategies for Fighting against the Emerging Superbug Candida auris. Microorganisms. 2021;9:634. doi: 10.3390/microorganisms9030634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Arastehfar A., Gabaldon T., Garcia-Rubio R., Jenks J.D., Hoenigl M., Salzer H.J.F., Ilkit M., Lass-Florl C., Perlin D.S. Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium. Antibiotics. 2020;9:877. doi: 10.3390/antibiotics9120877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Atriwal T., Azeem K., Husain F.M., Hussain A., Khan M.N., Alajmi M.F., Abid M. Mechanistic Understanding of Candida albicans Biofilm Formation and Approaches for Its Inhibition. Front. Microbiol. 2021;12:638609. doi: 10.3389/fmicb.2021.638609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Borghi E., Morace G., Borgo F., Rajendran R., Sherry L., Nile C., Ramage G. New strategic insights into managing fungal biofilms. Front. Microbiol. 2015;6:1077. doi: 10.3389/fmicb.2015.01077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Powell J.R., Ausubel F.M. Methods in Molecular Biology. Volume 415. Humana Press; Totowa, NJ, USA: 2008. Models of Caenorhabditis elegans infection by bacterial and fungal pathogens; pp. 403–427. [DOI] [PubMed] [Google Scholar]
- 8.Irazoqui J.E., Urbach J.M., Ausubel F.M. Evolution of host innate defence: Insights from Caenorhabditis elegans and primitive invertebrates. Nat. Rev. Immunol. 2010;10:47–58. doi: 10.1038/nri2689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mallo G.V., Kurz C.L., Couillault C., Pujol N., Granjeaud S., Kohara Y., Ewbank J.J. Inducible antibacterial defense system in C. elegans. Curr. Biol. 2002;12:1209–1214. doi: 10.1016/S0960-9822(02)00928-4. [DOI] [PubMed] [Google Scholar]
- 10.Troemel E.R., Chu S.W., Reinke V., Lee S.S., Ausubel F.M., Kim D.H. p38 MAPK regulates expression of immune response genes and contributes to longevity in C. elegans. PLoS Genet. 2006;2:e183. doi: 10.1371/journal.pgen.0020183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ewbank J.J. WormBook. 2006. [(accessed on 23 January 2006)]. Signaling in the immune response; pp. 1–12. Available online: http://www.wormbook.org/chapters/www_signalingimmuneresponse/signalingimmuneresponse.html. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bortolus C., Billamboz M., Charlet R., Lecointe K., Sendid B., Ghinet A., Jawhara S. A Small Aromatic Compound Has Antifungal Properties and Potential Anti-Inflammatory Effects against Intestinal Inflammation. Int. J. Mol. Sci. 2019;20:321. doi: 10.3390/ijms20020321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jawhara S., Poulain D. Saccharomyces boulardii decreases inflammation and intestinal colonization by Candida albicans in a mouse model of chemically-induced colitis. Med. Mycol. 2007;45:691–700. doi: 10.1080/13693780701523013. [DOI] [PubMed] [Google Scholar]
- 14.Sanguinetti M., Posteraro B., Lass-Florl C. Antifungal drug resistance among Candida species: Mechanisms and clinical impact. Mycoses. 2015;58:2–13. doi: 10.1111/myc.12330. [DOI] [PubMed] [Google Scholar]
- 15.Maubon D., Garnaud C., Calandra T., Sanglard D., Cornet M. Resistance of Candida spp. to antifungal drugs in the ICU: Where are we now? Intensive Care Med. 2014;40:1241–1255. doi: 10.1007/s00134-014-3404-7. [DOI] [PubMed] [Google Scholar]
- 16.Wang G., Li X., Wang J., Xie Z., Li L., Chen M., Chen S., Peng Y. Synthesis, molecular docking and alpha-glucosidase inhibition of 2-((5,6-diphenyl-1,2,4-triazin-3-yl)thio)-N-arylacetamides. Bioorg. Med. Chem. Lett. 2017;27:1115–1118. doi: 10.1016/j.bmcl.2017.01.094. [DOI] [PubMed] [Google Scholar]
- 17.Kumar R., Sirohi T.S., Singh H., Yadav R., Roy R.K., Chaudhary A., Pandeya S.N. 1,2,4-triazine analogs as novel class of therapeutic agents. Mini-Rev. Rev. Med. Chem. 2014;14:168–207. doi: 10.2174/1389557514666140131111837. [DOI] [PubMed] [Google Scholar]
- 18.Sangshetti J.N., Shinde D.B. One pot synthesis and SAR of some novel 3-substituted 5,6-diphenyl-1,2,4-triazines as antifungal agents. Bioorg. Med. Chem. Lett. 2010;20:742–745. doi: 10.1016/j.bmcl.2009.11.048. [DOI] [PubMed] [Google Scholar]
- 19.Ahuja P., Siddiqui N. Anticonvulsant evaluation of clubbed indole-1,2,4-triazine derivatives: A synthetic approach. Eur. J. Med. Chem. 2014;80:509–522. doi: 10.1016/j.ejmech.2014.04.043. [DOI] [PubMed] [Google Scholar]
- 20.Zhan P., Li X., Li Z., Chen X., Tian Y., Chen W., Liu X., Pannecouque C., De Clercq E. Structure-based bioisosterism design, synthesis and biological evaluation of novel 1,2,4-triazin-6-ylthioacetamides as potent HIV-1 NNRTIs. Bioorg. Med. Chem. Lett. 2012;22:7155–7162. doi: 10.1016/j.bmcl.2012.09.062. [DOI] [PubMed] [Google Scholar]
- 21.Krauth F., Dahse H.M., Ruttinger H.H., Frohberg P. Synthesis and characterization of novel 1,2,4-triazine derivatives with antiproliferative activity. Bioorg. Med. Chem. 2010;18:1816–1821. doi: 10.1016/j.bmc.2010.01.053. [DOI] [PubMed] [Google Scholar]
- 22.Reich M.F., Fabio P.F., Lee V.J., Kuck N.A., Testa R.T. Pyrido[3,4-e]-1,2,4-triazines and related heterocycles as potential antifungal agents. J. Med. Chem. 1989;32:2474–2485. doi: 10.1021/jm00131a010. [DOI] [PubMed] [Google Scholar]
- 23.Breinhold J., Ludvigsen S., Rassing B.R., Rosendahl C.N., Nielsen S.E., Olsen C.E. Oxysporidinone: A novel, antifungal N-methyl-4-hydroxy-2-pyridone from Fusarium oxysporum. J. Nat. Prod. 1997;60:33–35. doi: 10.1021/np9605596. [DOI] [PubMed] [Google Scholar]
- 24.Ando K., Matsuura I., Nawata Y., Endo H., Sasaki H., Okytomi T., Saehi T., Tamura G. Funiculosin, a new antibiotic. II. Structure elucidation and antifungal activity. J. Antibiot. 1978;31:533–538. doi: 10.7164/antibiotics.31.533. [DOI] [PubMed] [Google Scholar]
- 25.Gupta A.K., Skinner A.R. Ciclopirox for the treatment of superficial fungal infections: A review. Int. J. Dermatol. 2003;42:3–9. doi: 10.1046/j.1365-4362.42.s1.2.x. [DOI] [PubMed] [Google Scholar]
- 26.Vancraeyneste H., Charlet R., Guerardel Y., Choteau L., Bauters A., Tardivel M., Francois N., Dubuquoy L., Soloviev D., Poulain D., et al. Short fungal fractions of beta-1,3 glucans affect platelet activation. Am. J. Physiol. Heart Circ. Physiol. 2016;311:H725–H734. doi: 10.1152/ajpheart.00907.2015. [DOI] [PubMed] [Google Scholar]
- 27.Jawhara S., Mogensen E., Maggiotto F., Fradin C., Sarazin A., Dubuquoy L., Maes E., Guerardel Y., Janbon G., Poulain D. Murine model of dextran sulfate sodium-induced colitis reveals Candida glabrata virulence and contribution of beta-mannosyltransferases. J. Biol. Chem. 2012;287:11313–11324. doi: 10.1074/jbc.M111.329300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tampakakis E., Okoli I., Mylonakis E. A C. elegans-based, whole animal, in vivo screen for the identification of antifungal compounds. Nat. Protoc. 2008;3:1925–1931. doi: 10.1038/nprot.2008.193. [DOI] [PubMed] [Google Scholar]
- 29.Kamaladevi A., Balamurugan K. Role of PMK-1/p38 MAPK defense in Caenorhabditis elegans against Klebsiella pneumoniae infection during host-pathogen interaction. Pathog. Dis. 2015;73:ftv021. doi: 10.1093/femspd/ftv021. [DOI] [PubMed] [Google Scholar]
- 30.Souza A.C.R., Fuchs B.B., Alves V.S., Jayamani E., Colombo A.L., Mylonakis E. Pathogenesis of the Candida parapsilosis Complex in the Model Host Caenorhabditis elegans. Genes. 2018;9:401. doi: 10.3390/genes9080401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Engelmann I., Griffon A., Tichit L., Montanana-Sanchis F., Wang G., Reinke V., Waterston R.H., Hillier L.W., Ewbank J.J. A comprehensive analysis of gene expression changes provoked by bacterial and fungal infection in C. elegans. PLoS ONE. 2011;6:e19055. doi: 10.1371/journal.pone.0019055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jawhara S., Thuru X., Standaert-Vitse A., Jouault T., Mordon S., Sendid B., Desreumaux P., Poulain D. Colonization of mice by Candida albicans is promoted by chemically induced colitis and augments inflammatory responses through galectin-3. J. Infect. Dis. 2008;197:972–980. doi: 10.1086/528990. [DOI] [PubMed] [Google Scholar]
- 33.Charlet R., Pruvost Y., Tumba G., Istel F., Poulain D., Kuchler K., Sendid B., Jawhara S. Remodeling of the Candida glabrata cell wall in the gastrointestinal tract affects the gut microbiota and the immune response. Sci. Rep. 2018;8:3316. doi: 10.1038/s41598-018-21422-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Charlet R., Bortolus C., Barbet M., Sendid B., Jawhara S. A decrease in anaerobic bacteria promotes Candida glabrata overgrowth while beta-glucan treatment restores the gut microbiota and attenuates colitis. Gut Pathog. 2018;10:50. doi: 10.1186/s13099-018-0277-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Boisse T., Rigo B., Millet R., Hénichart J.P. From dicarbonylallene to 1-aryl-3,6-dimethyl-4-aminoaryl-2-pyridones: A one-pot versatile and uncatalyzed synthesis. Tetrahedron. 2007;63:10511–10520. doi: 10.1016/j.tet.2007.07.079. [DOI] [Google Scholar]
- 36.Bustamante B., Martins M.A., Bonfietti L.X., Szeszs M.W., Jacobs J., Garcia C., Melhem M.S.C. Species distribution and antifungal susceptibility profile of Candida isolates from bloodstream infections in Lima, Peru. J. Med. Microbiol. 2014;63:855–860. doi: 10.1099/jmm.0.071167-0. [DOI] [PubMed] [Google Scholar]
- 37.Mosmann T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods. 1983;65:55–63. doi: 10.1016/0022-1759(83)90303-4. [DOI] [PubMed] [Google Scholar]
- 38.Doyle T.C., Nawotka K.A., Kawahara C.B., Francis K.P., Contag P.R. Visualizing fungal infections in living mice using bioluminescent pathogenic Candida albicans strains transformed with the firefly luciferase gene. Microb. Pathog. 2006;40:82–90. doi: 10.1016/j.micpath.2005.11.003. [DOI] [PubMed] [Google Scholar]
- 39.Dumortier C., Charlet R., Bettaieb A., Jawhara S. H89 Treatment Reduces Intestinal Inflammation and Candida albicans Overgrowth in Mice. Microorganisms. 2020;8:2039. doi: 10.3390/microorganisms8122039. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data presented in this study are available in the article.