

Abstract
The EFSA Plant Health Panel performed a pest categorisation of Fusarium oxysporum f. sp. cubense Tropical Race 4 (Foc TR4), an ascomycete fungus causing Fusarium wilt (Panama disease) on Musa spp. Foc TR4 is pathogenic to the commercial banana varieties including those of the ‘Cavendish’ group and is considered as the most destructive among Foc haplotypes. Uncertainty exists on the host range of Foc TR4, since it has not been demonstrated whether it can infect plant species other than Musa spp., which were previously reported as hosts of other Foc races. Foc TR4 is morphologically and physiologically identical to other representatives of the Fusarium oxysporum Species Complex (FOSC), but all Foc TR4 isolates belong to a single clonal lineage within the vegetative compatibility groups 01213‐01216. Several PCR protocols are described in the literature, but their specificity has been questioned as they may generate false positives. The pathogen is not included in EU Commission Implementing Regulation 2019/2072 and is not reported as present in the EU territory. Several potential entry pathways and means of spread were identified, including host plants for planting other than vitroplants, fresh fruits and leaves of host plants, soil and other substrates originating in infested third countries. Host availability and climate suitability occurring in some areas of the EU are favourable for the establishment of Foc TR4. Being a soil‐borne pathogen, eradication of Foc TR4 once it enters a new area is very difficult. Therefore, effective quarantine measures are essential in pathogen‐free areas. Although not specifically targeting against Foc TR4, phytosanitary measures are currently available to prevent the introduction of the pathogen into the EU. Considering that banana‐growing EU countries account for over 12% of the EU banana supply, it is expected that the economic impact of Foc TR4 on the European banana production areas would be devastating. Foc TR4 satisfies the criteria that are within the remit of EFSA to assess for this pathogen to be regarded as a potential Union quarantine pest.
Keywords: Pest risk, plant health, plant pest, quarantine, banana, Fusarium wilt, Panama disease
1. Introduction
1.1. Background and Terms of Reference as provided by the requestor
1.1.1. Background
The new Plant Health Regulation (EU) 2016/2031, on the protective measures against pests of plants, is applying from 14 December 2019. Conditions are laid down in this legislation in order for pests to qualify for listing as Union quarantine pests, protected zone quarantine pests or Union regulated non‐quarantine pests. The lists of the EU regulated pests together with the associated import or internal movement requirements of commodities are included in Commission Implementing Regulation (EU) 2019/2072. Additionally, as stipulated in the Commission Implementing Regulation 2018/2019, certain commodities are provisionally prohibited to enter in the EU (high risk plants, HRP). EFSA is performing the risk assessment of the dossiers submitted by exporting to the EU countries of the HRP commodities, as stipulated in Commission Implementing Regulation 2018/2018. Furthermore, EFSA has evaluated a number of requests from exporting to the EU countries for derogations from specific EU import requirements.
In line with the principles of the new plant health law, the European Commission with the Member States are discussing monthly the reports of the interceptions and the outbreaks of pests notified by the Member States. Notifications of an imminent danger from pests that may fulfil the conditions for inclusion in the list of the Union quarantine pest are included. Furthermore, EFSA has been performing horizon scanning of media and literature.
As a follow‐up of the above‐mentioned activities (reporting of interceptions and outbreaks, HRP, derogation requests and horizon scanning), a number of pests of concern have been identified. EFSA is requested to provide scientific opinions for these pests, in view of their potential inclusion by the risk manager in the lists of Commission Implementing Regulation (EU) 2019/2072 and the inclusion of specific import requirements for relevant host commodities, when deemed necessary by the risk manager.
1.1.2. Terms of Reference
EFSA is requested, pursuant to Article 29(1) of Regulation (EC) No 178/2002, to provide scientific opinions in the field of plant health.
EFSA is requested to deliver 53 pest categorisations for the pests listed in Annex 1A, 1B, 1D and 1E (for more details see mandate M‐2021‐00027 on the Open.EFSA portal ). Additionally, EFSA is requested to perform pest categorisations for the pests so far not regulated in the EU, identified as pests potentially associated with a commodity in the commodity risk assessments of the HRP dossiers (Annex 1C; for more details see mandate M‐2021‐00027 on the Open.EFSA portal ). Such pest categorisations are needed in the case where there are not available risk assessments for the EU.
When the pests of Annex 1A are qualifying as potential Union quarantine pests, EFSA should proceed to phase 2 risk assessment. The opinions should address entry pathways, spread, establishment, impact and include a risk reduction options analysis.
Additionally, EFSA is requested to develop further the quantitative methodology currently followed for risk assessment, in order to have the possibility to deliver an express risk assessment methodology. Such methodological development should take into account the EFSA Plant Health Panel Guidance on quantitative pest risk assessment and the experience obtained during its implementation for the Union candidate priority pests and for the likelihood of pest freedom at entry for the commodity risk assessment of High Risk Plants.
1.2. Interpretation of the Terms of Reference
Fusarium oxysporum f. sp. cubense Tropical Race 4 is one of a number of pests listed in Annex 1 to the Terms of Reference (ToR) (Section 1.1.2.1) to be subject to pest categorisation to determine whether it fulfils the criteria of a potential Union quarantine pest for the area of the EU excluding Ceuta, Melilla and the outermost regions of Member States referred to in Article 355(1) of the Treaty on the Functioning of the European Union (TFEU), other than Madeira and the Azores, and so inform European Commission decision‐making as to its appropriateness for potential inclusion in the lists of pests of Commission Implementing Regulation (EU) 2019/ 2072. If a pest fulfils the criteria to be potentially listed as a Union quarantine pest, risk reduction options for relevant host commodities will be identified.
2. Data and methodologies
2.1. Data
2.1.1. Literature search
A literature search on Fusarium oxysporum f. sp. cubense Tropical Race 4 was conducted at the beginning of the categorisation in the ISI Web of Science bibliographic database, using the scientific name of the pest as search term. Papers relevant for the pest categorisation were reviewed, and further references and information were obtained from experts, as well as from citations within the references and grey literature.
2.1.2. Database search
Pest information, on host(s) and distribution, was retrieved from the European and Mediterranean Plant Protection Organization (EPPO) Global Database (EPPO, online), the CABI databases and scientific literature databases as referred above in Section 2.1.1.
Data about the import of commodity types that could potentially provide a pathway for the pest to enter the EU and about the area of hosts grown in the EU were obtained from EUROSTAT (Statistical Office of the European Communities).
The Europhyt and TRACES databases were consulted for pest‐specific notifications on interceptions and outbreaks. Europhyt was a web‐based network run by the Directorate General for Health and Food Safety (DG SANTÉ) of the European Commission as a subproject of PHYSAN (Phyto‐Sanitary Controls) specifically concerned with plant health information. TRACES is the European Commission's multilingual online platform for sanitary and phytosanitary certification required for the importation of animals, animal products, food and feed of non‐animal origin and plants into the European Union, and the intra‐EU trade and EU exports of animals and certain animal products. Up until May 2020, the Europhyt database managed the notifications of interceptions of plants or plant products that do not comply with EU legislation, as well as notifications of plant pests detected in the territory of the Member States and the phytosanitary measures taken to eradicate or avoid their spread. The recording of interceptions switched from Europhyt interceptions to TRACES in May 2020.
2.2. Methodologies
The Panel performed the pest categorisation for Fusarium oxysporum f. sp. cubense Tropical Race 4, following guiding principles and steps presented in the EFSA guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018), the EFSA guidance on the use of the weight of evidence approach in scientific assessments (EFSA Scientific Committee, 2017) and the International Standards for Phytosanitary Measures No. 11 (FAO, 2013) and No. 21 (FAO, 2004).
The criteria to be considered when categorising a pest as a potential Union quarantine pest (QP) is given in Regulation (EU) 2016/2031 Article 3 and Annex I, Section 1 to this Regulation. Table 1 presents the Regulation (EU) 2016/2031 pest categorisation criteria on which the Panel bases its conclusions. In judging whether a criterion is met the Panel uses its best professional judgement (EFSA Scientific Committee, 2017) by integrating a range of evidence from a variety of sources (as presented above in Section 2.1) to reach an informed conclusion as to whether or not a criterion is satisfied.
Table 1.
Pest categorisation criteria under evaluation, as defined in Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column)
| Criterion of pest categorisation | Criterion in Regulation (EU) 2016/2031 regarding Union quarantine pest (article 3) |
|---|---|
| Identity of the pest ( Section 3.1 ) | Is the identity of the pest established, or has it been shown to produce consistent symptoms and to be transmissible? |
| Absence/presence of the pest in the EU territory ( Section 3.2 ) |
Is the pest present in the EU territory? If present, is the pest widely distributed within the EU? Describe the pest distribution briefly |
| Pest potential for entry, establishment and spread in the EU territory ( Section 3.4 ) | Is the pest able to enter into, become established in, and spread within, the EU territory? If yes, briefly list the pathways |
| Potential for consequences in the EU territory ( Section 3.5 ) | Would the pests’ introduction have an economic or environmental impact on the EU territory? |
|
Available measures (Specific import requirements) ( Section 3.6 ) |
Are there measures available to prevent the entry into the EU such that the likelihood of introduction becomes mitigated? |
| Conclusion of pest categorisation ( Section 4 ) | A statement as to whether (1) all criteria assessed by EFSA above for consideration as a potential quarantine pest were met and (2) if not, which one(s) were not met. |
The Panel’s conclusions are formulated respecting its remit and particularly with regard to the principle of separation between risk assessment and risk management (EFSA founding regulation (EU) No 178/2002); therefore, instead of determining whether the pest is likely to have an unacceptable impact, deemed to be a risk management decision, the Panel will present a summary of the observed impacts in the areas where the pest occurs, and make a judgement about potential likely impacts in the EU. Whilst the Panel may quote impacts reported from areas where the pest occurs in monetary terms, the Panel will seek to express potential EU impacts in terms of yield and quality losses and not in monetary terms, in agreement with [insert appropriate reference to EFSA not reporting impacts in financial/monetary terms] Article 3 (d) of Regulation (EU) 2016/2031 refers to unacceptable social impact as a criterion for quarantine pest status. Assessing social impact is outside the remit of the Panel.
3. Pest categorisation
3.1. Identity and biology of the pest
3.1.1. Identity and taxonomy
Is the identity of the pest established, or has it been shown to produce consistent symptoms and/or to be transmissible?
Yes, the identity of Fusarium oxysporum f. sp. cubense Tropical Race 4 (Foc TR4) is well established and it has been shown to produce consistent symptoms and to be transmissible.
Fusarium oxysporum f. sp. cubense Tropical Race 4 (Foc TR4) is a plant pathogenic ascomycete fungus of the family Nectriaceae that causes Fusarium wilt (or Panama disease) of banana, the most destructive disease of banana plantations.
Fusarium oxysporum Schlechtend.:Fr. (Fo) belongs to a group of cosmopolitan, soil‐borne asexual filamentous fungi that are taxonomically collated in the Fusarium oxysporum Species Complex (FOSC; Baayen et al., 2000). The members of FOSC share a set of morphological features, principally the shape of the macroconidium, the structure of the phialide and the formation and arrangement of chlamydospores. The FOSC includes mainly saprophytic strains, but also many plant pathogens which are capable to induce diseases, ranging from vascular wilt, root and crown rot, and damping‐off on a wide range of plant species. The pathogenic strains possess a high level of host specificity and are classified on this basis into more than 150 host‐specific formae speciales, with those infecting bananas being placed in the forma specialis ‘cubense’ (Foc). Each forma specialis can be further subdivided into physiological races based on their virulence on differential host cultivars.
In the case of Foc, there are three well‐known races identified by their pathogenicity to specific clonal triploid banana hosts (Stover and Buddenhagen, 1986; Stover and Simmonds, 1987; Stover, 1990; Ploetz et al., 2015): (i) Foc Race 1, which destroyed the well‐established ‘Gros Michel’ cultivar (AAA genome group) banana industry that was grown in monoculture plantations in the Americas, Africa and in the Far East during the 1900s and can also cause disease on other cultivars, such as ‘Silk’ (AAB), ‘Pome’ (AAB), ‘Pisang Awak’ (ABB) and ‘Maqueno’ (AAB) (Ploetz, 2006); (ii) Foc Race 2, which infects cooking bananas especially those in the Bluggoe subgroup (ABB) (Su et al., 1977; Ploetz, 2006); and (iii) Foc Race 4, which has been known to cause disease on cultivars susceptible to race 1 and race 2, and more importantly, on cultivars of the ‘Cavendish’ subgroup (AAA) resistant to Foc race 1. Foc Race 4 has been further split into Subtropical Race 4 (SR4), which infects cultivars of the ‘Cavendish’ subgroup and cultivars susceptible to Foc race 1 and Foc race 2 in the subtropics, where plants are exposed to stress conditions, such as low temperatures and drought (Waite and Stover, 1960; Stover and Malo, 1972), and Foc Tropical Race 4 (Foc TR4), which infects many of the same cultivars as SR4 under both tropical and subtropical conditions without predisposing factors (Pegg et al., 1996; Ploetz et al., 2015). Race 3, which was reported in the past to infect Heliconia spp. (tropical American banana relatives) (Waite, 1963), is no longer recognised as a banana pathogen (Ploetz et al., 2015).
Panama disease or Fusarium wilt, caused by Foc, is the most important lethal disease of banana, and was first described in Australia in 1874 (Pegg et al., 2019). Fusarium wilt was first observed in South‐East Asia, then in Central America, in Costa Rica and Panama. Since the 1990s, it has been identified in every commercial banana‐growing area in the world. Only a few regions have remained free of Foc, including some islands in the South Pacific and Melanesia and a few countries bordering the Mediterranean. The pathogen destroyed the well‐established ‘Gros Michel’ cultivar banana industry. The cultivar ‘Gros Michel’ was subsequently replaced by a resistant varietal group discovered in South‐East Asia, ‘Cavendish’ (AAA), which includes commercially grown cultivars such as ‘Grand Naine’ and ‘Williams’, accounting for most of the present‐day world trade. However, the disease recurred in the 1970s in the south‐east Asian continent and Australia with the detection of a new race of Foc, namely Tropical Race 4 (TR4). TR4 is one of the six clonal lineages of Foc able to attack ‘Cavendish bananas’ of the species Musa acuminata (Ploetz, 2006). Foc TR4 was first observed in Taiwan in 1967, but its importance as an invasive pathogen was identified in the 1990s when it was isolated from diseased ‘Cavendish’ plants in Indonesia and Malaysia (Buddenhagen, 2009). Since then, Foc TR4 has been reported to cause wilt, foliar yellowing, vascular necrosis and eventually death on species of Musa (Pérez‐Vicente et al., 2014). Foc TR4 is strongly pathogenic to the ‘Cavendish’ group of banana cultivars, but is considered the most destructive Foc haplotype because it affects more Musa species and cultivars than the other haplotypes of Foc (Cheng et al., 2019; Curry, 2020). Foc TR4 is morphologically and physiologically identical to other representatives of the Fusarium oxysporum Species Complex (FOSC). It can be detected and identified within infected plant tissues using molecular tools (Dita et al., 2010; Lin et al., 2013). Nevertheless, reliable identification of Foc TR4 also requires the determination of the vegetative compatibility groups (VCGs) (see Section 3.1.5 Detection and identification of the pest).
The EPPO Global Database (EPPO, online) provide the following taxonomic identification for F. oxysporum f. sp. cubense Tropical Race 4 (Foc TR4):
Preferred Scientific Name: Fusarium oxysporum f. sp. cubense Tropical Race 4.
Order: Hypocreales.
Family: Nectriaceae.
Genus: Fusarium.
Species: Fusarium oxysporum f. sp. cubense Tropical Race 4.
Common names: Foc TR4, Panama disease Tropical Race 4, TR4.
The EPPO code 1 (Griessinger and Roy, 2015; EPPO, 2019) for F. oxysporum f. sp. cubense Tropical Race 4 is FUSAC4 (EPPO, online).
In 2019, Maryani et al. (2019) proposed to consider Fusarium oxysporum f. sp. cubense Tropical Race 4 as a distinct species called Fusarium odoratissimum. However, this proposal has been questioned by Torres Bedoya et al. (2021) who, based on the results of their studies, considered that this change was premature and not sufficiently substantiated. Therefore, in the present Pest categorisation, the scientific name Fusarium oxysporum f. sp. cubense Tropical Race 4 (Foc TR4) will be used, although the binomial F. odoratissimum was also included in the literature search.
3.1.2. Biology of the pest
Foc TR4, as any other members of the FOSC, produces three types of spores: microconidia (5–7 × 2.5–3 µm), which are oval‐shaped spores and the most commonly produced spore type within infected plants; macroconidia (22–36 × 4–5 µm), which are larger, thin‐walled spores, most frequently produced on the surface of infected plants; chlamydospores (9 × 7 µm), which are round, thick‐walled asexual resting spores formed in older hyphal cells (terminal or intercalary) or by the transformation of macroconidia cells (Wardlaw, 1961).
Foc grows in a range of temperatures between 9°C and 38°C under in vitro conditions, with an optimum growth between 23°C and 27°C (Pérez‐Vicente et al., 2014). Usually, the disease is more intense during the warmer and wetter months of the year (Pérez‐Vicente et al., 2014). Plant infection occurs as response to primary and secondary root exudates. Major roots and rhizomes are not usually infected directly. Upon germination of spores, hyphae adhere to and directly penetrate the epidermis; the pathogen then advances intracellularly through the cortex and reaches the xylem vessels, where it produces microconidia and toxins that move upstream in the plant sap, colonising neighbouring vessels and producing new fungal structures (Pérez‐Vicente et al., 2014). The xylem vessels become discoloured and blocked, interrupting the supply of water and thereby causing plant wilting (Curry, 2020). Toxin production and/or host defence response including tylose and gum production, and vessel shrinkage due to parenchymatic companion cell growth enhance the impairment of water and nutrient transport. In resistant cultivars, tyloses and gums are produced earlier and far more rapidly than in susceptible cultivars (Ploetz et al., 2015).
The first internal symptoms (a reddish‐brown discoloration of the xylem vessels) appear in feeder roots, which represent the initial sites of infection. Vascular discoloration then extends to the rhizome and proceeds by including large portions of the pseudostem (Ploetz, 2006). On plants that are older than 4 months, the external leaves turn yellow or split longitudinally at the base. At a later phase of the disease progress, younger leaves wilt and collapse until the entire canopy wilts. Systemic invasion of the pseudostem does not occur in tolerant cultivars, due to the obstruction of the pathogen by tyloses and gums. However, in susceptible cultivars, systemic colonisation proceeds in advance of the production of these host products (Ploetz et al., 2015). According to Rishbeth (1957), the pathogen may have a long (2–6 months) incubation period i.e. the period between infection and symptom development.
As for other phytopathogenic formae speciales of F. oxysporum, when the plant dies, the fungus grows outside the xylem, invades the surrounding tissues and develops masses of macroconidia and chlamydospores. Through the production of chlamydospores and (for some isolates) of sclerotia, Foc may survive in soil up to 30 years in the absence of banana crops (Stover, 1962; Buddenhagen, 2009). Noteworthily, chlamydospores of TR4 are produced once the banana host is invaded, even before any external symptoms are visible (Li et al., 2011), and not just after the death of the plant. This feature, along with the ability of Foc to colonise and grow saprophytically in debris, greatly increases pathogen persistence in the contaminated soil (Stover and Waite, 1960; Stover, 1962).
Weed species that are infected or colonised endophytically by the pathogen may act as inoculum reservoirs (Stover, 1962; Pittaway et al., 1999; Hennessy et al., 2005; Ploetz, 2006; Dita et al., 2018). Consequently, eradication of Foc (albeit not specifically of TR4) poses a serious challenge once soil becomes contaminated, and exclusion of the pathogen from banana plantations can be very difficult once it moves into a new area (Ploetz et al., 2015). For these reasons, effective quarantine and exclusion measures are essential in pathogen‐free regions.
The only means of active spread of the pathogen through the soil are from plant to plant by root proximity: roots of diseased banana plants grow into the root zone of the adjacent healthy plants, leading to mat‐to‐mat spread (Figure 1; Dita et al., 2018; Pegg et al., 2019). Since infected rhizomes are often symptomless, they are able to effectively spread the pathogen when used as propagative material (Ploetz, 2006). The pathogen is also able to spread through soil and running water, as well as organic residues on farming tools and machinery. It has been hypothesised that wind‐blown rain could disperse Foc TR4. However, there are no studies confirming this means of spread, although in the Caribbean countries, which are frequently affected by hurricanes, strong wind accompanied by heavy rain causing flooding is considered as an important means of Foc TR4 dissemination (Pérez‐Vicente et al., 2014). Besides rainfall or surface drainage water after rainfall, Foc TR4 may spread rapidly and most efficiently through irrigation water, especially in cases where a contaminated water reservoir is used to irrigate a disease‐free area (Ploetz, 2006). Vertebrate and invertebrate animals inhabiting banana plantations may potentially spread TR4 inoculum. For instance, Meldrum et al. (2013) demonstrated the presence of viable spores of Foc on the exoskeleton of the Banana weevil borer, Cosmopolites sordidus (Germar; Coleoptera: Curculionidae), the most important insect pest of bananas and plantains. Likewise, being a soil‐borne pathogen that can penetrate the host through wounds, any root damage caused by biotic (e.g. nematodes) or abiotic (e.g. agricultural machinery) factors may facilitate the infection of the plant by TR4. Domestic or wild animals may also contribute to farm‐to‐farm dissemination of the pathogen. Since the detection of Foc TR4, drastic campaigns to reduce feral pig populations were implemented in Australia, as these animals are recognised as efficient means of spread of soil‐borne fungal diseases, including Fusarium wilt of banana (Dita et al., 2018).
Figure 1.
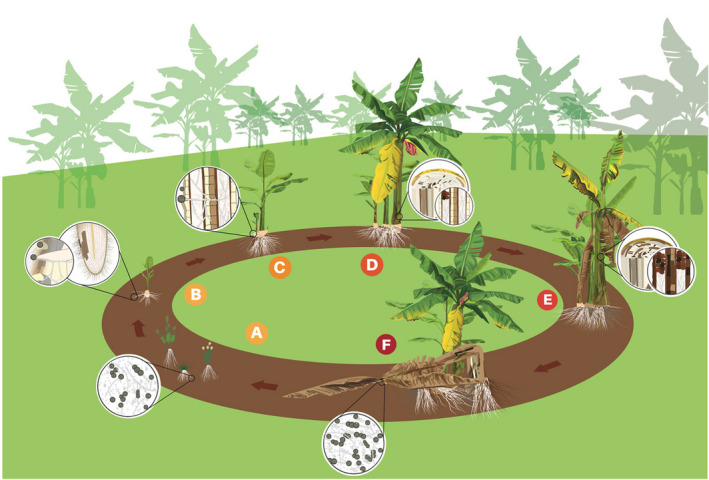
Life cycle of Fusarium oxysporum f. sp. cubense (Foc) in banana. (A) Spores (microconidia, macroconidia and chlamydospores) survive in the soil or on alternative hosts such as weeds. (B) Chlamydospores* germinate stimulated by root exudates and the germ tubes penetrate banana roots. (C) Foc grows through the cortex to the epidermis and mycelium invades the vascular system. (D) Conidia and chlamydospores are constantly produced in the vascular tissues. Conidia are rapidly distributed through the plant via transpiration system. Mycelium and gum block the vascular tissues and first symptoms of yellowing are observed in the older leaves. (E) Foc colonises and destroys more vascular tissues provoking intense wilting. (F) Infected plant dies and the daughter plant, which became infected through its vascular connection with the mother plant, shows initial symptoms. Mother plant eventually falls down and disease cycle starts again (from Dita et al., 2018; https://doi.org/10.3389/fpls.2018.01468).
*The panel is of the opinion that also conidia germinate stimulated by root exudates and the germ tubes penetrate banana roots
Despite Foc TR4 dispersal through banana fruits has been ruled out in the past (Pérez‐Vicente et al., 2014), Bai et al. (2020) have recently demonstrated that the pathogen can invade the peduncle via xylem vascular bundle causing discoloration. Foc TR4 isolates obtained from discoloured peduncles were able to infect ‘Cavendish’ banana plants (Bai et al., 2020).
3.1.3. Host range
According to the CABI datasheet on Foc TR4, the pathogen affects primarily species of Musa (banana, plantain), including M. acuminata. Moreover, Heliconia spp. and other banana relatives are indicated by CABI as ‘other hosts’. However, Heliconia spp. were reported in the past as hosts of Foc race 3, which is no longer recognised as a banana pathogen (Ploetz et al., 2015).
According to the EPPO Global Database, the main hosts of Foc TR4 are Musa sp. and M. acuminata (https://gd.eppo.int/taxon/FUSAC4).
In a review article by Pérez‐Vicente et al. (2014), other species are listed as potential hosts of Foc TR4: Amaranthus spp. (Pittaway et al., 1999), Chloris inflata = Chloris barbata (purpletop chloris; Hennessy et al., 2005), Commelina diffusa (spreading day flower; Wardlaw, 1972), Cyanthillium cinereum (little ironweed; Hennessy et al., 2005), Ensete ventricosum (enset, Abyssinian banana; Wardlaw, 1972), Euphorbia heterophylla (wild poinsettia; Hennessy et al., 2005), Tridax procumbens (coat buttons; Hennessy et al., 2005), Panicum purpurascens (Hennessy et al., 2005) and Paspalum spp. (Pittaway et al., 1999). According to Pérez‐Vicente et al. (2014), the epidemiological importance of plant species that do not belong to the genus Musa has not been documented. However, it has been hypothesised that weeds may act as inoculum reservoirs in the absence of the cultivated host species (Ploetz, 2006).
It should be noted that most of the reports on the host range of Foc reviewed by Pérez‐Vicente et al. (2014) cannot be considered as reliable, since they do not specifically refer to Foc TR4. Moreover, in Hennessy et al. (2005), Foc TR4 was identified (based on VCGs) on various weeds grown in banana plantations, but the Koch’s postulates were not applied and the pathogenicity of the obtained isolates was not tested on banana plants. The lack of application of the Koch’s postulates, along with the reported possibility that false‐positive reactions may occur by screening for the presence of Foc TR4 by PCR (see Section 3.1.5 Detection and identification of the pest) raises a high degree of uncertainty as per the real host range of this pathogen.
Given the high uncertainty on the reliability of studies and reports considering some weeds and non‐Musa plant species as hosts of the pathogen, this Pest categorisation will focus only on cultivated Musa species as major hosts.
3.1.4. Intraspecific diversity
Four pathogenic races have been designated in Foc based on pathogenicity to different reference varieties under field conditions (Stover and Waite, 1960; Stover, 1962; Moore et al., 1993). The following cultivars have been used as differentials: ‘Gros Michel’ for race 1; ‘Bluggoe’ for race 2; and ‘Cavendish’ cultivars for race 4. Since the Australian Heliconia strains do not affect banana, race 3 is now considered a distinct forma specialis of F. oxysporum, namely f. sp. eliconiae (Ploetz, 2006). Race 1 also affects abacá (M. textilis), ‘Maqueño’, ‘Silk’, ‘Pome’ and ‘Pisang Awak’; race 2 affects some bred tetraploids and enset; whereas race 4, race 1 and race 2 may be pathogenic on plantains, and assorted dessert and cooking cultivars (Ploetz, 2006).
Foc Race 4 (also known in the past as ‘subtropical race 4’) had been traditionally considered a subtropical problem, since cold temperatures occurring in the subtropical banana crops in Taiwan, Canary Islands, South Africa and the South of Australia and Brazil predispose the normally resistant Cavendish cultivars.
In the early 1990s, ‘Cavendish’ plantations grown in tropical Southeast Asia were severely hit by the new variant of Foc, named Tropical Race 4 or TR4 (Ploetz, 2006). Foc TR4 is also pathogenic to all banana cultivars susceptible to Foc races 1 and 2. Foc TR4 is a genetically distinct race, compared to the previous populations classified as subtropical race 4 (Pérez‐Vicente et al., 2014).
Various phenetic and genetic characters have been used to study variation in Foc. A significant progress to understand the population biology of Foc derives from the development of methods for determining vegetative compatibility groups (VCGs) that align with clonal lineages (Puhalla, 1985; Correll et al., 1987). Foc comprises 24 VCGs, distributed differentially across the globe (Fourie et al., 2009). At present, TR4 belongs to a restricted group of vegetative compatibility VCG 01213‐01216, whilst other Foc races span over different lineages (at least eight), with closely related VCGs, even when these are distributed over a wide geographic area. These relationships were confirmed by genetic analyses using RFLPs, AFLPs and RAPDs, electrophoretic karyotyping and phylogenies with multiple genes (reviewed in Ploetz, 2006 and Magdama et al., 2019), suggesting that the pathogen reproduces clonally.
The early assumption that Foc race TR4 in the subtropics was an artefact of environmental stress is supported by the presence of isolates suspected to be from Foc race TR4 in the subtropics within the same VCGs (0120 and 0124‐0125; see Section 3.1.5 Detection and identification) as those determined for race 1 isolates from the tropics (Ploetz and Pegg, 2000). Isolates of SR4 are also found in VCGs 0129 and 01211, whereas Foc TR4 affecting Cavendish cultivars in the tropics is characterised by VCG 01213‐01216 (Ploetz et al., 2015). However, DNA‐based studies suggest that Foc races are not genetically related to each other nor share a common ancestor: Foc is composed of several independent lineages, with a population structure that is mainly clonal (Koenig et al., 1997; O’Donnell et al., 1998; Fourie et al., 2009; Magdama et al., 2019).
3.1.5. Detection and identification of the pest
Are detection and identification methods available for the pest?
YES, detection and identification methods are available.
External symptoms of Foc TR4 are already visible in the field, starting with the yellowing of older leaves. However, these symptoms are the same as those caused by other Foc races, and include wilting and yellowing of leaves that begin from the margins of the older leaves, and eventually lead to the collapse of the entire canopy. In addition to the so‐called ‘yellow leaf’ syndrome, also a ‘green leaf’ syndrome may occur in some cultivars, whereby the leaves of affected plants remain predominantly green until the petioles bend and then collapse (Pérez‐Vicente et al., 2014). In general, younger leaves are the last to show symptoms, and remain unusually erect, showing a bristle‐like appearance. Growth does not stop in an infected plant and emerging leaves are of pale colour. The lamina of the emerging leaf can be reduced in size, shrivelled and distorted. The pseudostem eventually splits longitudinally at the plant base. A susceptible banana plant infected by Foc will rarely recover. In case it does, growth would be poor and the mother plant would produce many infected suckers before it dies (Pérez‐Vicente et al., 2014).
Noteworthily, the above‐mentioned symptoms are common to other Foc races; therefore, it is not possible to confirm that the infection is caused by Foc TR4 based on symptoms alone. A characteristic – albeit not exclusive ‐ symptom of Foc TR4 consists of vascular discoloration, which requires the cutting open of the pseudostem or rhizome. It is not useful to observe the fruit, as the pathogen does not cause symptoms on these organs and has never been reported to spread through banana fruit (finger). However, Bai et al. (2020) have recently demonstrated that Foc TR4 is able to colonise the peduncle via xylem vascular bundles. The ability of Foc TR4 to further colonise the xylem of the clusters and of the fingers has not been studied. But considering the vascular nature of the pathogen this cannot be excluded.
The wilt and leaf discoloration symptoms are similar to those caused by the so‐called ‘moko disease’ (incited by the bacterium Ralstonia solanacearum race 2) and by the ‘banana blood disease’ (caused by the bacterium Ralstonia syzygii subsp. celebensis), but these two bacterial pathogens also cause discoloration of the fruit (Plant Health Australia, 2013). In addition, the ‘yellow leaf’ syndrome, which is characterised by the yellowing at the margins of older leaves, can be confused with potassium deficiency, especially in drought and cold environments (Pérez‐Vicente et al., 2014).
In culture, isolates of F. oxysporum causing disease on different hosts cannot be distinguished morphologically. Fungal colonies grow 4–7 mm day–1 on potato dextrose agar (PDA) at 24°C, forming an abundant aerial, white to purple mycelium. Colonies on PDA have a variable morphology: Mycelia can be hairy to cottony, spaced or abundant and variable from white, salmon, to pale violet. Black to violet sclerotia can be produced by some Foc TR4 isolates (Pérez‐Vicente et al., 2014). Whilst F. oxysporum usually produces pale violet to dark red colour pigments in PDA, some Foc TR4 isolates mutate rapidly from pionnotal (i.e. with colonies lacking aerial mycelium, with abundant greasy or brilliant conidia aggregates) to a flat mycelium of white‐pale yellowish to peach colour. In a modified Komada medium, some isolates of Foc TR4 may develop laciniated radial colonies, which are not found in Foc isolates of races 1 and 2 (Pérez‐Vicente et al., 2014); however, these characteristics are not a determinant of an Foc TR4 diagnosis.
Sporodochia of Foc TR4 are tan to orange, and sclerotia are blue and submerged. Micro‐ and macroconidia are produced on branched and unbranched monophialides. Macroconidia (27–55 × 3.3–5.5 μm) are abundant, falcate to erect to almost straight, of thin walls, with 3–5 septa (usually 3 septa). Apical cell is attenuated or hook shaped in some isolates. Basal cells are foot shaped. Macroconidia are developed in single phialides. Microconidia (5–16 × 2.4–3.5 μm), usually without septa, can be oval, elliptic to kidney shaped and develop abundantly in false heads in short monophialides. Chlamydospores (7–11 μm diameter) are abundantly formed in hyphae or in conidia, single or in chains, usually in pairs, but their development can be slower in certain isolates (Pérez‐Vicente et al., 2014) (Figure 2).
Figure 2.
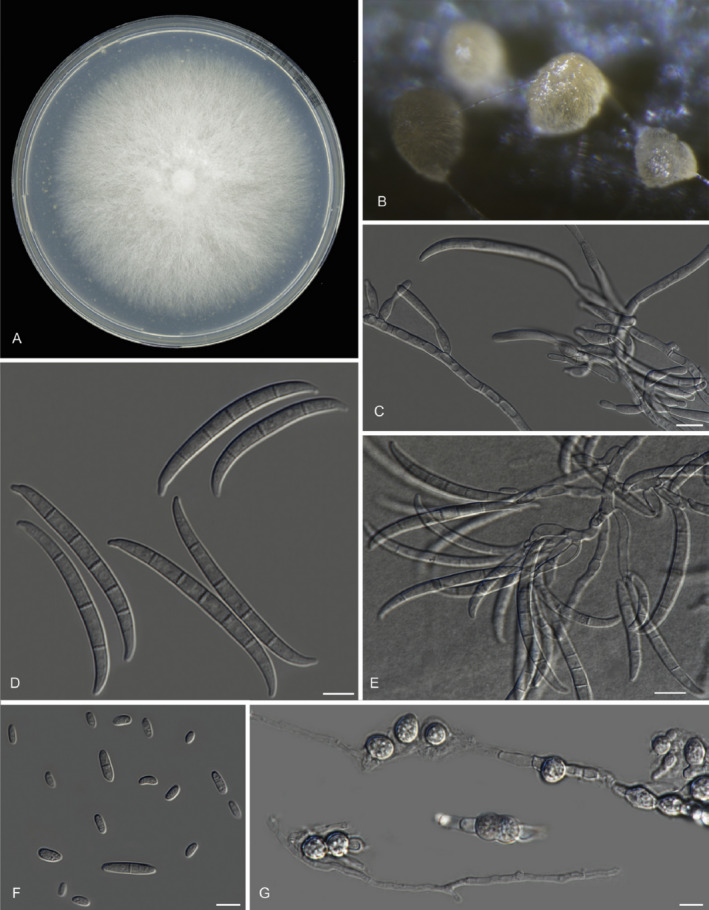
A. Culture of Fusarium oxysporum f. sp. cubense TR4 (referred to as Fusarium odoratissimum) grown on PDA. B. Sporodochia on carnation leaves. C. Monophialides with initial conidia being formed. D. Falcate‐shaped macroconidia. E. Branched conidiophores. F. Elliptical microconidia. G. Thick‐walled chlamydospores. Scale bars C–G = 10 µm (from Maryani et al., 2019, https://doi.org/10.1016/j.simyco.2018.06.003)
As previously mentioned, Foc TR4 has no known teleomorph (Ploetz, 2006).
The only means to differentiate Foc TR4 from other Foc races grounds on the use of PCR‐based diagnostic tests and on the analysis of the isolate to confirm it belongs to VCG 01213‐01216. The determination of VCGs requires substantial amount of work and time to generate nitrate‐deficient mutants and their crossing with nit‐M VCG tester strains representative of the known VCGs of Foc (Li et al., 2013).
According to Lin et al. (2009), race 4 of Foc, i.e. both STR4 and TR4 races, could be distinguished from races 1 and 2, and separated from other formae speciales of F. oxysporum. They first reported the primer set Foc‐1/Foc‐2, derived from the sequence of a random primer OPA‐02‐amplified fragment, that allowed the production of a 242‐bp DNA fragment allegedly specific to Foc race 4. The authors could detect small quantities of TR4 DNA as low as 10 pg in 50–2,000 ng host genomic DNA with high efficiency. However, according to Magdama et al. (2019), Lin et al. (2009) primers (as well as the other assays derived from the same sequence characterised amplified region, SCAR) also cross amplify DNA from a few F. oxysporum strains that are endophytes on ‘Cavendish’ banana plants.
Dita et al. (2010) reported another PCR diagnostic that uniquely amplifies a 463‐bp amplicon targeting an IGS sequence in isolates belonging to Foc VCG 01213, which encompasses Foc TR4. This diagnostic test was specific for Foc TR4 on pure‐culture DNAs of VCG 01213 isolates, including isolates from infected banana tissues of different origin. According to Dita et al. (2010), the Foc‐1⁄Foc‐2 primer set published by Lin et al. (2009), reacted with isolates of 10 different VCGs, including those belonging to the 01213 group (i.e. Foc TR4). According to Aguayo et al. (2017) and Magdama et al. (2019), Dita’s primers also cross amplify DNA from isolates belonging to VCGs 01215 from Foc race 4, 0126 and 0210 from race 1 of Foc, from F. oxysporum f. sp. canariensis and from a few F. oxysporum strains that are endophytes on ‘Cavendish’ banana plants, respectively.
Lin et al. (2013) have further described a real‐time PCR method based on SCAR primers FocSc‐1/FocSc‐2 for the specific detection of Foc race 4 (and therefore not specific to Foc TR4) in field‐infected banana. According to the authors, this method shows high reproducibility and allows to detect the pathogen in genomic DNA of symptomatic banana pseudostems and leaves. In the same paper, however, authors were able to isolate Foc colonies only from symptomatic banana tissues, but not from symptomless banana tissues, even though positive real‐time PCR results were obtained (Lin et al., 2013). This result raises some concern on the reliability of the entire diagnostic test.
There are several other detection assays available in the literature. Their specificity has been further addressed by the studies of Madgama et al. (2019) and Aguayo et al. (2017), confirming that the assays of Li et al. (2013) and Aguayo et al. (2017) are to date the most specific to Foc TR4 isolates. Both assays target a gene encoding a protein putatively involved in pathogenicity. According to Magdama et al. (2019), it is important to note that none of the available diagnostic methods when used alone is completely reliable and that a combination of phylogenetic analysis, VCG testing, sequence analysis and pathogenicity tests should be adopted for the unambiguous identification of the pathogen.
No EPPO Standard is available for the detection and identification of Foc TR4.
3.2. Pest distribution
3.2.1. Pest distribution outside the EU
F. oxysporum f. sp. cubense TR4 is reported from Asia, Oceania, Africa and America.
In Asia, Foc TR4 was first identified in Taiwan in 1967, but it was probably causing Fusarium banana wilt in Taiwan since the 1960s (Su et al., 1977; Molina et al., 2009; Li et al., 2011). In the 1990s, Foc TR4 was identified in Malaysia (Molina et al., 2009; Wong et al., 2019), Indonesia (Hermanto et al., 2009; Maryani et al., 2019), Borneo (Davis et al., 2000), Sumatra, Java, Kalimantan, Sulawesi and Papua (O’Neill et al. 2009; EPPO, 2021), where the strains isolated in Taiwan are thought to have originated from (Buddenhagen, 2009; Maryani et al., 2019). Foc TR4 has then spread to the entire Greater Mekong Subregion in Laos (Chittarath et al., 2018), Myanmar, Thailand (Zheng et al., 2018) and Vietnam (Hung et al., 2018; Zheng et al., 2018). The Foc TR4 is also reported from China (Qi et al., 2008; Zhang et al., 2009; Li et al., 2011; Huang et al., 2013; Wu et al., 2019), Philippines (Molina et al., 2008; Solpot et al., 2016), Pakistan (Ordoñez et al., 2016) and India (Damodaran et al., 2019; Thangavelu et al., 2019).
In the Middle East, Foc TR4 has been reported from Jordan (García‐Bastidas et al., 2014), Lebanon (Ordoñez et al., 2016), Israel (Maymon et al., 2018, 2020; EPPO, 2019), Oman (Dita et al., 2018) and Turkey (Özarslandan and Akgül, 2020). A banana cultivation area of 200 ha in Jericho, within the Palestinian Authority, needs to be re‐evaluated for the pest status as this production area is in close proximity to the borders with the Foc TR4‐infested Shooneh Janoobiyeh district in Jordan (Maymon et al., 2020).
In Oceania, an outbreak of Fusarium wilt occurred in 1997 on ‘Cavendish’ in the Northern Territory of Australia. This was the first detection of Foc TR4 in Australia (Conde and Pitkethley, 2001, 2001; Daly and Walduck, 2006). In March 2015, Foc TR4 was detected in Australia’s major banana production region near Tully in North Queensland (O’Neill et al., 2016).
In Africa, the presence of Foc TR4 has been reported in Mozambique (IPPC, 2013; Dita et al., 2018; Viljoen et al., 2020), and in the French Department of Mayotte, located in the Indian Ocean (Aguayo et al., 2021).
Foc TR4 is confirmed to have reached Latin America with a first report in Colombia (García‐Bastidas et al., 2020; Martínez‐Solórzano et al., 2020) and a second one in Peru (SENASA, 2021).
Details of the current distribution of Foc TR4 outside the EU are presented in Appendix A. An updated map on the global distribution of the pathogen is available in the EPPO Global Database and is presented in Figure 3.
Figure 3.
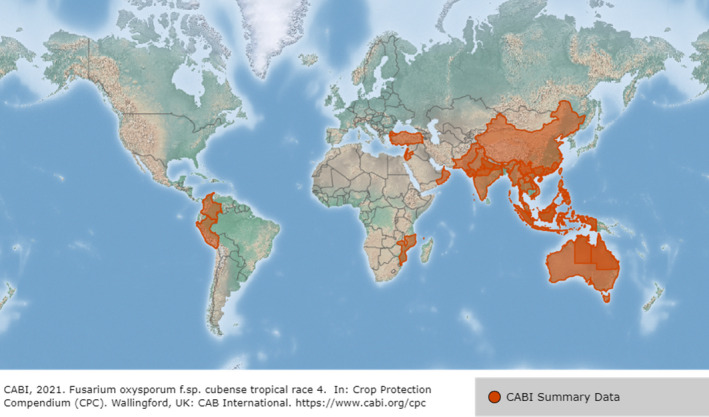
Global distribution of Fusarium oxysporum f. sp. cubense Tropical Race 4 (Source: CABI; Available online: https://www.cabi.org/isc/datasheet/59074053 [Accessed: 8 November 2021])
As the currently available PCR detection methods may generate false positives (see Section 3.1.5 Detection and identification of the pest), there is uncertainty on the geographic distribution of the pathogen unless confirmation of the VCG was performed.
3.2.2. Pest distribution in the EU
Is the pest present in the EU territory? If present, is the pest widely distributed within the EU?
No, Foc TR4 is not reported to be present in the EU.
The first and only report of Foc TR4 from an (administrative) EU region is from the French overseas Department of Mayotte, located in the Indian Ocean (Aguayo et al., 2021) which is not considered as EU territory in Regulation (EU) 2016/2031. Under such circumstances, the pathogen should be considered as absent from the EU territory.
3.3. Regulatory status
3.3.1. Commission Implementing Regulation 2019/2072
Fusarium oxysporum f. sp. cubense Tropical Race 4 is not listed in Annex II of Commission Implementing Regulation (EU) 2019/2072, an implementing act of Regulation (EU) 2016/2031.
3.3.2. Hosts of Fusarium oxysporum f. sp. cubense Tropical Race 4 that are prohibited from entering the Union from third countries
Musa spp., the hosts of F. oxysporum f. sp. cubense TR4, are not prohibited from entering the Union from third countries. However, special requirements exist for Musa spp. plants for planting imported from third countries where some species of Ralstonia are present [Annex VII of Implementing Regulation (EU) 2019/2072] (Table 2).
Table 2.
List of plants, plant products and other objects that are Fusarium oxysporum f. sp. cubense TR4 hosts whose introduction into the Union from certain third countries is prohibited (Source: Commission Implementing Regulation (EU) 2019/2072, Annex VI)
| List of plants, plant products and other objects whose introduction into the Union from certain third countries is prohibited | |||
|---|---|---|---|
| Description | CN Code | Third country, group of third countries or specific area of third country | |
| 19. |
Soil as such consisting in part of solid Organic substances |
ex 2530 90 00 ex 3824 99 93 |
Third countries other than Switzerland |
| 20. |
Growing medium as such, other than Soil, consisting in whole or in part of solid organic substances, other than that composed entirely of peat or fibre of Cocos nucifera L., previously not used for growing of plants or for any agricultural purposes |
ex 2530 10 00 ex 2530 90 00 ex 2703 00 00 ex 3101 00 00 ex 3824 99 93 |
Third countries other than Switzerland |
3.4. Entry, establishment and spread in the EU
3.4.1. Entry
Is the pest able to enter into the EU territory? If yes, identify and list the pathways
Yes, the pathogen could potentially enter the EU territory via the host plants for planting, other than vitroplants, the fresh fruits and leaves of host plants, and the soil and other substrates pathways.
Comment on plants for planting as a pathway
Host plants for planting, other than vitroplants, is the main pathway for the entry of the pathogen into the EU territory.
The PLH Panel identified the following potential pathways for the entry of the pathogen into the EU territory (see also Table 3 for a summary of potential pathways of entry):
host plants for planting, other than vitroplants
fresh fruits of host plants, soil and other substrates originating in infested third countries (Table 3).
Table 3.
Potential pathways for Fusarium oxysporum f. sp. cubense Tropical Race 4 into the EU 27
| Pathways | Life stage | Relevant mitigations [e.g. prohibitions (Annex VI) or special requirements (Annex VII) within Implementing Regulation 2019/2072] |
|---|---|---|
| Description (e.g. host/intended use/source) | ||
| Plants for planting of Musa L. | Microconidia, macroconidia, chlamydospores, mycelium |
Annex VII (22.) requires official statement that: a) the plants originate in areas which have been found free from Ralstonia solanacearum (Smith) Yabuuchi et al. emend. Safni et al., Ralstonia pseudosolanacearum Safni et al., Ralstonia syzygii subsp. celebensis Safni et al. and Ralstonia syzygii subsp. indonesiensis Safni et al. or b) no symptoms of Ralstonia solanacearum (Smith) Yabuuchi et al. emend. Safni et al., Ralstonia pseudosol anacearum Safni et al., Ralstonia syzygii subsp. celebensis Safni et al. and Ralstonia syzygii subsp. indonesiensis Safni et al. have been observed on the plants at the place of production since the beginning of the last complete cycle of vegetation. |
| Fresh fruits of Musa spp. (bananas, plantains) traded in bunches with peduncle | Microconidia, macroconidia, mycelium | |
| Fresh banana/plantain leaves used for cooking, food wrapping and food serving | Microconidia, macroconidia, mycelium | |
| Soil and other substrates associated or not with host plants for planting | Chlamydospores, sclerotia | Annex VI (19. & 20.) bans the introduction of soil and growing media as such into the Union from third countries other than Switzerland |
The major pathway for the entry of Foc TR4 into the EU territory is plant propagation material of Musa spp., other than in vitro‐grown plants, originated in infested third countries. In Eurostat, there is no import data specific for Musa spp. plants for planting. Detailed data on the EU 27 imports of live indoor and outdoor plants from third countries where the pathogen is present can be found in Appendix C.
Moreover, the recent demonstration that the pathogen can invade the fruit peduncle via xylem vascular bundle (Bai et al., 2020) indicates that Foc TR4 could potentially enter the EU through the fresh banana bunches imported from infested third countries, which may carry part of the potentially infected peduncle. However, uncertainty exists on the ability of Foc TR4 to invade xylem vessels in the terminal clusters and in the banana fingers (fruits).
The increasing use of banana leaves in eco‐friendly packaging and the traditional use of banana/plantain leaves for cooking, food wrapping and food serving in a wide range of countries may represent a potential pathway of entry of the pathogen into the EU territory.
Foc TR4 is a common soil inhabitant, able to survive saprophytically for long periods in soil (see Section 3.1.2). Therefore, soil and other substrates associated or not with host plants for planting imported into the EU from infested third countries represent a potential pathway of entry of the pathogen into the EU territory.
The pathogen is unlikely to enter the EU by natural means (wind, rain, insect vectors, surface water, etc.) because of the long distance between the infested third countries and the EU Member States.
Table 4 shows the imported quantities of Musa spp. fresh (or dried) fruit (bananas and plantains) between 2016 and 2020 from third countries where Foc TR4 is known to occur. No import data specific to Musa spp. plants for planting is available in Eurostat (only aggregated data on imports of live indoor and outdoor plants). Detailed data on the EU 27 imports of fresh (or dried) banana and plantain fruits and live indoor and outdoor plants from countries where the pathogen is present can be found in Appendix C.
Table 4.
EU 27 annual imports of fresh or dried produce of Musa spp. (bananas and plantains) from countries where Fusarium oxysporum f. sp. cubense TR4 is present, 2016–2020 (in 100 kg) Source: EUROSTAT accessed on 23/10/2021
| Commodity | HS code | 2016 | 2017 | 2018 | 2019 | 2020 |
|---|---|---|---|---|---|---|
| Bananas and plantain, fresh or dried | 0803 | 11,262,222 | 12,762,575 | 12,552,145 | 12,613,494 | 13,225,070 |
| Live indoor plants* | 06029099 | 167,407 | 302,528 | 369,253 | 425,115 | 460,379 |
| Live outdoor plants* | 06029050 | 9,696 | 9,105 | 8,506 | 10,446 | 12,512 |
| Sum | 11,439,325 | 13,074,208 | 12,929,904 | 13,049,055 | 13,697,961 |
The import of live indoor and outdoor plants from the countries where the pest occurs is aggregated data which can potentially also include Musa species. No specific data for the import of Musa plants is available in EUROSTAT.
Notifications of interceptions of harmful organisms began to be compiled in Europhyt in May 1994 and in TRACES in May 2020. As at 22 October 2021, there were no interceptions of Fusarium oxysporum f. sp. cubense TR4 in the Europhyt and TRACES databases.
3.4.2. Establishment
Is the pest able to become established in the EU territory?
Yes. The pest has not been reported to be present in the EU territory. However, both the biotic (host availability) and abiotic (climate suitability) factors suggest that Fusarium oxysporum f. sp. cubense Tropical Race 4 could potentially establish in the risk assessment area, particularly in those areas where banana crops are grown.
Given its ability to survive saprophytically, Foc TR4 could potentially be transferred from the host plants for planting pathway to the Musa spp. plants grown in the EU territory via the following routes: (i) suckers collected from infected plants and used as propagation material, (ii) roots of diseased banana plants growing into the root zone of the adjacent healthy plants, (iii) surface (rain or irrigation) water, (iv) wind‐disseminated and splash‐dispersed spores, (v) contaminated tools, footwear, vehicles/agricultural machinery and (vi) invertebrate and vertebrate animals.
The relevance of the transfer via fresh fruits traded in clusters with peduncle or via fresh banana leaves will depend on the volume and frequency of imported commodities, their final destination (e.g. retailers, packinghouses), the proximity to the banana hosts and the management of fruit/leaf waste.
3.4.2.1. EU distribution of main host plants
As noted above and shown in Appendix B, Foc TR4 has a very high level of host specificity with its main host being Musa spp. (banana and plantain). Banana crops are confined to the warmer southern Europe: Greece, Spain (Canary Islands), Italy, Cyprus and Portugal (Madeira) (source Promusa, data based on FAOSTAT; https://www.promusa.org/Banana‐producing+countries+portal#Production_by_country; Accessed: 3 November 2021).
The harvested area of the main host of Foc TR4 cultivated in the EU 27 in recent years is shown in Table 5. Eurostat also provides data of harvested area in France and Spain. However, these data have not been included in Table 5, as banana is grown only in the French Overseas Departments and Canary Islands, which are not considered EU territory, according to Regulation (EU) 2016/2031.
Table 5.
Harvested area of Fusarium oxysporum f. sp. cubense TR4 main host (banana) in EU 27, 2016–2020 (1,000 ha). Source EUROSTAT (Accessed: 25 October 2021) https://ec.europa.eu/eurostat/databrowser/view/apro_cpsh1/default/table?lang=en
| Country | 2016 | 2017 | 2018 | 2019 | 2020 |
|---|---|---|---|---|---|
| Greece | 0.08 | 0.09 | 0.09 | 0.10 | 0.10 |
| Cyprus | 0.29 | 0.21 | 0.22 | 0.21 | 0.21 |
| Portugal | 1.04 | 1.04 | 1.05 | 1.12 | 1.12 |
| EU 27 | 1.41 | 1.34 | 1.36 | 1.43 | 1.43 |
3.4.2.2. Climatic conditions affecting establishment
Foc TR4 has been reported from Asia, Oceania, Africa and South America. In some cases, the climatic zones where the pathogen is present are comparable to climatic zones within the EU (Figure 4).
Figure 4.

Fusarium oxysporum f.sp. cubense TR4 climate suitability analysis based on the Köppen–Geiger climate classification. Regions with black borders indicate countries/regions where the pest is present. Red dots indicate point locations where the pest was found. Climates not present in EU27 are not mapped. The legend shows the list of Köppen–Geiger climates. Only climates relevant for the pest are coloured
The global Köppen–Geiger climate zones (Kottek et al., 2006) describe terrestrial climate in terms of average minimum winter temperatures and summer maxima, amount of precipitation and seasonality (rainfall pattern). Foc TR4 occurs in several climate zones, such as BSh, BSk, Cfa, Cfb, Cfc, Csa, Csb. These climate zones also occur in the EU territory, where the main host (Musa spp.) of the pathogen is grown.
Based on the above, it may be assumed that the climatic conditions occurring in some parts of the EU territory (mainly in southern EU Member States) are favourable for the establishment of Foc TR4. The possibility of establishment of the pathogen in glasshouse‐grown banana crops at higher latitudes in the EU, as well as the possibility that warming of high latitude regions due to climate change may lead to northward expansion of tropical crops (King et al., 2018), should not be ruled out.
3.4.3. Spread
Describe how the pest would be able to spread within the EU territory following establishment?
Following establishment, TR4 could spread within the EU territory by natural and human‐assisted means. Trading of host plants for planting is the main means of long‐distance spread of the pathogen.
Following establishment, long distance dispersal could occur through anthropogenic‐related factors (e.g. asymptomatic, albeit infected, suckers used as planting material; fresh fruits commercialised in bunches with the peduncle and collected from infected plants, farm equipment, clothes, footwear, tools, containers, etc., which have been used in Foc TR4‐infested areas).
Dispersal at short distances may be associated with both anthropogenic and natural factors, such as mat to mat spread, water run‐off, animal movement or contaminated soil (see Section 3.1.2 Biology of the pest).
3.5. Impacts
Would the pests’ introduction have an economic or environmental impact on the EU territory? (Yes or no)
Yes, the introduction of Foc TR4 will have severe economic impact on some parts of the EU territory where the host is grown.
In the areas of its current distribution, Foc TR4 affects a wide range of banana cultivars, including the economically important Cavendish (AAA) banana cultivars, which are commonly grown in large monocultures. The spread of Foc TR4 to these plantations can destroy the entire crop, thereby causing significant economic impact to commercial banana growers and exporters (Mostert et al., 2017).
The annual cost of Foc TR4 has been calculated for some countries that have experienced epidemics of Fusarium wilt caused by this pathogenic fungus. In Malaysia, for instance, the annual loss was calculated as 14.1 million USD (Malik et al., 2013). In Indonesia, the assessed value was 121 million USD (Hermanto et al., 2009), whereas annual losses on ‘Cavendish’ bananas in Taiwan were estimated at 253.3 million USD (Peng et al., 2013).
As a paradigmatic example, in the 1960s, Taiwan exported 60,000 12‐kg boxes of ‘Giant Cavendish’. The first appearance of the Foc TR4 pathogen has been reported on Cavendish cultivars in 1967: The disease has spread so rapidly, that the number of affected plants increased to 5,536 within 3 years. In 1976, 1,200 ha was infested, representing approximately 500,000 banana plants (Hwang and Ko, 2004).
A dynamic model has been developed to estimate the potential economic impact of Foc TR4 on the Australia banana industry if it continues to spread via natural and anthropogenic means, and annual losses of > 138 million AUD were predicted (Cook et al., 2015).
The economic impact on the European banana production would be devastating should the pathogen be introduced and spread into the EU territory.
3.6. Available measures and/or potential specific import requirements and limits of mitigation measures
Are there measures available to prevent the entry into the EU such that the risk becomes mitigated?
Yes. Although not specifically targeted against Foc TR4, existing phytosanitary measures (see Sections 3.3.2 and 3.4.1) mitigate the likelihood of the pathogen’s entry into the EU territory. Potential additional measures also exist to further mitigate the risk of entry (see Section 3.6.1).
Phytosanitary measures are currently applied to the soil pathway (see Section 3.3 for prohibitions and Section 3.4.1 for specific requirements). However, current regulations applied to the Musa spp. plants for planting pathway (see Section 3.4.1), do not specifically consider Foc TR4. As plants for planting and fresh fruit and leaves of Musa spp. (bananas and plantains) are not currently prohibited for import into the EU territory, additional measures are listed in Table 6.
Table 6.
Selected control measures (a full list is available in EFSA PLH Panel, 2018) for pest entry/establishment/spread/impact in relation to currently unregulated hosts and pathways. Control measures are measures that have a direct effect on pest abundance
| Control measure/Risk reduction option (Blue underline = Zenodo doc, Blue = WIP) | RRO summary | Risk element targeted (entry/establishment/spread/impact) |
|---|---|---|
| Require pest freedom | Plants (other than vitroplants) or fresh leaves and fruits must come from a country officially free from the pest, or from a pest‐free area or from a pest‐free place of production. | Entry/Spread |
| Growing plants in isolation | Growing transplants in isolation from Musa spp. crops may represent an effective control measure. | Entry/Establishment/Spread/Impact |
| Managed growing conditions |
The use of pathogen‐free propagative material (vitroplants), proper field drainage, avoidance of contaminated water for irrigation, plant distancing and removal of any infected plants and plant residues in the field may all represent effective strategies to reduce the spread of Foc TR4. Additional mitigating recommendations include: using NO3‐based fertilisers and avoiding the use of urea and NH4 as fertiliser; controlling root and rhizome pests, which are potential vectors of the pathogen. |
Entry/Establishment/Spread/Impact |
| Crop rotation, associations and density, weed/volunteer control |
Foc TR4 is well adapted to long‐term survival in soil and it is virtually impossible to eliminate chlamydospores from infested soil by crop rotation or by bare fallowing. Although uncertainty exists on the role of specific weeds (e.g. Amaranthus spp., Chloris inflata = Chloris barbata, Commelina diffusa, Cyanthillium cinereum, Euphorbia heterophylla, Tridax procumbens, Panicum purpurascens and Paspalum spp.) as alternative host plants in Foc TR4 survival, control of these weeds might reduce inoculum sources. |
Establishment/Impact |
| Use of resistant and tolerant plant species/varieties | Tolerant Cavendish clones from Taiwan, China, Australia and Indonesia are available. However, resistance is not complete and growing such plants may represent an epidemiological risk, as they may mask the presence and spread of Foc TR4 | Establishment/Impact |
| Roguing and pruning | Effectiveness of roguing against the pathogen is uncertain and pruning is not practiced in banana crops. | Impact |
| Cleaning and disinfection of facilities, tools and machinery | Cleaning, disinfection and disinfestation (sanitation) of equipment and facilities (including premises, storage areas) are good cultural and handling practices employed in the production and marketing of any commodity and may contribute to mitigate likelihood of entry or spread of TR4. | Entry/Spread |
| Limits on soil |
Plants, plant products other pathway agents (e.g. used farm machinery) to be free from soil or growing medium; Growing medium is pest free e.g. the growing medium is free from soil and organic matter and had not been previously used for growing plants or for any other agricultural purposes, or was composed entirely of peat or fibre, or was subjected to effective fumigation to ensure freedom from pests; |
Entry/Spread |
| Soil treatment |
Soil fumigation can be effective in reducing the disease incidence upon treatment. However, fumigated areas are easily re‐invaded by the pathogen. Soil flooding, where applicable, has been shown to reduce the incidence of Panama disease, but in the case of Foc TR4, this practice is not advisable as it may help spreading the pathogen over long distances. |
Spread/Impact |
| Use of non‐contaminated water | Movement and survival of Foc TR4 in water can be significant and outbreaks of the disease commonly occur in association with irrigation and/or flooding. Therefore, chemical and physical treatment of water to eliminate pathogen’s propagules is advisable, albeit limited to banana grown under glasshouse. | Spread |
| Waste management | Waste management (incineration, production of bio‐energy) takes place in authorised facilities and official restriction on the movement of infected material is in force to prevent the pest from escaping. | Spread |
| Conditions of transport | When potentially infected/contaminated material must be transported (including proper disposal of infested waste material), specific transport conditions (kind of packaging/protection, time of transport, transport mean) should be defined to prevent the pest from escaping. | Entry/Spread |
| Controlled atmosphere | Controlled atmosphere is unlikely to represent an effective mean of control against Foc TR4. | |
| Post‐entry quarantine and other restrictions of movement in the importing country | Recommended for Musa spp. plants for planting other than vitroplants. Imports of Musa plants for planting should use the route of intermediate quarantine stations. Those materials should be adequately indexed and identified as free of Foc TR4. | Entry/Establishment/Spread |
3.6.1. Identification of potential additional measures
Additional potential risk reduction options and supporting measures are shown in Sections 3.6.1.1 (Table 7) and 3.6.1.2 (Table 8).
Table 7.
Selected supporting measures (a full list is available in EFSA PLH Panel, 2018) in relation to currently unregulated hosts and pathways. Supporting measures are organisational measures or procedures supporting the choice of appropriate risk reduction options that do not directly affect pest abundance
| Supporting measure | Summary | If pest not known to occur EU: Risk element targeted (entry/establishment/spread/impact) |
|---|---|---|
| Inspection and trapping |
Symptoms of Foc TR4‐induced banana wilt are similar to those caused by other races of Foc or other diseases and disorders. Signs of the pathogen (micro‐ and macroconidia, etc.) are difficult to be detected visually, and moreover, they are similar to those of other races of Foc. Therefore, the pathogen cannot be detected based on visual inspection. However, vascular discoloration on plants for planting and on the petioles should prompt immediate laboratory testing. |
Entry/Establishment/Spread |
| Laboratory testing | Laboratory testing of host plants and products for Foc TR4. | Entry/Establishment/Spread |
| Sampling | Necessary as part of other RROs. | Entry/Establishment/Spread |
| Phytosanitary certificate and plant passport | Recommended for Musa spp. plants, including fruits | Entry/Spread |
| Certified and approved premises | If plant material originates from an approved premise, e.g. from a pest‐free area, the likelihood of commodity being infected is assumed to be reduced. | Entry/Spread |
| Certification of reproductive material (voluntary/official) | Plants come from within an approved propagation scheme and are certified pest free (level of infestation) following testing; Used to mitigate against pests that are included in a certification scheme | Entry/Spread |
| Delimitation of Buffer zones | Foc TR4 is not present in the EU. | Spread |
| Surveillance | Foc TR4 is not present in the EU. | Spread |
Table 8.
The Panel’s conclusions on the pest categorisation criteria defined in Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column)
| Criterion of pest categorisation | Panel’s conclusions against criterion in Regulation (EU) 2016/2031 regarding Union quarantine pest | Key uncertainties |
|---|---|---|
| Identity of the pest ( Section 3.1 ) | The identity of Fusarium oxysporum f. sp. cubense (Foc) Tropical Race 4 (TR4) is well established and the pathogen has been shown to produce consistent symptoms and to be transmissible. | None |
| Absence/presence of the pest in the EU ( Section 3.2 ) | The pathogen has not been reported to be present in the EU territory. | None. |
| Regulatory status ( Section 3.3 ) | Foc TR4 has not been reported to be present in the EU and it is currently not regulated. | None. |
| Pest potential for entry, establishment and spread in the EU ( Section 3.4 ) |
The pathogen is able to enter into, become established in, and spread within the EU territory through the following pathways:
The biotic and abiotic factors occurring in some parts of the EU territory suggest that Foc TR4 could potentially establish in the risk assessment area. Following establishment, Foc TR4 could potentially spread within the EU territory by natural and human‐assisted means. Trading/movement of host plants for planting is the main means of long‐distance spread of the pathogen. |
Uncertainty on the real host range of the pathogen, since it has not been demonstrated whether FocTR4 isolates can infect plant species other than Musa spp. that were previously reported as hosts of other Foc races. |
| Potential for consequences in the EU ( Section 3.5 ) | The pathogen’s introduction would have a severe economic impact on the main banana production areas in the EU territory. | None. |
| Available measures ( Section 3.6 ) | Although not specifically targeted against Foc TR4, existing phytosanitary measures mitigate the likelihood of the pathogen’s entry into the EU territory. Potential additional measures also exist to further mitigate the risk of entry into, establishment and spread of the pathogen within the EU. | None. |
| Conclusion ( Section 4 ) | All criteria assessed by EFSA for consideration of Foc TR4 as a potential Union quarantine pest are met. | None. |
| Aspects of assessment to focus on/scenarios to address in future if appropriate: | Specific studies are needed to ascertain the host range of Foc TR4, whose presence in species other than Musa spp. should be validated by VCG analysis and fulfilment of Koch’s postulates. Available PCR protocols may generate false‐positive reactions with non‐pathogenic endophytic strains of F. oxysporum. Therefore, until a quick one step PCR test will be developed, a combination of phylogenetic analysis, VCG testing, sequence analysis and pathogenicity tests is needed to identify the pathogen unambiguously. | |
3.6.1.1. Additional supporting measures
Potential additional supporting measures are listed in Table 7.
3.6.1.2. Biological or technical factors limiting the effectiveness of measures to prevent the entry (and spread when applicable) of the pest
The effectiveness of measures to prevent the entry of the pest into the EU might be limited by the following biological or technical factors:
Latently infected plants and plant products are unlikely to be detected by visual inspection.
The similarity of symptoms and signs caused by F. oxysporum f. sp. cubense TR4 with those of other races of the same forma specialis or other Fusarium spp. makes it impossible to detect the pathogen based solely on symptomatology and morphology.
The available PCR detection protocols may generate false‐positive reactions in the presence of non‐pathogenic endophytic strains of F. oxysporum.
The ability to survive in soil may favour the unintentional introduction of the pathogen by tourists traveling from infested areas (e.g. through contaminated soil particles adhering to footwear).
3.7. Uncertainty
A major uncertainty concerns the host range of Foc TR4, since it has not been demonstrated whether isolates of this race can infect plant species other than Musa spp. that were previously reported as hosts of other Foc races (e.g. Ensete ventricosum and Heliconia spp.).
Further uncertainty resides on the real potential of pathogen introduction through banana fresh fruits (fingers) commercialised in clusters, as Foc TR4 has been recently demonstrated to be able to infect and survive in the peduncle vessels.
As the currently available PCR detection methods may generate false positives, there is uncertainty on the geographic distribution of the pathogen unless confirmation of the VCG was performed.
Nonetheless, these uncertainties do not affect the conclusions of the pest categorisation. Considering the devastating effects of Foc TR4 reported from currently infested areas, there are no uncertainties on the fact that this pathogen represents ‘the most imminent threat’ to banana production (GLOBAL G.A.P.).
4. Conclusions
Fusarium oxysporum f. sp. cubense (Foc) TR4 has not been reported to be present in the EU territory as defined in Regulation (EU) 2016/2031. Foc TR4 satisfies the criteria that are within the remit of EFSA to assess for this pathogen to be regarded as a potential Union quarantine pest.
Abbreviations
- EPPO
European and Mediterranean Plant Protection Organization
- FAO
Food and Agriculture Organization
- IPPC
International Plant Protection Convention
- ISPM
International Standards for Phytosanitary Measures
- MS
Member State
- PLH
EFSA Panel on Plant Health
- PZ
Protected Zone
- TFEU
Treaty on the Functioning of the European Union
- ToR
Terms of Reference
Glossary
- Containment (of a pest)
Application of phytosanitary measures in and around an infested area to prevent spread of a pest (FAO, 2018)
- Control (of a pest)
Suppression, containment or eradication of a pest population (FAO, 2018)
- Entry (of a pest)
Movement of a pest into an area where it is not yet present, or present but not widely distributed and being officially controlled (FAO, 2018)
- Eradication (of a pest)
Application of phytosanitary measures to eliminate a pest from an area (FAO, 2018)
- Establishment (of a pest)
Perpetuation, for the foreseeable future, of a pest within an area after entry (FAO, 2018)
- Greenhouse
A walk‐in, static, closed place of crop production with a usually translucent outer shell, which allows controlled exchange of material and energy with the surroundings and prevents release of plant protection products (PPPs) into the environment.
- Impact (of a pest)
The impact of the pest on the crop output and quality and on the environment in the occupied spatial units
- Introduction (of a pest)
The entry of a pest resulting in its establishment (FAO, 2018)
- Pathway
Any means that allows the entry or spread of a pest (FAO, 2018)
- Phytosanitary measures
Any legislation, regulation or official procedure having the purpose to prevent the introduction or spread of quarantine pests, or to limit the economic impact of regulated non‐quarantine pests (FAO, 2018)
- Quarantine pest
A pest of potential economic importance to the area endangered thereby and not yet present there, or present but not widely distributed and being officially controlled (FAO, 2018)
- Risk reduction option (RRO)
A measure acting on pest introduction and/or pest spread and/or the magnitude of the biological impact of the pest should the pest be present. A RRO may become a phytosanitary measure, action or procedure according to the decision of the risk manager
- Spread (of a pest)
Expansion of the geographical distribution of a pest within an area (FAO, 2018)
Appendix A – Distribution of Fusarium oxysporum f. sp. cubense Tropical Race 4
Distribution records based on the following sources: EPPO Global Database (EPPO online), CABI Invasive Species Compendium, Web of Science, Scopus, Google Scholar.
| Region | Country | Subnational (e.g. State) | Status |
|---|---|---|---|
| South America | Colombia | Guajira Department | Present, no details |
| Peru | Querecotillo District (Sullana Province, Piura Department) | Present, no details | |
| Africa | Mayotte |
Poroani, south west Mayotte Kongou, Bouéni |
Present, no details |
| Mozambique | Monapo District, Nampula District | Present, no details | |
| Asia | China |
Fujian Province, Guandong Province (Panyu district, Zhongshen), Guanxi Autonomous Region, Hainan Province, Yunnan Province |
Present, no details |
| India | Bihar State, Gujarat State, Madhya Pradesh State, Uttar Pradesh State | Present, no details | |
| Indonesia | Tanah Bumbu Regency (Kalimantan Province), Tana Jampea, Sulawesi | Present, no details | |
| Israel | Shfeya, Ein Gev, Masada, Gesher, Degania | Present, under eradication | |
| Java | Bogol | Present, no details | |
| Jordan | Jordan Valley | Present, no details | |
| Laos | Luang Namtha Province, Bokeo Province, Vientiane Province | Present, no details | |
| Lebanon | Mansouri | Present, no details | |
| Malaysia | Seremban, Port Dickson, Tampin, Jelebu, Jempol, Selangor, Melaka, Perak, Kuantan, Pasir Raja, Dungan, Trenggann, Kuala Krai, Kota Bharu, Kampung Terasil, Tanah Merah | Present, no details | |
| Myanmar | N.A. | Present, no details | |
| Oman | N.A. | Present, no details | |
| Pakistan | Sindh Province | Present, no details | |
| West bank | Jericho | Present, no details | |
| Papua | N.A. | Present, no details | |
| Philippines | Calinan, Davao District, Mindanao Island | Present, no details | |
| Sumatra | Lampung District | Present, no details | |
| Republic of China (Taiwan) | Present, no details | ||
| Thailand | Chiang Rai Province | Present, no details | |
| Turkey | Alanya, Anamur, Gazipasa | Present, no details | |
| Vietnam | Hanoi, Hung Yen, Lao Cai Province | ||
| Australia |
Northern Territory (Berry Springs, Lambell’s Lagoon, Middle Point, Humpty Doo) Queensland (Tully) |
Present, no details | |
| Oceania |
N.A.: Not Available.
Appendix B – Fusarium oxysporum f. sp. cubense Tropical Race 4 host plants
Sources: EPPO Global Database (EPPO, online), CABI Invasive Species Compendium, Web of Science, Scopus, Google Scholar.
| Host status | Host name | Plant family | Common name | Reference |
|---|---|---|---|---|
| Cultivated hosts | Musa acuminata | Musaceae | Cavendish Banana | Stover (1962) |
| Musa balbisiana | Musaceae | Plantain | Stover (1962) | |
| Musa × paradisiaca | Musaceae | Edible banana | Stover (1962) | |
| Musa schizocarpa | Musaceae | Not reported | Stover (1962) | |
| Musa textilis | Musaceae | Abacà, Manila Hemp | Stover (1962) | |
| Wild weed hosts | Chloris inflata= Chloris barbata | Poaceae | Purpletop Chloris | Hennessy et al. (2005) |
| Cyanthillium cinereum | Asteraceae | Little Ironweed | Hennessy et al. (2005) | |
| Tridax procumbens | Asteraceae | Coat Buttons | Hennessy et al. (2005) |
Appendix C – EU 27 annual imports of fresh and dried produce and live indoor and outdoor plants from countries where Fusarium oxysporum f. sp. cubense TR4 is present, 2016–2020 (in 100 kg)
Source: EUROSTAT accessed on 23/10/2021
| 2016 | 2017 | 2018 | 2019 | 2020 | ||
|---|---|---|---|---|---|---|
| Bananas and plantains fresh and dried | Colombia | 10,120,590.13 | 11,594,479.46 | 11,286,833.38 | 11,524,355.75 | 12,214,425.97 |
| Peru | 1,137,458.40 | 1,155,371.63 | 1,259,803.18 | 1,084,569.14 | 1,006,730.16 | |
| Mozambique | 0.00 | 2,010.72 | 664.56 | |||
| China | 252.64 | 188.73 | 390.56 | 545.74 | 854.93 | |
| India | 515.19 | 445.99 | 571.13 | 607.74 | 1,418.91 | |
| Jordan | 0.00 | |||||
| Israel | 2.10 | 0.00 | 0.75 | |||
| Indonesia | 0.01 | 37.27 | 14.72 | 64.17 | ||
| Lebanon | 170.23 | 0.00 | 0.05 | |||
| Malaysia | 0.00 | 8.02 | ||||
| Pakistan | 0.00 | 2.60 | 49.70 | |||
| Philippines | 2,480.90 | 11,415.47 | 1,674.92 | 2,160.35 | 1,240.80 | |
| Thailand | 550.44 | 674.34 | 603.32 | 526.15 | 334.58 | |
| Turkey | 202.06 | 0.00 | 210.60 | 0.14 | ||
| Australia | 0.00 | 0.01 | ||||
| Sum | 11,262,222.09 | 12,762,575.63 | 12,552,145.70 | 12,613,494.04 | 13,225,070.28 |
| 2016 | 2017 | 2018 | 2019 | 2020 | ||
|---|---|---|---|---|---|---|
| Live indoor plants | Colombia | 2.59 | 8.74 | 2.80 | 2.39 | 18.63 |
| Peru | 4.19 | 2.75 | 2.70 | 79.30 | 0.27 | |
| Mozambique | 0.45 | 0.00 | ||||
| China | 113,690.36 | 209,159.22 | 256,728.45 | 309,102.76 | 357,880.19 | |
| India | 43.29 | 60.91 | 21.66 | 410.13 | 5.43 | |
| Jordan | 0.00 | 8.34 | ||||
| Israel | 346.85 | 1,842.10 | 4,767.40 | 2,663.21 | 2,160.19 | |
| Indonesia | 2,278.41 | 2,861.74 | 7,516.78 | 11,962.96 | 11,340.52 | |
| Lebanon | 1.15 | |||||
| Malaysia | 9,663.04 | 25,680.63 | 32,199.49 | 33,201.71 | 29,123.07 | |
| Pakistan | 0.00 | 0.36 | ||||
| Philippines | 14,936.73 | 22,468.30 | 26,969.42 | 27,487.39 | 21,029.65 | |
| Thailand | 25,353.58 | 38,355.74 | 40,476.79 | 39,918.52 | 38,160.26 | |
| Turkey | 287.95 | 1,900.77 | 376.12 | 280.83 | 652.27 | |
| Australia | 800.22 | 186.08 | 191.58 | 6.37 | 0.40 | |
| Sum | 167,407.66 | 302,528.13 | 369,253.19 | 425,115.93 | 460,379.22 |
| 2016 | 2017 | 2018 | 2019 | 2020 | ||
|---|---|---|---|---|---|---|
| Live outdoor plants | Colombia | 2.99 | 11.31 | 2.26 | 7.31 | |
| Peru | 0.00 | 0.42 | 0.26 | |||
| China | 2,008.78 | 1,820.48 | 1,757.70 | 2,402.93 | 2,226.18 | |
| India | 34.54 | 350.65 | 508.69 | 389.92 | 172.96 | |
| Jordan | 0.08 | |||||
| Israel | 2,768.08 | 1,959.35 | 2,190.75 | 3,758.35 | 2,292.20 | |
| Indonesia | 25.67 | 283.34 | 103.93 | 149.10 | 12.92 | |
| Lebanon | 4.30 | 2.90 | 2.10 | 1.50 | 0.10 | |
| Malaysia | 0.00 | 0.04 | 1.00 | 10.40 | ||
| Pakistan | 0.00 | 81.60 | ||||
| Philippines | 0.00 | 0.35 | ||||
| Thailand | 349.33 | 180.15 | 220.06 | 556.39 | 57.93 | |
| Turkey | 4,001.04 | 4,494.07 | 2,774.75 | 3,132.68 | 7,427.19 | |
| Australia | 501.98 | 3.06 | 948.13 | 52.00 | 223.80 | |
| Sum | 9,696.71 | 9,105.31 | 8,506.15 | 10,446.90 | 12,512.93 |
Suggested citation: EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard C, Baptista P, Chatzivassiliou E, Di Serio F, Gonthier P, Jaques Miret JA, Justesen AF, MacLeod A, Magnusson CS, Milonas P, Navas‐Cortes JA, Parnell S, Potting R, Stefani E, Thulke H‐H, Van der Werf W, Civera AV, Yuen J, Zappalà L, Migheli Q, Vloutoglou I, Maiorano A, Streissl F and Reignault PL, 2022. Scientific Opinion on the pest categorisation of Fusarium oxysporum f. sp. cubense Tropical Race 4. EFSA Journal 2022;20(1):7092, 32 pp. 10.2903/j.efsa.2021.7092
Requestor: European Commission
Question number: EFSA‐Q‐2021‐00546
Panel members: Claude Bragard, Paula Baptista, Elisavet Chatzivassiliou, Francesco Di Serio, Paolo Gonthier, Josep Anton Jaques Miret, Annemarie Fejer Justesen, Alan MacLeod, Christer Sven Magnusson, Panagiotis Milonas, Juan A Navas‐Cortes, Stephen Parnell, Roel Potting, Philippe L. Reignault, Hans‐Hermann Thulke, Wopke Van der Werf, Antonio Vicent, Jonathan Yuen and Lucia Zappalà.
Reproduction of the images listed below is prohibited and permission must be sought directly from the copyright holder: Figure 1: © 2018 Dita, Barquero, Heck, Mizubuti and Staver, Figure 2: © https://creativecommons.org/licenses/by‐nc‐nd/4.0/, Figure 3: © CABI
Adopted: 19 May 2021
Notes
An EPPO code, formerly known as a Bayer code, is a unique identifier linked to the name of a plant or plant pest important in agriculture and plant protection. Codes are based on genus and species names. However, if a scientific name is changed, the EPPO code remains the same. This provides a harmonised system to facilitate the management of plant and pest names in computerised databases, as well as data exchange between IT systems (Griessinger & Roy, 2015; EPPO, 2019).
References
- Aguayo J, Mostert D, Fourrier‐Jeandel C, Cerf‐Wendling I, Hostachy B, Viljoen A and Ioos R, 2017. Development of a hydrolysis probe‐based real‐time assay for the detection of tropical strains of Fusarium oxysporum f. sp. cubense race 4. PLoS One, 12, e0171767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aguayo J, Cerf‐Wendling I, Folscher AB, Fourrier‐Jeandel C, Ioos R, Mathews MC, Viljoen A, 2021. First report of Fusarium oxysporum f. sp. cubense tropical race 4 (TR4) causing banana wilt in the Island of Mayotte. Plant Disease, 105, 219. [Google Scholar]
- Baayen RP, O'Donnell K, Bonants PJ, Cigelnik E, Kroon LP, Roebroeck EJ and Waalwijk C, 2000. Gene genealogies and AFLP analyses in the Fusarium oxysporum complex identify monophyletic and nonmonophyletic formae speciales causing wilt and rot disease. Phytopathology, 90, 891–900. 10.1094/PHYTO.2000.90.8.891 [DOI] [PubMed] [Google Scholar]
- Bai T, Qin M, Li X, Fan H, Xu S, Zeng L and Zheng SJ, 2020. An Additional threat to ‘cavendish’ banana growers and traders: the infection of banana peduncles by Fusarium oxysporum f. sp. cubense Tropical Race 4 (TR4). Plant Health Progress, 21, 312–316. 10.1094/PHP-08-20-0067-RS [DOI] [Google Scholar]
- Buddenhagen I, 2009. Understanding strain diversity in Fusarium oxysporum f. sp. cubense and history of introduction of ‘Tropical Race 4’ to better manage banana production. In III International Symposium on Banana: ISHS‐ProMusa Symposium on Recent Advances in Banana Crop Protection for Sustainable 828 (pp. 193–204). [Google Scholar]
- Cheng C, Liu F, Sun X, Tian N, Mensah RA, Li D and Lai Z, 2019. Identification of Fusarium oxysporum f. sp. cubense tropical race 4 (TR4) responsive miRNAs in banana root. Scientific Reports, 9, 1–16. 10.1038/s41598-019-50130-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chittarath K, Mostert D, Crew KS, Viljoen A, Kong G, Molina AB and Thomas JE, 2018. First report of Fusarium oxysporum f. sp. cubense tropical race 4 (VCG 01213/16) associated with Cavendish bananas in Laos. Plant Disease, 102, 449. 10.1094/PDIS-08-17-1197-PDN [DOI] [Google Scholar]
- Conde BD and Pitkethley RN, 2001. Discovery, identification and management of banana fusarium wilt outbreaks in the Northern Territory of Australia. In International Workshop on the Banana Fusarium Wilt Disease, Genting Highlands Resort (Malaysia), 18–20 Oct 1999. Abstract available online: https://agris.fao.org/agris‐search/search.do?recordID=PH2001101415
- Conde BD and Pitkethley RN, 2001. Discovery, identification and management of banana fusarium wilt outbreaks in the Northern Territory of Australia. In International Workshop on the Banana Fusarium Wilt Disease, Genting Highlands Resort (Malaysia), 18–20 October 1999. Abstract available online: https://agris.fao.org/agris‐search/search.do?recordID=PH2001101415
- Cook DC, Taylor AS, Meldrum RA and Drenth A, 2015. Potential economic impact of Panama disease (tropical race 4) on the Australian banana industry. Journal of Plant Diseases and Protection, 122, 229–237. https://link.springer.com/content/pdf/ 10.1007/BF03356557.pdf [DOI] [Google Scholar]
- Correll JC, Klittich CJR and Leslie JF, 1987. Nitrate nonutilizing mutants of Fusarium oxysporum and their use in vegetative compatibility tests. Phytopathology, 77, 1640–1646. [Google Scholar]
- Curry C, 2020. Fusarium oxysporum f. sp. cubense tropical race 4 (TR4). Crop Protection Compendium, CABI, Wallingford, UK. 10.1079/CPC.59074053.20210102104. Available online: https://www.cabi.org/cpc/datasheet/59074053 [DOI] [Google Scholar]
- Daly A and Walduck G, 2006. Fusarium wilt of bananas (Panama Disease)(Fusarium oxysporum f. sp. cubense). Department of Primary Industry. Fisheries and Mines, Agnote, 151, 1–5. [Google Scholar]
- Damodaran T, Mishra VK, Jha SK, Gopal R, Rajan S and Ahmed I, 2019. First report of Fusarium wilt in banana caused by Fusarium oxysporum f. sp. cubense tropical race 4 in India. Plant Disease, 103, 1022. 10.1094/PDIS-07-18-1263-PDN [DOI] [Google Scholar]
- Davis RI, Moore NY, Bentley S, Gunua TG and Rahamma S, 2000. Further records of Fusarium oxysporum f. sp. cubense from New Guinea. Australasian Plant Pathology, 29, 224. 10.1071/AP00041 [DOI] [Google Scholar]
- Dita MA, Waalwijk C, Buddenhagen IW, Souza MT Jr and Kema GHJ, 2010. A molecular diagnostic for tropical race 4 of the banana fusarium wilt pathogen. Plant Pathology, 59, 348–357. 10.1111/j.1365-3059.2009.02221.x [DOI] [Google Scholar]
- Dita M, Barquero M, Heck D, Mizubuti ES and Staver CP, 2018. Fusarium wilt of banana: current knowledge on epidemiology and research needs toward sustainable disease management. Frontiers in Plant Science, 9, 1468. 10.3389/fpls.2018.01468 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger M, Bragard C, Caffier D, Candresse T, Chatzivassiliou E, Dehnen‐Schmutz K, Gregoire J‐C, Jaques Miret JA, MacLeod A, Navajas Navarro M, Niere B, Parnell S, Potting R, Rafoss T, Rossi V, Urek G, Van Bruggen A, Van Der Werf W, West J, Winter S, Hart A, Schans J, Schrader G, Suffert M, Kertesz V, Kozelska S, Mannino MR, Mosbach‐Schulz O, Pautasso M, Stancanelli G, Tramontini S, Vos S and Gilioli G, 2018. Guidance on quantitative pest risk assessment. EFSA Journal 2018;16(8):5350, 86 pp. 10.2903/j.efsa.2018.5350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EPPO (European and Mediterranean Plant Protection Organization) , online. EPPO Global Database. Available online: https://gd.eppo.int [Accessed: December 2021].
- EPPO publication 2019/059 . Update on the situation of Fusarium oxysporum f.sp. cubense tropical race 4 in Israel. Available online: https://gd.eppo.int/reporting/article‐6489
- FAO (Food and Agriculture Organization of the United Nations) , 2004. ISPM (International Standards for Phytosanitary Measures) 21—Pest risk analysis of regulated non‐quarantine pests. FAO, Rome, 30 pp. Available online: https://www.ippc.int/sites/default/files/documents//1323945746_ISPM_21_2004_En_2011‐11‐29_Refor.pdf
- FAO (Food and Agriculture Organization of the United Nations) , 2013. ISPM (International Standards for Phytosanitary Measures) 11—Pest risk analysis for quarantine pests. FAO, Rome, 36 pp. Available online: https://www.ippc.int/sites/default/files/documents/20140512/ispm_11_2013_en_2014‐04‐30_201405121523‐494.65%20KB.pdf
- FAO (Food and Agriculture Organization of the United Nations) , 2018. International Standards for Phytosanitary Measures. ISPM 5 Glossary of phytosanitary terms. Revised version adopted CPM 13, April 2018. FAO, Rome. Available online: https://www.ippc.int/en/publications/621/
- Fourie G, Steenkamp ET, Gordon TR and Viljoen A, 2009. Evolutionary relationships among the Fusarium oxysporum f. sp. cubense vegetative compatibility groups. Applied and Environmental Microbiology, 75, 4770–4781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- García‐Bastidas F, Ordóñez N, Konkol J, Al‐Qasim M, Naser Z, Abdelwali M, Kema GHJ, 2014. First report of Fusarium oxysporum f. sp. cubense tropical race 4 associated with Panama disease of banana outside Southeast Asia. Plant Disease, 98, 694. 10.1094/PDIS-09-13-0954-PDN [DOI] [PubMed] [Google Scholar]
- Griessinger D and Roy A‐S, 2015. EPPO codes: a brief description. Available online: https://www.eppo.int/media/uploaded_images/RESOURCES/eppo_databases/A4_EPPO_Codes_2018.pdf
- Hennessy C, Walduck G, Daly A and Padovan A, 2005. Weed hosts of Fusarium oxysporum f. sp. cubense tropical race 4 in northern Australia. Australasian Plant Pathology, 34, 115–117. 10.1071/AP04091 [DOI] [Google Scholar]
- Hermanto C, Sutanto A, Edison HS, Daniells JW, O'Neill WT, Sinohin V, Molina AB and Taylor P, 2009. Incidence and distribution of Fusarium wilt disease of banana in Indonesia. In V International Symposium on Banana: ISHS‐ProMusa Symposium on Global Perspectives on Asian Challenges 897 (pp. 313–322). 10.17660/ActaHortic.2011.897.43 [DOI]
- Huang S, Mo J, Guo T, Li Q and Pan C, 2013. Occurrence of Fusarium oxysporum f. sp. cubense Tropical Race 4 and resistance identification of banana varieties in Guangxi. Journal of Southern . Agriculture, 44, 769–772. Available online: https://www.cabdirect.org/cabdirect/abstract/20143040359 [Google Scholar]
- Hung TN, Hung NQ, Mostert D, Viljoen A, Chao CP and Molina AB, 2018. First report of Fusarium wilt on Cavendish bananas, caused by Fusarium oxysporum f. sp. cubense tropical race 4 (VCG 01213/16), in Vietnam. Plant Disease, 102, 448. Available online: https://apsjournals.apsnet.org/doi/full/ 10.1094/PDIS-08-17-1140-PDN [DOI] [Google Scholar]
- Hwang SC and Ko WH, 2004. Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan. Plant Disease, 88, 580–588. 10.1094/PDIS.2004.88.6.580 [DOI] [PubMed] [Google Scholar]
- IPPC , 2013. New banana disease found in Mozambique (Fusarium oxysporum f. sp. cubense Tropical Race 4). IPPC Official Pest ReportNo. MOZ‐03/1, FAO, Rome, Italy. Available online: https://www.cabi.org/isc/datasheet/59074053
- King M, Altdorff D, Li P, Galagedara L, Holden J and Unc A, 2018. Northward shift of the agricultural climate zone under 21 st‐century global climate change. Scientific Reports, 8, 1–10. Available online: https://www.nature.com/articles/s41598‐018‐26321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koenig RL, Ploetz RC and Kistler HC, 1997. Fusarium oxysporum f. sp. cubense consists of a small number of divergent and globally distributed clonal lineages. Phytopathology, 87, 915–923. [DOI] [PubMed] [Google Scholar]
- Kottek M, Grieser J, Beck C, Rudolf B and Rubel F, 2006. World map of the Köppen_Geiger climate classification updated. Meteorologische Zeitschrift, 15, 259–263. 10.1127/0941-2948/2006/0130 [DOI] [Google Scholar]
- Li M, Shi J, Xie X, Leng Y, Wang H, Xi P, Jiang Z, 2013. Identification and application of a unique genetic locus in diagnosis of Fusarium oxysporum f. sp. cubense tropical race 4. Canadian Journal of Plant Pathology, 35, 482–493. [Google Scholar]
- Li MH, Yang B, Leng Y, Chao CP, Liu JM, He ZF and Zhong S, 2011. Molecular characterization of Fusarium oxysporum f. sp. cubense race 1 and 4 isolates from Taiwan and Southern China. Canadian Journal of Plant Pathology, 33, 168–178. 10.1080/07060661.2011.559176 [DOI] [Google Scholar]
- Lin YH, Chang JY, Liu ET, Chao CP, Huang JW and Chang PFL, 2009. Development of a molecular marker for specific detection of Fusarium oxysporum f. sp. cubense race 4. European Journal of Plant Pathology, 123, 353–365. 10.1007/s10658-008-9372-4 [DOI] [Google Scholar]
- Lin YH, Su CC, Chao CP, Chen CY, Chang CJ, Huang JW and Chang PFL, 2013. A molecular diagnosis method using real‐time PCR for quantification and detection of Fusarium oxysporum f. sp. cubense race 4. European Journal of Plant Pathology, 135, 395–405. 10.1007/s10658-012-0096-0 [DOI] [Google Scholar]
- Magdama F, Monserrate‐Maggi L, Serrano L, Sosa D, Geiser DM and Jiménez‐Gasco MDM, 2019. Comparative analysis uncovers the limitations of current molecular detection methods for Fusarium oxysporum f. sp. cubense race 4 strains. PLoS One, 14, e0222727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malik TMTA, Roff MM, Rozeita L and Maimun T, 2013. Socio‐economic impact, research and development and policy making/regulatory of Fusarium wilt on banana in Malaysia. Available at https://www.researchgate.net/profile/Vida‐Grace‐Sinohin/publication/320466356_Proceedings_of_the_Consultation‐Workshop_on_the_Socio‐Economic_Impacts_of_Fusarium_Wilt_Disease_of_Cavendish_Banana_in_the_Asia_Pacific_Region/links/59e71145aca2721fc2304d3f/Proceedings‐of‐the‐Consultation‐Workshop‐on‐the‐Socio‐Economic‐Impacts‐of‐Fusarium‐Wilt‐Disease‐of‐Cavendish‐Banana‐in‐the‐Asia‐Pacific‐Region.pdf#page=32 [Accessed: 26 December 2019].
- Martínez‐Solórzano GE, Rey‐Brina JC, Pargas‐Pichardo RE and Manzanilla EE, 2020. Fusarium wilt by tropical race 4: Current status and presence in the American continent. Agronomía Mesoamericana, 31, 259–276. Available online: https://revistas.ucr.ac.cr/index.php/agromeso/article/download/37925/41072?inline=1 [Google Scholar]
- Maryani N, Lombard L, Poerba YS, Subandiyah S, Crous PW and Kema GHJ, 2019. Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin. Studies in Mycology, 92, 155–194. 10.1016/j.simyco.2018.06.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maymon M, Shpatz U, Harel YM, Levy E, Elkind G, Teverovsky E, Freeman S, 2018. First report of Fusarium oxysporum f. sp. cubense tropical race 4 causing Fusarium wilt of Cavendish bananas in Israel. Plant Disease, 102, 2655. [Google Scholar]
- Maymon M, Sela N, Shpatz U, Galpaz N and Freeman S, 2020. The origin and current situation of Fusarium oxysporum f. sp. cubense tropical race 4 in Israel and the Middle East. Scientific Reports, 10, 1–11. 10.1038/s41598-020-58378-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meldrum RA, Daly AM, Tran‐Nguyen LTT and Aitken EAB, 2013. Are banana weevil borers a vector in spreading Fusarium oxysporum f. sp. cubense tropical race 4 in banana plantations? Australasian Plant Pathology, 42, 543–549. 10.1007/s13313-013-0214-2 [DOI] [Google Scholar]
- Molina AB, Fabregar EG, Sinohin V, Fourie G and Viljoen A, 2008. Tropical race 4 of Fusarium oxysporum f. sp cubense causing new Panama wilt epidemics in Cavendish varieties in the Philippines. In Phytopathology (Vol. 98, No. 6, pp. S108–S108). 3340 PILOT KNOB ROAD, ST PAUL, MN 55121 USA: AMER PHYTOPATHOLOGICAL SOC.
- Molina AB, Fabregar E, Sinohin VG, Yi G and Viljoen A, 2009. Recent occurrence of Fusarium oxysporum f. sp. cubense tropical race 4 in Asia. In III International Symposium on Banana: ISHS‐ProMusa Symposium on Recent Advances in Banana Crop Protection for Sustainable 828 (pp. 109–116).
- Moore NY, Pegg KG, Allen RN and Irwin JAG, 1993. Vegetative compatibility and distribution of Fusarium oxysporum f. sp. cubense in Australia. Australian Journal of Experimental Agriculture, 33, 797–802. [Google Scholar]
- Mostert D, Molina AB, Daniells J, Fourie G, Hermanto C, Chao C‐P, Fabregar E, Sinohin VG, Masdek N, Thangavelu R, Li C, Yi G, Mostert L and Viljoen A, 2017. The distribution and host range of the banana Fusarium wilt fungus, Fusarium oxysporum f. sp. cubense, in Asia. PLoS One, 12, e0181630. 10.1371/journal.pone.0181630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Donnell K, Kistler HC, Cigelnik E and Ploetz RC, 1998. Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proceedings of the National Academy of Sciences, 95, 2044–2049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Neill WT, Pattison AB, Daniells JW, Hermanto C and Molina AB, 2009. Vegetative compatibility group analysis of Indonesian Fusarium oxysporum f. sp. cubense isolates. In V International Symposium on Banana: ISHS‐ProMusa Symposium on Global Perspectives on Asian Challenges 897 (pp. 345–351). [Google Scholar]
- O’Neill WT, Henderson J, Pattemore JA, O’Dwyer C, Perry S, Beasley DR, Tan YP, Smyth AL, Goosem CH, Thomson KM, Hobbs RL, Grice KRE, Trevorrow P, Vawdrey LL, Pathania N and Shivas RG, 2016. Detection of Fusarium oxysporum f. sp. cubense tropical race 4 strain in northern Queensland. Australasian Plant Disease . Notes, 11, 1–3. 10.1007/s13314-016-0218-1 [DOI] [Google Scholar]
- Ordoñez N, García‐Bastidas F, Laghari HB, Akkary MY, Harfouche EN, Al Awar BN and Kema GHJ, 2016. First report of Fusarium oxysporum f. sp. cubense tropical race 4 causing Panama disease in Cavendish bananas in Pakistan and Lebanon. Plant Disease, 100, 209. 10.1094/PDIS-12-14-1356-PDN [DOI] [Google Scholar]
- Özarslandan M and Akgül DS, 2020. First report of Fusarium oxysporum f. sp. cubense race 4 causing fusarium wilt disease of banana in Turkey. Plant Disease, 104, 974. Available online: https://apsjournals.apsnet.org/doi/full/ 10.1094/PDIS-09-19-1881-PDN [DOI] [Google Scholar]
- Pegg KG, Coates LM, O’Neill WT and Turner DW, 2019. The epidemiology of Fusarium wilt of banana. Frontiers in Plant Science, 10, 1395. 10.3389/fpls.2019.01395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pegg KG, Moore NY and Bentley S, 1996. Fusarium wilt of banana in Australia: a review. Australian Journal of Agricultural Research, 47, 637–650. [Google Scholar]
- Peng KC, Tai CF and Chao CP, 2013. Socio‐economic impact of Fusarium wilt on Cavendish banana in Taiwan. In Consultation‐Workshop on the Socio‐economic Impacts of Fusarium Wilt Disease of Cavendish Banana in the Asia‐Pacific Region. Waterfront Insular Hotel, Davao City, Philippines (pp. 11–15).
- Pérez‐Vicente L, Dita MA and Martínez‐de la Parte E, 2014. Prevention and diagnostic of Fusarium wilt (Panama disease) of banana caused by Fusarium oxysporum f. sp. cubense. Tropical Race 4 (TR4). Available online: https://www.cabdirect.org/cabdirect/abstract/20207200009 [Google Scholar]
- Pittaway PA, Pittaway PA, Nasir N and Pegg KG, 1999. Soil receptivity and host–pathogen dynamics in soils naturally infested with Fusarium oxysporum f. sp. cubense, the cause of Panama disease in bananas. Australian Journal of Agricultural Research, 50, 623–628. Available online: https://www.publish.csiro.au/CP/A98152 [Google Scholar]
- Plant Health Australia , 2013. Blood Disease, Plant Health Australia, Canberra, Australia. 2 pp. [Google Scholar]
- Ploetz RC, 2006. Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. cubense. Phytopathology, 96, 653–656. 10.1094/PHYTO-96-0653 [DOI] [PubMed] [Google Scholar]
- Ploetz R, Freeman S, Konkol J, Al‐Abed A, Naser Z, Shalan K, Barakat R and Israeli Y, 2015. Tropical race 4 of Panama disease in the Middle East. Phytoparasitica, 43, 283–293. 10.1007/s12600-015-0470-5 [DOI] [Google Scholar]
- Ploetz RC and Pegg KG, 2000. Fusarium wilt. Diseases of banana, abaca and enset. CABI Publishing.[Google Scholar], Wallingford, UK. [Google Scholar]
- Puhalla JE, 1985. Classification of strains of Fusarium oxysporum on the basis of vegetative compatibility. Canadian Journal of Botany, 63, 179–183. Available online: https://cdnsciencepub.com/doi/abs/ 10.1139/b85-020 [DOI] [Google Scholar]
- Qi YX, Zhang X, Pu JJ, Xie YX, Zhang HQ and Huang SL, 2008. Race 4 identification of Fusarium oxysporum f. sp. cubense from Cavendish cultivars in Hainan province, China. Australasian Plant Disease Notes, 3, 46–47. Available online: https://www.publish.csiro.au/DN/DN08018 [Google Scholar]
- Rishbeth J, 1957. Fusarium wilt of bananas in Jamaica. II. Some aspects of host parasite relationships. Annals of Botany, 21, 215–245. 10.1093/oxfordjournals.aob.a083561 [DOI] [Google Scholar]
- SENASA (SENASA confirma brote de Fusarium Raza 4 Tropical en Piura) , 2021. Peru: Servicio Nacional de Sanidad Agraria del Perú (SENASA). Available online: https://www.gob.pe/institucion/senasa/noticias/429832‐senasa‐confirma‐brote‐de‐fusarium‐raza‐4‐tropical‐en‐piura
- Solpot TC, Pangga IB, Baconguis RD and Cumagun CJR, 2016. Occurrence of Fusarium oxysporum f. sp. cubense Tropical Race 4 and other genotypes in Banana in South‐Central Mindanao, Philippines. Philippine Agricultural, Scientist, 99, 370–378. Available online: https://www.researchgate.net/profile/Christian‐Joseph‐Cumagun/publication/313823070_Occurrence_of_Fusarium_oxysporum_f_sp_cubense_Tropical_Race_4_and_Other_Genotypes_in_Banana_in_South‐Central_Mindanao_Philippines/links/58b66611aca27261e51665e9/Occurrence‐of‐Fusarium‐oxysporum‐f‐sp‐cubense‐Tropical‐Race‐4‐and‐Other‐Genotypes‐in‐Banana‐in‐South‐Central‐Mindanao‐Philippines.pdf [Google Scholar]
- Stover RH, 1962. Fusarial wilt (Panama Disease) of bananas and other Musa species. Fusarial wilt (Panama disease) of bananas and other Musa species.
- Stover RH and Malo SE, 1972. The occurrence of fusarial wilt in normally resistant “Dwarf Cavendish” banana.
- Stover RH and Waite BH, 1960. Studies on Fusarium wilt of bananas: V. Pathogenicity and distribution of F. oxysporum f. cubense races 1 and 2. Canadian Journal of Botany, 38, 51–61. [Google Scholar]
- Su HJ, Chuang TY and Kong WS, 1977. Physiological race of fusarial wilt fungus attacking Cavendish banana of Taiwan. Taiwan Banana Research Instance Special Publications, 2, 1–21. [Google Scholar]
- Thangavelu R, Mostert D, Gopi M, Devi PG, Padmanaban B, Molina AB and Viljoen A, 2019. First detection of Fusarium oxysporum f. sp. cubense tropical race 4 (TR4) on Cavendish banana in India. European Journal of Plant Pathology, 154, 777–786. [Google Scholar]
- Torres Bedoya E, Bebber D and Studholme DJ, 2021. Taxonomic revision of the banana Fusarium wilt TR4 pathogen is premature. Phytopathology, (ja). 10.1094/PHYTO-03-21-0089-LE [DOI] [PubMed] [Google Scholar]
- Viljoen A, Mostert D, Chiconela T, Beukes I, Fraser C, Dwyer J, Murray H, Amisse J, Matabuana EL, Tazan G, Amugoli OM, Mondjana A, Vaz A, Pretorius A, Bothma S, Rose LJ, Beed F, Dusunceli F, Chao C‐P and Molina A, 2020. Occurrence and spread of the banana fungus Fusarium oxysporum f. sp. cubense TR4 in Mozambique. South African Journal of Science, 116, 1–11. 10.17159/sajs.2020/8608 [DOI] [Google Scholar]
- Waite BH, 1963. Wilt of Heliconia spp. caused by Fusarium oxysporum f. sp. cubense race 3. Tropical Agriculture, 40, 299–305. [Google Scholar]
- Waite BH and Stover RH, 1960. Studies on Fusarium wilt of bananas: VI. Variability and the cultivar concept in Fusarium oxysporum f. cubense. Canadian Journal of Botany, 38, 985–994. [Google Scholar]
- Wardlaw CW, 1961. Banana diseases, including plantains and abaca. Banana diseases, including plantains and abaca.
- Wardlaw CW, 1972. Banana Diseases including Plantains and Abaca (No. Ed. 2).
- Wong CKF, Vadamalai G, Saidi NB and Zulperi D, 2019. Research progress, challenges and future perspectives on the management of fusarium wilt of banana in Malaysia: a review. Malaysian Journal of Science, 38, 47–66. 10.22452/mjs.vol38no2.4 [DOI] [Google Scholar]
- Wu K‐L, Chen W‐Z, Yang S, Wen YA, Zheng Y‐R, Anjago WM, Yun Y‐Z and Wang Z‐H, 2019. Isolation and identification of Fusarium oxysporum f. sp. cubense in Fujian Province, China. Journal of Integrative Agriculture, 18, 1905–1913. 10.1016/S2095-3119(18)62149-5 [DOI] [Google Scholar]
- Zhang X, Zhang H, Xie Y, Pu J, Qi Y and Lu Y, 2009. Monitoring Fusarium wilt in Hainan Province and preliminary pathogen characterisation. In V International Symposium on Banana: ISHS‐ProMusa Symposium on Global Perspectives on Asian Challenges 897 (pp. 471–473). (abstract Available online: https://www.actahort.org/books/897/897_64.htm).
- Zheng SJ, García‐Bastidas FA, Li X, Zeng L, Bai T, Xu S and Kema GH, 2018. New geographical insights of the latest expansion of Fusarium oxysporum f. sp. cubense tropical race 4 into the greater Mekong subregion. Frontiers in Plant Science, 9, 457. 10.3389/fpls.2018.00457 [DOI] [PMC free article] [PubMed] [Google Scholar]