

Abstract
Glucagon‐like peptide 1 (GLP‐1) is well known as a gut hormone and also acts as a neuropeptide, produced in a discrete population of caudal brainstem neurons that project widely throughout the brain. GLP‐1 receptors are expressed in many brain areas of relevance to energy balance, and stimulation of these receptors at many of these sites potently suppresses food intake. This review surveys the current evidence for effects mediated by GLP‐1 receptors on feeding behaviour at a wide array of brain sites and discusses behavioural and neurophysiological mechanisms for the effects identified thus far. Taken together, it is clear that GLP‐1 receptor activity in the brain can influence feeding by diverse means, including mediation of gastrointestinal satiation and/or satiety signalling, suppression of motivation for food reward, induction of nausea and mediation of restraint stress‐induced hypophagia, but many questions about the organization of this system remain.
LINKED ARTICLES
This article is part of a themed issue on GLP1 receptor ligands (BJP 75th Anniversary). To view the other articles in this section visit http://onlinelibrary.wiley.com/doi/10.1111/bph.v179.4/issuetoc
Keywords: feeding behavior, food reward, GLP‐1, gut‐brain communication, satiation, satiety
Abbreviations
- AP
area postrema
- AAV
adeno‐associated virus
- BNST
bed nucleus of the stria terminalis
- CCK
cholecystokinin
- CeA
central amygdala
- CPP
conditioned place preference
- CRH
corticotropin‐releasing hormone
- CTA
conditioned taste aversion
- DR
dorsal raphe
- DREADD
designer receptors exclusively activated by designer drugs
- GLP‐1
glucagon‐like peptide 1
- LDTg
lateral dorsal tegmental area
- LH
lateral hypothalamus
- mPFC
medial prefrontal cortex
- NAc
nucleus accumbens
- NTS
nucleus of the solitary tract
- PPG
preproglucagon
- PR
progressive ratio
- PVN
paraventricular nucleus of the hypothalamus
- PVT
paraventricular nucleus of the thalamus
- Sim1
single‐minded homologue 1
- vHP
ventral hippocampus
- VTA
ventral tegmental area
1. INTRODUCTION
Glucagon‐like peptide 1 (GLP‐1) is well known as an incretin hormone, and the anorexigenic effects of GLP‐1 itself, as well as long‐acting GLP‐1 receptor agonists have been a prominent focus for the last several decades. Many of the physiological effects of GLP‐1 and at least some of its behavioural effects are likely mediated by GLP‐1 receptors expressed in the periphery, but a large body of research has shown that the GLP‐1 receptors in a number of different brain regions potently affect feeding and related behaviours. Two influential papers published in 1996 demonstrated that i.c.v. injection of GLP‐1 strongly suppressed food intake in rats (Tang‐Christensen et al., 1996; Turton et al., 1996), at doses far below those required for effects when GLP‐1 is administered systemically, supporting the idea that the hypophagia was mediated by GLP‐1 receptors in the brain. These receptors are widely expressed and found in many brain regions well known to influence food intake (Merchenthaler et al., 1999). Since those initial reports, a large body of literature has focused on identifying brain regions in which GLP‐1 receptors contribute to feeding control, determining whether endogenous activation of GLP‐1 receptors plays a role and unravelling the neurophysiological and behavioural mechanisms for the effects of GLP‐1 receptor stimulation on feeding (see Figure 1). For each location at which GLP‐1 receptors are now known to affect feeding, the answers to the subsequent questions have varied, and possible species and sex differences further complicate the picture. Some regions have been examined in considerably more depth than others. However, the available data support the broad conclusion that brain GLP‐1 receptors influence ingestive behaviour in a number of ways, including effects on satiation and satiety, food reward, and mediation of stress and illness responses. This review will discuss work on GLP‐1 receptor action in brain regions from caudal to rostral and consider what these findings tell us about the role of the brain GLP‐1 system.
FIGURE 1.
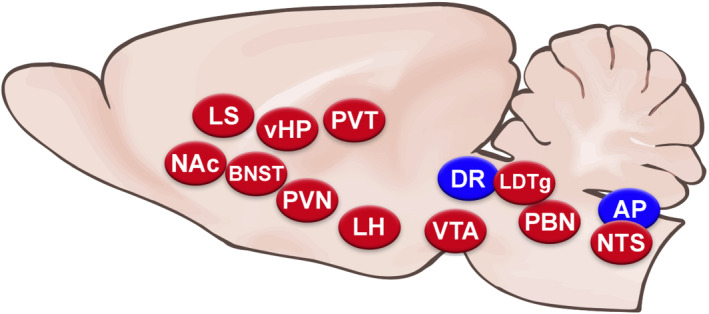
Diagram of a sagittal section of rodent brain showing the brain nuclei where GLP‐1 receptors have been shown to affect food intake. Blue ovals represent those at which GLP‐1 receptor agonist application suppresses feeding, whereas red ovals represent those at which both (1) GLP‐1 receptor agonist application suppresses feeding and (2) antagonist injection or receptor knockdown increases feeding, providing evidence that endogenous GLP‐1 receptor stimulation at these sites plays a role in feeding control
In discussing the effects of stimulating the brain GLP‐1 receptors, this review assumes the endogenous source of ligand for these receptors to originate from caudal brainstem preproglucagon (PPG) neurons, in most if not all cases. The reasons for this are discussed in depth in another review in this issue (Trapp & Brierley, 2021), but some of the evidence will be explained briefly here. First, active GLP‐1 in the circulation is so rapidly metabolized to the inactive form that little to no active GLP‐1 released by the intestine is likely to arrive in the brain, given the relatively low maximum concentrations this hormone reaches after meals (Holst & Deacon, 2005). Second, measurement of active GLP‐1 content in the brain of mice confirms concentrations to be many‐fold higher in brain tissue than in the systemic circulation and only detectable in brain regions that receive PPG neuron projections (Holt et al., 2019). In that study, selective lesion of PPG neurons in the nucleus of the solitary tract (NTS) substantially reduced active GLP‐1 measured in those brain regions that do receive PPG projections. These findings support the idea that the active GLP‐1 found in the brain comes from PPG neurons and not from the periphery. It is also consistent with earlier pharmacological data showing that in rats, blockade of brain GLP‐1 receptors by i.c.v. delivery of the antagonist, Exendin (9‐39) (Ex9), can block the ability of i.c.v.‐injected GLP‐1 to suppress feeding but not i.p. GLP‐1 (Williams et al., 2009). This distinction suggests that even if some amount of intestine‐derived GLP‐1 does access the brain, it does not stimulate GLP‐1 receptors sufficiently to alter feeding. Therefore, the behaviourally relevant endogenous ligand of most brain GLP‐1 receptors must come from PPG neurons.
It has been proposed that peripheral GLP‐1 stimulates neuronal GLP‐1 release, but recent data refute this and suggest that instead, these are two distinct systems. First, PPG neurons do not express GLP‐1 receptors and do not show an electrophysiological response to GLP‐1 receptor agonists (Card et al., 2018; Hisadome et al., 2010), so peripheral GLP‐1 could not stimulate PPG neurons directly. Instead, some have suggested that peripheral GLP‐1 activates vagal afferents, which then excite PPG neurons (e.g., Müller et al., 2019). However, Brierley et al. (2021) have presented compelling evidence that this is not the case. In that study, they showed that there is little innervation of NTS PPG neurons by GLP‐1 receptor‐expressing vagal afferents. They also reported that NTS PPG neurons received very little innervation from GLP‐1 receptor‐expressing neurons in the area postrema (AP), a circumventricular organ where circulating GLP‐1 or systemic long‐acting GLP‐1 receptor agonists could act, eliminating a second possible mechanism for peripheral–central interaction. Together, the evidence suggests that the peripheral and central GLP‐1 systems suppress feeding behaviour independently.
Degradation‐resistant GLP‐1 receptor agonists are now in use in humans for treatment of Type 2 diabetes and obesity (Aroda, 2018; Grill, 2020), and whether these long‐acting GLP‐1 receptor agonist therapies suppress food intake through the GLP‐1 receptors in the CNS is a separate question (also discussed in the review by Trapp & Brierley, 2021). When labelled versions of these compounds are delivered peripherally, they can be observed to access circumventricular organs and some hypothalamic nuclei (Gabery et al., 2020; Secher et al., 2014). Blockade or knockout of GLP‐1 receptors in the CNS can attenuate the hypophagic effect of peripheral degradation‐resistant agonist treatment (Kanoski et al., 2011; Sisley et al., 2014), suggesting that stimulation of the brain GLP‐1 receptors plays some role in the feeding response. Similar to speculation about the mechanism of action of intestine‐derived GLP‐1, it has been suggested that these degradation‐resistant GLP‐1 receptor agonists could engage the central GLP‐1 system by activating PPG neurons, but this does not appear to be the case. As noted above, NTS PPG neurons do not express GLP‐1 receptors, so a direct effect of these drugs on PPG neurons is not possible, and neither Exendin‐4 (Ex4) or semaglutide induced c‐fos, a measure of neuronal activation, in PPG neurons (Brierley et al., 2021; Holt et al., 2020). Therefore, it seems that if long‐acting GLP‐1 receptors agonists work, even in part, through GLP‐1 receptors in the CNS, they are likely to do so by accessing some of these receptors directly. Further research is needed to determine the most relevant brain locations, identify specific cell types involved and determine whether central pathways recruited by long‐acting GLP‐1 receptors agonists interact with those engaged by PPG neurons and neuronal GLP‐1 action.
2. HINDBRAIN AND MIDBRAIN REGIONS
2.1. Nucleus of the solitary tract
Application of GLP‐1 agonists into the fourth ventricle effectively suppresses feeding (Hayes et al., 2008), and because of the rostro‐caudal flow of CSF in the ventricular system, we can assume that these effects are mediated by receptors in the caudal brainstem. GLP‐1 receptors are expressed at high density in the NTS (Cork et al., 2015; Merchenthaler et al., 1999), and this nucleus has an established role in the control of food intake (Grill & Hayes, 2012), so it was a natural candidate for investigation. Many studies have shown that injection of the degradation‐resistant GLP‐1 receptor agonist exendin‐4 directly into the NTS suppresses food intake (Alhadeff & Grill, 2014; Hayes et al., 2011; Richard et al., 2015), although this could be a pharmacological effect and does not necessarily reflect a physiological role for these receptors. Loss‐of‐function studies resolve this. Injection of the antagonist, exendin‐9, directly into the caudal NTS, at a dose subthreshold for effect when delivered to the ventricle, increases food intake in rats, supporting the hypothesis that endogenous stimulation of GLP‐1 receptors in the NTS limits feeding (Hayes et al., 2009). Taking a different loss‐of‐function approach in a subsequent study, Alhadeff et al. (2017) knocked down GLP‐1 receptor expression in the NTS with adeno‐associated virus (AAV) delivery of shRNA. This manipulation caused a small but significant increase in food intake in rats 2 weeks after AAV delivery relative to subjects that received a control AAV. Together, these findings offer strong support for a physiological role of GLP‐1 receptors in the NTS in suppressing food intake.
What are the behavioural mechanisms through which GLP‐1 receptors in the NTS exert this effect? As the first stop in the brain for vagal afferents carrying information about meal‐related, gastrointestinal (GI) signals, the NTS is positioned to play a prominent role in satiation (the process of meal termination) and satiety (the continued inhibition of feeding after a meal has ended). PPG neurons in the NTS, themselves, are activated by vagal afferents and signals including the prototypical satiation signal cholecystokinin (CCK) (Hisadome et al., 2010), so it is reasonable to hypothesize that GLP‐1 receptors in the NTS play a mediatory role in satiation and/or satiety, and the evidence bears this out. Knockdown of NTS expression of GLP‐1 receptors was shown to selectively increase meal size, suggesting that these receptors normally promote satiation (Alhadeff et al., 2017). Hayes et al. (2009) asked whether caudal brainstem GLP‐1 receptors mediate the intake‐suppressive effects of either intra‐duodenal nutrient infusion or gastric balloon distention, which both strongly promote satiation and satiety. In that study, fourth‐i.c.v. pretreatment with exendin‐9 attenuated the effect of gastric distention but not the duodenal nutrient load. Although this pharmacological manipulation was not restricted to the NTS, we can assume that fourth‐i.c.v. delivery effectively blocked GLP‐1 receptors in the NTS, and therefore, these data suggest that these GLP‐1 receptors mediate the effects of some, though not all, GI negative feedback signals.
Nausea is a well‐known side effect of GLP‐1 agonists in humans (Bettge et al., 2017; also discussed in another review in this issue, Borner et al., 2021), and evidence of visceral illness as a consequence of GLP‐1 receptor stimulation can be observed in rodent models, as well, using approaches including conditioned taste aversion (CTA) and measurement of pica. For any treatment that seems to promote satiation or satiety, but especially for one such as this, it is important to ask whether that intake suppression may be a result of nausea or aversion rather than a normal physiological process. This has been investigated for the GLP‐1 receptors in the NTS and the results are inconsistent. Injection of exendin‐4 into the medial NTS induced pica and supported the formation of a CTA (Kanoski et al., 2012). However, others have shown that at least some doses of exendin‐4 delivered to the NTS can suppress food intake without inducing pica (Alhadeff & Grill, 2014; Richard et al., 2015). Therefore, although the GLP‐1 receptors in the NTS may be involved in mediation of visceral illness responses, this can be uncoupled from food intake effects.
Although the NTS has traditionally been considered to have a role in homeostatic control of feeding, responding to meal‐related GI signals and adiposity signals such as leptin (Grill & Hayes, 2012), this nucleus has direct and poly‐synaptic links to brain regions commonly considered to mediate reward and motivation. More recent data have shown that indeed, the NTS can influence food reward, and GLP‐1 receptors in the NTS alter ingestive behaviour through this mechanism. One approach to assessing food reward focuses on motivation by examining operant responding for a palatable food reinforcer on a progressive ratio (PR) schedule. The ratio requirement increases with each subsequent reinforcer, and the breakpoint—the last completed ratio before the subject ‘gives up’—is taken as a measure of motivation. A second common approach is to assess conditioned place preference (CPP), in which subjects are trained to associate a specific location with palatable food. After training, preference for that versus a location that has never been paired with food is evaluated. CPP involves associative learning, memory and motivated approach behaviour, so interpretation is not always straightforward, but it can be instructive to compare results obtained in this task with others. Two groups have shown that NTS activation of GLP‐1 receptors can alter behaviour in both of these tests; NTS injection of exendin‐4 suppressed PR responding for sucrose pellets as well as expression of a high‐fat palatable food‐induced CPP (Alhadeff & Grill, 2014; Richard et al., 2015). Importantly, the endogenous activation of the GLP‐1 receptors in the NTS seems to alter food reward, because shRNA‐mediated knockdown of GLP‐1 receptors in the NTS increased PR responding for sucrose (Alhadeff et al., 2017).
Several mechanisms for these effects of caudal hindbrain GLP‐1 receptors on feeding have been identified. The GLP‐1 receptor is a GPCR, and a great deal of investigation of GLP‐1 receptor signalling in pancreatic cells shows that it couples primarily to Gαs and increases cAMP (Pabreja et al., 2014). It is perhaps unsurprising, then, that in the hindbrain, application of GLP‐1 receptor agonists increases PKA activity. In a series of experiments using a variety of kinase inhibitors and promoters, Hayes et al. (2011) showed that the intake‐suppressive effects of hindbrain GLP‐1 receptor activation are mediated through PKA‐induced suppression of AMPK and activation of MAPK, and a subsequent study from this group showed that GLP‐1 receptors induced PI3K/PIP3 ‐induced translocation of Akt to the membrane plays a role as well (Rupprecht et al., 2013). The neurochemical phenotypes of some of the NTS neurons involved in mediating GLP‐1 receptor effects have also been identified. Richard et al. (2015) showed that PPG neuron fibres have close appositions with noradrenergic A2 neurons in the NTS, many of which project to the ventral tegmental area (VTA), the major source of dopamine in the mesolimbic reward pathway, and that NTS application of exendin‐4 potently increases tyrosine hydroxylase and D2 receptor expression in the VTA, possibly through this pathway. Recently, GABA neurons in the NTS were shown to express GLP‐1 receptors, and acute silencing of these neurons via a selectively expressed inhibitory designer receptors exclusively activated by designer drugs (DREADD) approach attenuated the effects of GLP‐1 receptor agonist treatment (Fortin et al., 2020). GLP‐1 receptors are not exclusively expressed on neurons and there is evidence that GLP‐1 receptor signalling in NTS astrocytes affects feeding, as well (Reiner et al., 2016; covered in depth in another review in this issue, Hayes & Stein, 2021).
2.2. Area postrema
Neighbouring the NTS, neurons of the AP also express GLP‐1 receptors (Cork et al., 2015; Merchenthaler et al., 1999), and this nucleus has a role in mediating the effects of satiation and satiety signals as well. It is therefore a reasonable candidate for mediation of feeding effects of GLP‐1 receptor stimulation and, as a circumventricular organ, has been suggested as a potential mediatory site for peripheral GLP‐1 agonist treatment. This is a difficult nucleus to isolate in pharmacological studies, because its location and small size render attempts at injection directly into the AP likely to deliver drug to the NTS, as well. Indirect evidence of AP mediation of effects comes from studies in the rat showing that peripheral degradation‐resistant agonist administration induces c‐Fos in AP neurons, many of which are catecholaminergic and project to brain areas associated with autonomic control (Yamamoto et al., 2003). This could be a secondary effect, not necessarily mediated by direct stimulation of GLP‐1 receptors on AP neurons, but electrophysiologic data in mice demonstrate that GLP‐1 directly activates AP neurons through AC/cAMP pathways (Kawatani et al., 2018). Few studies have attempted to examine the contribution of GLP‐1 receptors in the AP to control of food intake and data from experiments focusing on the AP are mixed, rendering it difficult to draw a firm conclusion. In one study, lesion of the AP attenuated the feeding‐suppression following GLP‐1 infusion into the hepatic portal vein (Punjabi et al., 2014). However, in another report, AP lesions did not attenuate the effect of systemic liraglutide, the long‐acting GLP‐1 agonist (Fortin et al., 2020). It may be that the AP has a role in effects of GLP‐1 itself and that the effects of degradation‐resistant agonists are mediated elsewhere. Recent work from Zhang et al. (2020) suggests that GLP‐1 receptor‐expressing AP neurons are one of several AP cell types that mediate nausea.
2.3. Parabrachial nucleus
Expression of GLP‐1 receptors in the parabrachial nucleus (PBN) (Cork et al., 2015; Merchenthaler et al., 1999) and PPG neuron projections to this area (Alhadeff et al., 2014; Richard et al., 2014) in both rats and mice have made this nucleus another target for investigation. The PBN is a complex nucleus with many subdivisions that play a role in a wide variety of physiological and behavioural functions. Of most relevance here, it is a key node for integrating information about GI signals and taste, receiving projections from both rostral and caudal NTS (Baird et al., 2001). The lateral PBN (lPBN) in particular has been examined as a site for GLP‐1 receptor‐mediated effects on feeding and, indeed, delivery of ventricle‐subthreshold doses of exendin‐4 at this location can suppress intake of chow, high‐fat diet, chocolate pellets and saccharine, without inducing pica (Alhadeff et al., 2014; Richard et al., 2014). Conversely, in these reports, blockade of GLP‐1 receptors in the lPBN with site‐specific exendin‐9 injection increased chow and high‐fat diet intake, showing that endogenous activation of these receptors plays a role in limiting ingestion. The behavioural mechanisms for these effects on feeding have not been as thoroughly investigated as for the NTS, so it is unclear whether the GLP‐1 receptors in the lPBN may be involved in satiation or satiety. However, Alhadeff et al. (2014) showed that exendin‐4 administration into the lPBN reduced PR responding for chocolate pellets, suggesting that modulation of motivation for food may mediate the effects of these GLP‐1 receptors on feeding behaviour. Richard et al. (2014) investigated potential neurophysiological mechanisms for these effects and reported that bath application of exendin‐4 to brain slices increased the firing rate of lPBN neurons. In that study, lateral‐i.c.v. injection of exendin‐4 increased expression of the mRNA for CGRP and IL‐6 in the lPBN, effects that could be indirect given that this manipulation would have accessed GLP‐1 receptors throughout the brain. However, they also reported that PPG neuron fibres were observed in close apposition to CGRP neurons in the lPBN, so it is plausible that these neurons mediate at least some of the behavioural effects observed.
2.4. Dorsal raphe
5‐HT neurons and 5‐HT receptors in the brain have long been thought to play a role in energy balance based in part on the appetite suppressive effects of 5‐HT agonists and reuptake inhibitors in both humans and rodent models (Berger et al., 2009; Burke & Heisler, 2015). The dorsal raphe (DR) is home to the largest 5‐HT neuron population, which projects widely throughout the brain into many other nuclei known to influence ingestive behaviour. GLP‐1 receptors are expressed on 5‐HT neurons in the DR (Cork et al., 2015; Merchenthaler et al., 1999), though PPG neuron projections to this location are not dense and vary by subregion (Anderberg et al., 2017; Llewellyn‐Smith et al., 2013). Limited data are available to support a physiological role of the GLP‐1 receptors in the DR in ingestive behaviour, but one report showed that injection of GLP‐1 or exendin‐4 into the DR of rats strongly suppressed feeding (Anderberg et al., 2017). Ex4 application to brain slices increased excitability of 5‐HT neurons in the DR, suggesting a potential neurophysiological mechanism for these effects. Without more detailed analysis of the feeding effects, it is difficult to speculate on the underlying behavioural mechanisms for this intake suppression, and malaise has not been ruled out for this site. It is also unclear whether the reported effects reflect a physiological role for these receptors or if this is entirely pharmacological in nature.
2.5. Lateral dorsal tegmental area
The lateral dorsal tegmental area (LDTg) is not a brain area traditionally associated with control of food intake but rather has been implicated in effects of stimulant drugs of abuse, such as methamphetamine and nicotine, through modulation of the dopaminergic neurons of the VTA (Dobbs & Cunningham, 2014; Lodge & Grace, 2006). GLP‐1 receptors are expressed in this nucleus (Merchenthaler et al., 1999), and Reiner et al. (2018) showed that in the rat, the LDTg does receive projections from a small number of PPG neurons. They also reported that intra‐LDTg injection of exendin‐4 suppressed feeding by selective reduction in meal size, with no induction of pica (Reiner et al., 2018). Importantly, blockade of the GLP‐1 receptors in the LDTg attenuated the food intake suppression normally induced by a nutrient preload. Together, these results suggest that endogenous stimulation of these LDTg receptors has a role in mediating the satiation response to GI signals. Based on the connectivity of this nucleus with the mesolimbic reward pathway, it would be reasonable to hypothesize an effect on motivation for palatable food. However, one study has shown that intra‐LDTg injection of exendin‐4 can suppress cocaine‐seeking operant responding whereas the same doses have no effect on sucrose‐seeking behaviour (Hernandez et al., 2020). There may be a role for the GLP‐1 receptors in the LDTg in a different aspect of food reward or this dissociation may reflect different roles for GLP‐1 action in this nucleus in food and drug reward.
2.6. Ventral tegmental area
As the location of the dopamine neurons whose projections form the mesolimbic reward pathway, the VTA is well known for its role in reward and motivation. This nucleus influences responses to drugs of abuse and also natural rewards, including palatable food (Meye & Adan, 2014). In rats, PPG neurons project to the VTA (Alhadeff et al., 2012; Rinaman, 2010) and a moderate level of GLP‐1 receptor expression is observed there (Merchenthaler et al., 1999). Intra‐VTA injection of exendin‐4, at low doses, has been shown to suppress intake of chow, high‐fat diet and sucrose, without inducing pica (Alhadeff et al., 2012; Dickson et al., 2012). The finding that blockade of GLP‐1 receptors in the VTA increases high‐fat diet intake (Alhadeff et al., 2012) supports the idea that this is a physiological, and not just a pharmacological, effect of these receptors. Consistent with the role of the VTA in motivation, activation of GLP‐1 receptors here suppressed PR responding for sucrose (Dickson et al., 2012). Meal pattern analysis in rats consuming high‐fat food showed that exendin‐4 injection into the VTA suppressed feeding primarily through reduction in meal size, supporting a possible role for these receptors in satiation (Mietlicki‐Baase et al., 2013). The same paper described electrophysiological evidence that GLP‐1 receptors in the VTA act pre‐synaptically via AMPA receptors to increase glutamatergic transmission to dopaminergic neurons in the VTA. A recent paper examining the effect of i.c.v. administration of exendin‐4 supports the idea that GLP‐1 receptor stimulation modulates activity of VTA dopaminergic neurons, specifically by suppressing their activity in response to palatable food‐associated cues (Konanur et al., 2020). Given that i.c.v. delivery would access GLP‐1 receptors throughout the brain, this is not necessarily a direct effect of VTA GLP‐1 receptors and this result is not consistent with the previous finding that GLP‐1 receptor stimulation targeted exclusively to this site increases excitatory drive to VTA dopamine neurons, so further research is needed to elucidate the mechanisms at play here.
3. HYPOTHALAMIC AND THALAMIC REGIONS
3.1. Hypothalamic nuclei
The hypothalamus has naturally been a target of considerable investigation, because of its longstanding importance in energy balance and because GLP‐1 receptors are expressed at high density in many hypothalamic nuclei (Cork et al., 2015; Merchenthaler et al., 1999). Because another review in this issue covers hypothalamic GLP‐1 action (K. Williams et al., 2021), this review will discuss only some of this work in brief.
The paraventricular nucleus (PVN) was one of the first hypothalamic nuclei to be targeted, probably because GLP‐1 receptors are expressed at high density here, and PPG neurons strongly innervate this nucleus (Cork et al., 2015; Gu et al., 2013; Merchenthaler et al., 1999; Rinaman, 2010). Indeed, intra‐PVN injection of GLP‐1 suppressed food intake in rats, at doses that did not support the formation of a CTA (McMahon & Wellman, 1998), and PVN injection of exendin‐9 produced a small but significant elevation of food intake (Katsurada et al., 2014), supporting a role for endogenous activation of GLP‐1 receptors at this site. The evidence in mouse models is mixed. Selective knockout of GLP‐1 receptors in single‐minded homologue 1 (Sim1) neurons, mainly found in the PVN, has no effect on food intake (Burmeister, Ayala, et al., 2017). That lack of effect could be due to developmental compensation, because Liu et al. (2017) showed that virally mediated knockdown of GLP‐1 receptors the PVN in the adult mouse increased daily food intake. PPG neurons are strongly activated by stress (Rinaman, 1999; Terrill et al., 2019), so a number of studies have examined brain GLP‐1 receptor mediation of stress responses. The PVN is an obvious candidate for such investigations, and indeed, knockout of GLP‐1 receptors in Sim1 neurons attenuates a number of physiological responses to stress (Ghosal et al., 2017). In that study, mice lacking GLP‐1 receptors in Sim1 neurons did not show the typical body weight loss induced by chronic stress, suggesting that they may be protected from stress‐induced anorexia (Ghosal et al., 2017). Corticotrophin‐releasing hormone (CRH) neurons of the PVN were identified as targets of GLP‐1 action decades ago (Larsen et al., 1997). More recent work shows that GLP‐1 receptor stimulation in the PVN increased excitatory synaptic strength in CRH neurons, through a PKA‐mediated increase in AMPA receptor trafficking to the membrane (Liu et al., 2017), suggesting that this as a possible mechanism for the behavioural effects of GLP‐1 receptors in the PVN. This would be consistent with an earlier finding that in rats that were deprived of food for 24 h before the food intake test, third‐i.c.v. pretreatment with the non‐selective CRH receptor antagonist, α‐helical CRH, completely prevented the usual intake suppression seen after GLP‐1 administration into the third ventricle (Gotoh et al., 2005).
The lateral hypothalamus (LH) is another candidate site for GLP‐1 receptor action on ingestive behaviour, especially compelling because of its well‐established involvement in food intake as well as reward and motivated behaviour in general (Stuber & Wise, 2016). Research on the effects of GLP‐1 receptors in the LH suggests complex sex differences in brain GLP‐1 function, which have largely gone unstudied. Very few reports on the effects of brain GLP‐1 receptors have used females or compared sexes. Evidence of possible sex differences in the effects of brain GLP‐1 receptors effects were originally shown with i.c.v. agonist administration. Thus, oestrogens increased sensitivity to lateral‐i.c.v. GLP‐1 effects on chow intake in female rats (Maske et al., 2017), and female rats were more sensitive to lateral‐i.c.v. exendin‐4‐induced suppression in motivation for food (Richard et al., 2016). In the LH, GLP‐1 receptor agonist injection suppressed motivation for food as measured by PR responding for sucrose, but surprisingly, female rats required a higher dose to show this effect than males (López‐Ferreras et al., 2018). Moreover, loss of function of the GLP‐1 receptors in the LH through antagonist injection or shRNA‐mediated knockdown increased PR responding for food in males but had no effect in females (López‐Ferreras et al., 2018).
3.2. Paraventricular nucleus of the thalamus
The paraventricular nucleus of the thalamus (PVT) has been implicated in arousal, stress responses and reward processes including motivation for food. In rats and mice, PPG neurons project to the PVT, and neurons here express GLP‐1 receptors (Cork et al., 2015; Gu et al., 2013; Llewellyn‐Smith et al., 2013; Merchenthaler et al., 1999; Rinaman, 2010). At this location, injection of GLP‐1 receptor agonists suppressed chow and high‐fat diet intake in rats, without inducing pica (Ong et al., 2017). Blockade of GLP‐1 receptors in the PVT increased food intake primarily through an increase in meal size (Ong et al., 2017). Furthermore, intra‐PVT injection of exendin‐4 suppressed PR responding for sucrose and also blocked the expression of a CPP induced by high‐fat diet (Ong et al., 2017), suggesting a role for these GLP‐1 receptors in food reward. Interestingly, in evaluating potential downstream mechanisms for the effects of the GLP‐1 receptors in the PVT, Ong et al. (2017) determined that GLP‐1 receptor agonist application to brain slices had an inhibitory effect on neurons projecting from the PVT to the nucleus accumbens (NAc), and this was mediated both pre‐ and post‐synaptically. The GLP‐1 receptors are often considered an excitatory GPCR because of its established signalling through Gαs, but these and other data (Cork et al., 2015; Hällbrink et al., 2001; Williams et al., 2018) suggest that GLP‐1 receptor signalling should be considered more heterogeneous.
4. LIMBIC AND OTHER FOREBRAIN REGIONS
4.1. Bed nucleus of the stria terminalis
The bed nucleus of the stria terminalis (BNST), part of the extended amygdala, is a complex nucleus that has been implicated in a wide array of behaviours including stress responses, drug and alcohol seeking, appetite and mating (Ch'ng et al., 2018). In both rats and mice, PPG neurons project to the BNST and GLP‐1 receptors are expressed here (Cork et al., 2015; Merchenthaler et al., 1999; Rinaman, 2010; Williams et al., 2018). In mice, intra‐BNST delivery of GLP‐1 suppressed, and exendin‐9 increased chow intake (Williams et al., 2018). In that study, when mice were maintained on high‐fat diet, the effect of intra‐BNST application of GLP‐1 was attenuated and exendin‐9 no longer had any effect. This contrasts with the ability of these treatments to strongly impact high‐fat diet intake when targeted to many other brain areas described here, and these findings suggest that modulation of GLP‐1 receptor function in the BNST could contribute to the overconsumption typically seen on high‐fat diets.
Consistent with its known involvement in stress responses, blockade of GLP‐1 receptors in the BNST can attenuate acute restraint stress‐induced hypophagia in the mouse (Williams et al., 2018), and data suggest a similar function of these GLP‐1 receptors in the rat. Zheng et al. (2019) reduced expression of GLP‐1 receptors in the anterolateral BNST of adult rats using virally delivered shRNA and found that this attenuated the hypophagia induced by the mild stressors of novelty and elevated open platform exposure and also attenuated some but not all other behavioural stress responses tested. In that same study, rats with BNST knockdown of GLP‐1 receptors showed no alteration in daily home cage food intake, suggesting that at least in rats, these GLP‐1 receptors may not have a physiological role in feeding under non‐stressed conditions. It is possible that the main behavioural role of GLP‐1 receptors in this nucleus is in responding to stress, including stress‐related modulation of feeding.
Cellular mechanisms for these GLP‐1 receptor‐mediated effects in BNST neurons have not been fully elucidated, but we have some clues. In rat anterolateral BNST, GLP‐1 receptors were found to be expressed by a subset of GABA neurons, and in the dorsal part of the anterolateral BNST, GLP‐1 receptor mRNA was co‐expressed with CRH mRNA, suggesting these cell types as candidate mediators of the effects of GLP‐1 in this nucleus (Zheng et al., 2019). Electrophysiological evidence suggests heterogeneous mechanisms. In slice preparations, 40% of identified GLP‐1 receptor‐expressing neurons in mouse BNST had an excitatory response to bath application of GLP‐1, whereas 60% of these cells were inhibited (Williams et al., 2018). Direction of response did not correlate with location within the BNST or other electrophysiological properties of the neurons but, interestingly, was correlated with response to dopamine. In that study, the projections of BNST neurons expressing GLP‐1 receptors were assessed with virally mediated cell type‐specific anterograde tracing, and fibres from these neurons were observed in many established BNST target sites, including the lateral septum (LS), PVN and central nucleus of the amygdala (CeA). Which of these projections, if any, mediate the feeding behavioural effects of activating the GLP‐1 receptors in the BNST is as yet unknown.
4.2. Nucleus accumbens
The NAc is the recipient of the VTA dopamine neuron projections that form the mesolimbic reward pathway, and this nucleus has a known role in controlling motivation for food and intake of palatable foods (Kelley, 2004). There is documented expression of GLP‐1 receptors in the caudal part of the NAc in both rats and mice, though lower density than many other locations (Cork et al., 2015; Merchenthaler et al., 1999). Around the same time that effects of GLP‐1 receptors in the VTA were first investigated, several labs examined NAc effects, as well, finding that intra‐NAc administration of GLP‐1 or exendin‐4 could suppress intake of chow, high‐fat diet and sucrose (Alhadeff et al., 2012; Dickson et al., 2012; Dossat et al., 2011), without inducing pica (Alhadeff et al., 2012). Increased intake after exendin‐9 treatment suggests that this is a function of endogenous GLP‐1 receptor stimulation in the NAc core subregion in particular (Alhadeff et al., 2012; Dossat et al., 2011). Notably, the contribution of GLP‐1 receptors in the NAc may differ in mice, because intra‐NAc injection of doses of exendin‐9 that increased feeding when delivered to other brain nuclei had no effect in mice (Williams et al., 2018).
Further analysis of the behavioural mechanisms for GLP‐1 receptor action in the NAc suggests involvement in mediation of GI satiation/satiety signalling as well as multiple aspects of food reward. Blockade of GLP‐1 receptors in the NAc core subregion increased meal size when rats consumed palatable liquid sweetened condensed milk or sucrose solutions, without increasing meal frequency, and intra‐NAc exendin‐9 attenuated the satiating effect of intragastric nutrient infusion (Dossat et al., 2013). Site‐specific injection of exendin‐4 into the NAc suppressed PR responding for sucrose reinforcers (Dickson et al., 2012), supporting a role in motivation, though it is unclear whether this is a function of endogenous GLP‐1 receptor stimulation at this site. Food palatability is another aspect of reward, which interacts with but is distinct from motivation for food. In rodents, analysis of licking behaviour can provide clues about hedonic evaluation, and in particular, variables such as the number of licks in the first minute of a meal and the number of licks per licking burst (defined as a series of licks separated by no more than 1 s) are associated with sucrose concentration (Johnson, 2018). In experiments examining how licking is affected by intra‐NAc core injection of exendin‐9, increased licks in the first minute and increased licks per burst during the early part of a sucrose solution meal suggested that blocking those GLP‐1 receptors led rats to treat this sucrose solution as though it were a higher concentration and tasted sweeter (Dossat et al., 2013). There has been relatively little investigation of the neurophysiological underpinnings of the effects on feeding of GLP‐1 receptors in the NAc, but Mietlicki‐Baase et al. (2014) determined that effects of GLP‐1 receptor stimulation in the core subregion of the NAc are at least partly mediated by the same pre‐synaptic, AMPA/kainate receptor mechanisms that were described for the VTA.
4.3. Hippocampus
The hippocampus is most closely associated with learning and memory, not food intake control, but there is a substantial body of work implicating hippocampal involvement in cognitive controls of eating (Davidson et al., 2019). The ventral hippocampus (vHP), in particular, has been identified as a site for GLP‐1 receptor effects on feeding, and this location is distinguished from all the others discussed here because although GLP‐1 receptors are expressed in the vHP, PPG neurons do not project here (Cork et al., 2015; Hsu et al., 2015; Merchenthaler et al., 1999). Intra‐vHP injection of ventricle‐subthreshold doses of exendin‐4 suppressed intake of chow and ‘Western’ diet relatively high in fat and sugar, and conversely, blockade of GLP‐1 receptors in the vHP increased intake (Hsu et al., 2015). This group also showed that the food intake effects of GLP‐1 receptors in the vHP are accounted for by suppression of meal size and that intra‐vHP exendin‐4 administration does not support the formation of a CTA (Hsu et al., 2015, 2018), suggesting that GLP‐1 receptors in the vHP are engaging normal satiation mechanisms as opposed to illness responses.
Hippocampal GLP‐1 receptors were also implicated in motivation for food in experiments in which intra‐vHP exendin‐4 suppressed PR responding for moderately high fat and sucrose pellets, and conversely, virally mediated GLP‐1 receptor knockdown in vHP increased operant responding for these reinforcers (Hsu et al., 2015, 2018). Interestingly, exendin‐4 given to the vHP had no effect in a different assessment of food reward, palatable food‐induced CPP (Hsu et al., 2015). At least some of the feeding effects of stimulating the GLP‐1 receptors in the vHP appear to be mediated through glutamatergic vHP neurons that project to the medial prefrontal cortex (mPFC), because inhibitory DREADD‐mediated silencing of this vHP glutamate projection or mPFC NMDA receptor antagonist treatment prevented vHP exendin‐4 from suppressing food intake (Hsu et al., 2018).
The results of the loss‐of‐function experiments clearly suggest that endogenous activation of GLP‐1 receptors in the vHP plays a physiological role in meal size control and motivation for food, but how does the endogenous agonist reach these receptors? Hsu et al. (2015) proposed a volume transmission route, with PPG neurons releasing GLP‐1 into the nearby lateral ventricle, through which that GLP‐1 ultimately reaches the hippocampus. This mechanism could play a role for other sites, as well, and may be especially relevant for those areas where GLP‐1 receptors are expressed, but there appears to be only weak or no PPG neuron fibre presence.
4.4. Lateral septum
The LS, which has been implicated in drug reward and stress responses, has dense expression of GLP‐1 receptors, especially in the dorsal subregion, and receives some PPG projections (Cork et al., 2015; Merchenthaler et al., 1999; Williams et al., 2018). In the rat, exendin‐4 given intra‐LS, reduced high‐fat diet intake primarily through an effect on meal size, with no induction of pica (Terrill et al., 2016). In mice, intra‐LS GLP‐1 injection decreases and exendin‐9 injection increases the size of liquid Ensure meals (Terrill et al., 2019). In both rats and mice, intra‐LS blockade of GLP‐1 receptors attenuates the satiating effect of an Ensure preload on subsequent chow intake, providing strong support for the idea that GLP‐1 receptors in this nucleus mediate at least some of the satiating effects of GI nutrients (Terrill et al., 2016, 2019). Blockade of the GLP‐1 receptors in the LS increased PR responding for sucrose reinforcers in both species, as well, though the effect was more substantial in mice, and intra‐LS injection of exendin‐9 blunted the hypophagic response to restraint stress in rats and mice (Terrill et al., 2018, 2019), suggesting multiple mechanisms of action for GLP‐1 receptors at this site.
The identity of the LS neurons through which GLP‐1 receptor activation affects these feeding behaviours has not been well studied, but one report suggests that many of these are neurotensin‐expressing neurons, and they may modulate feeding through a projection to the LH (Azevedo et al., 2020). In that study, selective chemogenetic activation of LS neurons expressing GLP‐1 receptors suppressed feeding in mice, and the authors showed that many of these GLP‐1 receptors and neurotensin neurons in the LS project to the LH. The activity of these neurons was linked with stressful situations in which the animal could attempt active escape (Azevedo et al., 2020), so this may be a pathway involved in brain GLP‐1's mediation of stress responses. Another study, focused on the role of these LS GLP‐1 receptors in responses to cocaine, showed that in some parts of the LS, GLP‐1 receptor‐expressing neurons overlap with fields of dopamine fibres, which originate from the VTA, raising the possibility of interaction between these two systems at this site (Reddy et al., 2016). In a slice preparation, Reddy et al. (2016) found that GLP‐1 application increased dopamine transporter surface expression and dopamine uptake in the LS, supporting this hypothesis. However, it remains to be investigated whether these interactions with dopamine in the LS mediate any of the food intake or motivation effects of GLP‐1 receptor activation at this site.
5. CONCLUSIONS
The evidence discussed in this survey of brain regions where GLP‐1 receptors affect feeding allows us to draw several conclusions about the brain GLP‐1 system, and about brain control of ingestive behaviour in general. These data also raise important questions for future research.
Taken together, it is clear that stimulation of GLP‐1 receptors throughout the brain suppresses food intake through diverse behavioural mechanisms, which are in some but not all cases shared across brain regions (see Figure 2). GLP‐1 receptors in the CNS can mediate nausea, aversion and responses to stressors such as restraint, but these can be dissociated from feeding effects in some brain regions. A number of brain structures in which GLP‐1 receptor stimulation reduces eating have been discussed, without evidence of viscerosensory malaise, and in at least one location, the CeA, stimulation of GLP‐1 receptors supports the formation of a CTA but has no effect on food intake (Kinzig et al., 2002). In the majority of sites where meal pattern effects have been examined, stimulation of GLP‐1 receptors suppresses food intake through a reduction in meal size, and at some locations, endogenous GLP‐1 receptor activation plays a mediatory role in the intake‐suppressive effects of meal‐related GI signals. Suppression of food reward is a common finding across many regions, though effects on specific behavioural tasks are not always consistent across brain sites (e.g., food‐CPP affected by exendin‐4 injection into some but not other nuclei). Therefore, the specific behavioural mechanisms underlying brain GLP‐1 receptor effects on feeding behaviour should not be considered homogenous across brain sites, and each location deserves its own thorough investigation.
FIGURE 2.

Summary of behavioural mechanisms through which brain GLP‐1 receptors (GLP‐1R) appear to suppress food intake (amplifying satiation; suppressing food reward; inducing nausea; and promoting stress responses), along with the brain areas implicated in each of these types of responses thus far. *GLP‐1 receptor stimulation in the central nucleus of the amygdala (CeA) appears to induce nausea without hypophagia
More broadly, studies examining the behavioural effects of GLP‐1 receptor stimulation have contributed to a shift in view of how the brain controls feeding. In the past, sharp distinctions were often made between ‘hedonic’ and ‘homeostatic’ controls of eating and the brain regions thought to underlie these mechanisms, with limited appreciation of crosstalk between these systems. This perspective was challenged by findings that GLP‐1 receptor action in brain areas like the NTS and lPBN, more often considered to mediate homeostatic feeding, could affect food reward and that GLP‐1 receptor action in traditionally reward‐associated areas like the VTA and NAc could mediate GI satiation/satiety signalling. This evidence, in addition to other research in the field, blurs the lines between hedonic and homeostatic feeding and has advanced our understanding of how the brain responds to external and internal signals to control ingestive behaviour.
This review focused on the ingestive behavioural effects of brain GLP‐1 receptors but a significant body of research shows that these receptors have other behavioural effects, as well, and these data should be taken into consideration. GLP‐1 receptors in the CNS seem to be involved in drug and alcohol seeking (Jerlhag, 2019; also reviewed in this issue, Fink‐Jensen), drinking behaviour (McKay et al., 2014; McKay & Daniels, 2013), mating behaviour (Vestlund & Jerlhag, 2020a, 2020b) and a variety of stress responses (Ghosal et al., 2017; Maniscalco & Rinaman, 2017; also reviewed in this issue, Holt & Rinaman). We do not yet know if separate populations of PPG neurons, the endogenous source of the ligand, are activated by stressors, GI signals, drugs of abuse or other stimuli, but given the numbers of activated PPG neurons identified in such studies, substantial overlap is most likely. Some target GLP‐1 receptor populations may mediate multiple types of behavioural and physiological responses, whereas others may be involved in only one.
There is also likely to be crosstalk between GLP‐1 receptor‐expressing neurons located in different brain nuclei, given that reciprocal projections exist between many of these brain sites. A clear example of these anatomical relationships can be seen in a study that used transgenic mice expressing cre recombinase and a fluorescent reporter in GLP‐1 receptor neurons, in which anterograde tracing from GLP‐1 receptor neurons in the BNST project to the LS, where their axons surround GLP‐1 receptor‐expressing neurons in the LS (Williams et al., 2018). It is not yet known if there are direct synaptic connections from one GLP‐1 receptor neuron to another, but even if indirect, interactions between GLP‐1‐receptive brain regions could affect the behavioural outcome of GLP‐1 receptor activation.
Brain GLP‐1 effects may depend on the environmental context and/or internal state of the animal, as well. A few findings point to interoceptive state as an important modulator. Overnight food deprivation blocks the effects of CCK, restraint stress and elevated platform exposure to induce c‐fos in PPG neurons (Maniscalco et al., 2015; Maniscalco & Rinaman, 2013), suggesting that in a fasting state, PPG neurons are less likely to be activated and release GLP‐1 at projection targets. Fasting also impairs the ability of GLP‐1 given into the third ventricle to suppress food intake (Sandoval et al., 2012), and some evidence suggests that this change in sensitivity may be mediated through brain glucose sensing (Burmeister, Brown, et al., 2017; Sandoval et al., 2012). This suggests that GLP‐1 receptor signalling or downstream mediation of its effects is altered, as well as the availability of the endogenous ligand. It is easy to imagine this modulation by fasting to be adaptive; if food is scarce, the motivation to find and consume food should remain high even in the face of stimuli that normally suppress food seeking and eating.
In almost all laboratory research on the effects of GLP‐1 receptors, experiments were designed to isolate one or another behavioural response, so it is unclear how the brain GLP‐1 system functions in more naturalistic situations in which animals face a more varied environment with many behavioural options. The majority of research has put the food intake suppression effects of stimulating the GLP‐1 receptors in the CNS at the forefront, but this bears rethinking as we continue to learn more about the role of these receptors in other behavioural responses. Ultimately, it may be more accurate to consider brain GLP‐1 receptors as part of a network that enables certain interoceptive (e.g., GI signals and homeostatic challenges) and environmental cues (e.g., signals of possible danger) to broadly suppress motivated behaviour, including but not limited to food intake, in the interest of survival.
5.1. Nomenclature of targets and ligands
Key protein targets and ligands in this article are hyperlinked to corresponding entries in the IUPHAR/BPS Guide to PHARMACOLOGY (http://www.guidetopharmacology.org) and are permanently archived in the Concise Guide to PHARMACOLOGY 2019/20 (Alexander, Christopoulos et al., 2019; Alexander, Fabbro et al., 2019; Alexander, Mathie et al., 2019).
ACKNOWLEDGEMENTS
Work in the Williams laboratory is supported by NIH (National Institute of Diabetes and Digestive and Kidney Diseases) grant R01DK095757 to DLW.
Williams, D. L. (2022). The diverse effects of brain glucagon‐like peptide 1 receptors on ingestive behaviour. British Journal of Pharmacology, 179(4), 571–583. 10.1111/bph.15535
Funding information NIH (National Institute of Diabetes and Digestive and Kidney Diseases), Grant/Award Number: R01DK095757
DATA AVAILABILITY STATEMENT
Data sharing is not applicable to this article because no new data were created or analysed in this study.
REFERENCES
- Alexander, S. P. H. , Christopoulos, A. , Davenport, A. P. , Kelly, E. , Mathie, A. , Peters, J. A. , Veale, E. L. , Armstrong, J. F. , Faccenda, E. , Harding, S. D. , Pawson, A. J. , Sharman, J. L. , Southan, C. , Davies, J. A. , & CGTP Collaborators . (2019). The Concise Guide to PHARMACOLOGY 2019/20: G protein‐coupled receptors. British Journal of Pharmacology, 176(Suppl 1), S21–S141. 10.1111/bph.14748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Fabbro, D. , Kelly, E. , Mathie, A. , Peters, J. A. , Veale, E. L. , Armstrong, J. F. , Faccenda, E. , Harding, S. D. , Pawson, A. J. , Sharman, J. L. , Southan, C. , Davies, J. A. , & CGTP Collaborators . (2019). The Concise Guide to PHARMACOLOGY 2019/20: Enzymes. British Journal of Pharmacology, 176, S297–S396. 10.1111/bph.14752 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Mathie, A. , Peters, J. A. , Veale, E. L. , Striessnig, J. , Kelly, E. , Armstrong, J. F. , Faccenda, E. , Harding, S. D. , Pawson, A. J. , Sharman, J. L. , & CGTP Collaborators . (2019). The Concise Guide to PHARMACOLOGY 2019/20: Ion channels. British Journal of Pharmacology, 176, S142–S228. 10.1111/bph.14749 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alhadeff, A. L. , Baird, J.‐P. , Swick, J. C. , Hayes, M. R. , & Grill, H. J. (2014). Glucagon‐like peptide‐1 receptor signaling in the lateral parabrachial nucleus contributes to the control of food intake and motivation to feed. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 39(9), 2233–2243. 10.1038/npp.2014.74 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alhadeff, A. L. , & Grill, H. J. (2014). Hindbrain nucleus tractus solitarius glucagon‐like peptide‐1 receptor signaling reduces appetitive and motivational aspects of feeding. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology, 307(4), R465–R470. 10.1152/ajpregu.00179.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alhadeff, A. L. , Mergler, B. D. , Zimmer, D. J. , Turner, C. A. , Reiner, D. J. , Schmidt, H. D. , Grill, H. J. , & Hayes, M. R. (2017). Endogenous glucagon‐like peptide‐1 receptor signaling in the nucleus tractus solitarius is required for food intake control. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 42(7), 1471–1479. 10.1038/npp.2016.246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alhadeff, A. L. , Rupprecht, L. E. , & Hayes, M. R. (2012). GLP‐1 neurons in the nucleus of the solitary tract project directly to the ventral tegmental area and nucleus accumbens to control for food intake. Endocrinology, 153(2), 647–658. 10.1210/en.2011-1443 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderberg, R. H. , Richard, J. E. , Eerola, K. , López‐Ferreras, L. , Banke, E. , Hansson, C. , Nissbrandt, H. , Berqquist, F. , Gribble, F. M. , Reimann, F. , Wernstedt Asterholm, I. , Lamy, C. M. , & Skibicka, K. P. (2017). Glucagon‐like peptide 1 and its analogs act in the dorsal raphe and modulate central serotonin to reduce appetite and body weight. Diabetes, 66(4), 1062–1073. 10.2337/db16-0755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aroda, V. R. (2018). A review of GLP‐1 receptor agonists: Evolution and advancement, through the lens of randomised controlled trials. Diabetes, Obesity & Metabolism, 20, 22–33. 10.1111/dom.13162 [DOI] [PubMed] [Google Scholar]
- Azevedo, E. P. , Tan, B. , Pomeranz, L. E. , Ivan, V. , Fetcho, R. , Schneeberger, M. , Doerig, K. R. , Liston, C. , Friedman, J. M. , & Stern, S. A. (2020). A limbic circuit selectively links active escape to food suppression. eLife, 9, e58894. 10.7554/eLife.58894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baird, J. P. , Travers, S. P. , & Travers, J. B. (2001). Integration of gastric distension and gustatory responses in the parabrachial nucleus. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology, 281(5), R1581–R1593. 10.1152/ajpregu.2001.281.5.R1581 [DOI] [PubMed] [Google Scholar]
- Berger, M. , Gray, J. A. , & Roth, B. L. (2009). The expanded biology of serotonin. Annual Review of Medicine, 60, 355–366. 10.1146/annurev.med.60.042307.110802 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bettge, K. , Kahle, M. , Abd El Aziz, M. S. , Meier, J. J. , & Nauck, M. A. (2017). Occurrence of nausea, vomiting and diarrhoea reported as adverse events in clinical trials studying glucagon‐like peptide‐1 receptor agonists: A systematic analysis of published clinical trials. Diabetes, Obesity & Metabolism, 19(3), 336–347. 10.1111/dom.12824 [DOI] [PubMed] [Google Scholar]
- Brierley, D. I. , Holt, M. K. , Singh, A. , de Araujo, A. , McDougle, M. , Vergara, M. , Afaghani, M. H. , Lee, S. J. , Scott, K. , Maske, C. , Langhans, W. , Krause, E. , de Kloet, A. , Gribble, F. M. , Reimann, F. , Rinaman, L. , de Lartigue, G. , & Trapp, S. (2021). Central and peripheral GLP‐1 systems independently suppress eating. Nature Metabolism, 3(2), 258–273. 10.1038/s42255-021-00344-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burke, L. K. , & Heisler, L. K. (2015). 5‐Hydroxytryptamine medications for the treatment of obesity. Journal of Neuroendocrinology, 27(6), 389–398. 10.1111/jne.12287 [DOI] [PubMed] [Google Scholar]
- Burmeister, M. A. , Ayala, J. E. , Smouse, H. , Landivar‐Rocha, A. , Brown, J. D. , Drucker, D. J. , Stoffers, D. A. , Sandoval, D. A. , Seeley, R. J. , & Ayala, J. E. (2017). The hypothalamic glucagon‐like peptide 1 receptor is sufficient but not necessary for the regulation of energy balance and glucose homeostasis in mice. Diabetes, 66(2), 372–384. 10.2337/db16-1102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burmeister, M. A. , Brown, J. D. , Ayala, J. E. , Stoffers, D. A. , Sandoval, D. A. , Seeley, R. J. , & Ayala, J. E. (2017). The glucagon‐like peptide‐1 receptor in the ventromedial hypothalamus reduces short‐term food intake in male mice by regulating nutrient sensor activity. American Journal of Physiology. Endocrinology and Metabolism, 313(6), E651–E662. 10.1152/ajpendo.00113.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Card, J. P. , Johnson, A. L. , Llewellyn‐Smith, I. J. , Zheng, H. , Anand, R. , Brierley, D. I. , Trapp, S. , & Rinaman, L. (2018). GLP‐1 neurons form a local synaptic circuit within the rodent nucleus of the solitary tract. The Journal of Comparative Neurology, 526(14), 2149–2164. 10.1002/cne.24482.Epub2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ch'ng, S. , Fu, J. , Brown, R. M. , McDougall, S. J. , & Lawrence, A. J. (2018). The intersection of stress and reward: BNST modulation of aversive and appetitive states. Progress in Neuro‐Psychopharmacology & Biological Psychiatry, 87(Pt A), 108–125. 10.1016/j.pnpbp.2018.01.005 [DOI] [PubMed] [Google Scholar]
- Cork, S. C. , Richards, J. E. , Holt, M. K. , Gribble, F. M. , Reimann, F. , & Trapp, S. (2015). Distribution and characterisation of glucagon‐like peptide‐1 receptor expressing cells in the mouse brain. Molecular Metabolism, 4(10), 718–731. 10.1016/j.molmet.2015.07.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davidson, T. L. , Jones, S. , Roy, M. , & Stevenson, R. J. (2019). The cognitive control of eating and body weight: It's more than what you “think”. Frontiers in Psychology, 10, 62. 10.3389/fpsyg.2019.00062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickson, S. L. , Shirazi, R. H. , Hansson, C. , Bergquist, F. , Nissbrandt, H. , & Skibicka, K. P. (2012). The glucagon‐like peptide 1 (GLP‐1) analogue, exendin‐4, decreases the rewarding value of food: A new role for mesolimbic GLP‐1 receptors. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 32(14), 4812–4820. 10.1523/JNEUROSCI.6326-11.2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dobbs, L. K. , & Cunningham, C. L. (2014). The role of the laterodorsal tegmental nucleus in methamphetamine conditioned place preference and locomotor activity. Behavioural Brain Research, 265, 198–202. 10.1016/j.bbr.2014.02.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dossat, A. M. , Diaz, R. , Gallo, L. , Panagos, A. , Kay, K. , & Williams, D. L. (2013). Nucleus accumbens GLP‐1 receptors influence meal size and palatability. American Journal of Physiology. Endocrinology and Metabolism, 304(12), E1314–E1320. 10.1152/ajpendo.00137.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dossat, A. M. , Lilly, N. , Kay, K. , & Williams, D. L. (2011). Glucagon‐like peptide 1 receptors in nucleus accumbens affect food intake. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 31(41), 14453–14457. 10.1523/JNEUROSCI.3262-11.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fortin, S. M. , Lipsky, R. K. , Lhamo, R. , Chen, J. , Kim, E. , Borner, T. , Schmidt, H. D. , & Hayes, M. R. (2020). GABA neurons in the nucleus tractus solitarius express GLP‐1 receptors and mediate anorectic effects of liraglutide in rats. Science Translational Medicine, 12(533), eaay8071. 10.1126/scitranslmed.aay8071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gabery, S. , Salinas, C. G. , Paulsen, S. J. , Ahnfelt‐Rønne, J. , Alanentalo, T. , Baquero, A. F. , Buckley, S. T. , Farkas, E. , Fekete, C. , Frederiksen, K. S. , Helms, H. C. C. , Jeppesen, J. F. , John, L. M. , Pyke, C. , Nøhr, J. , Lu, T. T. , Polex‐Wolf, J. , Prevot, V. , Raun, K. , … Knudsen, L. B. (2020). Semaglutide lowers body weight in rodents via distributed neural pathways. JCI Insight, 5(6), e133429. 10.1172/jci.insight.133429 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosal, S. , Packard, A. E. B. , Mahbod, P. , McKlveen, J. M. , Seeley, R. J. , Myers, B. , Ulrich‐Lai, Y. , Smith, E. P. , D'Alessio, D. A. , & Herman, J. P. (2017). Disruption of glucagon‐like peptide 1 signaling in Sim1 neurons reduces physiological and behavioral reactivity to acute and chronic stress. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 37(1), 184–193. 10.1523/JNEUROSCI.1104-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gotoh, K. , Fukagawa, K. , Fukagawa, T. , Noguchi, H. , Kakuma, T. , Sakata, T. , & Yoshimatsu, H. (2005). Glucagon‐like peptide‐1, corticotropin‐releasing hormone, and hypothalamic neuronal histamine interact in the leptin‐signaling pathway to regulate feeding behavior. The FASEB Journal, 19(9), 1131–1133. 10.1096/fj.04-2384fje [DOI] [PubMed] [Google Scholar]
- Grill, H. J. (2020). A role for GLP‐1 in treating hyperphagia and obesity. Endocrinology, 161(8), 1–14. 10.1210/endocr/bqaa093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grill, H. J. , & Hayes, M. R. (2012). Hindbrain neurons as an essential hub in the neuroanatomically distributed control of energy balance. Cell Metabolism, 16(3), 296–309. 10.1016/j.cmet.2012.06.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu, G. , Roland, B. , Tomaselli, K. , Dolman, C. S. , Lowe, C. , & Heilig, J. S. (2013). Glucagon‐like peptide‐1 in the rat brain: Distribution of expression and functional implication. The Journal of Comparative Neurology, 521(10), 2235–2261. 10.1002/cne.23282 [DOI] [PubMed] [Google Scholar]
- Hällbrink, M. , Holmqvist, T. , Olsson, M. , Ostenson, C. G. , Efendic, S. , & Langel, U. (2001). Different domains in the third intracellular loop of the GLP‐1 receptor are responsible for Gαs and Gαi/Gαo activation. Biochimica et Biophysica Acta, 1546(1), 79–86. 10.1016/s0167-4838(00)00270-3 [DOI] [PubMed] [Google Scholar]
- Hayes, M. R. , Bradley, L. , & Grill, H. J. (2009). Endogenous hindbrain glucagon‐like peptide‐1 receptor activation contributes to the control of food intake by mediating gastric satiation signaling. Endocrinology, 150(6), 2654–2659. 10.1210/en.2008-1479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes, M. R. , Leichner, T. M. , Zhao, S. , Lee, G. S. , Chowansky, A. , Zimmer, D. , De Jonghe, B. C. , Kanoski, S. E. , Grill, H. J. , & Bence, K. K. (2011). Intracellular signals mediating the food intake‐suppressive effects of hindbrain glucagon‐like peptide‐1 receptor activation. Cell Metabolism, 13(3), 320–330. 10.1016/j.cmet.2011.02.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes, M. R. , Skibicka, K. P. , & Grill, H. J. (2008). Caudal brainstem processing is sufficient for behavioral, sympathetic, and parasympathetic responses driven by peripheral and hindbrain glucagon‐like‐peptide‐1 receptor stimulation. Endocrinology, 149(8), 4059–4068. 10.1210/en.2007-1743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernandez, N. S. , Weir, V. R. , Ragnini, K. , Merkel, R. , Zhang, Y. , Mace, K. , Rich, M. T. , Christopher Pierce, R. , & Schmidt, H. D. (2020). GLP‐1 receptor signaling in the laterodorsal tegmental nucleus attenuates cocaine seeking by activating GABAergic circuits that project to the VTA. Molecular Psychiatry. 10.1038/s41380-020-00957-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hisadome, K. , Reimann, F. , Gribble, F. M. , & Trapp, S. (2010). Leptin directly depolarizes preproglucagon neurons in the nucleus tractus solitarius: Electrical properties of glucagon‐like peptide 1 neurons. Diabetes, 59(8), 1890–1898. 10.2337/db10-0128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holst, J. J. , & Deacon, C. F. (2005). Glucagon‐like peptide‐1 mediates the therapeutic actions of DPP‐IV inhibitors. Diabetologia, 48(4), 612–615. 10.1007/s00125-005-1705-7 [DOI] [PubMed] [Google Scholar]
- Holt, M. K. , Cook, D. R. , Brierley, D. I. , Richards, J. E. , Reimann, F. , Gourine, A. V. , Marina, N. , & Trapp, S. (2020). PPG neurons in the nucleus of the solitary tract modulate heart rate but do not mediate GLP‐1 receptor agonist‐induced tachycardia in mice. Molecular Metabolism, 39, 101024. 10.1016/j.molmet.2020.101024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holt, M. K. , Richards, J. E. , Cook, D. R. , Brierley, D. I. , Williams, D. L. , Reimann, F. , Gribble, F. M. , & Trapp, S. (2019). Preproglucagon neurons in the nucleus of the solitary tract are the main source of brain GLP‐1, mediate stress‐induced hypophagia, and limit unusually large intakes of food. Diabetes, 68(1), 21–33. 10.2337/db18-0729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu, T. M. , Hahn, J. D. , Konanur, V. R. , Lam, A. , & Kanoski, S. E. (2015). Hippocampal GLP‐1 receptors influence food intake, meal size, and effort‐based responding for food through volume transmission. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 40(2), 327–337. 10.1038/npp.2014.175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu, T. M. , Noble, E. E. , Liu, C. M. , Cortella, A. M. , Konanur, V. R. , Suarez, A. N. , Reiner, D. J. , Hahn, J. D. , Hayes, M. R. , & Kanoski, S. E. (2018). A hippocampus to prefrontal cortex neural pathway inhibits food motivation through glucagon‐like peptide‐1 signaling. Molecular Psychiatry, 23(7), 1555–1565. 10.1038/mp.2017.91 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jerlhag, E. (2019). Gut‐brain axis and addictive disorders: A review with focus on alcohol and drugs of abuse. Pharmacology & Therapeutics, 196, 1–14. 10.1016/j.pharmthera.2018.11.005 [DOI] [PubMed] [Google Scholar]
- Johnson, A. W. (2018). Characterizing ingestive behavior through licking microstructure: Underlying neurobiology and its use in the study of obesity in animal models. International Journal of Developmental Neuroscience: The Official Journal of the International Society for Developmental Neuroscience, 64, 38–47. 10.1016/j.ijdevneu.2017.06.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanoski, S. E. , Fortin, S. M. , Arnold, M. , Grill, H. J. , & Hayes, M. R. (2011). Peripheral and central GLP‐1 receptor populations mediate the anorectic effects of peripherally administered GLP‐1 receptor agonists, liraglutide and exendin‐4. Endocrinology, 152(8), 3103–3112. 10.1210/en.2011-0174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanoski, S. E. , Rupprecht, L. E. , Fortin, S. M. , De Jonghe, B. C. , & Hayes, M. R. (2012). The role of nausea in food intake and body weight suppression by peripheral GLP‐1 receptor agonists, exendin‐4 and liraglutide. Neuropharmacology, 62(5–6), 1916–1927. 10.1016/j.neuropharm.2011.12.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katsurada, K. , Maejima, Y. , Nakata, M. , Kodaira, M. , Suyama, S. , Iwasaki, Y. , Kario, K. , & Yada, T. (2014). Endogenous GLP‐1 acts on paraventricular nucleus to suppress feeding: Projection from nucleus tractus solitarius and activation of corticotropin‐releasing hormone, nesfatin‐1 and oxytocin neurons. Biochemical and Biophysical Research Communications, 451(2), 276–281. 10.1016/j.bbrc.2014.07.116 [DOI] [PubMed] [Google Scholar]
- Kawatani, M. , Yamada, Y. , & Kawatani, M. (2018). Glucagon‐like peptide‐1 (GLP‐1) action in the mouse area postrema neurons. Peptides, 107, 68–74. 10.1016/j.peptides.2018.07.010 [DOI] [PubMed] [Google Scholar]
- Kelley, A. E. (2004). Ventral striatal control of appetitive motivation: Role in ingestive behavior and reward‐related learning. Neuroscience and Biobehavioral Reviews, 27(8), 765–776. 10.1016/j.neubiorev.2003.11.015 [DOI] [PubMed] [Google Scholar]
- Kinzig, K. P. , D'Alessio, D. A. , & Seeley, R. J. (2002). The diverse roles of specific GLP‐1 receptors in the control of food intake and the response to visceral illness. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 22(23), 10470–10476. 10.1523/JNEUROSCI.22-23-10470.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konanur, V. R. , Hsu, T. M. , Kanoski, S. E. , Hayes, M. R. , & Roitman, M. F. (2020). Phasic dopamine responses to a food‐predictive cue are suppressed by the glucagon‐like peptide‐1 receptor agonist Exendin‐4. Physiology & Behavior, 215, 112771. 10.1016/j.physbeh.2019.112771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larsen, P. J. , Tang‐Christensen, M. , & Jessop, D. S. (1997). Central administration of glucagon‐like peptide‐1 activates hypothalamic neuroendocrine neurons in the rat. Endocrinology, 138(10), 4445–4455. 10.1210/endo.138.10.5270 [DOI] [PubMed] [Google Scholar]
- Liu, J. , Conde, K. , Zhang, P. , Lilascharoen, V. , Xu, Z. , Lim, B. K. , Seeley, R. J. , Zhu, J. J. , Scott, M. M. , & Pang, Z. P. (2017). Enhanced AMPA receptor trafficking mediates the anorexigenic effect of endogenous glucagon‐like peptide‐1 in the paraventricular hypothalamus. Neuron, 96(4), 897–909.e5. 10.1016/j.neuron.2017.09.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llewellyn‐Smith, I. J. , Gnanamanickam, G. J. E. , Reimann, F. , Gribble, F. M. , & Trapp, S. (2013). Preproglucagon (PPG) neurons innervate neurochemically identified autonomic neurons in the mouse brainstem. Neuroscience, 229, 130–143. 10.1016/j.neuroscience.2012.09.071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lodge, D. J. , & Grace, A. A. (2006). The laterodorsal tegmentum is essential for burst firing of ventral tegmental area dopamine neurons. Proceedings of the National Academy of Sciences of the United States of America, 103(13), 5167–5172. 10.1073/pnas.0510715103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- López‐Ferreras, L. , Richard, J. E. , Noble, E. E. , Eerola, K. , Anderberg, R. H. , Olandersson, K. , Taing, L. , Kanoski, S. E. , Hayes, M. R. , & Skibicka, K. P. (2018). Lateral hypothalamic GLP‐1 receptors are critical for the control of food reinforcement, ingestive behavior and body weight. Molecular Psychiatry, 23(5), 1157–1168. 10.1038/mp.2017.187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maniscalco, J. W. , & Rinaman, L. (2013). Overnight food deprivation markedly attenuates hindbrain noradrenergic, glucagon‐like peptide‐1, and hypothalamic neural responses to exogenous cholecystokinin in male rats. Physiology & Behavior, 121, 35–42. 10.1016/j.physbeh.2013.01.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maniscalco, J. W. , & Rinaman, L. (2017). Interoceptive modulation of neuroendocrine, emotional, and hypophagic responses to stress. Physiology & Behavior, 176, 195–206. 10.1016/j.physbeh.2017.01.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maniscalco, J. W. , Zheng, H. , Gordon, P. J. , & Rinaman, L. (2015). Negative energy balance blocks neural and behavioral responses to acute stress by “silencing” central glucagon‐like peptide 1 signaling in rats. The Journal of Neuroscience, 35(30), 10701–10714. 10.1523/JNEUROSCI.3464-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maske, C. B. , Jackson, C. M. , Terrill, S. J. , Eckel, L. A. , & Williams, D. L. (2017). Estradiol modulates the anorexic response to central glucagon‐like peptide 1. Hormones and Behavior, 93, 109–117. 10.1016/j.yhbeh.2017.05.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKay, N. J. , & Daniels, D. (2013). Glucagon‐like peptide‐1 receptor agonist administration suppresses both water and saline intake in rats. Journal of Neuroendocrinology, 25(10), 929–938. 10.1111/jne.12086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKay, N. J. , Galante, D. L. , & Daniels, D. (2014). Endogenous glucagon‐like peptide‐1 reduces drinking behavior and is differentially engaged by water and food intakes in rats. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 34(49), 16417–16423. 10.1523/JNEUROSCI.3267-14.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McMahon, L. R. , & Wellman, P. J. (1998). PVN infusion of GLP‐1‐(7‐36) amide suppresses feeding but does not induce aversion or alter locomotion in rats. The American Journal of Physiology, 274(1), R23–R29. 10.1152/ajpregu.1998.274.1.R23 [DOI] [PubMed] [Google Scholar]
- Merchenthaler, I. , Lane, M. , & Shughrue, P. (1999). Distribution of pre‐pro‐glucagon and glucagon‐like peptide‐1 receptor messenger RNAs in the rat central nervous system. The Journal of Comparative Neurology, 403(2), 261–280. [DOI] [PubMed] [Google Scholar]
- Meye, F. J. , & Adan, R. A. H. (2014). Feelings about food: The ventral tegmental area in food reward and emotional eating. Trends in Pharmacological Sciences, 35(1), 31–40. 10.1016/j.tips.2013.11.003 [DOI] [PubMed] [Google Scholar]
- Mietlicki‐Baase, E. G. , Ortinski, P. I. , Reiner, D. J. , Sinon, C. G. , McCutcheon, J. E. , Pierce, R. C. , Roitman, M. F. , & Hayes, M. R. (2014). Glucagon‐like peptide‐1 receptor activation in the nucleus accumbens core suppresses feeding by increasing glutamatergic AMPA/kainate signaling. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 34(20), 6985–6992. 10.1523/JNEUROSCI.0115-14.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mietlicki‐Baase, E. G. , Ortinski, P. I. , Rupprecht, L. E. , Olivos, D. R. , Alhadeff, A. L. , Pierce, R. C. , & Hayes, M. R. (2013). The food intake‐suppressive effects of glucagon‐like peptide‐1 receptor signaling in the ventral tegmental area are mediated by AMPA/kainate receptors. American Journal of Physiology. Endocrinology and Metabolism, 305(11), E1367–E1374. 10.1152/ajpendo.00413.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller, T. D. , Finan, B. , Bloom, S. R. , D'Alessio, D. , Drucker, D. J. , Flatt, P. R. , Fritsche, A. , Gribble, F. , Grill, H. J. , Habener, J. F. , Holst, J. J. , Langhans, W. , Meier, J. J. , Nauck, M. , Perez‐Tilve, D. , Pocai, A. , Reimann, F. , Sandoval, D. A. , Schwartz, T. W. , & Tschöp, M. H. (2019). Glucagon‐like peptide 1 (GLP‐1). Molecular Metabolism, 30, 72–130. 10.1016/j.molmet.2019.09.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ong, Z. Y. , Liu, J.‐J. , Pang, Z. P. , & Grill, H. J. (2017). Paraventricular thalamic control of food intake and reward: Role of glucagon‐like peptide‐1 receptor signaling. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 42(12), 2387–2397. 10.1038/npp.2017.150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pabreja, K. , Mohd, M. A. , Koole, C. , Wootten, D. , & Furness, S. G. B. (2014). Molecular mechanisms underlying physiological and receptor pleiotropic effects mediated by GLP‐1R activation. British Journal of Pharmacology, 171(5), 1114–1128. 10.1111/bph.12313 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Punjabi, M. , Arnold, M. , Rüttimann, E. , Graber, M. , Geary, N. , Pacheco‐López, G. , & Langhans, W. (2014). Circulating glucagon‐like peptide‐1 (GLP‐1) inhibits eating in male rats by acting in the hindbrain and without inducing avoidance. Endocrinology, 155(5), 1690–1699. 10.1210/en.2013-1447 [DOI] [PubMed] [Google Scholar]
- Reddy, I. A. , Pino, J. A. , Weikop, P. , Osses, N. , Sørensen, G. , Bering, T. , Valle, C. , Bluett, R. J. , Erreger, K. , Wortwein, G. , Reyes, J. G. , Graham, D. , Stanwood, G. D. , Hackett, T. A. , Patel, S. , Fink‐Jensen, A. , Torres, G. E. , & Galli, A. (2016). Glucagon‐like peptide 1 receptor activation regulates cocaine actions and dopamine homeostasis in the lateral septum by decreasing arachidonic acid levels. Translational Psychiatry, 6, e809. 10.1038/tp.2016.86 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reiner, D. J. , Leon, R. M. , McGrath, L. E. , Koch‐Laskowski, K. , Hahn, J. D. , Kanoski, S. E. , Mietlicki‐Baase, E. G. , & Hayes, M. R. (2018). Glucagon‐like peptide‐1 receptor signaling in the lateral dorsal tegmental nucleus regulates energy balance. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 43(3), 627–637. 10.1038/npp.2017.225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reiner, D. J. , Mietlicki‐Baase, E. G. , McGrath, L. E. , Zimmer, D. J. , Bence, K. K. , Sousa, G. L. , Konanur, V. R. , Krawczyk, J. , Burk, D. H. , Kanoski, S. E. , Hermann, G. E. , Rogers, R. C. , & Hayes, M. R. (2016). Astrocytes regulate GLP‐1 receptor‐mediated effects on energy balance. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 36(12), 3531–3540. 10.1523/JNEUROSCI.3579-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richard, J. E. , Anderberg, R. H. , Göteson, A. , Gribble, F. M. , Reimann, F. , & Skibicka, K. P. (2015). Activation of the GLP‐1 receptors in the nucleus of the solitary tract reduces food reward behavior and targets the mesolimbic system. PLoS ONE, 10(3), e0119034. 10.1371/journal.pone.0119034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richard, J. E. , Anderberg, R. H. , López‐Ferreras, L. , Olandersson, K. , & Skibicka, K. P. (2016). Sex and estrogens alter the action of glucagon‐like peptide‐1 on reward. Biology of Sex Differences, 7, 1–16. 10.1186/s13293-016-0059-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richard, J. E. , Farkas, I. , Anesten, F. , Anderberg, R. H. , Dickson, S. L. , Gribble, F. M. , Reimann, F. , Jansson, J.‐O. , Liposits, Z. , & Skibicka, K. P. (2014). GLP‐1 receptor stimulation of the lateral parabrachial nucleus reduces food intake: Neuroanatomical, electrophysiological, and behavioral evidence. Endocrinology, 155(11), 4356–4367. 10.1210/en.2014-1248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rinaman, L. (1999). Interoceptive stress activates glucagon‐like peptide‐1 neurons that project to the hypothalamus. The American Journal of Physiology, 277(2), R582–R590. 10.1152/ajpregu.1999.277.2.R582 [DOI] [PubMed] [Google Scholar]
- Rinaman, L. (2010). Ascending projections from the caudal visceral nucleus of the solitary tract to brain regions involved in food intake and energy expenditure. Brain Research, 1350, 18–34. 10.1016/j.brainres.2010.03.059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupprecht, L. E. , Mietlicki‐Baase, E. G. , Zimmer, D. J. , McGrath, L. E. , Olivos, D. R. , & Hayes, M. R. (2013). Hindbrain GLP‐1 receptor‐mediated suppression of food intake requires a PI3K‐dependent decrease in phosphorylation of membrane‐bound Akt. American Journal of Physiology. Endocrinology and Metabolism, 305(6), E751–E759. 10.1152/ajpendo.00367.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandoval, D. , Barrera, J. G. , Stefater, M. A. , Sisley, S. , Woods, S. C. , D'Alessio, D. D. , & Seeley, R. J. (2012). The anorectic effect of GLP‐1 in rats is nutrient dependent. PLoS ONE, 7(12), e51870. 10.1371/journal.pone.0051870 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Secher, A. , Jelsing, J. , Baquero, A. F. , Hecksher‐Sørensen, J. , Cowley, M. A. , Dalbøge, L. S. , Hansen, G. , Grove, K. L. , Pyke, C. , Raun, K. , Schäffer, L. , Tang‐Christensen, M. , Verma, S. , Witgen, B. M. , Vrang, N. , & Bjerre Knudsen, L. (2014). The arcuate nucleus mediates GLP‐1 receptor agonist liraglutide‐dependent weight loss. The Journal of Clinical Investigation, 124(10), 4473–4488. 10.1172/JCI75276 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sisley, S. , Gutierrez‐Aguilar, R. , Scott, M. , D'Alessio, D. A. , Sandoval, D. A. , & Seeley, R. J. (2014). Neuronal GLP1R mediates liraglutide's anorectic but not glucose‐lowering effect. The Journal of Clinical Investigation, 124(6), 2456–2463. 10.1172/JCI72434 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stuber, G. D. , & Wise, R. A. (2016). Lateral hypothalamic circuits for feeding and reward. Nature Neuroscience, 19(2), 198–205. 10.1038/nn.4220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang‐Christensen, M. , Larsen, P. J. , Göke, R. , Fink‐Jensen, A. , Jessop, D. S. , Møller, M. , & Sheikh, S. P. (1996). Central administration of GLP‐1‐(7‐36) amide inhibits food and water intake in rats. The American Journal of Physiology, 271(4 Pt 2), R848–R856. 10.1152/ajpregu.1996.271.4.R848 [DOI] [PubMed] [Google Scholar]
- Terrill, S. J. , Holt, M. K. , Maske, C. B. , Abrams, N. , Reimann, F. , Trapp, S. , & Williams, D. L. (2019). Endogenous GLP‐1 in lateral septum promotes satiety and suppresses motivation for food in mice. Physiology & Behavior, 206, 191–199. 10.1016/j.physbeh.2019.04.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terrill, S. J. , Jackson, C. M. , Greene, H. E. , Lilly, N. , Maske, C. B. , Vallejo, S. , & Williams, D. L. (2016). Role of lateral septum glucagon‐like peptide 1 receptors in food intake. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology, 311(1), R124–R132. 10.1152/ajpregu.00460.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terrill, S. J. , Maske, C. B. , & Williams, D. L. (2018). Endogenous GLP‐1 in lateral septum contributes to stress‐induced hypophagia. Physiology & Behavior, 192, 17–22. 10.1016/j.physbeh.2018.03.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turton, M. D. , O'Shea, D. , Gunn, I. , Beak, S. A. , Edwards, C. M. , Meeran, K. , Choi, S. J. , Taylor, G. M. , Heath, M. M. , Lambert, P. D. , Wilding, J. P. , Smith, D. M. , Ghatei, M. A. , Herbert, J. , & Bloom, S. R. (1996). A role for glucagon‐like peptide‐1 in the central regulation of feeding. Nature, 379(6560), 69–72. 10.1038/379069a0 [DOI] [PubMed] [Google Scholar]
- Vestlund, J. , & Jerlhag, E. (2020a). The glucagon‐like peptide‐1 receptor agonist, exendin‐4, reduces sexual interaction behaviors in a brain site‐specific manner in sexually naïve male mice. Hormones and Behavior, 124, 104778. 10.1016/j.yhbeh.2020.104778 [DOI] [PubMed] [Google Scholar]
- Vestlund, J. , & Jerlhag, E. (2020b). Glucagon‐like peptide‐1 receptors and sexual behaviors in male mice. Psychoneuroendocrinology, 117, 104687. 10.1016/j.psyneuen.2020.104687 [DOI] [PubMed] [Google Scholar]
- Williams, D. L. , Baskin, D. G. , & Schwartz, M. W. (2009). Evidence that intestinal glucagon‐like peptide‐1 plays a physiological role in satiety. Endocrinology, 150(4), 1680–1687. 10.1210/en.2008-1045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams, D. L. , Lilly, N. A. , Edwards, I. J. , Yao, P. , Richards, J. E. , & Trapp, S. (2018). GLP‐1 action in the mouse bed nucleus of the stria terminalis. Neuropharmacology, 131, 83–95. 10.1016/j.neuropharm.2017.12.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto, H. , Kishi, T. , Lee, C. E. , Choi, B. J. , Fang, H. , Hollenberg, A. N. , Drucker, D. J. , & Elmquist, J. K. (2003). Glucagon‐like peptide‐1‐responsive catecholamine neurons in the area postrema link peripheral glucagon‐like peptide‐1 with central autonomic control sites. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 23(7), 2939–2946. 10.1523/JNEUROSCI.23-07-02939.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, C. , Kaye, J. A. , Cai, Z. , Wang, Y. , Prescott, S. L. , & Liberles, S. D. (2020). Area postrema cell types that mediate nausea‐associated behaviors. Neuron, 109, 461–472.e5. 10.1016/j.neuron.2020.11.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng, H. , Reiner, D. J. , Hayes, M. R. , & Rinaman, L. (2019). Chronic suppression of glucagon‐like peptide‐1 receptor (GLP1R) mRNA translation in the rat bed nucleus of the stria terminalis reduces anxiety‐like behavior and stress‐induced hypophagia, but prolongs stress‐induced elevation of plasma corticosterone. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 39(14), 2649–2663. 10.1523/JNEUROSCI.2180-18.2019 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing is not applicable to this article because no new data were created or analysed in this study.