

Abstract
In humans, age-related declines in vision, hearing, and touch coincide with changes in amplitude and latency of sensory-evoked potentials. These age-related differences in neural activity may be related to a common deterioration of supra-modal brain areas (e.g., PFC) that mediate activity in sensory cortices or reflect specific sensorineural impairments that may differ between sensory modalities. To distinguish between these two possibilities, we measured neuroelectric brain activity while 37 young adults (18-30 years, 18 males) and 35 older adults (60-88 years, 20 males) were presented with a rapid randomized sequence of lateralized auditory, visual, and somatosensory stimuli. Within each sensory domain, we compared amplitudes and latencies of sensory-evoked responses, source activity, and functional connectivity (via phase-locking value) between groups. We found that older adults' early sensory-evoked responses were greater in amplitude than those of young adults in all three modalities, which coincided with enhanced source activity in auditory, visual, and somatosensory cortices. Older adults also showed stronger neural synchrony than young adults between superior prefrontal and sensory cortices; and in older adults, the degree of phase synchrony was positively correlated with the magnitude of source activity in sensory areas. Critically, older adults who showed enhanced neural activity in one sensory domain also showed enhanced activity in other modalities. Together, these findings support the common cause hypothesis of aging and highlight the role of prefrontal regions in exerting top-down control over sensory cortices.
SIGNIFICANCE STATEMENT A prominent theory of aging posits that age-related declines in sensory processing across domains are related to a single common neurobiological mechanism. However, the neural evidence supporting this common cause hypothesis has remained elusive. Our study revealed robust age-related changes in three sensory domains across a range of neural metrics. Importantly, older adults who showed increased neural activity within one sensory domain also showed enhanced neural activity in the other two sensory modalities. No such relation among activity in sensory cortices was observed in young adults. Age-related increases in neural activity in sensory cortices coincided with enhanced neural synchrony between the PFC and sensory cortices, underlining the importance of the PFC in regulating sensory processing.
Keywords: aging, common cause, EEG, evoked potential, prefrontal, sensory
Introduction
Age-related declines in vision, hearing, and touch are well documented (Humes et al., 2013; Gadkaree et al., 2016; Cavazzana et al., 2018) and are often observed with changes in amplitude and latency of sensory-evoked potentials measured with EEG. For instance, the early auditory-evoked response peaking at ∼50 ms after sound onset is usually more prominent in older adults than in young adults, whereas the subsequent response peaking at ∼200 ms is frequently delayed in older adults relative to young adults (Amenedo and Diaz, 1998; Bertoli et al., 2005; Alain et al., 2012). Similarly, visual evoked responses between 80 and 250 ms after stimulus at occipital and parieto-occipital sites often increase in amplitude and latency with age (Amenedo and Diaz, 1998; Curran et al., 2001; Fernandez et al., 2013). Additionally, sensory-evoked responses elicited by somatosensory stimuli at central-parietal sites show increased response latency with age (Bolton and Staines, 2012; Strommer et al., 2017), with some studies reporting an age-related increase in amplitude (Strommer et al., 2017) and others reporting an age-related decrease in amplitude (Bolton and Staines, 2012).
The larger amplitude and longer latencies of auditory-evoked potentials (AEPs), visual-evoked potentials (VEPs), and somatosensory-evoked potentials (SEPs) may represent a common feature of aging (the common cause hypothesis) (Lindenberger and Baltes, 1994; de la Fuente et al., 2020). Notably, prefrontal damages have been associated with changes in auditory-, visual-, and somatosensory-evoked response amplitude (Knight et al., 1989; Yamaguchi and Knight, 1990), which suggest that the prefrontal cortex (PFC) exerts modulatory control over the activity in sensory cortices (Knight et al., 1999). However, fMRI studies have identified subregions within the PFC that show modality-specific attention effects (Tobyne et al., 2018; Assem et al., 2021). Evidence from fMRI studies also suggests a fractionation of executive control functions within the PFC, which showed different sensitivity to aging (Kievit et al., 2014). These more recent findings from these studies raise the possibility that age differences in sensory-evoked responses could be associated with ineffective top-down regulation arising from subregions within the PFC that are also differently affected by aging.
Research assessing age differences in sensory processing has typically focused on a single sensory domain. In the few studies that measured sensory-evoked responses in more than one modality, age-related differences were often observed in more than one sensory domain (Allison et al., 1984; Lille et al., 1991; Ceponiene et al., 2008; Price et al., 2017). These studies, however, did not examine whether the observed age differences across sensory domains were driven by the same or different individuals within the sample.
The present study sought to evaluate age-related differences in auditory-, visual-, and somatosensory-evoked responses and to assess whether age-related changes in one modality were associated with those in other modalities within the same sample of participants. First, we elected to confirm the age-related increases in auditory-, visual-, and somatosensory-evoked responses by: (1) measuring evoked potentials in the absence of an explicit behavioral task to minimize top-down attention effects; and (2) using a data-driven analysis approach to identify patterns of sensory-evoked activity that best capture age differences in sensory processing. Then, we examined age differences in the strength of sensory-evoked responses from sensory cortices using distributed source modeling. Finally, we measured functional connectivity using phase-locking values (PLVs) to assess age differences in putative synchronization between oscillatory activity in the PFC and sensory cortices.
According to the common cause hypothesis, age-related differences in sensory-evoked responses should be observed in all three modalities. If correct, these differences should also be correlated such that individuals with an age-related amplitude increase in one modality would also show increases in the other sensory modalities. Last, if older adults recruit or engage different neural resources to increase stimulus detection and inhibition efficiency, then we should observe age differences in phase synchrony between brain sources located in prefrontal and sensory cortices.
Materials and Methods
Participants
Thirty-seven young adults and 37 healthy older adults were recruited from the Rotman Research Institute participant database. Data were excluded from one healthy older adult because of attrition between study visits and from one other healthy older adult because of insert earphones that did not properly fit. Our final sample included 37 young adults (18-30 years, 18 males) and 35 older adults (60-88 years, 20 males). The young and older groups did not differ as a function of biological sex (χ2 (1, N = 72) = 0.521, p = 0.471), but older adults had significantly more years of education (M = 16.34, SD = 2.29) than young adults (M = 15.19, SD = 2.07) (t(70) = 2.248, p = 0.028), with many younger adults still enrolled in postsecondary education at the time of testing. The study protocol was approved by the Research Ethics Board of the Baycrest Center. All participants provided informed written consent and were paid a modest stipend for their participation and out-of-pocket expenses (e.g., parking).
Participants included in this study were part of a larger project that included recruitment of young adults, healthy older adults, and older adults with amnestic mild cognitive impairment. They were all native English speakers or learned English before the age of 5. Participants were excluded if they had a history of myocardial infarction, coronary artery disease, or bypass surgery. Participants were also excluded if they were taking medication known to affect cognitive functioning, including antidepressants, anticonvulsants, neuroleptics, or if they consumed recreational drugs either currently or within the year before testing. An inclusion criterion for older adults was scoring above the cutoff on the Telephone Interview for Cognitive Status-Modified (Welsh et al., 1993). They were invited for further testing if they had no history of degenerative conditions, glaucoma, cataracts (significant enough to impede vision or color blindness), and reported no history of learning disabilities, stroke, transient ischemic attack, traumatic brain injury with loss of consciousness >5 min, substance abuse disorder, neurodegenerative disease, history of intracranial surgery, and any other diagnosis of major neurologic or psychiatric disorder.
No objective measures of sensory function were obtained (i.e., auditory, visual, vibration perception thresholds) from the participants. However, none of the participants reported having hearing, visual, or cutaneous sensation problems on their fingertips, nor did they report difficulties perceiving a sample of auditory, visual, and tactile stimuli used in the study. Participants were asked whether they used a hearing aid or wore eyeglasses. Those using a hearing aid, even occasionally, were excluded from the study, whereas those using glasses were allowed to use them during the experiment.
All older adult participants were administered a battery of standardized neuropsychological tests in the domains of intellectual functioning, memory, language, processing speed, and executive functioning. These tests were used to distinguish individuals with amnestic mild cognitive impairment from healthy older adults. Only healthy older adults were included in the present study, and the cognitive profiles of all healthy older adult participants were verified by a registered neuropsychologist (N.D.A.). The neuropsychological assessment and measurement of sensory-evoked responses took place on separate days to prevent fatigue effects. Demographic, neuropsychological, and clinical data are displayed in Table 1.
Table 1.
Participant demographic and neuropsychological dataa
| Variable | Young adult [mean (SD)] (n = 37) | Older adult [mean (SD)] (n = 35) |
|
|---|---|---|---|
| Raw | Scaled | ||
| Demographics | — | ||
| Age (yr) | 22.51 (3.59) | 75.86 (6.99) | — |
| Education (yr) | 15.19 (2.07) | 16.34 (2.29) | — |
| Sex (F:M) | 19:18 | 15:20 | — |
| TICS-m | — | 37.03 (3.04) | — |
| MoCA | — | 27.09(2.34) | — |
| Estimates of IQ | |||
| WAIS III Matrix Reasoning | 24.14 (5.26) | 14.40 (2.67) | |
| Shipley Vocabulary | 35.94 (3.28) | 12.54 (2.70) | |
| Memory | |||
| CVLT-II Learning | — | 49.29 (9.77) | 13.00 (2.75) |
| CVLT-II Short Delay FR | — | 11.74 (2.42) | 11.16 (3.19) |
| CVLT-II Long Delay FR | — | 10.57 (3.32) | 11.45 (2.88) |
| WMS-R Visual PA I | — | 11.97 (3.29) | 11.71 (2.47) |
| WMS-R Visual PA II | — | 4.97 (1.40) | 11.97 (1.72) |
| WMS-R Verbal PA I | — | 16.03 (2.96) | 9.94 (2.25) |
| WMS-R Verbal PA II | — | 6.77 (1.17) | 11.54 (2.49) |
| WAIS III Digit Symbol IL FR | — | 7.31 (0.93) | 10.43 (1.07) |
| WAIS III Digit Symbol IL PR | — | 11.89 (4.32) | 10.40 (1.42) |
| Language | |||
| BNT-15 | — | 53.71 (4.93) | 11.06 (2.77) |
| Phonemic Fluency (FAS) | — | 47.40 (13.16) | 11.46 (3.46) |
| Semantic Fluency (Animals) | — | 19.00 (5.27) | 10.47 (3.59) |
| WAIS-III Digit Symbol | — | 60.97 (14.42) | 12.80 (2.77) |
| D-KEFS Trails Numbers | — | 38.97 (12.18) | 12.94 (2.18) |
| D-KEFS Trails Letters | — | 37.74 (11.41) | 12.89 (1.64) |
| D-KEFS Trails N-L Switch | — | 98.97 (37.95) | 12.03 (2.51) |
| D-KEFS Color | — | 30.55 (5.59) | 11.54 (2.22) |
| D-KEFS Word | — | 22.85 (4.56) | 11.17 (2.93) |
| D-KEFS Inhibition | — | 57.59 (13.69) | 12.91 (1.90) |
| Alpha Span | — | 28.51 (9.28) | 10.60 (3.10) |
| WCST Perseverative Errors % | — | 14.12 (10.19) | 13.20 (4.41) |
| WCST Categories | — | 4.66 (1.94) | — |
| Questionnaires | |||
| HADS Anxiety | — | 4.69 (2.89) | — |
| HADS Depression | — | 2.51 (2.16) | — |
| EPW | — | 6.69 (2.96) | — |
| PSQI | — | 5.76 (2.99) | — |
aTICS-m, modified Telephone Interview of Cognitive Status (raw score out of 50); MoCA, Montreal Cognitive Assessment; WAIS, Wechsler Adult Intelligence Scale; CVLT, California Verbal Learning Test; FR, Free Recall; WMS-R, Wechsler Memory Scale-Revised; PA, Paired Associates; PR, Paired Recall; IL, Incidental Learning; BNT, Boston Naming Test; FAS, phonemic fluency to the letters F, A, and S; D-KEFS, Delis Kaplan Executive Functioning System; N-L, Number-Letter; HADS, Hospital Anxiety and Depression Scale; EPW, Epworth Sleepiness Scale; PSQI, Pittsburgh Sleep Quality Index; WCST, Wisconsin Card Sorting Test.
Stimuli
The visual stimulus was a black and white 5 × 10 square checkerboard pattern presented for 50 ms in duration. It spanned the entire height, and ∼40% of the width of a 19 inch computer monitor and extended from either the left or right edge. The entire checkerboard stimulus subtended a visual angle of 14.3° × 28.1°, with each checkerboard square subtending a visual angle of 2.9° × 2.9°. The auditory stimulus was a monaural harmonic complex tone (50 ms in duration, 5 ms rise/fall time). It comprised the first five harmonics of the fundamental frequency of 200 Hz at sine phase and equal intensity. The tone was presented to the left or right ear through ER-3A insert earphones (Etymotic Research). The sound intensity was set at 85 decibels (dB) sound pressure level (SPL) measured by a Larson Davis 824 SPL meter using a 2 ml coupler. The somatosensory stimulus consisted of pneumatic somatosensory stimulation, simultaneously delivered to the tips of all four fingers (thumb excluded) of either the left or right hand. The somatosensory stimulation was delivered from a puff-type pressure stimulator from a compressed air tank with electromagnetic air valves kept constant at 30 psi. This stimulator drove pressurized air through plastic tubes to 8 pneumatically driven inflatable circular plastic membranes with a diameter of 1 cm each. Each membrane was worn on the participant's fingers over and slightly above the distal interphalangeal joint. During somatosensory stimulation, the valves were activated to periodically inflate the membrane and deliver brief pressure pulses of 50 ms duration to the fingertips.
Procedure
We used a paradigm in which lateralized visual, auditory, and somatosensory stimuli were presented sequentially in a randomized order to measure sensory-evoked potentials. The paradigm was administered with participants seated in a sound-attenuated booth. Before recording sensory-evoked potentials, participants were briefed on what the paradigm entailed, including details about the length of the experiment and the modes of stimulation involved. During a familiarization phase, all participants were presented with a 10 s sample of the paradigm to ensure that they could perceive the lateralized auditory, visual, and somatosensory stimulation. After the familiarization phase, participants were probed for their self-reported comfort level, specifically, whether the auditory stimulation was perceived as too loud, and whether the tactile stimulation was perceived as too strong on their fingertips. All participants reported no issues after the familiarization phase and after the experiment. Participants were also monitored from outside the sound-attenuated booth with a webcam during both the familiarization and experimental phases to ensure compliance and that no participants fell asleep.
During stimulus presentation, participants were instructed to maintain their gaze on a fixation dot 0.60 cm in diameter at the center of the computer screen set at 100% contrast level and placed 60 cm from participants. The stimulus onset asynchrony varied between 200 and 340 ms (20 ms steps, rectangular distribution). The sequence lasted ∼12 min, with a total of 1440 stimuli (480 trials per sensory modality) and evenly distributed between left and right laterality. Stimuli were presented using Presentation software (version 13, Neurobehavioral Systems). Our multisensory-evoked potential paradigm was inspired from prior clinical studies aiming to quickly assess the integrity of multiple sensory systems (Cohen et al., 1982; Werner and Vanderzant, 1991; Iodice et al., 2016).
Data acquisition and preprocessing
Neuroelectric brain activity was recorded continuously using a 76-channel ActiveTwo acquisition system (BioSemi) with a sampling rate of 512 Hz. Sixty-six electrodes were positioned on the scalp using a BioSemi head cap according to the standard 10-20 system with a Common Mode Sense active electrode and Driven Right Leg (DRL) ground electrode. Ten additional electrodes were placed below the hairline (both mastoids, pre-auricular points, two lateral ocular sites, two inferior ocular sites, and two additional frontolateral electrodes) to monitor eye movements and cover the whole scalp evenly. EEG recordings were preprocessed offline using Brain Electrical Source Analysis software (BESA Research version 7.1; MEGIS).
EEG preprocessing
The EEG data were visually inspected to identify segments contaminated by defective electrodes. Noisy electrodes were interpolated using data from the surrounding electrodes, and no more than eight electrodes were interpolated per participant. The EEG was then rereferenced to the average of all electrodes and digitally filtered with a 1 Hz high-pass filter (forward, 6 dB/octave) and 40 Hz low-pass filter (zero phase, 24 dB/octave). For each participant, a set of ocular movements was identified from the continuous EEG recording and used to generate spatial components to best account for eye movement artifacts. The spatial topographies were then subtracted from the continuous EEG to correct for lateral and vertical eye movements as well as for eye blinks. The data were parsed into 500 ms epochs that were time-locked to stimulus onset, including 100 ms of prestimulus activity. Epochs with EEG signal exceeding ±60 µV were marked and excluded from further analysis. This excluded, on average, 2.78% of epochs per participant for the young adult group and 3.94% of epochs for the older adult group. The remaining epochs were averaged according to the left- and right-lateralized AEPs, VEPs, and SEPs. Each average was then baseline-corrected with respect to the 100 ms prestimulus baseline interval. We did not have a priori hypotheses regarding possible age-related differences as a function of stimulus lateralization. Hence, as a dimension reduction method, and to improve the signal-to-noise ratio, epochs of lateralized sensory-evoked potentials were averaged together in a transposed montage such that right-lateralized potentials (elicited by left-lateralized stimuli) were kept untransposed and left-lateralized potentials (elicited by right-lateralized stimuli) were transposed across laterality. This transposition method produced epochs such that the sensory-evoked potentials, regardless of whether elicited by left or right-lateralized stimuli, occurred over the right laterality of the electrode montage.
Distributed source analysis
We used an iterative application of Low-Resolution Electromagnetic Tomography (LORETA) to model the neural sources of sensory-evoked responses, which reduces the source space in each iteration. This imaging approach, termed Classical LORETA Analysis Recursively Applied (CLARA), provides more focal localization of brain activity and can separate sources in close vicinity (Dimitrijevic et al., 2013; Beniczky et al., 2016; Zimmermann et al., 2020). Distributed source analysis with CLARA was performed on sensory-evoked responses using BESA software with a voxel size of 7 mm in the Talairach space; we found that this default setting was appropriate for the distributed images in most situations (Shen et al., 2018; Zimmermann et al., 2020). The regularization parameters that account for the noise in the data were set with a singular value decomposition cutoff at 0.01%. We used a four-shell ellipsoidal head model with a head radius of 85 mm, and thickness for scalp, bone, and CSF of 6, 7, and 1 mm, respectively. The relative conductivities were 0.33, 0.33, 0.0042, and 1 S/m for brain, scalp, bone, and CSF, respectively.
Functional connectivity
We used PLV, a measure of phase synchrony between two different time series, to examine the synchronization in oscillatory activity between brain sources (Lachaux et al., 1999). PLV can range from 0 to 1, with values close to 1 indicating strong synchrony (i.e., perfect phase locking), while values close to 0 indicate substantial phase variation between the two signals, and thus low synchronicity between the two regions (i.e., no phase synchrony and randomly dispersed phases).
To transform EEG data from scalp electrode space into source space, we used surrogate source montages provided by BESA Research software that have been specifically designed to model sensory-evoked responses elicited by auditory, visual, and somatosensory stimuli. These source montages act as spatial filters that optimally capture scalp-evoked responses with a subset of discrete brain sources, which are then used to extract time series.
To calculate PLV, the continuous EEG data were converted to the time-frequency domain with BESA Research software. The continuous EEG data were filtered at a 1 Hz high-pass filter (forward, 6 dB/octave) and no low-pass filter. The analysis epoch for the time-frequency decomposition consisted of 500 ms of prestimulus activity and 500 ms of poststimulus activity time-locked to stimulus onset. A complex demodulation method with 1-Hz-wide frequency bins and 50 ms time resolution in the range of 2 and 50 Hz was used for decomposing the single-trial EEG data into time-frequency representations. To minimize possible contamination from cross-modal phase resetting of oscillatory activity, we also calculated PLV for non–stimulus-locked epochs by inserting triggers at 2 s intervals starting at the beginning of the sequence. These triggers were not time-locked to any stimulus, and occurred at random intervals relative to a stimulus onset. The average PLV for the non–stimulus-locked activity was then used as baseline. As an additional precaution, we excluded trials that were preceded or followed by the target modality. For instance, for the auditory modality, the non–stimulus-locked average included only trials that were preceded and followed, at random intervals, by a visual or somatosensory stimulus.
We used a seed in the right auditory, visual, and somatosensory cortex for auditory, visual, and somatosensory stimulation, respectively. For the sources in the right auditory cortex, we used the dipole with the tangential orientation to best capture the early auditory P1 deflection. For the sources in the right visual cortex, we used the radial component to best capture the visual P1 and N1 deflections. For the sources in the right somatosensory cortex, we used the tangential dipole (vertical orientation), which best accounted for the somatosensory P1 and N1 deflections.
Statistical analyses
Measures of sensory-evoked response amplitude, source strength, and PLV were subjected to nonparametric cluster-based permutation testing between age groups using BESA Statistics software (Statistics 2.1, MEGIS). In a preliminary step, the software used a parametric test to identify clusters both in time (adjacent time points) and space (adjacent electrodes) where the two groups show pronounced differences in evoked response amplitude, source strength, or PLV. For each cluster in this initial step, the software calculated a cluster value that represents the sum of all t values of all data points in temporal and spatial domains. In a second subsequent stage, the software used permutation tests with a Monte-Carlo resampling technique (Maris and Oostenveld, 2007) to identify those clusters that had higher cluster values than a specified threshold (i.e., a cluster α) and then compared with a random null distribution of cluster values generated by random permutation of the data. This latter nonparametric permutation statistic is not subject to the multiple comparisons problem (for an in-depth overview of permutation statistics as implemented in BESA Statistics, see Maris and Oostenveld, 2007). Separate unpaired t tests between groups were run for each sensory modality. A time interval was set from stimulus onset (0 ms) to 300 ms after stimulus. This interval was chosen because it minimizes the contribution from possible stimuli occurring at the shortest stimulus onset asynchrony, and yet it encompasses the time window of interest for sensory-evoked responses shared among all three modalities. For analysis of sensory-evoked response amplitude, a cluster α of 0.01 was used, and the number of permutations was set at 3000. For the analyses of source activity and PLV, we used a cluster α of 0.001 and 0.01, respectively, in a time window between 0 and 300 ms after stimulus and 3000 as the number of permutations for hypothesis testing.
For scalp-recorded sensory-evoked responses, significant clusters identified by cluster-based permutation testing may not necessarily indicate a group difference in amplitude, as they can also be driven by group differences in latency of a response. Therefore, post hoc mixed model ANOVAs were performed for each sensory-evoked response ERP component identified by permutation testing to determine whether significant clusters were attributed to group differences in amplitude, latency, or both. These analyses were performed on peak amplitude and latency measurements extracted from a prespecified time window and a subset of five electrodes that best captured the sensory-evoked response of interest. These peak amplitudes and latencies were extracted using BESA Research software and subjected to a two-way mixed model ANOVA with group (young, older adults) as a between-subjects factor, and electrode as a within-subjects factor (with each electrode as a level). As the main effect of electrode and group by electrode interaction were not of interest, only group main effects are reported. An α value of 0.05 was used throughout, and the effect size was measured with partial η2.
To examine the relationship of sensory-evoked potentials across sensory modalities, bivariate two-tailed Pearson correlation analyses were run between each pair of sensory domains (i.e., visual and auditory, visual and somatosensory, auditory and somatosensory) for measures of sensory-evoked response amplitude and source strength. Separate correlation analyses were run for each age group. To further examine whether the degree of phase synchronization between the PFC and sensory cortices was related to the magnitude of source activity from sensory cortices, another set of correlation analyses were conducted. For each sensory modality, bivariate two-tailed Pearson correlation analyses were run between prefrontal-to-sensory cortex phase synchrony at low-to-middle frequency ranges on one end and source strength at each of the primary visual, auditory, and somatosensory cortices on the other end. Similarly, correlation analyses were run for each age group separately. Post hoc ANOVAs and correlation analyses were run using IBM SPSS Statistics version 26.
Results
We first demonstrated age-related differences in auditory-, visual-, and somatosensory-evoked response amplitude using data-driven cluster-based permutation testing. Figure 1 shows the group mean sensory-evoked responses elicited by auditory, visual, and somatosensory stimuli. A summary of significant clusters revealed by permutation testing is displayed in Table 2.
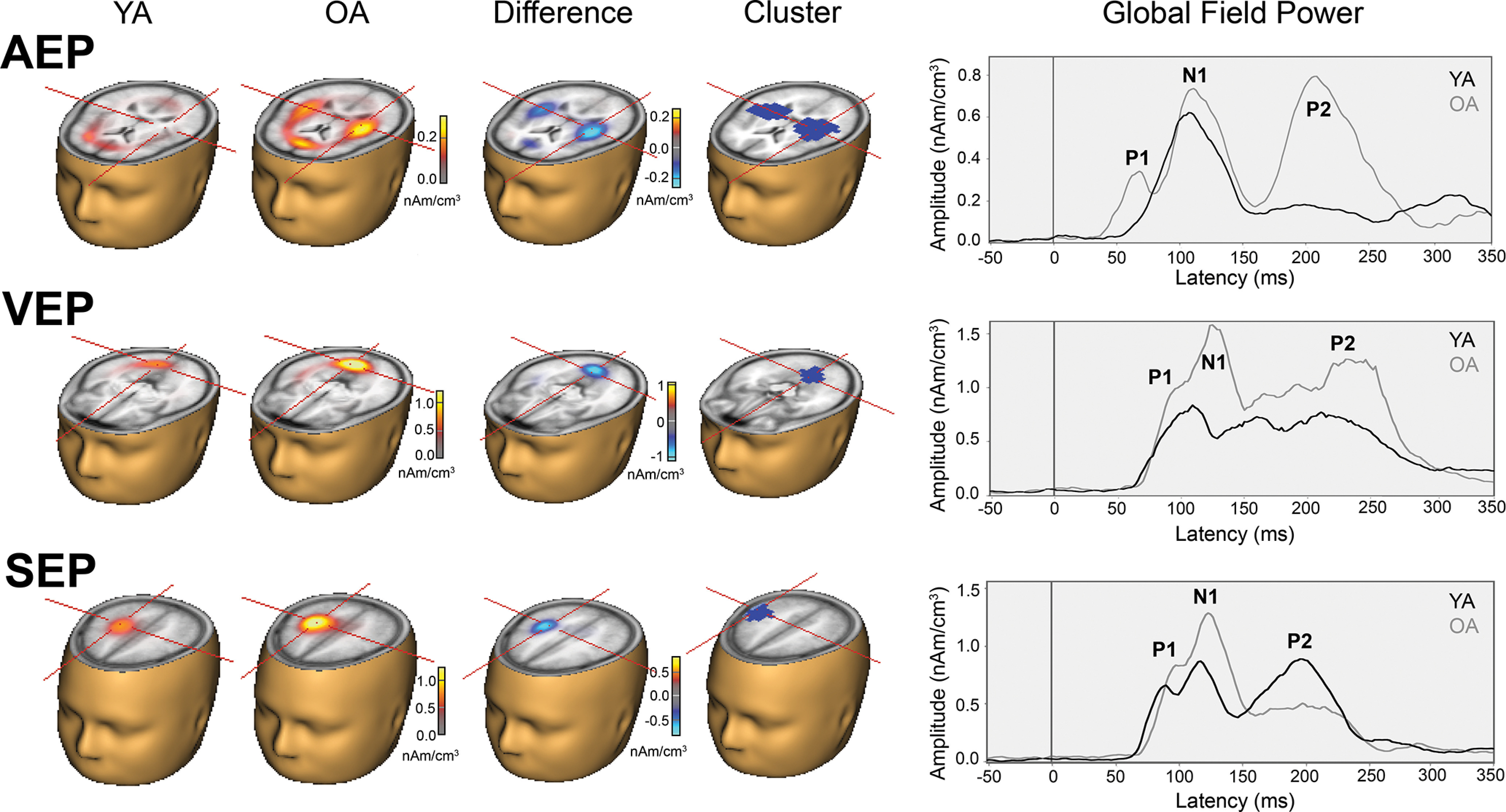
Figure 1.

Group mean AEPs, VEPs, and SEPs in young (YA) and older (OA) adults. Right, the iso-contour maps highlighting the P1, N1, and P2 deflections in all three sensory modalities. FCz = midline frontal central; PO4 = right parietal-occipital; CP4 = right central-parietal.
Table 2.
Summary of cluster-based statistics on sensory-evoked potential waveforms
| Electrode cluster | Time (ms) | Peak latency/electrode | p | |
|---|---|---|---|---|
| AEPs | ||||
| 1 | FP1, AF7, AF3, F1, F3, F5, F7, FC5, FC3, FC1, C1, C3, C5, CP3, CP1, CPz, FPz, FP2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FC6, FC4, FC2, FCz, Cz, C2, C4, C6, CP4, CP2 | 174-338 | 266 ms, C2 | <0.001 |
| 2 | T7, TP7, CP5, P3, P5, P7, P9, PO7, PO3, O1, Iz, Oz, POz, T8, TP8, P6, P8, P10, PO8, PO4, O2, CB1, CB2, TP9, TP10 | 178-350 | 271 ms, Iz | <0.001 |
| 3 | FP1, AF7, AF3, F1, F3, F5, F7, FT7, FC5, FC3, FC1, C1, C3, C5, CP5, CP3, CP1, P1, P3, POz, Pz, CPz, FPz, FP2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FC6, FC4, FC2, FCz, Cz, C2, C4, C6, CP6, CP4, CP2, P2 | 29-100 | 59 ms, C4 | <0.001 |
| 4 | TP7, P5, P7, P9, PO7, PO3, O1, Iz, Oz, POz, TP8, P6, P8, P10, PO8, PO4, O2, CB1, CB2, TP9, TP10 | 0-80 | 57 ms, Iz | 0.001 |
| 5 | FT9, F9, LO1, IO1 | 193-334 | 268 ms, FT9 | 0.032 |
| 6 | FP1, AF7, AF3, F3, F5, F7, FC5, FPz, FP2, AF8, FT9, FT10, F9, F10, LO1, LO2, IO1, IO2 | 43-107 | 59 ms, FT10 | 0.044 |
| 7 | FT10, F10, LO2, IO2 | 201-328 | 267 ms, FT10 | 0.049 |
| VEPs | ||||
| 1 | FP1, AF7, AF3, F1, F3, F5, F7, FT7, FC5, FC3, FC1, C1, C3, C5, T7, TP7, CP5, P1, P3, P5, P7, P9, PO7, PO3, O1, Iz, Oz, POz, CPz, FPz, FP2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FT8, FC6, FC4, FC2, FCz, Cz, C2, C4, C6, T8, CP6, CP4, CP2, P2, P4, P6, P8, PO8, PO4, O2, CB1, TP9, FT10, F9, F10, LO2, IO1, IO2 | 115-350 | 139 ms, P9 | <0.001 |
| 2 | FP1, AF7, F5, F7, FT7, FC5, C5, T7, TP7, CP5, CP3, P1, P3, P5, P7, P9, PO7, PO3, O1, Iz, Oz, POz, Pz, P2, P4, P6, P8, P10, PO8, PO4, O2, CB1, CB2, TP9, TP10, FT9, F9, LO1, IO1 | 166-350 | 293 ms, Oz | <0.001 |
| 3 | FP1, AF3, F1, F3, F5, FC3, FC1, C1, C3, CP1, CPz, FPz, FP2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FT8, FC6, FC4, FC2, FCz, Cz, C2, C4, C6, T8, CP6, CP4, CP2 | 115-170 | 139 ms, C4 | <0.001 |
| 4 | AF3, F1, F3, F5, FC5, FC3, FC1, FPz, FP2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FT8, FC6, FC4, FC2, FCz, C4, C6, T8, TP8, CP6, P10, TP10, FT10, F10, LO2, IO2 | 51-111 | 98 ms, FC6 | 0.003 |
| 5 | TP7, CP5, CP3, CP1, P1, P3, P5, P7, P9, PO7, PO3, O1, Iz, Oz, POz, Pz, P2, P4, PO4, O2, CB1 | 49-109 | 96 ms, POz | 0.006 |
| SEPs | ||||
| 1 | FP1, AF7, AF3, F3, F5, F7, FT7, FC5, C5, T7, TP7, CP5, CP3, P1, P3, P5, P7, P9, PO7, PO3, O1, Oz, POz, Pz, FPz, FP2, AF8, AF4, AFz, F6, F8, FT8, FC6, C4, C6, T8, TP8, CP6, P2, P4, P6, P8, P10, PO8, PO4, O2, CB1, CB2, TP9, TP10, FT9, FT10, F9, F10, LO1, LO2, IO1, IO2 | 160-350 | 205 ms, CP5 | <0.001 |
| 2 | FP1, AF7, AF3, F1, F3, F5, F7, FT7, FC5, FC3, FC1, T7, FPz, FP2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FT8, FC6, FC4, FC2, FCz, C2, C4, C6, T8, TP8, CP6, CP4, CP2, P2, P4, P6, FT9, FT10, F9, F10, LO1, LO2, IO1, IO2 | 0-123 | 80 ms, F2 | <0.001 |
| 3 | F1, FC3, FC1, C1, C3, CP3, CP1, P1, P3, P5, PO7, PO3, O1, Oz, POz, Pz, CPz, Fz, F2, FC4, FC2, FCz, Cz, C2, C4, CP6, CP4, CP2, P2, P4, P6, P8, PO8, PO4, O2 | 250-350 | 345 ms, PO4 | <0.001 |
| 4 | FP1, AF7, AF3, F1, F3, F5, FC3, FC1, FPz, FP2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FC6, FC4, FC2, FCz, Cz, C2 | 166-248 | 212 ms, F2 | <0.001 |
| 5 | AF7, AF3, F3, F5, F7, FC5, C5, T7, TP7, CP5, P5, P7, P9, PO7, PO3, O1, Iz, Oz, POz, Pz, C4, C6, TP8, CP6, CP4, CP2, P2, P4, P6, P8, P10, PO8, PO4, O2, CB1, CB2, TP9, TP10, LO1, IO1 | 66-123 | 80 ms, PO8 | <0.001 |
AEPs
In both groups, lateralized auditory stimuli generated AEPs with large negative (N1) and positive (P2) deflections that peaked at ∼100 and 200 ms after stimulus, respectively, over the frontocentral scalp area. In older adults, the N1 and P2 waves were preceded by an early positive deflection (P1) with a group mean peak latency of ∼60 ms after sound onset (Fig. 1). Cluster-based permutation testing identified significant clusters demonstrating larger amplitude in older than young adults in the time interval that encompassed the P1 and P2 wave. As the P1 wave was virtually absent in young adults, no post hoc ANOVAs were performed for the P1 amplitude or latency. Post hoc ANOVAs for the P2 revealed both an age-related increase in P2 amplitude and a prolongation of the P2 deflection in older adults. The P2 peak latency measured between 150 and 300 ms at five frontocentral electrodes (Fz, FC1, FCz, FC2, and Cz) was significantly longer in older adults (M = 222 ms, SEM = 3 ms) than in young adults (M = 197 ms, SEM = 3 ms) (F(1,70) = 31.25, p < 0.001, ηp2 = 0.308). Additionally, P2 peak amplitudes measured in the same time window and electrode cluster were significantly greater in older adults (M = 2.91 μV, SEM = 0.19 μV) than in young adults (M = 2.06 μV, SEM = 0.18 μV) (F(1,70) = 10.40, p = 0.002, η2p = 0.129). Permutation testing did not reveal a significant difference in the N1 wave between young and older adults.
VEPs
In both young and older adults, VEPs were characterized by positive (P1), negative (N1), and positive (P2) deflections that peaked at ∼85, 150, and 230 ms, respectively, over occipital and parieto-occipital areas (Fig. 1). VEPs were largest over the hemisphere contralateral to the stimulus. Cluster-based permutation testing revealed larger P1, N1, and P2 amplitudes in older than in young adults (Fig. 1; Table 2); therefore, post hoc ANOVAs for both amplitude and latency measures were run for all three deflections. The P1 peak amplitude and peak latency between 50 and 150 ms were quantified at five electrodes over the parietal and occipital region contralateral to the stimulus presentation (P4, POz, PO4, PO8, and O2). As expected, older adults showed significantly greater P1 amplitudes (M = 3.26 μV, SEM = 0.30 μV) than young adults (M = 1.61 μV, SEM = 0.29 μV) (F(1,70) = 15.18, p < 0.001, η2p = 0.065), but the groups did not significantly differ in P1 latencies (F(1,70) = 0.99, p = 0.323, ηp2 = 0.014). As for the visual N1 deflection, peak amplitude and peak latency between 125 and 225 ms were quantified over the same five occipital and parieto-occipital electrodes. The ANOVA revealed greater N1 amplitudes in older (M = −2.83 μV, SEM = 0.35 μV) than young adults (M = −0.99 μV, SEM = 0.34 μV) (F(1,70) = 14.41, p < 0.001, η2p = 0.171), but the groups did not significantly differ in N1 latencies (F(1,70) = 0.03, p = 0.870, η2p < 0.001). Like the AEPs, the post hoc ANOVA for the VEP P2 revealed an age-related increase in amplitude and in latency. The P2 peak amplitude and peak latency between 190 and 320 ms was quantified over the same cluster of electrodes. Older adults showed significantly larger P2 amplitudes (M = 3.10 μV, SEM = 0.27 μV) than young adults (M = 2.30 μV, SEM = 0.26 μV) (F(1,70) = 4.84, p = 0.038, η2p = 0.060). Older adults also showed significantly longer P2 latencies (M = 268 ms, SEM = 5 ms) than young adults (M = 245 ms, SEM = 5 ms) (F(1,70) = 10.65, p = 0.002, η2p = 0.132).
SEPs
In both groups, SEPs comprised a positive deflection peaking at ∼90 ms after stimulus onset at central-parietal and parietal sites over the hemisphere contralateral to the stimulated hand. This deflection was inverted in polarity at midline right frontal scalp sites, consistent with an anterior, posterior tangential source located at the somatosensory cortex (Fig. 1). The P1 at central-parietal and parietal sites was followed by a negative deflection peaking at ∼130 ms (N1) and a positive wave peaking at ∼250 ms after stimulus onset (P2). Cluster-based permutation testing showed larger amplitude and delayed P1 latency in older adults than in young adults. The P1 peak latency and amplitude measured between 50 and 125 ms at five electrodes centered around the mid central-parietal sites (CP2, C4, CP4, P4, and CP6) revealed significantly longer P1 latencies in older adults (M = 98 ms, SEM = 1 ms) than in young adults (M = 90 ms, SEM = 1 ms) (F(1,70) = 26.70, p < 0.001, η2p = 0.276). The P1 peak amplitude was also larger in older (M = 2.39 μV, SEM = 0.18 μV) than in young adults (M = 1.76 μV, SEM = 0.17 μV) (F(1,70) = 6.63, p = 0.012, η2p = 0.087). The analysis also revealed significant clusters (Clusters 1, 3, and 4) that encompassed the P2 interval. Therefore, the P2 peak latency and amplitude were measured between 190 and 300 ms at the same electrode cluster. However, the post hoc ANOVA did not find a significant group main effect in P2 amplitude (F(1,70) = 0.84, p = 0.362, η2p = 0.012) or in P2 latency (F(1,70) = 2.56, p = 0.114, η2p = 0.035). No significant clusters were found to indicate group differences in the somatosensory N1.
Summary of sensory-evoked potentials
In summary, we found age-related differences in the amplitude of early scalp-recorded evoked potentials in all three sensory modalities (i.e., auditory P1, visual P1, and somatosensory P1). Older adults showed larger amplitude in all three sensory domains than did young adults. Older adults also showed longer response latencies in all three sensory domains than young adults (i.e., auditory P2, visual P2, and somatosensory P1).
Next, we determined whether these age-related differences in scalp-recorded ERPs were related to source activity from sensory cortices. To do so, we modeled the scalp-recorded responses generated by auditory, visual, and somatosensory stimuli.
Brain source activity
The scalp-recorded evoked potentials reflect the aggregation of neural responses generated in distributed cortical sources. To determine whether the age-related increase in early sensory-evoked potentials reflects increased excitability in sensory cortices, we used distributed source modeling (i.e., CLARA) to localize generators of scalp-recorded evoked potentials for each sensory modality at each time point per participant. Then, we compared the source strength at each time point from stimulus onset to 300 ms after stimulus using cluster-based permutation testing of distributed source space data. A summary of significant clusters revealed by cluster-based permutation testing is displayed in Table 3.
Table 3.
Summary of cluster-based permutation statistics on source activitya
| Cluster | p | YA mean source strength | OA mean source strength | Window (ms) | Peak latency (ms) | BA | x | y | z |
|---|---|---|---|---|---|---|---|---|---|
| Auditory | |||||||||
| 1 | <0.001 | 0.081 | 0.257 | 175-265 | 209 | 48 | 32 | −17 | 10 |
| 2 | <0.001 | 0.125 | 0.020 | 142-228 | 197 | 7 | −11 | −45 | 52 |
| 3 | 0.001 | 0.052 | 0.207 | 41-84 | 55 | 13 | −32 | −24 | 10 |
| 4 | 0.003 | 0.149 | 0.028 | 277-300 | 292 | NA | 18 | −3 | 17 |
| 5 | 0.046 | 0.113 | 0.276 | 90-101 | 94 | 48 | −46 | 4 | −4 |
| Visual | |||||||||
| 1 | <0.001 | 0.099 | 0.304 | 168-300 | 199 | 20 | 32 | −17 | −11 |
| 2 | 0.001 | 0.133 | 0.015 | 205-267 | 212 | 48 | −32 | 4 | 10 |
| 3 | 0.003 | 0.264 | 0.072 | 134-162 | 148 | 19 | −18 | −59 | −4 |
| 4 | 0.012 | 0.196 | 0.594 | 117-133 | 123 | NA | −4 | −59 | −11 |
| 5 | 0.026 | 0.299 | 0.078 | 150-170 | 158 | 20 | 25 | −17 | −18 |
| 6 | 0.037 | 0.053 | 0.176 | 285-300 | 298 | 11 | 18 | 25 | −4 |
| 7 | 0.050 | 0.150 | 0.391 | 82-107 | 94 | NA | 17 | −59 | 10 |
| Somatosensory | |||||||||
| 1 | <0.001 | 0.340 | 0.092 | 162-300 | 195 | 6 | 32 | −10 | 38 |
| 2 | 0.001 | 0.177 | 0.063 | 70-99 | 86 | 32 | 4 | 11 | 38 |
| 3 | 0.003 | 0.027 | 0.117 | 191-240 | 214 | 7 | 32 | −66 | 31 |
| 4 | 0.026 | 0.147 | 0.405 | 117-140 | 125 | 3 | 53 | −24 | 45 |
| 5 | 0.030 | 0.026 | 0.124 | 189-214 | 193 | 11 | 25 | 32 | −4 |
| 6 | 0.035 | 0.019 | 0.078 | 72-82 | 76 | 38 | 46 | 11 | −4 |
aYA, Young adults; OA, older adults; BA, Brodmann area; coordinates in Talairach space.
For the auditory modality, the between-group contrast yielded greater source activity in bilateral auditory cortices along the superior temporal gyrus during the two intervals that encompass the P1 (Cluster 3) and P2 (Cluster 1) deflections (Fig. 2). Young adults showed stronger source activity in the mid parietal region (Cluster 2) between 142 and 228 ms and right insula areas (Cluster 4) between 275 and 300 ms after stimulus.
Figure 2.
Group mean distributed source activity for AEPs, VEPs, and SEPs in young (YA) and older (OA) adults. Right, the strength of source activity over time.
Compared with young adults, older adults showed enhanced source activity in visual cortices between 80 and 130 ms after stimulus (Cluster 4 and 7). They also showed enhanced source activity in the right anterior and medial temporal region (Cluster 1). We observed, however, weaker source activity in older adults in the visual areas contralateral to the stimulus (Cluster 3) between 135 and 160 ms after stimulus. Young adults also showed stronger activity in the mid and superior PFC than older adults (Clusters 2 and 5).
For the somatosensory modality, older adults also showed enhanced source activity in somatosensory cortices between 117 and 140 ms after stimulus (Cluster 4). Additionally, older adults showed greater source activity than young adults in the right parietal (Cluster 6) and right inferior frontal areas (Cluster 5). We also observed greater source activity in young than older adults in the right somatosensory cortex, peaking at 195 ms after stimulus onset (Cluster 1).
In summary, older adults showed enhanced source activity in sensory areas compared with young adults. The time course of source activity encompassed the early auditory-, visual-, and somatosensory-evoked responses (i.e., 40-140 ms). We also observed weaker source activity in older adults, primarily in supramodal areas, such as parietal and prefrontal cortices.
Functional connectivity
We tested whether age differences in functional connectivity contribute to age differences in sensory-evoked responses by comparing PLV between discrete brain sources in sensory cortices and between parietal and prefrontal cortices. The summary of cluster-based permutation test statistics is presented in Table 4. The analysis of PLV revealed stronger synchrony in older than in young adults between the mid PFC and right auditory and visual cortices (Fig. 3). The analysis of PLV for the somatosensory stimuli trended toward significance, with weaker synchrony between the mid PFC and the right somatosensory cortex. For visual stimuli, older adults also showed stronger PLV between the right and left visual cortices, between the right visual cortex and the mid parietal area, and between the right visual cortex and the right anterior temporal cortex.
Table 4.
Summary of cluster-based permutation statistics on functional connectivity of sensory-evoked responsesa
| Cluster | p | YA Mean PLV | OA mean PLV | Window (ms) | Frequency range (Hz) | Latency at maximum (ms) | Frequency at maximum (Hz) | Source at maximum |
|---|---|---|---|---|---|---|---|---|
| Auditory | ||||||||
| 1 | 0.017 | 0.028 | 0.087 | 150-300 | 3-5 | 200 | 4 | FR |
| 2 | 0.018 | 0.047 | 0.111 | 100-300 | 3-5 | 250 | 4 | FpM |
| 3 | 0.030 | 0.048 | 0.113 | 100-250 | 2-6 | 200 | 4 | FL |
| 4 | 0.048 | 0.020 | 0.077 | 200-300 | 3-5 | 250 | 4 | FM |
| Visual | ||||||||
| 1 | <0.001 | 0.061 | 0.206 | 100-300 | 2-6 | 250 | 4 | VClR |
| 2 | <0.001 | 0.040 | 0.131 | 0-300 | 2-6 | 300 | 4 | VClL |
| 3 | 0.001 | 0.043 | 0.132 | 100-300 | 3-7 | 250 | 5 | VCbR |
| 4 | 0.006 | −0.027 | 0.013 | 100-300 | 10-14 | 150 | 11 | FM |
| 5 | 0.014 | −0.050 | −0.002 | 100-300 | 10-14 | 250 | 12 | TAR |
| 6 | 0.016 | 0.060 | 0.156 | 150-300 | 3-6 | 300 | 5 | PM |
| 7 | 0.020 | 0.020 | 0.072 | 50-300 | 3-5 | 100 | 5 | TAR |
| Somatosensory | ||||||||
| 1 | 0.108 | 0.037 | −0.001 | 100-300 | 10-11 | 300 | 10 | FM |
aYA, Young adults; OA, older adults; FR, frontal right; FpM, fronto-polar middle; FL, frontal left; FM, frontal middle; VClR, visual cortex lateral right; VClL, visual cortex lateral left; VCbR, visual cortex basal right; TAR, temporal anterior right; PM, parietal middle.
Figure 3.
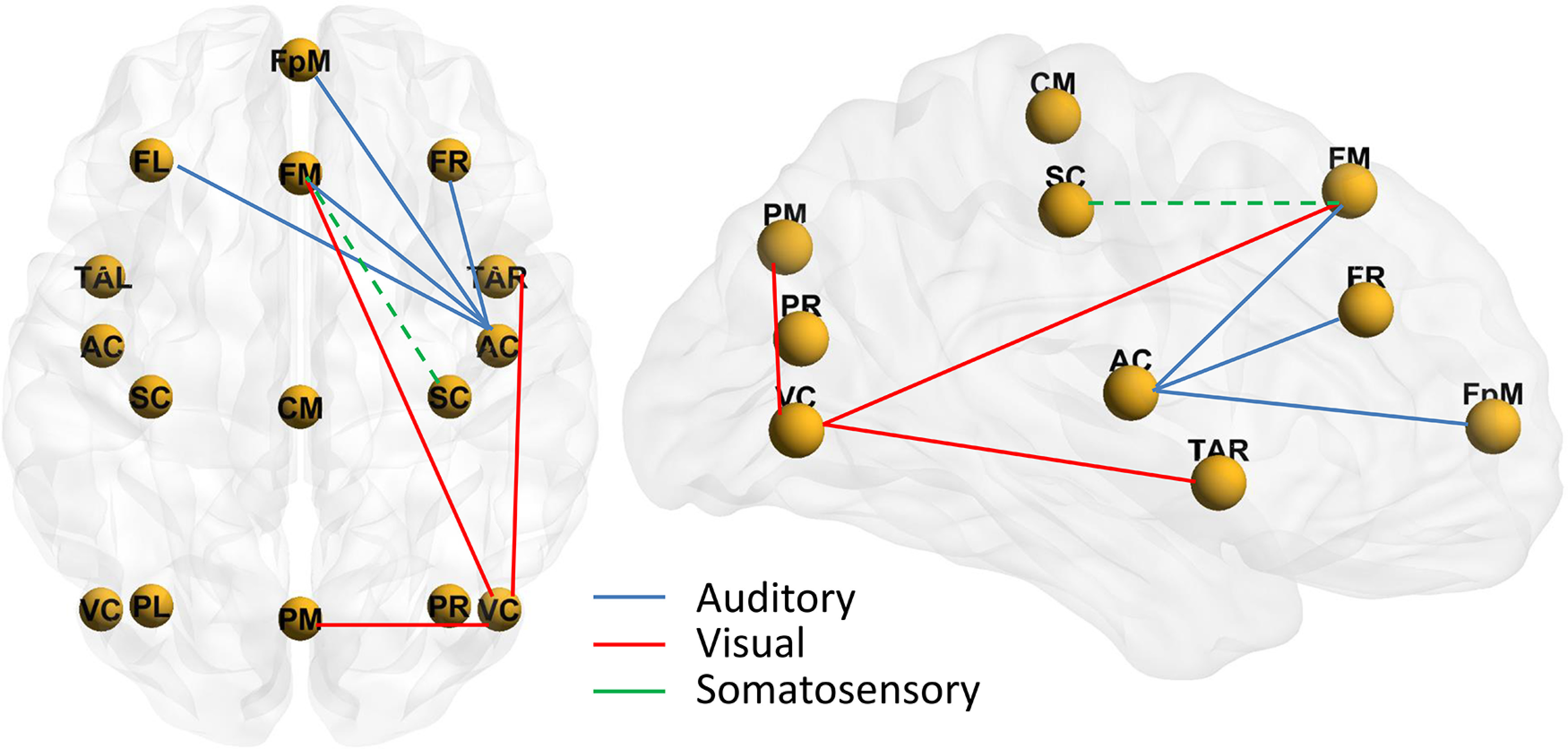
Schematic showing the functional connectivity between the sensory cortices and the prefrontal cortices. Color lines indicate age differences in PLVs between the nodes. Solid lines indicate stronger PLV in older than young adults. Dashed lines indicate weaker PLV in older than in young adults. AC, Auditory cortex; VC, visual cortex; SC, somatosensory cortex; TAR, temporal anterior right; PL, parietal left; PM, parietal midline; PR, parietal right; CM, central midline; FL, frontal left; FM, frontal midline; FR, frontal right; FpM, fronto-polar midline.
Correlation analyses
Given the age-related increase in early sensory-evoked response amplitude (i.e., P1) in all three modalities, we conducted correlation analyses to test the premise that age-related changes observed in one sensory modality would be related to changes in other sensory modalities. In older adults, the scalp-recorded auditory P1 amplitude was significantly correlated with the visual P1 amplitude (r = 0.644, p < 0.001), such that larger auditory P1 amplitudes were associated with larger visual P1 amplitudes. The auditory P1 was also positively correlated with the somatosensory P1 amplitude (r = 0.434, p = 0.009). The correlation between visual P1 and somatosensory P1 amplitude was also significant (r = 0.549, p = 0.001). For latency measures, there were no significant correlations between the three sensory modalities (p > 0.16 in all cases). In young adults, there was a significant negative correlation between the auditory P1 amplitude and visual P1 amplitude (r = –0.342, p = 0.039). No other correlations between P1 amplitude and latency among sensory systems were found to be significant in young adults.
We also examined whether source strength in sensory areas correlated with one another (Fig. 4). For each participant, we used the mean source activity that encompassed the P1 deflection from each sensory domain (i.e., Cluster 3 for AEP, Cluster 7 for VEP, and Cluster 4 for SEP). Older adults who showed enhanced source activity in bilateral auditory cortices also showed enhanced source activity in visual and somatosensory cortices (r = 0.572 and r = 0.342, respectively, p < 0.05 in both cases). Similarly, older adults who showed enhanced source activity in the visual cortex also showed enhanced activity in somatosensory areas (r = 0.635, p < 0.001). By contrast, in young adults, there was no significant correlation between the source strength in auditory, visual, or somatosensory cortices (r < 0.163, p > 0.15 in all cases).
Figure 4.
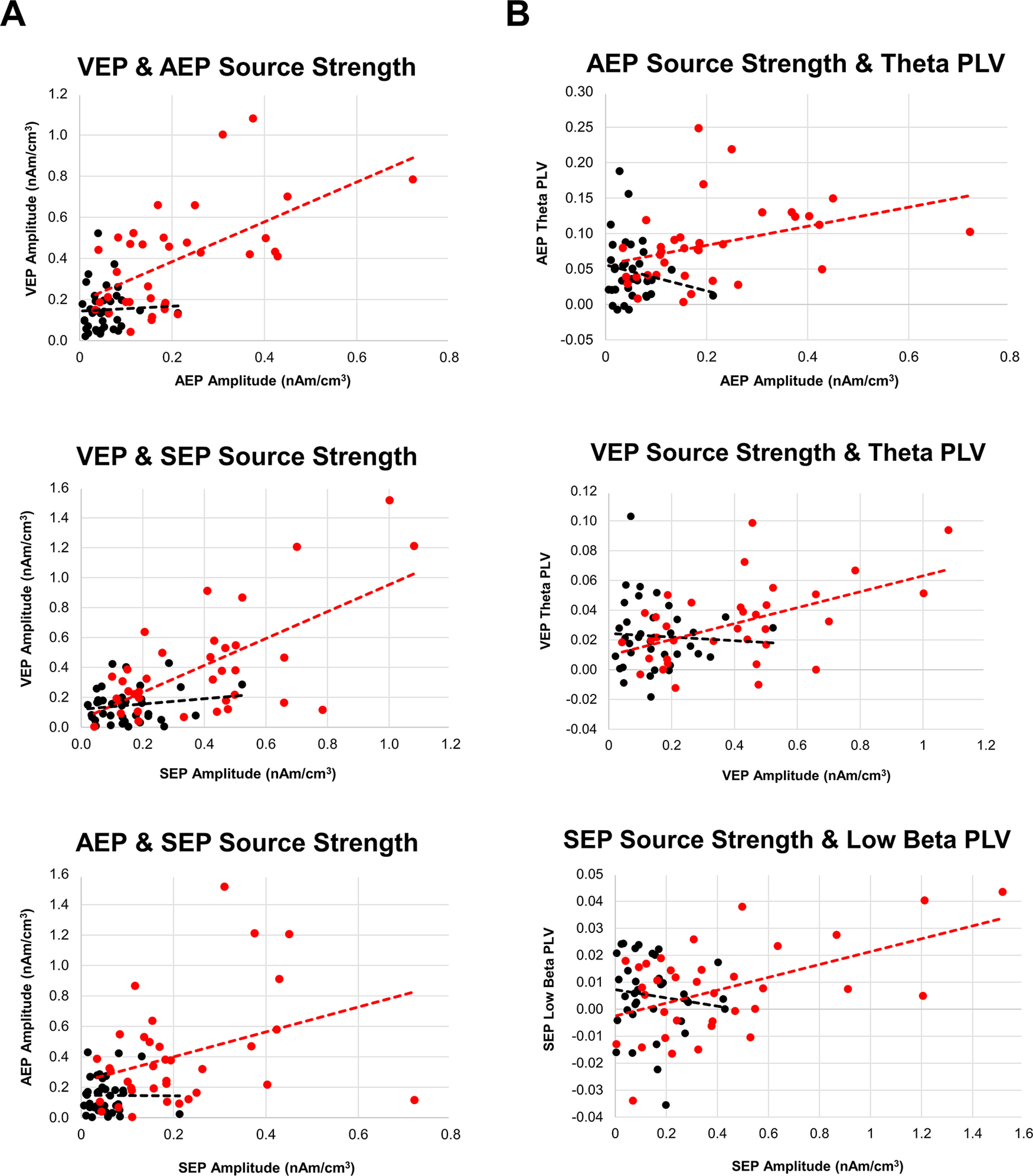
A, Scatterplots showing the relationship between the source strength activity from auditory, visual, and somatosensory cortices. Measures derived from the statistically significant cluster from permutation statistics of CLARA source activity that represented the sensory P1 from each domain: Cluster 3 for AEP, Cluster 7 for VEP, and Cluster 4 for SEP. B, Scatterplots showing the relationship between prefrontal-to-sensory cortex synchrony and source activity. Measures derived from the statistically significant cluster from permutation statistics of CLARA imaging that represented the sensory P1 from each domain: Cluster 3 for AEP, Cluster 7 for VEP, and Cluster 4 for SEP. Some PLV values are negative. This is because of the relative contrast used (stimulus-locked minus non–stimulus-locked epochs) to estimate neural synchrony between brain sources in young and older adults.
Last, we examined whether the degree of phase synchronization between the PFC and sensory cortices was related to the magnitude of source activity. For each sensory modality, we averaged PLV from the left, mid-central, and right PFC for the 50-250 ms interval, and then conducted correlation analyses for the theta, α, and β bands. The correlational analyses were limited to lower and mid-range frequency bands because these were showing age differences in PLV between sensory cortices and PFC. In older adults, the strength of auditory source activity (Fig. 2) was positively correlated with theta PLV (r = 0.361, p = 0.033), such that higher synchrony was associated with stronger source activity (Fig. 4). A similar pattern was observed for activity in the visual cortex, with greater theta PLV being associated with stronger source activity (r = 0.506, p = 0.002). Last, the strength of activity in somatosensory cortex was positively correlated with low β PLV (r = 0.501, p = 0.002), such that increased synchrony in the low beta band was associated with enhanced source activity. In young adults, source activity in auditory, visual, and somatosensory cortex was not related to the degree of synchrony between the PFC and sensory cortex. Together, the findings from these correlational analyses suggest that age differences in sensory-evoked responses amplitude may be linked to ineffective top-down regulation from supramodal prefrontal executive regions.
A Fisher's z statistic was used to determine whether the correlations observed in older adults differed from those of the young adults. For the P1 amplitude, older adults exhibited significantly stronger correlations than young adults between auditory and visual modalities (z = −4.553, p < 0.001), and the auditory and somatosensory modalities (z = −1.968, p = 0.025). The contrast between the visual and somatosensory modality trended toward significance (z = −1.618, p = 0.053). For the source strength in sensory cortices, older adults also showed significantly stronger correlations than young adults between auditory and visual cortices (z = −2.462, p = 0.007) and between the visual and somatosensory cortices (z = −2376, p = 0.009). The contrast between the auditory and somatosensory modality trended toward significance (z = −1.475, p = 0.070). For the relation between prefrontal-to-sensory cortex synchrony and sensory source activity, older adults showed significantly stronger correlations than young adults for auditory (z = 2.240, p = 0.013), visual (z = 2.495, p = 0.006), and somatosensory stimulation (z = 2.758, p = 0.003).
Discussion
The present study is the first to show age-related differences in early sensory-evoked responses in terms of latency, amplitude, source activity, and functional connectivity across three sensory domains: auditory, visual, and somatosensory modalities. The age-related increase in AEP, VEP, and SEP amplitude coincided in latency with an age-related enhancement of source activity in the corresponding sensory cortices. We observed similar age differences in all three sensory modalities in terms of functional connectivity, with greater phase synchrony between prefrontal and sensory cortices in older than young adults in middle-high frequencies. Notably, in older adults, the sensory-evoked response amplitude measures were correlated between sensory domains. Older adults showing larger response amplitude in one modality also showed larger amplitudes in other modalities. Likewise, measures of source strength were correlated between sensory modalities. In older adults, the degree of phase synchronization between prefrontal and sensory cortices was related to the magnitude of source activity in sensory areas. To summarize, the analyses revealed clear age-related changes in auditory-, visual-, and somatosensory-evoked responses, which may be mitigated by age-related changes in prefrontal-to-sensory cortex synchrony.
Age-related changes in sensory-evoked response latency were not homogeneous across modalities. That is, older adults showed prolonged P2 latency following auditory and visual stimuli, whereas somatosensory stimuli generated longer P1 latency in older than in young adults. The latency of evoked responses from sensory cortices is thought to index both speed of nerve conduction and integration of sensory information. In older adults, the increase P1 latency elicited by somatosensory stimuli is consistent with prior studies (e.g., Bolton and Staines, 2012; Strommer et al., 2017), and could indicate prolongation of peripheral and central nerve conduction. The age difference in auditory and visual P2 latency is also consistent with prior studies (e.g., Amenedo and Diaz, 1998; Bertoli et al., 2005; Alain et al., 2012; Fernandez et al., 2013), and could indicate a longer time for older adults to process and encode sensory information into sensory memory.
Age-related increases in sensory-evoked response amplitude were consistent across all three sensory modalities. The larger amplitude of stimulus-evoked responses may represent an adaptive neural response to healthy aging. Compared with young adults, older adults may recruit more or engage different neural resources to increase stimulus detection and sensory processing efficiency. The age-related increase in sensory-evoked responses may also be related to reduced sensory adaptation, with young adults showing a stronger repetition effect (adaptation) than older adults (e.g., Walrath and Hallman, 1984; Leung et al., 2013). Our findings are consistent with the common cause hypothesis, in which age-related increases in early cortical evoked responses may reflect changes in sensory regulation mediated by the PFC (Knight et al., 1999).
Evidence from behavioral studies (Stuss, 2006; Picton et al., 2007), fMRI studies (Gazzaley and D'Esposito, 2007; Amer et al., 2016), and other electrophysiological studies (Bolton and Staines, 2014; Bidet-Caulet et al., 2015) suggests that the PFC plays an important role in attentional control and in regulating neural activity in sensory areas. The PFC may facilitate and/or suppress sensory processing via thalamocortical connections to the thalamic reticular nucleus (Scannell et al., 1999; Zikopoulos and Barbas, 2006; Nakajima et al., 2019), which has extensive connections with the medial geniculate body, lateral geniculate nucleus, and ventral-posterior medial thalamic nucleus (i.e., the bottleneck of afferent input to auditory, visual, and somatosensory cortices, respectively). Prefrontal modulations of sensory processing may also occur via corticocortical connections to primary sensory cortices via associative sensory areas (Fuster, 2001). Anatomical tracing and neurophysiological studies also provide evidence for dense corticocortical connections between sensory and ventrolateral and dorsolateral prefrontal cortices (Diehl and Romanski, 2014; Plakke and Romanski, 2014, 2016). Furthermore, healthy aging is also associated with decreased gray matter density, which is more pronounced in prefrontal areas than in sensory areas (Raz et al., 1997). Notably, white and gray matter changes associated with age have been related to increases in auditory and visual evoked response latency (Price et al., 2017). Together, these findings support the PFC's role in downregulating sensory input, which may account for a shared factor common to the age-related changes in sensory-evoked responses observed in the present study.
Evidence also suggests a link between sensory-evoked response amplitude, particularly with prefrontal executive control mechanisms as measured with neuropsychological tests. For instance, a higher percentage of perseverative errors on the Wisconsin Card Sorting Test (WCST) can indicate worse cognitive flexibility, a subtype of executive functions mediated by prefrontal regions (Gruber et al., 2010; Kim et al., 2011). Chao and Knight (1997) demonstrated that older adults who showed larger amplitude in early AEPs also showed a greater percentage of WCST preservative errors. The PFC's putative role in regulating sensory-evoked response amplitude is further supported by research using noninvasive brain stimulation. For instance, transcranial magnetic stimulation to the dorsolateral PFC has been shown to modulate the amplitude of VEPs and SEPs (Adams et al., 2019). Transcranial direct current stimulation over the dorsolateral PFC also modulates the auditory P1 amplitude (Terada et al., 2015). Our findings are also consistent with studies showing that stroke-related prefrontal injury is often associated with increased amplitude and longer latencies for AEPs (Woods and Clayworth, 1986; Knight et al., 1989) and SEPs (Yamaguchi and Knight, 1990). This is in stark contrast with injury localized to temporal, parietal, or occipital regions, which are usually associated with decreased sensory-evoked response amplitude (Knight et al., 1980; Kileny et al., 1987; Woods et al., 1987; Alain et al., 1998; Haigh et al., 2018; Mammadkhanli et al., 2020).
Our functional connectivity analysis revealed age-related differences in neural synchrony (i.e., greater PLV in older than young adults) between the superior PFC and the auditory and visual cortices. The present study is the first to our knowledge to investigate age-related changes in three sensory domains with a metric of functional connectivity. These age differences in phase coherence observed in the present study may reflect age differences in the allocation of attention, with older adults' potentially allocating more attention to the sensory stimuli than young adults. Indeed, prior research has found that young adults may exhibit greater mind-wandering than older adults on cognitive tasks, whereas older adults may have greater focused attention on task stimuli (Maillet et al., 2018, 2020). Future research may examine how the variability of attentional states may modulate sensory-evoked responses during passive sensory stimulation.
In the present study, we show in older adults a link between sensory-evoked responses elicited by auditory, visual, and tactile stimuli across a range of analyses. Although these results provide support for the common-cause hypothesis, some limitations should be kept in mind while interpreting and generalizing findings. For instance, in the present study, there was a small group difference in the number of years of education; and while small differences in education may appear unlikely to account for group differences in bottom-up passive sensory-evoked responses, the possibility should be ruled out in future studies. The neuropsychological assessment was administered only for older adults, making it difficult to compare the cognitive profiles of young and older adults and to assess how this relates to sensory-evoked response amplitude and latency. Future studies should also include structural MRI data to further enhance the precision of the source and functional connectivity analyses. The present study did not include behavioral measures of sensory functions, making it difficult to determine whether young and older adults did similarly experience the stimuli. For instance, our sensory stimuli may have appeared more salient to older than young older adults, which could have contributed to enhancing sensory-evoked response. Consequently, in future studies, it may be important to include questionnaires to evaluate the participants' subjective experience during data collection to ensure that both young and older adults are comparable across this variable. Future research should also consider additional measures of sensory acuity that could be used as moderator variables.
In conclusion, the present study shows that healthy aging is associated with increased amplitude, latency, and source activity of early sensory-evoked responses that appear to be shared in all three sensory modalities. Within older adults, these age-related increases in early sensory processing were found correlated between modalities. With our functional connectivity measures, we also demonstrate age-related differences in phase synchrony between prefrontal regions and sensory cortices, suggesting that age-related changes in prefrontal functioning may account for deficits in sensory inhibition in healthy older adults.
Footnotes
This work was supported by Natural Sciences and Engineering Research Council of Canada Grant RGPIN-2016-05523, Alzheimer Society of Canada, and Lorraine Johnson Foundation to C.A.; National Natural Science Foundation of China 31600798 and 61761166001, and the 111 Project from Ministry of Education of China (B12027) to D.Y. R.R. was supported by Natural Sciences and Engineering Research Council postdoctoral fellowship and Alzheimer Society of Canada. M.F. was supported by Saul A. Silverman Family Foundation as a Canada International Scientific Exchange Program, and Morris Kerzner Memorial Fund. We thank Elizabeth Baker-Sullivan, Haley Gabel, Farnosh Hakimy, Zakia Hussain, Brooke Koblinsky, Cherie Lin, Shahier Paracha, Karishma Ramdeo, Brahm Sanger, Serah Seo, Mariam Sidrak, Parinaz Soghrati, Parnian Tajbakhsh, and Kevin Tang for assistance in participant recruitment and data preparation; and all participants for involvement in this study.
The authors declare no competing financial interests.
References
- Adams MS, Andrew D, Staines WR (2019) The contribution of the prefrontal cortex to relevancy-based gating of visual and tactile stimuli. Exp Brain Res 237:2747–2759. 10.1007/s00221-019-05633-9 [DOI] [PubMed] [Google Scholar]
- Alain C, Woods DL, Knight RT (1998) A distributed cortical network for auditory sensory memory in humans. Brain Res 812:23–37. 10.1016/s0006-8993(98)00851-8 [DOI] [PubMed] [Google Scholar]
- Alain C, McDonald K, Van Roon P (2012) Effects of age and background noise on processing a mistuned harmonic in an otherwise periodic complex sound. Hear Res 283:126–135. 10.1016/j.heares.2011.10.007 [DOI] [PubMed] [Google Scholar]
- Allison T, Hume AL, Wood CC, Goff WR (1984) Developmental and aging changes in somatosensory, auditory and visual evoked potentials. Electroencephalogr Clin Neurophysiol 58:14–24. 10.1016/0013-4694(84)90196-2 [DOI] [PubMed] [Google Scholar]
- Amenedo E, Diaz F (1998) Aging-related changes in processing of non-target and target stimuli during an auditory oddball task. Biol Psychol 48:235–267. 10.1016/s0301-0511(98)00040-4 [DOI] [PubMed] [Google Scholar]
- Amer T, Anderson JA, Campbell KL, Hasher L, Grady CL (2016) Age differences in the neural correlates of distraction regulation: a network interaction approach. Neuroimage 139:231–239. 10.1016/j.neuroimage.2016.06.036 [DOI] [PubMed] [Google Scholar]
- Assem M, Shashidhara S, Glasser MF, Duncan J (2021) Precise topology of adjacent domain-general and sensory-biased regions in the human brain. bioRxiv. Advance online publication. Retrieved September 05, 2021. 10.1101/2021.02.21.431622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beniczky S, Rosenzweig I, Scherg M, Jordanov T, Lanfer B, Lantz G, Larsson PG (2016) Ictal EEG source imaging in presurgical evaluation: high agreement between analysis methods. Seizure 43:1–5. 10.1016/j.seizure.2016.09.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertoli S, Smurzynski J, Probst R (2005) Effects of age, age-related hearing loss, and contralateral cafeteria noise on the discrimination of small frequency changes: psychoacoustic and electrophysiological measures. J Assoc Res Otolaryngol 6:207–222. 10.1007/s10162-005-5029-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bidet-Caulet A, Buchanan KG, Viswanath H, Black J, Scabini D, Bonnet-Brilhault F, Knight RT (2015) Impaired facilitatory mechanisms of auditory attention after damage of the lateral prefrontal cortex. Cereb Cortex 25:4126–4134. 10.1093/cercor/bhu131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolton DA, Staines WR (2012) Age-related loss in attention-based modulation of tactile stimuli at early stages of somatosensory processing. Neuropsychologia 50:1502–1513. 10.1016/j.neuropsychologia.2012.03.002 [DOI] [PubMed] [Google Scholar]
- Bolton DA, Staines WR (2014) Attention-based modulation of tactile stimuli: a comparison between prefrontal lesion patients and healthy age-matched controls. Neuropsychologia 57:101–111. 10.1016/j.neuropsychologia.2014.03.003 [DOI] [PubMed] [Google Scholar]
- Cavazzana A, Rohrborn A, Garthus-Niegel S, Larsson M, Hummel T, Croy I (2018) Sensory-specific impairment among older people: an investigation using both sensory thresholds and subjective measures across the five senses. PLoS One 13:e0202969. 10.1371/journal.pone.0202969 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceponiene R, Westerfield M, Torki M, Townsend J (2008) Modality-specificity of sensory aging in vision and audition: evidence from event-related potentials. Brain Res 1215:53–68. 10.1016/j.brainres.2008.02.010 [DOI] [PubMed] [Google Scholar]
- Chao LL, Knight RT (1997) Prefrontal deficits in attention and inhibitory control with aging. Cereb Cortex 7:63–69. 10.1093/cercor/7.1.63 [DOI] [PubMed] [Google Scholar]
- Cohen SN, Potvin A, Syndulko K, Pettler-Jennings P, Potvin JH, Tourtellotte WW (1982) Multimodality evoked potentials: clinical applications and assessment of utility. Bull Los Angeles Neurol Soc 47:55–61. [PubMed] [Google Scholar]
- Curran T, Hills A, Patterson MB, Strauss ME (2001) Effects of aging on visuospatial attention: an ERP study. Neuropsychologia 39:288–301. 10.1016/s0028-3932(00)00112-3 [DOI] [PubMed] [Google Scholar]
- de la Fuente J, Moreno-Agostino D, de la Torre-Luque A, Prina AM, Haro JM, Caballero FF, Ayuso-Mateos JL (2020) Development of a combined sensory-cognitive measure based on the common cause hypothesis: heterogeneous trajectories and associated risk factors. Gerontologist 60:e357–e366. 10.1093/geront/gnz066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diehl MM, Romanski LM (2014) Responses of prefrontal multisensory neurons to mismatching faces and vocalizations. J Neurosci 34:11233–11243. 10.1523/JNEUROSCI.5168-13.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dimitrijevic A, Pratt H, Starr A (2013) Auditory cortical activity in normal hearing subjects to consonant vowels presented in quiet and in noise. Clin Neurophysiol 124:1204–1215. 10.1016/j.clinph.2012.11.014 [DOI] [PubMed] [Google Scholar]
- Fernandez R, Monacelli A, Duffy CJ (2013) Visual motion event related potentials distinguish aging and Alzheimer's disease. J Alzheimers Dis 36:177–183. 10.3233/JAD-122053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuster JM (2001) The prefrontal cortex—an update: time is of the essence. Neuron 30:319–333. 10.1016/s0896-6273(01)00285-9 [DOI] [PubMed] [Google Scholar]
- Gadkaree SK, Sun DQ, Li C, Lin FR, Ferrucci L, Simonsick EM, Agrawal Y (2016) Does sensory function decline independently or concomitantly with age? Data from the Baltimore Longitudinal Study of Aging. J Aging Res 2016:1865038. 10.1155/2016/1865038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gazzaley A, D'Esposito M (2007) Top-down modulation and normal aging. Ann NY Acad Sci 1097:67–83. 10.1196/annals.1379.010 [DOI] [PubMed] [Google Scholar]
- Gruber AJ, Calhoon GG, Shusterman I, Schoenbaum G, Roesch MR, O'Donnell P (2010) More is less: a disinhibited prefrontal cortex impairs cognitive flexibility. J Neurosci 30:17102–17110. 10.1523/JNEUROSCI.4623-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haigh SM, Robinson AK, Grover P, Behrmann M (2018) Differentiation of types of visual agnosia using EEG. Vision (Basel) 2:44. 10.3390/vision2040044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humes LE, Busey TA, Craig J, Kewley-Port D (2013) Are age-related changes in cognitive function driven by age-related changes in sensory processing? Atten Percept Psychophys 75:508–524. 10.3758/s13414-012-0406-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iodice R, Carotenuto A, Dubbioso R, Cerillo I, Santoro L, Manganelli F (2016) Multimodal evoked potentials follow up in multiple sclerosis patients under fingolimod therapy. J Neurol Sci 365:143–146. 10.1016/j.jns.2016.04.026 [DOI] [PubMed] [Google Scholar]
- Kievit RA, Davis SW, Mitchell DJ, Taylor JR, Duncan J, Cam C, Henson RN, Cam C, Cam-CAN Research Team (2014) Distinct aspects of frontal lobe structure mediate age-related differences in fluid intelligence and multitasking. Nat Commun 5:5658. 10.1038/ncomms6658 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kileny P, Paccioretti D, Wilson AF (1987) Effects of cortical lesions on middle-latency auditory-evoked responses (MLR). Electroencephalogr Clin Neurophysiol 66:108–120. 10.1016/0013-4694(87)90180-5 [DOI] [PubMed] [Google Scholar]
- Kim C, Johnson NF, Cilles SE, Gold BT (2011) Common and distinct mechanisms of cognitive flexibility in prefrontal cortex. J Neurosci 31:4771–4779. 10.1523/JNEUROSCI.5923-10.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knight RT, Hillyard SA, Woods DL, Neville HJ (1980) The effects of frontal and temporal-parietal lesions on the auditory-evoked potential in man. Electroencephalogr Clin Neurophysiol 50:112–124. 10.1016/0013-4694(80)90328-4 [DOI] [PubMed] [Google Scholar]
- Knight RT, Scabini D, Woods DL (1989) Prefrontal cortex gating of auditory transmission in humans. Brain Res 504:338–342. 10.1016/0006-8993(89)91381-4 [DOI] [PubMed] [Google Scholar]
- Knight RT, Staines WR, Swick D, Chao LL (1999) Prefrontal cortex regulates inhibition and excitation in distributed neural networks. Acta Psychol (Amst) 101:159–178. 10.1016/s0001-6918(99)00004-9 [DOI] [PubMed] [Google Scholar]
- Lachaux JP, Rodriguez E, Martinerie J, Varela FJ (1999) Measuring phase synchrony in brain signals. Hum Brain Mapp 8:194–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung AW, He Y, Grady CL, Alain C (2013) Age differences in the neuroelectric adaptation to meaningful sounds. PLoS One 8:e68892. 10.1371/journal.pone.0068892 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lille F, Hassine L, Margules S (1991) Evoked potentials and age: different aging by sex? Neurophysiol Clin 21:459–472. 10.1016/s0987-7053(05)80336-3 [DOI] [PubMed] [Google Scholar]
- Lindenberger U, Baltes PB (1994) Sensory functioning and intelligence in old age: a strong connection. Psychol Aging 9:339–355. 10.1037/0882-7974.9.3.339 [DOI] [PubMed] [Google Scholar]
- Maillet D, Beaty RE, Jordano ML, Touron DR, Adnan A, Silvia PJ, Kwapil TR, Turner GR, Spreng RN, Kane MJ (2018) Age-related differences in mind-wandering in daily life. Psychol Aging 33:643–653. 10.1037/pag0000260 [DOI] [PubMed] [Google Scholar]
- Maillet D, Yu L, Hasher L, Grady CL (2020) Age-related differences in the impact of mind-wandering and visual distraction on performance in a go/no-go task. Psychol Aging 35:627–638. 10.1037/pag0000409 [DOI] [PubMed] [Google Scholar]
- Mammadkhanli O, Bozkurt M, Caglar YS (2020) Assessment of functional results for the lesions located in eloquent areas with using intraoperative cortical-subcortical stimulation and cortical mapping. Turk Neurosurg 30:854–863. [DOI] [PubMed] [Google Scholar]
- Maris E, Oostenveld R (2007) Nonparametric statistical testing of EEG- and MEG-data. J Neurosci Methods 164:177–190. 10.1016/j.jneumeth.2007.03.024 [DOI] [PubMed] [Google Scholar]
- Nakajima M, Schmitt LI, Halassa MM (2019) Prefrontal cortex regulates sensory filtering through a basal ganglia-to-thalamus pathway. Neuron 103:445–458.e410. 10.1016/j.neuron.2019.05.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picton TW, Stuss DT, Alexander MP, Shallice T, Binns MA, Gillingham S (2007) Effects of focal frontal lesions on response inhibition. Cereb Cortex 17:826–838. 10.1093/cercor/bhk031 [DOI] [PubMed] [Google Scholar]
- Plakke B, Romanski LM (2014) Auditory connections and functions of prefrontal cortex. Front Neurosci 8:199. 10.3389/fnins.2014.00199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plakke B, Romanski LM (2016) Neural circuits in auditory and audiovisual memory. Brain Res 1640:278–288. 10.1016/j.brainres.2015.11.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price D, Tyler LK, Neto Henriques R, Campbell KL, Williams N, Treder MS, Taylor JR, Henson RN, Cam-CAN (2017) Age-related delay in visual and auditory-evoked responses is mediated by white- and grey-matter differences. Nat Commun 8:15671. 10.1038/ncomms15671 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz N, Gunning FM, Head D, Dupuis JH, McQuain J, Briggs SD, Loken WJ, Thornton AE, Acker JD (1997) Selective aging of the human cerebral cortex observed in vivo: differential vulnerability of the prefrontal gray matter. Cereb Cortex 7:268–282. 10.1093/cercor/7.3.268 [DOI] [PubMed] [Google Scholar]
- Scannell JW, Burns GA, Hilgetag CC, O'Neil MA, Young MP (1999) The connectional organization of the cortico-thalamic system of the cat. Cereb Cortex 9:277–299. 10.1093/cercor/9.3.277 [DOI] [PubMed] [Google Scholar]
- Shen D, Vuvan DT, Alain C (2018) Cortical sources of the auditory attentional blink. J Neurophysiol 120:812–829. 10.1152/jn.00007.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strommer JM, Poldver N, Waselius T, Kirjavainen V, Jarvelainen S, Bjorksten S, Tarkka IM, Astikainen P (2017) Automatic auditory and somatosensory brain responses in relation to cognitive abilities and physical fitness in older adults. Sci Rep 7:13699. 10.1038/s41598-017-14139-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stuss DT (2006) Frontal lobes and attention: processes and networks, fractionation and integration. J Int Neuropsychol Soc 12:261–271. 10.1017/S1355617706060358 [DOI] [PubMed] [Google Scholar]
- Terada H, Kurayama T, Nakazawa K, Matsuzawa D, Shimizu E (2015) Transcranial direct current stimulation (tDCS) on the dorsolateral prefrontal cortex alters P50 gating. Neurosci Lett 602:139–144. 10.1016/j.neulet.2015.07.003 [DOI] [PubMed] [Google Scholar]
- Tobyne SM, Somers DC, Brissenden JA, Michalka SW, Noyce AL, Osher DE (2018) Prediction of individualized task activation in sensory modality-selective frontal cortex with 'connectome fingerprinting.' Neuroimage 183:173–185. 10.1016/j.neuroimage.2018.08.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walrath LC, Hallman LE (1984) Habituation of the event-related potential in elderly and young subjects. Ann NY Acad Sci 425:391–397. 10.1111/j.1749-6632.1984.tb23561.x [DOI] [PubMed] [Google Scholar]
- Welsh KA, Breitner JC, Magruder-Habib KM (1993) Detection of dementia in the elderly using telephone screening of cognitive status. Cogn Behav Neurol 6:103–110. [Google Scholar]
- Werner RA, Vanderzant CW (1991) Multimodality evoked potential testing in acute mild closed head injury. Arch Phys Med Rehabil 72:31–34. [PubMed] [Google Scholar]
- Woods DL, Clayworth CC (1986) Age-related changes in human middle latency auditory-evoked potentials. Electroencephalogr Clin Neurophysiol 65:297–303. 10.1016/0168-5597(86)90008-0 [DOI] [PubMed] [Google Scholar]
- Woods DL, Clayworth CC, Knight RT, Simpson GV, Naeser MA (1987) Generators of middle- and long-latency auditory-evoked potentials: implications from studies of patients with bitemporal lesions. Electroencephalogr Clin Neurophysiol 68:132–148. 10.1016/0168-5597(87)90040-2 [DOI] [PubMed] [Google Scholar]
- Yamaguchi S, Knight RT (1990) Gating of somatosensory input by human prefrontal cortex. Brain Res 521:281–288. 10.1016/0006-8993(90)91553-s [DOI] [PubMed] [Google Scholar]
- Zikopoulos B, Barbas H (2006) Prefrontal projections to the thalamic reticular nucleus form a unique circuit for attentional mechanisms. J Neurosci 26:7348–7361. 10.1523/JNEUROSCI.5511-05.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmermann J, Ross B, Moscovitch M, Alain C (2020) Neural dynamics supporting auditory long-term memory effects on target detection. Neuroimage 218:116979. 10.1016/j.neuroimage.2020.116979 [DOI] [PubMed] [Google Scholar]