

Abstract
Viruses are omnipresent, yet the knowledge on drivers of viral prevalence in wild host populations is often limited. Biotic factors, such as sympatric managed host species, as well as abiotic factors, such as climatic variables, are likely to impact viral prevalence. Managed and wild bees, which harbor several multi-host viruses with a mostly fecal–oral between-species transmission route, provide an excellent system with which to test for the impact of biotic and abiotic factors on viral prevalence in wild host populations. Here we show on a continental scale that the prevalence of three broad host viruses: the AKI-complex (Acute bee paralysis virus, Kashmir bee virus and Israeli acute paralysis virus), Deformed wing virus, and Slow bee paralysis virus in wild bee populations (bumble bees and solitary bees) is positively related to viral prevalence of sympatric honey bees as well as being impacted by climatic variables. The former highlights the need for good beekeeping practices, including Varroa destructor management to reduce honey bee viral infection and hive placement. Furthermore, we found that viral prevalence in wild bees is at its lowest at the extreme ends of both temperature and precipitation ranges. Under predicted climate change, the frequency of extremes in precipitation and temperature will continue to increase and may hence impact viral prevalence in wild bee communities.
Subject terms: Virus-host interactions, Viral epidemiology
Introduction
Even though the presence of viral pathogens often goes unnoticed, they form an indispensable facet of ecosystems1–3. However, when the natural dynamic interactions between hosts and their viral pathogens are disturbed they can have devastating effects on their hosts, often modulated by host shifts, as recently demonstrated by the SARS-CoV-2 pandemic4. The interactions between viruses and their hosts are a complex and can be affected by biotic factors, such as contact between wild and domesticated animals or humans5,6, as well as abiotic factors, such as temperature or precipitation7–13. Hence, to fully understand what drives viral prevalence and what triggers its negative effects on host populations, a thorough understanding of the role of biotic and abiotic factors is crucial for both wildlife as well as human welfare.
The virosphere of bees is very diverse, yet most knowledge on bee viruses and pathogens in general originates from studies of the managed Western honey bee, Apis mellifera14. Data for managed honey bees show that the currently documented viruses appear to have a global distribution14,15. In this managed species the viral landscape is highly impacted by the presence of the ectoparasitic mite, Varroa destructor, an effective vector of several viruses found in honey bees16,17. In the last decade, sparked by reported global declines of wild bees, research focus has shifted to viruses in wild bees18–20. As is true for the majority of pathogens21,22, most viruses in bees can be detected in multiple hosts16,23,24. Furthermore, co-infections with multiple pathogens are common, where different pathogens can impact one another inside the host20,25–28. Moreover, the bee microbiome may modulate within host pathogen dynamics29–31. Various studies have also highlighted the impact of agrochemical exposure of bees on their interaction with pathogens (reviewed in32).
One omnipresent factor impacting the whole bee community and its interactions with pathogens is climate. Albeit, several studies having shown that climatic variables can impact both the honey bee as a host as well as the interactions with its (viral) pathogens33–38, these studies are restricted to local scales and no study has investigated these impacts at large geographical scales. To understand the impact of viral pathogens on bees, we need to understand that a multitude of pathogens interact with different bee species within the pollinator community and how these interactions are influenced by climatic conditions across large spatial scales. The influence of climatic conditions can either be direct, affecting a host’s lifestyle, or indirect, by affecting pathogen transmission. Since most bee pathogens are transmitted via the fecal–oral route, enabling inter- and intra-species transmission via shared flowers16,39–42, climatic variables, such as UV-exposure, temperature and precipitation could potentially influence pathogen survival on flowers and thereby their transmission40. Furthermore, climatic variables affect vegetation phenology43, flower attractiveness44 and diversity45, which may alter the transmission network via flowers23, as well as quality, quantity of floral resources45–47, which may impact host immunity48,49 and consequently pathogen susceptibility and transmission.
Here, we performed a pan-European assessment of the virosphere of 12 bee communities, each consisting of three bee groups: bumble bees, solitary bee species and sympatric managed honey bees, on a continental scale and across different climatic zones (Fig. 1). Using an AICc-based multi-model inference approach50, we related viral prevalence, i.e. fraction of specimens with positive virus detection per bee group, in wild bees to that of managed honey bees, climatic conditions, i.e. temperature and precipitation, and vegetation phenology and tested for their interactions, particularly addressing the following two hypotheses:
Figure 1.
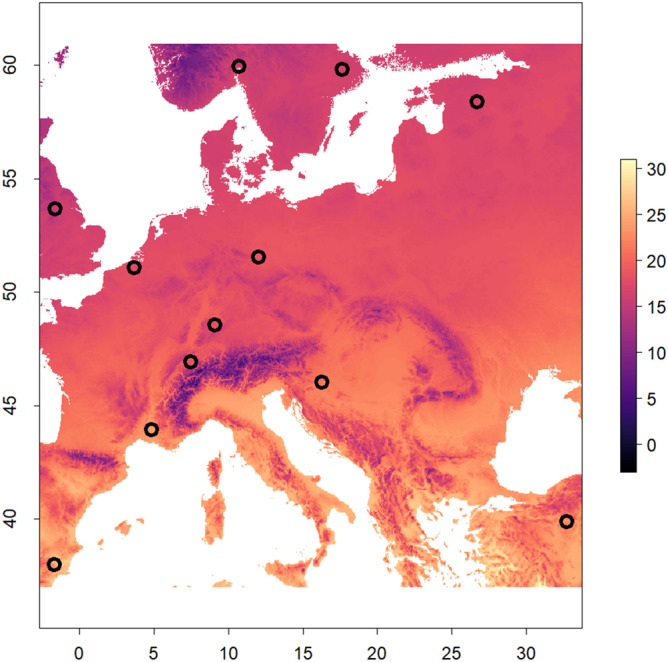
Map with the geographical distribution of the 12 sample sites (indicated with black circles) across Europe with color representing the climatic gradients (mean temperature of the warmest month [°C], as an example). Map created using R (version 4.0.4)51.
H1 Viral prevalence in wild bees is entirely determined by abiotic climatic conditions.
H2 Viral prevalence in wild bees is related to both biotic factors, namely viral prevalence in sympatric managed honey bees, as well as abiotic climatic conditions.
Results
In total, we screened 1227 bee specimens (495 honey bees, 476 bumble bees, 256 solitary bees) for the following three virus species: Deformed wing virus (DWV), Slow bee paralysis virus (SBPV), AKI-virus complex (i.e. Acute bee paralysis virus (ABPV), Kashmir bee virus (KBV) and Israeli acute paralysis virus (IAPV)) and calculated viral prevalence for each of the three bee groups. Across Europe (see Fig. 1) and all three bee groups, DWV was the most prevalent virus, while all viruses showed considerable geographic variation in their prevalence (Supplementary Fig. 1). An exclusive impact of climatic conditions or length of the vegetation period (i.e., approximation for temporal extent of flower availability) on the observed geographic variation in virus prevalence (H1) was not supported. While virus prevalence in honey bees was not related to environmental conditions (see Supplementary Table 1), there was a strong positive relation with viral prevalence in wild bees (Table 1; column ‘Apis’ present each model, with a positive coefficient), providing support for H2, i.e. a strong relationship between viral prevalence in honey bees and wild bees. The relationship with viral prevalence in honey bees was similar for both bumble bees and solitary bees (there was no interaction effect of species group). However, the relation was different for the three viruses (see Fig. 2). While the slope of this relationship was steep for the AKI-virus complex and SBPV, it was shallower for DWV, i.e., only higher prevalence of DWV in honey bees was reflected by an increase in DWV prevalence in wild bees (Fig. 2). In addition to the strong relation with viral prevalence in honey bees, wild bee viral prevalence depended also on climatic and vegetation conditions (Table 1), further supporting H2. The viral prevalence in wild bees generally declined across the year (Table 1; column ‘Phen’ indicated by a negative coefficient in all models). We also found virus species-specific responses to the temperature of the warmest quarter. This impact was negligible for DWV and SBPV, while the AKI-virus complex prevalence decreased with the temperature of the warmest quarter (Supplementary Fig. 3). The impact of precipitation of the warmest and driest quarter (Fig. 3A,B) and temperature of the driest quarter (Fig. 3C) was hump-shaped. Viral prevalence in wild bees was particularly low at both high and low levels of precipitation and temperature but without differences among the viruses.
Table 1.
Multi-model inference results for separate analyses of climatic and vegetation phenological conditions ordered with increasing AICc.
| Apis | Phen | Env | Env2 | Virus | Apis:Env | Apis:Virus | Env:Virus | AICc | Delta | Weight | R2m | R2c |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Precipitation warmest quarters | ||||||||||||
| 1.29 | − 0.78 | − 1.04 | 228.94 | 0.00 | 0.65 | 0.33 | 0.67 | |||||
| 1.30 | − 0.84 | 0.15 | − 0.98 | 231.60 | 2.66 | 0.17 | 0.35 | 0.67 | ||||
| 1.45 | 232.84 | 3.90 | 0.09 | 0.27 | 0.58 | |||||||
| 1.53 | − 0.41 | 232.85 | 3.92 | 0.09 | 0.31 | 0.59 | ||||||
| Temperature warmest quarter | ||||||||||||
| 2.52 | − 0.55 | − 1.40 | + | + | 230.89 | 0.00 | 0.18 | 0.51 | 0.64 | |||
| 1.34 | − 0.75 | − 0.92 | 231.83 | 0.94 | 0.11 | 0.26 | 0.66 | |||||
| 2.68 | − 1.16 | + | + | 232.15 | 1.27 | 0.10 | 0.53 | 0.66 | ||||
| 2.78 | − 0.56 | − 1.22 | − 0.34 | + | + | 232.64 | 1.76 | 0.07 | 0.56 | 0.66 | ||
| 1.45 | 232.84 | 1.95 | 0.07 | 0.27 | 0.58 | |||||||
| 1.53 | − 0.41 | 232.85 | 1.97 | 0.07 | 0.31 | 0.59 | ||||||
| 2.38 | − 0.58 | − 1.57 | + | 0.30 | + | 233.00 | 2.11 | 0.06 | 0.52 | 0.64 | ||
| 3.00 | − 0.48 | − 1.55 | + | + | + | 233.17 | 2.28 | 0.06 | 0.56 | 0.66 | ||
| 3.10 | − 1.52 | + | + | + | 233.32 | 2.43 | 0.05 | 0.55 | 0.66 | |||
| 1.36 | − 0.73 | − 0.91 | 0.17 | 233.59 | 2.71 | 0.05 | 0.28 | 0.65 | ||||
| 2.68 | − 0.57 | − 1.40 | − 0.52 | + | 0.43 | + | 233.83 | 2.95 | 0.04 | 0.58 | 0.67 | |
| 2.71 | − 1.06 | − 0.25 | + | + | 234.32 | 3.44 | 0.03 | 0.54 | 0.66 | |||
| 1.33 | − 0.79 | − 0.91 | − 0.16 | 234.39 | 3.50 | 0.03 | 0.26 | 0.67 | ||||
| 2.44 | − 1.25 | + | 0.25 | + | 234.55 | 3.66 | 0.03 | 0.51 | 0.64 | |||
| 2.16 | − 0.48 | + | 234.67 | 3.79 | 0.03 | 0.42 | 0.63 | |||||
| 1.37 | − 0.27 | 234.73 | 3.84 | 0.03 | 0.23 | 0.59 | ||||||
| Precipitation driest quarter | ||||||||||||
| 1.36 | − 0.54 | − 1.02 | 231.02 | 0.00 | 0.30 | 0.33 | 0.63 | |||||
| 1.34 | − 0.71 | 232.72 | 1.71 | 0.13 | 0.28 | 0.59 | ||||||
| 1.45 | 232.84 | 1.82 | 0.12 | 0.27 | 0.58 | |||||||
| 1.53 | − 0.41 | 232.85 | 1.84 | 0.12 | 0.31 | 0.59 | ||||||
| 1.40 | − 0.68 | 0.24 | − 0.90 | 233.46 | 2.45 | 0.09 | 0.36 | 0.62 | ||||
| 2.32 | − 0.89 | 0.60 | + | 233.65 | 2.63 | 0.08 | 0.50 | 0.66 | ||||
| 1.60 | − 0.73 | 0.47 | 233.89 | 2.87 | 0.07 | 0.37 | 0.61 | |||||
| 1.29 | − 0.32 | − 0.98 | 234.35 | 3.33 | 0.06 | 0.27 | 0.62 | |||||
| 2.16 | − 0.48 | + | 234.67 | 3.66 | 0.05 | 0.42 | 0.63 | |||||
| Temperature driest quarter | ||||||||||||
| 1.41 | − 0.75 | − 0.98 | 231.19 | 0.00 | 0.32 | 0.30 | 0.62 | |||||
| 1.45 | 232.84 | 1.65 | 0.14 | 0.27 | 0.58 | |||||||
| 1.53 | − 0.41 | 232.85 | 1.67 | 0.14 | 0.31 | 0.59 | ||||||
| 1.43 | − 0.81 | 0.41 | − 1.29 | 233.54 | 2.36 | 0.10 | 0.32 | 0.61 | ||||
| 1.45 | − 0.45 | − 0.50 | 234.16 | 2.97 | 0.07 | 0.28 | 0.61 | |||||
| 1.39 | − 0.34 | 234.48 | 3.30 | 0.06 | 0.25 | 0.58 | ||||||
| 1.40 | − 0.35 | 234.59 | 3.41 | 0.06 | 0.24 | 0.59 | ||||||
| 1.83 | − 0.72 | − 0.79 | + | 234.59 | 3.41 | 0.06 | 0.37 | 0.61 | ||||
| 2.16 | − 0.48 | + | 234.67 | 3.49 | 0.06 | 0.42 | 0.63 | |||||
| Length of vegetation period | ||||||||||||
| 1.44 | − 1.24 | 1.12 | 231.69 | 0.00 | 0.26 | 0.40 | 0.67 | |||||
| 1.45 | 232.84 | 1.15 | 0.15 | 0.27 | 0.58 | |||||||
| 1.53 | − 0.41 | 232.85 | 1.17 | 0.15 | 0.31 | 0.59 | ||||||
| 1.38 | − 1.39 | 0.65 | 0.49 | 233.76 | 2.07 | 0.09 | 0.33 | 0.66 | ||||
| 1.46 | − 1.25 | 1.15 | − 0.04 | 234.37 | 2.68 | 0.07 | 0.41 | 0.67 | ||||
| 2.16 | − 0.48 | + | 234.67 | 2.98 | 0.06 | 0.42 | 0.63 | |||||
| 1.87 | − 1.07 | 0.86 | + | 234.88 | 3.20 | 0.05 | 0.44 | 0.65 | ||||
| 1.49 | − 0.11 | 235.09 | 3.41 | 0.05 | 0.29 | 0.59 | ||||||
| 1.94 | − 0.55 | + | + | 235.28 | 3.59 | 0.04 | 0.49 | 0.64 | ||||
| 1.94 | + | 235.31 | 3.62 | 0.04 | 0.36 | 0.60 | ||||||
| 1.44 | 0.05 | 235.34 | 3.65 | 0.04 | 0.26 | 0.58 | ||||||
Apis, viral prevalence in Apis mellifera; Phen, vegetation phenology during sampling; Env, linear term of respective environmental variable; Env2, quadratic term of respective environmental variable; Virus, virus species; colon indicates interaction terms. Numbers are coefficient estimates of continuous variables; ‘+’ indicates relevance of categorical variables. AICc, Akaike information criterion corrected for small sample sizes; Delta; delta AICc; Weight, Akaike weight; R2m, marginal pseudo-R2 (only fixed effects52); R2c, conditional pseudo-R2 (fixed and random effects). Models with delta AICc < 2 indicate strong support and are marked with grey background. White background: models with delta AICc between 2 and 4, indicating weaker support.
Figure 2.

Relationship between viral prevalence in managed honey bees and wild bees split by virus. Analysis indicated a strong relation between the viral prevalence in honey bees and wild bees, the slope of the relation differs, however, between the viruses. For both SBPV (top panel) and the AKI-complex (bottom left panel) we see a clear increase in viral prevalence in wild bees as the viral prevalence in managed honey bees increases. For DWV (bottom right panel) we see a similar relation, yet only when the viral prevalence in honey bees increases above 50%. Shaded blue indicates the 95% confidential interval.
Figure 3.

Relationship between viral prevalence (all viruses together) in wild bees (y- axis) and mean precipitation of the warmest quarter (A), mean precipitation of the driest quarter (B) and mean temperature of the driest quarter (C). Analysis indicated that the viral prevalence in wild bees was significantly affected by these three climatic variables. Precipitation and temperature means were obtained over a period of more than two decades at a resolution of 1–3 km. Shaded blue indicates the 95% confidential interval.
The impacts of vegetation period or environmental conditions on the relation between viral prevalence in managed honey bees and wild bees were only weakly supported (2 < ΔAICc < 4; Table 1: column ‘Apis: Env). Of all environmental conditions, only temperature of the warmest quarter had an impact on the relation in viral prevalence between honey bees and wild bees (Supplementary Fig. 2a) Further, we found that the relation between honey bee viral prevalence and viral prevalence in wild bees was less steep in areas with shorter vegetation periods, while it increased with the length of the vegetation period (Supplementary Fig. 2b).
Discussion
Within this study we specifically address viral prevalence in wild bees with respect to viral prevalence of sympatric honey bees as a biotic factor, as well as temperature and precipitation as climatic abiotic factors. We find that wild bee viral prevalence is affected by both the biotic factor as well as the abiotic factors, supporting our second hypothesis. We find only a weakly supported interaction effect between honey bee viral prevalence and the climatic factors on wild bee viral prevalence (see Table 1 column ‘Apis: Env’; the models with an interaction term all have ΔAICc > 2).
The viral prevalence of multi-host viruses in the bee community is, amongst others, determined by the presence and density of different host species. As bee hosts are all somehow connected through their shared floral resources and viral transmission can be multidirectional for multi-host viruses in the bee community, we cannot demonstrate the directionality of viral transmission in our study. Nevertheless, we find a strong link between the viral prevalence in wild bees and that of sympatric honey bees. This result confirms earlier studies that demonstrate that viral prevalence in wild bees is related to viral prevalence of sympatric honey bees19,20,53. While these former studies were restricted to local53 and national scales19,20, we expand these relationships to a continental scale.
Interestingly, the effects of managed honey bee viral prevalence on wild bee viral prevalence did not differ between bumble bees and solitary bees, despite their different lifestyles and social organization, thereby underlining the importance of interspecific pathogen transmission, e.g. via shared floral resources16,39,42,54,55. Even though some solitary bee species are dietary specialists, visiting flowers of only a few plant species, the presence of generalist species such as honey bees connect many species in one network module, increasing the likelihood of viral transmission to all bee species present in the same environment56–60. However, we found that the relation between viral prevalence in honey bees and wild bees differs between viruses. There is a tight relationship for both AKI-virus complex and SBPV, with increasing wild bee prevalence when honey bee prevalence increases. Yet, although a similar relation was found for DWV, this relation only becomes apparent when viral prevalence in honey bees was high (see Fig. 2).
In contrast to AKI-virus complex and SBPV, DWV is a virus which is predominantly found in honey bees20,59,61. This discrepancy in viral prevalence is also reflected in our results, where DWV is always more prevalent in honey bees at our different sampling sites compared to the other viruses, namely the AKI-virus complex and SBPV. Experimental infection assays have shown that all three viruses can infect both wild bees as well as honey bees19,62–65. However, in contrast to wild bees, honey bees face the additional pressure of the parasitic mite Varroa destructor, which besides its virus-vectoring capacity can also weaken colonies, increasing the impact of viral infections66,67. A recent study highlighted the role of V. destructor in the transmission dynamics of DWV55. Incorporation of within-honey bee colony transmission, mediated by V. destructor vectoring and bee-to-bee transfer into a transmission model could explain the higher DWV prevalence in managed honey bees compared to wild bumble bees observed in the field55. These results may also explain why we only observed DWV in wild bees when its prevalence in honey bees was high and the environmental viral load is likely to have been high.
Although both the AKI-virus complex and SBPV can also be vectored and amplified to lethal levels in honey bee colonies by V. destructor, these viruses are more commonly found in wild bees20,59,61,68,69, suggesting key differences in host susceptibility and competence between these three viruses. The strong relations found in our study between viral prevalence in honey bees and wild bees together with the results from previous studies19,55 highlight the need for good beekeeping practices, which include proactive varroa monitoring and control by beekeepers as well as appropriate hive placement, where the beekeeper takes into account the presence of endangered wild bees, to prevent managed honey bee colonies becoming sources of virus amplification and dissemination to wild bees19,55,70.
In spite of the high variation in viral prevalence in managed honey bees across Europe (Supplementary Fig. 1), we found no relation between viral prevalence in managed honey bees and the investigated environmental conditions, temperature and precipitation (Supplementary Table 1). In contrast to most wild bee species, honey bees are highly social insects, who actively control the climate within the hive71, and can maintain large food reserves to overcome poor foraging conditions; these traits may make their viral dynamics independent from environmental conditions. Moreover, the active management of honey bees may contribute to the absence of an environmental effect on viral prevalence. Beekeeping practices, such as hive placement and water provisioning as well as varroa mite control and nutritional support72, could have a large impact on the health status and viral prevalence in colonies, hence they may obscure potentially smaller effects of environmental conditions in modulating viral prevalence.
Contrary to honey bees, we found that viral prevalence in wild bees was related to environmental conditions, whereby intermediate temperatures and intermediate precipitation of the warmest and driest quarters led to the highest viral prevalence in wild bees.
The impact of climatic conditions, in particular those related to heat and drought stress (precipitation in the driest and in the warmest quarter, temperature in the driest quarter), was consistent across all three viruses. For these environmental conditions, viral prevalence in wild bees was highest in the most moderate (intermediate) conditions and lowest at the extreme ends. Here one might argue that these extremes of both precipitation in the warmest and driest quarter as well as temperature in the driest quarter are likely suboptimal for most pollinator species as well as their floral resources. This is either due to a lack of rain-free days for foraging or drought or a combination of drought and heat, which may have a negative impact on the availability and attractiveness of floral resources44,47. These suboptimal conditions can induce physiological and nutritional stress, which can either result in reduced host populations73,74 and hence lower viral transmission and prevalence or result in an increased virulence of viral infections due to stressed hosts63. Malnutrition has been shown to negatively affect a host’s defense system75, which can increase virus-induced mortality76, as sown for Slow bee paralysis virus and DWV infections in bumble bees63,77, resulting in lower viral prevalence78,79. In the light of climate change, which has been shown to impact host–pathogen interaction80,81, we anticipate that these extremes in both temperature and precipitation will increase82,83. This, together with the ongoing loss of suitable naturally occurring floral resources84, can further increase the nutritional stress on wild bees, which may amplify pathogen stress85, and hence warrants further research. Besides their direct effect on hosts, these environmental conditions may also affect the viruses themselves. Heat, UV-exposure and drought may impact virus viability on the flowers, and increased rainfall may increase wash-off or dilute the virus below its minimal infection dose, leading to lower viral transmission at flowers.
We found only weak support for environmental modulation of the relationship between viral prevalence in honey bees and wild bees. Here the link between viral prevalence in honey bees and wild bees increased with the length of the vegetation period (Supplementary Fig. 2b), which may be attributed to prolonged foraging activity, facilitating transmission across a longer period of time together with the increase in viral prevalence in honey bee colonies as the foraging season progresses14,86.
Overall, our study shows at a continental scale that wild bee viral prevalence is affected by both biotic factors, namely viral prevalence in sympatric honey bees, and abiotic factors, specifically climatic conditions. The role of managed honey bees as an impact on wild bee viral prevalence has been identified before on a smaller scale, yet the link with climate has to our knowledge not been addressed at a continental scale. Temperature and precipitation extremes will likely continue to increase in the coming years due to climate change, and pathogens can exert negative effects when their interaction dynamics change85. Understanding pathogen prevalence in an environmental context, especially that of threatened species, can improve conservation strategies and hence deserves more attention.
Methods
The bee sampling, identification, processing and virus detection protocols were performed as described in Miranda et al.87, and described in short here below. Sample collection, identification, processing, and cDNA synthesis was all done by the local partner. The virus assays were conducted by the partners in Belgium, France, Germany, Sweden, and Switzerland.
Bee collection
Foraging adults were collected at 12 sites, in 11 countries across a climatic gradient in Europe (Fig. 1). The field sampling was performed according to a standardized sampling protocol87, where each partner sampled in their respective country. At each site 30 honey bees and 30 wild bees were collected in the order they were encountered as well an additional 15 specimens of the most common wild bee. The presence of wild honey bees is close to non-existing in Europe, with the exception of a few records. It is assumed that these wild colonies disappeared due to the introduced parasitic mite Varroa destructor and its viral vectoring capacity88,89. It is therefore reasonable to presume that nearly all the caught honey bees in this study originate from managed colonies and not from wild ones. At each site, all specimens were collected on the same day and in the same flower-rich area (ca. 100 m2) and then stored individually on ice after capture. Upon return to the laboratory, all samples were stored immediately at − 80 °C until further analysis. Sampling date varied between sites as bees were collected between April and September on a day with high foraging activity.
RNA extraction from individual bees and cDNA production
For RNA extraction of the bees, we used the protocol described by de Miranda et al.87. For each bee we only used the abdomen, which was dissected sterile and crushed in a TBS-buffer (50 mM TRIS.HCl PH 7.4, 150 mM NaCl). The volume of TBS buffer used differed depending on the size of the abdomen, i.e., 800 µl for bumble bee sized abdomens, 500 µl for honey bee sized abdomens and 200 µl for small sweat bee sized abdomens. The homogenate was centrifuged to spin down the exoskeleton then 100 µl of supernatant was mixed with 350 µl RLT buffer supplemented with 1% beta-mercaptoethanol. RNA extraction was done using the Qiagen Plant RNeasy Plant kit (Qiagen) according to manufacturer’s protocol and was eluted with 50 µl (for large bees) or 30 µl (for small bees) nuclease free water. RNA concentrations were measured using NanoDrop and adjusted to 100 ng/µl using nuclease free water. cDNA was prepared using random hexamer primers and 1 µg RNA and cDNA kit (#K1612, ThermoFisher Scientific, Waltham, Massachusetts, United States of America) containing M-MLV reverse transcriptase and RNAse inhibitor in a 20 µl volume according to the manufacturer’s protocol.
Virus screening
All bees were screened for three common bee virus targets, namely the AKI-virus complex (acute bee paralysis virus, Kashmir bee virus and Israeli acute paralysis virus), deformed wing virus and slow bee paralysis virus.
These viruses were chosen based on their wide distribution, relative high prevalence, and broad host range where they are detected in both wild bees and honey bees, which was a perquisite to test our hypothesis on the impact of honey bees as a biotic factor on wild bee viral prevalence.
Each PCR reaction contained 2 µl of diluted (1/10) cDNA and 18 µl qPCR mixture, containing 0.2 µl of both forward and reverse primers. For primer sequences, see Supplementary Table 2. Although the used primers are broad-range, capable of detecting several viral strains and species within a complex, we are aware that they are not flawless and may have missed some viral strains/variants due to the high mutation and recombination rates of these RNA-viruses.
Statistical analysis
Viral prevalence was calculated for honey bees and wild bees, aggregated into two taxonomic groups, bumble bees and solitary bees, as the fraction of specimens scored positive in virus detection. We used a Cq threshold of 35 to define positive viral detection (Cq < 35) per species group. As we report the detection of viruses and classify a bee as positive if Cq values are below the set threshold, we are aware that we cannot state that positive bees are truly infected. This would require the detection of replicating virus (e.g. by detecting the negative strand of these positive strand RNA viruses), which we have not done. All analysis were performed using R (version 4.0.4)51 and the following packages (rgdal, raster, sp, dplyr, Hmisc, glmmTMB, MuMIn, ggplot2, effects)90–98.
Explanatory variables
We selected an initial set of the following nine bioclimatic variables covering relevant temperature and precipitation conditions: annual mean temperature (BIO1), temperature seasonality (BIO4), mean temperature of driest quarter (BIO9), mean temperature of warmest quarter (BIO10), mean temperature of coldest quarter (BIO11), annual precipitation (BIO12), precipitation seasonality (BIO15), precipitation of driest quarter (BIO17), and precipitation of warmest quarter (BIO18).
Bioclimatic variables were obtained from CHELSA (v1.2) at a resolution of 30 arc seconds (about 1 km) averaged across the years 1979–201399,100. To analyze the impact of activity period and time of sampling in the respective sampling year, we extracted data on the start and length of the vegetation period for each site as a proxy for the time of flower availability. Calculation of the vegetation period was based on the Normalized Difference Vegetation Index (NDVI) and provided by the Vegetation Index and Phenology Lab at a 3 arc min (about 5.6 km) resolution (https://vip.arizona.edu/). The actual sampling date was related to the start of the vegetation period per site (number of days since start of the vegetation period) and is referred to as ‘sampling phenology’.
All variables were tested for collinearity with a hierarchical cluster analysis based on pairwise Spearman rank correlation coefficients using UPGMA (unweighted pair-group method with arithmetic averages) agglomeration. Only non-collinear variables were retained (Spearman r < 0.7). Selection from clusters of collinear variables was based on ecological relevance (e.g. mean temperature of the warmest quarter was selected instead of annual mean temperature). After selection, the following six variables remained: mean temperature of driest quarter, mean temperature of warmest quarter, precipitation of driest quarter, precipitation of warmest quarter, length of vegetation period, and sampling phenology.
Model development
To test the impact of climate and phenology on honey bee viral prevalence, we used a generalized linear mixed effects model (GLMM) with a binomial error structure (prevalence between 0 and 1) and a logit link, weighted by sample size. Climatic variables were included with their linear and quadratic terms. All three viruses were considered in one model to test for differential responses among the viruses, i.e., interaction between virus type and environmental variable. As crossed random effects, we considered virus type to avoid pseudo-replication, assay laboratory to account for potential systematic differences among the analyzing laboratories and an observer term to address overdispersion where necessary.
To test the relationship between viral prevalence in wild bees and honey bees and the, potentially modulating, impact of environmental variables, we used the same approach as described above but added main effects, two-way and three-way interactions of viral prevalence in honey bees, environment, and virus type. Bumble bees and solitary bees were considered in one model. Group-specific responses to viral prevalence in honey bees were initially tested with interaction effects but without environmental variables to avoid model overfitting. No interaction was evident and thus it was excluded from subsequent analyses. We also added bee taxon (bumble bees, solitary bees) as an additional random effect.
To avoid model overfitting, we limited the number of fixed effects and each environmental variable was tested separately while keeping sampling phenology as a covariate to control for potential differences in viral prevalence due to differences in the time of sampling across the year. We used a multi-model inference approach based on the Akaike Information Criterion corrected for small sample sizes (AICc)101 for model simplification. We considered models with a delta AICc lower than 2, towards the best model, having strong support and models with delta AICc between 2 and 4 as having weaker support. We disregarded a set of models for a particular environmental variable if the intercept-only model was within the respective subset (set of models with delta AICc < 2 and set of models with delta 2 < AICc < 4).
Supplementary Information
Acknowledgements
We thank the referees for valuable and insightful comments which helped improve the clarity of the manuscript. The authors acknowledge the support of the EU COST-Action FA1307 Sustainable pollination in Europe-joint research on bees and other pollinators (Super-B) for providing the platform to initiate this project, as well as internal funders from each country and institute for financing the sample collection and processing costs. We also would like to thank the technicians and students at the collecting-processing labs and the assaying labs for their invaluable help and competence. RJP thanks Kerstin Gößel and Anja Manigk for technical help. JRM would like to thank the excellent technical support provided by MSc student Sofie Tinggren, BMA student Nellie Svedin and BMA Emilia Semberg during the course of this project. GS, NP and IM would like to thank student Tian Tian for her practical assistance and the Research Foundation-Flanders (FWO-Vlaanderen) for funding. PG would like to thank Crystal Frost for technical assistance. AD would like to thank Maxime Thomasson and Elodie Boeuf for their technical support, Bernard E. Vaissière and Laurent Guilbaud for their support in taxonomy for wild bees, and the scientific department SPE of INRAE for funding. RJP thanks the DFG (Pa632/10) for funding.
Author contributions
All authors participated in the conception of the project. IM and RJP designed the sample collection strategy. J.R.M., I.M., G.S., P.N. and O.Y. designed the initial sample processing methods. J.R.M., I.M., O.Y., G.S., N.P. and P.N. designed the initial pathogen assay methods. R.J.P. and J.R.M. designed the barcoding identification method. All authors contributed to the optimization and validation of the protocols. All authors contributed to the sample collection and processing. J.R.M., I.M., O.Y., S.M., P.N., R.R., A.D., L.J. and V.D. performed the pathogen assays. JRM curated the data. O.S. performed the statistical analyses. N.P. wrote the first draft of the manuscript. All authors contributed to editing the manuscript. All authors read and approved the final manuscript.
Funding
Open access funding provided by Swedish University of Agricultural Sciences.
Data availability
The dataset is included as a supplementary file.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Niels Piot, Email: niels.piot@ugent.be.
Robert J. Paxton, Email: robert.paxton@zoologie.uni-halle.de
Joachim R. de Miranda, Email: joachim.de.miranda@slu.se
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-022-05603-2.
References
- 1.Lafferty KD, Dobson AP, Kuris AM. Parasites dominate food web links. Proc. Natl. Acad. Sci. USA. 2006;103:11211–11216. doi: 10.1073/pnas.0604755103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wood CL, Johnson PT. A world without parasites: Exploring the hidden ecology of infection. Front. Ecol. Environ. 2015;13:425–434. doi: 10.1890/140368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Preston DL, Mischler JA, Townsend AR, Johnson PTJ. Disease ecology meets ecosystem science. Ecosystems. 2016;19:737–748. doi: 10.1007/s10021-016-9965-2. [DOI] [Google Scholar]
- 4.Petersen E, et al. Comparing SARS-CoV-2 with SARS-CoV and influenza pandemics. Lancet Infect. Dis. 2020;20:e238–e244. doi: 10.1016/S1473-3099(20)30484-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Daszak P. Emerging infectious diseases of wildlife—Threats to biodiversity and human health. Science (80-). 2000;287:443–449. doi: 10.1126/science.287.5452.443. [DOI] [PubMed] [Google Scholar]
- 6.Mack R. The great African cattle plague epidemic of the 1890s. Trop. Anim. Health Prod. 1970;2:210–219. doi: 10.1007/BF02356441. [DOI] [Google Scholar]
- 7.Van Hemert C, Pearce JM, Handel CM. Wildlife health in a rapidly changing North: Focus on avian disease. Front. Ecol. Environ. 2014;12:548–556. doi: 10.1890/130291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hueffer K, O’Hara TM, Follmann EH. Adaptation of mammalian host-pathogen interactions in a changing arctic environment. Acta Vet. Scand. 2011;53:17. doi: 10.1186/1751-0147-53-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Musgrave K, Bartlow AW, Fair JM. Long-term variation in environmental conditions influences host–parasite fitness. Ecol. Evol. 2019;9:7688–7703. doi: 10.1002/ece3.5321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Poisot T, Guéveneux-Julien C, Fortin MJ, Gravel D, Legendre P. Hosts, parasites and their interactions respond to different climatic variables. Glob. Ecol. Biogeogr. 2017;26:942–951. doi: 10.1111/geb.12602. [DOI] [Google Scholar]
- 11.Elliot SL, Blanford S, Thomas MB. Host–pathogen interactions in a varying environment: Temperature, behavioural fever and fitness. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002;269:1599–1607. doi: 10.1098/rspb.2002.2067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lambertini C, et al. Biotic and abiotic determinants of Batrachochytrium dendrobatidis infections in amphibians of the Brazilian Atlantic Forest. Fungal Ecol. 2021;49:100995. doi: 10.1016/j.funeco.2020.100995. [DOI] [Google Scholar]
- 13.Hoarau AOG, Mavingui P, Lebarbenchon C. Coinfections in wildlife: Focus on a neglected aspect of infectious disease epidemiology. PLOS Pathog. 2020;16:e1008790. doi: 10.1371/journal.ppat.1008790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Beaurepaire A, et al. Diversity and global distribution of viruses of the western honey bee, Apis mellifera. Insects. 2020;11:239. doi: 10.3390/insects11040239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Boncristiani H, et al. World honey bee health: The global distribution of western honey bee (Apis mellifera L.) pests and pathogens. Bee World. 2021;98:1–5. doi: 10.1080/0005772X.2020.1800330. [DOI] [Google Scholar]
- 16.Yañez O, et al. Bee viruses: Routes of infection in hymenoptera. Front. Microbiol. 2020;11:1–22. doi: 10.3389/fmicb.2020.00943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Martin SJ, et al. Global honey bee viral landscape altered by a parasitic mite. Science (80-). 2012;336:1304–1306. doi: 10.1126/science.1220941. [DOI] [PubMed] [Google Scholar]
- 18.Ravoet J, et al. Widespread occurrence of honey bee pathogens in solitary bees. J. Invertebr. Pathol. 2014;122:55–58. doi: 10.1016/j.jip.2014.08.007. [DOI] [PubMed] [Google Scholar]
- 19.Fürst MA, McMahon DP, Osborne JL, Paxton RJ, Brown MJF. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature. 2014;506:364–366. doi: 10.1038/nature12977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.McMahon DP, et al. A sting in the spit: Widespread cross-infection of multiple RNA viruses across wild and managed bees. J. Anim. Ecol. 2015;84:615–624. doi: 10.1111/1365-2656.12345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rigaud T, Perrot-Minnot M-J, Brown MJF. Parasite and host assemblages: Embracing the reality will improve our knowledge of parasite transmission and virulence. Proc. R. Soc. B Biol. Sci. 2010;277:3693–3702. doi: 10.1098/rspb.2010.1163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rodrigues RAL, et al. An anthropocentric view of the virosphere–host relationship. Front. Microbiol. 2017;8:1–11. doi: 10.3389/fmicb.2017.01673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Daughenbaugh KF, et al. Metatranscriptome analysis of sympatric bee species identifies bee virus variants and a new virus, Andrena-associated bee virus-1. Viruses. 2021;13:291. doi: 10.3390/v13020291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Olgun T, Everhart SE, Anderson T, Wu-Smart J. Comparative analysis of viruses in four bee species collected from agricultural, urban, and natural landscapes. PLoS ONE. 2020;15:e0234431. doi: 10.1371/journal.pone.0234431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Carrillo-Tripp J, et al. In vivo and in vitro infection dynamics of honey bee viruses. Sci. Rep. 2016;6:22265. doi: 10.1038/srep22265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Remnant EJ, Mather N, Gillard TL, Yagound B, Beekman M. Direct transmission by injection affects competition among RNA viruses in honeybees. Proc. Biol. Sci. 2019;286:20182452. doi: 10.1098/rspb.2018.2452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jones CM, Brown MJF. Parasites and genetic diversity in an invasive bumblebee. J. Anim. Ecol. 2014 doi: 10.1111/1365-2656.12235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Faurot-Daniels C, et al. Longitudinal monitoring of honey bee colonies reveals dynamic nature of virus abundance and indicates a negative impact of Lake Sinai virus 2 on colony health. PLoS ONE. 2020;15:e0237544. doi: 10.1371/journal.pone.0237544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Engel P, et al. The bee microbiome: Impact on bee health and model for evolution and ecology of host-microbe interactions. MBio. 2016;7:e02164–e2215. doi: 10.1128/mBio.02164-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Maebe K, et al. The holobiont as a key to the adaptation and conservation of wild bees in the anthropocene. Front. Ecol. Evol. 2021;9:1–5. doi: 10.3389/fevo.2021.781470. [DOI] [Google Scholar]
- 31.McMenamin AJ, Brutscher LM, Glenny W, Flenniken ML. Abiotic and biotic factors affecting the replication and pathogenicity of bee viruses. Curr. Opin. Insect Sci. 2016;16:14–21. doi: 10.1016/j.cois.2016.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Harwood GP, Dolezal AG. Pesticide–virus interactions in honey bees: Challenges and opportunities for understanding drivers of bee declines. Viruses. 2020;12:566. doi: 10.3390/v12050566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Dalmon A, Peruzzi M, Le Conte Y, Alaux C, Pioz M. Temperature-driven changes in viral loads in the honey bee Apis mellifera. J. Invertebr. Pathol. 2019;160:87–94. doi: 10.1016/j.jip.2018.12.005. [DOI] [PubMed] [Google Scholar]
- 34.Le Conte Y, Navajas M. Climate change: Impact on honey bee populations and diseases. Rev. Sci. Tech. 2008;27(485–497):499–510. [PubMed] [Google Scholar]
- 35.Switanek M, Crailsheim K, Truhetz H, Brodschneider R. Modelling seasonal effects of temperature and precipitation on honey bee winter mortality in a temperate climate. Sci. Total Environ. 2017;579:1581–1587. doi: 10.1016/j.scitotenv.2016.11.178. [DOI] [PubMed] [Google Scholar]
- 36.Flores JM, et al. Effect of the climate change on honey bee colonies in a temperate Mediterranean zone assessed through remote hive weight monitoring system in conjunction with exhaustive colonies assessment. Sci. Total Environ. 2019;653:1111–1119. doi: 10.1016/j.scitotenv.2018.11.004. [DOI] [PubMed] [Google Scholar]
- 37.Martin-Hernandez R, et al. Effect of temperature on the biotic potential of honeybee microsporidia. Appl. Environ. Microbiol. 2009;75:2554–2557. doi: 10.1128/AEM.02908-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Appler R, Frank S, Tarpy D. Within-colony variation in the immunocompetency of managed and feral honey bees (Apis mellifera L.) in different urban landscapes. Insects. 2015;6:912–925. doi: 10.3390/insects6040912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Alger SA, Burnham PA, Brody AK. Flowers as viral hot spots: Honey bees (Apis mellifera) unevenly deposit viruses across plant species. PLoS ONE. 2019;14:e0221800. doi: 10.1371/journal.pone.0221800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Figueroa LL, et al. Bee pathogen transmission dynamics: Deposition, persistence and acquisition on flowers. Proc. R. Soc. B Biol. Sci. 2019;286:20190603. doi: 10.1098/rspb.2019.0603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.McArt SH, Koch H, Irwin RE, Adler LS. Arranging the bouquet of disease: Floral traits and the transmission of plant and animal pathogens. Ecol. Lett. 2014;17:624–636. doi: 10.1111/ele.12257. [DOI] [PubMed] [Google Scholar]
- 42.Durrer S, Schmid-Hempel P. Shared use of flowers leads to horizontal pathogen transmission. Proc. R. Soc. B Biol. Sci. 1994;258:299–302. doi: 10.1098/rspb.1994.0176. [DOI] [Google Scholar]
- 43.Zhang X, Tarpley D, Sullivan JT. Diverse responses of vegetation phenology to a warming climate. Geophys. Res. Lett. 2007;34:L19405. doi: 10.1029/2007GL031447. [DOI] [Google Scholar]
- 44.Descamps C, Jambrek A, Quinet M, Jacquemart A-L. Warm temperatures reduce flower attractiveness and bumblebee foraging. Insects. 2021;12:493. doi: 10.3390/insects12060493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Suggitt AJ, Lister DG, Thomas CD. Widespread effects of climate change on local plant diversity. Curr. Biol. 2019;29:2905–2911.e2. doi: 10.1016/j.cub.2019.06.079. [DOI] [PubMed] [Google Scholar]
- 46.Descamps C, Quinet M, Jacquemart A-L. Climate change-induced stress reduce quantity and alter composition of nectar and pollen from a bee-pollinated species (Borago officinalis, Boraginaceae) Front. Plant Sci. 2021;12:2264. doi: 10.3389/fpls.2021.755843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rering CC, Franco JG, Yeater KM, Mallinger RE. Drought stress alters floral volatiles and reduces floral rewards, pollinator activity, and seed set in a global plant. Ecosphere. 2020;11:e03254. doi: 10.1002/ecs2.3254. [DOI] [Google Scholar]
- 48.Alaux C, Ducloz F, Crauser D, Le Conte Y. Diet effects on honeybee immunocompetence. Biol. Lett. 2010;6:562–565. doi: 10.1098/rsbl.2009.0986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Roger N, Michez D, Wattiez R, Sheridan C, Vanderplanck M. Diet effects on bumblebee health. J. Insect Physiol. 2017;96:128–133. doi: 10.1016/j.jinsphys.2016.11.002. [DOI] [PubMed] [Google Scholar]
- 50.Burnham KP, Anderson DR. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach. Springer; 2002. [Google Scholar]
- 51.R Core Team. R A Lang. Environ. Stat. Comput. R Found. Stat.Comput. Vienna, Austria.http://www.R-project.org (2021).
- 52.Nakagawa S, Johnson PCD, Schielzeth H. The coefficient of determination R 2 and intra-class correlation coefficient from generalized linear mixed-effects models revisited and expanded. J. R. Soc. Interface. 2017;14:20170213. doi: 10.1098/rsif.2017.0213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Alger SA, Burnham PA, Boncristiani HF, Brody AK. RNA virus spillover from managed honeybees (Apis mellifera) to wild bumblebees (Bombus spp.) PLoS ONE. 2019;14:e0217822. doi: 10.1371/journal.pone.0217822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Singh R, et al. RNA viruses in hymenopteran pollinators: evidence of inter-taxa virus transmission via pollen and potential impact on non-apis hymenopteran species. PLoS ONE. 2010;5:e14357. doi: 10.1371/journal.pone.0014357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Burnham PA, et al. Flowers as dirty doorknobs: Deformed wing virus transmitted between Apis mellifera and Bombus impatiens through shared flowers. J. Appl. Ecol. 2021 doi: 10.1111/1365-2664.13962. [DOI] [Google Scholar]
- 56.Figueroa LL, et al. Landscape simplification shapes pathogen prevalence in plant-pollinator networks. Ecol. Lett. 2020;23:1212–1222. doi: 10.1111/ele.13521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Piot N, Smagghe G, Meeus I. Network centrality as an indicator for pollinator parasite transmission via flowers. Insects. 2020;11:872. doi: 10.3390/insects11120872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Truitt LL, McArt SH, Vaughn AH, Ellner SP. Trait-based modeling of multihost pathogen transmission: Plant–pollinator networks. Am. Nat. 2019;193:E149–E167. doi: 10.1086/702959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Dalmon A, et al. Possible spillover of pathogens between bee communities foraging on the same floral resource. Insects. 2021;12:122. doi: 10.3390/insects12020122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Rasmussen C, et al. Evaluating competition for forage plants between honey bees and wild bees in Denmark. PLoS ONE. 2021;16:e0250056. doi: 10.1371/journal.pone.0250056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Manley R, et al. Knock-on community impacts of a novel vector: Spillover of emerging DWV-B from Varroa-infested honeybees to wild bumblebees. Ecol. Lett. 2019 doi: 10.1111/ele.13323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Gusachenko ON, Woodford L, Balbirnie-Cumming K, Ryabov EV, Evans DJ. Evidence for and against deformed wing virus spillover from honey bees to bumble bees: A reverse genetic analysis. Sci. Rep. 2020;10:16847. doi: 10.1038/s41598-020-73809-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Manley R, Boots M, Wilfert L. Condition-dependent virulence of slow bee paralysis virus in Bombus terrestris: Are the impacts of honeybee viruses in wild pollinators underestimated? Oecologia. 2017;184:305–315. doi: 10.1007/s00442-017-3851-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Meeus I, de Miranda JR, de Graaf DC, Wäckers F, Smagghe G. Effect of oral infection with Kashmir bee virus and Israeli acute paralysis virus on bumblebee (Bombus terrestris) reproductive success. J. Invertebr. Pathol. 2014;121:64–69. doi: 10.1016/j.jip.2014.06.011. [DOI] [PubMed] [Google Scholar]
- 65.Wang H, Meeus I, Piot N, Smagghe G. Systemic Israeli acute paralysis virus (IAPV) infection in bumblebees (Bombus terrestris) through feeding and injection. J. Invertebr. Pathol. 2018;151:158–164. doi: 10.1016/j.jip.2017.11.015. [DOI] [PubMed] [Google Scholar]
- 66.Genersch E, Aubert M. Emerging and re-emerging viruses of the honey bee (Apis mellifera L.) Veterinary Res. 2010;41:54. doi: 10.1051/vetres/2010027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Martin SJ. The role of Varroa and viral pathogens in the collapse of honeybee colonies: A modelling approach. J. Appl. Ecol. 2001;38:1082–1093. doi: 10.1046/j.1365-2664.2001.00662.x. [DOI] [Google Scholar]
- 68.Carreck NL, Ball BV, Martin SJ. Honey bee colony collapse and changes in viral prevalence associated with Varroa destructor. J. Apic. Res. 2010;49:93–94. doi: 10.3896/IBRA.1.49.1.13. [DOI] [Google Scholar]
- 69.de Miranda J, Cordoni G, Budge G. The Acute bee paralysis virus–Kashmir bee virus–Israeli acute paralysis virus complex. J. Invertebr. Pathol. 2010;103:S30–S47. doi: 10.1016/j.jip.2009.06.014. [DOI] [PubMed] [Google Scholar]
- 70.de Pereira K, et al. Managed bumble bees acquire parasites from their foraging environment: A case study on parasite spillback. J. Invertebr. Pathol. 2021;182:107583. doi: 10.1016/j.jip.2021.107583. [DOI] [PubMed] [Google Scholar]
- 71.Jones JC. Honey bee nest thermoregulation: Diversity promotes stability. Science (80-). 2004;305:402–404. doi: 10.1126/science.1096340. [DOI] [PubMed] [Google Scholar]
- 72.Döke MA, Frazier M, Grozinger CM. Overwintering honey bees: Biology and management. Curr. Opin. Insect Sci. 2015;10:185–193. doi: 10.1016/j.cois.2015.05.014. [DOI] [PubMed] [Google Scholar]
- 73.Rasmont P, Iserbyt S. The Bumblebees Scarcity Syndrome: Are heat waves leading to local extinctions of bumblebees (Hymenoptera: Apidae: Bombus)? Ann. la Société Entomol. Fr. 2012;48:275–280. doi: 10.1080/00379271.2012.10697776. [DOI] [Google Scholar]
- 74.Soroye P, Newbold T, Kerr J. Climate change contributes to widespread declines among bumble bees across continents. Science (80-). 2020;367:685–688. doi: 10.1126/science.aax8591. [DOI] [PubMed] [Google Scholar]
- 75.Brunner FS, Schmid-Hempel P, Barribeau SM. Protein-poor diet reduces host-specific immune gene expression in Bombus terrestris. Proc. R. Soc. B Biol. Sci. 2014;281:20140128. doi: 10.1098/rspb.2014.0128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Dolezal AG, et al. Interacting stressors matter: Diet quality and virus infection in honeybee health. R. Soc. Open Sci. 2019;6:181803. doi: 10.1098/rsos.181803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Tehel A, Streicher T, Tragust S, Paxton RJ. Experimental infection of bumblebees with honeybee-associated viruses: No direct fitness costs but potential future threats to novel wild bee hosts. R. Soc. Open Sci. 2020;7:200480. doi: 10.1098/rsos.200480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.De Castro F, Bolker B. Mechanisms of disease-induced extinction. Ecol. Lett. 2004;8:117–126. doi: 10.1111/j.1461-0248.2004.00693.x. [DOI] [Google Scholar]
- 79.Swinton J, Harwood J, Grenfell BT, Gilligan CA. Persistence thresholds for phocine distemper virus infection in harbour seal Phoca vitulina metapopulations. J. Anim. Ecol. 1998;67:54–68. doi: 10.1046/j.1365-2656.1998.00176.x. [DOI] [Google Scholar]
- 80.Gallana M, Ryser-Degiorgis M-P, Wahli T, Segner H. Climate change and infectious diseases of wildlife: Altered interactions between pathogens, vectors and hosts. Curr. Zool. 2013;59:427–437. doi: 10.1093/czoolo/59.3.427. [DOI] [Google Scholar]
- 81.Zhan J, Ericson L, Burdon JJ. Climate change accelerates local disease extinction rates in a long-term wild host–pathogen association. Glob. Chang. Biol. 2018;24:3526–3536. doi: 10.1111/gcb.14111. [DOI] [PubMed] [Google Scholar]
- 82.Trenberth KE. Changes in precipitation with climate change. Clim. Res. 2011;47:123–138. doi: 10.3354/cr00953. [DOI] [Google Scholar]
- 83.Fischer EM, Knutti R. Anthropogenic contribution to global occurrence of heavy-precipitation and high-temperature extremes. Nat. Clim. Change. 2015;5:560–564. doi: 10.1038/nclimate2617. [DOI] [Google Scholar]
- 84.Goulson D, et al. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science. 2015;347:1255957. doi: 10.1126/science.1255957. [DOI] [PubMed] [Google Scholar]
- 85.Meeus I, Pisman M, Smagghe G, Piot N. Interaction effects of different drivers of wild bee decline and their influence on host–pathogen dynamics. Curr. Opin. Insect Sci. 2018;26:136–141. doi: 10.1016/j.cois.2018.02.007. [DOI] [PubMed] [Google Scholar]
- 86.Tentcheva D, et al. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004;70:7185–7191. doi: 10.1128/AEM.70.12.7185-7191.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.de Miranda JR, et al. Protocols for assessing the distribution of pathogens in individual Hymenopteran pollinators. Protoc. Exch. 2021 doi: 10.21203/rs.3.pex-1453/v1. [DOI] [Google Scholar]
- 88.Potts SG, et al. Declines of managed honey bees and beekeepers in Europe. J. Apic. Res. 2010;49(15):22. [Google Scholar]
- 89.Moritz RFA, Kraus FB, Kryger P, Crewe RM. The size of wild honeybee populations (Apis mellifera) and its implications for the conservation of honeybees. J. Insect Conserv. 2007;11:391–397. doi: 10.1007/s10841-006-9054-5. [DOI] [Google Scholar]
- 90.Bivand, R., Keitt, T. & Rowlingson, B. rgdal: Bindings for the ‘geospatial’ data abstraction library. (2019).
- 91.Hijmans, R. J. raster: Geographic data analysis and modeling. (2019).
- 92.Pebesma, E. J. & Bivand, R. S. Classes and methods for spatial data in R. (2005).
- 93.Wickham, H., François, R., Henry, L. & Müller, K. dplyr: A grammar of data manipulation. (2020).
- 94.Harrell, F. E. Hmisc: Harrell miscellaneous. (2020).
- 95.Brooks ME, et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017;9:378–400. doi: 10.32614/RJ-2017-066. [DOI] [Google Scholar]
- 96.Barton, K. MuMIn: Multi-model inference. (2020).
- 97.Wickham H. ggplot2: Elegant Graphics for Data Analysis. Springer Verlag; 2016. [Google Scholar]
- 98.Fox J, Weisberg S. An R Companion to Applied Regression. Sage publishing; 2019. [Google Scholar]
- 99.Karger DN, et al. Climatologies at high resolution for the earth’s land surface areas. Sci. Data. 2017 doi: 10.1038/sdata.2017.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Karger DN, et al. Data from: Climatologies at high resolution for the earth’s land surface areas. Dryad Digit. Repos. 2018 doi: 10.5061/dryad.kd1d4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Burnham KP, Anderson DR, Huyvaert KP. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011;65:23–35. doi: 10.1007/s00265-010-1029-6. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The dataset is included as a supplementary file.