

Abstract
Decades of research in attention have shown that salient distractors (e.g., a color singleton) tend to capture attention. However, in most studies, singleton distractors are just as likely to be present as absent. We therefore have little knowledge of how probabilistic expectations of the salient distractor’s occurrence and features affect suppression. In three experiments, we explored this question by manipulating the frequency of a singleton distractor and the variability of its color within a search display. We found that increased expectations regarding the occurrence of the singleton distractor eliminated the singleton RT cost and reduced the number of first saccades to the singleton. In contrast, expectations regarding variability in the singleton color did not affect singleton capture. This was surprising and suggests the ability to suppress second order salience over and above that of first order features. We next inserted the probe display that included a to-be-reported letter inside each shape between search trials to measure if attention went to multiple objects. The letter in the singleton location was reported less often in the high frequency condition, suggesting proactive suppression of expected singleton. Additionally, we found that trial-to-trial repetitions of a singleton (irrespective of its color and location) facilitated performance (i.e., singleton repetition priming), but repetitions of its specific color or location did not. Together our findings demonstrate that attentional capture by a color singleton distractor is attenuated by probabilistic expectations of its occurrence, but not of its color and location.
Keywords: attentional capture, distractor suppression, visual search, probabilistic expectation, repetition priming
Introduction
In visually complex environments, the role of visual attention is to selectively guide sensory processing towards goal relevant information and to avoid attentional perseveration on task-irrelevant distractions (Folk, Remington, & Johnston, 1992; Folk & Remington, 1998; Folk & Remington, 2008; Wolfe, Cave, & Franzel, 1989; Egeth & Yantis, 1997; Bacon & Egeth, 1994; Folk, Leber, & Egeth, 2002). Thus, the success of information selection depends on the dual mechanisms of target selection and distractor suppression (Chun & Marois, 2002). The mechanisms of target selection have been well-studied and shown to depend on how the target is defined. For example, selective enhancements occur for specific target features (i.e., feature-based selection; Desimone & Duncan, 1995; Duncan, Humphreys, & Ward, 1997; Martinez-Trujillo & Treue, 2004; Maunsell & Treue, 2006) or second order properties such as the target being a shape singleton (i.e., singleton detection mode; Bacon & Egeth, 1994; Balan & Gottlieb, 2006; Proulx & Serences, 2006). However, the mechanisms of distractor suppression are less well understood.
One reason for this may be that there is still disagreement about what defines the saliency of a non-target distractor and whether salient distractors necessarily capture attention. Even within widely used paradigms, such as the additional singleton paradigm (Theeuwes, 1992) in which targets are defined by a unique shape (e.g., circle amongst squares) and salient distractors are defined by a unique color (e.g., red amongst green objects), different manipulations have led to opposite conclusions. For example, Bacon and Egeth (1994) found singleton distractors no longer captured attention when search for the target switched from a “unique” shape (i.e., singleton-detection mode), to a specific feature such as diamond shape (i.e., engaging feature search mode). They concluded that the distraction was based not on distractor properties, but on the target search strategy, or “attentional set” (Bacon & Egeth, 1994; Folk, Remington, et al., 1992; Lamy & Egeth, 2003; Leber & Egeth, 2006). However, Theeuwes (2004, 2010) argued in response that the singleton distractor does automatically capture attention, and the first saccadic eye-movement, under feature search mode as long as spatial attention is broad and the target can be detected without the need for narrow serial search (Theeuwes, Kramer, Hahn, & Irwin, 1998; Born, Kerzel, & Theeuwes, 2011; Theeuwes, 1991, 1992, 2013; Theeuwes, Kramer, Hahn, et al., 1998; Franconeri & Simons, 2003; Yantis & Jonides, 1984; Lavie, 1995; Lavie, Ro, & Russell, 2003). These studies have led to a rich debate regarding the automaticity of attentional capture by stimulus saliency, but they have primarily pitted attentional goals based on the target search strategy against perceptual saliency (e.g., feature uniqueness) that cannot be predicted. However, more recent work has shown that probabilistic expectations about distractors can reduce, or in some cases even eliminate, behavioral costs associated with attentional capture when all task and stimulus properties are held constant. Such expectations can be explicit, analogous to attentional facilitation by target cueing (Posner, 1980). For example, Moher, Abrams, Egeth, Yantis, and Stuphorn (2011), used a singleton search paradigm in which subjects were asked to make a first saccade to a uniquely shaped (circle or square) green target amongst uniformly shaped (opposite shape to the target) green distractors; critically, prior to each search display, a probabilistic cue (“90%” or “10%”) appeared and indicated the likelihood of one distractor being a red singleton in the upcoming display. Despite the fact that the stimulus properties were identical across probability conditions, the red singleton distractor was more likely to capture the first saccade following the “10%” cue than the “90%” cue. This suggests that the strength of distractor suppression was modulated by explicitly formed expectations. Similar enhancements in distractor suppression have been found in response to an explicit spatial cue of an upcoming distractor’s location or cues of a distractor feature (Arita, Carlisle, & Woodman, 2012; Moher & Egeth, 2012, Cunningham & Egeth, 2016, Moher et al., 2011; van Diepen, Miller, Mazaheri, & Geng, 2016).
In addition to explicit cueing, expectations shaped by short- and long- term statistical experiences that are known to guide selective attention for targets (Maljkovic & Nakayama, 1994, 1996; Geng & Behrman, 2002, 2005; Jiang, Swallow, Rosenbaum, & Herzig, 2013; Kristjansson & Campana, 2010; Failing & Theeuwes, 2014; 2017), and have been more recently found to also guide distractor suppression (Ferrante et al., 2017, Geyer, Müller, & Krummenacher, 2008; Wang & Theeuwes, 2018a, 2018b). For example, Geyer et al. (2008) asked participants to find a green circle among eight green diamonds. On some trials, one diamond was colored red (i.e., color singleton). They manipulated the frequency of trials with the color singleton (20%, 50%, and 80%) and found that the size of attentional capture reduced as the probability of singleton increased. In the 80% block, attentional capture was completely eliminated. A subsequent study (Muller, Geyer, Zehetleitner, & Krummenacher, 2009), also used a high probability of color singleton distractors, but introduced a “rare” singleton color in addition to the frequent singleton color. Even though the frequent singleton color was again entirely suppressed, the novel singleton color produced a performance cost, albeit a smaller one than when all singletons were rare. Vatterot and Vecera (2012) found similar results in a feature-search paradigm when new color singletons were introduced after a period of color stability using only 50% singleton-present displays. From these studies, it appears that singleton suppression is sensitive to the likelihood of occurrence and involves the suppression of specific features rather than the suppression of second order “salience” (Moher et al., 2011; Theeuwes, 2004; Gaspelin et al., 2017; Gaspelin & Luck, 2017, 2018).
More recently, Vatterott, Mozer, and Vecera (2018) examined the generalizability of suppression to singletons of varying colors. Subjects searched for a circle-shaped target amongst distractors of the same color but that varied in shape; color singletons appeared on 50% of trials. In contrast to previous studies, there was one condition in which singletons were all one color and another in which four (or six) singleton colors alternated. Both conditions were followed by blocks with new singleton colors. They found that exposure to one singleton color early on resulted in a performance cost (i.e., RT increase) when a new color singleton appeared, but exposure to four (or six) singleton colors led to the generalization of suppression to new colors – there was no increase in attentional capture by the new singleton colors when they were introduced. Thus, the appearance of a never-before-seen singleton captured attention when subjects were expecting consistent singleton colors, but was suppressed when the observer had been exposed to color variability (Horstmann, 2002, 2005; Baldi & Itti, 2010). Interestingly, Vatterot et al. (2018) did not argue that the generalization was due to suppression of second order salience, but rather due to a combination of slow decaying memory traces for each singleton color and generalization of that suppression to feature-similar colors. Their conclusions converge well with a recent finding in which exposure to a consistent group of distractors during visual search “training” led to generalized suppression to broadly similar new “test” distractors, but not dissimilar distractors (Won & Geng, 2018).
Together, these previous results suggest that distractor suppression may be more flexible than once thought and highly sensitive to probabilistic expectations. But they leave open the question of whether feature generalization is sensitive to frequency manipulations based on repeated exposure to specific exemplars only, or if it can also extend to the second order representations of “salience” (Moher et al., 2012, Gaspelin & Luck, 2017, 2018). In the current studies, we explore this question by simultaneously manipulating expectations about the singleton distractor’s frequency of occurrence and its variability of color using the additional singleton paradigm; importantly, the target is always a shape-singleton, which precludes the use of feature-search mode and makes it more difficult to ignore color singleton distractors (Bacon & Egeth, 1994). We hypothesized that expectations of the singleton’s occurrence and color contribute independently to distractor suppression and therefore performance should be best when the singleton distractor is consistently one color and appears frequently; and performance should be worst when the singleton distractor is rare and of varying colors. However, if it is possible to suppress singletons based on second order salience and not just direct experience with specific feature, then performance will be equal when singleton colors are fixed and mixed, while still being modulated by frequency of occurrence.
Experiment 1
In Experiment 1, we test the influence of a singleton distractor’s frequency of occurrence and color variability on attentional capture. The frequency of singletons was manipulated in the high frequency condition (singleton on 80% of trials) and the low frequency condition (singleton on 20% of trials). Variability in singleton color was manipulated in a mixed color condition (randomly colored singletons) and a fixed color condition (only one singleton color shown). We hypothesize that the predictability of the singleton distractor’s occurrence and color would increase the ease with which it is suppressed. However, it is also possible that feature expectations can be based on second order salience rather than specific features, in which case, color variability may have limited impact on performance while preserving the effect of frequency.
Method
Participants.
Twenty undergraduates from UC Davis (mean age=21.35 years, SD=2.3, range=18–27; 8 males; 4 left-handed) participated for course credit. The primary a priori analyses of interest involve the effects of singleton frequency and color variability. However, the most similar prior studies using probability manipulations of singleton frequency did not report effect sizes (Geyer et al., 2008; Moher et al., 2011). We therefore used a combination of related studies to estimate sample size. First, Wang and Theeuwes (2018) used a spatial probability manipulation of singleton distractors and reported a partial eta-squared of .74 using a sample size of 20, resulting in estimated power of .98. Second, Gaspelin and Luck (2017, Experiment 1) found a Cohen’s d = .789 for the “singleton presence cost” using a sample size of 24, resulting in power of .95. We therefore used a sample size of twenty as a conservative estimate. Consistent with this expectation, the calculated effect size from the most similar contrast in our current study resulted in Cohen’s d = 1.107. All provided informed consent in accordance with NIH guidelines provided through the UCD Institutional Review Board.
Stimuli and apparatus.
Each participant sat in a sound-attenuated, dimly lit room, 60 cm from the monitor. Stimuli were displayed on a 27-in. Asus LCD monitor (2560 × 1440 pixels), with a refresh rate of 60 Hz. Stimuli were generated using (www.mathworks.com), with Psychtoolbox extensions (Brainard, 1997; Pelli, 1997). Gaze position of the right eye was monitored using an EyeLink 1000 desktop mount eye-tracker sampling at 500 Hz (SR Research, Mississauga, Ontario, Canada). Search displays contained six shapes drawn in gray (36.3 cd/m2), five diamonds, (1.1° ×1.1°) and one circle (.9° in diameter) or one diamond and five circles on a black background (.21 cd/m2). The eccentricity (center of each item to the center of screen) was 4°. Each shape contained a black line subtending .05° × .3° that was randomly tilted 45° to the left or right. The target shape appeared equally often in six positions. Importantly, on some trials, one distractor was drawn in color, i.e., the “color singleton”. Note that hereafter, “singleton” refers a uniquely colored distractor object (cf. the target defined shape-singleton).
Singleton colors were based on two sets of four colors (adopted from Experiment 4 in Gaspelin & Luck, 2017). The first set was orange (22.5 cd/m2), pink (21.0 cd/m2), green (22.5 cd/m2), and blue (22.3 cd/m2). The second set was teal (23.1 cd/m2), gold (22.2 cd/m2), red (21.5 cd/m2), and purple (22.5 cd/m2). The colors within a set were chosen to be maximally different from each other in hue. The color set assignment was counterbalanced across subjects. Participants were encouraged to ignore the color singleton. The singleton location was equally, but randomly chosen to be next to the target, one distractor away from the target, or the opposite to the target. A white fixation cross (.4°x.4°) appeared before each search display.
Design and procedure.
The experiment was composed of four main experimental blocks corresponding to the singleton frequency (low, high) x singleton color (mixed, fixed) experimental design. In the “low–mixed” block, the singleton appeared on 20% of trial and varied color randomly between three colors trial-by trial; in the “low–fixed” block, the singleton appeared on 20% of trials and was always only one fixed color; in the “high–mixed” block, the singleton appeared on 80% of trials and varied color randomly between three colors trial-by trial; and finally, in the “high–fixed” block, the singleton appeared on 80% of trials, but was always one fixed color. The order of these four blocks was randomized across participants.
Each block began with a 13-point eye-tracker calibration procedure. Each trial began with a fixation cross at the center of the screen, which participants were asked to fixate. When participants continuously fixated on the cross at least 150-ms, search display appeared after 350-ms fixation display. We used this gaze-contingent display to force the participants to start search from the center. Once the search display appeared, participants searched for the target, defined as the unique shape, called “shape singleton”. The target was either a circle among five identical diamonds or a diamond among five identical circles. The task was to report whether the bar inside the target shape was tilted to the left or right by pressing “z” or “x” for those who were left-handed, or by pressing “.” or “?” for those who were right-handed. Upon response or after 5-sec without any responses, the display was removed and auditory and visual feedback given (Figure 1). Auditory feedback was composed of an up-chirp tone for correct responses and a low-pitch tone for incorrect responses; visual feedback consisted of a gray fixation for correct responses and white fixation plus a 2-sec delay for incorrect responses. Participants completed 60 trials in each of the four experimental blocks. An additional mini block of 30 search trials that did not have any singleton distractors occurred between experimental blocks in order to reduce carry-over expectations regarding the singleton.
Figure 1.
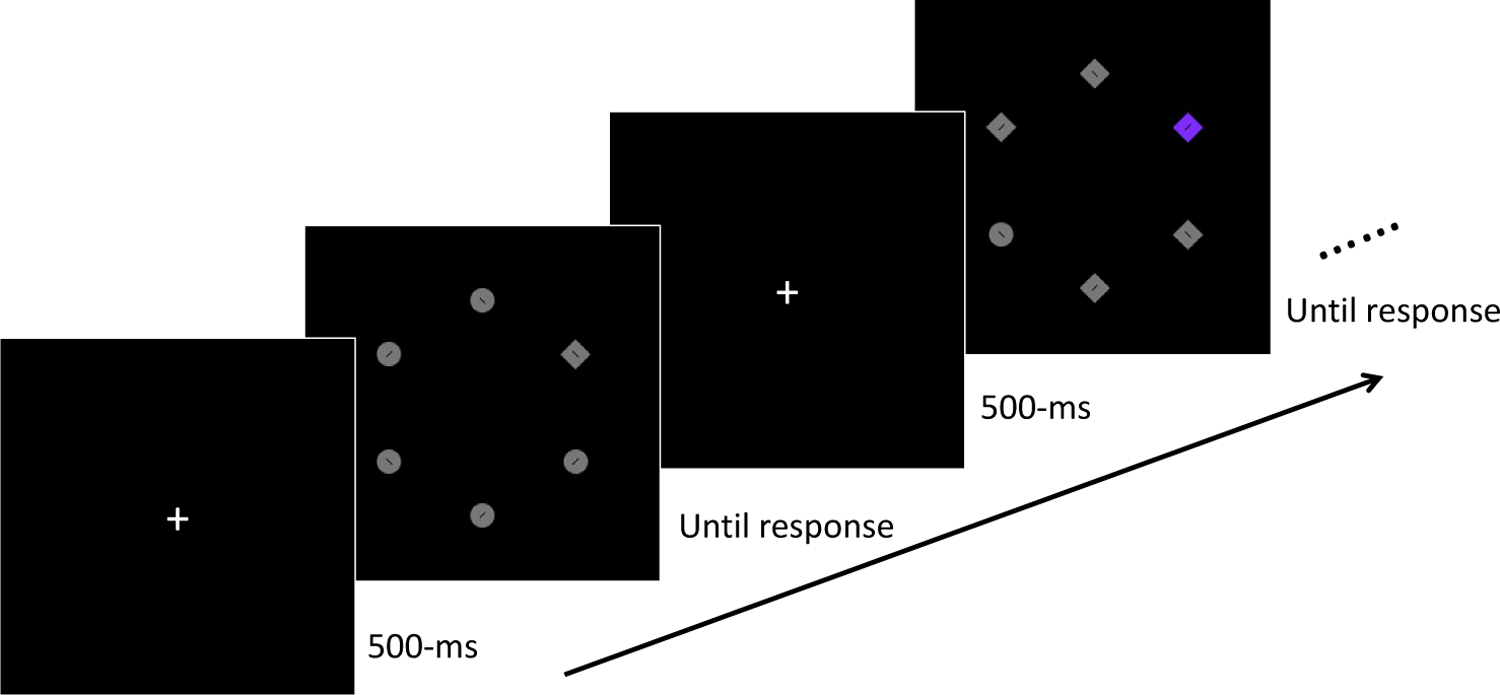
Experiment 1 trial procedure. Participants were asked to search for a uniquely shaped target, and report the orientation of bar inside (left or right). A colored distractor (i.e., singleton) appeared on some trials (in this example, on the second trial).
Analyses.
The destination of the first saccade on each trial was classified as being in one of seven interest areas (IA; 2°× 2° each). Each IA was centered over each of the six items and the fixation point. Saccade end points were classified as being to the target, the singleton, or any non-singleton item based on its the nearest IA. Trials were excluded from analysis if any of the following were true: there was no eye movement (.6%), the first fixation landed more than 1.75° from the edge of the nearest IA (3.1%), or the response was inaccurate or the RT was longer than 3SD above the block mean (3.9%). This resulted in the exclusion of 7.5% of the data. The p value of post hoc comparisons following analyses of variance (ANOVAs) were Greenhouse–Geisser corrected. We also provide the Bayes factor corresponding to the t test (Rouder, Speckman, Sun, Morey, & Iverson, 2009). Bayes factors quantify the relative likelihood of obtaining the observed data under the null hypothesis compared to the alternative hypothesis and have the advantage of being equally well suited to quantify the evidence for and against the null hypothesis. Evidence in favor of the null hypothesis is denoted as BF01 and in favor of the alternative hypothesis as BF10. Overall accuracy was high (mean: 97.2%; SD: 2%), therefore we only focused RT in subsequent analyses. All raw data are publicly available in https://osf.io/v8ct4/?view_only=e810dc0301d941c3b32502d828b0f2ce.
Results and discussion
Visual search RT.
The primary purpose of this experiment was to determine whether attentional capture by the singleton distractor would decrease with increasing frequency and color stability. We conducted a three-way ANOVA with singleton frequency (low, high), singleton color (mixed, fixed), and singleton presence (present, absent) (Figure 2). Search RTs were overall longer for the mixed color condition than fixed color condition (difference: 55-msec), F(1, 19) = 5.50, p = .030, η2 = .22, and for the singleton present condition than the singleton absent condition (difference: 121-msec), F(1, 19) = 66.96, p < .001, η2 = .78. Search RTs for the two singleton frequency conditions did not differ (13-msec), F < 1. Importantly, there was a significant two-way interaction between singleton frequency and singleton presence, F(1, 19) = 64.50, p < .001, η2 = .77. Other interactions were not significant (smallest p = .345).
Figure 2.
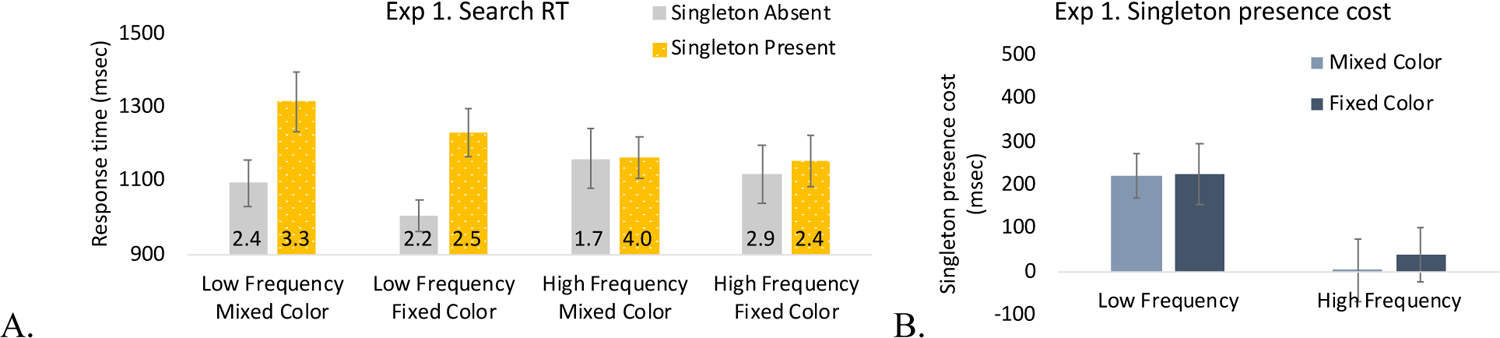
Results from Experiment 1. A. Search RTs. The number in each bar indicates the error rate (%). B. Singleton presence costs (calculated by RT singleton present trials minus singleton absent trials) by singleton frequency and color variability. All error bars shown here and in subsequent figures represent within-subject 95% confidence intervals (Loftus & Masson, 1994).
Post-hoc pairwise t-tests indicated that the significant two-way interaction was due to a significant singleton presence cost (i.e., longer RT for singleton present than absent trials) in the low frequency block, t(19) = 11.25, p < .001, Cohen’s d = 2.52, Log(BF10) = 16.43, but no singleton presence cost in the high frequency block, t(19) = 1.07, p = .30, Cohen’s d = .24, BF01 = 2.61 (Figure 2)1. Note that in the low frequency condition, the BF shows that the data is 1.370e+7 times more likely to be observed if the alternative hypothesis were true than if the null hypothesis were true, whereas in the high frequency condition the data was 2.61 times more likely to be observed if the null hypothesis were true than if the alternative hypothesis were true (Figure 2). Notably, the lack of an interaction between singleton color and singleton presence (F < 1) suggests that having mixed (compared to fixed) singleton colors did not selectively modulate the strength of attentional capture by the singleton. Thus, surprisingly, expectations of the specific singleton color did not improve singleton suppression, but stronger expectations about the immediate appearance of any singleton did.
Repetition priming.
Because frequency effects are often intertwined with repetition priming, we next compared search RTs on each trial (N) as a function of the previous trial (N-1) for singleton repetitions and color repetitions. In order to maximize the number of trials included for analysis, the data from different blocks were collapsed (the average number of singleton repetition trials was 76; the average number of singleton non-repetition trials was 37), although, most trials will inevitably come from high frequency blocks. Consistent with the finding that RTs were shorter in the high frequency blocks compared to low frequency blocks, there was a significant color-nonspecific singleton repetition priming effect, t(19) = 2.86, p = .01, Cohen’s d = .64, BF10 = 5.14. When a singleton was present on a second consecutive trial (irrespective of color), RTs were shorter. However, the RT benefit from a singleton repetition (~55-ms) was less than the overall benefit of the high frequency manipulation (~200-ms). This suggests that the probabilistic expectations operated over and above that of single-trial repetition priming alone2.
In contrast, there was no evidence for repetition priming based on the singleton color, (the average number of color repetition trials was 49.9; the average number of color non-repetition trials was 26.1), t < 1, BF01 = 4.27, which is consistent with the lack of RT differences between mixed and fixed color blocks (Figure 3). In addition to these effects, we further tested repetitions of the singleton location. Interestingly, there was also no evidence for significant singleton location priming (the average number of location repetition trials was 13.2; the average number of location non-repetition trials was 62.9), t < 1, BF01 = 2.983. In sum, there was robust repetition priming on trials with consecutive singletons, but there was no priming based on the singleton color or location. The finding that distractor singleton repetition suppression can be location- and color-general, suggests the mechanisms of distractor suppression may operate directly on the second order property of being a “singleton” stimulus.
Figure 3.
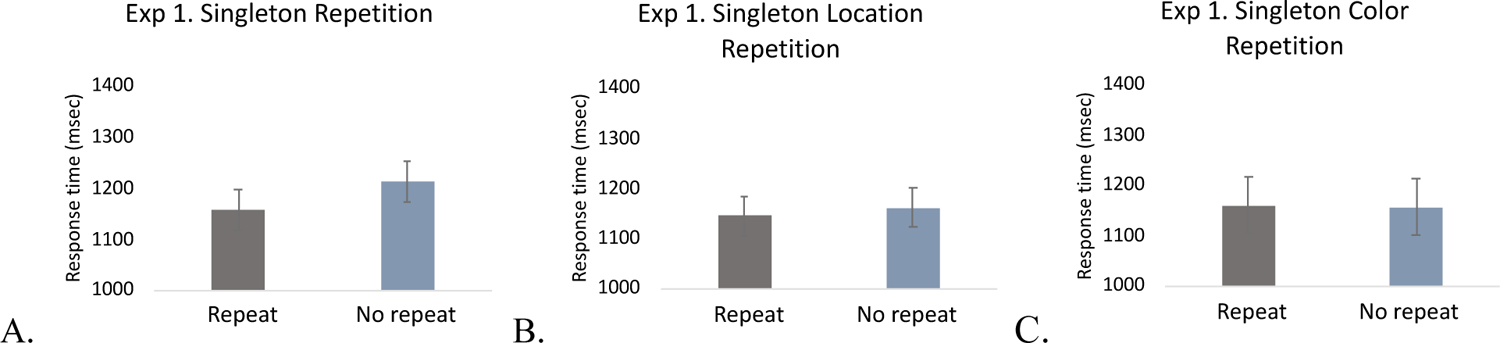
Singleton repetition priming in Experiment 1. A. Singleton presence, B. Singleton location, and C. Singleton color repetition priming. The error bars represent within-subject 95% confidence intervals.
First saccades.
The RT results demonstrated a greater singleton presence cost in the low frequency condition, but this could be due to either greater attentional capture by the singleton or a greater filtering cost (Becker, 2007; Folk & Remington, 1998) – a preattentive delay in the allocation of spatial attention as the system resolves the competition between the two “pop-out” objects (i.e., the target and singleton, DiQuattro & Geng, 2017). We therefore used eye-movements as an indicator of spatial attention to test between these two hypotheses. If the greater singleton presence cost in the low frequency condition was due to attentional capture, we would expect more first saccades to the singleton distractor in the low frequency condition than the high frequency condition. Further, since the singleton presence cost was equivalent between the mixed and fixed color conditions, we expected no differences in saccade behavior in the fixed and mixed color conditions. However, if the singleton presence cost is due to an overall filtering cost, we would expect no differences in first saccades between any conditions.
The proportion of trials with a first saccade to the singleton (compared to the target or non-singleton distractors) was entered into a frequency (low, high) x color (mixed, fixed) repeated measures ANOVA. The results showed that the first saccade was more likely to be directed toward the singleton location in the low frequency condition than high frequency condition (difference: 14.9%), F(1, 19) = 21.66, p < .001, η2 = .53, but there was no difference between the two color conditions (difference: 3.9%), F < 1. The frequency x color interaction was also not significant, F(1, 19) = 1.25, p = .28, η2 = .06, despite apparent trends. Overall, the eye-data are consistent with the RT results and demonstrate that the frequency of the singleton had a large impact on the likelihood of attentional capture, but the variability of the singleton color did not (Figure 4).
Figure 4.
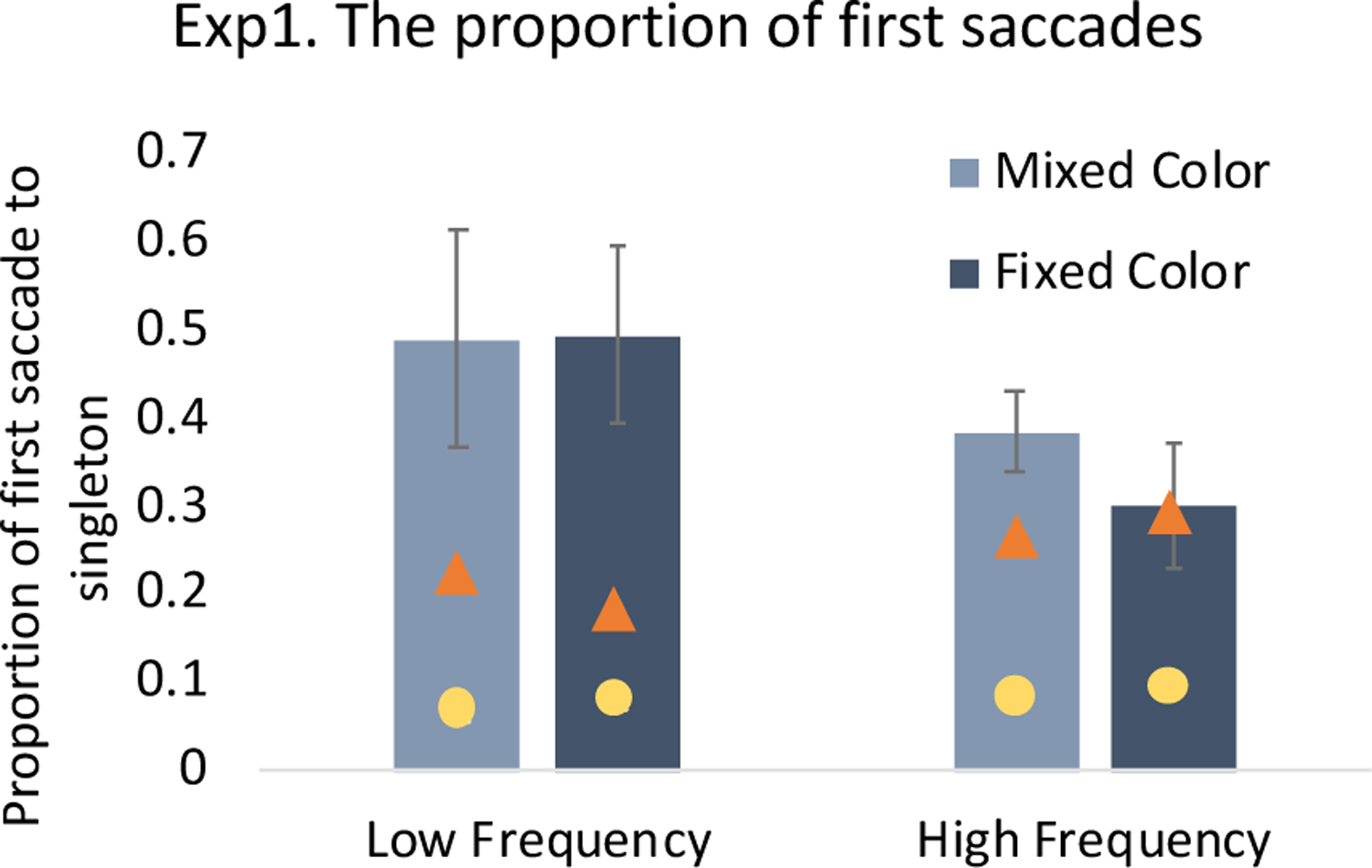
Proportion of trials with first saccades to the color singleton in Experiment 1. The triangle indicates the proportion of trials with first saccades to the target and the circle indicates the proportion of trials with first saccades to a non-singleton distractor. The error bars represent within-subject 95% confidence intervals.
Experiment 2
In Experiment 1, there was no effect of singleton color variability on singleton suppression (i.e., no difference in RT or first saccades on blocks of trials with only one fixed color singleton vs. three color singletons). However, it is possible that three colors were sufficiently few for participants to build feature-specific distractor templates to suppress each singleton (Vatterot et al., 2018; Won & Geng, 2018). Thus, color variability in Experiment 1 may not play a significant role in attentional capture, not because variability doesn’t matter, but because we failed to introduce sufficient variability. In Experiment 2, we rule out this alternative explanation by increasing the number of singleton colors to 192 different colors in the mixed singleton color condition. If the lack of singleton variability in Experiment 1 is due to the mixed condition containing too few colors, we should now observe a significant influence of singleton color variability in attentional capture.
Methods
Participants.
Twenty undergraduates from UC Davis (mean age=19.6 years, SD=1.6, range=18–24; 4 males; 1 left-handed) participated for course credit. This sample size was selected a priori to match the sample size of Experiment 1. All provided informed consent in accordance with NIH guidelines provided through the UCD Institutional Review Board.
Stimuli & apparatus.
The stimuli were identical to Experiment 1 except for the following changes: The fixed singleton color condition only had one singleton color per block and the mixed singleton condition had 48 different singleton colors per block. 196 colors (4 colors for the 4 fixed singleton color blocks and 192 colors for 4 mixed color blocks) were randomly chosen from 360 CIE Lab color wheel (radius = 39, luminance = 70, a* = 0, b* = 0, Figure 5A). Other aspects were the same as Experiment 1.
Figure 5.
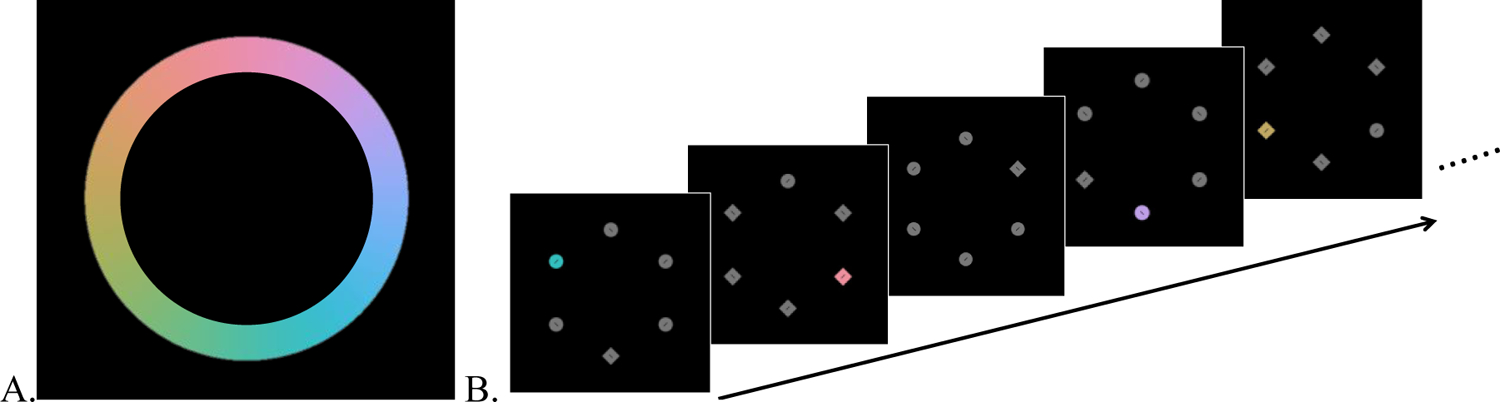
A. Color wheel from which singleton colors were selected. 196 colors were randomly chosen for use as color singletons. B. Examples of search displays. Inter-trial fixation displays are not included, for illustrative clarity.
Design & procedure.
The procedure was identical with Experiment 1 except that the singleton frequency was always 80% and for both the fixed and mixed singleton color blocks (Figure 5B). There were eight blocks (60 trials per block) composed from four mixed color blocks and four fixed color blocks. The order of two types of blocks was randomized.
Analyses.
Eye movement and RT data were analyzed identically to Experiment 1. Trials with no eye movement data (.2%), trials with a fixation landing more than 1.75° from the nearest IA borders (2.8%), and trials with inaccurate and RTs longer than 3SD above each individual’s mean (4.6%) were excluded from analyses. Post hoc analyses based on results from the analyses of variance (ANOVAs) report Greenhouse–Geisser corrected p value. Overall search accuracy was high (mean: 96.9%; SD: 1.9) and therefore only RT data were included in subsequent analyses.
Results and discussion
Visual search RT.
A singleton color (mixed, fixed) x singleton presence (present, absent) ANOVA revealed longer RTs on singleton present trials than singleton absent trials (difference: 32-msec), F(1, 19) = 10.72, p = .004, η2 = .36, but there was no significant difference between the mixed and fixed color conditions (BF01 = 3.22), nor was there a significant interaction, Fs < 1. Unlike Experiment 1, there was no overall slowing in the mixed compared to fixed blocks. However, the BFs were relatively weak (BF10 = 1.80 in Experiment 1) in both cases rendering interpretation of the apparent discrepancy difficult (Figure 6). Most importantly, the data for the mixed and fixed conditions were more 3.22 times more likely if the null hypothesis is true than the alternative, suggesting that singleton suppression was identical regardless of whether the singleton was always a fixed color or if it varied between 192 colors.
Figure 6.
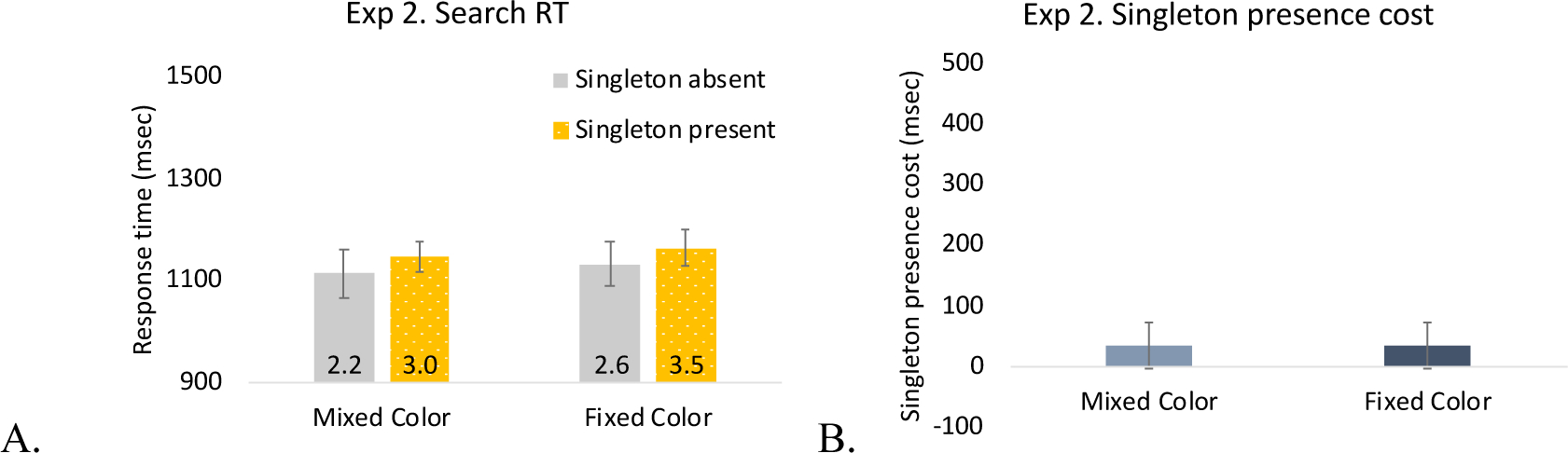
Results of Experiment 2. A. Search RT. The number in each bar indicates the error rate (%). B. Singleton-presence cost. The error bars represent within-subject 95% confidence intervals.
Repetition priming.
We again tested repetition priming for the singleton color, and found very similar results with Experiment 1. As in Experiment 1, there was a significant singleton repetition priming effect such that RTs were shorter when the singleton appeared on a second consecutive trial than when it did not appear in the previous trial (the average number of singleton repetition trials was 286.7; the average number of singleton non-repetition trials was 73.8), t(19) = 2.65, p = .016, Cohen’s d = .59, BF10 = 3.524. However, there was no repetition effect of singleton location (the average number of location repetition trials was 48.7; the average number of location non-repetition trials was 238), t(19) = 1.262, p = .22, Cohen’s d = .28, BF01 = 2.15. Similarly, there was no priming effect associated with the specific color (the average number of color repetition trials was 143; the average number of color non-repetition trials was 143.7), t(19) = 1.08, p = .294. Cohen’s d = .24, BF01 = 2.58 (Figure 7). These results replicate those from Experiment 1 in showing that singleton repetition priming effect was applied to the singleton stimulus, not its specific color or location suggesting that it was the suppression that was primed, not processing of specific stimulus characteristics.
Figure 7.
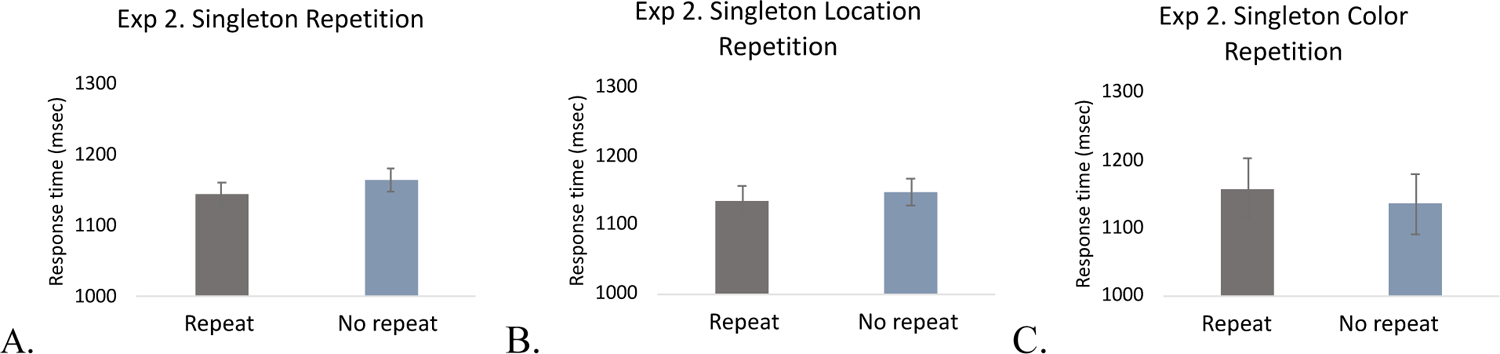
Singleton repetition priming in Experiment 2. A. Singleton presence, B. Singleton location, and C. Singleton color repetition priming. The error bars represent within-subject 95% confidence intervals.
First saccade.
The proportion of trials with first saccades to the singleton in the mixed color condition (30.4%) did not differ from that in the fixed color condition (29.0%), t(19) = 1.00, p = .328. Cohen’s d = .225, BF01 = 2.76. This replicates the results from Experiment 1 and suggests that expectations regarding color variability do not modulate attentional capture (Figure 8). Note that the rate of capture by the singleton in this experiment is similar to that in Experiment 1 (mixed block: 38.2% and fixed block: 30.1%).
Figure 8.

Proportion of trials having with first saccades to the color singleton in in Experiment 2. The triangle indicates the proportion of trials with first saccades to the target and the circle indicates the proportion of trials with first saccades to a non-singleton distractor. The error bars represent within-subject 95% confidence intervals.
Experiment 3
The results from Experiment 1 and 2 suggest that high frequency singletons are well suppressed. However, it remains unclear what was the source of the better suppression. One possibility is that the frequency manipulation led to differential engagement of proactive vs. reactive suppression mechanisms. The distinction between proactive and reactive mechanisms were originally proposed in the cognitive control literature (Braver, 2012; Theeuwes, Atchley, & Kramer, 2000), but also apply to attentional control mechanisms (Marini et al., 2016; Mazaheri, DiQuattro, Bengson, & Geng, 2011; for review, Geng, 2014). In these studies, stronger probabilistic expectations may have led to greater use of proactive suppression mechanisms that prevent attentional capture by the singleton in the first place. But because proactive suppression is more effortful (Braver, 2012), it may be sufficient to rely on reactive mechanisms when singletons are rare. In Experiment 3, we test whether participants adopt different suppression strategies based on singleton frequency by using the capture-probe paradigm (Gaspelin et al., 2015, Gaspelin & Luck, 2017). If a proactive suppression mechanism is engaged, subjects should be unable to report the letter from the probe at the location of the singleton, but if reactive mechanisms are engaged, the letter at the singleton location should be reported even if the target letter is also reported.
Method
Participants.
Twenty undergraduates from UC Davis (mean age=21.0 years, SD=1.4, range=19–24; 10 males; 1 left-handed) participated for course credit. This sample size was selected a priori to match the sample size of Experiment 1 and Experiment 2. All provided informed consent in accordance with NIH guidelines provided through the UCD Institutional Review Board.
Stimuli & apparatus.
Stimulus displays were identical to those of Experiment 1. Each of the four colors from one color set was randomly assigned as the singleton color in four different blocks. The color set assignment was counterbalanced across subjects (see method in Experiment 1 in detail). Importantly, this experiment includes new probe sequences composed of a stimulus display followed by a response display. The first display was composed of the same objects as the search display, but instead of oriented bars within each object, there was an upper-case white alphanumeric letter (.4° tall). This was immediately followed by a response display with the English alphabet in white. Subjects clicked on all the letters that they saw in the previous display (up to six letters). A click on the “ok” button terminated the trial. Probe displays always had one color singleton.
Design & procedure.
The procedure was identical to Experiment 1 except for the following changes: The frequency of singleton occurrence (low, high) was manipulated across four blocks - two low frequency blocks and two high frequency blocks. The order of these blocks was randomized. The singleton colors were always “fixed”, but given the results from the previous two Experiments, we would expect the results to be the same for “mixed” singleton color conditions. Most importantly, on some trials, a probe sequence occured (see above). On “early probe” trials, the probe display replaced the search trial entirely, lasted for 200-ms, and was followed by the response display; on “late probe” trials, the search display appeared for 200-ms before being replaced by the probe display for 200-ms, after which the response display appeared.
On all probe trials, participants were asked to report as many letters as they could remember from the probe display by clicking on letters (up to six letters) on the response display. The total number for trials for low frequency condition was 216 trials that consisted of 144 singleton absent trials, 24 singleton present trials, 24 early probe trials, and 24 late probe trials. For the high-frequency condition, there were also 216 trials that consisted of 48 singleton absent trials, 120 singleton present trials, 24 early probe trials, and 24 late probe trials (Figure 9).
Figure 9.
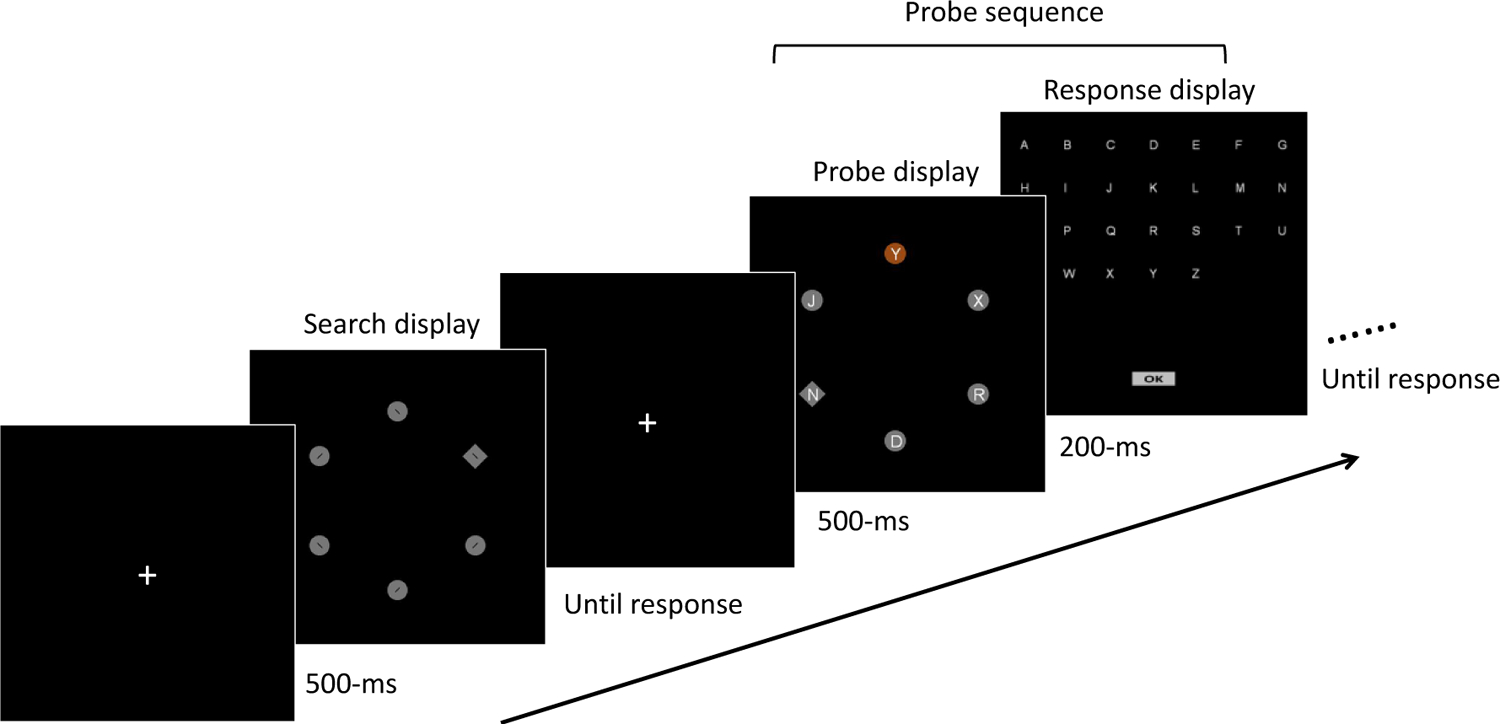
The trial procedure in Experiment 3. The procedure is the same as previous experiments, but now includes probe sequence trials. In this example, the second trial is a probe trial (i.e., a probe display followed by a response display). On response displays, participants clicked all the letters that they remembered from the probe display.
RT, Eye movement, and probe analyses.
As in previous experiments, we excluded inaccurate trials and trials with the fixation landing more than 1.75° from the nearest IA border (2.9%), trials with no eye movement data (.1%), and incorrect trials or ones with RTs longer than 3SD above each individual’s mean (7.0%). Altogether, 10.0% of trials were excluded. When needed for post hoc analyses, we report the Greenhouse–Geisser corrected p value. Overall search accuracy was high (mean: 94.3%; SD: 5.7), and therefore we only use RT in subsequent analyses.
Results and discussion
Probe report.
The main question of interest is whether the report of letters located on the target, singleton, and non-singleton shapes differ between the two frequency conditions. The rate of letter reports was independent at each location because subjects were asked to report as many letters as possible. A 3-way ANOVA with letter location (target, singleton, non-singleton), singleton frequency (low, high), and probe type (early, late) showed three significant main effects (letter location, F(1.17, 22.19) = 110.42, p < .001, η2 = .85, singleton frequency, F(1, 19) = 4.54, p = .046, η2 = .19, and probe type, F(1, 19) = 168.84, p < .001, η2 = .90). Letter location significantly interacted with the singleton frequency, F(1.41, 26.81) = 14.43, p < .001, η2 = .43, and with the probe type, F(1.21, 23.02) = 54.88, p < .001, η2 = .74, but the interaction between singleton frequency x probe type was not significant, F(1, 19) = 2.76, p = .113. The three-way interaction was also significant, F(2, 38) = 21.58, p < .001, η2 = .53.
Because the three-way interaction was significant, we conducted two 2-way ANOVAs, dividing the data based on probe type (early and late) to clarify the source of the 3-way interaction. Within the early probe trials, there was a significant main effect of letter location, F(1.05, 19.96) = 33.27, p < .001, η2 = .64, but neither a main effect of singleton frequency nor 2-way interaction was significant, Fs < 1. The effect of letter location was due to more reports of the singleton letter than of the target, t(19) = 7.47, pBonf < .001, Cohen’s d = 1.23, or non-singleton, t(19) = 8.29, pBonf < .001, Cohen’s d = 1.38; report of the target letter and the non-singleton were not different, t(19) = 1.92, pBonf = .19. Early on, there is no benefit for singleton suppression in the high probability condition, suggesting that it takes some time to initiate. However, because participants reported very few letters (.69 per trial) on early probe trials compared to late-probe trials (1.12 per trial), t(19) = 10.24, p < .001, Cohen’s d = 2.29, Log(BF10) = 15, we interpret these results with caution.
Next, we conducted the same two-way ANOVA with letter location (target, singleton, non-singleton) and singleton frequency (low, high) on the late probe trials. We found significant main effects of letter location, F(2, 38) = 163.36, p < .001, η2 = .90, and singleton frequency, F(1, 19) = 5.42, p = .03, η2 = .22, and a significant interaction between letter location and singleton frequency, F(2, 38) = 23.05, p < .001, η2 = .55. The interaction was due to a double dissociation such that the letter at the target location was reported more often in the high frequency condition than low frequency condition, t(19) = 4.27, p < .001, Cohen’s d = .95, BF10 = 78.38, but the letter at the singleton location was reported more often in the low frequency condition than high frequency condition, t(19) = 4.61, p < .001, Cohen’s d = 1.03, BF10 = 155.66. This suggests that in the high frequency condition, the singleton distractor was better suppressed than in the low frequency condition, which allowed better selection of the target. Non-singleton distractors were reported on less than 10% of trials and there was no difference between the two frequency conditions, t(19) = 1.48, p = .156, Cohen’s d = .331, BF01 = 1.69.
More reports of the target letter and fewer reports of the letter in the singleton in the high frequency condition suggests that participants were better able to use a strategy of proactive suppression when they expected frequent singleton distractors. Proactive suppression reduced the proportion of trials when attention was captured by the singleton (indicated by singleton letter reports) and increased the proportion of target letter reports. However, it is interesting that even in the high frequency condition, the rate of singleton letter reports was still higher than target location reports, indeed it was almost twice as high (.54 vs. .21). This suggests that increasing singleton frequency facilitates distractor suppression, but does not eliminate attentional capture by all singleton distractors. In contrast, when expectations of the singleton were low, participants relied more on a reactive strategy to suppress the distractor and disengage attention after capture by the singleton (Figure 10).
Figure 10.
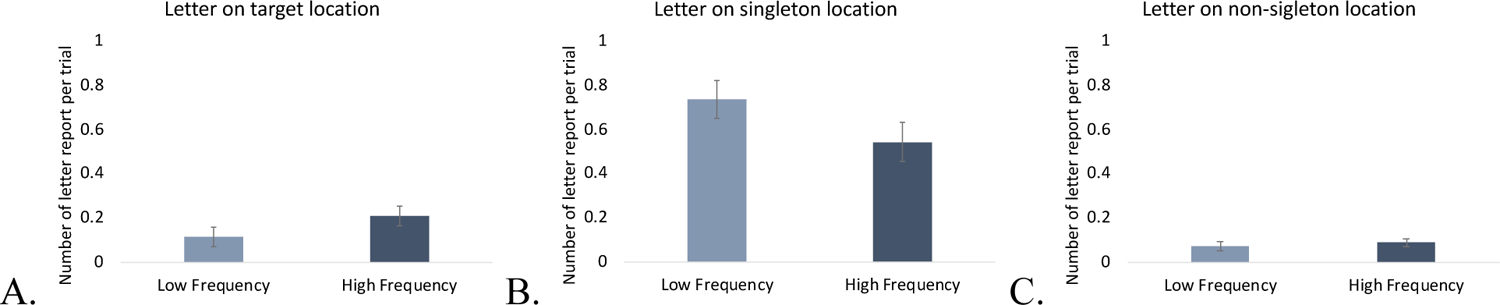
Probe report in Experiment 3. A. The number of letter reports per trial on target location, B. The number of letter reports per trial on singleton location, C. The number of letter reports per trial on any of non-singleton locations. The error bars represent within-subject 95% confidence intervals.
Visual search RTs.
We conducted a two-way ANOVA with singleton frequency (low, high) and singleton presence (present, absent). The results replicated those of Experiment 1: longer RTs were found for singleton present trials than singleton absent trials (difference: 244-msec), F(1, 19) = 181.73, p < .001, η2 = .91; the overall RTs between high and low frequency conditions did not differ, F(1, 19) = 2.24, p = .151, but importantly, there was a significant 2-way interaction between singleton frequency and singleton presence, F(1, 19) = 64.88, p < .001, η2 = .77. Consistent with the results of Experiment 1, the interaction was due to a larger singleton presence cost in the low frequency condition than the high frequency condition, although post-hoc pairwise t-tests found significant singleton presence costs for both conditions (low frequency: t(19) = 12.80, p < .001, Cohen’s d = 2.86, Log(BF10) = 18.47; high frequency condition, t(19) = 7.54, p < .001, Cohen’s d = 1.69, Log(BF10) = 10.64) (Figure11).
Figure 11.
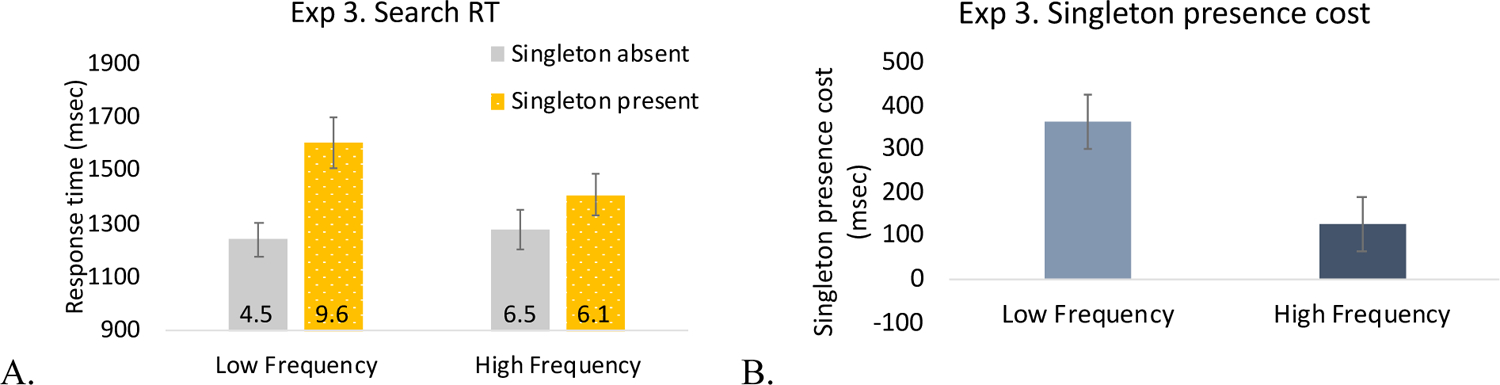
Results of Experiment 3. A. Search RT. B. Singleton-presence cost. The number in each bar indicates the error rate (%). The error bars represent within-subject 95% confidence intervals.
Repetition priming.
We again analyzed the repetition priming effects of the singleton and replicated the significant singleton repetition priming effect (the average number of singleton repetition trials was 58.3; the average number of singleton non-repetition trials was 72.8), t(19) = 3.96, p < .001, Cohen’s d = .89, BF10 = 42.59. In contrast, repetitions of singleton location did not reach to significance (the average number of location repetition trials was 10.1; the average number of location non-repetition trials was 48.2), t(19) = 1.810, p = .086, Cohen’s d = .41, BF01 = 1.09 (Figure 12). These results replicate those of Experiments 1 and 2, providing strong evidence that singleton suppression was facilitated by repetitions even when the specific properties of the singleton (location and color) do not.
Figure 12.
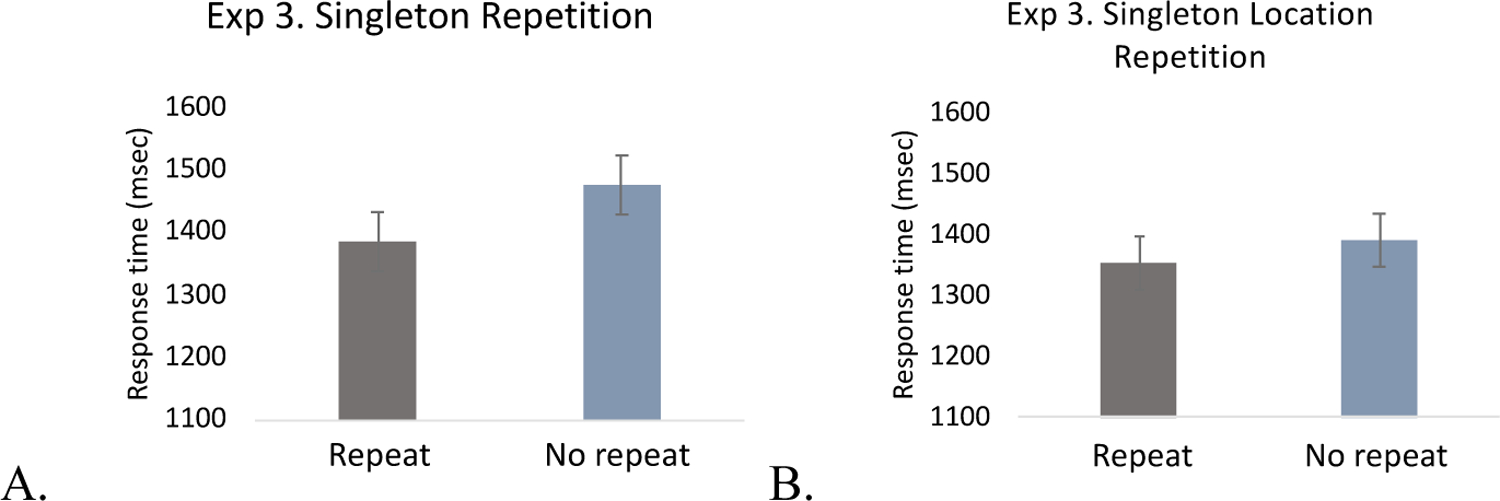
Singleton repetition priming in Experiment 3. A. Singleton presence, B. Singleton location. The error bars represent within-subject 95% confidence intervals.
First saccade destination.
The difference in proportion of first saccades to the singleton distractor between two frequency conditions replicated the results of Experiment 1: more first saccades were went to the singleton distractor in the low frequency condition than in the high frequency condition (difference = 10.2%), t(19) = 2.80, p = .011, Cohen’s d = .626, BF10 =4.57 (Figure 13). Notably, however, there were still more first saccades directed to the singleton than the target (values).
Figure 13.
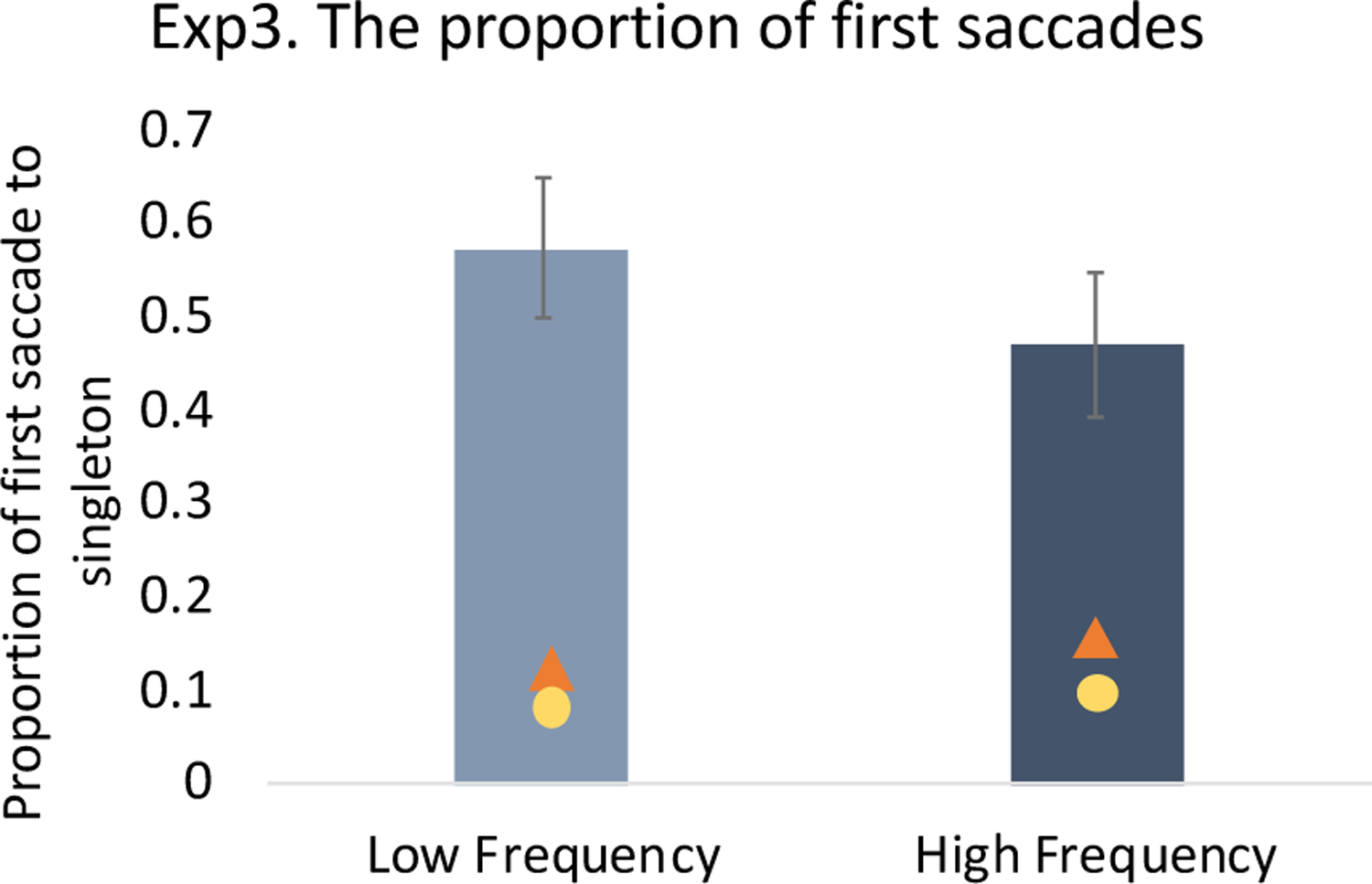
Proportion of trials having with first saccades to the color singleton in Experiment 3. The triangle indicates the proportion of trials having with first saccades to the target and the circle indicates the proportion of trials having with first saccades to a non-singleton distractor. The error bars represent within-subject 95% confidence intervals.
General Discussion
In a series of three experiments, we examined how probabilistic expectations modulate singleton suppression. Two types of expectations – singleton occurrence and singleton color – were manipulated by frequency and color variability, respectively. In Experiment 1, both singleton frequency (low or high) and color variability (mixed or fixed) were manipulated: We found that increasing singleton frequency strongly reduced attentional capture by the singleton, but changing color variability had no effect on attentional capture. These results suggest that having strong expectations for the presence of a singleton enhances suppression mechanisms that are sensitive to second order salience information. This means that subjects are able to adopt a generalizable “singleton” suppression mode rather than a specific feature-based suppression mode. In Experiment 2, we ruled out the possibility that generalized singleton suppression was due to insufficient variability of singleton colors by using new singleton colors on every trial (6 colors in Experiment 1 vs. 192 colors in Experiment 2). Finally, in Experiment 3, we used the capture probe paradigm (Gaspelin et al., 2015; Gaspelin & Luck, 2017), and found that the likelihood of attentional capture by the singleton 200–400-msec after its onset is much higher in the low frequency condition than the high frequency condition, although attentional capture is not eliminated in the high frequency condition. This suggests that the low frequency of singleton relies more heavily on reactive suppression mechanisms that disengage attention from singleton quickly in order to locate the target, but that proactive suppression mechanisms that prevent attention capture from singleton in the first place are more often engaged in the high frequency condition. Subjects appear to exploit proactive mechanisms to suppress salient distractor when their occurrence can be expected, but also still sometimes relied on reactive mechanisms.
In addition to the RT findings that singleton suppression generalized over specific singleton color values, we also found singleton repetition priming effects albeit some of the effects were weaker than others. Importantly, the repetition priming effect was neither location specific nor color specific. RTs were shorter on trials when a singleton appeared in the previous trial, even when the location and color of the singleton did not repeat. This suggests that singleton priming effects, similar to our search RT results, were based on processing of second order salience, or singleton status, rather than a specific feature or location. The consistency between priming effects and overall behavior in showing sensitivity to the singleton rather than its specific color or location, supports the notion that suppression mechanisms in this study operated at the level of second order salience and not specific features.
One might argue that our frequency effects were due to enhancement of the gray target in the high frequency blocks, rather than increased suppression of color singleton distractors. If this was true, we would have observed better (i.e., faster and more accurate) performance for the singleton absent trials in the high frequency condition than that the low frequency condition. However, search performance was not different between conditions when the singleton was absent.
A couple of interesting questions remain to be answered. The results in Experiment 3 suggest that subjects were better at suppressing singleton distractors in the high frequency blocks because stronger expectations allowed them to better exploit proactive suppression of second order salience. However, some might argue that attention was actually engaged by the singleton, but it was too rapidly disengaged for subject to recognize the letter on the singleton. This is possible and requires another experimental design with finer temporal resolution. Another interesting open question is whether there are limits to the generalization of singleton suppression based on the range of color exposure. In these studies we only used either 1, 6, or 192 singleton colors. It therefore remains unclear where the boundary is between the decision to use suppression for a feature-based singleton vs. suppression of second order singleton salience. It may be that the boundary depends on working memory capacity limits, but this remains to be tested.
In summary, the current results, in combination with previous studies (Bacon & Egeth, 1994; Moher et al., 2012; Gaspelin & Luck, 2017), suggest that distractor suppression can operate on both specific features (presumably held in a distractor template within working memory) as well as second order singleton defined saliency. Both in RT and repetition priming effects, the strength of suppression was not affected by expectations of the singleton’s location or color, but only by its appearance. Interestingly, second order salience suppression was possible for a color singleton distractor even though the target was also defined by a singleton, albeit in the shape dimension. This suggests that singleton search mode may be dimensional constrained (Müller, Reimann, & Krummenacher, 2003). Additionally, our results suggest that probabilistic expectations of singleton occurrence enhance suppression by boosting the use of proactive over reactive mechanisms. Together these results indicate that distractor suppression, like target selection, can operate at different levels of stimulus processing and be flexibly attuned to the type of information expected within the current environment.
Significance.
The mechanisms of target selection have been well-studied whereas those of distractor suppression are less well understood. One of reasons may be that there is still disagreement about whether salient distractors necessarily capture attention, which has led to a rich debate regarding the automaticity of attentional capture by stimulus saliency. However, these previous studies have primarily pitted attentional goals based on the target search strategy against perceptual saliency that cannot be predicted. We therefore have little knowledge of how probabilistic expectations affect attentional capture by a singleton distractor, yet performance in real-world environments rely heavily on expectations. We address this question in a series of three studies. Our results demonstrate that increased expectations regarding the occurrence of the singleton distractor eliminated attentional capture (indexed by search RT and first saccades). In contrast, expectations regarding singleton feature (i.e., color) did not affect singleton capture, even when there were 192 different singleton colors used. These results suggest that attentional capture by a singleton distractor is attenuated by probabilistic expectations of its occurrence, but not its specific color and location. This indicates that distractor suppression, like target selection, can operate at different levels of stimulus processing and be flexibly attuned to the type of information expected within the current environment.
Acknowledgments
Support was provided by National Science Foundation Grant BCS-201502778 to Joy J. Geng. We thank Alexis Oyao and Aditi Venkatesh for help with data collection. We also thank Nicholas Gaspelin and Steven Luck for helpful comments. All data are publicly available in https://osf.io/v8ct4/?view_only=e810dc0301d941c3b32502d828b0f2ce
Footnotes
To investigate any possible statistical biases from uneven trials between two conditions, we randomly subsampled trials from the condition with more trials to match the condition with fewer, and then compared their means using a pair-wise t-test. We did this 10,000 times and created a permuted distribution of t-values. This was done for all conditions with unequal numbers of trials in all three experiments (e.g., singleton trials vs. subsample non-singleton trials in the low frequency conditions; subsampled singleton trials vs. non-singleton trials in the high frequency condition). In every comparison that is reported as significant in the manuscript, fewer than 5% of the permuted t-values was below the critical t-value (one- and two-tailed) unless otherwise noted. The distributions of t-values for comparisons that are reported as being non-significant were all centered around zero. This indicates that conditions with the smaller number of trials had sufficient numbers to accurately estimate the central tendency of responses in that condition.
The permutation analysis using matched numbers of trials in the two conditions showed more than 95% of t-values greater than the critical one-tailed t-value, but only 89% of the data above the critical two-tailed t-value. These results are consistent with our findings that singleton repetition suppression is a significant but weaker effect than the frequency effect in our study (BF10 = 5.14 vs. BF10 = 1.370e+7).
To test whether singleton location repetition acts within a broader spatial range (i.e., has poor spatial resolution), we expanded the repetition locations to the two adjacent locations, and found no significant singleton location repetition effects in Experiment 1, t(19) = 1.08, p = .29, BF01 = 2.57, or in Experiment 3, t(19) = .70, p = .49, BF01 = 3.45. However, a weak but significant repetition effect was found in Experiment 2, t(19) = 2.24, p = .037, BF10 = 1.75. Given the inconsistent effects, we refrain from drawing strong conclusions about the presence of broad spatial repetition suppression.
The permutation analysis using matched numbers of trials in the two conditions showed only 64% of t-values being greater than the critical one-tailed t-value, and 50% above the two-tailed critical t-value. Thus, this analysis may have been affected by the smaller number of trials included within the repetition condition and therefore the significant difference should be interpreted with caution.
References
- Arita JT, Carlisle NB, & Woodman GF (2012). Templates for rejection: Configuring attention to ignore task-irrelevant features. Journal of Experimental Psychology: Human Perception and Performance, 38(3), 580–584. doi: 10.1037/a0027885 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bacon WF & Egeth HE (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55(5), 485–496. [DOI] [PubMed] [Google Scholar]
- Balan PF & Gottlieb J (2006). Integration of exogenous input into a dynamic salience map revealed by perturbing attention. Journal of Neuroscience, 26(36), 9239–9249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldi P & Itti L (2010). Of bits and wows: A Bayesian theory of surprise with applications to attention. Neural Networks, 23, 649–666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becker SI (2007). Irrelevant singletons in pop-out search: Attentional capture or filtering costs? Journal of Experimental Psychology: Human Perception and Performance, 33(4), 764–787. doi: 10.1037/0096-1523.33.4.764 [DOI] [PubMed] [Google Scholar]
- Born S, Kerzel D, & Theeuwes J (2011). Evidence for a dissociation between the control of oculomotor capture and disengagement. Experimental Brain Research. 208(4), 621–31. doi: 10.1007/s00221-010-2510-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brainard DH (1997). The psychophysics toolbox. Spatial Vision, 10, 433–436. 10.1163/156856897X00357 [DOI] [PubMed] [Google Scholar]
- Braver TS (2012). The variable nature of cognitive control: a dual mechanisms framework. Trends in Cognitive Science, 16(2), 106–113. doi: 10.1016/j.tics.2011.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chun MM & Marois R (2002). The dark side of visual attention. Current Opinion of Neurobiology, 12(2), 184–189. [DOI] [PubMed] [Google Scholar]
- Cunningham CA, & Egeth HE (2016). Taming the White Bear. Psychological Science, 27(4), 476–485. doi: 10.1177/0956797615626564 [DOI] [PubMed] [Google Scholar]
- Desimone R, & Duncan J (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18(1), 193–222. doi: 10.1146/annurev.ne.18.030195.001205 [DOI] [PubMed] [Google Scholar]
- DiQuattro NE & Geng JJ (2017). Presaccadic target competition attenuates distraction. Attention, Perception and Psychophysics, 79(4), 1087–1096. doi: 10.3758/s13414-017-1288-7. [DOI] [PubMed] [Google Scholar]
- Duncan J, Humphreys G, & Ward R (1997). Competitive brain activity in visual attention. Current Opinion in Neurobiology, 7(2), 255–261. doi: 10.1016/S0959-4388(97)80014-1 [DOI] [PubMed] [Google Scholar]
- Egeth HE & Yantis S (1997). Visual attention: control, representation, and time course. Annual Review of Psychology. 48, 269–297. doi: 10.1146/annurev.psych.48.1.269 [DOI] [PubMed] [Google Scholar]
- Failing M & Theeuwes J (2014). Exogenous visual orienting by reward. Journal of Vision, 14(5):6, 1–9, doi: 10.1167/14.5.6 [DOI] [PubMed] [Google Scholar]
- Failing M & Theeuwes J (2017). Don’t let it distract you: how information about the availability of reward affects attentional selection. Attention, Perception, & Psychophysics, 79(8):2275–2298. doi: 10.3758/s13414-017-1376-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrante O, Patacca A, Di Caro V, Della Libera C, Santandrea E, & Chelazzi L (2017). Altering spatial priority maps via statistical learning of target selection and distractor filtering. Cortex. 102, 67–95. doi: 10.1016/j.cortex.2017.09.027 [DOI] [PubMed] [Google Scholar]
- Folk CL, Leber AB, & Egeth HE (2002). Made you blink! Contingent attentional capture produces a spatial blink. Perception & Psychophysics, 64(5), 741–753. 10.3758/BF03194741 [DOI] [PubMed] [Google Scholar]
- Folk CL, & Remington RW (2008). Bottom-up priming of top-down attentional control settings. Visual Cognition, 16(2–3), 215–231. 10.1080/13506280701458804 [DOI] [Google Scholar]
- Folk CL, & Remington RW (1998). Selectivity in distraction by irrelevant featural singletons: evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 24(3), 847–858. doi: 10.1037/0096-1523.24.3.847 [DOI] [PubMed] [Google Scholar]
- Folk CL, Remington RW, & Johnston JC (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18(4), 1030–1044. doi: 10.1037/0096-1523.18.4.1030 [DOI] [PubMed] [Google Scholar]
- Franconeri SL & Simons DJ (2003). Moving and looming stimuli capture attention. Perception & Psychophysics, 65(7), 999–1010. [DOI] [PubMed] [Google Scholar]
- Gaspelin N, Leonard CJ, & Luck SJ (2015). Direct evidence for active suppression of salient-but-irrelevant sensory inputs. Psychological Science, 26(11), 1740–1750. 10.1177/0956797615597913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaspelin N, Leonard CJ, & Luck SJ (2017). Suppression of overt attentional capture by salient-but-irrelevant color singletons. Attention, Perception, & Psychophysics, 79(1), 45–62. 10.3758/s13414-016-1209-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaspelin N, & Luck SJ (2017). Distinguishing among potential mechanisms of singleton suppression. Journal of Experimental Psychology: Human Perception and Performance, 44(4), 626–644. doi: 10.1037/xhp0000484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaspelin N, & Luck SJ (2018). The role of inhibition in avoiding distraction by salient stimuli. Trends in Cognitive Sciences, 22(1), 79–92. 10.1016/j.tics.2017.11.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geng JJ (2014). Attentional mechanisms of distractor suppression. Current Directions in Psychological Science, 23, 147–153. [Google Scholar]
- Geng JJ, & Behrmann M (2002). Probability cuing of target location facilitates visual search implicitly in normal participants and patients with hemispatial neglect. Psychological Science, 13, 520–525. doi: 10.1111/1467-9280.00491 [DOI] [PubMed] [Google Scholar]
- Geng JJ, & Behrmann M (2005). Spatial probability as an attentional cue in visual search. Perception & Psychophysics, 67(7), 1252–1268. doi: 10.3758/BF03193557 [DOI] [PubMed] [Google Scholar]
- Geyer T, Müller HJ, & Krummenacher J (2008). Expectancies modulate attentional capture by salient color singletons. Vision Research, 48, 1315–1326. doi: 10.1016/j.visres.2008.02.006 [DOI] [PubMed] [Google Scholar]
- Horstmann G (2002). Evidence for attentional capture by a surprising color-singleton in visual search. Psychological Science, 13(6), 499–505. [DOI] [PubMed] [Google Scholar]
- Horstmann G (2005). Attentional capture by an unannounced color- singleton depends on expectation discrepancy. Journal of Experimental Psychology: Human Perception and Performance, 31, 1039–1060. [DOI] [PubMed] [Google Scholar]
- JASP Team. (2017). JASP (Version 0.8.1.2) [Computer software]. Retrieved from https://jasp-stats.org/faq/how-do-i-cite-jasp/
- Jiang YV, Swallow KM, Rosenbaum GM, & Herzig C (2013). Rapid acquisition but slow extinction of an attentional bias in space. Journal of Experimental Psychology: Human Perception and Performance, 39(1), 87–99. doi: 10.1037/a0027611 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kristjánsson Á & Campana G (2010). Where perception meets memory: A review of repetition priming in visual search tasks. Attention, Perception, & Psychophysics, 72(1), 5–18. doi: 10.3758/APP.72.1.5 [DOI] [PubMed] [Google Scholar]
- Lamy D & Egeth HE (2003). Attentional capture in singleton-detection and feature-search modes. Journal of Experimental Psychology: Human Perception and Performance, 29(5), 1003–1020. doi: 10.1037/0096-1523.29.5.1003 [DOI] [PubMed] [Google Scholar]
- Lavie N (1995). Perceptual load as a necessary condition for selective attention. Journal of Experimental Psychology: Human Perception and Performance, 21(3), 451–468. doi: 10.1037/0096-1523.21.3.451 [DOI] [PubMed] [Google Scholar]
- Lavie N, Ro T, & Russell C (2003). The role of perceptual load in processing distractor faces. Psychological Science, 14(5), 510–515, doi: 10.1111/1467-9280.03453 [DOI] [PubMed] [Google Scholar]
- Leber AB, & Egeth HE (2006). Attention on autopilot: Past experience and attentional set. Visual Cognition, 14(4–8), 565–583. [Google Scholar]
- Leonard CJ & Luck SJ (2011). The role of magnocellular signals in oculomotor attentional capture. Journal of Vision, 11(13):11, 1–12. doi: 10.1167/11.13.11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loftus GR, & Masson MEJ (1994). Using confidence intervals in within-subject designs. Psychonomic Bulletin and Review, 1, 476–490. doi: 10.3758/BF03210951 [DOI] [PubMed] [Google Scholar]
- Maljkovic V, & Nakayama K (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22(6), 657–672. doi: 10.3758/BF03209251 [DOI] [PubMed] [Google Scholar]
- Maljkovic V & Nakayama K (1996). Priming of pop-out: II. The role of position. Perception & Psychophysics. 58, 977–991. doi: 10.3758/BF03206826 [DOI] [PubMed] [Google Scholar]
- Marini F, Demeter E, Roberts KC, Chelazzi L, & Woldorff MG (2016). Orchestrating proactive and reactive mechanisms for filtering distracting information: Brain-behavior relationships revealed by a mixed-design fMRI study. The Journal of Neuroscience, 36, 988–1000. doi: 10.1523/JNEUROSCI.2966-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez-Trujillo JC, & Treue S (2004). Feature-based attention increases the selectivity of population responses in primate visual cortex. Current Biology, 14, 744–751. doi: 10.1016/j.cub.2004.04.028 [DOI] [PubMed] [Google Scholar]
- Maunsell JH & Treue S (2006). Feature-based attention in visual cortex. Trends in Neuroscience. 29(6), 317–322. [DOI] [PubMed] [Google Scholar]
- Mazaheri A, DiQuattro NE, Bengson J, & Geng JJ (2011). Pre-stimulus activity predicts the winner of top-down vs. bottom-up attentional selection. PLoS ONE, 6(2), e16243. doi: 10.1371/journal.pone.0016243 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moher J, Abrams J, Egeth HE, Yantis S, & Stuphorn V (2011). Trial-by-trial adjustments of top-down set modulate oculomotor capture. Psychonomic Bulletin & Review, 18(5), 897–903. doi: 10.3758/s13423-011-0118-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moher J & Egeth HE (2012). The ignoring paradox: cueing distractor features leads first to selection, then to inhibition of to- be-ignored items. Attention, Perception and Psychophysics, 74, 1590–1605. [DOI] [PubMed] [Google Scholar]
- Müller HJ, Geyer T, Zehetleitner M, and Krummenacher J (2009). Attentional capture by salient color singleton distractors is modulated by top-down dimensional set. Journal of Experimental Psychology: Human Perception and Performance, 35, 1–16. doi: 10.1037/0096-1523.35.1.1 [DOI] [PubMed] [Google Scholar]
- Müller HJ, Reimann B, & Krummenacher J (2003). Visual search for singleton feature targets across dimensions: Stimulus- and expectancy-driven effects in dimensional weighting. Journal of Experimental Psychology: Human Perception and Performance, 29(5), 1021–1035. doi: 10.1037/0096-1523.29.5.1021 [DOI] [PubMed] [Google Scholar]
- Pelli DG (1997). The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10, 437–442. 10.1163/156856897X00366 [DOI] [PubMed] [Google Scholar]
- Posner MI (1980). Orienting of attention. The Quarterly Journal of Experimental Psychology, 32(1), 3–25. [DOI] [PubMed] [Google Scholar]
- Proulx MJ & Serences JT (2006). Searching for an oddball: neural correlates of singleton detection mode in parietal cortex. Journal of Neuroscience. 26(49), 12631–12632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rouder JN, Speckman PL, Sun D, Morey RD, & Iverson G (2009). Bayesian t tests for accepting and rejecting the null hypothesis. Psychonomic Bulletin & Review, 16, 225–237. doi: 10.3758/PBR.16.2.225 [DOI] [PubMed] [Google Scholar]
- Theeuwes J (1991). Cross-dimensional perceptual selectivity. Perception & Psychophysics, 50, 184. doi: 10.3758/BF03212219 [DOI] [PubMed] [Google Scholar]
- Theeuwes J (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51(6), 599–606. [DOI] [PubMed] [Google Scholar]
- Theeuwes J (2004) Top-down search strategies cannot override attentional capture. Psychonomic Bulletin & Review, 11, 65–70 [DOI] [PubMed] [Google Scholar]
- Theeuwes J (2010). Top-down and bottom-up control of visual selection. Acta Psychologica, 135(2), 77–99. doi: 10.1016/j.actpsy.2010.02.006 [DOI] [PubMed] [Google Scholar]
- Theeuwes J (2013). Feature-based attention: it is all bottom-up priming. Philosophical Transactions of the Royal Society B: Biological Sciences, 368(1628), 20130055. doi: 10.1098/rstb.2013.0055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theeuwes J, Atchley P, & Kramer AF (2000). On the time course of top-down and bottom-up control of visual attention. In Monsell S & Driver J (Eds.), Attention and performance XVIII: Control of cognitive performance (pp. 105–125). Cambridge, MA: MIT Press. [Google Scholar]
- Theeuwes J, Kramer AF, Hahn S, & Irwin DE (1998). Our eyes do not always go where we want them to go: Capture of the eyes by new objects. Psychological Science, 9(5), 379–385. [Google Scholar]
- Van Diepen RM, Miller LM, Mazaheri A, & Geng JJ (2016). The role of alpha activity in spatial and feature-based attention. eNeuro, 3(5), ENEURO.0204–16.2016. doi: 10.1523/ENEURO.0204-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vatterott DB, Mozer MC, & Vecera SP (2018). Rejecting salient distractors: Generalization from experience. Attention, Perception & Psychophysics, 80(2), 485–499. doi: 10.3758/s13414-017-1465-8. [DOI] [PubMed] [Google Scholar]
- Vatterott DB, & Vecera SP (2012). Experience-dependent attentional tuning of distractor rejection. Psychonomic Bulletin & Review, 19(5), 871–878. doi 10.3758/s13423-012-0280-4 [DOI] [PubMed] [Google Scholar]
- Wang B & Theeuwes J (2018a). Statistical regularities modulate attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 44(1), 13–17. doi: 10.1037/xhp0000472 [DOI] [PubMed] [Google Scholar]
- Wang B & Theeuwes J (2018b) How to inhibit a distractor location? Statistical learning versus active, top-down suppression. Attention, Perception, & Psychophysics, 80(4), 860–870. doi: 10.3758/s13414-018-1493-z [DOI] [PubMed] [Google Scholar]
- Wolfe JM, Cave KR, & Franzel SL (1989). Guided search: An alternative to the feature integration model for visual search. Journal of Experimental Psychology: Human Perception and Performance, 15(3), 419–433. doi: 10.1037/0096-1523.15.3.419 [DOI] [PubMed] [Google Scholar]
- Won B-Y & Geng JJ (2018). Learned suppression for multiple distractors in visual search. Journal of Experimental Psychology: Human Perception and Performance, [Epub ahead of print] doi: 10.1037/xhp0000521 [DOI] [PubMed] [Google Scholar]
- Yantis S, & Jonides J (1984). Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10(5), 601–621. [DOI] [PubMed] [Google Scholar]