

Abstract
The highly heterogeneous symptomatology and unpredictable progress of COVID-19 triggered unprecedented intensive biomedical research and a number of clinical research projects. Although the pathophysiology of the disease is being progressively clarified, its complexity remains vast. Moreover, some extremely infrequent cases of thrombotic thrombocytopenia following vaccination against SARS-CoV-2 infection have been observed. The present study aimed to map the signaling pathways of thrombocytopenia implicated in COVID-19, as well as in vaccine-induced thrombotic thrombocytopenia (VITT). The biomedical literature database, MEDLINE/PubMed, was thoroughly searched using artificial intelligence techniques for the semantic relations among the top 50 similar words (>0.9) implicated in COVID-19-mediated human infection or VITT. Additionally, STRING, a database of primary and predicted associations among genes and proteins (collected from diverse resources, such as documented pathway knowledge, high-throughput experimental studies, cross-species extrapolated information, automated text mining results, computationally predicted interactions, etc.), was employed, with the confidence threshold set at 0.7. In addition, two interactomes were constructed: i) A network including 119 and 56 nodes relevant to COVID-19 and thrombocytopenia, respectively; and ii) a second network containing 60 nodes relevant to VITT. Although thrombocytopenia is a dominant morbidity in both entities, three nodes were observed that corresponded to genes (AURKA, CD46 and CD19) expressed only in VITT, whilst ADAM10, CDC20, SHC1 and STXBP2 are silenced in VITT, but are commonly expressed in both COVID-19 and thrombocytopenia. The calculated average node degree was immense (11.9 in COVID-19 and 6.43 in VITT), illustrating the complexity of COVID-19 and VITT pathologies and confirming the importance of cytokines, as well as of pathways activated following hypoxic events. In addition, PYCARD, NLP3 and P2RX7 are key potential therapeutic targets for all three morbid entities, meriting further research. This interactome was based on wild-type genes, revealing the predisposition of the body to hypoxia-induced thrombosis, leading to the acute COVID-19 phenotype, the 'long-COVID syndrome', and/or VITT. Thus, common nodes appear to be key players in illness prevention, progression and treatment.
Keywords: SARS-CoV-2, COVID-19, thrombocytopenia, vaccine-induced thrombotic thrombocytopenia, interactions network, HLA system, enzymes, lymphocytes, autoimmunity, artificial intelligence, cytokine storm, natural language processing
Introduction
The current SARS-CoV-2-induced pandemic has raised a number of public health policy and scientific queries, related to the virus origin, transmission, activity, contamination, pathophysiologic effects and treatment. As of May 3, 2021, almost 188 million cases had been confirmed, while 4.05 million deaths had been registered under the cause of death: 'COVID-19'. Although this may underline an apogee of the third phase of the pandemic in some countries, or may have been the result of certain interventions. Public health policy approaches, communication campaigns, pharmacological approaches, surveillance, and prevention practices have been suggested.
The highly varying symptomatology and the unpredictable global progress of COVID-19 have triggered an unprecedentedly intensive activity in biomedical research and public policy decisions. Furthermore, although the pathophysiology of the disease is being progressively clarified, its complexity remains vast, and preventive care approaches or treatments, although both have significantly improved, remain unsatisfactory.
Notably, the extremely rare yet highly unpredictable and occasionally lethal vaccination-induced thrombotic thrombocytopenia (VITT) syndrome has emphasized the gaps in the current knowledge of certain unsuspected pathophysiological pathways. The VITT morbid entity is of particular importance given the generally mild and to a certain extent expected vaccination side-effects, namely chills, fever, diarrhea, fatigue, muscle pain, headache and mildly increased blood coagulability (1,2). As of April 2021, 16 vaccination options were available: Two RNA vaccines [BNT162b2 (Comirnaty) by Pfizer-BioNTech, mRNA.1273 (Spikevax) by Moderna], seven conventional inactivated ones (CoronaVac, Covaxin, BBIBP-CorV, WIBP-CorV, Minhai-Kangtai, QazVac, CovIran Bakerat), five viral vector-employing ones (Covishield and Vaxzevria by Oxford Astra-Zeneca, the Janssen COVID-19 vaccine by Johnson & Johnson, the Sputnik V and Sputnik Light by the Gamaleya Research Institute of Epidemiology and Microbiology in Russia, and the AD5-nCOV-Convidencia by CanSino Biologics Inc.), and two protein subunit vaccines (EpiVacCorona and RDB-dimer). Vaccination programs have been implemented so as to reach 'herd immunity', in every country. According to national health authority reports, as of August 30, 2021, 5.27 billion doses had been administered globally. This is equal to 39.7% of the population on the planet (where, however, only 1.6% of individuals in the low-income countries had received at least one dose), having been fully vaccinated (3). As of August 30, 2021, 55.15% of the Greek population had been fully vaccinated (3).
The aim of the present study was to illustrate the signaling pathways implicated in SARS-CoV-2 infection, including those of the extremely rare, yet severe VITT syndrome.
Data and methods
The scientific literature database, MEDLINE/PubMed (https://pubmed.ncbi.nlm.nih.gov/), was searched thoroughly for genes or gene products implicated in COVID-19 infection and VITT syndrome. Searches were conducted in the PubTator article collection (4) (https://www.ncbi.nlm.nih.gov/research/pubtator/) from the LitCovid database (5), using i) ('COVID19' OR 'SARS-CoV-2') AND ('VITT' OR 'vaccine-induced thrombotic thrombocytopenia'); ii) ('COVID19' OR 'SARS-CoV-2') AND ('thrombocytopenia' OR 'thrombopenia') key words to obtain relevant articles. Of the 495 candidate articles, 190 met the inclusion criteria which were as follows: i) written in English; ii) include an abstract; and iii) contain adequate information in their text for processing (Fig. 1).
Figure 1.
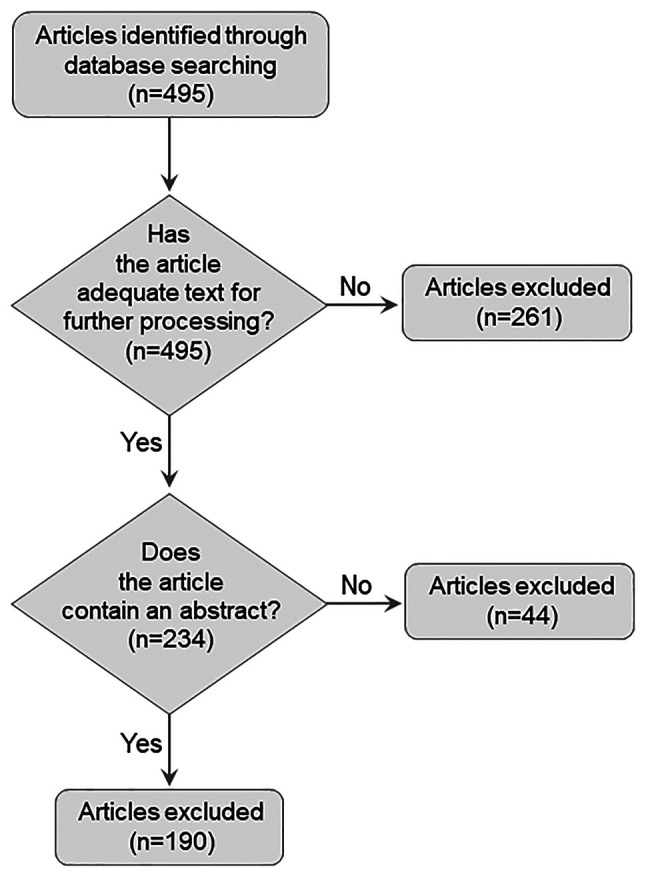
Flowchart of the process followed for the acquisition of eligible articles containing relevant data.
The natural language toolkit (NLTK: https://www.nltk.org/), a freely accessible Python platform, was used for text processing, including tokenization, parsing and stemming. Word2vec embeddings module in the open-source Python library Gensim (https://pypi.org/project/gensim/) was implemented to train word vectors of processed text. A list of all word-to-word distances was extracted. To calculate the similarity distances between each word pair, the Word2Vec.most_similar function in Gensim Word2vec model was used. The top 50 detected entries were included in the present study. The work flow is presented in Fig. 1. The search results are illustrated in Fig. 2.
Figure 2.

Networks depicting the semantic relations of the top 50 most similar words to the query (A) COVID-19 or VITT, and (B) thrombocytopenia. Only those word pairs with a cosine similarity score of each word vector >0.9 are shown. The nodes represent the words and the edges denote the semantic associations between them. The size of the nodes indicates the frequency of occurrence of the given term. VITT, vaccine-induced thrombotic thrombocytopenia.
Furthermore, the interactions among the retrieved genes/proteins were investigated by employing the Search Tool for Retrieval of Interacting Genes/Proteins (STRING) database v11.0 (6,7), a database containing both primary and predicted, physical and functional association data among genes or proteins. These data are collected from diverse resources, such as documented pathway knowledge, high-throughput experimental studies, cross-species extrapolated information, automated text mining results, computationally predicted interactions, etc. The confidence threshold value for displaying interactions was set to 'high' (i.e., 0.7). The interactions in the generated network were manipulated and visualized through Cytoscape (http://www.cytoscape.org/) (8), a software platform for network processing and statistical analyses; the Edge Betweenness mode was used to detect the number of the shortest paths that pass-through a given edge in the COVID-19 network.
Results
Main findings
The constructed networks presented in Fig. 2 provide noteworthy information on how diverse terms are closely interlinked within the context of thrombocytopenia induced by SARS-CoV-2 infection or through vaccination. The term thrombocytopenia appears with a rather high frequency in the COVID-19/VITT network (Fig. 2A). Similarly, the term VITT is included in the COVID-19/thrombocytopenia network (Fig. 2B). COVID-19 and VITT share several comorbidities implicating vascular and epithelial dysfunction and thrombocytopenia. The nodes represent the top 50 words with a cosine similarity score of each word vector >0.9.
Interactome construction
Subsequently, two interactomes were constructed: The first one involving 119 nodes is described in Table I and illustrated in Fig. 3. Collectively, 119 nodes are involved in COVID-19, while 57 are implicated in thrombocytopenia [the latter profits from an unpublished work of ours (unpublished data). Of these, 14 nodes were common in both entities (Figs. 3 and 4), namely AIM2, IFI16, NOD2, CD8A, IL-1B, 1L-6, JAK2, NCAM1, HLA-DRB1, SERPINE1, TGFB1, TLR2, TNF and VWF. The major hubs detected are displayed in the center of the constructed circular network, while the less connected nodes are shown at the periphery of the circle (Fig. 3). The thrombocytopenia-related nodes are represented in square bullets, and the COVID-19-related ones are presented in circles, whilst the common nodes are depicted in rhomboids. The calculated average node degree of the entire interactome was extremely high (11.9).
Table I.
| Gene symbol | Gene name | Main function with brief description (Refs.) | Figure(s) | Entitya |
|---|---|---|---|---|
| ACE2 | Angiotensin I converting enzyme 2 | Transmembrane protein-entry point of SARS-CoV-2 (22-24,28) | 3 | C |
| ADAM10 | ADAM metallopeptidase domain 10 | Sheddase with strong specificity for peptide hydrolysis reactions (68-70) | 3 | T |
| ADAM17 | ADAM metallopeptidase domain 17 | Sheddase triggering release of cytokines receptors, ligands, etc. (68,69,71) | 3,4 | V, T |
| ADAMTS13 | ADAM metallopeptidase with throm bospondin type 1 motif 13 | Enzyme that cleaves von Willebrand factor (68,69) | 3,4 | V, T |
| ADRA2C | Adrenoceptor alpha 2C | Mediators in catecholamine-induced inhibition of adenylate cyclase through the action of G proteins (72) | 3 | C |
| ADRB1 | Adrenoceptor beta 1 | Renin release/lipolysis/Increases heart rate with chrono/inotropic effect (73) | 3 | C |
| ADRB2 | Adrenoceptor beta 2 | Facilitating respiration (74) | 3 | C |
| AGER | Advanced glycosylation end-product specific receptor | Mediates interactions of advanced glycosylation end products (75) | 3 | C |
| AIM2 | Interferon-inducible protein AIM2 | AIM2 inflammasome plays a crucial role in the defense against viral infection (76) | 3 | C, T |
| ANGPT1 | Angiopoietin 1 | Receptor of advanced glycosylation end products of proteins, mediating amyloid beta peptide effect on neurons and microglia (77) | 3 | C |
| ANGPT2 | Angiopoietin 2 | Binds to TEK/TIE2, competing for the ANGPT1 binding site, and modulating ANGPT1 signaling (78) | 3 | C |
| AURKA | Serine/threonine-protein kinase 6 | Orchestrate an exit from mitosis by contributing to the completion of cytokinesis the process through which the cytoplasm of the parent cell is split into two daughter cells (79) | 4 | V |
| C4B | Complement C4B (Chido blood group) | Mediator of local inflammatory process, inducing the contraction of smooth muscle, increasing vascular permeability and causing histamine release from mast cells and basophilic leukocytes (80) | 3,4 | V, T |
| C5 | Complement C5 | Involved in the complement system (81) | 3,4 | V, T |
| C6 | Complement C6 | Causes cell lysis (82) | 3,4 | V, T |
| C7 | Complement C7 | Creates a hole on pathogen surfaces leading to cell lysis (82) | 3,4 | V, T |
| C9 | Complement C9 | Cell lysis and death contributor (82) | 3,4 | V, T |
| CASP1 | Caspase 1 | Inflammatory response initiator (83) | 3,4 | C, V |
| CASP10 | Caspase 10 | Cell apoptosis (84) | 3 | C |
| CASP9 | Caspase 9 | Innate immunity, mitochondrial apoptosis (85) | 3 | C |
| CCL2 | C-C motif chemokine ligand 2 | Induces a strong chemotactic response and mobilization of intracellular calcium ions (86,87) | 3 | C |
| CCL3 | Chemokine (C-C motif) ligand 3 | Pyrogenic, attracting macrophages, monocytes and neutrophils (88) | 3 | C |
| CCN2 | Cellular communication network factor 2 | Cell adhesion, apoptosis, migration, proliferation, differentiation, apoptosis, survival and senescence (89) | 3 | C |
| CD3D | CD3d molecule | Cell differentiation and adaptive immune response (90) | 3 | C |
| CD3E | CD3e molecule | Cell differentiation and adaptive immune response (90) | 3 | C |
| CD3G | CD3g molecule | Cell differentiation and adaptive immune response (90) | 3 | C |
| CD4 | CD4 molecule | Cell differentiation and adaptive immune response (91) | 3 | C |
| CD40LG | CD40 ligand | Acts as a ligand for integrins which have cell-type dependent effects, such as B-cell activation, NF-κB signaling and anti-apoptotic signaling (92,93) | 3 | T |
| CD8A | CD8a molecule | Multiple functions in responses against both ex/internal offenses (91) | 3 | C, T |
| CD19 | B-lymphocyte antigen CD19 | Decreases B-cell receptor pathways (94,95) | 4 | V |
| CD40LG | Cluster of differentiation 40 | Mediates many immune and inflammatory responses including T-cell-dependent immunoglobulin class switching, memory B cell development, and germinal center formation (96) | 3,4 | T, V |
| CD46 | CD46 complement regulatory protein | Activates T-lymphocytes following vaccination (97,98) | 4 | V |
| CDC20 | Cell division cycle 20 | Regulates the formation of synaptic vesicle clustering at active zone to the presynaptic membrane in post-mitotic neurons; Cdc20-apc/ c-induced degradation of neurod2 induces presynaptic differentiation (91) | 3,4 | V, T |
| CDCA3 | Cell division cycle associated 3 | Involves in protein ubiquitination (99) | 3,4 | V, T |
| CRP | C-reactive protein | Mitotic initiator (100) | 3 | C |
| CSF1R | Colony stimulating factor 1 receptor | Controls the production, differentiation, and function macrophages (93,101) | 3,4 | V, T |
| CSF2 | Colony stimulating factor 2 | Cytokine affecting mostly eosinophils and macrophages (102) | 3,4 | V, T |
| CXCL10 | C-X-C motif chemokine ligand 10 | Chemoattraction for T- and NK cells, monocytes (87,93,103,104) | 3 | C |
| CXCL8 | C-X-C motif chemokine ligand 8 | Neutrophil chemotactic factor increasing respiratory burst (87,105) | 3,4 | C, V |
| CYP11B2 | Cytochrome P450 family 11 subfamily B member 2 | Aldosterone synthesis (87,106) | 3 | C |
| CYP2C19 | Cytochrome P450 family 2 subfamily C member 19 | Part of cytochrome P450, involved in drug and lipid metabolism (107) | 3 C | |
| CYP2C9 | Cytochrome P450 family 2 subfamily C member 9 | Part of cytochrome P450, involved in drug and lipid metabolism (107) | 3 C | |
| DDX58 | Retinoic acid-inducible gene I | Activates interferon and cytokines production after viral infection (108) | 3 | C |
| EDN1 | Endothelin 1 | Potent vasoconstrictor (106,109) | 3 | C |
| EPO | Erythropoietin | Stimulation of erythropoiesis, vasoconstriction, angiogenesis (106) | 3,4 | V, T |
| F2 | Coagulation factor II, thrombin | Activates the coagulation cascade inhibition (110) | 3,4 | V, T |
| FCGR1A | Fc fragment of IgG receptor Ia | Complex activation or inhibitory effects on cell functions (111) | 3,4 | V, T |
| FCGR1B | Fc fragment of IgG receptor Ib | Humoral immune response (112) | 3,4 | V, T |
| FCGR2A | Fc fragment of IgG receptor IIa | Humoral immune response to pathogens, phagocytosis of opsonized antigens (113) | 3,4 | V, T |
| FCGR2B | Fc fragment of IgG receptor IIb | Phagocytosis of immune complexes and regulation of antibody production (114) | 3,4 | V, T |
| FCGR3A | Fc fragment of IgG receptor IIIa | Mediates antibody-dependent cellular cytotoxicity and phagocytosis (115) | 3,4 | V, T |
| FCGR3B | Fc fragment of IgG receptor IIIb | Captures immune complexes in the peripheral circulation (116) | 3,4 | V, T |
| FGF7 | Fibroblast growth factor 7 | Cell growth, morphogenesis and tissue repair (117) | 3,4 | C, V |
| FKBP1A | FKBP prolyl isomerase 1A | Immunoregulation and basic cellular processes involving protein folding and trafficking (118) | 3,4 | V, T |
| FN1 | Fibronectin 1 | Cell growth, morphogenesis and tissue repair (70) | 3 | C |
| FOS | Fos proto-oncogene, AP-1 transcription factor subunit | Signal transduction, cell proliferation and differentiation (119) | 3 | C |
| GNB3 | G protein subunit beta 3 | Integrates signals between receptor and effector proteins (120) | 3 | C |
| GZMA | Granzyme A | Common component necessary for lysis of target cells by cytotoxic T-lymphocytes and natural killer cells (24) | 3 | C |
| GZMB | Granzyme B | Recognize specific infected target cells (121) | 3 | C |
| GZMH | Granzyme H | Suppresses viral replication (122) | 3 | C |
| HLA-A | Major histocompatibility complex, class I, A | Sole link between the immune system and what happens inside cells (123) | 3,4 | C, V |
| HLA-B | Major histocompatibility complex, class I, B | Helps the immune system distinguish the endo-from exogenous proteins (123) | 3,4 | C, V |
| HLA-DRB1 | HLA class II histocompatibility antigen, DRB1 beta chain | Triggers response to viral infections (41) | 3,4 | C, V, T |
| ICAM1 | Intercellular adhesion molecule 1 | Signal transduction (92,93) | 3,4 | V, T |
| IFI16 | Interferon gamma inducible protein 16 | Recognizes RNA viral infection, enhancing DDX58 production (124) | 3 | C, T |
| IFNA1 | Interferon alpha 1 | Antiviral and immunomodulator (125) | 3 | C |
| IFNG (IFN-γ) | Interferon gamma | Antiviral antibacterial and immunomodulatory effects (104) | 3,4 | V, T |
| IFNL1 | Interferon lambda 1 | Antiviral antibacterial and immunomodulatory effects (126) | 3 | C |
| IFNL2 | Interferon lambda 2 | Antiviral antibacterial and immunomodulatory effects (126) | 3 | C |
| IFNL3 | Interferon lambda 3 | Antiviral antibacterial and immunomodulatory effects (126) | 3 | C |
| IFNLR1 | Interferon lambda receptor 1 | Antiviral antibacterial and immunomodulatory effects (126) | 3 | C |
| IKBKG | Inhibitor of nuclear factor kappa B kinase regulatory subunit gamma | Antiviral activity through JAK/STAT signaling activation (127) | 3 | C |
| IL10 | Interleukin 10 | Multiple, pleiotropic effects in immunoregulation, limits excessive infected tissue disruption (92) | 3 | C |
| IL10RB | Interleukin 10 receptor subunit beta | JAK1 and STAT2-mediated phosphorylation of STAT3 (128) | 3 | C |
| IL12A | Interleukin 12A | Induces IFNG (92) | 3 | C |
| IL12B | Interleukin 12B | Induces IFNG by resting PBMC (92) | 3 | C |
| IL17A | Interleukin 17A | Mediates protective innate immunity to pathogens or contributes to pathogenesis of inflammatory diseases (87) | 3 | C |
| IL18 | Interleukin 18 | Potent inducer of inflammatory cytokines, especially IFNG (129) | 3 | C |
| IL1A | Interleukin 1 alpha | Promotion of intimal inflammation, fever, sepsis and atherogenesis (41) | 3 | C |
| IL1B | Interleukin 1 beta | Promotion of fever, development of diabetes mellitus, apoptosis of pancreatic β-cells (87,105) | 3,4 | C, V, T |
| IL1RAP | Interleukin 1 receptor accessory protein | Induces synthesis of acute phase and proinflammatory proteins during infection, tissue damage, or stress (130) | 3 | C |
| IL3 | Interleukin 3 | Growth and differentiation of hematopoietic progenitor cells regulator and functional activator of mature neutrophils or macrophages (131) | 3,4 | V, T |
| IL33 | Interleukin 33 | Gene transcription regulator, released after cell necrosis triggering immune response and stress (132) | 3 | C |
| IL36G | Interleukin 36 gamma | Inflammasome dependent, involved in systemic inflammation (133) | 3 | C |
| IL4 | Interleukin 4 | Hematopoiesis, antibody production, inflammation response (117) | 3,4 | V, T |
| IL5 | Interleukin 5 | Eosinophil migration, activation survival (134) | 3,4 | V, T |
| IL6 | Interleukin 6 | Innate and adaptive immune response to infections (135) | 3,4 | C, V, T |
| INS | Insulin | Blood sugar regulator (136) | 3 | C |
| ITGA2B | Integrin subunit alpha 2b | Coagulation (137,138) | 3,4 | V, T |
| JAK1 | Janus kinase 1 | Cell growth survival, development differentiation of various cell types (139) | 3 | C |
| JAK2 | Janus kinase 2 | Cell growth and proliferation (139) | 3 | C, T |
| JUN | Jun proto-oncogene, AP-1 transcription factor subunit | Gene expression regulator (92) | 3 | C |
| KCNE1 | Potassium voltage-gated channel subfamily E regulatory subunit 1 | Potassium channels regulator (140,141) | 3 | C |
| KCNH2 | Potassium voltage-gated channel subfamily H member 2 | Electrical signals transmission (141) | 3 | C |
| KCNJ2 | Potassium inwardly rectifying channel subfamily J member 2 | Muscle movement (heart or skeletal) (142) | 3 | C |
| KCNQ1 | Potassium voltage-gated channel subfamily Q member 1 | Electrical signals generation and transmission (143) | 3 | C |
| LCN2 | Lipocalin 2 | Sequesters iron and preventing its use by bacteria, thus limiting their growth (144) | 3 | C |
| MMP1 | Matrix metallopeptidase 1 | Degrades collagen type I and II (145,146) | 3 | C |
| MMP2 | Matrix metallopeptidase 2 | Extracellular matrix (146) | 3 | C |
| MPL | MPL proto-oncogene, thrombopoietin receptor | Proliferator of cells involved in blood clotting (147) | 3,4 | V, T |
| MS4A1 | Membrane spanning 4-domains A1 | Regulator of cellular calcium influx necessary for the B-lymphocytes activation (148) | 3,4 | C, V |
| MS4A3 | Membrane spanning 4-domains A3 | Marker of immature circulating neutrophils, a cellular population associated to COVID-19 severe disease (148) | 3 | C |
| MUC1 | Mucin 1, cell surface associated | High viscosity of airway mucus and sputum retention in the small airway of COVID-19 patients (149) | 3 | C |
| MYD88 | MYD88 innate immune signal transduction adaptor | Initiates early immune responses (150) | 3 | C |
| NCAM1 | Neural cell adhesion molecule 1 | Molecular mimicry between NCAM-1 and the SARS-CoV-2 envelope protein (151) | 3 | C, T |
| NFAT5 | Nuclear factor of activated T-cells 5 | Protects cells against harmful effects of stress (137) | 3 | C |
| NFATC1 | Nuclear factor of activated T-cells 1 | Transcription factor (137) | 3,4 | C, V |
| NFATC2 | Nuclear factor of activated T-cells 2 | Neuroinflammatory factor (137) | 3 | C |
| NFATC3 | Nuclear factor of activated T-cells 3 | Involved in proliferation of human pulmonary fibroblasts after hypoxic stimulus (137) | 3 | C |
| NFATC4 | Nuclear factor of activated T-cells 4 | Transcriptional regulator in naive T-cells and differentiated effector T-cells, dependent on calcium/PLCγ/calmodulin/calcineurin signaling (137) | 3 | C |
| NFKB1 | Nuclear factor kappa B subunit 1 | Activated by various intra/extra-cellular stimuli as viruses (92) | 3,4 | C, V |
| NLRP3 | NLR family pyrin domain containing 3 | Intracellular sensor that detects a broad range of pathogen motifs (59) | 3,4 | C, V |
| NOD2 | Inflammatory bowel disease protein 1 | Activates NFKB1, negatively regulates TLR2 (152,153) | 3 | C, T |
| NOS1 | Nitric oxide synthase 1 | Chemical messenger (154,155) | 3 | C |
| NOS1AP | Nitric oxide synthase 1 adaptor protein | Inhibitor of Nnos (156) | 3 | C |
| NOS3 | Nitric oxide synthase 3 | Regulating vascular tone, cellular proliferation leucocyte adhesion and platelet aggregation (157,158) | 3 | C |
| NTRK1 | Neurotrophic receptor tyrosine kinase 1 | Development and survival of neurons (159) | 3,4 | V, T |
| NTRK2 | Neurotrophic receptor tyrosine kinase 2 | Development and maturation of the central and the peripheral nervous systems (159) | 3,4 | V, T |
| NTRK3 | Neurotrophic receptor tyrosine kinase 3 | Development of heart and nervous (159) | 3,4 | V, T |
| OLFM4 | Olfactomedin 4 | Facilitates cell adhesion, most probably through interaction with cell surface lectins and cadherin (160) | 3 | C |
| P2RX1 | Purinergic receptor P2X 1 | Ligand-gated ion channel with relatively high calcium permeability (161) | 3 | C |
| P2RX7 | Purinergic receptor P2X 7 | Receptor for ATP that acts as a ligand-gated ion channel (162) | 3,4 | C, V |
| PDGFA | Platelet derived growth factor subunit A | Wound healing (163) | 3 | C |
| PECAM1 | Platelet and endothelial cell adhesion molecule 1 | Cell adhesion (164) | 3 | C |
| PLAUR | Plasminogen activator, urokinase receptor | Localizing and promoting plasmin formation (165) | 3 | C |
| PPP3CB | Protein phosphatase 3 catalytic subunit beta | Transduction of intracellular Ca(2+)-mediated signals (166) | 3 | C |
| PRF1 | Perforin 1 | Defense against virus-infected cells (122) | 3 | C |
| PTGS2 | Prostaglandin-endoperoxide synthase 2 | Role in the inflammatory response (167) | 3 | C |
| PTPN11 | Protein tyrosine phosphatase non-receptor type 11 | Positively regulates MAPK signal transduction pathway (168,169) | 3 | C |
| PYCARD | PYD and CARD domain containing | Key mediator in apoptosis and inflammation (170,171) | 3,4 | C, V |
| REN | Renin | Angiotensin I from angiotensinogen generator in the plasma, initiating a cascade of reactions that produce an elevation of blood pressure and increased sodium retention by the kidney (172,173) | 3 | C |
| SCL11A2 | Natural resistance-associated macrophage protein 2 | Important in metal transport and their insertion into mitochondria (174) | 3,4 | V, T |
| SCN5A | Sodium voltage-gated channel alpha subunit 5 | Responsible for the initial upstroke of the action potential in an electrocardiogram (175) | 3 | C |
| SELE | Selectin E | Immunoadhesion (176) | 3,4 | V, T |
| SELP | Selectin P | Mediates rapid rolling of leukocyte rolling over vascular surfaces during the initial steps in inflammation through interaction with SELPLG (177) | 3,4 | V, T |
| SERPINE1 | Serpin family E member 1 | Alveolar type 2 cells senescence in the lung (178) | 3 | C, T |
| SERPINE2 | Serpin family E member 2 | Serine protease inhibitor with activity toward thrombin, trypsin, and urokinase (40) | 3 | C |
| SFTPC | Surfactant protein C | Lowering the surface tension at the air-liquid interface in the peripheral air spaces (179) | 3 | C |
| SFTPD | Surfactant protein D | May participate in the extracellular reorganization or turnover of pulmonary surfactant, regulates immune response (180) | 3 | C |
| SHC1 | SHC adaptor protein 1 | Signaling adapter that couples activated growth factor receptors to signaling pathways (181) | 3 | T |
| SIGIRR | Single Ig and TIR domain containing | Inflammation immune, response modulator (182) | 3 | C |
| SLC11A2 | Solute carrier family 11-member 2 | Metal transporter (183) | 3 | T |
| SOCS1 | Suppressor of cytokine signaling 1 | Exerts the viral virulence effect via inhibition of type I and type II interferon (IFN) function (184) | 3,4 | V, T |
| STXBP2 | Syntaxin binding protein 2 | Involved in cytolytic pathway (185) | 3 | T |
| TBK1 | TANK binding kinase 1 | Regulator of inflammatory responses to foreign agents (186) | 3 | C |
| TF | Transferrin | Transports of iron from sites of absorption and heme degradation to those of storage and utilization (187) | 3,4 | V, T |
| TFPI | Tissue factor pathway inhibitor | Anticoagulant protein blocking the initiation of blood coagulation by inhibiting TF-f VIIa and early forms of prothrombinase (188) | 3,4 | V, T |
| TFRC | Transferrin receptor | Erythropoiesis and neurologic development (189) | 3,4 | V, T |
| TGFB1 | Transforming growth factor beta 1 | Gene expression proliferation (70) | 3 | C, T |
| THPO | Thrombopoietin | Regulates platelets and macrophages differentiation (190) | 3,4 | V, T |
| TICAM1 | Toll-like receptor adaptor molecule 1 | Native immunity against invading pathogens (191) | 3 | C |
| TLR2 | Toll-like receptor 2 | Pathogen recognition-potential therapeutic target (192-194) | 3 | C, T |
| TLR4 | Toll-like receptor 4 | Upregulated after SARS-CoV-2 infection (195) | 3 | C |
| TNF | Tumor necrosis factor | Biomarker of COVID-19 severity (104) | 3,4 | C, V, T |
| TNFRSF1A | TNF receptor superfamily member 1A | Contributes to the induction of non-cytocidal TNF effects including anti-viral state and activation of the acid sphingomyelinase (93,104) | 3 | C |
| TNFRSF1B | TNF receptor superfamily member 1B | Regulates TNF-α function by antagonizing its biological activity (93,104) | 3 | C |
| TRAF3 | TNF receptor associated factor 3 | Regulates pathways leading to a NFKB and MAP kinases activation, and B-cell survival (196) | 3 | C |
| TYK2 | Tyrosine kinase 2 | Antiviral activity (197) | 3 | C |
| VCAM1 | Vascular cell adhesion molecule 1 | Mediates the adhesion of lymphocytes, monocytes, eosinophils and basophils to vascular endothelium (198) | 3,4 | V, T |
| VEGFA | Vascular endothelial growth factor A | Dominant inducer to blood vessels growth (increases their permeability) (199) | 3 | C |
| VKORC1 | Vitamin K epoxide reductase complex subunit 1 | Reduces inactive vitamin K 2,3-epoxide to active vitamin K (200) | 3 | C |
| VWF | von Willebrand factor | Involved in hemostasis and thrombosis (201) | 3,4 | C, V, T |
Entities: C, COVID-19; V, vaccine-induced thrombotic thrombocytopenia; T, thrombocytopenia.
Figure 3.

COVID-19 and thrombocytopenia interaction network. COVID-19 molecules are represented by circles; thrombocytopenia-related molecules are represented by squares; common molecules are represented by rhomboids.
Figure 4.

Overlaps between and amid all three morbid entities in Venn diagrams: (A) between COVID-19 and thrombocytopenia, (B) between VITT and thrombocytopenia, (C) between COVID-19 and VITT, and (D) amid all. VITT, vaccine-induced thrombotic thrombocytopenia.
The second one including 61 molecules, is described in Table I and illustrated in Fig. 5. Of these, 47 are common with thrombocytopenia (indicated by a polygon), and 16 with COVID-19 (represented by circles). The VITT-related molecules are denoted with triangles.
Figure 5.

COVID-19 and thrombocytopenia interactions network. VITT-related molecules are depicted by triangles; common with COVID-19 molecules are encircled; thrombocytopenia-related molecules are depicted by squares; VITT common with thrombocytopenia molecules are depicted by polygons.
Venn diagrams were further created to illustrate the nodes that are common between thrombocytopenia and COVID-19 or VITT (Fig. 4A and B, respectively), between COVID-19 and VITT (Fig. 4C), and amid the three morbid entities (Fig. 4D). The common nodes are listed in each diagram in detail.
All included molecules herein are listed in Table I. The figure (network) in which each molecule is implicated is also noted in a separate column in Table I.
Discussion
Epidemics were already identified as entities in antiquity by Hippocrates and named by him in his Treatises 'On Epidemics' (9,10). Viral epidemics were described therein and in other works of the Hippocratic Corpus (11,12). On the other hand, Aristotle, the ancient Greek physician and philosopher (4th century B.C.) wrote that 'the creativeness of nature focuses on qualities rather than quantities and description rather than measurements' (13,14). This concept was rejected by Newton's determinism and reductionism and was since forgotten, until it was re-established by Wulff in 1999 (15). Indeed, subtle change in qualities may trigger phase shift alterations with unpredictable consequences, as the Chaos theory of dynamic systems recently confirmed (16). According to this concept, the systems theory was coined as representing a rapid, cost and time-effective method of research (17). It may integrate basic, preclinical and clinical research, and both human and animal results to unravel new insights in complex and often unpredictable issues. In the case of the COVID-19 pandemic, the urgency, and certain ethical issues, make such an in silico approach a sine qua non research method.
The human-to-human transmission of SARS-CoV-2 is either mediated by respiratory droplets via sneezing/coughing or even just breathing, while the disease demonstrates an incubation period of 5-7 days (18). The clinical outcomes range from asymptomatic to influenza-like, or to even pneumonia and severe acute respiratory distress syndrome (ARDS) (19), and thromboembolic events (20,21), pointing to the lung tropism of this virus. Dissimilarities in patients' profiles are attributed to genetic and/or epigenetic variations and underlying pathologies. Dissimilarities in severity may be attributed to the aforementioned factors, but also to the size of the viral inoculum and/or viral mutations.
COVID-19 and the thrombocytopenia interactions network
Ariadne's thread appears to be the angiotensin I converting enzyme 2 (ACE2), which clearly plays a crucial role. SARS-CoV-2, via its spike S protein, a surface glycoprotein that surrounds the spherical virus, is attached to ACE2 and this is followed by entry into cells of the host (22-27). ACE2 is expressed in cells of a number of human organs (including the skin, nasal and oral mucosa, lung, nasopharynx, brain, lymph nodes, thymus, stomach, small intestine, colon, bone marrow, spleen, liver and kidneys). Additionally, its expression in lung alveoli (type 2 pneumonocytes) and small intestine endothelium, as well as in the arterial and other tissue smooth muscle epithelium (28), may trigger the release of anaphylatoxin (29). There is clinical evidence to confirm the aforementioned knowledge of COVID-19 (29).
In the generated network illustrated in Fig. 2, ACE2 interacts with CYP11B2 and with IL-6. The latter is the greatest hub in this vastly interconnected network, with 63 interactions, confirming that the progress of SARS-CoV-2-induced infection would profit from therapeutic blockade of IL-6. As noted by Mazzoni et al (24), blocking this mechanism would 'suppress noxious systemic inflammation but also restore the protective antiviral potential'. It has been established that innate immunity via natural killer (NK) cells exerts the frontline defense, with CD8+ T-lymphocytes being important for the long-term surveillance against viruses, while adaptive immune responses play a key role in the control of viral infections (28). Both responses are mediated either via cytotoxicity or by IF-γ, IL-12 and IL-18. Virus-induced cytotoxicity is primarily moderated by perforin and granzymes. Increased severity in viral infections may lead to dysregulated immunity and tissue/organ damage (30). Clinical evidence in SARS-CoV-2 infection has demonstrated that high IL-6 levels in patients in intensive care units, are inversely associated with the concentration of NK cells (24,31).
The network included dense interactions illustrating clearly that SARS-CoV-2-specific T-cells are critical for the extended damage caused by the 'cytokine storm' (or 'cytokine release syndrome') (30,32) (Fig. 3). This excessive inflammatory response may be lethal for some patients (29,33). Although the phenomenon may manifest in other inflammatory conditions, including bacterial sepsis, pneumonia, sterile inflammation, etc., the extent in the secretion of several specific cytokines is different in COVID-19-related storm (29). Of note, COVID-19 infection has been associated with changes in the blood coagulation mechanisms, with differing manifestations in different patients, in distinct phases of the disease, and independently of disease severity.
Autoimmune destruction of platelets, cytokine release and high consumption of coagulation factors and platelets have been observed in patients with SARS-CoV-2 infection (Geronikolou et al, unpublished data) and initial hypercoagulability (34). Thromboembolic events increase by 31% in patients with COVID-19 admitted in intensive care units (35,36); the phenomenon may be interpreted by the 'two way activation theory' (20,37), i.e., thrombogenesis via inflammation-relevant pathways, with parallel occurrence of release of VWF large polymers. The coagulation and platelet profiles of patients with COVID-19 are then rather normal, unlike in patients with sepsis where platelets are activated and consumed, with the occurrence of thrombocytopenia (38). Only a few patients may then survive, particularly of those with extensive disseminated intravascular coagulation (38). Thrombosis has been observed in situ in the lungs, as well as in a systemic manner, in a similar fashion with classic sepsis and acute respiratory distress syndrome. Reported thromboembolic complications include mostly venous pulmonary embolism (38), aortic graft thrombosis, and mesenteric ischemia; coronary and cerebral thrombosis cases have been reported, although these are rare. The so-called 'COVID toe' is a sign of thrombosis accompanied by arterial and venous clots, urgent oxygen demand and multiple organ dysfunction (20,36,39).
COVID-19 and thrombocytopenia interactomes share only 14 nodes (AIM2, IFI16, TLR2, NOD2, NKAM1, IL-6, TNF, JAK2, IL-1B, SERPINE1, HLA-DRB1, TGFB1, CD8A, and VWF) (Fig. 3), most of which serve as major hubs (IL-6, TNF, JAK2, IL-1B, SERPINE1, TGFB1, CD8A and VWF) in the herein presented interactome (Figs. 1 and 2).
Cytokines, such as IL-1B, 1L-6 and TNF contribute to the so-called cytokine storm, as aforementioned. Moreover, JAK2 is a kinase suspected to be implicated in thrombocytopenia via reduced levels of thrombopoietin or via decreased expression levels of their cognate receptors (cMpl receptors). JAK2 mutations (V617F) that are present in the majority of patients with myeloproliferative disease, may increase hematopoietic cell sensitivity to erythropoietin and thrombopoietin. NKAM1 or CD56 is a homophilic binding glycoprotein expressed on the surface of neurons, glia cells and skeletal muscles. NKAM1 is a prototypic marker of NK cells, also present in CD8+ T-cells. These cell types exhibit diminished antiviral ability and cytotoxic impairment during COVID-19 infection (24). CD8A1 is a cytotoxic marker for T-cell populations. SERPINE1 or plasminogen activator inhibitor-1 is a protein encoded by the SERPINE1 gene, which participates in both thrombosis and atherogenesis (40).
TGFB1 is a multifunctional peptide, with diverse activities, including the control of cell growth, proliferation, differentiation, and apoptosis. It can also down-regulate the activity of immune cells via decreasing the expression levels of cytokine receptors, such as that of IL-2. Several types of T-cells secrete TGFB1, so as to inhibit cytotoxicity and the secretion of certain cytokines, such as interferon-γ, TNF-α and various interleukins, such as IL-6. This makes this molecule a potential target of therapeutic value. On the other hand, the hemostatic VWF is detected in blood plasma, endothelium and megakaryocytes, as well as in subendothelial connective tissue. This factor appears to be also increased and implicated in autoimmune diseases, such as thrombotic thrombocytopenic purpura, as well as in stroke and atrial fibrillation, due to the platelet clots that are potentially formed when its levels are elevated.
Recent literature has further revealed that an HLA class I and II molecule, that is, HLA-DRB1, which is common in COVID-19 and in thrombocytopenia networks (Fig. 2), may play a role in the observed COVID-19 individual and ethnic diversity in clinical severity and/or response to therapy or vaccination (41-44). Of note, HLA-DRB1 is interconnected with the lymphocyte function markers CD3D, CD3E, CD3G, CD4, lymphocyte regulation positive FCGR1A, FCGR1B, HLA class I and II molecules, such as HLA-A, HLA-B, similar to the NCAM1, PTPN1, SHC1 and VCAM1 molecules that have been implicated in thrombosis and atherosclerosis. NCAM1 is involved in cell-cell adhesion in neural-muscle cells in the embryonic phase and later, and more notably, in the responsiveness to viral infections (rabies virus and papilloma virus) (45). PTPN1 is a potential therapeutic target of obesity and type 2 diabetes mellitus as well (46); SHC1 is implicated in reactive oxygen species regulation, thus, in the oxidative stress response (47), while VCAM1 is directly involved in thrombosis and atherogenesis and acute respiratory syndrome (48-51).
VITT and thrombocytopenia interactome
Various coagulation mechanisms have been implicated in VITT: High levels of D-dimers and low levels of fibrinogen have been observed in patients (2,52,53). On the other hand, early reports of VITT described a higher incidence of the syndrome in young women, exhibiting both age-dependence and sexual dimorphism. VITT, though very rare, is of utmost importance. Yet, in March, 2021, the European Medicines Agency (EMA) issued a statement noting that the thromboembolic events of VITT in vaccinated populations were not higher than in general population (54). Subsequently, the 'risk vs. benefit' equilibrium was weighed by the World Health Organization (WHO), promoting the benefit of the vaccination vs. the extremely low risk of thromboembolic risk of VITT in the general population (55).
VITT is currently termed 'thrombosis with thrombocytopenia syndrome (TTS)' by the Centers for Disease Control and Prevention (CDC) and the US Food and Drug Administration (FDA) (56), and is characterized by arterial and venous thrombosis at unexpected sites (i.e., cerebral venous sinuses, splanchnic vessels of variant severity and/or positive platelet factor (PF) 4-heparin ELISA ('HIT' ELISA) syndrome (52), exhibiting both age dependence and sexual dimorphism (more frequent in individuals <50 years old and of the female sex) (2). The laboratory and clinical features of this syndrome are similar to those of the heparin-induced thrombocytopenia (HIT) syndrome and/or the HIT-like autoimmune thrombosis with thrombocytopenia syndrome (2,52,53), both of which have already been reported following surgery, the uptake of certain pharmaceuticals, or during some infections in patients that are not being treated with heparin. The therapeutic suggestions of this recently coined syndrome include early initiation of non-heparin anticoagulation, high-dose IVIG, and/or prednisolone (57).
The genetic basis of the VITT syndrome appears to be closely intertwined with that of the COVID-19 disease and, as such, they share 16 nodes: CASP1, CXCL8, FGF7, HLA-A, HLA-B, IL1B, IL6, MS4A1, NFATC1, NFKB1, NLP3, P2RX7, PYCARD, TNF, TFP1, VWF (Figs. 3-5). The purpose of the vaccine is to inhibit pathways that mediate this condition (52,58). More importantly, the relevant research is ongoing with the extremely rare cases of this syndrome, as VITT incidence is ~0.74-1 cases per 100,000 vaccinated subjects (52). Of note, the anti-COVID-19 vaccines do not cause illness and the two morbid entities (COVID-19 and VITT) are by no means identical, with the etiopathology of the latter being actually autoimmune, with auto-antibodies against platelet factor 4. More explicitly, COVID-19 network shares 14 nodes with thrombocytopenia (AIM2, CD8A, HLA-DRB1, IFI16, IL1B, IL6, JAK2, NCAM1, NOD2, SERPINE1, TGFB1, TLR2, TNF and VWF), while VITT (which is a type of thrombocytopenia) shares 46 nodes with thrombocytopenia (Figs. 3-5). Notably, SHC1, STXBP2, CDC20 and ADAM10 are silenced in VITT, while AURKA, CD46, CD19 are uniquely expressed following vaccination (apparently not expressed in common thrombocytopenia or in COVID-19) (Figs. 3-5). These molecules were not previously identified as VITT-related and are, thus, a novel finding, at least to the best of our knowledge.
It is known that the NLP3 inflammasome is implicated in both COVID-19 and VITT, apart from its participation in other inflammatory reactions (59). It has also been previously demonstrated that acute thrombotic events may manifest during hypoxia, as shown in COVID-19, due to an early proinflammatory state in the venous milieu, mediated by a HIF-induced NLP3 inflammasome complex (60,61). In the network constructed in the present study, NLP3 connects with CASP1, IL-IB, IL17A, CXCL8, IL-6, MYD88, NFKB1, P2RX7, PYCARD and TNF.
P2RX7 exhibits sexually dimorphic and contrasting roles in the pathogenesis of thrombosis, depending on the pathogen type, the severity of infection, the cell type infected and the level of tissue activation (62). In the thrombocytopenia/ COVID-19/VITT cases, the viral load, the cell-type infected and the infecting virus strain or certain vaccine types have been associated with NLP3 hyperactivation, which in the presence of comorbidities, such as liver, renal, gut or respiratory tract illnesses, diabetes mellitus, previous infections, exposure to pollutants, and/or lifestyle factors, such as smoking and obesity, may upend the roles of P2RX7 and PYCARD to those of tissue-damaging, or even lethal factors (62,63). More importantly, the persistent neurological effects ('long-COVID-19') observed in a large percentage of patients with COVID-19 may be explained via the activation of these pathways. Thus, P2RX7 antagonists may be promising therapeutics in the management of both VITT and 'long-COVID-19' (62,64), as P2RX7 receptor stimulation has been implicated in lung damage, psychiatric disorders and pathological inflammation (65,66). In the COVID-19 interactome, P2RX7 directly interacts with NLP3, CASP1 and P2RX1. On the contrary, in the VITT network, P2RX7 directly interacts only with NLP3, IL1B and CASP1. Accordingly, PYCARD interacts with NLP3, CASP1, IL1B, IL18 and IKBKG in COVID-19, and with NLP3, CASP1 and IL1B in the VITT syndrome (Table II). The common node in all possible combinations, as shown in Table II, is CASP1, a downstream event of the NLP3 inflammasome; CASP1 activation promotes IL1B production, which may be prevented by a pan-caspase inhibitor or by glyburide treatment (67).
Table II.
Common direct connections between 'PYCARD' or 'P2RX7' and 'COVID-19' or 'VITT'.
| Gene | COVID-19 | VITT | COMMON direct connections |
|---|---|---|---|
| PYCARD | NLP3, CASP1, IL1B, IL18, IKBKG | NLP3, CASP1, IL1B | NLP3, CASP1, IL1B |
| P2RX7 | NLP3, CASP1, P2RX1 | CASP1, IL1B | CASP1 |
| COMMON direct connections | NLP3, CASP1 | CASP1, IL1B | CASP1 |
To this end, the present study investigated the aforementioned issues through the construction of molecular networks and the detection of at least one known COVID/VITT/thrombocytopenia molecule that confirmed that endothelial dysfunction and blood thrombosis are the key players of both COVID-19 and VITT morbid entities. One limitation of the present study is that it included only wild-type genes and their products. To the best of our knowledge, however, this is the first effort made at providing a comprehensive network map of the molecules involved in the underlying mechanisms of COVID-19, long COVID-19 and/or VITT pathophysiology.
In conclusion, the interactomes presented herein revealed therapeutic and vaccination modification targets (i.e., SHC1, NCAM1, HLAs, CD8A, PTPN1, VWF and TBP1). It was also demonstrated that: i) NCAM1 is involved in SARS-CoV-2 infection responsiveness, apart from papilloma and rabies virus infections, and may be responsible for relevant vaccination side effects; ii) NLP3, P2RX7 and PYCARD contribution may help explain (partly or mostly) VITT and/or 'long COVID-19 side-effects'; iii) furthermore, the antagonism of these latter nodes should focus on potential pharmacological targets in the context of SARS-CoV-2 infection and/or vaccine immunization responsiveness. In conclusion, network construction is a powerful tool, which may be used to elucidate the physiology and pathophysiology of different states in clinical investigation. The highly interconnected network presented herein highlights the complexity of COVID-19/VITT pathophysiology, mapping the key role of cytokines, enzymes and immune response markers (lymphocyte regulators and human leucocyte antigens) that may be potential drug or vaccine targets. It was constructed using wild-type genes and gene products, revealing the body's predisposition to COVID-19 infection or VITT. Of note, the COVID-19 and thrombocytopenia common nodes appear to be key players in the natural history of the illness.
Acknowledgments
Not applicable.
Funding Statement
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current study are available throughout the manuscript.
Authors' contributions
SAG and MM were involved in the conceptualization of the study. SAG was involved in the study methodology. SAG, AP and MM were involved in data validation. SAG and AP was involved in formal analysis and in the investigative aspects of the study. SAG was involved in the provision of resources (study material). SAG, IT and AP was involved in data curation. IT provided the software used in this study. SAG, IT, GPC and AP were involved in the interpretation of the data, and in the writing and preparation of the original draft. SAG, AP, MM and GPC were involved in the writing, reviewing and editing of the manuscript. MM and GPC supervised the study. SAG and GPC were involved in project administration. All authors confirm the authenticity of the raw data and have read and agreed to the published version of the manuscript.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.World Health Organization (WHO) Coronovirus disease (COVID-19): Vaccines safety. WHO; Geneva: 2021. [Google Scholar]
- 2.Greinacher A, Thiele T, Warkentin TE, Weisser K, Kyrle PA, Eichinger S. Thrombotic thrombocytopenia after ChAdOx1 nCov-19 vaccination. N Engl J Med. 2021;384:2092–2101. doi: 10.1056/NEJMoa2104840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mathieu E, Ritchie H, Ortiz-Ospina E, Roser M, Hasell J, Appel C, Giattino C, Rodés-Guirao L. A global database of COVID-19 vaccinations. Nat Hum Behav. 2021;5:947–953. doi: 10.1038/s41562-021-01122-8. [DOI] [PubMed] [Google Scholar]
- 4.Wei CH, Allot A, Leaman R, Lu Z. PubTator central: Automated concept annotation for biomedical full text articles. Nucleic Acids Res. 2019;47(W1):W587–W593. doi: 10.1093/nar/gkz389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chen Q, Allot A, Lu Z. LitCovid: An open database of COVID-19 literature. Nucleic Acids Res. 2021;49(D1):D1534–D1540. doi: 10.1093/nar/gkaa952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork P, et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019;47(D1):D607–D613. doi: 10.1093/nar/gky1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Szklarczyk D, Gable AL, Nastou KC, Lyon D, Kirsch R, Pyysalo S, Doncheva NT, Legeay M, Fang T, Bork P, et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021;49(D1):D605–D612. doi: 10.1093/nar/gkaa1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13:2498–2504. doi: 10.1101/gr.1239303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Smith Wesley D., editor. Loeb Classical Library 477. Harvard University Press; Cambridge, MA: 1994. Hippocrates: Epidemics 2, 4-7. [Google Scholar]
- 10.Jouanna J. Hippocrates. John Hopkins University Press; Baltimore, MD: 1999. [Google Scholar]
- 11.Mammas IN, Spandidos DA. Paediatric virology in the Hippocratic corpus. Exp Ther Med. 2016;12:541–549. doi: 10.3892/etm.2016.3420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pappas G, Kiriaze IJ, Falagas ME. Insights into infectious disease in the era of Hippocrates. Int J Infect Dis. 2008;12:347–350. doi: 10.1016/j.ijid.2007.11.003. [DOI] [PubMed] [Google Scholar]
- 13.Misselbrook D. Aristotle, hume and the goals of medicine. J Eval Clin Pract. 2016;22:544–549. doi: 10.1111/jep.12371. [DOI] [PubMed] [Google Scholar]
- 14.Wulff HR. The concept of disease: From Newton back to Aristotle. Lancet. 1999;354(Suppl):SIV50. doi: 10.1016/S0140-6736(99)90393-8. [DOI] [PubMed] [Google Scholar]
- 15.Wulff HR. The concept of disease: From Newton back to Aristotle. Lancet. 1999;54:354. doi: 10.1016/s0140-6736(99)90393-8. [DOI] [PubMed] [Google Scholar]
- 16.Lorenz EN. Deterministic nonperiodic flow. J Atmos Sci. 1963;20:130–141. doi: 10.1175/1520-0469(1963)020<0130:DNF>2.0.CO;2. [DOI] [Google Scholar]
- 17.Barabási AL, Gulbahce N, Loscalzo J. Network medicine: A network-based approach to human disease. Nat Rev Genet. 2011;12:56–68. doi: 10.1038/nrg2918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Li Q, Guan X, Wu P, Wang X, Zhou L, Tong Y, Ren R, Leung KSM, Lau EHY, Wong JY, et al. Early transmission dynamics in Wuhan, China, of novel coronavirus-infected pneumonia. N Engl J Med. 2020;382:1199–1207. doi: 10.1056/NEJMoa2001316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Raoult D, Zumla A, Locatelli F, Ippolito G, Kroemer G. Coronavirus infections: Epidemiological, clinical and immunological features and hypotheses. Cell Stress. 2020;4:66–75. doi: 10.15698/cst2020.04.216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mondal S, Quintili AL, Karamchandani K, Bose S. Thromboembolic disease in COVID-19 patients: A brief narrative review. J Intensive Care. 2020;8:70. doi: 10.1186/s40560-020-00483-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Xu P, Zhou Q, Xu J. Mechanism of thrombocytopenia in COVID-19 patients. Ann Hematol. 2020;99:1205–1208. doi: 10.1007/s00277-020-04019-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li W, Moore MJ, Vasilieva N, Sui J, Wong SK, Berne MA, Somasundaran M, Sullivan JL, Luzuriaga K, Greenough TC, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426:450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ge XY, Li JL, Yang XL, Chmura AA, Zhu G, Epstein JH, Mazet JK, Hu B, Zhang W, Peng C, et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature. 2013;503:535–538. doi: 10.1038/nature12711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mazzoni A, Salvati L, Maggi L, Capone M, Vanni A, Spinicci M, Mencarini J, Caporale R, Peruzzi B, Antonelli A, et al. Impaired immune cell cytotoxicity in severe COVID-19 is IL-6 dependent. J Clin Invest. 2020;130:4694–4703. doi: 10.1172/JCI138554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sama IE, Ravera A, Santema BT, van Goor H, Ter Maaten JM, Cleland JGF, Rienstra M, Friedrich AW, Samani NJ, Ng LL, et al. Circulating plasma concentrations of angiotensin-converting enzyme 2 in men and women with heart failure and effects of renin-angiotensin-aldosterone inhibitors. Eur Heart J. 2020;41:1810–1817. doi: 10.1093/eurheartj/ehaa373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Diaz JH. Hypothesis: Angiotensin-converting enzyme inhibitors and angiotensin receptor blockers may increase the risk of severe COVID-19. J Travel Med. 2020;27:taaa041. doi: 10.1093/jtm/taaa041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A, et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;18:271–280.e8. doi: 10.1016/j.cell.2020.02.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hamming I, Timens W, Bulthuis ML, Lely AT, Navis G, van Goor H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J Pathol. 2004;203:631–637. doi: 10.1002/path.1570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gao T, Hu M, Zhang X, Li H, Zhu L, Liu H, Dong Q, Zhang Z, Wang Z, Hu Y, et al. Highly pathogenic coronavirus N protein aggravates lung injury by MASP-2-mediated complement over-activation. medRxiv. 2020 ppmedrxiv-20041962. [Google Scholar]
- 30.Cao X. COVID-19: Immunopathology and its implications for therapy. Nat Rev Immunol. 2020;20:269–270. doi: 10.1038/s41577-020-0308-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Channappanavar R, Perlman S. Pathogenic human coronavirus infections: Causes and consequences of cytokine storm and immunopathology. Semin Immunopathol. 2017;39:529–539. doi: 10.1007/s00281-017-0629-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhao J, Zhao J, Perlman S. T cell responses are required for protection from clinical disease and for virus clearance in severe acute respiratory syndrome coronavirus-infected mice. J Virol. 2010;84:9318–9325. doi: 10.1128/JVI.01049-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Meduri GU, Kohler G, Headley S, Tolley E, Stentz F, Postlethwaite A. Inflammatory cytokines in the BAL of patients with ARDS. Persistent elevation over time predicts poor outcome. Chest. 1995;108:1303–1314. doi: 10.1378/chest.108.5.1303. [DOI] [PubMed] [Google Scholar]
- 34.Tang N, Li D, Wang X, Sun Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J Thromb Haemost. 2020;18:844–847. doi: 10.1111/jth.14768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Helms J, Tacquard C, Severac F, Leonard-Lorant I, Ohana M, Delabranche X, Merdji H, Clere-Jehl R, Schenck M, Fagot Gandet F, et al. High risk of thrombosis in patients with severe SARS-CoV-2 infection: A multicenter prospective cohort study. Intensive Care Med. 2020;46:1089–1098. doi: 10.1007/s00134-020-06062-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Klok FA, Kruip MJHA, van der Meer NJM, Arbous MS, Gommers DAMPJ, Kant KM, Kaptein FHJ, van Paassen J, Stals MAM, Huisman MV, Endeman H. Incidence of thrombotic complications in critically ill ICU patients with COVID-19. Thromb Res. 2020;191:145–147. doi: 10.1016/j.thromres.2020.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chang JC. Hemostasis based on a novel 'two-path unifying theory' and classification of hemostatic disorders. Blood Coagul Fibrinolysis. 2018;29:573–584. doi: 10.1097/MBC.0000000000000765. [DOI] [PubMed] [Google Scholar]
- 38.Chang JC. Sepsis and septic shock: Endothelial molecular pathogenesis associated with vascular microthrombotic disease. Thromb J. 2019;17:10. doi: 10.1186/s12959-019-0198-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Seirafianpour F, Sodagar S, Pour Mohammad A, Panahi P, Mozafarpoor S, Almasi S, Goodarzi A. Cutaneous manifestations and considerations in COVID-19 pandemic: A systematic review. Dermatol Ther. 2020;33:e13986. doi: 10.1111/dth.13986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Vaughan DE. PAI-1 and atherothrombosis. J Thromb Haemost. 2005;3:1879–1883. doi: 10.1111/j.1538-7836.2005.01420.x. [DOI] [PubMed] [Google Scholar]
- 41.Badary OA. Pharmacogenomics and COVID-19: Clinical implications of human genome interactions with repurposed drugs. Pharmacogenomics J. 2021;21:275–284. doi: 10.1038/s41397-021-00209-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chen MR, Kuo HC, Lee YJ, Chi H, Li SC, Lee HC, Yang KD. Phenotype, susceptibility, autoimmunity, and immunotherapy between Kawasaki disease and coronavirus disease-19 associated multisystem inflammatory syndrome in children. Front Immunol. 2021;12:632890. doi: 10.3389/fimmu.2021.632890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Romero-López JP, Carnalla-Cortés M, Pacheco-Olvera DL, Ocampo-Godínez JM, Oliva-Ramírez J, Moreno-Manjón J, Bernal-Alferes B, López-Olmedo N, García-Latorre E, Domínguez-López ML, et al. A bioinformatic prediction of antigen presentation from SARS-CoV-2 spike protein revealed a theoretical correlation of HLA-DRB1*01 with COVID-19 fatality in Mexican population: An ecological approach. J Med Virol. 2021;93:2029–2038. doi: 10.1002/jmv.26561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Anzurez A, Naka I, Miki S, Nakayama-Hosoya K, Isshiki M, Watanabe Y, Nakamura-Hoshi M, Seki S, Matsumura T, Takano T, et al. Association of HLA-DRB1*09:01 with severe COVID-19. HLA. 2021;98:37–42. doi: 10.1111/tan.14256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Rotondo JC, Bosi S, Bassi C, Ferracin M, Lanza G, Gafà R, Magri E, Selvatici R, Torresani S, Marci R, et al. Gene expression changes in progression of cervical neoplasia revealed by microarray analysis of cervical neoplastic keratinocytes. J Cell Physiol. 2015;230:806–812. doi: 10.1002/jcp.24808. [DOI] [PubMed] [Google Scholar]
- 46.Combs AP. Recent advances in the discovery of competitive protein tyrosine phosphatase 1B inhibitors for the treatment of diabetes, obesity, and cancer. J Med Chem. 2010;53:2333–2344. doi: 10.1021/jm901090b. [DOI] [PubMed] [Google Scholar]
- 47.Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing. Nature. 2000;408:239–247. doi: 10.1038/35041687. [DOI] [PubMed] [Google Scholar]
- 48.Choi YM, Kwon HS, Choi KM, Lee WY, Hong EG. Short-term effects of beraprost sodium on the markers for cardiovascular risk prediction in type 2 diabetic patients with microalbuminuria. Endocrinol Metab (Seoul) 2019;34:398–405. doi: 10.3803/EnM.2019.34.4.398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Nomura S, Taniura T, Shouzu A, Omoto S, Suzuki M, Okuda Y, Ito T. Effects of sarpogrelate, eicosapentaenoic acid and pitavastatin on arterioslcerosis obliterans-related biomarkers in patients with type 2 diabetes (SAREPITASO study) Vasc Health Risk Manag. 2018;14:225–232. doi: 10.2147/VHRM.S171143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zheng Y, Liu SQ, Sun Q, Xie JF, Xu JY, Li Q, Pan C, Liu L, Huang YZ. Plasma microRNAs levels are different between pulmonary and extrapulmonary ARDS patients: A clinical observational study. Ann Intensive Care. 2018;8:23. doi: 10.1186/s13613-018-0370-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Attia EF, Jolley SE, Crothers K, Schnapp LM, Liles WC. Soluble vascular cell adhesion molecule-1 (sVCAM-1) is elevated in bronchoalveolar lavage fluid of patients with acute respiratory distress syndrome. PLoS One. 2016;11:e0149687. doi: 10.1371/journal.pone.0149687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Cines DB, Bussel JB. SARS-CoV-2 vaccine-induced immune thrombotic thrombocytopenia. N Engl J Med. 2021;384:2254–2256. doi: 10.1056/NEJMe2106315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Schultz NH, Sørvoll IH, Michelsen AE, Munthe LA, Lund-Johansen F, Ahlen MT, Wiedmann M, Aamodt AH, Skattør TH, Tjønnfjord GE, Holme PA. Thrombosis and thrombocytopenia after ChAdOx1 nCoV-19 vaccination. N Engl J Med. 2021;384:2124–2130. doi: 10.1056/NEJMoa2104882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.European Medicines Agency (EMA) COVID-19 Vaccine AstraZeneca: PRAC investigating cases of thromboembolic events-vaccine's benefits currently still outweigh risks-update. 2021 [Google Scholar]
- 55.World Health Organization (WHO) Statement of the WHO global advisory committee on vaccine safety (GACVS) COVID-19 subcommittee on safety signals related to the AstraZeneca COVID-19 vaccine. WHO; Geneva: 2021. [Google Scholar]
- 56.Bussel JB, Connors JM, Cines DB, Dunbar CE, Michaelis LC, Kreuziger LB, Lee AYY, Pabinger-Fasching I. Thrombosis with thrombocytopenia syndrome (also termed vaccine-induced thrombotic thrombocytopenia) American Society of Haematology; Washington, DC: 2021. [Google Scholar]
- 57.Thaler J, Ay C, Gleixner KV, Hauswirth AW, Cacioppo F, Grafeneder J, Quehenberger P, Pabinger I, Knöbl P. Successful treatment of vaccine-induced prothrombotic immune thrombocytopenia (VIPIT) J Thromb Haemost. 2021;19:1819–1822. doi: 10.1111/jth.15346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Smadja DM, Mentzer SJ, Fontenay M, Laffan MA, Ackermann M, Helms J, Jonigk D, Chocron R, Pier GB, Gendron N, et al. COVID-19 is a systemic vascular hemopathy: Insight for mechanistic and clinical aspects. Angiogenesis. 2021;24:755–788. doi: 10.1007/s10456-021-09805-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kashir J, Ambia AR, Shafqat A, Sajid MR, AlKattan K, Yaqinuddin A. Scientific premise for the involvement of neutrophil extracellular traps (NETs) in vaccine-induced thrombotic thrombocytopenia (VITT) J Leukoc Biol. 2021 Sep 1; doi: 10.1002/JLB.5COVR0621-320RR. Epub ahead of prin. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Gupta N, Sahu A, Prabhakar A, Chatterjee T, Tyagi T, Kumari B, Khan N, Nair V, Bajaj N, Sharma M, Ashraf MZ. Activation of NLRP3 inflammasome complex potentiates venous thrombosis in response to hypoxia. Proc Natl Acad Sci USA. 2017;114:4763–4768. doi: 10.1073/pnas.1620458114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Salaro E, Rambaldi A, Falzoni S, Amoroso FS, Franceschini A, Sarti AC, Bonora M, Cavazzini F, Rigolin GM, Ciccone M, et al. Involvement of the P2X7-NLRP3 axis in leukemic cell proliferation and death. Sci Rep. 2016;6:26280. doi: 10.1038/srep26280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ribeiro DE, Oliveira-Giacomelli Á, Glaser T, Arnaud-Sampaio VF, Andrejew R, Dieckmann L, Baranova J, Lameu C, Ratajczak MZ, Ulrich H. Hyperactivation of P2X7 receptors as a culprit of COVID-19 neuropathology. Mol Psychiatry. 2021;26:1044–1059. doi: 10.1038/s41380-020-00965-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Savio LEB, de Andrade Mello P, da Silva CG, Coutinho-Silva R. The P2X7 receptor in inflammatory diseases: Angel or demon. Front Pharmacol. 2018;9:52. doi: 10.3389/fphar.2018.00052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Pacheco PAF, Faria RX. The potential involvement of P2X7 receptor in COVID-19 pathogenesis: A new therapeutic target? Scand J Immunol. 2021;93:e12960. doi: 10.1111/sji.12960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ortiz GG, Pacheco-Moisés FP, Macías-Islas M, Flores-Alvarado LJ, Mireles-Ramírez MA, González-Renovato ED, Her nández-Nava r ro VE. Role of the blood-brain barrier in multiple sclerosis. Arch Med Res. 2014;45:687–697. doi: 10.1016/j.arcmed.2014.11.013. [DOI] [PubMed] [Google Scholar]
- 66.Di Virgilio F, Tang Y, Sarti AC, Rossato M. A rationale for targeting the P2X7 receptor in coronavirus disease 19. Br J Pharmacol. 2020;177:4990–4994. doi: 10.1111/bph.15138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Ferreira AC, Soares VC, de Azevedo-Quintanilha IG, Dias SDSG, Fintelman-Rodrigues N, Sacramento CQ, Mattos M, de Freitas CS, Temerozo JR, Teixeira L, et al. SARS-CoV-2 engages inflammasome and pyroptosis in human primary monocytes. Cell Death Discov. 2021;7:43. doi: 10.1038/s41420-021-00428-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Moss ML, Bartsch JW. Therapeutic benefits from targeting of ADAM family members. Biochemistry. 2004;43:7227–7235. doi: 10.1021/bi049677f. [DOI] [PubMed] [Google Scholar]
- 69.Souza JSM, Lisboa ABP, Santos TM, Andrade MVS, Neves VBS, Teles-Souza J, Jesus HNR, Bezerra TG, Falcão VGO, Oliveira RC, Del-Bem LE. The evolution of ADAM gene family in eukaryotes. Genomics. 2020;112:3108–3116. doi: 10.1016/j.ygeno.2020.05.010. [DOI] [PubMed] [Google Scholar]
- 70.Xu J, Xu X, Jiang L, Dua K, Hansbro PM, Liu G. SARS-CoV-2 induces transcriptional signatures in human lung epithelial cells that promote lung fibrosis. Respir Res. 2020;21:182. doi: 10.1186/s12931-020-01445-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Katneni UK, Alexaki A, Hunt RC, Schiller T, DiCuccio M, Buehler PW, Ibla JC, Kimchi-Sarfaty C. Coagulopathy and thrombosis as a result of severe COVID-19 infection: A microvascular focus. Thromb Haemost. 2020;120:1668–1679. doi: 10.1055/s-0040-1715841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Tian J, Sun D, Xie Y, Liu K, Ma Y. Network pharmacology-based study of the molecular mechanisms of Qixuekang in treating COVID-19 during the recovery period. Int J Clin Exp Pathol. 2020;13:2677–2690. [PMC free article] [PubMed] [Google Scholar]
- 73.Boron WF, Boulpaep EL. Medical physiology: A cellular and molecular approach. Saunders Elsevier; Philadelphia, PA: 2012. [Google Scholar]
- 74.Fitzpatrick D, Purves D, Augustine G. Neuroscience. 3rd edition. Sinauer Associates, Inc; Sunderland, MA: 2004. [Google Scholar]
- 75.Wang Q, Zhu W, Xiao G, Ding M, Chang J, Liao H. Effect of AGER on the biological behavior of non-small cell lung cancer H1299 cells. Mol Med Rep. 2020;22:810–818. doi: 10.3892/mmr.2020.11176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Man SM, Karki R, Kanneganti TD. AIM2 inflammasome in infection, cancer, and autoimmunity: Role in DNA sensing, inflammation, and innate immunity. Eur J Immunol. 2016;46:269–280. doi: 10.1002/eji.201545839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Bafunno V, Firinu D, D'Apolito M, Cordisco G, Loffredo S, Leccese A, Bova M, Barca MP, Santacroce R, Cicardi M, et al. Mutation of the angiopoietin-1 gene (ANGPT1) associates with a new type of hereditary angioedema. J Allergy Clin Immunol. 2018;141:1009–1017. doi: 10.1016/j.jaci.2017.05.020. [DOI] [PubMed] [Google Scholar]
- 78.PubMed Gene database: ANGPT2 angiopoietin 2 [Homo sapiens (human)] https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=ShowDetailView&TermToSearch=285 Accessed December 12, 2020.
- 79.Marumoto T, Honda S, Hara T, Nitta M, Hirota T, Kohmura E, Saya H. Aurora-A kinase maintains the fidelity of early and late mitotic events in HeLa cells. J Biol Chem. 2003;278:51786–51795. doi: 10.1074/jbc.M306275200. [DOI] [PubMed] [Google Scholar]
- 80.Li N, Zhang J, Liao D, Yang L, Wang Y, Hou S. Association between C4, C4A, and C4B copy number variations and susceptibility to autoimmune diseases: A meta-analysis. Sci Rep. 2017;7:42628. doi: 10.1038/srep42628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Horiuchi T, Tsukamoto H. Complement-targeted therapy: Development of C5- and C5a-targeted inhibition. Inflamm Regen. 2016;36:11. doi: 10.1186/s41232-016-0013-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Hobart MJ, Fernie BA, DiScipio RG. Structure of the human C7 gene and comparison with the C6, C8A, C8B, and C9 genes. J Immunol. 1995;154:5188–5194. [PubMed] [Google Scholar]
- 83.Xia S, Zhang Z, Magupalli VG, Pablo JL, Dong Y, Vora SM, Wang L, Fu TM, Jacobson MP, Greka A, et al. Gasdermin D pore structure reveals preferential release of mature interleukin-1. Nature. 2021;593:607–611. doi: 10.1038/s41586-021-03478-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Cohen GM. Caspases: The executioners of apoptosis. Biochem J. 1997;326:1–16. doi: 10.1042/bj3260001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Avrutsky MI, Troy CM. Caspase-9: A multimodal therapeutic target with diverse cellular expression in human disease. Front Pharmacol. 2021;12:701301. doi: 10.3389/fphar.2021.701301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Singh S, Anshita D, Ravichandiran V. MCP-1: Function, regulation, and involvement in disease. Int Immunopharmacol. 2021;101:107598. doi: 10.1016/j.intimp.2021.107598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Coperchini F, Chiovato L, Ricci G, Croce L, Magri F, Rotondi M. The cytokine storm in COVID-19: Further advances in our understanding the role of specific chemokines involved. Cytokine Growth Factor Rev. 2021;58:82–91. doi: 10.1016/j.cytogfr.2020.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Guan E, Wang J, Norcross MA. Identification of human macrophage inflammatory proteins 1alpha and 1beta as a native secreted heterodimer. J Biol Chem. 2001;276:12404–12409. doi: 10.1074/jbc.M006327200. [DOI] [PubMed] [Google Scholar]
- 89.Charrier A, Brigstock DR. Regulation of pancreatic function by connective tissue growth factor (CTGF, CCN2) Cytokine Growth Factor Rev. 2013;24:59–68. doi: 10.1016/j.cytogfr.2012.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Garcillán B, Fuentes P, Marin AV, Megino RF, Chacon-Arguedas D, Mazariegos MS, Jiménez-Reinoso A, Muñoz-Ruiz M, Laborda RG, Cárdenas PP, et al. CD3G or CD3D knockdown in mature, but not immature, T lymphocytes similarly cripples the human TCRαβ complex. Front Cell Dev Biol. 2021;9:608490. doi: 10.3389/fcell.2021.608490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Heritable gene regulation in the CD4:CD8 T cell lineage choice. Front Immunol. 2017;8:291. doi: 10.3389/fimmu.2017.00291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Sharma P, Pandey AK, Bhattacharyya DK. Determining crucial genes associated with COVID-19 based on COPD findings✶,✶✶. Comput Biol Med. 2021;128:104126. doi: 10.1016/j.compbiomed.2020.104126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Zou M, Su X, Wang L, Yi X, Qiu Y, Yin X, Zhou Z, Niu X, Wang L, Su M. The molecular mechanism of multiple organ dysfunction and targeted intervention of COVID-19 based on time-order transcriptomic analysis. Front Immunol. 2021;12:729776. doi: 10.3389/fimmu.2021.729776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Jing Y, Luo L, Chen Y, Westerberg LS, Zhou P, Xu Z, Herrada AA, Park CS, Kubo M, Mei H, et al. SARS-CoV-2 infection causes immunodeficiency in recovered patients by downregulating CD19 expression in B cells via enhancing B-cell metabolism. Signal Transduct Target Ther. 2021;6:345. doi: 10.1038/s41392-021-00749-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Badbaran A, Mailer RK, Dahlke C, Woens J, Fathi A, Mellinghoff SC, Renné T, Addo MM, Riecken K, Fehse B. Digital PCR to quantify ChAdOx1 nCoV-19 copies in blood and tissues. Mol Ther Methods Clin Dev. 2021;23:418–423. doi: 10.1016/j.omtm.2021.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Grewal IS, Flavell RA. CD40 and CD154 in cell-mediated immunity. Annu Rev Immunol. 1998;16:111–135. doi: 10.1146/annurev.immunol.16.1.111. [DOI] [PubMed] [Google Scholar]
- 97.Riley-Vargas RC, Gill DB, Kemper C, Liszewski MK, Atkinson JP. CD46: Expanding beyond complement regulation. Trends Immunol. 2004;25:496–503. doi: 10.1016/j.it.2004.07.004. [DOI] [PubMed] [Google Scholar]
- 98.Lundstrom K, Barh D, Uhal BD, Takayama K, Aljabali AAA, Abd El-Aziz TM, Lal A, Redwan EM, Adadi P, Chauhan G, et al. COVID-19 vaccines and thrombosis-roadblock or dead-end street? Biomolecules. 2021;11:1020. doi: 10.3390/biom11071020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Chen J, Goyal N, Dai L, Lin Z, Del Valle L, Zabaleta J, Liu J, Post SR, Foroozesh M, Qin Z. Developing new ceramide analogs and identifying novel sphingolipid-controlled genes against a virus-associated lymphoma. Blood. 2020;136:2175–2187. doi: 10.1182/blood.2020005569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Dementyeva E, Kryukov F, Kubiczkova L, Nemec P, Sevcikova S, Ihnatova I, Jarkovsky J, Minarik J, Stefanikova Z, Kuglik P, Hajek R. Clinical implication of centrosome amplification and expression of centrosomal functional genes in multiple myeloma. J Transl Med. 2013;11:77. doi: 10.1186/1479-5876-11-77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Martinez FO, Combes TW, Orsenigo F, Gordon S. Monocyte activation in systemic Covid-19 infection: Assay and rationale. EBioMedicine. 2020;59:102964. doi: 10.1016/j.ebiom.2020.102964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Root RK, Dale DC. Granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor: Comparisons and potential for use in the treatment of infections in nonneutropenic patients. J Infect Dis. 1999;179(Suppl 2):S342–S352. doi: 10.1086/513857. [DOI] [PubMed] [Google Scholar]
- 103.Zhang N, Zhao YD, Wang XM. CXCL10 an important chemokine associated with cytokine storm in COVID-19 infected patients. Eur Rev Med Pharmacol Sci. 2020;24:7497–7505. doi: 10.26355/eurrev_202007_21922. [DOI] [PubMed] [Google Scholar]
- 104.Bergamaschi C, Terpos E, Rosati M, Angel M, Bear J, Stellas D, Karaliota S, Apostolakou F, Bagratuni T, Patseas D, et al. Systemic IL-15, IFN-γ, and IP-10/CXCL10 signature associated with effective immune response to SARS-CoV-2 in BNT162b2 mRNA vaccine recipients. Cell Rep. 2021;36:109504. doi: 10.1016/j.celrep.2021.109504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Du HX, Zhu JQ, Chen J, Zhou HF, Yang JH, Wan HT. Revealing the therapeutic targets and molecular mechanisms of emodin-treated coronavirus disease 2019 via a systematic study of network pharmacology. Aging (Albany NY) 2021;13:14571–14589. doi: 10.18632/aging.203098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Lombardero M, Kovacs K, Scheithauer BW. Erythropoietin: A hormone with multiple functions. Pathobiology. 2011;78:41–53. doi: 10.1159/000322975. [DOI] [PubMed] [Google Scholar]
- 107.Petrović J, Pešić V, Lauschke VM. Frequencies of clinically important CYP2C19 and CYP2D6 alleles are graded across Europe. Eur J Hum Genet. 2020;28:88–94. doi: 10.1038/s41431-019-0480-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Kell AM, Gale M., Jr RIG-I in RNA virus recognition. Virology. 2015;479-480:110–121. doi: 10.1016/j.virol.2015.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Boron WF, Boulpaep EL. Medical physiology: A cellular and molecular approach. 2nd edition. Saunders Elsevier; Philadelphia, PA: 2009. [Google Scholar]
- 110.Devreese KMJ. COVID-19-related laboratory coagulation findings. Int J Lab Hematol. 2021;43(Suppl 1):S36–S42. doi: 10.1111/ijlh.13547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Patel KR, Roberts JT, Barb AW. Multiple variables at the leukocyte cell surface impact Fc γ receptor-dependent mechanisms. Front Immunol. 2019;10:223. doi: 10.3389/fimmu.2019.00223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Kelton JG, Smith JW, Santos AV, Murphy WG, Horsewood P. Platelet IgG Fc receptor. Am J Hematol. 1987;25:299–310. doi: 10.1002/ajh.2830250310. [DOI] [PubMed] [Google Scholar]
- 113.Qiao J, Al-Tamimi M, Baker RI, Andrews RK, Gardiner EE. The platelet Fc receptor, FcγRIIa. Immunol Rev. 2015;268:241–252. doi: 10.1111/imr.12370. [DOI] [PubMed] [Google Scholar]
- 114.Fearon DT, Carroll MC. Regulation of B lymphocyte responses to foreign and self-antigens by the CD19/CD21 complex. Annu Rev Immunol. 2000;18:393–422. doi: 10.1146/annurev.immunol.18.1.393. [DOI] [PubMed] [Google Scholar]
- 115.Hartwig JH, Barkalow K, Azim A, Italiano J. The elegant platelet: Signals controlling actin assembly. Thromb Haemost. 1999;82:392–398. doi: 10.1055/s-0037-1615858. [DOI] [PubMed] [Google Scholar]
- 116.Viertlboeck BC, Schweinsberg S, Hanczaruk MA, Schmitt R, Du Pasquier L, Herberg FW, Göbel TW. The chicken leukocyte receptor complex encodes a primordial, activating, high-affinity IgY Fc receptor. Proc Natl Acad Sci USA. 2007;104:11718–11723. doi: 10.1073/pnas.0702011104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Tan Y, Tang F. SARS-CoV-2-mediated immune system activation and potential application in immunotherapy. Med Res Rev. 2021;41:1167–1194. doi: 10.1002/med.21756. [DOI] [PubMed] [Google Scholar]
- 118.Hotchkiss KM, Clark NM, Olivares-Navarrete R. Macrophage response to hydrophilic biomaterials regulates MSC recruitment and T-helper cell populations. Biomaterials. 2018;182:202–215. doi: 10.1016/j.biomaterials.2018.08.029. [DOI] [PubMed] [Google Scholar]
- 119.Springer S, Menzel LM, Zieger M. Google trends provides a tool to monitor population concerns and information needs during COVID-19 pandemic. Brain Behav Immun. 2020;87:109–110. doi: 10.1016/j.bbi.2020.04.073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Brockmeyer NH, Potthoff A, Kasper A, Nabring C, Jöckel KH, Siffert W. GNB3 C825T polymorphism and response to anti-retroviral combination therapy in HIV-1-infected patients-a pilot study. Eur J Med Res. 2005;10:489–494. [PubMed] [Google Scholar]
- 121.Uddin MN, Akter R, Li M, Abdelrahman Z. Expression of SARS-COV-2 cell receptor gene ACE2 is associated with immunosuppression and metabolic reprogramming in lung adenocarcinoma based on bioinformatics analyses of gene expression profiles. Chem Biol Interact. 2021;335:109370. doi: 10.1016/j.cbi.2021.109370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Bieberich F, Vazquez-Lombardi R, Yermanos A, Ehling RA, Mason DM, Wagner B, Kapetanovic E, Di Roberto RB, Weber CR, Savic M, et al. A single-cell atlas of lymphocyte adaptive immune repertoires and transcriptomes reveals age-related differences in convalescent COVID-19 patients. Front Immunol. 2021;12:701085. doi: 10.3389/fimmu.2021.701085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Fricke-Galindo I, Falfán-Valencia R. Genetics insight for COVID-19 susceptibility and severity: A review. Front Immunol. 2021;12:622176. doi: 10.3389/fimmu.2021.622176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Jiang Z, Wei F, Zhang Y, Wang T, Gao W, Yu S, Sun H, Pu J, Sun Y, Wang M, et al. IFI16 directly senses viral RNA and enhances RIG-I transcription and activation to restrict influenza virus infection. Nat Microbiol. 2021;6:932–945. doi: 10.1038/s41564-021-00907-x. [DOI] [PubMed] [Google Scholar]
- 125.Kennedy RB, Poland GA, Ovsyannikova IG, Oberg AL, Asmann YW, Grill DE, Vierkant RA, Jacobson RM. Impaired innate, humoral, and cellular immunity despite a take in smallpox vaccine recipients. Vaccine. 2016;34:3283–3290. doi: 10.1016/j.vaccine.2016.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Kotenko SV. IFN-λs. Curr Opin Immunol. 2011;23:583–590. doi: 10.1016/j.coi.2011.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Wu UI, Holland SM. Host susceptibility to non-tuberculous mycobacterial infections. Lancet Infect Dis. 2015;15:968–980. doi: 10.1016/S1473-3099(15)00089-4. [DOI] [PubMed] [Google Scholar]
- 128.Voloudakis G, Hoffman G, Venkatesh S, Lee KM, Dobrindt K, Vicari JM, Zhang W, Beckmann ND, Jiang S, Hoagland D, et al. IL10RB as a key regulator of COVID-19 host susceptibility and severity. medRxiv. doi: 10.1101/2021.05.31.21254851. [DOI] [Google Scholar]
- 129.Vecchié A, Bonaventura A, Toldo S, Dagna L, Dinarello CA, Abbate A. IL-18 and infections: Is there a role for targeted therapies. J Cell Physiol. 2021;236:1638–1657. doi: 10.1002/jcp.30008. [DOI] [PubMed] [Google Scholar]
- 130.Peters VA, Joesting JJ, Freund GG. IL-1 receptor 2 (IL-1R2) and its role in immune regulation. Brain Behav Immun. 2013;32:1–8. doi: 10.1016/j.bbi.2012.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Bénard A, Jacobsen A, Brunner M, Krautz C, Klösch B, Swierzy I, Naschberger E, Podolska MJ, Kouhestani D, David P, et al. Interleukin-3 is a predictive marker for severity and outcome during SARS-CoV-2 infections. Nat Commun. 2021;12:1112. doi: 10.1038/s41467-021-21310-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Zizzo G, Cohen PL. Imperfect storm: Is interleukin-33 the Achilles heel of COVID-19? Lancet Rheumatol. 2020;12:e779–e790. doi: 10.1016/S2665-9913(20)30340-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Walsh PT, Fallon PG. The emergence of the IL-36 cytokine family as novel targets for inflammatory diseases. Ann NY Acad Sci. 2018;1417:23–34. doi: 10.1111/nyas.13280. [DOI] [PubMed] [Google Scholar]
- 134.Nussbaum JC, Van Dyken SJ, von Moltke J, Cheng LE, Mohapatra A, Molofsky AB, Thornton EE, Krummel MF, Chawla A, Liang HE, Locksley RM. Type 2 innate lymphoid cells control eosinophil homeostasis. Nature. 2013;502:245–248. doi: 10.1038/nature12526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Coomes EA, Haghbayan H. Interleukin-6 in Covid-19: A systematic review and meta-analysis. Rev Med Virol. 2020;30:1–9. doi: 10.1002/rmv.2141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Das UN. Bioactive lipids in COVID-19-further evidence. Arch Med Res. 2021;52:107–120. doi: 10.1016/j.arcmed.2020.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Islam ABMMK, Khan MA, Ahmed R, Hossain MS, Kabir SMT, Islam MS, Siddiki AMAMZ. Transcriptome of nasopharyngeal samples from COVID-19 patients and a comparative analysis with other SARS-CoV-2 infection models reveal disparate host responses against SARS-CoV-2. J Transl Med. 2021;19:32. doi: 10.1186/s12967-020-02695-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.O'Brien JR. Shear-induced platelet aggregation. Lancet. 1990;335:711–713. doi: 10.1016/0140-6736(90)90815-M. [DOI] [PubMed] [Google Scholar]
- 139.Langmuir P, Yeleswaram S, Smith P, Knorr B, Squier P. Design of clinical trials evaluating ruxolitinib, a JAK1/JAK2 inhibitor, for treatment of COVID-19-associated cytokine storm. Dela J Public Health. 2020;6:50–54. doi: 10.32481/djph.2020.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Melman YF, Krummerman A, McDonald TV. KCNE regulation of KvLQT1 channels: Structure-function correlates. Trends Cardiovasc Med. 2002;12:182–187. doi: 10.1016/S1050-1738(02)00158-5. [DOI] [PubMed] [Google Scholar]
- 141.Gouas L, Nicaud V, Berthet M, Forhan A, Tiret L, Balkau B, Guicheney P, D.E.S.I.R. Study Group Association of KCNQ1, KCNE1, KCNH2 and SCN5A polymorphisms with QTc interval length in a healthy population. Eur J Hum Genet. 2005;13:1213–1222. doi: 10.1038/sj.ejhg.5201489. [DOI] [PubMed] [Google Scholar]
- 142.Lazzerini PE, Acampa M, Laghi-Pasini F, Bertolozzi I, Finizola F, Vanni F, Natale M, Bisogno S, Cevenini G, Cartocci A, et al. Cardiac arrest risk during acute infections: Systemic inflammation directly prolongs QTc interval via cytokine-mediated effects on potassium channel expression. Circ Arrhythm Electrophysiol. 2020;13:e008627. doi: 10.1161/CIRCEP.120.008627. [DOI] [PubMed] [Google Scholar]
- 143.Szendrey M, Guo J, Li W, Yang T, Zhang S. COVID-19 drugs chloroquine and hydroxychloroquine, but not azithromycin and remdesivir, block hERG potassium channels. J Pharmacol Exp Ther. 2021;377:265–272. doi: 10.1124/jpet.120.000484. [DOI] [PubMed] [Google Scholar]
- 144.Dahl SL, Woodworth JS, Lerche CJ, Cramer EP, Nielsen PR, Moser C, Thomsen AR, Borregaard N, Cowland JB. Lipocalin-2 functions as inhibitor of innate resistance to mycobacterium tuberculosis. Front Immunol. 2018;9:2717. doi: 10.3389/fimmu.2018.02717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Vincenti MP, Brinckerhoff CE. Transcriptional regulation of collagenase (MMP-1, MMP-13) genes in arthritis: Integration of complex signaling pathways for the recruitment of gene-specific transcription factors. Arthritis Res. 2002;4:157–164. doi: 10.1186/ar401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Jabłońska-Trypuć A, Matejczyk M, Rosochacki S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J Enzyme Inhib Med Chem. 2016;31(Suppl 1):S177–S183. doi: 10.3109/14756366.2016.1161620. [DOI] [PubMed] [Google Scholar]
- 147.Pisano TJ, Hakkinen I, Rybinnik I. Large vessel occlusion secondary to COVID-19 hypercoagulability in a young patient: A case report and literature review. J Stroke Cerebrovasc Dis. 2020;29:105307. doi: 10.1016/j.jstrokecerebrovasdis.2020.105307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Apostolidis SA, Kakara M, Painter MM, Goel RR, Mathew D, Lenzi K, Rezk A, Patterson KR, Espinoza DA, Kadri JC, et al. Cellular and humoral immune responses following SARS-CoV-2 mRNA vaccination in patients with multiple sclerosis on anti-CD20 therapy. Nat Med. 2021;27:1990–2001. doi: 10.1038/s41591-021-01507-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Lu W, Liu X, Wang T, Liu F, Zhu A, Lin Y, Luo J, Ye F, He J, Zhao J, et al. Elevated MUC1 and MUC5AC mucin protein levels in airway mucus of critical ill COVID-19 patients. J Med Virol. 2021;93:582–584. doi: 10.1002/jmv.26406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Conti P, Ronconi G, Caraffa A, Gallenga CE, Ross R, Frydas I, Kritas SK. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by Coronavirus-19 (COVI-19 or SARS-CoV-2): Anti-inflammatory strategies. J Biol Regul Homeost Agents. 2020;34:327–331. doi: 10.23812/CONTI-E. [DOI] [PubMed] [Google Scholar]
- 151.Morsy S. NCAM protein and SARS-COV-2 surface proteins: In-silico hypothetical evidence for the immunopathogenesis of Guillain-Barré syndrome. Med Hypotheses. 2020;145:110342. doi: 10.1016/j.mehy.2020.110342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Root-Bernstein R. Innate receptor activation patterns involving TLR and NLR synergisms in COVID-19, ALI/ARDS and sepsis cytokine storms: A review and model making novel predictions and therapeutic suggestions. Int J Mol Sci. 2021;22:2108. doi: 10.3390/ijms22042108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Watanabe T, Kitani A, Murray PJ, Strober W. NOD2 is a negative regulator of Toll-like receptor 2-mediated T helper type 1 responses. Nat Immunol. 2004;5:800–808. doi: 10.1038/ni1092. [DOI] [PubMed] [Google Scholar]
- 154.Esposito E, Cuzzocrea S. The role of nitric oxide synthases in lung inflammation. Curr Opin Investig Drugs. 2007;8:899–909. [PubMed] [Google Scholar]
- 155.Gamkrelidze M, Intskirveli N, Vardosanidze K, Goliadze L, Chikhladze KH, Ratiani L. Myocardial dysfunction during septic shock (review) Georgian Med News. 2014;237:40–46. [PubMed] [Google Scholar]
- 156.Zang X, Li S, Zhao Y, Chen K, Wang X, Song W, Ma J, Tu X, Xia Y, Zhang S, Gao C. Systematic meta-analysis of the association between a common NOS1AP genetic polymorphism, the QTc interval, and sudden death. Int Heart J. 2019;60:1083–1090. doi: 10.1536/ihj.19-024. [DOI] [PubMed] [Google Scholar]
- 157.Guan SP, Seet RCS, Kennedy BK. Does eNOS derived nitric oxide protect the young from severe COVID-19 complications. Ageing Res Rev. 2020;64:101201. doi: 10.1016/j.arr.2020.101201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Thom SR, Fisher D, Xu YA, Garner S, Ischiropoulos H. Role of nitric oxide-derived oxidants in vascular injury from carbon monoxide in the rat. Am J Physiol. 1999;276:H984–H992. doi: 10.1152/ajpheart.1999.276.3.H984. [DOI] [PubMed] [Google Scholar]
- 159.Valent A, Danglot G, Bernheim A. Mapping of the tyrosine kinase receptors trkA (NTRK1), trkB (NTRK2) and trkC(NTRK3) to human chromosomes 1q22, 9q22 and 15q25 by fluorescence in situ hybridization. Eur J Hum Genet. 1997;5:102–104. doi: 10.1159/000484742. [DOI] [PubMed] [Google Scholar]
- 160.Liu W, Chen L, Zhu J, Rodgers GP. The glycoprotein hGC-1 binds to cadherin and lectins. Exp Cell Res. 2006;312:1785–1797. doi: 10.1016/j.yexcr.2006.02.011. [DOI] [PubMed] [Google Scholar]
- 161.Hennigs JK, Lüneburg N, Stage A, Schmitz M, Körbelin J, Harbaum L, Matuszcak C, Mienert J, Bokemeyer C, Böger RH, et al. The P2-receptor-mediated Ca2+ signalosome of the human pulmonary endothelium-implications for pulmonary arterial hypertension. Purinergic Signal. 2019;15:299–311. doi: 10.1007/s11302-019-09674-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Russo MV, McGavern DB. Immune surveillance of the CNS following infection and injury. Trends Immunol. 2015;36:637–650. doi: 10.1016/j.it.2015.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Tuuminen R, Nykänen A, Keränen MA, Krebs R, Alitalo K, Koskinen PK, Lemström KB. The effect of platelet-derived growth factor ligands in rat cardiac allograft vasculopathy and fibrosis. Transplant Proc. 2006;38:3271–3273. doi: 10.1016/j.transproceed.2006.10.056. [DOI] [PubMed] [Google Scholar]
- 164.Blum E, Margalit R, Levy L, Getter T, Lahav R, Zilber S, Bradfield P, Imhof BA, Alpert E, Gruzman A. A Potent leukocyte transmigration blocker: GT-73 showed a protective effect against LPS-induced ARDS in mice. Molecules. 2021;26:4583. doi: 10.3390/molecules26154583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Rovina N, Akinosoglou K, Eugen-Olsen J, Hayek S, Reiser J, Giamarellos-Bourboulis EJ. Soluble urokinase plasminogen activator receptor (suPAR) as an early predictor of severe respiratory failure in patients with COVID-19 pneumonia. Crit Care. 2020;24:187. doi: 10.1186/s13054-020-02897-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Kumar S, Jain A, Choi SW, da Silva GPD, Allers L, Mudd MH, Peters RS, Anonsen JH, Rusten TE, Lazarou M, Deretic V. Mammalian Atg8 proteins and the autophagy factor IRGM control mTOR and TFEB at a regulatory node critical for responses to pathogens. Nat Cell Biol. 2020;22:973–985. doi: 10.1038/s41556-020-0549-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Hoxha M. What about COVID-19 and arachidonic acid pathway. Eur J Clin Pharmacol. 2020;76:1501–1504. doi: 10.1007/s00228-020-02941-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Keikha M, Ghazvini K, Eslami M, Yousefi B, Casseb J, Yousefi M, Karbalaei M. Molecular targeting of PD-1 signaling pathway as a novel therapeutic approach in HTLV-1 infection. Microb Pathog. 2020;144:104198. doi: 10.1016/j.micpath.2020.104198. [DOI] [PubMed] [Google Scholar]
- 169.Coggeshall KM. Negative signaling in health and disease. Immunol Res. 1999;19:47–64. doi: 10.1007/BF02786476. [DOI] [PubMed] [Google Scholar]
- 170.de Souza JG, Starobinas N, Ibañez O. Unknown/enigmatic functions of extracellular ASC. Immunology. 2021;163:377–388. doi: 10.1111/imm.13375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Larabi A, Barnich N, Nguyen HTT. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in IBD. Autophagy. 2020;16:38–51. doi: 10.1080/15548627.2019.1635384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Brown MJ. Renin: Friend or foe? Heart. 2007;93:1026–1033. doi: 10.1136/hrt.2006.107706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Amraei R, Rahimi N. COVID-19, renin-angiotensin system and endothelial dysfunction. Cells. 2020;9:1652. doi: 10.3390/cells9071652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Yanatori I, Yasui Y, Noguchi Y, Kishi F. Inhibition of iron uptake by ferristatin II is exerted through internalization of DMT1 at the plasma membrane. Cell Biol Int. 2015;39:427–434. doi: 10.1002/cbin.10403. [DOI] [PubMed] [Google Scholar]
- 175.Denham NC, Pearman CM, Ding WY, Waktare J, Gupta D, Snowdon R, Hall M, Cooper R, Modi S, Todd D, Mahida S. Systematic re-evaluation of SCN5A variants associated with Brugada syndrome. J Cardiovasc Electrophysiol. 2019;30:118–127. doi: 10.1111/jce.13740. [DOI] [PubMed] [Google Scholar]
- 176.Smadja DM, Guerin CL, Chocron R, Yatim N, Boussier J, Gendron N, Khider L, Hadjadj J, Goudot G, Debuc B, et al. Angiopoietin-2 as a marker of endothelial activation is a good predictor factor for intensive care unit admission of COVID-19 patients. Angiogenesis. 2020;23:611–620. doi: 10.1007/s10456-020-09730-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Bongiovanni D, Klug M, Lazareva O, Weidlich S, Biasi M, Ursu S, Warth S, Buske C, Lukas M, Spinner CD, et al. SARS-CoV-2 infection is associated with a pro-thrombotic platelet phenotype. Cell Death Dis. 2021;12:50. doi: 10.1038/s41419-020-03333-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Wu D, Yang XO. Dysregulation of pulmonary responses in severe COVID-19. Viruses. 2021;13:957. doi: 10.3390/v13060957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Katzen J, Beers MF. Contributions of alveolar epithelial cell quality control to pulmonary fibrosis. J Clin Invest. 2020;130:5088–5099. doi: 10.1172/JCI139519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Nandy D, Sharma N, Senapati S. Systematic review and meta-analysis confirms significant contribution of surfactant protein D in chronic obstructive pulmonary disease. Front Genet. 2019;10:339. doi: 10.3389/fgene.2019.00339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Di Lisa F, Kaludercic N, Carpi A, Menabò R, Giorgio M. Mitochondria and vascular pathology. Pharmacol Rep. 2009;61:123–130. doi: 10.1016/S1734-1140(09)70014-3. [DOI] [PubMed] [Google Scholar]
- 182.Dinarello CA, Nold-Petry C, Nold M, Fujita M, Li S, Kim S, Bufler P. Suppression of innate inflammation and immunity by interleukin-37. Eur J Immunol. 2016;46:1067–1081. doi: 10.1002/eji.201545828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Montalbetti N, Simonin A, Kovacs G, Hediger MA. Mammalian iron transporters: families SLC11 and SLC40. Mol Aspects Med. 2013;34:270–287. doi: 10.1016/j.mam.2013.01.002. [DOI] [PubMed] [Google Scholar]
- 184.Schulert GS, Blum SA, Cron RQ. Host genetics of pediatric SARS-CoV-2 COVID-19 and multisystem inflammatory syndrome in children. Curr Opin Pediatr. 2021;33:549–555. doi: 10.1097/MOP.0000000000001061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Pachlopnik Schmid J, de Saint Basile G. Angeborene hämophagozytische lymphohistiozytose (HLH) Klin Padiatr. 2010;222:345–350. doi: 10.1055/s-0029-1246165. [DOI] [PubMed] [Google Scholar]
- 186.Shi JH, Xie X, Sun SC. TBK1 as a regulator of autoimmunity and antitumor immunity. Cell Mol Immunol. 2018;15:743–745. doi: 10.1038/cmi.2017.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Zimecki M, Actor JK, Kruzel ML. The potential for Lactoferrin to reduce SARS-CoV-2 induced cytokine storm. Int Immunopharmacol. 2021;95:107571. doi: 10.1016/j.intimp.2021.107571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Chabot PR, Raiola L, Lussier-Price M, Morse T, Arseneault G, Archambault J, Omichinski JG. Structural and functional characterization of a complex between the acidic transactivation domain of EBNA2 and the Tfb1/p62 subunit of TFIIH. PLoS Pathog. 2014;10:e1004042. doi: 10.1371/journal.ppat.1004042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Speeckaert MM, Speeckaert R, Delanghe JR. Biological and clinical aspects of soluble transferrin receptor. Crit Rev Clin Lab Sci. 2010;47:213–228. doi: 10.3109/10408363.2010.550461. [DOI] [PubMed] [Google Scholar]
- 190.Bg S, Gosavi S, Ananda Rao A, Shastry S, Raj SC, Sharma A, Suresh A, Noubade R. Neutrophil-to-lymphocyte, lymphocyte-to-monocyte, and platelet-to-lymphocyte ratios: Prognostic significance in COVID-19. Cureus. 2021;13:e12622. doi: 10.7759/cureus.12622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Campbell GR, To RK, Hanna J, Spector SA. SARS-CoV-2, SARS-CoV-1, and HIV-1 derived ssRNA sequences activate the NLRP3 inflammasome in human macrophages through a non-classical pathway. iScience. 2021;24:102295. doi: 10.1016/j.isci.2021.102295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Borrello S, Nicolò C, Delogu G, Pandolfi F, Ria F. TLR2: a crossroads between infections and autoimmunity. Int J Immunopathol Pharmacol. 2011;24:549–556. doi: 10.1177/039463201102400301. [DOI] [PubMed] [Google Scholar]
- 193.Zheng M, Karki R, Williams EP, Yang D, Fitzpatrick E, Vogel P, Jonsson CB, Kanneganti TD. TLR2 senses the SARS-CoV-2 envelope protein to produce inflammatory cytokines. Nat Immunol. 2021;22:829–838. doi: 10.1038/s41590-021-00937-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Khan S, Shafiei M, Longoria C, Schoggins JW, Savani R, Zaki H. SARS-CoV-2 spike protein induces inflammation via TLR2-dependent activation of the NF-κB pathway. Elife. 2021;10:e68563. doi: 10.7554/eLife.68563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Sohn KM, Lee SG, Kim HJ, Cheon S, Jeong H, Lee J, Kim IS, Silwal P, Kim YJ, Paik S, et al. COVID-19 patients upregulate toll-like receptor 4-mediated inflammatory signaling that mimics bacterial sepsis. J Korean Med Sci. 2020;35:e343. doi: 10.3346/jkms.2020.35.e343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Guven-Maiorov E, Keskin O, Gursoy A, VanWaes C, Chen Z, Tsai CJ, Nussinov R. TRAF3 signaling: Competitive binding and evolvability of adaptive viral molecular mimicry. Biochim Biophys Acta. 2016;1860:2646–2655. doi: 10.1016/j.bbagen.2016.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Callaway E. The quest to find genes that drive severe COVID. Nature. 2021;595:346–348. doi: 10.1038/d41586-021-01827-w. [DOI] [PubMed] [Google Scholar]
- 198.Kaur S, Tripathi DM, Yadav A. The enigma of endothelium in COVID-19. Front Physiol. 2020;11:989. doi: 10.3389/fphys.2020.00989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Rovas A, Osiaevi I, Buscher K, Sackarnd J, Tepasse PR, Fobker M, Kühn J, Braune S, Göbel U, Thölking G, et al. Microvascular dysfunction in COVID-19: The MYSTIC study. Angiogenesis. 2021;24:145–157. doi: 10.1007/s10456-020-09753-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Holcomb D, Alexaki A, Hernandez N, Hunt R, Laurie K, Kames J, Hamasaki-Katagiri N, Komar AA, DiCuccio M, Kimchi-Sarfaty C. Gene variants of coagulation related proteins that interact with SARS-CoV-2. PLoS Comput Biol. 2021;17:e1008805. doi: 10.1371/journal.pcbi.1008805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Shahidi M. Thrombosis and von Willebrand factor. Adv Exp Med Biol. 2017;906:285–306. doi: 10.1007/5584_2016_122. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available throughout the manuscript.