

Abstract
Bitter taste perception plays vital roles in animal behavior and fitness. By signaling the presence of toxins in foods, particularly noxious defense compounds found in plants, it enables animals to avoid exposure. In vertebrates, bitter perception is initiated by TAS2Rs, a family of G protein-coupled receptors expressed on the surface of taste buds. There, oriented toward the interior of the mouth, they monitor the contents of foods, drinks and other substances as they are ingested. When bitter compounds are encountered, TAS2Rs respond by triggering neural pathways leading to sensation. The importance of this role placed TAS2Rs under selective pressures in the course of their evolution, leaving signatures in patterns of gene gain and loss, sequence polymorphism, and population structure consistent with vertebrates' diverse feeding ecologies. The protective value of bitter taste is reduced in modern humans because contemporary food supplies are safe and abundant. However, this is not always the case. Some crops, particularly in the developing world, retain surprisingly high toxicity and bitterness remains an important measure of safety. Bitter perception also shapes health through its influence on preference driven behaviors such as diet choice, alcohol intake and tobacco use. Further, allelic variation in TAS2Rs is extensive, leading to individual differences in taste sensitivity that drive these behaviors, shaping susceptibility to disease. Thus, bitter taste perception occupies a critical intersection between ancient evolutionary processes and modern human health.
Keywords: bitter, molecular evolution, senses, taste, genetics, diet
Lay Summary
Bitter tastes warn animals about noxious substances in the environment, especially toxins found in plants. This has placed the genes controlling bitter taste under eons of pressure from natural selection. Signatures of these pressures remain in taste genes today, shaping food preferences, consumption, and health in animals and humans alike.
INTRODUCTION
The diets of humans, their hominid ancestors and their primate relatives are dominated by plants, and for good reason [1]. Plants are abundant, productive and immobile, making them a potentially bountiful and accessible source of nutrition. However, plants are not as vulnerable as they seem. In a classic evolutionary arms race, they have adapted to herbivores, equipping themselves with an array of physical, reproductive and chemical defenses [2]. Substantial herbivore behavior and energy are dedicated to circumventing obstacles such as spines and thorns, shells and husks, synchronized fruiting and symbioses with animal defenders such as ants. Humans’ cognitive sophistication and adaptability make them uniquely successful at evading and even exploiting these protections, a talent catalyzing their dispersal from tropical Africa into virtually every ecosystem worldwide in the span of less than 75 000 years.
Perhaps the most formidable line of anti-herbivore defense in plants is their use of toxins. Nearly all plant species produce noxious compounds aimed at deterring predators [3, 4]. Some are notorious; alkaloids from poison hemlock were famously used to execute Socrates, and strychnine was used by an unknown assassin to kill Jane Stanford, co-founder of Stanford University (Fig. 1) [5]. However, the full repertoire of defense compounds used by plants reaches into the hundreds of thousands, with most receiving little notice because they are either not deadly or are rarely encountered [3]. For instance, urushiol, found in poison ivy, is irritating but not dangerous and abrin, found in rosary pea, is lethal at minute concentrations but encapsulated in easily recognized, undigestible seeds [6, 7]. Other notable examples among many include cyclopamine, an alkaloid in corn lily that interferes with embryogenesis, oleandrin, a cardiac glycoside in oleander that disrupts heartbeat, and cicutoxin, a polyacetylene in water hemlock that causes respiratory paralysis.
Figure 1.

The Death of Socrates, by Jacques-Louis David (1787). Several lines of evidence suggest that Socrates was executed using extracts of hemlock (Conium maculatum) containing coniine and related alkaloids. Descriptions of Socrates's symptoms as he died to raise the possibility that additional compounds were included in the mixture but their identities remain unknown [5]
Herbivores have evolved a range of strategies for circumventing plants’ toxic defenses such as learned avoidance, metabolic denaturation and even symbiosis with microbiota [8, 9]. Among these, a particularly effective and widespread mechanism in vertebrates is bitter taste perception [10]. It has long been recognized that numerous plant toxins are perceived as bitter, and this provides a powerful means of reducing exposure. By signaling the presence of toxins in foods before they are fully ingested, bitter perception allows avoidance. Agriculture renders this role mostly redundant today. However, some crops retain toxicity despite domestication, and bitterness can be an important indicator of safety. Such is the case with cassava, a tropical crop containing neurotoxins that damage sensory, motor and cognitive function [11]. Other crops contain compounds that are bitter yet non-hazardous. This is the case with cruciferous vegetables, which contain the thyroid inhibitor goitrin [12, 13]. Goitrin is harmless in the quantities present in domesticated crucifers, yet its bitterness discourages consumption [14–17].
The ubiquity of bitter perception across vertebrates and its complex role in humans have raised questions about its evolution since the 1930s. Early studies comparing humans and chimpanzees concluded that natural selection has maintained variation in the two species, but inferences were limited by a lack of information about the specific genes involved [18, 19]. However, advancements have revealed the transduction pathways underlying bitter taste as well as the genes encoding them [20–27]. These are making possible the first high-resolution studies of molecular evolution in taste mechanisms, shedding light on the origins of bitter perception, variation in taste abilities within and among species, and their relationships with animal ecology and behavior [28, 29]. They are also providing new tools for dissecting the underpinnings of taste phenotypes, consumption behaviors and their nutritional consequences in humans. These lines of research continue to diversify and expand, painting an integrated portrait of connections between ancient evolutionary processes and modern human health.
MECHANISMS OF BITTER PERCEPTION
Bitter taste perception begins, of course in the mouth, whose surface is composed of epithelial tissue [30]. Embedded within the tissue on the tongue, soft palate and pharynx lie taste buds, clusters of receptor cells exposed to the mouth's interior through pores (Fig. 2). Each of the major taste qualities in humans (bitter, sweet, umami, sour and salty) is mediated by cells expressing corresponding receptor proteins, with salty and sour cells expressing ion channels and bitter, sweet and umami cells expressing G protein-coupled receptors (GPCRs) [31]. The receptors specifically responsible for initiating bitter perception were identified roughly 20 years ago and designated T2Rs, TAS2Rs, Tas2rs, or Tas2Rs, according to species (for convenience we refer to all using the human term, TAS2R, here) [24, 25, 32, 33]. This revealed their relationships with other GPCRs involved in environmental sensing including olfactory receptors (ORs) and opsins (RHO and OPNs), which operate through similar molecular mechanisms [34].
Figure 2.

Tongue anatomy. Taste receptor cells are clustered in bundles beneath the surface of papillae, where they are exposed to the interior of the mouth through pores. TAS2Rs in the apical portion of the cells are poised to detect compounds in foods, smoke, pharmaceuticals and other ingested substances (©Casey Henley, CC BY-NC-SA 4.0 International License)
TAS2Rs localizes to the surface of bitter receptor cells, where they are exposed to chemicals in foods and other substances passing through the oral cavity. When stimulated by compatible compounds, TAS2Rs trigger a transduction cascade composed of two parallel pathways (Fig. 3) [35]. The first operates through the activation of phospholipase C β2 (PLCB2) by the β and γ subunits of TAS2Rs' cognate G protein. The second, which plays a modulatory role, operates through the activation of phosphodiesterase 1A (PDE1A) by the G protein's α subunit, α-gustducin. Downstream, ATP release is facilitated by activation of a calcium homeostasis modulator channel, CALHM1/3 [36–38]. Together these pathways depolarize the receptor cell, producing a signal conveyed to the central nervous system.
Figure 3.
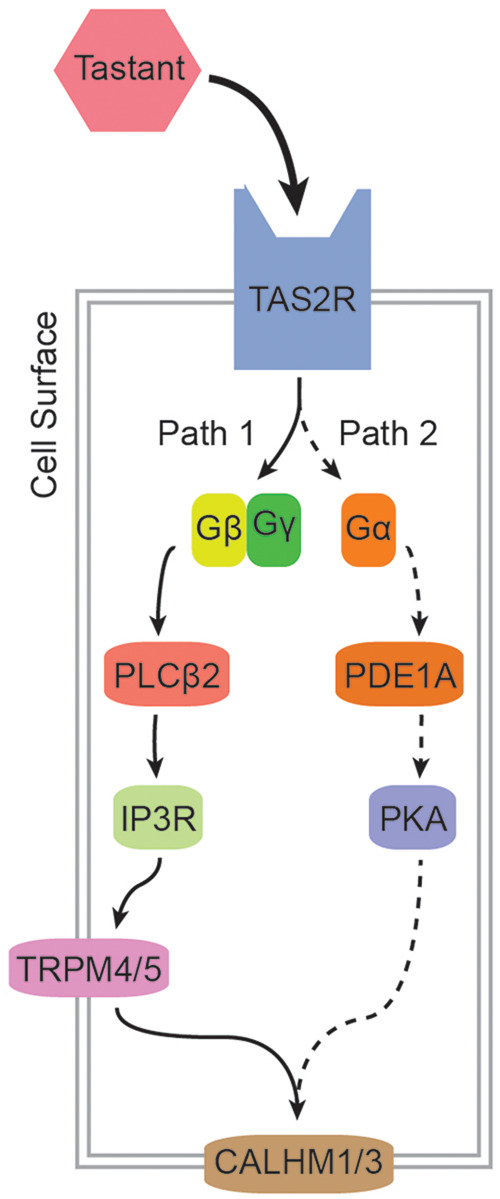
Bitter taste transduction cascade. TAS2Rs initiate the transduction process when exposed to compatible compounds. These interactions determine which substances are perceived as bitter and which are not, placing TAS2Rs under selective pressures that vary according to diet
The discovery that TAS2Rs are bitter receptors was nearly simultaneous with the completion of the human genome project, which uncovered the complete set of genes encoding bitter transduction mechanisms. Remarkably, they included not one but 33 TAS2Rs, with 25 functional loci and 8 pseudogenes occurring in clusters on chromosomes 5, 7 and 12 [24, 25, 39]. This makes the TAS2R family intermediate in size among sensory GPCR families such as opsins, of which there are four (OPNs and RHO), and olfactory receptors (ORs), of which there are hundreds [34]. TAS2Rs were also found to be simple structurally, with each composed of a single exon ∼1kb in length (Table 1). The remaining components of the transduction cascade are represented by one gene each with the exception of phosphokinase A (PKA), a tetramer encoded by seven. The genes encoding transduction components are also more complex than TAS2Rs, averaging 15 exons. Later findings in non-humans revealed that most vertebrates utilize the same system, with TAS2Rs initiating transductional cascades mediated by PLC β2.
Table 1.
Human TAS2R genes, coordinates, transcript lengths and product sizes in the GRCh37 human genome map
| Gene | GRCh 37 Coordinates | Transcript length | # Codons |
|---|---|---|---|
| TAS2R1 | 5: 9629109–9630463 | 1355 | 300 |
| TAS2R3 | 7: 141463897–141464997 | 1101 | 317 |
| TAS2R4 | 7: 141478242–141479235 | 994 | 300 |
| TAS2R5 | 7: 141490017–141491166 | 1150 | 300 |
| TAS2R7 | 12: 10954131–10955226 | 1096 | 319 |
| TAS2R8 | 12: 10958650–10959892 | 1243 | 310 |
| TAS2R9 | 12: 10961693–10962767 | 1075 | 313 |
| TAS2R10 | 12: 10977916–10978957 | 1042 | 308 |
| TAS2R13 | 12: 11060525–11062161 | 1637 | 304 |
| TAS2R14 | 12: 11090005–11091862 | 1858 | 318 |
| TAS2R16 | 7: 122634759–122635754 | 996 | 292 |
| TAS2R19 | 12: 11174218–11175219 | 1002 | 300 |
| TAS2R20 | 12: 11149094–11150474 | 1381 | 310 |
| TAS2R30 | 12: 11285557–11287243 | 1687 | 320 |
| TAS2R31 | 12: 11182986–11184006 | 1021 | 310 |
| TAS2R38 | 7: 141672431–141673573 | 1143 | 334 |
| TAS2R39 | 7: 142880512–142881528 | 1017 | 339 |
| TAS2R40 | 7: 142919130–142920162 | 1033 | 324 |
| TAS2R41 | 7: 143174966–143175889 | 924 | 308 |
| TAS2R42 | 12: 11338599–11339543 | 945 | 315 |
| TAS2R43 | na | na | na |
| TAS2R45 | na | na | na |
| TAS2R46 | 12: 11213964–11214893 | 930 | 310 |
| TAS2R50 | 12: 11138512–11139511 | 1000 | 300 |
| TAS2R60 | 7: 143140546–143141502 | 957 | 319 |
| TAS2R2P | 7: 12530721–12531630 | 910 | ∼303 |
| TAS2R12P | 12: 11047542–11048481 | 940 | ∼313 |
| TAS2R15P | 12: 11117024–11117951 | 928 | ∼309 |
| TAS2R18P | 12: 11311375–11312293 | 919 | ∼303 |
| TAS2R62P | 7: 143134127–143135066 | 940 | ∼313 |
| TAS2R63P | 12: 11200931–11201855 | 925 | ∼308 |
| TAS2R64P | 12: 11229915–11230841 | 927 | ∼309 |
| TAS2R67P | 12: 11332272–11333061 | 790 | ∼263 |
Coordinates of TAS2R43 and TAS2R45 are denoted na because their coordinates are ambiguous in GRCh37.
Like the hierarchical organization of the bitter transduction cascade, the expression of genes encoding the bitter transduction cascade highlights the specialization of TAS2Rs relative to other transduction components. Among the genes encoding the pathway, the most circumscribed expression pattern is that of TAS2Rs. They are highly expressed in bitter receptor cells in taste buds as well as in chemosensory cells in gut and bronchial smooth muscle, where they detect ingested and inhaled agonists [33, 40–42]. A critical feature of TAS2Rs in taste bud cells is that they are coexpressed, with the average cell in humans expressing a random set of ∼5 to 10 of the 25 possible [40]. As a result, responsiveness to any particular compound can vary from cell to cell. Like TAS2Rs, α-gustducin is required for fully functional bitter perception. However, unlike TAS2Rs, it is also highly expressed in receptor cells responsive to sweet and savory compounds where it interacts with TAS1R receptors [43, 44]. Genes encoding other components of the pathway are expressed more broadly still, with some being highly generic to GPCR signaling processes and ubiquitously present across cell and tissue types [45–56].
While TAS2Rs are best known for their importance to perception, they play additional roles as well. They appear to be particularly important chemical sensors in the gut and airways, where they mediate responses to nutrients and inhaled substances [57–60]. For instance, stimulation of gut-expressed TAS2Rs triggers endocrine responses as well as processes such as gastric emptying [42]. Evidence that TAS2Rs can eavesdrop on bacterial quorum sensing activity in the gut suggests that they may also mediate responses to microbial conditions, which are both affected by and driven by health factors [59, 61]. Recent findings point to another function of TAS2Rs in the gut, the detection of parasites secreting-excreting TAS2R agonists [62]. Similar patterns are found in the airways, where TAS2Rs trigger responses to compounds originating from both inside and outside the body and likely mediate immune responses [59, 60].
TAS2RS AND THEIR AGONISTS
The initial discovery that TAS2Rs are bitter taste receptors was based on their responses to a small number of well-defined compounds such as cycloheximide, denatonium and 6-n-propylthiouracil [32]. However, dozens of TAS2R agonists are now known, including numerous plant toxins as well as bitter yet non-toxic substances. In the most comprehensive study performed to date, Meyerhof et al. [63] used in vitro assays to determine the responses of all 25 human TAS2Rs to a library of 58 structurally diverse compounds, with an emphasis on toxins found in plants (Fig. 4). The results revealed that of the 25 tested TAS2Rs, 15 were activated by at least one of the 58 tested compounds. Conversely, of the 58 compounds, 43 were activated at least one TAS2R. In addition, across compounds tested, the number of responsive TAS2Rs ranged from 0 (no receptors for curcumin, digitonin, α-solanine and others were found) to 9 (quinine), and the number of compounds to which a given TAS2R was responsive ranged from 0 (TAS2R3, TAS2R5 and TAS2R9 and others responded to none of the 58) to 19 (TAS2R46). Moreover, when different TAS2Rs were responsive to the same compound they frequently exhibited different levels of sensitivity. These findings indicate that perception of most bitter compounds is not a simple function of compatibility between an agonist and a single receptor, it is a complex function of the full set of TAS2Rs to which the compound is agonistic, their individual patterns of response, and their variable expression across cells.
Figure 4.
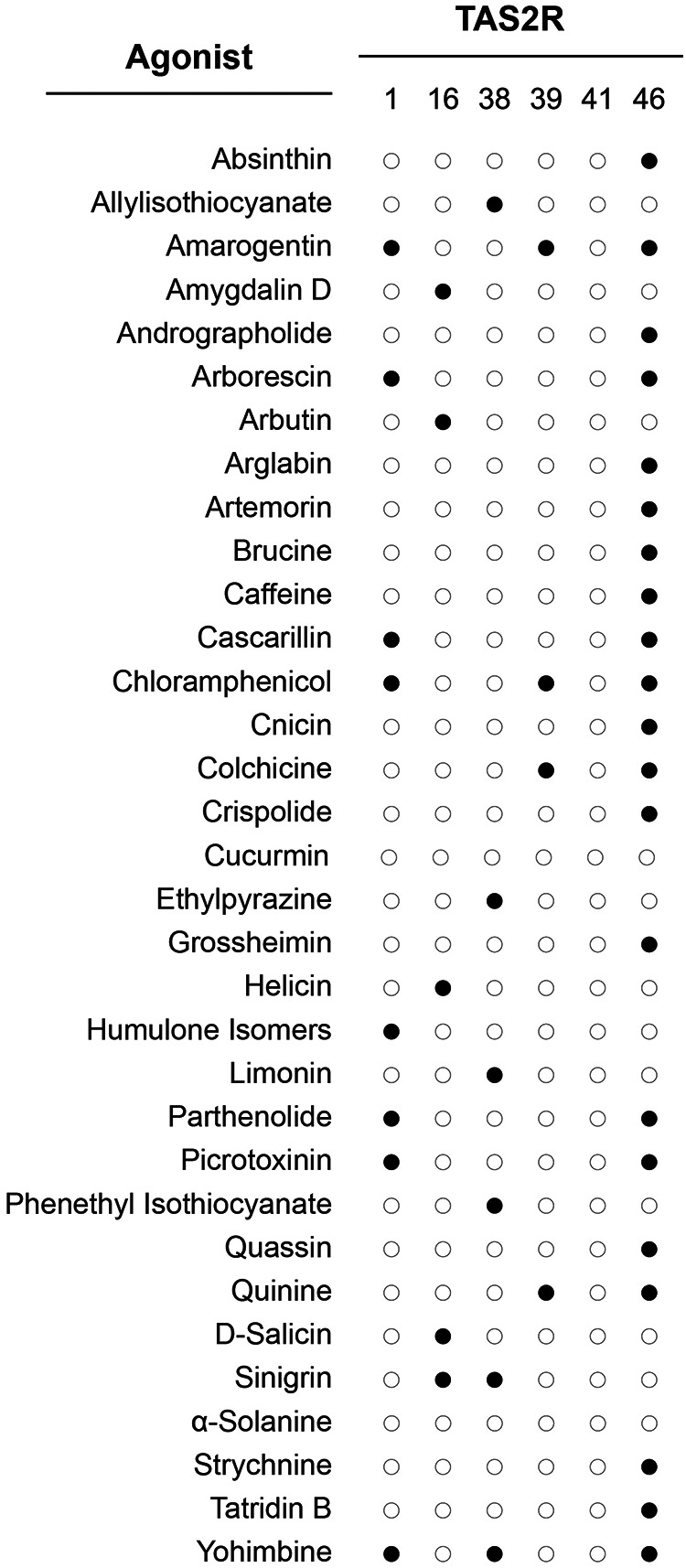
Example interactions between TAS2Rs and plant secondary compounds described by Meyerhof et al. [63]. Filled circles indicate activation, empty circles indicate no activation. Two key patterns are that most TAS2Rs are responsive to multiple compounds, and many compounds activate multiple TAS2Rs. These patterns extend across the TAS2R family in humans
Surveys of receptor–agonist interactions in non-humans have revealed patterns similar to those in humans, with most TAS2Rs responding to multiple compounds and most compounds eliciting responses from multiple TAS2Rs [64]. Remarkably, many compounds agonistic to human TAS2Rs are also agonists of non-human TAS2Rs, and some compounds elicit responses from TAS2Rs in every species tested to date. This is surprising given the level of variability in diet among taxa, which would seem to predict that species carry receptors specifically targeting compounds encountered when foraging. A potential explanation for the overlap is that plants do not produce an unlimited diversity of secondary compounds. Most belong to families, such as alkaloids, terpenes and phenolics [3]. For instance, numerous plants produce glycosides, with the specific molecules varying across species but all sharing the chemical characteristics that make them glycosides. Hence, if a TAS2R is responsive to one compound it is likely responsive to others. Likewise, if a compound elicits responses from one TAS2R it is likely to elicit responses from TAS2Rs responsive to compounds with similar chemical structures. This phenomenon is documented for TAS2R16, which is broadly responsive to β-D-glucopyranosides, and TAS2R38, which is broadly responsive to isothiocyanates [17, 65–69]. Investigations using computational methods have yielded additional insights, suggesting that compounds with small globular structures tend to activate more TAS2Rs than do compounds with spacious and flat structures [70].
The evolution of bitter receptors
The rapidly rising availability of whole-genome sequencing is providing, for the first time, perspectives on the evolution of TAS2Rs as a family. One of the most remarkable revelations from comparisons across vertebrates is that TAS2Rs are a relatively recent evolutionary innovation. Whereas they are present in the bony fishes, lobe-finned fishes and tetrapods, they are not found in cartilaginous fishes [64]. Thus, TAS2Rs must first have appeared in an aquatic species ∼430 million years ago [71]. This makes them the most recently derived of the chemosensory receptor families in vertebrates [64, 72, 73]. The pressures driving TAS2Rs' earliest origins can only be speculated upon, but, notably, vascular plants underwent an explosive radiation ∼430 million years ago likened to the Cambrian explosion in animals [74, 75]. The coincidence suggests that the origin of TAS2Rs may mark a pivotal transition in plant–herbivore interactions. However, there are compelling alternatives. In particular, evidence that TAS2Rs play roles outside of taste, especially nutrient sensing in the gut, suggests that TAS2Rs might have evolved to play those roles first with their function as taste receptors coming later.
Following its initial emergence, the TAS2R family diversified both within and among taxa. The estimated size of TAS2R repertoires continues to be refined as genome assemblies increase in precision, and trends are already evident (Fig. 5). The most conspicuous aspect of diversification is in the number of TAS2R loci in species’ genomes, which varies substantially. This is exemplified in fishes, in which gene counts range from one in Amazon molly (Poecilia formosa) to 74 in coelacanth (Latimeria chalumnae) [76–83]. However, most fishes carry an intermediate number, such as the 6 in Atlantic cod (Gadus morhua), 7 in zebra danio (Danio rerio) and 21 in blind cave fish (Astyanax mexicanus) [82]. Fish genomes also contain TAS2R pseudogenes. One is found in the spotted green puffer fish (Tetraodon nigroviridis) and three are found in A. mexicanus [82]. The presence of multiple TAS2Rs in most species in conjunction with the presence of both functional and non-functional loci suggest that birth-death processes are a key aspect of TAS2R evolution, although little is currently known about them.
Figure 5.

Size of bitter taste receptor gene repertoires in bony fish, amphibia, reptilia, aves and mammalia. Gene count is proportional to image height. Across species studied to date, the number ranges from 0 (in some marine mammals and birds) to 136 (in frogs). The large number of TAS2Rs in frogs may be due to their dietary reliance on insects, which often sequester secondary metabolites from consumed plants
Variation in the number of TAS2R genes occurs outside the fishes, as well. Among amphibians, cecilians have the smallest number (18–28) and anurans the largest of any recorded vertebrate (50–136) [76–79, 81, 84, 85]. Numbers in axolotl range from 35 to 49 depending on the study [76, 85]. The sauria, a highly heterogeneous taxon that includes the crocodilians, lizards, snakes and birds, are similarly variable. Snakes have low numbers (0–2) whereas crocodiles and turtles have more (5–9 and 5–11, respectively), and lizards have 36–50, making them comparable to many fishes [76, 77, 79, 84, 86, 87]. TAS2R repertoires in birds, which are highly diverse ecologically, are highly diverse as well. Strikingly, a survey of five penguin species revealed that none bear functional TAS2Rs yet other avians such as zebra finch (Taeniopygia guttata) and white-throated sparrow (Zonotrichia albicollis) do have them, hosting 7 and 17, respectively [88]. Mammals have intermediate numbers of TAS2Rs compared to other taxa, such as the 7 in platypus, 16–27 in marsupials and ∼35 in rodents [28, 79]. However, like penguins, the cetacea lack or nearly lack TAS2Rs altogether, with 0 or 1 depending on the species [89, 90]. Most higher primates have roughly 25 intact TAS2Rs and 10 pseudogenes (Fig. 6). The lowest number is found in gibbon (Nomascus leucogenys), which possesses 18 intact and 8 pseudogenes. Humans' closest relatives, chimpanzee, gorilla and orangutan possess 26, 24 and 23 intact loci, respectively.
Figure 6.

Intact genes and pseudogenes in primates. All primates studied to date harbor both intact genes and pseudogenes. However, their numbers vary even across closely related taxa, such as humans and chimpanzees. This suggests that gene birth–death processes are an important mechanism of adaptation to toxin exposure. Redrawn from Ref. [79]
While toxins are a threat to any animal eating plants, species are not equally vulnerable. For instance, carnivores rely minimally on plants and have little need to manage exposure. In contrast, leaf eating monkeys are constantly exposed and must adapt accordingly. This suggests that TAS2Rs have evolved under different selective pressures in different species. Evidence for such differences is reflected in the size of TAS2R repertoires, which correlate with the proportion of species' diets composed of plants [79, 91]. The most conspicuous trend is that repertoires are larger in herbivores than in carnivores. The most extreme case, the complete absence of TAS2Rs, is found in obligate carnivores that gulp rather than chew their food including whales, penguins and pinnipeds [79, 89, 91–93] (Hawaiian monk seal genome assembly neoSch1, 2017). Explanations for the opposite end of the repertoire size spectrum, in anurans, are less obvious. However, it may relate to their being insectivores. Many insects sequester secondary compounds from plants in their diets, coopting them for use in their own defense systems [94]. Examples abound, with the classic being monarch butterflies, which sequester cardenolides found in their food plant, milkweed. Insects' use of plant toxins suggests that even though they are not herbivores anurans encounter a broad spectrum of phytotoxins indirectly and benefit from being able to detect overly hazardous prey.
Although selective pressures on taste perception are the most obvious potential driver of diversification in TAS2Rs, they are probably not the only driver. The extraoral roles of TAS2Rs point to pressures arising from their other functions, as well. For instance, because bitter perception shapes diet, which in turn shapes the gut microbiome, selective pressures on TAS2Rs could originate from both roles simultaneously. Likewise, ambient air conditions vary from environment to environment, which could exert pressures on TAS2Rs mediating respiratory responses. Thus, selective pressures on TAS2Rs likely arise from their diverse roles in multiple systems, not solely from their role in perception.
Population genetics
TAS2Rs exhibit variability within species as well as between them. The earliest evidence for intraspecific variation was reported in 1932 by Fox [66], who observed that PTC sensitivity varies profoundly among individuals in most populations. Blakeslee [95] confirmed shortly thereafter that PTC perception is indeed heritable and is best explained by the presence of dominant and recessive alleles of a single gene, although the specific gene involved would remain unknown for nearly 70 years. These went on to be recognized as the famous PTC “taster” and “non-taster” alleles. The identity of the locus underlying PTC sensitivity was finally determined by Kim et al. [96], who discovered that two alleles of TAS2R38 explain 50–80% of variance in the trait. Further, the two alleles differed by three amino acid changes, which often alter receptor function. And, as predicted by their amino acid differences, the two alleles exhibit divergent responses to PTC in vitro [97], corresponding to the long hypothesized taster and non-taster types. The finding that several amino acid variants occur in TAS2R38, along with the discovery that humans carry not one but 25 TAS2R loci, suggested that variation in TAS2Rs and their associated phenotypes could be far reaching.
Owing to its early discovery and role in shaping a classic phenotype, TAS2R38 is the most intensively investigated and best-understood TAS2R from a population genetic standpoint. In the first study aimed at documenting global sequence variation at TAS2R38, Wooding et al. [98] identified five variable amino acid positions in 165 subjects from Africa, Asia, Europe and North America, and they were configured into seven alleles. Notably, the canonical taster and non-taster alleles mapped by Kim et al. [96] were present at roughly equal frequencies and jointly accounted for 96% of the sample, with the remaining five accounting for just 4%. These findings confirmed that TAS2R38 harbors a number of amino acid substitutions that might affect taste phenotypes but diversity worldwide is dominated by the three distinguishing the classic taster and non-taster variants. In the largest study of TAS2R38 to date, Risso et al. [99] examined >5500 worldwide subjects and reported findings congruent with previous results. Twenty-four alleles were present, with the taster and non-taster variants distributed worldwide and a third common in Africa but rare elsewhere.
Insights into the population genetics of TAS2Rs are no longer limited to TAS2R38. It is now known that allelic variation is abundant across the TAS2R family as a whole. In a study of all 25 TAS2Rs in 55 subjects, Kim et al. [100] identified 144 nucleotide changes with a range from 1 (in TAS2R13) to 12 (in TAS2R48), 108 (75%) of which result in amino acid changes. In a survey of all TAS2Rs across 22 subjects, Wang et al. [101] obtained similar results, observing 105 changes with a range of 0 (TAS2R7) to 12 (TAS2R49), 72 (70%) of which result in amino acid changes. These results demonstrate that all TAS2Rs harbor variation likely to shape perception, and it is distributed widely across populations.
Studies of natural selection in additional TAS2Rs further illuminate the evolutionary complexity of bitter perception. An instructive example is TAS2R16, which is responsive to glucopyranosides [102, 103]. Like TAS2R38, TAS2R16 is represented by three primary alleles, which are defined by two amino acid changes. Also like TAS2R38, TAS2R16 harbors two globally distributed alleles, along with a third one common in Africans but rare in non-Africans. However, TAS2R16 contains a greater proportion of low-frequency nucleotide variants than does TAS2R38, a pattern expected under positive selection, not balancing selection [102, 103]. In addition, within Africa, some human alleles are unexpectedly similar to those of chimpanzees, which is indicative of strong purifying selection [102]. This suggests that reduced ability to perceive glucopyranosides is for some reason favored there. A speculative explanation offered by Soranzo et al. [103] is that low sensitivity to glucopyranosides could be favored if it results in greater consumption of them, which is thought to protect against malaria. However, this hypothesis was not supported by the results of Campbell et al. [102], who found no correlation between malaria prevalence and the frequency of the proposed protective allele. These findings reiterate the trends in TAS2R38, that selection on TAS2Rs appears to be relaxed for some haplotypes but not others, and in some populations but not others. Similar complexity likely spans the rest of the TAS2R family, but it remains to be investigated.
Health importance of bitter taste
Contemporary human diets rely principally on crops domesticated in the last 10 000 years, which were selectively bred to maximize productivity while minimizing the hazards of consumption [104]. Many are derived from ancestors that had low toxicity such as grasses, and selective breeding mainly improved calorie content. Such crops include rice, corn and wheat, which originated from species that were not toxic but were laborious to gather and process. However, some crops do originate from toxic predecessors. The most familiar example is potato, whose wild varieties produce alkaloids that render them inedible. Other examples include squashes, whose wild forms produce cucurbitacin, and beans, which produce phytohemagglutinin. Domestication of these species reduced toxicity to levels safe to consume with little preparation. However, plant toxins do remain in human diets, in some cases at high levels, and their bitterness is an important gauge of food safety.
The model example of bitter taste’s significance to food safety and disease involves cassava (Manihot esculenta). Though poorly known in industrialized countries, cassava is a major global crop. It ranks seventh in tonnage produced among crops worldwide and is a staple source of calories for ∼500 million people throughout the tropics [105]. However, cassava’s wide use belies a threat—it utilizes a highly toxic anti-herbivore defense system. When macerated, as by chewing, cassava releases neurotoxic nitriles and cyanide when an enzyme-substrate pair produced by the plant, linamarase and linamarin, react [11, 106]. Cassava's toxicity is valuable from an agricultural standpoint because it allows cultivation without pesticides or the manual labor needed to manage more delicate crops, reducing work and financial burden. However, cassava’s defense system is toxic to humans as well as pests. High dose exposures lead to quick death, and chronic exposures lead to nerve damage permanently impairing dexterity, locomotion and cognition [107]. Poisoning is so common in some regions of Africa that the resulting syndrome has a name, konzo (bound legs, in the Yaka language) (Fig. 7). Konzo is commonly found at frequencies ∼1–3% in affected areas and has been documented at frequencies >15% in extreme cases [108–111].
Figure 7.

Cassava toxicity and konzo. (A) Konzo patients in the Democratic Republic of Congo. Note foot position of child and crutch used by an adult. Photograph courtesy of Dr Thorkild Tylleskär, University of Bergen. (B) Plot of cyanogenic glucoside content and perceived bitterness of samples tasted by farmers in Malawi. Higher taste score indicates higher perceived bitterness. Redrawn from Ref. [112]
Cassava's toxicity can be eliminated prior to consumption through grinding and shredding, which drive its reactions to completion rapidly, or by peeling and soaking or drying, which do so more slowly. However, these processes are time consuming and energy intensive. This imposes countervailing pressures on growers, who must aim for toxicity high enough to protect the crop but low enough to avoid unnecessary labor removing it. Bitter taste helps resolve the conflict. Cassava cultivars vary in bitterness, and bitterness predicts toxicity [112]. Moreover, cassava’s bitterness is imparted by linamarin, the metabolic precursor to cassava's toxins, so taste and toxicity are directly connected [113]. Experienced growers are experts at assessing the toxicity of their crops based on taste, which they use for selecting which cultivars to grow and anticipating the effort required to process them (Fig. 7). Bitterness can also be used to monitor the level of residual toxicity as cassava is processed, ensuring the end product is safe to eat.
A second example of connections between taste, toxicity and health involves the traditional cultivation of potato (Solanum tuberosum) in Andean populations [114]. Potatoes are in the plant family Solanaceae, which is noted for its many toxic members including nightshade (Atropa spp.), jimsonweed (Datura spp.) and tobacco (Nicotiana spp.). Like many species in the family, potatoes naturally synthesize the glycoalkaloids α-solanine and α-chaconine as a mechanism of self-defense. Here again, bitter taste plays a protective role. In a study of taste perception and potato use in the Aymara people of Bolivia, Johns and Keen [115] found that growers have an elaborate vocabulary for describing potato characteristics. Two key terms are luq’i (bitter) and ch’oke (non-bitter) [115, 116]. Luq’i and ch’oke correlate with glycoalkaloid content and express whether content is too high or acceptable, and how the potatoes must be processed to make them safely edible [115].
While cassava and potatoes illustrate bitter perception’s protective functions, they are unusual in the current world. Today’s food supplies are overwhelmingly safe, and not only are bitter foods usually harmless in the quantities consumed by humans they are sometimes preferred. Such foods demonstrate that aversions driven by bitterness are not always adaptive, and can be maladaptive if they result in the rejection of viable sources of nutrition [117]. For instance, bitter melon (Momordica charantia) is widely used in African and Asian cuisines and consumption shows no association with disease [118]. Similarly, citrus fruits (Citrus spp.) are often bitter but non-toxic [119]. Rejecting these foods results in the rejection of the calories, vitamins and other nutrients they contain.
The volume and quality of modern diets make the rejection of many foods inconsequential. However, this is not always the case. The classic example of bitter perception undermining health involves cruciferous vegetables, which include common table offerings such as cabbage and Brussels sprouts. Vegetables are an essential part of the diet, yet they are chronically under-consumed worldwide [120–122]. The reasons for this are complex. However, in the case of crucifers, bitter taste is an important contributor. Crucifers utilize a defense system analogous to that of cassava in which a glucoside (sinigrin) stored by the plant is hydrolyzed by an enzyme (myrosinase) when tissue damage occurs, producing a burst of noxious compounds [123, 124]. The products of sinigrin–myrosinase reactions are easily tolerated even by children and reports of overexposure are rare. Furthermore, some products of sinigrin–myrosinase reactions have anti-cancer properties [125–127]. However, one of the products of glucosinolate–myrosinase reactions, goitrin, is intensely bitter [17]. As a result, while crucifers are an abundant and readily available source of nutrients lacking in most diets, and may even have unique advantages, they are often rejected as unpalatable [13, 15, 128, 129].
A seemingly paradoxical observation about bitterness is that it is not always aversive, and can even be attractive. For instance, tonic water, which contains quinine, is a bitter yet popular drink. Other examples include bitter vegetables such as kale, coffee, tobacco smoke and many beers. The factors driving food preferences as opposed to perception alone are complex and the basis for bitter attraction is still being unraveled [10]. However, general observations offer explanations. First, humans' cognitive abilities enable them to learn from one another, and learned information about a bitter food's being safe may override instinctive aversions. Second, humans are adept at learning through trial and error. If a bitter food is tentatively consumed without ill effects, it may be consumed again. Further, an observation offered by Reed and Knaapila is that beyond being harmless some bitter substances have rewarding effects, with caffeine and nicotine being examples, which could also drive attraction [10]. These alternatives are not mutually exclusive. In the case of coffee, for instance, people may be prompted to try it by observing its popularity, continue drinking it once it is discovered to be not only harmless but pleasurable, and eventually come to prefer it.
Another seeming paradox surrounding attractions to plant toxins is their traditional use as pharmaceuticals [130]. Myriad plant species are consumed for their healing properties, and many are noted for their bitterness, which can be a sign of potency [131, 132]. The compounds responsible are usually unknown because isolating plants' active constituents and testing their efficacy is such a challenge. Moreover, many medicines are derived from multiple species, which may contain compounds with synergistic effects. However, the specific plant compounds involved are known in a growing number of cases, and TAS2Rs responsive to them are known for a handful. The clearest example is salicin, a relative of aspirin found in willow (Salix spp.) that has pain killing and anti-fever properties and is an agonist of TAS2R16 [63, 133]. Others include papaverine, from poppy (Papaver spp.) (vasodilator; an agonist of TAS2R7, -10 and -14) and quinine (anti-malarial; agonist of multiple TAS2Rs).
Remarkably, the use of bitter plants as medicines extends to non-humans, which exhibit zoopharmognosy (animal knowledge of pharmaceuticals) [134]. Many instances of animals ingesting toxic plants for their pharmacological effects are documented [134]. The plants are frequently bitter, suggesting that taste is a signal of therapeutic value to animals just as it is for humans. In addition, the ingested plants frequently harbor compounds in classes eliciting responses from TAS2Rs, such as glycosides [134]. Some are also known to have specific health effects, such as anti-parasite activity. An especially well-documented connection between bitter taste and animal self-medication is in chimpanzees (Pan troglodytes) [135]. In an investigation of chimpanzee populations in Tanzania, Koshimizu et al. [135] found that a selectively ingested plant (Vernonia amygdalina) harbors bitter compounds with potent effects on intestinal parasites. The implicated compounds included sesquiterpene lactones and glucosides, which are often agonists of human TAS2Rs, implying they are also TAS2R agonists in chimpanzees. Consumption of other plants by chimpanzees have been found to reduce parasite load in the gut, and it seems likely they have similarities to Vernonia amygdalina with respect to taste and medicinal effect [136].
Genetic variation and health
Potential connections between variation in bitter taste sensitivity and dietary preferences were recognized early on in taste research and continue to be the focus of much investigation [10, 137, 138]. Initial efforts to dissect taste–diet relationships took advantage of the high heritability of sensitivity to PTC and its close relative 6-n-propylthiouracil (PROP), which justified their use as genetic markers prior to the discovery of TAS2R38 [137, 139–141]. Treating PTC sensitivity as a genetic marker was a successful approach, but it was constrained by several factors including the facts that PTC and PROP are synthetic and never encountered in nature, they are imperfect indicators of genotype, and they are weak predictors of variation in the perception of other compounds. Current approaches are capitalizing on genotypic data obtained directly from TAS2Rs, which provide a more precise tool for ascertaining genetic effects on the perception of specific bitter compounds as well as foods and drinks.
The range of bitter compounds known to elicit variable taste responses due to polymorphism in TAS2Rs is rising rapidly, particularly with respect to substances found in the diet. Most have been established using a combination of association studies in subjects and functional analyses in vitro. For instance, the perceived bitterness of goitrin is now known to associate with variants of TAS2R38, which encodes receptor isoforms exhibiting functionally divergent responses to goitrin in vitro [17]. Similarly, taste responses to the artificial sweeteners acesulfame K and saccharin associate with SNPs in TAS2R31, and responses to stevioside, a natural non-caloric sweetener, associate with SNPs in TAS2R4 and TAS2R14 [142–144]. Other examples include grosheimin (found in artichoke; TAS2R19, -31 and -50), and compounds in hops (TAS2R1, -14 and -40) [145, 146]. Quinine, an ingredient of tonic water, exhibits a notoriously complex relationship with TAS2Rs. It activates nine in vitro and taste responses in subjects associate with variants in two, TAS2R19 and -31 [63, 147, 148]. Compounds rare in the diet have also been investigated. Perception of aristolochic acid, a food contaminant originating from Aristolochia clematitis in Eastern Europe, associates with polymorphism in TAS2R43, and salicin perception associates with variants in TAS2R16 [102, 144, 149]. Additional substances show genotype–phenotype associations that are compelling but not yet validated such as capsaicin, piperine and ethanol (which associate with SNPs in TAS2R3, -4 and -5) [150]. Evidence that polymorphism in TAS2Rs shapes sensitivity to specific compounds suggests they shape the preferences for foods containing them, as well, with consequences for consumption patterns and health.
The earliest persuasive evidence for connections between genetic variation in taste, diet and health was reported by Greene in 1974 [151]. In an investigation of endemic goiter in Andean populations, Greene found that PTC sensitivity was highly variable and associated with disease, with low sensitivity associating with high susceptibility. In addition, the Andeans’ diets were rich in bitter vegetables implicated as being goitrogenic. Greene concluded that the PTC taster allele confers aversion to goitrogen containing vegetables, providing protection against their negative effects. Thus, the observed association between taste and goiter had a simple explanation. It also raised an evolutionary question: if taste sensitivity to goitrogens confers protection against exposure, why is the taster allele not fixed in the population? Greene speculated that it arises from changes in goiter susceptibility through the lifespan, with children being more susceptible than adults. Under these conditions heterozygotes with respect to the taster and non-taster alleles might have a reproductive advantage if they exhibit intermediate taste sensitivity. This would maintain both alleles in the population.
The most extensively investigated relationship between genetic variation, perception of foods and diet choices connects TAS2R38 genotype and vegetable intake. It is well established that the perceived bitterness of vegetables, particularly cruciferous vegetables, is negatively associated with liking and intake [14]. In addition, the perceived bitterness of goitrin, a major constituent of cruciferous vegetables, associates with alleles of TAS2R38 [17]. TAS2R38 genotype also associates with perceived bitterness and liking of cruciferous vegetables themselves, not just goitrin [129, 152]. Further, although TAS2R38 genotype associates with the perceived bitterness of cruciferous vegetables, it does not associate with the perceived bitterness of non-cruciferous vegetables, which is expected if goitrin content is driving the relationship [153]. The patterns are further supported by evidence that the associations are weaker for vegetables overall than for cruciferous vegetables in particular, which follows because not all vegetables are crucifers [154, 155]. Together these findings support the long-held but difficult to test hypothesis that heritable variation in bitter taste sensitivity drives preferences and consumption.
Insights into relationships between TAS2R38 genotype, diet and health are well developed due to PTC sensitivity’s high heritability and early discovery. However, similar relationships involving other TAS2Rs and health behaviors are being uncovered. Some involve compounds not encountered by humans until recently. For instance, variation in the perceived bitterness and intake of alcohol associates with polymorphism in TAS2R3, -4, -5, -16 and -19 as well as -38 [150, 156–159]. Ethanol is not completely absent from the natural environment (it sometimes occurs in fermenting fruits), but it is rare and seems unlikely to be specifically targeted by TAS2Rs. A simpler explanation is that ethanol stimulates some TAS2R isoforms incidentally, imparting bitterness. Similarly, variation in TAS2R38 associated with habitual tobacco use, another recent phenomenon [160–163]. TAS2R38 could not have evolved under selective pressures arising from tobacco use. However, smoke contains hundreds of combustion products that could act as TAS2R agonists incidentally, driving associations. Relationships with more distant connections to health are also known, such as the association of TAS2R16 alleles with the perceived bitterness of espresso coffee and grapefruit juice [164]. The implication of findings like these is that although variation in TAS2Rs is a holdover from ancient processes, and rarely influences exposure to toxins encountered through foraging, it retains its importance to modern patterns of perception and behavior.
Evidence that TAS2Rs mediate health behaviors such as alcohol and tobacco use suggests that they associate with their downstream consequences, as well. However, evidence for such effects has been elusive. To be detectable, not only does variation in bitter perception have to affect behavior, the effect of the behavior on health has to be evident amidst myriad other factors. The challenge is illustrated by patterns of association between bitter perception and body fat. The role of TAS2Rs in shaping food preferences suggests that bitter perception is likely associated with measures such as body mass index (BMI), and such associations do exist. For instance, taste sensitivity to PROP associates with measures of body fat in some populations [165–169]. However, the associations are not universal [155, 170–172]. Moreover, while TAS2R38 genotypes are strongly associated with taste sensitivity to PROP, which associates with BMI, associations between TAS2R38 genotypes themselves and BMI are weak or absent [172–174]. The simplest explanation for these patterns is that PROP sensitivity is determined by factors in addition to TAS2R38 genotype and it is overall taste sensitivity, not TAS2R38-mediated sensitivity alone, that drives the associations [175].
Here again, it is important to note the potential importance of extraoral TAS2Rs. The roles of TAS2Rs in hormonal responses and bronchodilation in the gut and lungs, imply that genetic variation in TAS2Rs could exert health effects via those mechanisms. Findings from genotype-phenotype association studies targeting TAS2Rs emphasize their potential importance. Consistent with evidence that ingestion of bitter compounds can produce hormonal responses in gut cells, Dotson et al. [176] found that variation in TAS2R9 associates with glucose dysregulation and Clark et al. [177] found that variation in TAS2R42 associates with thyroid hormone levels. Similarly, in an investigation of variation of associations between TAS2R38 variants and respiratory health, Lee et al. [178] found that variation in TAS2R38 associates with susceptibility to infection.
It is important to recognize that the elusiveness of direct associations between genotypic variation in TAS2Rs and complex health measures such as BMI does not imply that they are unimportant. First, associations between variation in TAS2Rs and behaviors such as vegetable consumption and tobacco use are consistent. These behaviors have health impacts, and understanding the role of TAS2Rs in shaping them will reveal strategies for modifying them, improving health outcomes. Second, even if associations between TAS2R variation and complex traits such as BMI are weak statistically, they are potentially important on population scales, which are the primary focus of public health. That is, effects so small they are difficult to detect in samples of hundreds or thousands of subjects could be large enough to merit intervention in populations composed of hundreds of thousands or millions of individuals.
CONCLUSIONS
Insights into the evolutionary processes underlying bitter taste perception continue to shed light on the complex connections between ancient evolutionary processes and modern human health. They also continue to expand. While early studies focused on single TAS2R loci, and then on TAS2R repertoires in select taxa, developments in genomics are accelerating the pace at which data are emerging. Whole-genome sequencing, in particular, is providing a basis for comparing TAS2R genes and gene repertoires within and across species at high resolution. As the focal point of biomedical and health research, human data are being generated at an especially high rate. This is fostering population genetic research with a global outlook and association analyses in the largest phenotyped cohorts studied to date, offering opportunities for research on new scales.
The power of coming evolutionary studies is illustrated by the Vertebrate Genomes Project (VGP), an initiative to sequence the complete genomes of ∼70 000 vertebrate species (vertebrategenomesproject.org) [179]. The VGP and related efforts are providing access to data not only from large numbers of species but from species outside the research mainstream, which typically focuses on mammals, especially primates. Gene birth–death processes are a particularly promising line of research. Given that the number of functional TAS2Rs varies from 0 to more than 100 across species it is clear that birth–death processes are an essential feature of TAS2Rs' evolution, yet little is known about them. Genomic data from novel species also provide inventories of homologous but functionally divergent TAS2R isoforms, which are a resource in efforts to dissect the molecular basis of receptor response. Similarly, large-scale resequencing projects within humans, which now extend beyond the 1000 Genomes Project, are providing genetic portraits of global populations at unprecedented levels of detail.
Like evolutionary studies, efforts to define taste–health connections are capitalizing on the advent of genomic technologies. While early efforts were limited to tens or hundreds of subjects, the economics of data collection are producing datasets composed of tens of thousands or even hundreds of thousands of subjects [96, 180, 181]. Data on this scale offer high statistical power, making them a promising resource for resolving relationships between TAS2R polymorphism and health traits. They are also potentially able to incorporate evolutionary evidence for the functional importance of specific sites to make prior predictions about phenotypic effects. However, most large scale studies to date have focused on associations between genome-wide markers and health traits generally, rather than on TAS2Rs and taste phenotypes specifically. An essential step moving forward will be the establishment of cohorts with phenotyping components directly aimed at sensory phenotypes, behaviors and outcomes.
Conflict of interest: None declared.
REFERENCES
- 1.Ungar P, Teaford M.. Perspectives on the evolution of human diet. In: Ungar P., Teaford M., (eds). Human Diet: Its Origins and Evolution. Westport, CT: Bergen & Garvey, 2002, 1–6. [Google Scholar]
- 2.Walters D. Plant Defense: Warding off Attack by Pathogens, Herbivores, and Parasitic Plants. Chichester, UK: John Wiley & Sons Ltd., 2011. [Google Scholar]
- 3.Crozier A, Clifford MN, Asihara H.. Plant Secondary Metabolites: Occurrence, Structure, and Role in the Human Diet. Oxford, UK: Blackwell Publishing, Ltd, 2006, 25–46. [Google Scholar]
- 4.Kessler A, Kalske A.. Plant secondary metabolite diversity and species interactions. Annu Rev Ecol Evol Syst 2018;49:115–38. [Google Scholar]
- 5.Dayan AD. What killed Socrates? Toxicological considerations and questions. Postgrad Med J 2009;85:34–7. [DOI] [PubMed] [Google Scholar]
- 6.Bradberry SM, Dickers KJ, Rice P. et al. Ricin poisoning. Toxicol Rev 2003;22:65–70. [DOI] [PubMed] [Google Scholar]
- 7.Dickers KJ, Bradberry SM, Rice P. et al. Abrin poisoning. Toxicol Rev 2003;22:137–42. [DOI] [PubMed] [Google Scholar]
- 8.Freeland W, Janzen D.. Strategies in herbivory by mammals: the role of plant secondary compounds. Am Nat 1974;108:269–89. [Google Scholar]
- 9.Kohl KD, Weiss RB, Cox J. et al. Gut microbes of mammalian herbivores facilitate intake of plant toxins. Ecol Lett 2014;17:1238–46. [DOI] [PubMed] [Google Scholar]
- 10.Reed DR, Knaapila A.. Genetics of taste and smell: poisons and pleasures. Prog Mol Biol Transl Sci 2010;94:213–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Llorens J, Soler-Martín C, Saldaña-Ruíz S. et al. A new unifying hypothesis for lathyrism, konzo and tropical ataxic neuropathy: nitriles are the causative agents. Food Chem Toxicol 2011;49:563–70. [DOI] [PubMed] [Google Scholar]
- 12.Astwood EB, Greer MA, Ettlinger MG.. L-5-vinyl-2-thiooxazolidone, an antithyroid compound from yellow turnip and from Brassica seeds. J Biol Chem 1949;181:121–30. [PubMed] [Google Scholar]
- 13.Fenwick GR, Griffiths NM.. The identification of the goitrogen (-)5-vinyloxazolidine-2-thione (goitrin), as a bitter principle of cooked brussels sprouts (Brassica oleracea L. var. gemmifera). Z Lebensm Unters Forsch 1981;172:90–2. [DOI] [PubMed] [Google Scholar]
- 14.Dinehart ME, Hayes JE, Bartoshuk LM. et al. Bitter taste markers explain variability in vegetable sweetness, bitterness, and intake. Physiol Behav 2006;87:304–13. [DOI] [PubMed] [Google Scholar]
- 15.Drewnowski A, Gomez-Carneros C.. Bitter taste, phytonutrients, and the consumer: a review. Am J Clin Nutr 2000;72:1424–35. [DOI] [PubMed] [Google Scholar]
- 16.Johnston CS, Taylor CA, Hampl JS.. More Americans are eating “5 a day” but intakes of dark green and cruciferous vegetables remain low. J Nutr 2000;130:3063–7. [DOI] [PubMed] [Google Scholar]
- 17.Wooding S, Gunn H, Ramos P, et al. Genetics and bitter taste responses to goitrin, a plant toxin found in vegetables. Chem Senses 2010;35:685–92. [DOI] [PubMed] [Google Scholar]
- 18.Fisher RA. Personal letter to Edmund Brisco Ford, September 26, 1939, in Sir Ronald Aylmer Fisher, Papers 1911-94, Correspondence Series 1, 1939.
- 19.Wooding S. Phenylthiocarbamide: a 75-year adventure in genetics and natural selection. Genetics 2006;172:2015–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Roper SD. The cell biology of vertebrate taste receptors. Annu Rev Neurosci 1989;12:329–53. [DOI] [PubMed] [Google Scholar]
- 21.Margolskee R. The molecular biology of taste transduction. Bioessays 1993;15:645–50. [DOI] [PubMed] [Google Scholar]
- 22.Kinnamon S, Margolskee R.. Mechanisms of taste transduction. Curr Opin Neurobiol 1996;6:506–13. [DOI] [PubMed] [Google Scholar]
- 23.Herness MS, Gilbertson TA.. Cellular mechanisms of taste transduction. Annu Rev Physiol 1999;61:873–900. [DOI] [PubMed] [Google Scholar]
- 24.Hoon MA, Adler E, Lindemeier J. et al. Putative mammalian taste receptors: a class of taste-specific GPCRs with distinct topographic selectivity. Cell 1999;96:541–51. [DOI] [PubMed] [Google Scholar]
- 25.Adler E, Hoon MA, Mueller KL. et al. A novel family of mammalian taste receptors. Cell 2000;100:693–702. [DOI] [PubMed] [Google Scholar]
- 26.Gilbertson T, Damak S, Margolskee R.. The molecular physiology of taste transduction. Curr Opin Neurobiol 2000;10:519–27. [DOI] [PubMed] [Google Scholar]
- 27.Kim U-K, Breslin PAS, Reed D. et al. Genetics of human taste perception. J Dent Res 2004;83:448–53. [DOI] [PubMed] [Google Scholar]
- 28.Shi P, Zhang J, Yang H. et al. Adaptive diversification of bitter taste receptor genes in mammalian evolution. Mol Biol Evol 2003;20:805–14. [DOI] [PubMed] [Google Scholar]
- 29.Wooding S, Ramirez V.. Taste. In: Fritzsch B., Meyerhof W. (eds). The Senses: A Comprehensive Reference. Cambridge, MA: Elsevier, Academic Press, 2020, 264–79. [Google Scholar]
- 30.Witt M. Anatomy and development of the human taste system. Handb Clin Neurol 2019;164:147–71. [DOI] [PubMed] [Google Scholar]
- 31.Roper SD, Chaudhari N.. Taste buds: cells, signals and synapses. Nat Rev Neurosci 2017;18:485–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chandrashekar J, Mueller KL, Hoon MA. et al. T2Rs function as bitter taste receptors. Cell 2000;100:703–11. [DOI] [PubMed] [Google Scholar]
- 33.Matsunami H, Montmayeur JP, Buck LB.. A family of candidate taste receptors in human and mouse. Nature 2000;404:601–4. [DOI] [PubMed] [Google Scholar]
- 34.Julius D, Nathans J.. Signaling by sensory receptors. Cold Spring Harb Perspect Biol 2012;4:a005991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Roper S. Signal transduction and information processing in mammalian taste buds. Pflugers Arch 2007;454:759–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ma Z, Tanis JE, Taruno A. et al. Calcium homeostasis modulator (CALHM) ion channels. Pflugers Arch 2016;468:395–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kinnamon SC, Finger TE.. Recent advances in taste transduction and signaling. F1000Res 2019;8:2117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Taruno A, Vingtdeux V, Ohmoto M. et al. CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes. Nature 2013;495:223–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lander ES, Linton LM, Birren B. et al. International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature 2001;409:860–921. [DOI] [PubMed] [Google Scholar]
- 40.Behrens M, Foerster S, Staehler F. et al. Gustatory expression pattern of the human TAS2R bitter receptor gene family reveals a heterogenous population of bitter responsive taste receptor cells. J Neurosci 2007;27:12630–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Deshpande DA, Wang WCH, McIlmoyle EL. et al. Bitter taste receptors on airway smooth muscle bronchodilate by localized calcium signaling and reverse obstruction. Nat Med 2010;16:1299–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Egan JM, Margolskee RF.. Taste cells of the gut and gastrointestinal chemosensation. Mol Interv 2008;8:78–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Jang H-J, Kokrashvili Z, Theodorakis MJ. et al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc Natl Acad Sci USA 2007;104:15069–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.McLaughlin S, McKinnon P, Margolskee R.. Gustducin is a taste-cell-specific G protein closely related to the transducins. Nature 1992;357:563–9. [DOI] [PubMed] [Google Scholar]
- 45.Azevedo MF, Faucz FR, Bimpaki E. et al. Clinical and molecular genetics of the phosphodiesterases (PDEs). Endocr Rev 2014;35:195–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Goldlust IS, Hermetz KE, Catalano LM. et al. Unique Rare Chromosome Disorder Support Group. Mouse model implicates GNB3 duplication in a childhood obesity syndrome. Proc Natl Acad Sci USA 2013;110:14990–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Huang L, Max M, Margolskee RF. et al. G protein subunit Gγ13 is coexpressed with Gαo, Gβ3, and Gβ4 in retinal ON bipolar cells. J Comp Neurol 2003;455:1–10. [DOI] [PubMed] [Google Scholar]
- 48.Kashio M, Wei-Qi G, Ohsaki Y. et al. CALHM1/CALHM3 channel is intrinsically sorted to the basolateral membrane of epithelial cells including taste cells. Sci Rep 2019;9:2681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kaske S, Krasteva G, König P. et al. TRPM5, a taste-signaling transient receptor potential ion-channel, is a ubiquitous signaling component in chemosensory cells. BMC Neurosci 2007;8:49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ma Z, Taruno A, Ohmoto M. et al. CALHM3 is essential for rapid ion channel-mediated purinergic neurotransmission of GPCR-mediated tastes. Neuron 2018;98:547–61.e10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mao GF, Vaidyula VR, Kunapuli SP. et al. Lineage-specific defect in gene expression in human platelet phospholipase C-β2 deficiency. Blood 2002;99:905–11. [DOI] [PubMed] [Google Scholar]
- 52.Maranto AR. Primary structure, ligand binding, and localization of the human type 3 inositol 1,4,5-trisphosphate receptor expressed in intestinal epithelium. J Biol Chem 1994;269:1222–30. [PubMed] [Google Scholar]
- 53.Mathar I, Jacobs G, Kecskes M. et al. TRPM4. Handb Exp Pharmacol 2014;222:461–87. [DOI] [PubMed] [Google Scholar]
- 54.Ogurtsov AY, Mariño-Ramírez L, Johnson GR. et al. Expression patterns of protein kinases correlate with gene architecture and evolutionary rates. PLoS One 2008;3:e3599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ritchey ER, Bongini RE, Code KA. et al. The pattern of expression of guanine nucleotide-binding protein β3 in the retina is conserved across vertebrate species. Neuroscience 2010;169:1376–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wettschureck N, Offermanns S.. Mammalian G proteins and their cell type specific functions. Physiol Rev 2005;85:1159–204. [DOI] [PubMed] [Google Scholar]
- 57.Calvo SS, Egan JM.. The endocrinology of taste receptors. Nat Rev Endocrinol 2015;11:213–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.An SS, Liggett SB.. Taste and smell GPCRs in the lung: evidence for a previously unrecognized widespread chemosensory system. Cell Signal 2018;41:82–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Cohen NA. The genetics of the bitter taste receptor T2R38 in upper airway innate immunity and implications for chronic rhinosinusitis. Laryngoscope 2017;127:44–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tran HTT et al. Allyl isothiocyanate: a TAS2R38 receptor-dependent immune modulator at the interface between personalized medicine and nutrition. Front Immunol 2021;12:669005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Fan Y, Pedersen O.. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol 2021;19:55–71. [DOI] [PubMed] [Google Scholar]
- 62.Luo X-C, Chen Z-H, Xue J-B. et al. Infection by the parasitic helminth Trichinella spiralis activates a Tas2r-mediated signaling pathway in intestinal tuft cells. Proc Natl Acad Sci USA 2019;116:5564–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Meyerhof W, Batram C, Kuhn C. et al. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem Senses 2010;35:157–70. [DOI] [PubMed] [Google Scholar]
- 64.Behrens M, Meyerhof W.. Vertebrate bitter taste receptors: keys for survival in changing environments. J Agric Food Chem 2018;66:2204–13. [DOI] [PubMed] [Google Scholar]
- 65.Bufe B, Hofmann T, Krautwurst D. et al. The human TAS2R16 receptor mediates bitter taste in response to beta-glucopyranosides. Nat Genet 2002;32:397–401. [DOI] [PubMed] [Google Scholar]
- 66.Fox AL. The relationship between chemical constitution and taste. Proc Natl Acad Sci USA 1932;18:115–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Suzuki N, Sugawara T, Matsui A. et al. Identification of non-taster Japanese macaques for a specific bitter taste. Primates 2010;51:285–9. [DOI] [PubMed] [Google Scholar]
- 68.Lossow K, Hübner S, Roudnitzky N. et al. Comprehensive analysis of mouse bitter taste receptors reveals different molecular receptive ranges for orthologous receptors in mice and humans. J Biol Chem 2016;291:15358–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Wooding S. Signatures of natural selection in a primate bitter taste receptor. J Mol Evol 2011;73:257–65. [DOI] [PubMed] [Google Scholar]
- 70.Di Pizio A, Niv MY.. Promiscuity and selectivity of bitter molecules and their receptors. Bioorg Med Chem 2015;23:4082–91. [DOI] [PubMed] [Google Scholar]
- 71.Brazeau MD, Friedman M.. The origin and early phylogenetic history of jawed vertebrates. Nature 2015;520:490–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Grus WE, Zhang J.. Origin of the genetic components of the vomeronasal system in the common ancestor of all extant vertebrates. Mol Biol Evol 2009;26:407–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Sharma K, Syed AS, Ferrando S. et al. The chemosensory receptor repertoire of a true shark is dominated by a single olfactory receptor family. Genome Biol Evol 2019;11:398–405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Harrison CJ, Morris JL.. The origin and early evolution of vascular plant shoots and leaves. Philos Trans R Soc Lond B Biol Sci 2018;373:20160496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Morris JL, Puttick MN, Clark JW. et al. The timescale of early land plant evolution. Proc Natl Acad Sci USA 2018;115: E2274–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Behrens M et al. At the root of T2R gene evolution: recognition profiles of coelacanth and zebrafish bitter receptors. Genome Biol Evol 2021;13. DOI: 10.1093/gbe/evaa264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Dong D, Jones G, Zhang S.. Dynamic evolution of bitter taste receptor genes in vertebrates. BMC Evol Biol 2009;9:12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Go Y; SMBE Tri-National Young Investigators. Proceedings of the SMBE Tri-National Young Investigators' Workshop 2005: lineage-specific expansions and contractions of the bitter taste receptor gene repertoire in vertebrates. Mol Biol Evol 2006;23:964–72. [DOI] [PubMed] [Google Scholar]
- 79.Li D, Zhang J.. Diet shapes the evolution of the vertebrate bitter taste receptor gene repertoire. Mol Biol Evol 2014;31:303–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Oike H, Nagai T, Furuyama A. et al. Characterization of ligands for fish taste receptors. J Neurosci 2007;27:5584–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Shi P, Zhang J.. Contrasting modes of evolution between vertebrate sweet/umami receptor genes and bitter receptor genes. Mol Biol Evol 2006;23:292–300. [DOI] [PubMed] [Google Scholar]
- 82.Shiriagin V, Korsching SI.. Massive expansion of bitter taste receptors in blind cavefish, Astyanax mexicanus. Chem Senses 2019;44:23–32. [DOI] [PubMed] [Google Scholar]
- 83.Syed AS, Korsching SI.. Positive Darwinian selection in the singularly large taste receptor gene family of an ‘ancient’ fish, Latimeria chalumnae. BMC Genomics 2014;15:650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Behrens M, Korsching SI, Meyerhof W.. Tuning properties of avian and frog bitter taste receptors dynamically fit gene repertoire sizes. Mol Biol Evol 2014;31:3216–27. [DOI] [PubMed] [Google Scholar]
- 85.Lu B, Jiang J, Wu H. et al. A large genome with chromosome-scale assembly sheds light on the evolutionary success of a true toad (Bufo gargarizans). Mol Ecol Resour 2021;21:1256–73. [DOI] [PubMed] [Google Scholar]
- 86.Zhong H, Shang S, Wu X. et al. Genomic evidence of bitter taste in snakes and phylogenetic analysis of bitter taste receptor genes in reptiles. PeerJ 2017;5:e3708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Zhong H, Shang S, Zhang H. et al. Characterization and phylogeny of bitter taste receptor genes (Tas2r) in Squamata. Genetica 2019;147:131–9. [DOI] [PubMed] [Google Scholar]
- 88.Davis JK, Lowman JJ, Thomas PJ. et al. NISC Comparative Sequencing Program. Evolution of a bitter taste receptor gene cluster in a New World sparrow. Genome Biol Evol 2010;2:358–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Feng P, Zheng J, Rossiter SJ. et al. Massive losses of taste receptor genes in toothed and baleen whales. Genome Biol Evol 2014;6:1254–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Zhu K, Zhou X, Xu S. et al. The loss of taste genes in cetaceans. BMC Evol Biol 2014;14:218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Jiang P, Josue J, Li X. et al. Major taste loss in carnivorous mammals. Proc Natl Acad Sci USA 2012;109:4956–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Liu Z, Liu G, Hailer F. et al. Dietary specialization drives multiple independent losses and gains in the bitter taste gene repertoire of Laurasiatherian Mammals. Front Zool 2016;13:28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Zhao H, Li J, Zhang J.. Molecular evidence for the loss of three basic tastes in penguins. Curr Biol 2015;25: R141–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Duffey DS. Sequestration of plant natural products by insects. Annu Rev Entomol 1980;25:447–77. [Google Scholar]
- 95.Blakeslee AF, Fox AL.. Our different taste worlds: P.T.C. as a demonstration of genetic differences in taste. J Hered 1932;23:97–107. [Google Scholar]
- 96.Kim U-k, Jorgenson E, Coon H. et al. Positional cloning of the human quantitative trait locus underlying taste sensitivity to phenylthiocarbamide. Science 2003;299:1221–5. [DOI] [PubMed] [Google Scholar]
- 97.Bufe B, Breslin PAS, Kuhn C. et al. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr Biol 2005;15:322–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Wooding S, Kim U-k, Bamshad MJ. et al. Natural selection and molecular evolution in PTC, a bitter-taste receptor gene. Am J Hum Genet 2004;74:637–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Risso DS, Mezzavilla M, Pagani L. et al. Global diversity in the TAS2R38 bitter taste receptor: revisiting a classic evolutionary PROPosal. Sci Rep 2016;6:25506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Kim U, Wooding S, Ricci D. et al. Worldwide haplotype diversity and coding sequence variation at human bitter taste receptor loci. Hum Mutat 2005;26:199–204. [DOI] [PubMed] [Google Scholar]
- 101.Wang X, Thomas SD, Zhang J.. Relaxation of selective constraint and loss of function in the evolution of human bitter taste receptor genes. Hum Mol Genet 2004;13:2671–8. [DOI] [PubMed] [Google Scholar]
- 102.Campbell MC, Ranciaro A, Zinshteyn D. et al. Origin and differential selection of allelic variation at TAS2R16 associated with salicin bitter taste sensitivity in Africa. Mol Biol Evol 2014;31:288–302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Soranzo N, Bufe B, Sabeti PC. et al. Positive selection on a high-sensitivity allele of the human bitter-taste receptor TAS2R16. Curr Biol 2005;15:1257–65. [DOI] [PubMed] [Google Scholar]
- 104.Stetter MG, Gates DJ, Mei W. et al. How to make a domesticate. Curr Biol 2017;27: R896–R900. [DOI] [PubMed] [Google Scholar]
- 105.FAO/IFAD. The World Cassava Economy: Facts, Trends, and Outlook. Rome, International Fund for Agricultural Development Food and Agriculture Organization of the United Nations, 2000. [Google Scholar]
- 106.McMohan J, WLB, Sayre W. et al. Cyanogenesis in cassava (Manihot esculenta Crantz). J Exp Bot 1995;46:731–41. [Google Scholar]
- 107.Tshala-Katumbay D, Mumba N, Okitundu L. et al. Cassava food toxins, konzo disease, and neurodegeneration in sub-Sahara Africans. Neurology 2013;80:949–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Banea J-P et al. Survey of the konzo prevalence of village people and their nutrition in Kwilu District, Bandundu Province, DRC. Afr J Food Sci 2015;9:43–50. [Google Scholar]
- 109.Boivin MJ, Okitundu D, Makila-Mabe Bumoko G. et al. Neuropsychological effects of konzo: a neuromotor disease associated with poorly processed cassava. Pediatrics 2013;131:e1231–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Kashala-Abotnes E, Okitundu D, Mumba D. et al. Konzo: a distinct neurological disease associated with food (cassava) cyanogenic poisoning. Brain Res Bull 2019;145:87–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Tylleskar T et al. Epidemiological evidence from Zaire for a dietary etiology of konzo, an upper motor neuron disease. Bull World Health Organ 1991;69:581–9. [PMC free article] [PubMed] [Google Scholar]
- 112.Chiwona-Karltun L, Brimer L, Kalenga Saka JD. et al. Bitter taste in cassava roots correlates with cyanogenic glucoside levels. J Sci Food Agric 2004;84:581–90. [Google Scholar]
- 113.King NLR, Bradbury JH.. Bitterness of Cassava: identification of a new apiosyl glucoside and other compounds that affect its bitter taste. J Sci Food Agric 1995;68:223–30. [Google Scholar]
- 114.Johns T. The Origins of Human Diet and Medicine. Phoenix, AZ: University of Arizona Press, 1996. [Google Scholar]
- 115.Johns T, Keen SL.. Taste evaluation of potato alkaloids by the Aymara: a case study in human chemical ecology. Hum Ecol 1986;14:437–52. [Google Scholar]
- 116.Hawkes J. On the origin and meaning of South American Indian potato names. Bot J Linn Soc 1947;53:205–50. [Google Scholar]
- 117.Glendinning JI. Is the bitter rejection response always adaptive? Physiol Behav 1994;56:1217–27. [DOI] [PubMed] [Google Scholar]
- 118.Tan SP, Kha TC, Parks SE, Roach PD.. Bitter melon (Momordica charantia L.) bioactive composition and health benefits: a review. Food Rev Int 2016;32:181–202. [Google Scholar]
- 119.Hasegawa S, Berhow MA, Fong CH.. Analysis of bitter principles in citrus. In: Linskens H.F. and Jackson J.F. (eds). Fruit Analysis. Berlin: Springer-Verlag, 1995, 59–80. [Google Scholar]
- 120.Hall JN, Moore S, Harper SB, Lynch JW.. Global variability in fruit and vegetable consumption. Am J Prev Med 2009;36:402–409.e5. [DOI] [PubMed] [Google Scholar]
- 121.Hung H-C, Joshipura KJ, Jiang R. et al. Fruit and vegetable intake and risk of major chronic disease. J Natl Cancer Inst 2004;96:1577–84. [DOI] [PubMed] [Google Scholar]
- 122.Yahia E. The contribution of fruit and vegetable consumption to human health. In: Yahia E. (ed.). Fruit and Vegetable Phytochemicals: Chemistry, Nutritional Value and Stability, Oxford, UK: Wiley-Blackwell, 2018, 3–52. [Google Scholar]
- 123.Fahey JW, Zalcmann AT, Talalay P.. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001;56:5–51. [DOI] [PubMed] [Google Scholar]
- 124.Kissen R, Rossiter J, Bones A.. The ‘mustard oil bomb’: not so easy to assemble?! Localization, expression and distribution of the components of the myrosinase enzyme system. Phytochem Rev 2009;8:69–86. [Google Scholar]
- 125.Shapiro TA et al. Chemoprotective glucosinolates and isothiocyanates of broccoli sprouts: metabolism and excretion in humans. Cancer Epidemiol Biomarkers Prev 2001;10:501–8. [PubMed] [Google Scholar]
- 126.Fahey JW, Zhang Y, Talalay P.. Broccoli sprouts: an exceptionally rich source of inducers of enzymes that protect against chemical carcinogens. Proc Natl Acad Sci USA 1997;94:10367–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Fahey JW, Haristoy X, Dolan PM. et al. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc Natl Acad Sci USA 2002;99:7610–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.van Doorn HE, van der Kruk GC, van Holst G-J. et al. The glucosinolates sinigrin and progoitrin are important determinants for taste preference and bitterness of Brussels sprouts. J Sci Food Agric 1998;78:30–8. [DOI] [PubMed] [Google Scholar]
- 129.Sandell M, Breslin P.. Variability in a taste-receptor gene determines whether we taste toxins in food. Curr Biol 2006;16: R792–4. [DOI] [PubMed] [Google Scholar]
- 130.Robles Arias DM, Cevallos D, Gaoue OG. et al. Non-random medicinal plants selection in the Kichwa community of the Ecuadorian Amazon. J Ethnopharmacol 2020;246:112220. [DOI] [PubMed] [Google Scholar]
- 131.Johns T. With Bitter Herbs They Shall Eat It: Chemical Ecology and the Origins of Human Diet and Medicine (Arizona Studies in Human Ecology). Tucson, AZ: University of Arizona Press, 1990, 356. [Google Scholar]
- 132.Vandebroek I, Moerman DE.. The anthropology of ethnopharmacology. In: Heinrich M., Jäger A.K. (eds). Ethnopharmacology. New York, NY: Wiley-Blackwell, 2015, 17–28. [Google Scholar]
- 133.Tawfeek N, Mahmoud MF, Hamdan DI. et al. Phytochemistry, pharmacology and medicinal uses of plants of the genus Salix: an updated review. Front Pharmacol 2021;12:593856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Huffman MA. Animal self-medication and ethno-medicine: exploration and exploitation of the medicinal properties of plants. Proc Nutr Soc 2003;62:371–81. [DOI] [PubMed] [Google Scholar]
- 135.Koshimizu K, Ohigashi H, Huffman M.. Use of Vernonia amygdalina by wild chimpanzee: possible roles of its bitter and related constituents. Physiol Behav 1994;56:1209–16. [DOI] [PubMed] [Google Scholar]
- 136.Huffman MA, Page JE, Sukhdeo MVK. et al. Leaf-swallowing by chimpanzees: a behavioral adaptation for the control of strongyle nematode infections. Int J Primatol 1996;17:475–503. [Google Scholar]
- 137.Drewnowski A, Henderson S, Barratt-Fornell A.. Genetic taste markers and food preferences. Drug Metab Dispos 2001;29:535–8. [PubMed] [Google Scholar]
- 138.Fox AL. A new approach to explaining food preferences. Abstracts of Papers of the American Chemical Society’s 126th Meeting, Division of Agriculture and Food Chemistry, 1954, 14.
- 139.Duffy V, Bartoshuk L.. Food acceptance and genetic variation in taste. J Am Diet Assoc 2000;100:647–55. [DOI] [PubMed] [Google Scholar]
- 140.Tepper BJ. 6-n-Propylthiouracil: a genetic marker for taste, with implications for food preference and dietary habits. Am J Hum Genet 1998;63:1271–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Drewnowski A, Rock CL.. The influence of genetic taste markers on food acceptance. Am J Clin Nutr 1995;62:506–11. [DOI] [PubMed] [Google Scholar]
- 142.Roudnitzky N, Bufe B, Thalmann S. et al. Genomic, genetic, and functional dissection of bitter taste responses to artificial sweeteners. Hum Mol Genet 2011;20:3437–49. [DOI] [PubMed] [Google Scholar]
- 143.Risso D, Morini G, Pagani L. et al. Genetic signature of differential sensitivity to stevioside in the Italian population. Genes Nutr 2014;9:401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Pronin AN, Xu H, Tang H. et al. Specific alleles of bitter receptor genes influence human sensitivity to the bitterness of aloin and saccharin. Curr Biol 2007;17:1403–8. [DOI] [PubMed] [Google Scholar]
- 145.Intelmann D, Batram C, Kuhn C. et al. Three TAS2R bitter taste receptors mediate the psychophysical responses to bitter compounds of Hops (Humulus lupulus L.) and Beer. Chemosensory Percept 2009;2:118–32. [Google Scholar]
- 146.Roudnitzky N, Behrens M, Engel A. et al. Receptor polymorphism and genomic structure interact to shape bitter taste perception. PLoS Genet 2015;11:e1005530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Hayes JE, Feeney EL, Nolden AA. et al. Quinine bitterness and grapefruit liking associate with allelic variants in TAS2R31. Chem Senses 2015;40:437–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Reed DR, Zhu G, Breslin PAS. et al. The perception of quinine taste intensity is associated with common genetic variants in a bitter receptor cluster on chromosome 12. Hum Mol Genet 2010;19:4278–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Wooding SP, Atanasova S, Gunn HC. et al. Association of a bitter taste receptor mutation with Balkan Endemic Nephropathy (BEN). BMC Med Genet 2012;13:2–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Nolden AA, McGeary JE, Hayes JE.. Differential bitterness in capsaicin, piperine, and ethanol associates with polymorphisms in multiple bitter taste receptor genes. Physiol Behav 2016;156:117–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Greene LS. Physical growth and development, neurological maturation and behavioral functioning in two Ecuadorian Andean communities in which goiter is endemic. II. PTC taste sensitivity and neurological maturation. Am J Phys Anthropol 1974;41:139–52. [DOI] [PubMed] [Google Scholar]
- 152.des Gachons CP, Beauchamp GK, Breslin PA.. The genetics of bitterness and pungency detection and its impact on phytonutrient evaluation. Ann N Y Acad Sci 2009;1170:140–4. [DOI] [PubMed] [Google Scholar]
- 153.Shen Y, Kennedy O, Methven L.. Exploring the effects of genotypical and phenotypical variations in bitter taste sensitivity on perception, liking and intake of brassica vegetables in the UK. Food Qual Preference 2016;50:71–81. [Google Scholar]
- 154.Duffy VB, Hayes JE, Davidson AC. et al. Vegetable intake in college-aged adults is explained by oral sensory phenotypes and TAS2R38 genotype. Chemosens Percept 2010;3:137–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Timpson NJ, Christensen M, Lawlor DA. et al. TAS2R38 (phenylthiocarbamide) haplotypes, coronary heart disease traits, and eating behavior in the British Women's Heart and Health Study. Am J Clin Nutr 2005;81:1005–11. [DOI] [PubMed] [Google Scholar]
- 156.Dotson CD, Wallace MR, Bartoshuk LM. et al. Variation in the gene TAS2R13 is associated with differences in alcohol consumption in patients with head and neck cancer. Chem Senses 2012;37:737–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Duffy VB, Davidson AC, Kidd JR. et al. Bitter receptor gene (TAS2R38), 6-n-propylthiouracil (PROP) bitterness and alcohol intake. Alcohol Clin Exp Res 2004;28:1629–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Ramos-Lopez O, Roman S, Martinez-Lopez E. et al. Association of a novel TAS2R38 haplotype with alcohol intake among Mexican-Mestizo population. Ann Hepatol 2015;14:729–34. [PubMed] [Google Scholar]
- 159.Wang JC, Hinrichs AL, Bertelsen S. et al. Functional variants in TAS2R38 and TAS2R16 influence alcohol consumption in high-risk families of African-American origin. Alcohol Clin Exp Res 2007;31:209–15. [DOI] [PubMed] [Google Scholar]
- 160.Cannon DS, Baker TB, Piper ME. et al. Associations between phenylthiocarbamide gene polymorphisms and cigarette smoking. Nicotine Tob Res 2005;7:853–8. [DOI] [PubMed] [Google Scholar]
- 161.Fox GL. The Archaeology of Smoking and Tobacco. Gainesville, FL: The University Press of Florida, 2015. [Google Scholar]
- 162.Mangold JE, Payne TJ, Ma JZ. et al. Bitter taste receptor gene polymorphisms are an important factor in the development of nicotine dependence in African Americans. J Med Genet 2008;45:578–82. [DOI] [PubMed] [Google Scholar]
- 163.Risso DS, Kozlitina J, Sainz E. et al. Genetic variation in the TAS2R38 bitter taste receptor and smoking behaviors. PLoS One 2016;11:e0164157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Hayes JE, Wallace MR, Knopik VS. et al. Allelic variation in TAS2R bitter receptor genes associates with variation in sensations from and ingestive behaviors toward common bitter beverages in adults. Chem Senses 2011;36:311–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Goldstein GL, Daun H, Tepper BJ.. Adiposity in middle-aged women is associated with genetic taste blindness to 6-n-propylthiouracil. Obes Res 2005;13:1017–23. [DOI] [PubMed] [Google Scholar]
- 166.Keller KL, Reid A, MacDougall MC. et al. Sex differences in the effects of inherited bitter thiourea sensitivity on body weight in 4-6-year-old children. Obesity (Silver Spring) 2010;18:1194–200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Padiglia A, Zonza A, Atzori E. et al. Sensitivity to 6-n-propylthiouracil is associated with gustin (carbonic anhydrase VI) gene polymorphism, salivary zinc, and body mass index in humans. Am J Clin Nutr 2010;92:539–45. [DOI] [PubMed] [Google Scholar]
- 168.Tepper BJ, Ullrich NV.. Influence of genetic taste sensitivity to 6-n-propylthiouracil (PROP), dietary restraint and disinhibition on body mass index in middle-aged women. Physiol Behav 2002;75:305–12. [DOI] [PubMed] [Google Scholar]
- 169.Tepper BJ, Koelliker Y, Zhao L. et al. Variation in the bitter-taste receptor gene TAS2R38, and adiposity in a genetically isolated population in Southern Italy. Obesity (Silver Spring) 2008;16:2289–95. [DOI] [PubMed] [Google Scholar]
- 170.Baranowski JC, Baranowski T, Beltran A. et al. 6-n-Propylthiouracil sensitivity and obesity status among ethnically diverse children. Public Health Nutr 2010;13:1587–92. [DOI] [PubMed] [Google Scholar]
- 171.Drewnowski A, Kristal A, Cohen J.. Genetic taste responses to 6-n-propylthiouracil among adults: a screening tool for epidemiological studies. Chem Senses 2001;26:483–9. [DOI] [PubMed] [Google Scholar]
- 172.Drewnowski A, Henderson SA, Cockroft JE.. Genetic sensitivity to 6-n-propylthiouracil has no influence on dietary patterns, body mass indexes, or plasma lipid profiles of women. J Am Diet Assoc 2007;107:1340–8. [DOI] [PubMed] [Google Scholar]
- 173.Deshaware S, Singhal R.. Genetic variation in bitter taste receptor gene TAS2R38, PROP taster status and their association with body mass index and food preferences in Indian population. Gene 2017;627:363–8. [DOI] [PubMed] [Google Scholar]
- 174.Ortega FJ, Agüera Z, Sabater M. et al. Genetic variations of the bitter taste receptor TAS2R38 are associated with obesity and impact on single immune traits. Mol Nutr Food Res 2016;60:1673–83. [DOI] [PubMed] [Google Scholar]
- 175.Hayes JE, Bartoshuk LM, Kidd JR. et al. Supertasting and PROP bitterness depends on more than the TAS2R38 gene. Chem Senses 2008;33:255–65. [DOI] [PubMed] [Google Scholar]
- 176.Dotson CD, Zhang L, Xu H. et al. Bitter taste receptors influence glucose homeostasis. PLoS One 2008;3:e3974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Clark AA, Dotson CD, Elson AET. et al. TAS2R bitter taste receptors regulate thyroid function. FASEB J 2015;29:164–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Lee RJ, Xiong G, Kofonow JM. et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J Clin Invest 2012;122:4145–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Rhie A, McCarthy SA, Fedrigo O. et al. Towards complete and error-free genome assemblies of all vertebrate species. Nature 2021;592:737–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Levey DF, Gelernter J, Polimanti R. et al. the Million Veteran Program. Reproducible genetic risk loci for anxiety: results from approximately 200,000 participants in the million veteran Program. Am J Psychiatry 2020;177:223–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Lindstrom S et al. A comprehensive survey of genetic variation in 20,691 subjects from four large cohorts. PLoS One 2017;12:e0173997. [DOI] [PMC free article] [PubMed] [Google Scholar]