

Abstract
Recent work has implicated the hypocretin (orexin) system in the genesis of narcolepsy. In the current study we demonstrate that systemically administered hypocretin-1 (Hcrt-1) produces an increase in activity level, longer waking periods, a decrease in REM sleep without change in nonREM sleep, reduced sleep fragmentation and a dose dependent reduction in cataplexy in canine narcoleptics. Repeated administration of single daily doses of Hcrt-1 led to consolidation of waking and sleep periods and to a complete loss of cataplexy for periods of three or more days after treatment in animals that were never asymptomatic under control conditions. Systemic administration of Hcrt-1 may be an effective treatment for narcolepsy.
CURRENT CLAIM: Systemic administration of Hcrt-1 may be an effective treatment for narcolepsy.
Narcoleptic patients experience cataplexy, which is a sudden loss of muscle tone most commonly in response to the sudden onset of strong emotions, excessive daytime sleepiness and fragmentation of sleep during the night. Current drug treatments can be dichotomized into those that are aimed at daytime sleepiness, typically using dopamine agonists or Modafinil (Provigil, by Cephalon, West Chester, PA), and those that are aimed at cataplexy, typically using tricyclic antidepressants (Siegel, 1999). Drug side effects, residual sleepiness and cataplexy episodes continue to be major problems for most treated narcoleptics (Aldrich, 1998).
A mutation in the gene responsible for the hypocretin-2 (orexin-2) receptor is the genetic cause of canine narcolepsy (Lin et al., 1999). A null mutation of the gene encoding the two known hypocretin (Hcrt) peptides produces aspects of the narcolepsy syndrome in mice (Chemelli et al., 1999). Human narcoleptics have reduced levels of Hcrt-1 in their cerebrospinal fluid (Nishino et al., 2000). This work suggests that administration of Hcrt might reverse symptoms of narcolepsy by compensating for either inefficient receptor transduction or diminished levels of the agonist. Basic research on the behavioral effects of the hypocretins has generally used intracerebroventricular or intra-parenchymal microinjection of the peptide (Hagan et al., 1999; Dube et al., 1999), and some studies have concluded that Herts administered systemically do not cross the blood-brain barrier (BBB) at sufficient levels to affect physiological function (Chen et al., 1999; Takahashi et al., 1999), making development of an Hcrt receptor agonist with good BBB permeability a high priority. However, a recent study suggested that non-saturable mechanisms for Hcrt-1 transport across the BBB exist, at least for iodinated Hcrt-1 (Kastin and Akerstrom, 1999). In the current study, we tested the effect of intravenous administration of Hcrt-1 on narcolepsy/cataplexy in canine narcoleptics.
METHODS
Six genetically narcoleptic Doberman pinschers (5 males and 1 female) served as subjects. Dogs were between 1.5 years and 10 years of age, with an average of 3.8 years. We analyzed the effect of systemically administered Hcrt-1 on cataplexy with a modified food-elicited cataplexy test (FECT) (Baker and Dement, 1985).
Hcrt-1 has a high affinity for both Hcrt-1 and Hcrt-2 receptors (Sakurai et al. 1998). Hcrt-1 is pharmacologically stable whereas Hcrt-2 is known to degrade rapidly (Kastin and Akerstrom, 1999). Because of this, we administered Hcrt-1. We determined the effect of Hcrt-1 administration on sleep organization using polygraph recording. We determined the effect of Hcrt-1 administration on activity levels and the duration of sleep-waking states over the 24-hour period with actigraphy.
One to 4 /ug/kg of Hcrt-1 (orexin-A, #003–30, Phoenix Pharmaceuticals, Mountain View, CA) dissolved in normal saline (100 /ug in 2 ml) was administered through the cephalic vein using a glass syringe. The glass syringe was pre-soaked in 1% BSA, rinsed in Milli-Q water, then dried at 60°C prior to use. This treatment combined with the large volume of the diluent used minimizes problems caused by the “stickiness” of the peptide. On control days, saline was administered in the same manner. Hcrt-1 or control injections were administered between 15:30 and 16:30 h.
Cataplexy Test
The FECT measures the time it takes to consume a fixed amount of food. A non-symptomatic animal eats the food in a minimal amount of time. However, a symptomatic animal has cataplectic attacks elicited by food consumption. These attacks interrupt food consumption, increasing the time taken to complete the FECT. The FECT was done by introducing a bowl of soft food (Pedigree, by Kalkan) in the home cage and counting the number of cataplectic attacks (including hind limb collapse and total cataplexies in which all four limbs collapse and the whole body contacts the floor) and total time required to eat the food (FECT time). All the FECTs began 4 min after the administration of Hcrt-1 or saline.
Sleep-Wake Study
Polygraphic Recording.
Electrodes for the assessment of sleep-wake parameters (EEG, EMG, EOG and hippocampal theta) were chronically implanted in two dogs as described earlier (Siegel et al., 1991). Polygraphic variables were recorded for 4 h after Hcrt-1 (3 /ug/kg) or saline injection.
Actigraphy.
The effects of Hcrt-1 on sleep-wake periods were monitored continuously for 24 h/day with collar mounted actigraphs (Actiwatch, Mini Mitter Inc, Sundriver, OR) while the animals remained in their home dog runs. Actigraphs were secured to a neck collar that was placed on the dogs throughout the period of study. Data were downloaded to a PC through an inductively coupled Actiwatch reader and further analyzed by a program of our design. The program could integrate total numbers of movements above a preset amplitude for a measurement of total level of activity in 5 minute epochs. For analysis of sleep state, actigraphs were first calibrated by placing them on an animal instrumented for conventional polygraphic recording. A threshold was determined for discriminating polygraphically defined waking and sleep in 30 second epochs. The durations of sleep periods were then counted and tabulated by computer. The sleep-wake bouts measured by actigraph correlated well with hypnograms obtained from polygraphic recording (r= 0.84, p<0.001).
Data Analysis
Data were analyzed with ANOVA, followed by post hoc comparisons using Newman-Keuls tests. Bonferroni Mests were done to compare the effect of Hcrt-1 on sleep stages measured with polygraphic recording. One sample Mests were performed to test the significance of number of cataplectic attacks and FECT time (expressed as a percentage of baseline) after Hcrt-1 within each dose.
RESULTS
Changes in Cataplexy After Hcrt-1 Treatment
Hcrt-1 administration had a significant effect on cataplexy in a dose dependent manner (number of cataplectic attacks, p<0.005, F=7.98, df=2, 14; FECT time, p<0.001, F=17.15, df=2, 14; ANOVA). The 1 and 2 /ug/kg doses of Hcrt-1 did not produce any change in cataplexy (Fig. 1a). The 3 /ug/kg dose produced a significant reduction in cataplexy (p<0.001, df=7; Mest) and a significant reduction in the FECT time (p<0.001, df=l\ Mest; [Fig. 1b]). The 4 /ug/kg dose of Hcrt-1 significantly increased the severity of cataplexy compared to saline control (p<0.01, df=7; t-test) and significantly increased the FECT time (p<0.05, df=7; t-test).
Figure 1. Treatment with Hcrt-1 produced short and longer-term changes in cataplexy.
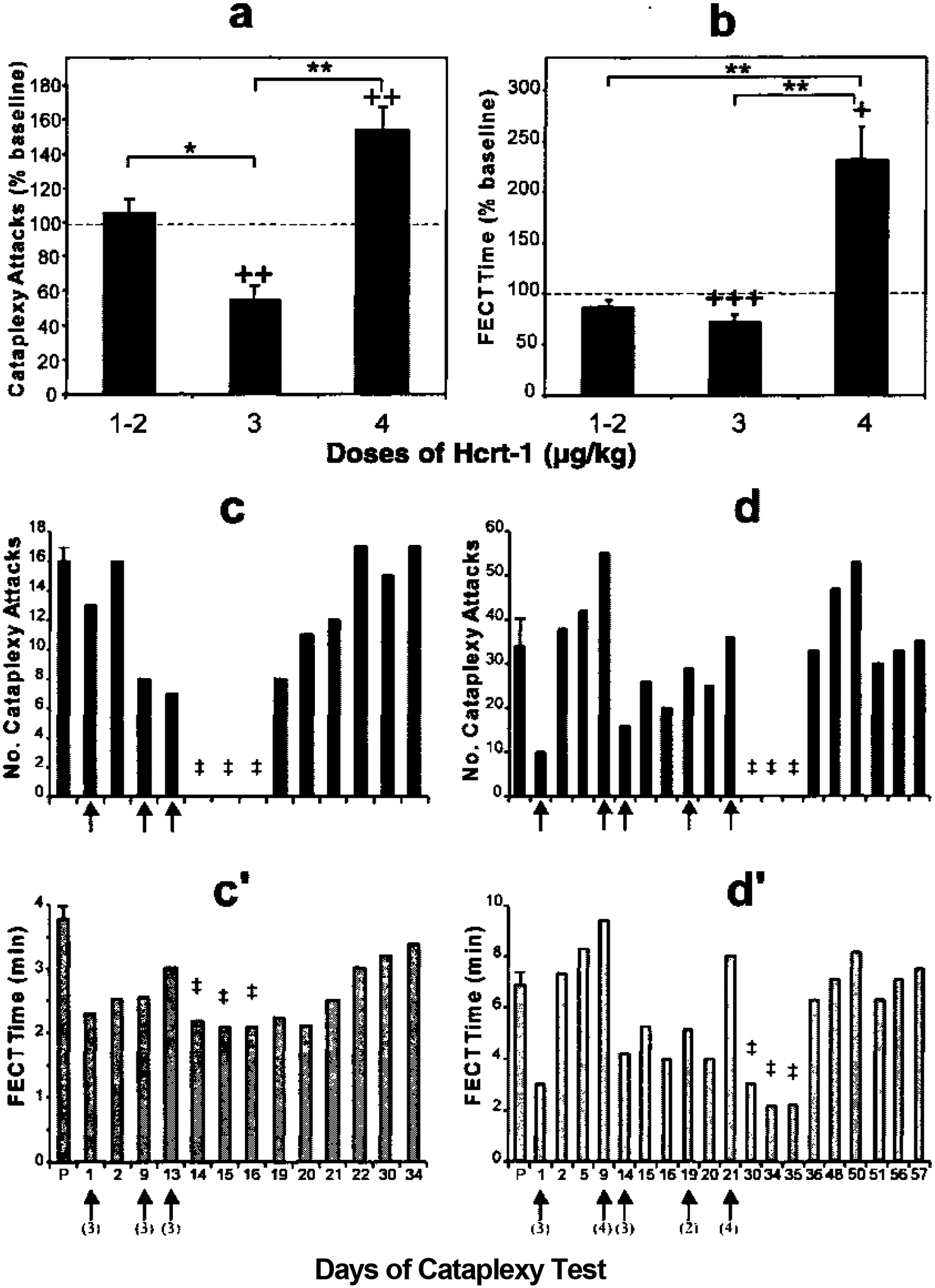
a: Intravenous injection of Hcrt-1 produced a dose dependent change in cataplexy. b: FECT time also showed a dose dependent change. Doses of 1 and 2 /ug/kg did not affect cataplexy. Doses of 3 /ug/kg produced a significant decrease in the number of falls (a) and in the time it took to consume the food (eating time + cataplexy time) (b). In contrast, 4 /ug/kg doses significantly increased the number of falls (a) and the time required to consume the food (b) relative to baseline (dotted line). c–d: In the two dogs tested with repeated doses of Hcrt-1, a complete suppression of cataplexy resulted following the last dose. c: Three doses of Hcrt-1 produced a complete suppression of cataplexy for three consecutive days. d: In a more severely affected dog, the administration of five doses of Hcrt-1 produced a complete suppression of cataplexy during the FECT observed on three tests given over a six day period. c’ and d’ show the FECT time on the days of the tests in c and d respectively. Arrows indicate days in which the FECT was done with Hcrt-1 treatment, and doses are indicated in parentheses. On all the other days animals were tested after the administration of the same volume of saline. All values in a and b are mean ±SE; P=average of pretreatment days, +p<0.05, ++p<0.01, +++p<0.001 compared to saline control, /-test; *p<0.05, **p<0.0l, between doses, Newman-Keuls test, t indicates no cataplexy attacks observed during the FECT.
Two of the three dogs treated with repeated doses of Hcrt-1 went for three or more days without any cataplexy after the administration of 3–5 doses of Hcrt-1 (Figs. 1c and 1d). A total absence of cataplexy had never been observed for even one day in any of the three dogs in 35 consecutive previous baseline tests in each animal. During the period without cataplexy the animals showed normal feeding during FECTs. The time taken to finish the food was significantly reduced due to the absence of cataplexy attacks (p<0.02, df=5; t-test). In both dogs, the severity of cataplexy gradually returned to pretreatment levels over a 3–4 day period (Figs. 1c’ and 1d’). The third of the dogs that received repeated administrations of Hcrt-1 showed a reduction but did not show a complete suppression of cataplexy.
Effect of Hcrt-1 on Sleep-Wake Periods and Activity Level
Polygraphic recording of sleep-wake parameters showed that the same dose of Hcrt-1 that induced a reduction in cataplexy produced a significant reduction in REM sleep (p<0.05, df=2; t-test) during the 4 h post-injection period as compared to saline controls (Fig. 2a).
Figure 2. Changes in sleep-wake stages after Hcrt-1 administration.
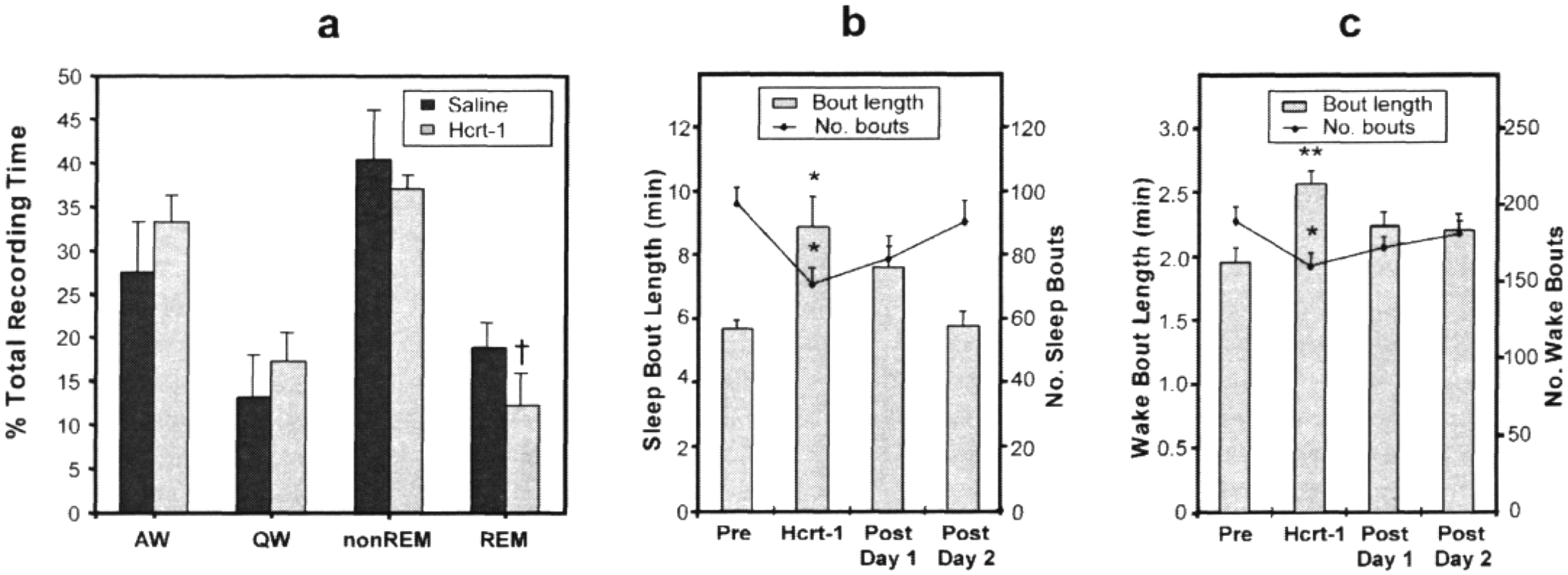
a:Polygraphic sleep-wake data for the 4 h periods after Hcrt-1 (3 /ug/kg) in comparison to the same periods after normal saline shows a significant decrease in REM sleep. b: Sleep-wake a\ctigraph data show a significant increase in sleep bout duration and a decrease in frequency during the dark period after Hcrt-1. c: Changes in wake bout duration and frequency (in 24-hour period). Pre=saline control before Hcrt-1; AW=active wake; QW=quite wake; nonREM=non-rapid eye movement sleep; REM=rapid eye movement sleep. All values are mean ±SE; tp<0.05 compared to saline control, t-test; *p<0.05, **p<0.01, compared to pre-treatment level, Newman-Keuls test.
Actigraph measurements were used to calculate the duration of sleep and waking states for the nights following injection. First, comparisons of actigraphic measurements and polygraphically recorded sleep states were made and used to determine thresholds for distinguishing sleep and wake periods. Then wake and sleep state periods were quantified starting 2 h following administration. We found that after a single dose of Hcrt-1 the mean duration of both sleep periods and wake periods increased. These effects lasted for more than 24 h (p<0.01, F=5.56, df=3, 15 and p<0.002, F=8.58, df=3, 15 for sleep and wake periods respectively). The frequency of sleep and wake bouts was reduced (p<0.05, F=3.35, df=3,15 and p<0.05, F=3.40, df=3, 15, respectively [Figs. 2b and 2c]). The total duration of sleep was increased after Hcrt-1 as compared to pre-drug levels, but not significantly (p=.055, df=5; t-test). During the periods of total cataplexy suppression following repeated Hcrt-1 doses, sleep was also consolidated (increased sleep bout length) relative to baseline conditions (p<0.05, df=5;t-test).
Hcrt-1 injection produced increased motor activity in the first 30 minutes after injection. The differences in amplitude of motor activity following Hcrt-1 and saline injection diminished over the following 60 minutes (Figs. 3a and 3b).
Figure 3. Acute changes in motor activity after Hcrt-1 administration.
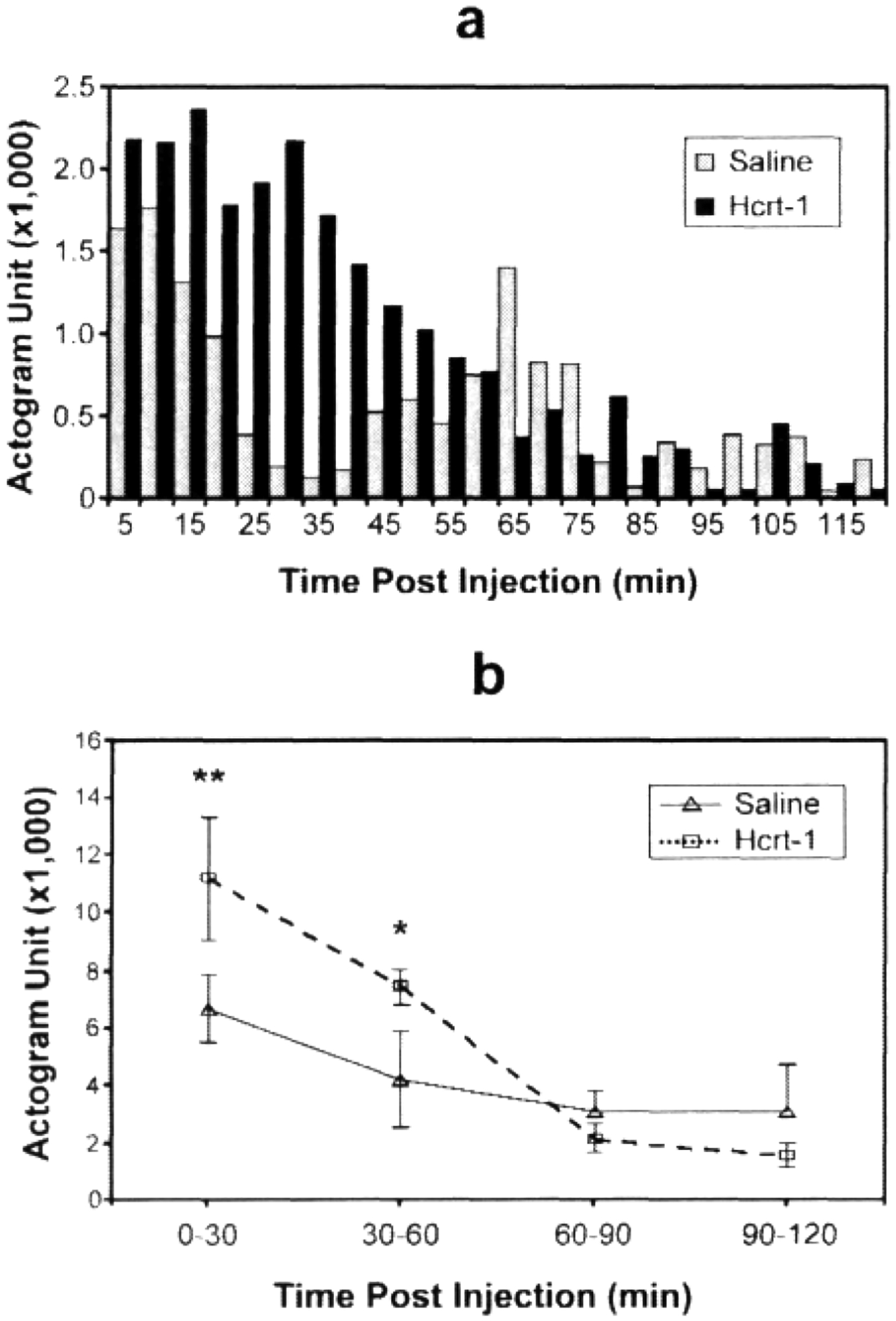
a: A representative actigraph record (5 min bins) showing the activity level of a dog during the 2 h period following Hcrt-1 (3 /ug/kg) and normal saline administration. Hcrt-1 produced an increase in motor activity within 5 minutes of injection that persisted for 60 minutes. b: The increase of motor activity was statistically significant at 0–30 and 30–60 minutes after Hcrt-1 administration (n=6), as compared to saline control (n=6). In b values are mean ±SE, *p<0.05; **p<0.01, Newman-Keuls test.
DISCUSSION
We found that Hcrt-1 administration at optimal doses can reduce or totally eliminate cataplexy for extended periods of time. We also found that high doses of Hcrt-1 produced a significant increase in cataplexy. This apparently paradoxical effect may be explained by a recent study of the role of Hcrt-1 on cataplexy, performed in our laboratory (Kiyashchenko et al., 2000). In this study, we found that microinjection of Hcrt-1 into the locus coeruleus (LC) of the rat produced an enhancement of muscle tone and blockade of experimental cataplexy. However, microinjection of the same doses of Hcrt-1 into areas ventral to LC produced an abrupt loss of muscle tone. We hypothesize that low doses of Hcrt-1 administered systemically activate monoaminergic brainstem systems (Horvath et al., 1999; Siegel, 1999; Wu et al., 1999) and other muscle tone facilitatory systems. However, higher doses act on both these facilitatory systems and on inhibitory systems (Lai and Siegel, 1988), resulting in a net suppression of muscle tone. If Hcrt-1 administration is to be used clinically, dose optimization will be critical in achieving control over cataplexy.
We saw a dramatic long-term suppression of cataplexy after repeated administrations of Hcrt-1 in two of the dogs that had never shown such a hiatus in cataplexy occurrence. During the period of suppression the dogs consumed their food in less time than in control conditions, demonstrating that the Hcrt-1 did not act by appetite suppression or by inducing illness. The dogs appeared in excellent health throughout the study. We saw no grooming, “wet dog shakes” or other abnormal behaviors that have been reported after central administration of high doses of Hcrt (Ida et al., 1999; Yamanaka et al., 1999).
A particularly striking finding was that Hcrt-1 administration caused a consolidation of both sleep and waking states. One of the cardinal signs of narcolepsy is daytime sleepiness, resulting in frequent intrusions of sleep into the waking period followed by disrupted nighttime sleep, with waking intrusion resulting in short mean sleep intervals (Mitler and Dement, 1977; Aldrich, 1992). This has been reported not only in human narcoleptics but also in canine narcoleptics (Mitler and Dement, 1977; Lucas et al., 1979). Hcrt-1 administration normalized both waking and sleep, resulting in longer waking periods, a higher level of activity and more continuous sleep periods. This linkage between reduction in cataplexy and consolidation of sleep-wake periods was seen not only on the days of Hcrt-1 administration, but also on the days of cataplexy cessation following repeated Hcrt-1 administrations.
The suppression of REM sleep seen after systemic Hcrt administration exactly mirrors the selective suppression of REM sleep seen after intracerebroventricular administration of Hcrt-1 (Hagan et al., 1999) and is further evidence for the central action of intravenously administered Hcrt-1. The suppression represents a normalization of REM sleep and cataplexy values, which are enhanced in Hcrt knockout mice (Chemelli et al., 1999) and narcoleptic dogs (Mitler and Dement, 1977; Lucas et al., 1979). The time course of changes in locomotor activity after Hcrt-1 administration approximates the time course of changes in brain concentration seen after systemic administration in one prior study (Kastin and Akerstrom, 1999). However, peripheral involvement in the effects of intravenously administered Hcrt-1 cannot be ruled out (Kirchgessner and Liu, 1999). ICV or intraparenchymal injection of Hcrt-1 has been used in most prior studies of the effects of Hcrt-1. The current study suggests that systemic administration will be a useful tool in exploring the behavioral and physiological effects of this compound.
Narcoleptic dogs are known to have a mutation in the gene that synthesizes the Hcrt-2 receptor (Lin et al., 1999). This mutation could either result in the receptor being nonfunctional or having altered function. The effectiveness of Hcrt-1 administration suggests that the receptor may be synthesized and remain responsive to its agonist, perhaps at a reduced level. However, it is more likely that the therapeutic effectiveness of Hcrt-1 administration is due to stimulation of the Hcrt-1 receptor or to activation of other as yet unidentified Hcrt receptors. We hypothesize that the longer-term reduction in symptoms, which followed repeated administrations of Hcrt-1, may be linked to downregulation of aminergic and cholinergic receptors, which are upregulated in both canine and human narcolepsy (Aldrich, 1992).
We find that, unlike most current pharmacological therapies, Hcrt-1 administration produces dramatic and correlated improvements in cataplexy, waking duration and sleep continuity. Systemic administration appears to be effective in the short and intermediate term. Further work is clearly necessary to establish the safety and efficacy of systemic administration of Hcrt-1. However, it appears to have the potential for being an effective treatment for the underlying abnormality in narcolepsy. Hcrt-1 administration may also prove useful in the treatment of other sleep disorders characterized by daytime sleepiness and interrupted nighttime sleep, such as sleep fragmentation in the elderly (Bliwise, 1994) and in other disorders of arousal.
ACKNOWLEDGMENTS
We thank Giselle B. Nguyen and Robert Nienhuis for their technical help. This work was supported by grants NS14610, HL41370 and HL60296 from the National Institutes of Health and the Medical Research Service of the Department of Veterans Affairs.
REFERENCES
- 1.Aldrich MS. Narcolepsy. Neurology 1992; 42(suppl 6): 34–43. [PubMed] [Google Scholar]
- 2.Aldrich MS. Diagnostic aspects of narcolepsy. Neurology 1998; 1950: S2–7. [DOI] [PubMed] [Google Scholar]
- 3.Baker TL, Dement WC. Canine narcolepsy-cataplexy syndrome: evidence for an inherited monoaminergic-cholinergic imbalance. In: McGinty DJ, Drucker-Colin R, Morrison A, Parmeggiani P, eds. Brain mechanisms of sleep. New York: Raven Press, 1985, pp. 199–233. [Google Scholar]
- 4.Bliwise D. Dementia. In: Kryger MH, Roth T, Dement WC, eds. The Principles and Practice of Sleep Medicine, Vol 2. Philadelphia: W.B. Saunders, 1994, pp. 790–800. [Google Scholar]
- 5.Chemelli RM, Willie JT, Sinton CM, Elmquist JK, Scammell T, Lee C, Richardson JA, Williams SC, Xiong Y, Kisanuki Y, Fitch TE, Nakazato M, Hammer RE, Saper CB, Yanagisawa M. Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell 1999; 98: 437–51. [DOI] [PubMed] [Google Scholar]
- 6.Chen C-T, Harrison TA, Dun SL, Hwang LL, Dun NJ, Chang J-K. Intracisternal administration of orexins increased blood pressure and heart rate in urethane anesthetized rats. Soc NeurosciAbst 1999; 25: 12 [Google Scholar]
- 7.Dube MG, Kalra SP, Kalra PS. Food intake elicited by central administration of orexin/hypocretins: identification of hypothalamic sites of action. Brain Res 1999; 842: 473–7. [DOI] [PubMed] [Google Scholar]
- 8.Hagan JJ, Leslie RA, Patel S, Evans ML, Wattam TA, Holmes S, Benham CD, Taylor SG, Routledge C, Hemmati P, Munton RP, Ashmeade TE, Shah AS, Hatcher JP, Hatcher PD, Jones DNC, Smith MI, Piper DC, Hunter AJ, Porter RA, Upton N. Orexin A activates locus coeruleus cell firing and increases arousal in the rat. Proc Natl Acad Sci USA 1999; 96: 10911–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Horvath TL, Peyron C, Dialno S, Ivanov A, Aston-Jones G, Kilduff TS, van den Pol AN. Hypocretin (Orexin) activation and synaptic innervation of the locus coeruleus noradrenergic system. J Comp Neurol 1999; 415: 145–59. [PubMed] [Google Scholar]
- 10.Ida T, Nakahara K, Katayama T, Murakami N, Nakazato M. Effect of lateral cerebroventricular injection of the appetite-stimulating neuropeptide, orexin and neuropeptide Y, on the various behavioral activities of rats. Brain Res 1999; 821: 526–9. [DOI] [PubMed] [Google Scholar]
- 11.Kastin AJ, Akerstrom V. Orexin A but not orexin B rapidly enters brain from blood by simple diffusion. J Pharmacol Exp Ther 1999; 289: 219–23. [PubMed] [Google Scholar]
- 12.Kirchgessner AL, Liu M. Orexin synthesis and response in the gut. Neuron 1999; 24: 941–51. [DOI] [PubMed] [Google Scholar]
- 13.Kiyashchenko LI, Mileykovskiy BY, Siegel JM. Hypocretin microinjections in the vicinity of locus coeruleus change muscle tone in decerebrate rats. Sleep 2000; 23(2): 811. [Google Scholar]
- 14.Lai YY, Siegel JM. Medullary regions mediating atonia. J Neurosci 1988; 8: 4790–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lin L, Faraco J, Li R, Kadotani H, Rogers W, Lin X-Y, Qiu X-H, de Jong PJ, Nishino S, Mignot E. The REM sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor gene. Cell 1999; 98: 365–76. [DOI] [PubMed] [Google Scholar]
- 16.Lucas EA, Foutz AS, Dement WC, Mitler MM. Sleep cycle organization in narcoleptic and normal dogs. Physiol Behav 1979; 23: 737–43. [DOI] [PubMed] [Google Scholar]
- 17.Mitler MM, Dement WC. Sleep studies on canine narcolepsy: pattern and cycle comparisons between affected and normal dogs. Electroencephalogr Clin Neurophysiol 1977; 43: 691–9. [DOI] [PubMed] [Google Scholar]
- 18.Nishino S, Ripley B, Overeem S, Lammers GJ, Mignot E. Hypocretin (orexin) deficiency in human narcolepsy. The Lancet 2000; 355: 39–40. [DOI] [PubMed] [Google Scholar]
- 19.Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, Tanaka H, Williams SC, Richardson JA, Kozlowski GP, Wilson S, Arch JR, Buckingham RE, Haynes AC, Carr SA, Annan RS, McNulty DE, Liu WS, Terrett JA, Elshourbagy NA, Bergsma DJ, Yanagisawa M. Orexin and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998; 92: 573–85 [DOI] [PubMed] [Google Scholar]
- 20.Siegel JM, Nienhuis R, Fahringer HM, Paul R, Shiromani P, Dement WC, Mignot E, Chiu C. Neuronal activity in narcolepsy: identification of cataplexy-related cells in the medial medulla. Science 1991; 252: 1315–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Siegel JM. Narcolepsy: A key role for hypocretins (Orexins). Cell 1999; 98: 409–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Takahashi N, Okumura T, Yamada H, Kohgo Y. Stimulation of gastric acid secretion by centrally administered orexin-A in conscious rats. Biochem Biophys Res Commun 1999; 254: 623–7. [DOI] [PubMed] [Google Scholar]
- 23.Wu M-F, Gulyani SA, Yau E, Mignot E, Phan B, Siegel JM. Locus coeruleus neurons: cessation of activity during cataplexy. Neuroscience 1999; 91: 1389–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yamanaka A, Sakurai T, Katsumoto T, Yanagisawa M, Goto K. Chronic intracerebroventricular administration of orexin-A to rats increases food intake in daytime, but has no effect on body weight. Brain Res 1999; 849: 248–52. [DOI] [PubMed] [Google Scholar]