

Abstract
The present study illustrates that different day intervals (DIs) between the sequential infestations of two pest species, the rice leaffolder (RLF) Cnaphalocrocis medinalis Guenée (Lepidoptera: Pyralidae) and the white-backed rice planthopper (WBPH) Sogatella furcifera Horváth (Hemiptera: Delphacidae), have a significant impact on the rice yield loss rate (YLR) and on the carbohydrate contents of rice plants. For WBPH release after RLF release (WRARR), the YLR decreased with the increasing DIs, and the YLR at the 24 DI was significantly lower compared to that at the 6 and 12 DIs and had a minimum value for a simultaneous infestation of the two pest species (SITS). In contrast, for RLF release after WBPH release (RRAWR), the YLR at the 24 DI had a maximum value and was significantly higher compared to that at the 6 and 12 DIs and the SITS. These findings indicate that damaged rice plants gradually recover, with an increase in the DI for WRARR. The above results were demonstrated by biochemical tests. Therefore, the sequential infestation of the two pest species and their DIs should be considered for integrated pest management (IPM) and control strategies for rice pests.
Keywords: Cnaphalocrocis medinalis, Sogata furcifera, sequential infestation, day interval, rice yield loss
Introduction
Rice (Oryza sativa L.) is one of the most important cultivated plants of the tropics and subtropics. Rice yield and quality are severely affected by many pests. The leaffolder insect Cnaphalocrocis medinalis (Guenée) (rice leaffolder (RLF)) is considered an important production constraint for rice and causes huge economic losses in south and southeast Asia and other rice regions of the world.1,2 C. medinalis (RLF) outbreaks have occurred in recent years in the rice-growing regions of southern China and in the middle and lower reaches of the Yangtze River. 3 The RLF larvae tie the edges of the tips of rice leaves together with silken threads, forming a protective cavity within which they feed by scraping mesophyll cells and epidermal tissue from the leaf blades. RLF infestation influences the photosynthesis rate of rice leaves. However, rice plants at the tillering stage have strong compensation mechanisms by increasing tillers; 4 the effect disappears at the later stage of rice growth. In addition, the white-backed planthopper Sogatella furcifera (Horváth) (white-backed rice planthopper (WBPH)) is widely distributed in Asia and the south Pacific regions and has acquired major economic importance because of its ability to cause leaves to dry out and tillers to wilt.5,6 Further, it is the most economically important rice pest in Asian countries, and ongoing outbreaks constitute a serious threat to the sustainability of rice production. 7 In southern China and in the middle and lower Yangtze River regions, the two pest species commonly occur simultaneously or there is a temporal sequence outbreak (WBPH infestation after RLF infestation or vice versa). However, the impacts of the temporal sequence outbreak and the day intervals (DIs)—the days from one pest infestation to another—on rice grain damage are not understood, even though rice plants at the tillering stage have a compensation mechanism for RLF infestations. Thus, investigating the effects of different DIs between the sequential infestations of the two pest species on rice yield losses has a practical significance for developing control strategies for the pests and integrated pest management (IPM).
Some studies have investigated the fact that the complex damage caused by multiple species is greater than that of any single species.8,9 Similarly, Selvaraj et al 10 established the yield loss as a function of damage to determine multiple pest economic injury levels (EILs). The EIL of a single pest species for RLF and the stem borer ranged from 2.9 to 6.4% folded leaves and 1.9 to 3.0% whiteheads, respectively, whereas that of the brown planthopper (BPH) Nilaparvata lugens (Stål) varied between 5.5 and 7.3 hoppers per hill. In addition, these joint incidence combinations indicated that although a single pest species can have a low EIL, the combination of multiple pests inflicts economic damage. 10 Therefore, we considered the idea that the impacts of temporal sequence infestations and the DIs of sequential infestations of two pest species on rice grain loss are closely associated with harmonizing chemical control for IPM. Thus, we conducted experiments to examine the effect of the temporal sequence infestation and its DIs on rice grain loss in 2011 and 2012. The objective of the present study was to determine the effects of different DIs between sequential infestations of RLF and WBPH on grain damage to provide a scientific basis for the rational application of pesticides and the reduction of pesticide use.
Materials and Methods
Rice Varieties and Insects
The Japonica rice variety, Ninjing 4, was used in these experiments. The seeds were sown into cement tanks at an experimental farm at Yangzhou University. Five leaf seedlings (25 days old) were selected from the tanks, and the soil was washed off from the plant roots with tap water. The seedlings were immediately transplanted into porcelain pots (height, 60 cm; diameter, 45 cm). Three seedlings were planted in each pot as a hill. Potted rice plants were covered with a cage, and the plants were used in the experiments at the tillering stage.
Second instar larvae, which hatched from the eggs laid by field-caught moths during the peak occurrence of C. medinalis (RLF) at the Yangzhou University experimental farm, and second instar WBPH nymphs, collected from a nursery maintained at natural conditions at an ecological laboratory (Yangzhou University), were used for these experiments.
The Experiments on Potted Rice
To examine the effects of simultaneous and sequential infestations of two pest species on rice grain loss, the experiments were designed in two parts: (1) the two pest species were simultaneously released onto rice plants at 30 days after rice transplanting (DAT) with RLF densities of 0, 2, 4, and 8 larvae per pot and WBPH nymph densities of 0, 20, 40, and 80 nymphs per pot to achieve four density combinations of 0 + 0, 2 + 20, 4 + 40, and 8 + 80 RLF larvae and WBPH nymphs, respectively; and (2) the two pest species were released in a temporal sequence, where the second pest species was released at 6, 12, and 24 DIs after the first pest species was released, which included two temporal sequences: (a) WBPH was released at 6, 12, and 24 days (ie, 36, 42, and 54 DATs) after RLF release (WRARR) at 30 DAT and (b) RLF was released at 6, 12, and 24 days after WBPH release (RRAWR) at 30 DAT The RLF larvae and WBPH nymph density combinations for the temporal sequence infestations were as follows: 2 larvae + 20 nymphs (2 + 20), 4 larvae + 40 nymphs (4 + 40), and 8 larvae + 80 nymphs (8 + 80) for WRARR; and 20 nymphs + 2 larvae (20 + 2), 40 nymphs + 4 larvae (40 + 4), and 80 nymphs + 8 larvae (80 + 8) for RRAWR. All the insects on the potted rice plants were removed after the infestations of a single generation for simultaneous and sequential infestations. All RLF pupae were manually removed from the rice plants after opening the cages. Before the laying of eggs (1–2 days after emergence), all WBPH adults were removed with a self-made sucking device. Each treatment was replicated three times for all experiments.
Determining the Components of Yield Loss
For the components of yield loss in the potted rice, the total grain weight of each potted rice hill from each treatment and a 1000 grain weight (1000 GW) from each treatment were weighed, and the blighted and filled grain rates were recorded. All yield samples were oven dried at 80°C for 48 h before being weighed. The grain loss included a decrease in the 1000 GW and an increase in the blighted grains.
Measurements of Sucrose and Starch Contents in the Rice Stems of the Potted Rice
To determine changes in the assimilates, we measured sucrose and starch contents in the plant stems at the heading stage. The sucrose and starch levels were measured using the methods described by Zhou and Li. 11
Calculation of 1000 GW, Rate of Yield Loss (RYL), and Blighted Grain Rate (BGR)
All spikes in each potted rice grain were cut down. Total grains and blighted grains were counted and weighed after drying for 2 days in an oven at 75°C. The 1000 GW and RYL were calculated as follows:
The normal distribution and homogeneity of variance (using a Bartlett test) were tested before using analysis of variance (ANOVA). Two-way ANOVA was performed twice (insect density as one way and DIs as another way or insect density as one way and sequential infestation as another way) to analyze the experimental data. Multiple comparisons of the means were conducted based on Fisher's protected least significant difference (PLSD) test. All analyses were conducted using the data processing system (DPS) of Tang and Feng. 12
Results
Effect of sequential infestations and their DIs between RLF and WBPH on the 1000 GW
For both sequential infestations, that is, WRARR and RLF release after WBPH release (RRAWR), 1000 GWs were significantly influenced by the DIs of the sequential infestations and the insect densities (Table 1). For WRARR, the multiple comparisons of the means showed that the 1000 GW was reduced with an increase in the insect density, and the weights for all infestation densities were significantly lower compared to the controls (without insect infestation) (F = 27.7, df = 3,32, P = 0.0001) (Fig. 1A). For the comparisons among DIs, the 1000 GWs for the infestations at 24 DIs were significantly lower compared to those for the 6 and 12 DIs and also lower than those for the simultaneous infestation of the two pest species (SITS), except for the 4 + 40 density (F = 135.7, df = 3.32, P = 0.0001) (Fig. 1B), indicating that the grain damage was more serious with an increase in DI. The 1000 GWs for all densities at all DIs, except for the 2 + 20 density at the 6 DI, were significantly lower compared to the control (F = 135.7, df = 3.32, P = 0.0001) (Fig. 1B). For RRAWR, multiple comparisons of means showed that the 1000 GW for all infestations, including SITS, were significantly lower compared to the control (Fig. 1C) (F = 11.5, df = 3,32, P = 0.0001). Interestingly, in contrast with WRARR, the 1000 GW at the 24 DI was higher than the 6 DI and SITS, and all DIs for the three infestation densities were significantly lower compared to the control (F = 119.9, df = 3,32, P = 0.0001) (Fig. 1D), indicating that grain damage is related to the temporal sequence of pest infestation and DI.
Table 1.
The F and P values of the two-way ANOVA for the components of the yield loss for two sequential infestations in potted rice: WBPH was released after the release of RLF (WRARR), and RLF was released after the release of WBPH (RRAWR).
| COMPONENTS OF YIELD LOSS | SOURCE OF VARIANCE | WRARR Df P-VALUE | F-VALUE | RRAWR Df P-VALUE | F-VALUE |
|---|---|---|---|---|---|
| 1000-grain weight | Day interval (A) | 3,32 | 27.7 | 3,32 | 11.5 |
| 0.0001 | 0.0001 | ||||
| Insect density (B) | 3,32 | 135.7 | 3,32 | 119.9 | |
| 0.0001 | 0.0001 | ||||
| A × B | 9,32 | 3.7 | 9,32 | 2.6 | |
| 0.003 | 0.02 | ||||
| Blighted grain | A | 3,32 | 41.8 | 3,32 | 48.9 |
| 0.0001 | 0.0001 | ||||
| B | 3,32 | 108.2 | 3,32 | 99.2 | |
| 0.0001 | 0.0001 | ||||
| A × B | 9,32 | 17.3 | 9,32 | 7.5 | |
| 0.0001 | 0.0001 | ||||
| Filled grain | A | 3,32 | 18.9 | 3,32 | 18.5 |
| 0.0001 | 0.0001 | ||||
| B | 3,32 | 76.3 | 3,32 | 113.5 | |
| 0.0001 | 0.0001 | ||||
| A × B | 9,32 | 5.1 | 9,32 | 2.6 | |
| 0.0003 | 0.002 | ||||
| Sucrose | A | 3,32 | 8.7 | 3,32 | 105.1 |
| 0.0002 | 0.0001 | ||||
| B | 3,32 | 206.4 | 3,32 | 365.0 | |
| 0.0001 | 0.0001 | ||||
| A × B | 9,32 | 3.2 | 9,32 | 16.7 | |
| 0.007 | 0.0001 | ||||
| Starch | A | 3,32 | 27.3 | 3,32 | 3.7 |
| 0.0001 | 0.02 | ||||
| B | 3,32 | 490.3 | 3,32 | 86.1 | |
| 0.0001 | 0.0001 | ||||
| A × B | 9,32 | 7.6 | 9,32 | 2.6 | |
| 0.00017 | 0.02 |
Figure 1.
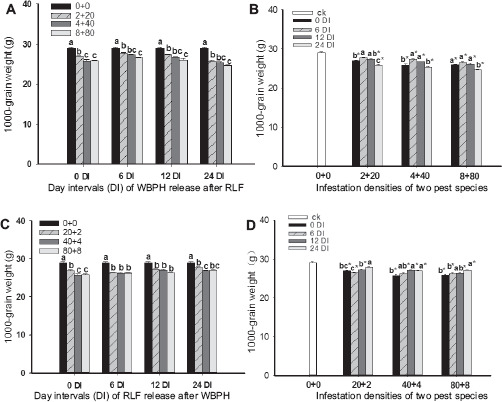
Effects of sequential infestation densities and the different DIs on 1000 GW. (A) The sequential infestation densities of WRARR, (B) different infestation densities for WRARR, (C) the sequential infestation densities of RRAWR, and (D) different infestation densities for RRAWR. The results are expressed as mean ± SE. Bars with different letters within the same DI (A or C) and within the same infestation density (B or D) represent significant differences at the 5% level. The “*” symbol represents a significant difference between the DI and control (0 + 0) (without insect infestation) at the 5% level.
Effect of Sequential Infestations and their DIs between RLF and WBPH on the BGR
The BGR was significantly influenced by the DIs of sequential infestations and insect densities (Table 1). For WRARR, multiple comparisons of means showed that the BGRs significantly increased with increases in the DI and insect density (Fig. 2A) (F = 41.8, df = 3.32, P = 0.0001). For comparisons among DIs, BRGs at the 24 DI for the 4 + 40 and 8 + 80 densities were significantly higher compared to the 0, 6, and 12 DI and the control; however, no significant differences between the 6 and 12 DIs were found (Fig. 2B) (F = 108.2, df = 3.32, P = 0.0001). For RRAWR, multiple comparisons of means showed that the BGRs significantly increased with the increasing insect density, except for the 20 + 2 density at the 12 and 24 DIs (F = 48.9, df = 3.32, P = 0.0001) (Fig. 2C). However, on comparisons among DIs, the BGRs reduced with an increase in the DI (F = 99.2, df = 3.32, P = 0.0001) (Fig. 2D), and those at SITS and 6 DI were significantly higher compared to the 12 and 24 DIs and control, with the maximum value observed at SITS.
Figure 2.
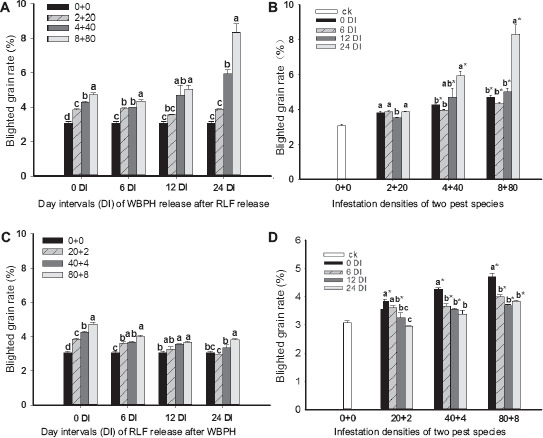
Effects of sequential infestation densities and the different DIs on BGR (%). (A) The sequential infestation densities of WRARR, (B) different infestation densities for WRARR, (C) the sequential infestation densities of RRAWR, and (D) different infestation densities for RRAWR. The results are expressed as mean ± SE. Bars with different letters within the same DI (A or C) and within the same infestation density (B or D) represent significant differences at the 5% level. The “*” symbol represents a significant difference between the DI and control (0 + 0) (without insect infestation) at the 5% level.
Effect of sequential Infestations and their DIs between RLF and WBPH on the NFG
The NFG was significantly influenced by the DIs of sequential infestations and insect densities (Table 1). The NFGs decreased with an increase in the infestation density for WRARR (F = 18.9, df = 3.32, P = 0.0001) (Fig. 3A). For comparisons among the DIs, the NFGs at the 12 and 24 DIs for the 4 + 40 and 8 + 80 densities and at the SITS and 6 DI for the 2 + 20 density were significantly lower compared to the control (F = 76.3, df = 3.32, P = 0.0001) (Fig. 3B). For RRAWR, NFGs significantly decreased with increases in the infestation density and DIs (Figs. 3C and D). For comparisons among the DIs, the NFGs at the 24 DI was significantly lower compared to the SITS and 6 DI, and the NFGs at all DIs for the three densities were significantly lower compared to the control (F = 113.5, df = 3.32, P = 0.0001) (Fig. 3D).
Figure 3.
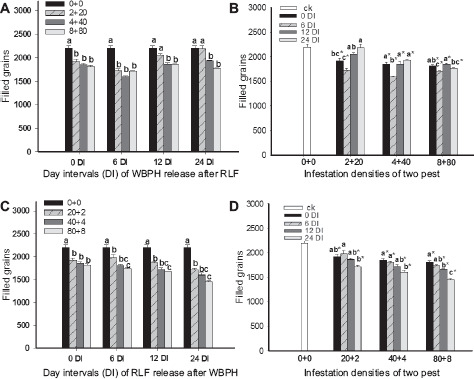
Effects of sequential infestation densities and the different DIs on filled grain numbers. (A) The sequential infestation densities of WRARR, (B) different infestation densities for WRARR, (C) the sequential infestation densities of RRAWR, and (D) different infestation densities for RRAWR. The results are expressed as mean ± SE. Bars with different letters within the same DI (A or C) and within the same infestation density (B or D) represent significant differences at the 5% level. The “*” symbol represents a significant difference between the DI and control (0 + 0) (without insect infestation) at the 5% level.
Effect of Sequential Infestations and Their DIs between RLF and WBPH on the Yield Loss
The yield loss rates (YLRs) were significantly influenced by the DIs of sequential infestations and insect densities (Table 1). For WRARR, YRL increased with the increasing insect density (F = 76.3, df = 3.32, P = 0.0001) (Fig. 4A). For comparisons among the DIs, YRLs at the 6, 12, and 24 DIs were significantly higher compared to the SITS (F = 18.9, df = 3.32, P = 0.0001) (Fig. 4B). For RRAWR, YRLs increased with the increasing insect density (Fig. 4C) (F = 113.5, df = 3.32, P = 0.0001). For comparisons among the DIs, YRLs at the 24 DI were significantly higher compared to the SITS, and 6 and 12 DIs (F = 18.5, df = 3.32, P = 0.0001) (Fig. 4D).
Figure 4.
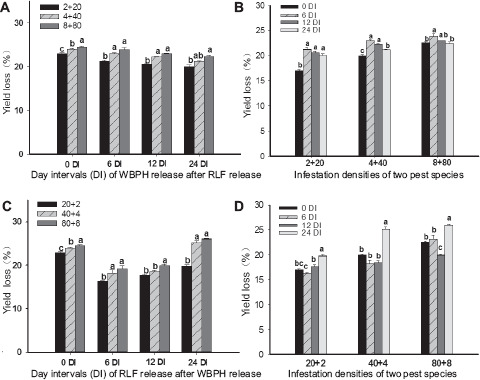
Effects of sequential infestation densities and the different DIs on yield loss (%). (A) The sequential infestation densities of WRARR, (B) different infestation densities for WRARR, (C) the sequential infestation densities of RRAWR, and (D) different infestation densities for RRAWR. The results are expressed as means ± SE. Bars with different letters within the same DI (A or C) and within the same infestation density (B or D) represent significant differences at the 5% level.
Changes in the Sucrose and Starch Contents
The sucrose and starch contents at the heading stage were significantly influenced by the DIs of sequential infestations and insect densities (Table 1). For WRARR, the sucrose and starch contents in the rice plants decreased with the increasing insect density (Figs. 5A and B), and these contents at all DIs for the three densities were significantly lower compared to the control (Figs. 5C and D). For comparisons among the DIs, the sucrose contents at the 12 and 24 DIs for the 2 + 20 density were significantly lower compared to the SITS and 6 DI, and those at the 12 and 24 DIs for the 4 + 40 density were significantly lower compared to the SITS. For the 8 + 80 density, we did not observe a regularity among the DIs (Fig. 5C). However, the starch contents among the DIs increased with the increasing DI. For RRAWR, the sucrose and starch contents in the rice plants decreased with the increasing insect density (Figs. 6A and B). On comparison among the DIs, the sucrose contents at the 6, 12, and 24 DIs for the 20 + 2 and 40 + 4 densities and at the 6 and 12 DIs for the 80 + 8 density were significantly higher compared to the SITS but significantly lower compared to the control, except at the 6 DI for the 20 + 2 and 40 + 4 densities (Fig. 6C). However, the starch contents for all of the DIs and densities were significantly lower compared to the control (Fig. 6D). The starch content at the 24 DI was significantly lower compared to the SITS for the 20 + 2 density, and the other densities showed fewer changes among the DIs.
Figure 5.
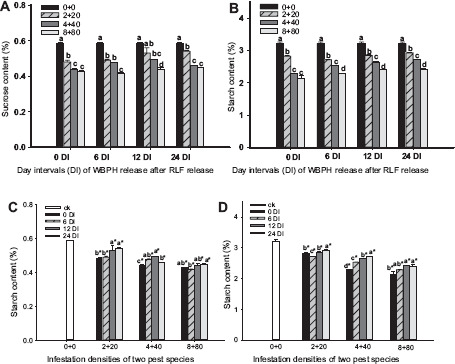
Effects of the sequential infestation densities of WRARR on the sucrose and starch contents in the rice plants under different DIs. (A) sucrose content, (B) starch content, (C) comparisons of the effects of the DIs on sucrose contents, and (D) comparisons of the effects of the DIs on starch contents. The results are expressed as mean ± SE. Bars with different letters within the same DI (A or B) and within the same infestation density (C or D) represent significant differences at the 5% level. The “*” symbol represents a significant difference between the DI and control (0 + 0) (without insect infestation) at the 5% level.
Figure 6.
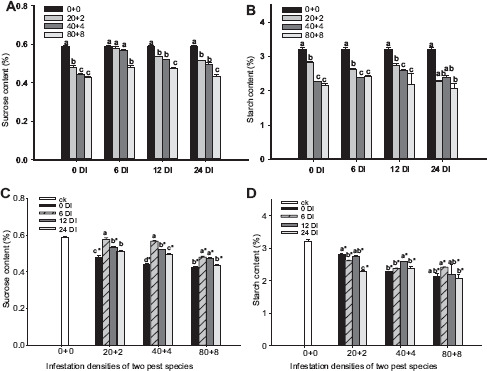
Effects of sequential infestation densities of RRAWR on the sucrose and starch contents in the rice plants under different DIs. (A) sucrose content, (B) starch content, (C) comparisons of the effects of the DIs on the sucrose contents, and (D) comparisons of the effects of the DIs on the starch contents. The results are expressed as mean ± SE. Bars with different letters within the same DI (A or B) and within the same infestation density (C or D) represent significant differences at the 5% level. The “*” symbol represents a significant difference between the DI and control (0 + 0) (without insect infestation) at the 5% level.
Discussion
It has previously been demonstrated that at early stages (approximately the tillering stage), rice plants have a strong compensation capacity against RLF infestations,4,38 and insecticides do not need to be applied at this stage.13,14 However, the effects of pest infestations at the middle or later stages of rice growth, or with different DIs between sequential infestations of two pest species on the yield loss after an early infestation of RLF are still not well understood. Understanding the effect of different DIs between sequential infestations on the yield loss is important for the balance between reducing insecticide applications and the maintenance of a high output. Many studies have indicated that multispecies EILs are complex and different from the sum of single species EILs.8,10,15,16 For example, WBPH and C. medinalis (RLF) infestations at the early stage of rice growth aggravate the damage of N. lugens infestations at the later stage. 8 Complex infestations or sequential infestations at lower densities of each species still result in significant yield losses even though the infestation density of any single species is below the EIL, 17 which may be due to the infestation of one species elevating the damage incidence of another species. Individual pests may be below their respective EILs; however, in combination, the pests may jointly inflict economic loss.15,18 An experiment examining marked rice plants in a field demonstrated that the yield loss of the complex infestation of the rice sheath blight (Pellicularia sasakii) and Chilo suppressalis Walker was higher compared to an infestation by either of the single pest species. 19 There are three possible outcomes of combined pest infestations on crop yield: (1) no interaction, (2) higher than additive (synergistic) interactions, and (3) lesser than additive (antagonistic) interactions. 20 According to our previous studies and experience with natural populations, the accurate determination of the EIL of combined pest infestations is quite complicated. Therefore, the qualitative determination of the effect of different DIs between sequential infestations on rice injury or yield loss may be more practical for developing strategies for IPM and the control of rice pests. The present findings indicate that there were significant differences in the effects of the sequential infestations of the two pest species and the DIs between infestations on the rice yield loss rate (YLR). For WRARR, the YLR decreased with the increasing DI. In contrast, for RRAWR, the YLR at the 24 DI had a maximum value and was significantly higher compared to that at the 6 and 12 DIs and the SITS. These findings indicate that the damaged rice plants gradually recovered with the increasing DI for WRARR. However, the plants that were previously damaged by WBPH were more seriously injured by RLF at the later stage of rice growth for RRAWR because we started to release insects at the tillering stage; that is, WBPH release at this stage can limit organ formation and differentiation, which results in the late biomass accumulation; 21 can cause decreases in rice tillers, and can result in a reduction in effective panicles. 22 However, rice can compensate for RLF infestation at this stage, indicating that the impact is not obvious on physiological and morphological indicators, even showing over-compensation for some insects. 23 In other words, RRAWR resulted in a higher yield loss compared to that of WRARR and SITS, especially at the 24 DI.
In addition, many studies have demonstrated that sucrose is one of the main products of plant photosynthesis. It is also the main component of rice plants, which can transport their carbon source over long distances when the plants are under adverse conditions, such as drought and low temperatures. 24 This carbohydrate can protect and stabilize cell protein structures by regulating osmosis to prevent enzyme inactivation. Many studies have indicated that plants can effectively counteract adverse stresses.25–27 There are many hypotheses regarding the relationship between the reducing sugar (sucrose) content of rice and BPH resistance; however, most studies suggest that sugar has a negative effect.28,29 There are a few studies regarding the sucrose content of rice after complex infestations. Our previous studies have demonstrated that the sucrose content of rice stems decreases with an increase in the infestation density at the booting stage, indicating that when rice is injured by pests, it will take the initiative to improve its resistance to reduce damage. Therefore, we hypothesize that rice plants have a strong compensation capacity for multispecies infestations in WRARR but not in RRAWR, and hence the rice plant could start to defend more against RLF than WBPH at the early stage.
Starch is the main stored photosynthate, and the starch content in the rice grain determines the final yield. The conversion from sucrose to starch is dynamic, and many studies have shown that the starch content in plants decreases under stress conditions.30–33 The present results show that the starch content in rice stems decreases significantly with an increase in the infestation density at the booting stage. The sucrose and starch contents decreased with increases in the insect density and DI for RRAWR, which may be because RLF infestations at the later stage influenced the leaf photosynthesis rate and the filling and accumulation of grain carbohydrates, resulting in reductions in grain yields. In contrast, the photosynthesis rate of the leaf remains normal for WRARR, even though WBPH removed a given amount of carbohydrates due to feeding. The effects of the sequential infestations of the multiple species on yield loss may vary with the combination of pests. The effect of infestations of the BPH N. lugens (Stål) at a later stage, after a RLF infestation at the early stage, of rice growth on yield loss may be different from that of WRARR because BPH infestations are more serious than WBPH infestations.8,34,35 Based on the present findings, we suggest that the control of RLF at the later stage of rice growth should be strictly managed, especially under the condition of WBPH infestation at the early stage. Therefore, the sequential infestation of pests and their DIs should be considered for IPM and control strategies for rice pests.
Most plants can compensate for herbivory to some extent, which depends on endogenous and exogenous factors. 39 Some plants may fully compensate for insect damage, whereas others may overcompensate and increase their yields or fitness. 36 The compensation effect can be generated at the level of vegetative organs and reproductive organs or at the physiological and biochemical levels. 37 The present study has demonstrated that the activities of sucrose phosphate synthase (SPS) and sucrose synthase (SS) in rice stems increased with an increase in the DI for RRAWR but not for WRARR (unpublished data), indicating that the rice plant can compensate for the damage caused by RLF infestations at the early stage of rice growth. In addition, the effect of the sequential infestations and the DIs of the natural populations of the two pest species on YLR in the rice yields needs to be further investigated. The relationship between the compensation capacity of the rice plants at an early stage of growth and the pest infestations or DIs of sequential infestations will be demonstrated in the future.
Conclusions
In summary, the present study demonstrated that the effect of different DIs between the sequential infestation of two pest species on the rice yield loss rate (YLR) and on the carbohydrate content of rice plants was significantly different. The damaged rice plants gradually recovered with an increase in the DI for WRARR. However, plants previously damaged by WBPH were more seriously injured by RLF at the later stage of rice growth during RRAWR. Therefore, the sequential infestation of pests and their DIs should be considered for IPM and control strategies for rice pests.
Author Contributions
Conceived and designed the experiments: L-BJ. Analyzed the data: L-BJ. Wrote the first draft of the manuscript: L-BJ. Contributed to the writing of the manuscript: JC, Z-FZ, L-QG. Agree with manuscript results and conclusions: G-QY, J-CW. Jointly developed the structure and arguments for the paper: JC, Z-FZ, L-QG. Made critical revisions and approved final version: J-CW. All authors reviewed and approved of the final manuscript.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests.
References
- 1.Dale D. Insect pests of the rice plants: their biology and ecology. In: Biology and Management of Rice Pests (Ed. By Heinrichs E.A.). Wiley Eastern Limited, New Delhi. 1994; 440–442. [Google Scholar]
- 2.Sardesai N., Rajyashri K.R., Behura S.K. et al. Genetic, physiological and molecular interactions of rice and its major dipteran pest, gall midgee. Plant Cell Tiss Org Cult. 2001; 64: 115–131. [Google Scholar]
- 3.Yang R.M., Zhu Y.J., Diao C.Y. et al. Causes of the outbreaks of Cnaphalocrocis medinalis in Jiangsu province in 2003 and its control strategies. China Plant Prot. 2004; 2: 11–14. [Google Scholar]
- 4.Jin D.R. Ability of compensation of rice to the larval injury by the rice leaffolder, Cnaphalocrocis medinalis (Guenée). Acta Phytophylacica Sin. 1984; 11: 1–7. [Google Scholar]
- 5.Qi G.F., Zhang X.F., Zhao X.Y. Endophytic Bacillus subtilis WH2 containing Pinellia ternate agglutinin showed insecticidal activity against whitebacked planthopper Sogatella furcifera. Bio Control. 2013; 58: 233–246. [Google Scholar]
- 6.Watanabe T., Kitagawa H. Photosynthesis and translocation of assimilates in rice plants following phloem feeding by the planthopper Nilaparvata lugens (Stål) (Homoptera: Delphacidae). J Econ Entomol. 2000; 93: 1192–1198. [DOI] [PubMed] [Google Scholar]
- 7.Catindig J.L.A., Arida G.S., Baehaki S.E. et al. Situation of planthoppers in Asia. In: Heong K.L., Hardy B., eds. Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia. Los Baños: International Rice Research Institute; 2009: 191–220. [Google Scholar]
- 8.Wu J.C., Liang J.R., Zhang F.J. et al. Studies on combined infestations of three insect pests and their effects on grain growth of rice. Acta Entomol Sin. 1995; 38: 30–38. [Google Scholar]
- 9.Qian S.M., Gao C.G., Fu G.Y. Study on the loss and control index of complex damage with rice stem borer and leafroller. Plant Prot Tech Ext. 2000; 20(4): 11–12. [Google Scholar]
- 10.Selvaraj K., Chander S., Sujithra M. Determination of multiple-species economic injury levels for rice insect pests. Crop Prot. 2012; 32: 150–160. [Google Scholar]
- 11.Zhou Z.F., Li Z.A. The guidance of plant physiology experiments. Beijing, China: Higher Education Press; 2005. [Google Scholar]
- 12.Tang Q.Y., Feng M.G. DPS data processing system for practical statistics. Beijing, China: Scientific Press; 2002. [Google Scholar]
- 13.Hu G.W. The principle and practice of reducing chemical application in paddy field. Entomol Knowl. 1996; 33: 3–7. [Google Scholar]
- 14.Hu G.W. The principle and practice of reducing chemical application in paddy field. Entomol Knowl. 1996; 33: 65–69. [Google Scholar]
- 15.Palis F.G., Pingali P.L., Litsinger J.A. A multi-pest economic threshold for rice production (A case study from the Philippines). In Teng P. (ed.), Crop Loss Assessment in Rice International Rice Institute: Los Banos, Laguna, Philippines. 1990. (pp. 229–242). [Google Scholar]
- 16.Zhang W.Q., Gu D.X., Pu Z.L. Studies on the multiple species injury loss of three major rice insect pests. Entomol Knowl. 1994; 31(6): 321–323. [Google Scholar]
- 17.Waibel H. The economics of integrated pest control in irrigated. Crop Protection Monograph. Springer-Verlag: Berlin; 1986. [Google Scholar]
- 18.Huang X., Wang X.Q. Analysis on control index of Sogatella furcifera and Cnaphalocrocis medinalis Güenée at the same time. Mod Agric Sci Tech. 2013; 6: 110–111. [Google Scholar]
- 19.He Z.Q., He S.L., He M. Research on mixture injuries by both sheath blight (Pellicularia sasakii) and stem borer (Chilo suppressalis) and their multiple yield losses on rice. Southwest China J Agric Sci. 1991; 4: 37–42. [Google Scholar]
- 20.Lamp W.O., Yeargan K.V., Nonis R.F. et al. Multiple pest interactions in alfalfa. In: Frisbie R.E., Adkinsson P.L., eds. CIPM-Integrated Pest Management on Major Agricultural Systems. Vol. 1616. Texas: Texas Agricultural Experiment Station Miscellaneous Publication, College Station; 1985: 345–364. [Google Scholar]
- 21.Hao S.G., Luo Y.J. On the yield losses of rice damaged by brown planthopper in different stages. Acta Phytophylacica Sin. 1997; 24(4): 321–325. [Google Scholar]
- 22.Liang J.R., Chen X.L. Studies on the path analysis of the white-backed planthopper damage to rice yield loss. Jiangsu Agric Sci. 1990; (5): 36–38. [Google Scholar]
- 23.Hu G.W., Guo Y.J., Shao S. Management of rice leaffolder in China. Beijing: China Agricultural Science and Technology Press; 1993. [Google Scholar]
- 24.Li H.S. The principles and techniques of plant physiological and biochemical experiment. Higher Educ Press. 2000; 194–195. [Google Scholar]
- 25.Singh K.B. Transcriptional regulation in plants: the importance of combinatorial control. Plant Physiol. 1998; 118: 1111–1120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sinha A.K., Hofmann M.G. Metabolizable and non-metabolizable sugars activate different signal transduction pathways in tomato. Plant Physiol. 2002; 128: 1480–1489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Oliver S.N., Dennis E.S., Dolferus R. ABA regulates apoplastic sugar transport and is a potential signal for cold-induced pollen sterility in rice. Plant Cell Physiol. 2007; 48: 1319–1330. [DOI] [PubMed] [Google Scholar]
- 28.Li R.Z., Ding J.H., Hu G.W. et al. Brown planthopper and its management. Shanghai: Fudan Press; 1996: 74–76. [Google Scholar]
- 29.Wu J.C., Xu J.F.X., Feng M. et al. Impacts of pesticides on physiology and biochemistry of rice. Sci Agric Sin. 2003; 3: 536–541. [Google Scholar]
- 30.Rijven AHGC. Heat inactivation of starch synthase in wheat endosperm tissue. Plant Physiol. 1986; 81: 448–453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Keeling P.L., Bacon P.J., Holt D.C. Elevated temperature reduces starch deposition in wheat endosperm by reducing the activity of soluble starch synthase. Planta. 1993; 191: 342–348. [Google Scholar]
- 32.Umemoto T., Nakamura Y.M., Ishikura N. Activity of starch synthase and the amylose content in rice endosperm. Phytochemistry. 1995; 40: 1613–1616. [Google Scholar]
- 33.Wang W., Cai Y.X., Cai K.Z. et al. Regulation of soil water deficits on stem-stored carbohydrate remobilization to grains of rice. Acta Phytoecol Sin. 2005; 29(5): 819–828. [Google Scholar]
- 34.Huang F.N., Cheng X.N. Studies on the damage of brown planthopper to yield components of rice. Chin J Rice Sci. 1990; 4: 117–121. [Google Scholar]
- 35.Qin H.G., Ye Z.X., Huang R.H. et al. Studies on the complex control index of Sogatella furcifera and Nilaparvata lugens at the spike stage of late rice. Sci Agric Sin. 1993; 26: 51–55. [Google Scholar]
- 36.Sheng C.F. Growth tediousness as an explanation of over-compensation of crops for insect feeding. Chin J Appl Ecol. 1990; 1: 26–30. [Google Scholar]
- 37.Hu Z.Y. Compensatory effects of pest plants. J Wenshan Teach Coll. 2007; 20: 106–109. [Google Scholar]
- 38.Fabellar L.T., Faballer N.G., Heong K.L. Simulating rice leaffolder feeding effects on yield using MACROS. Int Rice Res Notes. 1994; 19: 7–8. [Google Scholar]
- 39.Julien M.H., Bourne A.S. Compensatory branching and changes in nitrogen content in the aquatic weed Salvinia molesta in response to disbudding. Oecologia. 1986; 70: 250–257. [DOI] [PubMed] [Google Scholar]