

Abstract
Cataplexy, a symptom of narcolepsy, is a loss of muscle tone usually triggered by sudden, emotionally significant stimuli. We now report that locus coeruleus neurons cease discharge throughout cataplexy periods in canine narcoleptics. Locus coeruleus discharge rates during cataplexy were as low as or lower than those seen during rapid-eye-movement sleep. Prazosin, an α1 antagonist, and physostigmine, a cholinesterase inhibitor, both of which precipitate cataplexy, decreased locus coeruleus discharge rate.
Our results are consistent with the hypothesis that locus coeruleus activity contributes to the maintenance of muscle tone in waking, and that reduction in locus coeruleus discharge plays a role in the loss of muscle tone in cataplexy and rapid-eye-movement sleep. Our results also show that the complete cessation of locus coeruleus activity is not sufficient to trigger rapid-eye-movement sleep in narcoleptics.
Keywords: locus coeruleus, narcolepsy, cataplexy, neuronal activity, sleep, canine
Narcolepsy, hypothesized to be a disorder of the rapid-eye-movement (REM) sleep mechanism47,54 is characterized by a cluster of distinct symptoms, including hypnagogic hallucinations, sleepiness, sleep paralysis and cataplexy. Cataplexy is a complete loss of skeletal muscle tone that occurs when the narcoleptic is awake and alert. It may be triggered by athletic activity, laughter, anger or other emotions of sudden onset, or may occur without precipitating acts or emotions.2,22,23 The electroencephalogram (EEG) is activated during cataplexy periods, as in waking and REM sleep.2,22,23,53 The only time a loss of muscle tone comparable to cataplexy occurs in individuals without narcolepsy, is during REM sleep. Cataplexy allows us to separate the neuronal systems linked to muscle atonia from those linked to other aspects of REM sleep, including its dream mentation and absence of awareness of the environment.
The locus coeruleus (LC) provides noradrenergic input to the forebrain, cerebellum, brainstem and spinal cord.43,48 Research in monkeys, cats and rats has revealed that LC neurons are tonically active during both quiet and active waking.5,18,26,29 They phasically increase discharge during arousal, in response to stressors1,3 and with sensory stimulation.5,18,46 It has been hypothesized that the tonic activity of LC neurons facilitates information processing throughout the brain,6,45,51 and that it is required for attention to significant environmental stimuli,5–7,14,19,45 learning4,12,25,33 and the activation of immediate early genes in several brain regions in waking.11 LC neurons reduce their activity during non-REM sleep and cease discharge for an extended period only in REM sleep. It has been thought that the cessation of activity in LC and other aminergic cells is a key part of the mechanism triggering this state.6,26,29–31,45 In the current study we have recorded LC neurons during normal sleep states and during cataplexy in narcoleptic dogs to determine whether changes in LC activities are linked to cataplexy.
EXPERIMENTAL PROCEDURES
Surgery
LC cells were recorded from the dorsal pons of four narcoleptic Doberman Pinschers using microwire recording techniques as described previously.53 Under methoxyflurane or isoflurane anesthesia, microdrives containing 32 and 64 μm stainless steel microwires (California Fine Wire Co., Grover Beach, CA, U.S.A.) aimed at the LC area were implanted, with the tip of the electrodes 1 mm above the target area. The frontal sinus was opened and stainless steel screw electrodes were placed over sensorimotor cortex for EEG and into the posterior orbit for electrooculogram (EOG) recording. For the recording of hippocampal theta activity, tripolar twisted electrodes, made of three 256 μm Formvar-insulated stainless steel wires (California Fine Wire Co.), were placed in the dorsal hippocampus. Electromyogram (EMG) activity was recorded from the dorsal neck muscles with Teflon-coated multistranded stainless steel wires (Cooner Wire, Chatsworth, CA, U.S.A.).
Unit and physiological recording in sleep and cataplexy
The dogs were free to move around the chamber (1.2 × 1.2 × 1.2 m in size) during the recordings. Electrodes were moved in 80 μm steps until a cell or cells with signal-to-ratio of at least 2:1 were isolated. The activity of each cell was then characterized throughout sleep/waking states and during cataplexy. Unit pulses from the window discriminator along with the digitized action potentials, EEG, EOG, EMG and hippocampal activity were recorded continuously on a PC using Spike2 software (Cambridge Electronic Design, Cambridge, U.K.). Microwire signals were filtered, with roll off of signals below 300 Hz and above 10 kHz, and digitized at a frequency of 25 kHz. Cortical and hippocampal EEG, and EOG signals were filtered between 1 and 100 Hz and sampled at a rate of 200 Hz. EMG was filtered between 30 Hz and 3 kHz and sampled at 1 kHz. Polygraph records of the unit pulses and all the physiological recordings were taken simultaneously.
Baseline sleep/wake discharge profile for each cell was established across at least two complete sleep/wake cycles. Cataplexy was then induced either by the introduction of soft food (Pedigree, Vernon, CA, U.S.A.) or by giving novel toys or play objects to the dog. A minimum of five cataplexies (mean of >10 cataplexies) were observed during the recording of each cell. In some cases, the anti-cholinersterase physostigmine salicylate (50–100 μg/kg, i.v.; Forest Pharmaceuticals, St Louis, MO, U.S.A.) and/or the α1 antagonist prazosin hydrochloride (0.5 mg/kg, p.o.; UDL Laboratories, Rockford, IL, U.S.A.) was administered to increase the frequency and the intensity of cataplexy. Cells were held for recording for as long as needed to complete the experimental protocol, from between 6.5 h up to seven days, with an average of over four and a half days. Physostigmine administered i.v. produces increased cataplexy within 5 min of administration, while oral prazosin administration has its maximal effect after 1.5 h. Narcoleptic dogs display partial cataplexies in which only the hindlimbs collapse and complete cataplexies in which all four limbs collapse and the whole body contacts the floor. Partial cataplexies typically last less than 5 s while complete cataplexies may last from 10 s to a few minutes. Because of the short duration of the partial cataplexy episodes, only complete cataplexies were used for quantification of the change in unit discharge rate during this state. The effect of prazosin on cell activity was assessed between 1.5 to 2 h after drug administration during a quiet waking state. The effect of physostigmine was assessed during waking, 5–10 min after injection.
Cells were classified as noradrenergic based on the criteria reported for LC cells in the rat, cat and monkey.1,5,18 These criteria include: (i) slow (1–2 Hz) and regular firing in quiet waking, (ii) greatly reduced firing in REM sleep, (iii) suppression of firing by systemic administration of α2 agonist clonidine, (iv) long duration (>2 ms) action potentials, and (v) histological localization to the pontine region containing tyrosine hydroxylase (TH)-positive cells.
Clonidine hydrochloride was given either intravenously (Sigma, St Louis, MO, U.S.A.; 2 μg/kg, dissolved in saline) or orally in tablet form (UDL Laboratories; 0.2 mg/kg). The effect of clonidine on unit discharge was assessed 10–30 min after the intravenous injection or 1 h after oral administration during a quiet waking state. Spike duration was determined using the averaged waveform taken over 200 sweeps. To characterize the temporal relationship between unit and EMG activity, both unit and EMG activity were averaged around the onset and offset of cataplexy. The time of cataplexy onset and offset was determined from the digitized EMG. The onset of cataplexy was defined as the point at the beginning of a period in which the EMG became completely atonic for at least half a second. The offset of cataplexy was defined as the point where EMG activity resumed. Unit activity and EMG were averaged 20 s before and after the onset/offset points. A paired t-test was used to compare baseline and post-drug firing rates with clonidine, physostigmine and prazosin administration. To prevent drug interactions or contamination of normal sleep cycle observations we allowed a 3-h recovery from physostigmine and clonidine and a 12-h recovery from prazosin before any further observations.
Histology
At the completion of recording, iron was deposited by passing a 15-μA, 20-s anodal current through the electrodes from which cells had been recorded. Dogs were killed by first deeply anesthetizing them with sodium pentobarbital (50 mg/kg, i.v.), then perfusing transcardiacly with buffered saline and paraformaldehyde.
Sixty-micrometer brain sections were cut and processed for TH. Sections were rinsed three times in 0.1 M Trizma-buffered saline followed by a 48-h incubation in a 1/500 dilution of primary rabbit antiserum to TH (Eugene Tech International, Ridgefield Park, NJ, U.S.A.) at 4°C. The dilution was prepared with a solution of 1% normal goat serum and 0.25% Triton X-100 in 0.1 M Trizma-buffered saline. This was followed by a 2.5-h incubation in a 1/200 dilution of biotinylated goat anti-rabbit IgG (Vector Labs, Burlingame, CA, U.S.A.). The tissue was then incubated for 2 h with the avidin–biotin complex diluted 1/100 (Vector). Each incubation was followed by three rinses. Dilutions and rinses were with 1% normal goat serum in 0.1 M Trizma-buffered saline. The sections were then treated for 6 min with a 0.05% solution of 3,3′-diaminobenzidine and 0.01% hydrogen peroxide, rinsed and mounted on gel-coated slides. Sections were then rinsed in 5% potassium ferrocyanide for Prussian Blue reaction to stain for iron, counterstained with Neutral Red, cleared in xylene and coverslipped with Permount mounting medium. The electrode tracks and the TH-positive cells were visualized and plotted on a Nikon microscope equipped with plotting software (DiLog Instruments and Systems, FL, U.S.A.). The plotted images were then superimposed on to a dog brain atlas.37
RESULTS
Anatomical identification of recorded locus coeruleus cells
A total of 339 cells were recorded from the dorsal pontine area. The majority of these cells increased discharge during waking movements and also during REM sleep, relative to quiet waking and non-REM sleep values. Thirteen of the cells met the criteria for noradrenergic cells. They were recorded from three of the four dogs used (two, four and seven LC cells, respectively, for the three dogs). These cells were located in TH-positive cell regions (Figs 1, 2). TH-positive cells in the narcoleptic dogs were scattered along the medial portion of the superior cerebellar peduncle in the pons and in areas of LC proper and locus subcoeruleus (SC). Nine of the 13 noradrenergic cells were recorded from the rostral portion of the LC and SC, while the other four were recorded from the caudal part of the LC (Fig. 2). All of these cells showed similar behavior across the sleep cycle and in cataplexy.
Fig. 1.
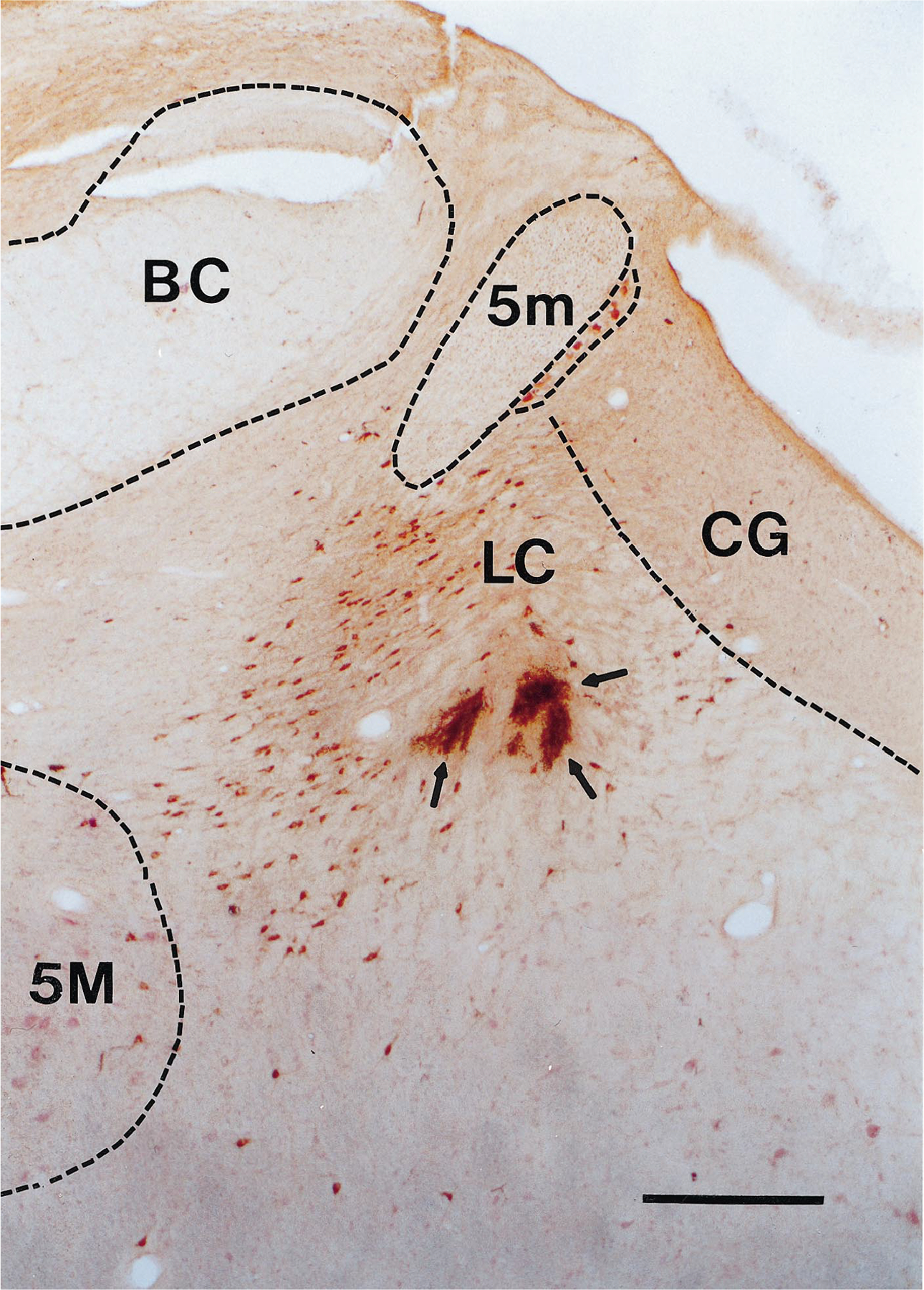
Coronal section at R3 level showing lesion marks made at the electrode tips (arrows) in a region of TH-positive cells (stained brown). Scale bar = 0.5 mm. BC, brachium conjunctivum; CG, central gray; 5M, motor trigeminal nucleus; 5m, tract of the mesencephalic trigeminal nucleus.
Fig. 2.
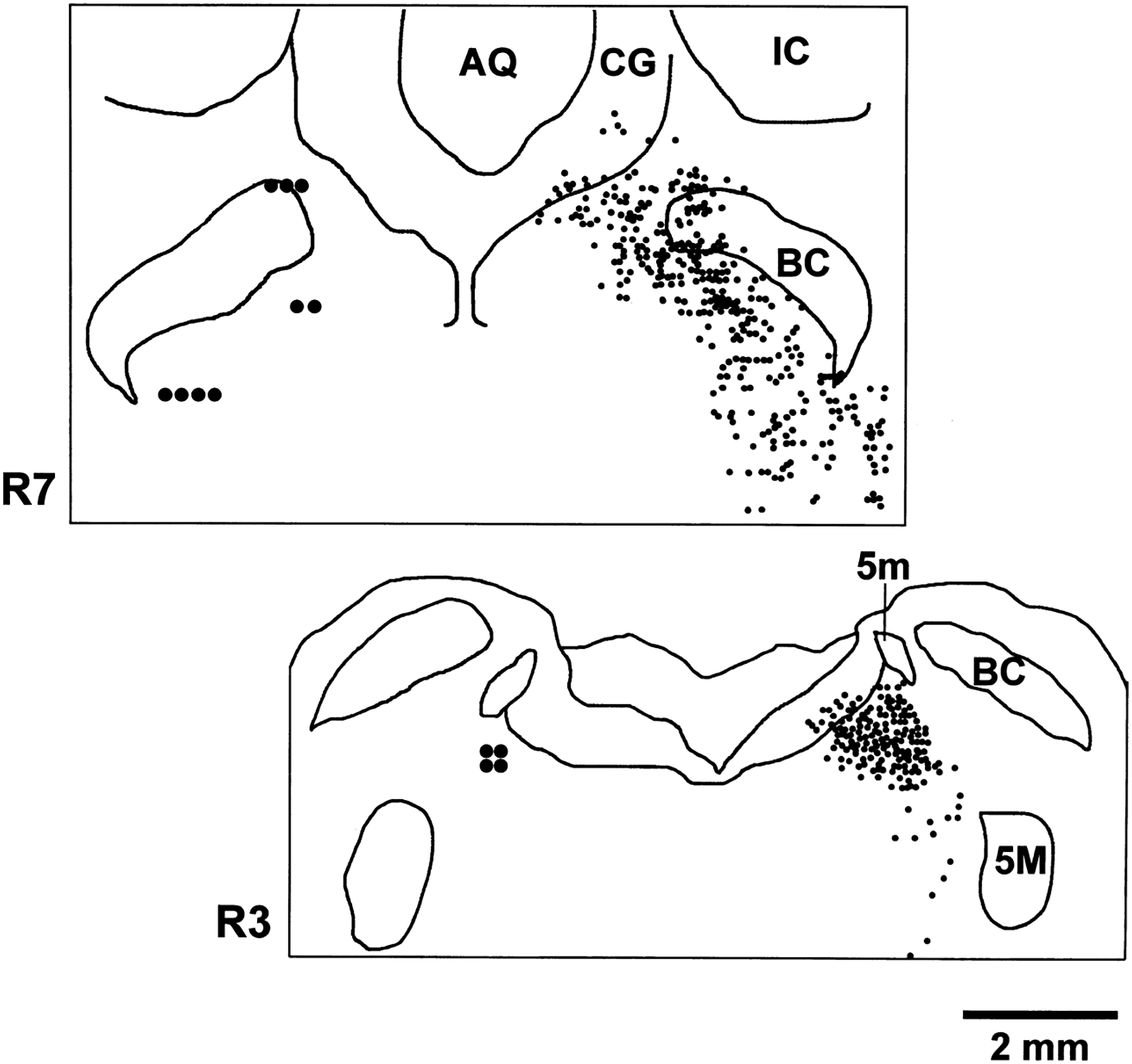
Coronal plots at R3 and R7 levels showing location of recorded REM-cataplexy-off cells on the left and TH-stained cells on the right. REM-cataplexy-off cells were all in regions with TH-positive cells. AQ, aqueduct; IC, inferior colliculus.
Physiological and behavioral characteristics of the locus coeruleus cells
All of the cells meeting the criteria for noradrenergic cells had spike duration greater than 2 ms, with an average of 2.83 ± 0.15 ms (Fig. 3). They all discharged slowly and regularly in quiet waking (QW), phasically increased firing during active waking (AW), slowed down considerably in non-REM sleep and had minimal discharge during REM sleep (average firing rate in mean Hz ± S.E.M.: AW, 1.20 ± 0.08; QW, 0.91 ± 0.09; non-REM sleep, 0.18 ± 0.03; REM sleep, 0.04 ± 0.01; Fig. 3). They ceased discharge before REM sleep onset and resumed activity at a low rate several seconds before the termination of REM sleep. In all cells, the return of discharge coincided with the cessation of hippocampal theta (Fig. 4).
Fig. 3.

Discharge patterns of a noradrenergic REM-cataplexy-off cell during quite waking (QW), non-REM sleep (nonREM) and REM sleep (REM). The insert depicts the spike waveform of the same cell. It has a spike width of 2.8 ms. Unit, computer triggered spike display; HIPP, hippocampal EEG.
Fig. 4.

Discharge pattern of a noradrenergic REM-cataplexy-off cell during the transition from REM sleep to waking. Note that discharge of the cell resumed about 16.6 s before the end of REM sleep, as indicated by the recovery of EMG activity. The cell began firing at a low rate coincident with the end of REM sleep hippocampal theta activity (arrow).
All of these cells responded to stimulation of several sensory modalities in both waking and sleep states. An example of response to auditory stimulation is shown in Fig. 5. Response latency to an 85-dB, 3.8-kHz tone pip was 40–60 ms. All the putative noradrenergic cells tested (seven of 13) reduced firing after clonidine administration. The inhibition occurred within minutes of intravenous injection and lasted for 30–60 min (Fig. 6). The discharge rate was reduced by an average of 78.1 ± 4.8% (from 1.23 ± 1.2 to 0.26 ± 0.06 Hz, P < 0.001) 10 min following clonidine injection (Fig. 6, insert).
Fig. 5.

Histogram showing the responses of an LC cell to 20-ms, 80-dB, 3.8-kHz tone pips. A total of 20 trials are delivered. LC cells respond to auditory stimuli with a latency of 40–60 ms.
Fig. 6.

Effects of clonidine (2 μg/kg, i.v.) on the spontaneous discharge of an LC cell. Clonidine produced an 86% reduction in the discharge of this cell 10 min after injection as compared to its immediate pre-drug level. Clonidine reduced discharge for about 45 min. The reduction of discharge by clonidine, a consequence of α2 receptor activation, is characteristic of noradrenergic cells. Insert: average discharge rate (mean ± S.E.M.) before and after clonidine administration in the seven cells tested. Baseline discharge rates were taken during the 10 min period before drug administration in QW. The effect of clonidine was assessed 10–30 min (i.v.; n = 6) or 1 h (oral; n = 1) after drug administration during QW. * denotes significant difference (P < 0.001, paired t-test) in firing rate between baseline and post-drug condition.
Behavior of locus coeruleus cells during cataplexy and sleep
Cataplexy was operationally defined as an abrupt loss of muscle tone in the alert waking state triggered during play or food intake. Cataplexy was accompanied by pronounced hippocampal theta activity similar to that seen in REM sleep. Eyes typically remained open and could follow moving objects. However, spontaneous eye movements were minimal. Apnea often occurred at cataplexy onset.52 Cataplexy normally terminated abruptly with the dog rising to its feet. In other cases cataplexy might be followed by eye closure and rapid eye movements signifying REM sleep. During cataplexy, all presumptive noradrenergic cells ceased discharge or reduced discharge to an extent comparable to or greater than that seen in REM sleep (Fig. 7). This relationship was highly consistent, occurring in every cell and in every cataplexy period. The cells reduced discharge 2–4 s before the loss of muscle tone (Fig. 7). The cells resumed firing at a low level more than 10 s before the termination of cataplexy, with a marked increase in discharge 2–4 s prior to the increase of muscle tone that terminated cataplexy (Fig. 7). They showed the same resumption of firing prior to the end of REM sleep, cf. Fig. 4. Cell activity during partial cataplexy (e.g., collapse of the hindlimbs; Fig. 8) was similar to that during complete cataplexy: reduction or cessation of discharge a few seconds before the onset of partial cataplexy, silence during the period of reduced tone, and resumption of discharge with or before the recovery of EMG activity. Figure 9 shows the discharge profile of individual cells during cataplexy and other sleep/waking states. Average firing rate during cataplexy was 0.018 ± 0.003 Hz, not significantly different from the rate in REM sleep (0.04 ± 0.01), but significantly different from that of AW (1.20 ± 0.08), QW (0.91 ± .0.09), and non-REM sleep (0.18 ± 0.03), all P < 0.001 with paired t-test.
Fig. 7.
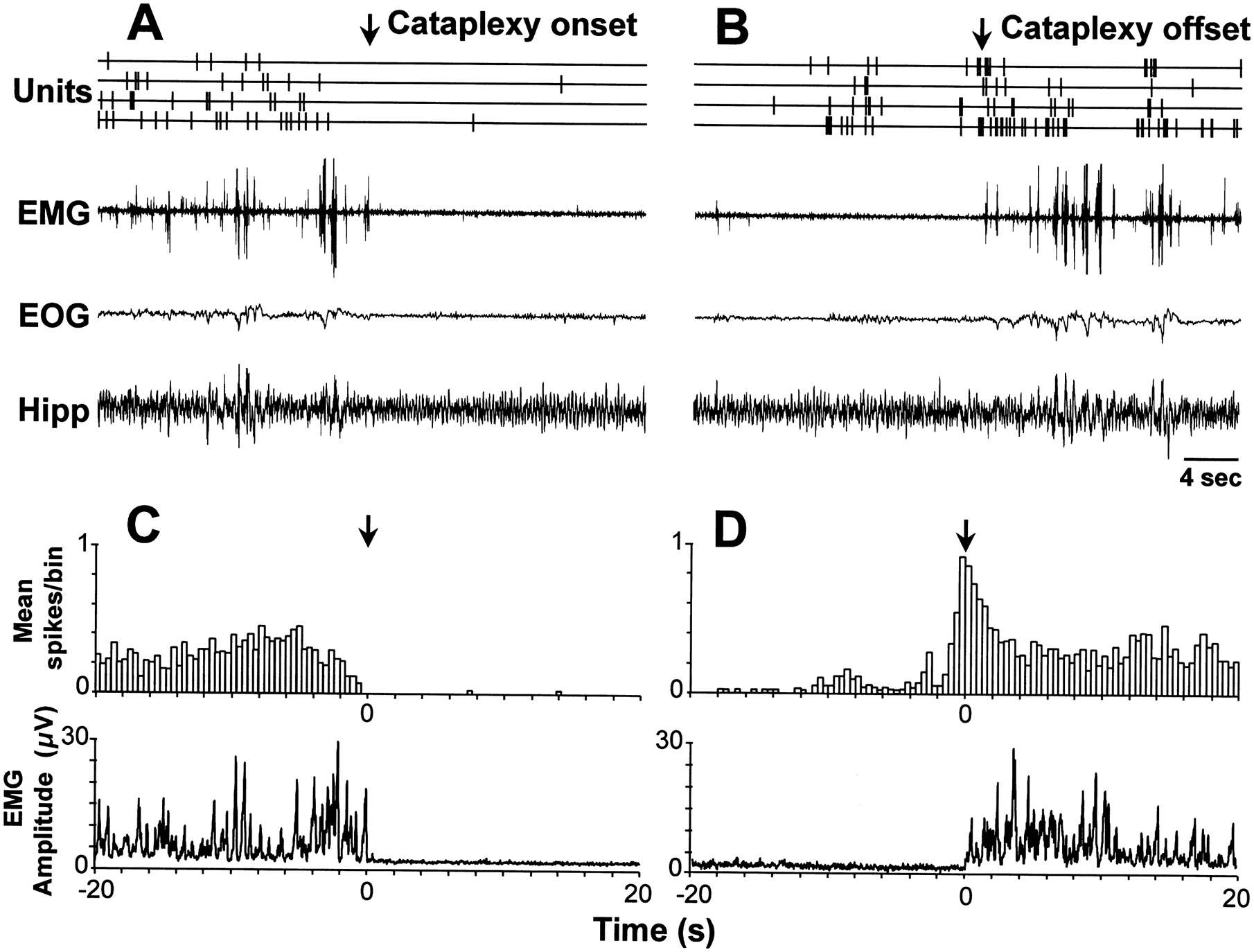
LC discharge during cataplexy. We were able to simultaneously record four REM-cataplexy-off cells throughout sleep/waking states and during episodes of cataplexy. (Top) Unit activity and polygraph traces during cataplexy onset (A) and offset (B). Note the cessation of discharge at cataplexy onset (A) and the resumption of discharge shortly before the termination of cataplexy (B). (Bottom) Averaged spike histogram and EMG before and after cataplexy onset (C) and offset (D). Twenty cataplexy episodes (mean duration 23.7 ± 3.7 s) were averaged across the four cells displayed above. The bin size of the histogram is 400 ms.
Fig. 8.

The activity of two simultaneously recorded REM-cataplexy-off cells during partial cataplexies (gray bars). These cells ceased activity before and during partial cataplexies and resumed firing at or before the recovery of EMG activity, similar to their behavior during complete cataplexies. The EMG signal was full-wave rectified and is displayed as a moving-time average with 500 ms time-constant.
Fig. 9.

Discharge rate profile of REM-cataplexy-off cells recorded across all sleep/waking states and during cataplexy. Rate is minimal in the atonic states of cataplexy and REM sleep, higher in non-REM sleep and highest in waking.
Drug effects on locus coeruleus discharge
Both prazosin and physostigmine at the doses used greatly increased the propensity for cataplexy. We found that prazosin reduced the firing rate of LC cells in QW by 59.4 ± 15.1% (P < 0.001, n = 6) at the time of its peak effect on cataplexy. Physostigmine also reduced the firing rate of three LC cells tested (34.8 ± 3.2%, P < 0.001), 5–10 min after i.v. administration during the period when the dogs began to show cataplexy.
DISCUSSION
The offset of LC activity coincident with cataplexy provides a cellular mechanism for cataplexy initiation and cessation, and can explain many of the pharmacological effects on cataplexy. All REM sleep-off cells located both rostrally and caudally in the LC ceased discharge in all cataplexy episodes. The α1 blocker prazosin dramatically increases cataplexy in both narcoleptic dogs and humans.2,22,23,42 We find that prazosin, at doses that increase cataplexy, reduced LC discharge. Therefore prazosin appears to have a dual effect of reducing norepinephrine release by discharge reduction, and reducing the postsynaptic effect of norepinephrine by blocking α1 receptors. The mechanism by which prazosin reduces the discharge rate of LC cells is unclear. Prazosin has no substantial inhibitory effect on LC cells in vitro.27,28 The drugs most clinically useful for blocking cataplexy are α2 antagonists and tri-cyclic antidepressants, both of which increase the availability of norepinephrine.42
Physostigmine in doses that consistently increase cataplexy also reduced the discharge of LC neurons. This effect is probably not the result of a direct cholinergic activation of muscarinic receptors on LC neurons, because both systemic injections of physostigmine or of an irreversible acetylcholinesterase inhibitor soman16,17 as well as iontophoretic applications of muscarinic agonists into LC8,16,24,34 increase LC discharge. These studies were done either in anesthetized animals or in vitro. The opposite effect seen in the current study could result from anesthesia used in some of the prior studies, a species difference or the effects of narcolepsy. However, it is more likely that both prazosin and physostigmine reduce LC activity indirectly through multisynaptic mechanisms. Physostigmine may stimulate cholinergic mechanisms in the pons that are known to be capable of precipitating a loss of muscle tone.51 Thus, it appears that cholinergic and noradrenergic systems both contribute to the action of systemically administered physostigmine.
The current finding demonstrates that the complete cessation of discharge in LC neurons during cataplexy is not sufficient to induce REM sleep. However, in some cases, sustained cataplexy attacks eventually terminate in REM sleep in dogs, as they do in humans. This indicates that the cessation of LC discharge, when combined with changes in the activity of cholinergic or other brainstem neuronal populations,53,54 can lead to REM sleep.
The major behavioral characteristic common to the states of cataplexy and REM sleep is muscle atonia. Lifting cats by the scruff of the neck, which causes a muscle atonia response and associated autonomic changes, is the only behavioral manipulation associated with a great reduction in LC discharge in waking.49 These findings suggest a strong correlation between LC activity and muscle tone. One third of LC neurons and the majority of non-LC noradrenergic cells have descending projections to spinal and brainstem motoneurons.32,39,43,48,50 A number of studies have demonstrated that norepinephrine released by the descending projections of LC neurons potently facilitates activity in spinal motoneurons, presumably acting through α1 adrenoceptors in the ventral horn.13,20,21,44,57 Unilateral or bilateral lesions of the LC, or chemical inactivation by clonidine microinjection, produce a reduction of ipsilateral or bilateral muscle tone.13 Thus cessation of LC discharge in REM sleep and cataplexy would disfacilitate motor activity. LC neurons also have strong projections to pontine and medullary regions involved in the circuit responsible for the hyperpolarization of motoneurons,9 a circuit that is activated both in cataplexy and in REM sleep.54 We have previously shown that cataplexy is accompanied by increased discharge in a population of cells in the nucleus magnocellularis of the medulla,53 the medullary inhibitory area. Stimulation of this area produces suppression of muscle tone.36 These results suggest that REM sleep atonia and the atonia of cataplexy result from aminergic disfacilitation,35 co-ordinated with active inhibition of motoneurons.10
LC cells have major ascending projections.31 These projections have been hypothesized to have a role in attention.7 The ascending projections of LC cells may help facilitate forebrain processing of sensory inputs generated by motor activity.
Blood pressure elevation, induced by intravenous infusion of norepinephrine or angiotensin II, causes an immediate reduction in LC cell discharge15 while nitroprusside-induced hypotension increases the discharge of LC neurons.58 Since cataplexy is normally triggered by emotional events, one may speculate that changes in blood pressure are linked to the cessation of LC activity during cataplexy. However, no consistent blood pressure change occurs during the onset of canine cataplexy.52 Furthermore, the transient increase of heart rate that we saw prior to cataplexy onset returns to baseline values52 and therefore cannot account for the prolonged inactivation of LC cells throughout the cataplexy period.
The loss of consciousness that accompanies slow-wave sleep can be seen as resulting from the occlusion of sensory processing by the synchronized rhythmicity of thalamocortical circuits.55,56 However, both REM sleep and waking are characterized by a low voltage EEG. This pattern, generated by thalamocortical interactions, is under the control of projections from brainstem cholinergic and aminergic cells. Despite the similarity of cortical EEG in waking and REM sleep, the organism is unconscious and experiences dream mentation only during REM sleep. Humans report unimpaired consciousness and memory for events transpiring during cataplexy.22 Narcoleptic dogs show their awareness of the environment during cataplexy by visually tracking objects of interest.41 The activity of aminergic cells groups in waking, and their inactivity during REM sleep was a possible explanation for the difference in the nature of consciousness in these two states. However, the current results demonstrate that awareness of the environment does not require the activity of LC cells in narcoleptic dogs. These results either reflect the independence of LC cell activity from the maintenance of awareness or represent a CNS adaptation in narcoleptics to the periodic loss of activity in LC cells. It remains to be seen if serotonergic and histaminergic cells, the other two groups known to be tonically active in waking and silent in REM sleep,38,40,59 are silent during cataleptic attacks.
CONCLUSION
The present results show a strong correlation between the cessation of LC activity and cataplexy. Drugs that enhance cataplexy reduced spontaneous LC activity during the time that the propensity for cataplexy was enhanced. These results suggest that cessation of LC activity and consequent disfacilitation of motoneurons is critical for the initiation of cataplexy. Cessation of discharge in the LC may act synergistically with activation of the medullary atonia system to produce motor inhibition in REM sleep and cataplexy.
Acknowledgements—
The authors thank Heidi Fahringer for her excellent histological assistance. This work is supported by the Medical Research Service of the Department of Veterans Affairs, USPHS NS14610 and NS23724.
Abbreviations:
- AW
active waking
- EEG
electroencephalogram
- EMG
electromyogram
- EOG
electrooculogram
- LC
locus coeruleus
- QW
quiet waking
- REM
rapid-eye-movement
- SC
locus subcoeruleus
- TH
tyrosine hydroxylase
REFERENCES
- 1.Abercrombie ED and Jacobs BL (1987) Single-unit response of noradrenergic neurons in the locus coeruleus of freely moving cats. I. Acutely presented stressful and nonstressful stimuli. J. Neurosci 7, 2837–2843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Aldrich MS (1990) Narcolepsy. New Engl. J. Med 323, 389–394. [DOI] [PubMed] [Google Scholar]
- 3.Amaral DG and Sinnamon HM (1977) The locus coeruleus: neurobiology of a central noradrenergic nucleus. Prog. Neurobiol 9, 147–196. [DOI] [PubMed] [Google Scholar]
- 4.Anlezark G, Crow T and Greenway A (1973) Impaired learning and decreased cortical norepinephrine after bilateral locus coeruleus lesions. Science 181, 682–684. [DOI] [PubMed] [Google Scholar]
- 5.Aston-Jones G and Bloom FE (1981) Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J. Neurosci 1, 876–886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Aston-Jones G, Foote SL and Bloom FE (1984) Norepinephrine. In Norepinephrine (eds Ziegler M and Lake R), pp. 92–116. Williams and Wilkins, Baltimore. [Google Scholar]
- 7.Aston-Jones G, Rajkowski J, Kubiak P and Alexinsky T (1994) Locus coeruleus neurons in monkey are selectively activated by attended cues in a vigilance task. J. Neurosci 14, 4467–4480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bird S and Kuhar MJ (1977) Iontophoretic application of opiates to the locus coeruleus. Brain Res. 122, 523–533. [DOI] [PubMed] [Google Scholar]
- 9.Bowden D, German D and Poynter W (1978) An autoradiographic, semistereotaxic mapping of major projections from locus coeruleus and adjacent nuclei in Macaca mulatta. Brain Res. 145, 257–276. [DOI] [PubMed] [Google Scholar]
- 10.Chase MH and Morales FR (1990) The atonia and myoclonia of active (REM) sleep. A. Rev. Psychol 41, 557–584. [DOI] [PubMed] [Google Scholar]
- 11.Cirelli C, Pompeiano M and Tononi G (1996) Neuronal gene expression in the waking state: a role for the locus coeruleus. Science 274, 1211–1217. [DOI] [PubMed] [Google Scholar]
- 12.Cornwell-Jones CA, Decker MW, Chang JW, Cole B, Goltz KM, Tran T and McGaugh JL (1989) Neonatal 6-hydroxydopa, but not DSP-4, elevates brainstem monoamines and impairs inhibitory avoidance learning in developing rats. Brain Res. 493, 258–268. [DOI] [PubMed] [Google Scholar]
- 13.D’Ascanio P, Pompeiano O and Stampacchia G (1988) Noradrenergic and cholinergic mechanisms responsible for the gain regulation of vestibulospinal reflexes. Prog. Brain Res 76, 361–374. [DOI] [PubMed] [Google Scholar]
- 14.Devauges V and Sara SJ (1990) Activation of the noradrenergic system facilitates an attentional shift in the rat. Behav. Brain Res 39, 19–28. [DOI] [PubMed] [Google Scholar]
- 15.Elam M, Yao T, Svensson TH and Thoren P (1984) Regulation of locus coeruleus neurons and splanchnic, sympathetic nerves by cardiovascular afferents. Brain Res. 290, 281–287. [DOI] [PubMed] [Google Scholar]
- 16.Engberg G and Svensson TH (1980) Pharmacological analysis of a cholinergic receptor mediated regulation of brain norepinephrine neurons. J. neural Transm 49, 137–150. [DOI] [PubMed] [Google Scholar]
- 17.Ennis M and Shipley MT (1992) Tonic activation of locus-coeruleus neurons by systemic or intracoerulear microinjection of an irreversible acetylcholinesterase inhibitor—increased discharge rate and induction of c-fos. Expl Neurol. 118, 164–177. [DOI] [PubMed] [Google Scholar]
- 18.Foote SL, Aston-Jones G and Bloom FE (1980) Impulse activity of locus coeruleus neurons in awake rats and monkeys is a function of sensory stimulation and arousal. Proc. natn. Acad. Sci. U.S.A 77, 3033–3037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Foote SL, Bloom FE and Aston-Jones G (1983) Nucleus locus coeruleus: new evidence of anatomical and physiological specificity. Physiol. Rev 63, 844–914. [DOI] [PubMed] [Google Scholar]
- 20.Fung SJ, Manzoni D, Chan JYH, Pompeiano O and Barnes CD (1991) Locus coeruleus control of spinal motor output. Prog. Brain Res 88, 395–409. [DOI] [PubMed] [Google Scholar]
- 21.Fung SJ, Pompeiano O and Barnes CD (1988) Coerulospinal influence on recurrent inhibition of spinal motonuclei innervating antagonistic hindleg muscles of the cat. Pflügers Arch. 412, 346–353. [DOI] [PubMed] [Google Scholar]
- 22.Guilleminault C (1976) Narcolepsy. In Narcolepsy (eds Guilleminault C, Dement WC and Passouant P), pp. 125–143. Spectrum, New York. [Google Scholar]
- 23.Guilleminault C (1994) Narcolepsy syndrome. In Principles and Practice of Sleep Medicine (eds Kryger MH, Roth T and Dement WC), pp. 145–162. W. B. Saunders, Philadelphia. [Google Scholar]
- 24.Guyenet PG and Aghajanian GK (1979) ACh, substance P and met-enkephalin in the locus coeruleus: pharmacological evidence for independent sites of action. Eur. J. Pharmac 53, 319–328. [DOI] [PubMed] [Google Scholar]
- 25.Harley CW (1987) A role for norepinephrine in arousal, emotion and learning?: limbic modulation by norepinephrine and the Kety hypothesis. Prog. Neuro-Psychopharmac. biol. Psychiat 11, 419–458. [DOI] [PubMed] [Google Scholar]
- 26.Hobson JA, McCarley RW and Wyzinski PW (1975) Sleep cycle oscillation: reciprocal discharge by two brainstem neuronal groups. Science 189, 55–58. [DOI] [PubMed] [Google Scholar]
- 27.Illes P and Norenberg W (1990) Blockade of a2-adrenoceptors increases opioid a-receptor-mediated inhibition of the firing rate of rat locus coeruleus neurons. Naunyn-Schmiedeberg’s Arch. Pharmac 342, 490–496. [DOI] [PubMed] [Google Scholar]
- 28.Ivanov A and Aston-Jones G (1995) Extranuclear dendrites of locus coeruleus neurons: activation by glutamate and modulation of activity by alpha adrenoceptors. J. Neurophysiol 74, 2427–2436. [DOI] [PubMed] [Google Scholar]
- 29.Jacobs BL (1986) Single unit activity of locus coeruleus neurons in behaving animals. Prog. Neurobiol 27, 183–194. [DOI] [PubMed] [Google Scholar]
- 30.Jacobs BL, Heym J and Trulson ME (1981) Behavioral and physiological correlates of brain serotoninergic unit activity. J. Physiol 77, 431–436. [PubMed] [Google Scholar]
- 31.Jones BE (1994) Basic mechanisms of sleep-wake states. In Principles and Practice of Sleep Medicine (eds Kryger M, Roth T and Dement WC), pp. 145–162. W. B. Saunders, Philadelphia. [Google Scholar]
- 32.Jones BE (1991) Noradrenergic locus coeruleus neurons: their distant connections and their relationship to neighboring (including cholinergic and GABAergic) neurons of the central gray and reticular formation. Prog. Brain Res 88, 15–30. [DOI] [PubMed] [Google Scholar]
- 33.Kasamatsu T and Pettigrew J (1976) Depletion of brain catecholamines: failure of ocular dominance shift after monocular occlusion in kittens. Science 194, 206–209. [DOI] [PubMed] [Google Scholar]
- 34.Koyama Y and Kayama Y (1993) Mutual interactions among cholinergic, noradrenergic and serotonergic neurons studied by ionophoresis of these transmitters in rat brainstem nuclei. Neuroscience 55, 1117–1126. [DOI] [PubMed] [Google Scholar]
- 35.Kubin L, Reignier C, Tojima H, Taguchi O, Pack AI and Davis RO (1994) Changes in serotonin level in the hypoglossal nucleus region during carbachol-induced atonia. Brain Res. 645, 291–302. [DOI] [PubMed] [Google Scholar]
- 36.Lai YY and Siegel JM (1988) Medullary regions mediating atonia. J. Neurosci 8, 4790–4796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lim RKS, Liu C-N and Moffitt RL (1960) A Stereotaxic Atlas of the Dog’s Brain. Charles C. Thomas, Springfield, IL. [Google Scholar]
- 38.Lin JS, Sakai K and Jouvet M (1986) Role of hypothalamic histaminergic systems in the regulation of vigilance states in cats. C. r. hebd Séanc. Acad. Sci., Paris 303, 469–474. [PubMed] [Google Scholar]
- 39.Lyons WE, Fritschy JM and Grzanna R (1989) The noradrenergic neurotoxin DSP-4 eliminates the coerulospinal projection but spares projections of the A5 and A7 groups to the ventral horn of the rat spinal cord. J. Neurosci 9, 1481–1489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.McGinty DJ and Harper RM (1976) Dorsal raphe neurons: depression of firing rates during sleep in cats. Brain Res. 101, 569–575. [DOI] [PubMed] [Google Scholar]
- 41.Mitler MM, Soave O and Dement WC (1976) Narcolepsy in seven dogs. J. Am. vet. med. Assoc 168, 1036–1038. [PubMed] [Google Scholar]
- 42.Nishino S and Mignot E (1997) Pharmacological aspects of human and canine narcolepsy. Prog. Neurobiol 52, 27–78. [DOI] [PubMed] [Google Scholar]
- 43.Nygren LG and Olson L (1977) A new major projection from locus coeruleus: the main source of noradrenergic nerve terminals in the ventral and dorsal columns of the spinal cord. Brain Res. 132, 85–93. [DOI] [PubMed] [Google Scholar]
- 44.Palmeri A and Wiesendanger M (1990) Concomitant depression of locus coeruleus neurons and of flexor reflexes by an alpha2-adrenergic agonist in rats: a possible mechanism for an alpha2-mediated muscle relaxation. Neuroscience 34, 177–187. [DOI] [PubMed] [Google Scholar]
- 45.Pineda JA, Foote SL and Neville HJ (1989) Effects of locus coeruleus lesions on auditory, long-latency, event-related potentials in monkey. J. Neurosci 9, 81–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rasmussen K, Morilak DA and Jacobs BL (1986) Single unit response of locus coeruleus neurons in the freely moving cat. I. During naturalistic behaviors and in response to simple and complex stimuli. Brain Res. 371, 324–334. [DOI] [PubMed] [Google Scholar]
- 47.Rechtschaffen A, Wolpert EA, Dement WC, Mitchel SA and Fisher C (1963) Nocturnal sleep of narcoleptics. Electroenceph. clin. Neurophysiol 15, 599–609. [DOI] [PubMed] [Google Scholar]
- 48.Reddy VK, Fung SJ, Zhuo H and Barnes CD (1989) Spinally projecting noradrenergic neurons of the dorsolateral pontine tegmentum: a combined immunocytochemical and retrograde labeling study. Brain Res. 491, 144–149. [DOI] [PubMed] [Google Scholar]
- 49.Reiner PB (1986) Correlational analysis of central noradrenergic neuronal activity and sympathetic tone in behaving cats. Brain Res. 378, 86–96. [DOI] [PubMed] [Google Scholar]
- 50.Satoh K, Tohyama M, Yamamoto KST and Shimizu N (1977) Noradrenaline innervation of the spinal cord studied by the horseradish peroxidase method combined with monoamine oxidase staining. Expl Brain Res. 30, 175–186. [DOI] [PubMed] [Google Scholar]
- 51.Siegel JM (1994) Brain mechanisms generating REM sleep. In Principles and Practice of Sleep Medicine (eds Kryger MH, Roth T and Dement WC), pp. 125–144. W. B. Saunders, Philadelphia. [Google Scholar]
- 52.Siegel JM, Fahringer H, Tomazewski KS, Kaitin K, Kilduff T and Dement WC (1986) Heart rate and blood pressure changes associated with cataplexy in canine narcolepsy. Sleep 9, 216–221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Siegel JM, Nienhuis R, Fahringer HM, Chiu C, Dement WC, Mignot E and Lufkin R (1992) Activity of medial mesopontine units during cataplexy and sleep-waking states in the narcoleptic dog. J. Neurosci 12, 1640–1646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Siegel JM, Nienhuis R, Fahringer HM, Paul R, Shiromani P, Dement WC, Mignot E and Chiu C (1991) Neuronal activity in narcolepsy: identification of cataplexy-related cells in the medial medulla. Science 252, 1315–1318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Steriade M (1996) Arousal: revisiting the reticular activating system. Science 272, 225–226. [DOI] [PubMed] [Google Scholar]
- 56.Steriade M, McCormick DA and Sejnowski TJ (1993) Thalamocortical oscillations in the sleeping and aroused brain. Science 262, 679–685. [DOI] [PubMed] [Google Scholar]
- 57.Tanabe M, Ono H and Fukuda H (1990) Spinal alpha 1- and alpha 2-adrenoceptors mediate facilitation and inhibition of spinal motor transmission, respectively. Jap. J. Pharmac 54, 69–77. [DOI] [PubMed] [Google Scholar]
- 58.Valentino RJ, Page ME and Curtis AL (1991) Activation of noradrenergic locus coeruleus neurons by hemodynamic stress is due to local release of corticotrophin-releasing factor. Brain Res. 555, 25–34. [DOI] [PubMed] [Google Scholar]
- 59.Vanni-Mercier G, Sakai K and Jouvet M (1984) Waking-state specific neurons in the caudal hypothalamus of the cat. C. r. hebd Séanc. Acad. Sci., Paris 298, 195–200. [PubMed] [Google Scholar]