

Abstract
We performed a molecular and cytogenetic analysis on different Mantellinae species and revised the available chromosomal data on this group to provide an updated assessment of its karyological diversity and evolution. Using a fragment of the mitochondrial 16S rRNA, we performed a molecular taxonomic identification of the samples that were used for cytogenetic analyses. A comparative cytogenetic analysis, with Giemsa’s staining, Ag-NOR staining and sequential C-banding + Giemsa + CMA + DAPI was performed on eight species: Gephyromantis sp. Ca19, G.striatus (Vences, Glaw, Andreone, Jesu et Schimmenti, 2002), Mantidactylus (Chonomantis) sp. Ca11, M. (Brygoomantis) alutus (Peracca, 1893), M. (Hylobatrachus) cowanii (Boulenger, 1882), Spinomantispropeaglavei “North” (Methuen et Hewitt, 1913), S.phantasticus (Glaw et Vences, 1997) and S. sp. Ca3. Gephyromantisstriatus, M. (Brygoomantis) alutus and Spinomantispropeaglavei “North” have a karyotype of 2n = 24 chromosomes while the other species show 2n = 26 chromosomes. Among the analysed species we detected differences in the number and position of telocentric elements, location of NOR loci (alternatively on the 6th, 7th or 10th pair) and in the distribution of heterochromatin, which shows species-specific patterns. Merging our data with those previously available, we propose a karyotype of 2n = 26 with all biarmed elements and loci of NORs on the 6th chromosome pair as the ancestral state in the whole family Mantellidae. From this putative ancestral condition, a reduction of chromosome number through similar tandem fusions (from 2n = 26 to 2n = 24) occurred independently in Mantidactylus Boulenger, 1895 (subgenus Brygoomantis Dubois, 1992), Spinomantis Dubois, 1992 and Gephyromantis Methuen, 1920. Similarly, a relocation of NORs, from the putative primitive configuration on the 6th chromosome, occurred independently in Gephyromantis, Blommersia Dubois, 1992, Guibemantis Dubois, 1992, Mantella Boulenger, 1882 and Spinomantis. Chromosome inversions of primitive biarmed elements likely generated a variable number of telocentric elements in Mantellanigricans Guibé, 1978 and a different number of taxa of Gephyromantis (subgenera Duboimantis Glaw et Vences, 2006 and Laurentomantis Dubois, 1980) and Mantidactylus (subgenera Brygoomantis, Chonomantis Glaw et Vences, 1994, Hylobatrachus Laurent, 1943 and Ochthomantis Glaw et Vences, 1994).
Keywords: Amphibia, chromosome evolution, karyotype, Madagascar, NORs
Introduction
Madagascar is one of the richest biodiversity hotspots and an ideal region to study evolutionary dynamics (Myers et al. 2000; Ganzhorn et al. 2001; Vences et al. 2009). The native amphibians of Madagascar belong to four distinct anuran families: Hyperoliidae, Mantellidae, Microhylidae and Ptychadenidae (Glaw and Vences 2007). Among them, the family Mantellidae includes ca 230 described species (AmphibiaWeb 2021; Frost et al. 2021), representing the most species-rich amphibian group of the island.
Mantellidae are characterized by an extraordinary ecological and morphological diversity (Glaw and Vences 2007; Wollenberg et al. 2011; AmphibiaWeb 2021) and are subdivided into three subfamilies: Laliostominae with an overall low species diversity, including the genera Laliostoma Glaw, Vences et Böhme, 1998 (1 species) and Aglyptodactylus Boulenger, 1919 (6 species); Boophinae, a species-rich clade of about 80 described species of tree frogs, all belonging to the genus Boophis Tschudi, 1838, and Mantellinae, which is by far the most species-rich group including nine genera and more than 140 described species (Glaw and Vences 2007; AmphibiaWeb 2021).
The last three decades have seen the flourishing of the use of molecular techniques, with numerous taxonomic and systematic studies that clarified the relationships among the major groups within this subfamily (Glaw et al. 1998; Vences et al. 1998; Richards et al. 2000; Glaw and Vences 2006; Wollenberg et al. 2011; Kaffenberger et al. 2012). Similarly, these tools have been used in the identification of candidate species (Vieites et al. 2009; Perl et al. 2014) and have later contributed to the formal description of many of them (e.g. Andreone et al. 2003; Crottini et al. 2011a; Cocca et al. 2020; AmphibiaWeb 2021).
However, in contrast to the fast-growing amount of molecular data on Mantellidae, the available chromosomal data remain limited, leaving the karyological diversification of the family mostly unexplored. In particular, available cytogenetic data on the subfamily Mantellinae, obtained using different methods, come from a handful of studies (Morescalchi 1967; Blommers-Schlösser 1978; Pintak et al. 1998; Odierna et al. 2001; Andreone et al. 2003), together providing the description of the karyotype of ca. 40 species. These studies revealed the occurrence of a conserved chromosome number in most genera (2n = 26), but a marked difference in chromosome morphology, location of NORs and heterochromatin distribution (see Odierna et al. 2001 and Andreone et al. 2003). Differences in chromosome number (2n = 24) were also identified, with five species of the subgenus Brygoomantis all sharing this state, thus suggesting that the state of 2n = 24 is a derived feature of the group (Blommers-Schlösser 1978).
Comparative cytogenetics, especially when linked to phylogenetic inference, offers the possibility to identify plesio- and apomorphic states, and recognizes different evolutionary lineages (see e.g. Mezzasalma et al. 2013, 2014, 2017a). However, both the limited taxon sampling and the outdated taxonomy used in most previous works limited the possibility to draw robust comparisons and consistent hypotheses on the evolution of chromosomal diversification in the subfamily.
In this study we performed a comparative cytogenetic analysis on eight mantellid species belonging to the genera Gephyromantis Methuen, 1920, Mantidactylus Boulenger, 1895 (subgenera Chonomantis, Brygoomantis and Hylobatrachus) and Spinomantis Dubois, 1992, using a combination of standard coloration and banding methods. We coupled cytogenetic analyses with the molecular taxonomic identification of the samples and synthesized previously available information on this subfamily to produce an overview of their chromosomal diversity. This, enable us to propose a hypothesis on the chromosome diversification in mantelline frogs.
Material and methods
Sampling
We studied 13 samples of eight mantelline species belonging to the genera Gephyromantis, Mantidactylus (subgenera Chonomantis, Brygoomantis and Hylobatrachus) and Spinomantis. These samples were collected between 1999 and 2004 and conserved as cell suspensions at the University of Naples Federico II.
The list of samples used in this study is provided in Table 1. To provide an overview of the chromosomal data on Malagasy mantelline frogs, we reviewed previously published karyotypes of the subfamily. A complete list of all the considered taxa and karyotypes is provided in Table 2.
Table 1.
Specimens analysed in this study. MRSN = Museo Regionale di Storia Naturale (Turin, Italy); ZMA = Zoological Museum Amsterdam (Amsterdam, Netherlands); FN and FAZC, field numbers of Franco Andreone; GA field numbers of Gennaro Aprea; FG/MV, field numbers of Frank Glaw and Miguel Vences.
| Species | Field Number | Sex | Locality |
|---|---|---|---|
| Gephyromantisstriatus | MRSN A1988 (FN 7645) | female | Ambatoledama Corridor: Beanjada |
| Gephyromantis sp. Ca19 | MRSN A2109 (FN 7630) | male | Ambatoledama Corridor: Beanjada |
| Gephyromantis sp. Ca19 | MRSN A2075 (FN 7903) | male | Ambatoledama Corridor: Andasin’i Governera |
| Gephyromantis sp. Ca19 | MRSN A2112 (FN 7890) | male | Ambatoledama Corridor: Andasin’i Governera |
| Gephyromantis sp. Ca19 | MRSN A2108 (FN 7566) | female | Ambatoledama Corridor: Beanjada |
| Mantidactylus (Brygoomantis) alutus (Peracca, 1893) | MRSN A3639 (FN 7945) | female | Ankaratra: Manjakatompo |
| Mantidactylus (Chonomantis) sp. Ca11 | MRSN A3708 (FN 7545) | male | Ambatoledama Corridor: Beanjada |
| Mantidactylus (Hylobatrachus) cowanii (Boulenger, 1882) | MRSN A2612 (FAZC 11370) | female | Antoetra: Soamazaka |
| Mantidactylus (Hylobatrachus) cowanii | GA 720 | male | Mandraka |
| Spinomantispropeaglavei “North” (Methuen et Hewitt, 1913) | MRSN A3563 (FN 7543) | male | Ambatoledama Corridor: Beanjada |
| Spinomantisphantasticus (Glaw et Vences, 1997) | ZMA 19627 (FG/MV 2002-970) | male | Vohidrazana |
| Spinomantis sp. Ca3 | MRSN A3998 (FN 7567) | male | Ambatoledama Corridor: Beanjada |
| Spinomantis sp. Ca3 | MRSN A3999 (FN 7629) | male | Ambatoledama Corridor: Beanjada |
Table 2.
Available karyological data on mantelline frogs. M = metacentric pairs; sm = submetacentric pairs; st = subtelocentric pairs; t = telocentric pairs; AN = arm number; [#] = NOR bearing chromosome pair; CB = C-banding; F = Fluorochrome; R = references; (1) = Morescalchi (1967); (2) = Blommers-Schlösser (1978); (3) = Pintak et al. (1998); (4) = Odierna et al. (2001); (5) = Andreone et al. (2003); (6) = this study. Nomenclature follows Vieites et al. (2009), updated in Perl et al. (2014).
| Genus/subgenus | Species | Karyotype | Banding | R |
|---|---|---|---|---|
| Mantella Boulenger, 1882 | aurantiaca Mocquard, 1900 | 2n = 26 10m 3sm; AN = 52 | (1) | |
| aurantiaca | 2n = 26 10m 3sm; AN = 52 | (2) | ||
| haraldmeieri Busse, 1981 | 2n = 26 9m 4sm; AN = 52 | (2) | ||
| ebenaui (Boettger, 1880) | 2n = 26 11m 2sm; AN = 52 | (2) | ||
| aurantiaca | 2n = 26 11m 2sm; AN = 52 | CB | (3) | |
| crocea Pintak et Böhme, 1990 | 2n = 26 11m 2sm; AN = 52 | CB | (3) | |
| baroni Boulenger, 1888 | 2n = 26 11m 2sm; AN = 52 | CB | (3) | |
| haraldmeieri | 2n = 26 11m 2sm; AN = 52 | CB | (3) | |
| ebenaui | 2n = 26 11m 2sm; AN = 52 | CB | (3) | |
| viridis Pintak et Böhme, 1988 | 2n = 26 11m 2sm; AN = 52 | CB | (3) | |
| laevigata Methuen et Hewitt, 1913 | 2n = 26 11m 2sm; AN = 52 | CB | (3) | |
| baroni | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| ebenaui | 2n = 26 11m 1sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| betsileo (Grandidier, 1872) | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| cowanii | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| expectata Busse et Böhme, 1992 | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| laevigata | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| madagascariensis (Grandidier, 1872) | 2n = 26 11m 1sm; 1st AN = 52 | Ag-NOR [2], CB | (4) | |
| nigricans Guibé, 1978 | 2n = 26 10m 2sm; 1t AN = 48 | Ag-NOR [2], CB | (4) | |
| pulchra Parker, 1925 | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| viridis | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| aurantiaca | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [2], CB | (4) | |
| Blommersia Dubois, 1992 | blommersae (Guibé 1975) | 2n = 26 12m 1sm; AN = 52 | (2) | |
| galani Vences, Köhler, Pabijan, et Glaw 2010 | 2n = 26 12m 1sm; AN = 52 | (2) | ||
| grandisonae (Guibé, 1974) | 2n = 26 10m 3sm; AN = 52 | Ag-NOR [1], CB, F | (5) | |
| Gephyromantis | ||||
| Asperomantis | asper (Boulenger, 1882) | 2n = 26 6m 3sm 4t; AN = 44 | (2) | |
| Duboimantis | granulatus (Boettger, 1881) | 2n = 26 8m 4sm 1t; AN050 | Ag-NOR [8], CB, F | (5) |
| Duboimantis | leucomaculatus (Guibé, 1975) | 2n = 26 6m 6sm 1t; AN = 50 | Ag-NOR [6], CB, F | (5) |
| Duboimantis | luteus (Methuen et Hewitt, 1913) | 2n = 26 6m 4sm 1st 2t; AN = 48 | (2) | |
| Duboimantis | prope luteus Methuen et Hewitt, 1913 | 2n = 26 6m 2sm 1st 4t; AN = 42 | Ag-NOR [11], CB, F | (5) |
| Duboimantis | propemoseri “Masoala” Glaw et Vences, 2002 | 2n = 26 6m 6sm 1t; AN = 52 | (5) | |
| Duboimantis | sp. Ca19 | 2n = 26 8m 5sm; AN = 52 | Ag-NOR [6], CB, F | (6) |
| Duboimantis | redimitus (Boulenger, 1889) | 2n = 26 7m 5sm 1t; AN = 50 | Ag-NOR [6], CB, F | (5) |
| Duboimantis | salegy (Andreone, Aprea, Vences et Odierna, 2003) | 2n = 26 5m 7sm 1st; AN = 52 | Ag-NOR [6], CB, F | (5) |
| Duboimantis | zavona (Vences, Andreone, Glaw et Randrianirina, 2003) | 2n = 26 9m 4sm; AN = 52 | Ag-NOR [6], CB, F | (5) |
| Laurentomantis | striatus | 2n = 24 6m 1sm 5t; AN = 38 | Ag-NOR [10], CB, F | (6) |
| Phylacomantis | pseudoasper (Guibé, 1974) | 2n = 26 7m 7sm; AN = 52 | Ag-NOR [9], CB, F | (5) |
| Guibemantis Dubois, 1992 | ||||
| Guibemantis | depressiceps (Boulenger, 1882) | 2n = 26 10m 3sm; AN = 52 | (2) | |
| Guibemantis | timidus (Vences et Glaw, 2005) | 2n = 26 11m 2sm; AN = 52 | (2) | |
| Pandanusicola | methueni (Angel, 1929) | 2n = 26 11m 2sm; AN = 52 | (2) | |
| Pandanusicola | bicalcaratus (Boettger, 1913) | 2n = 26 11m 2sm; AN = 52 | Ag-NOR [1], CB, F | (4) |
| Pandanusicola | prope bicalcaratus (Boettger, 1913) | 2n = 26 9m 4sm; AN = 52 | (2) | |
| Pandanusicola | liber (Peracca, 1893) | 2n = 26 11m 2sm; AN = 52 | (2) | |
| Pandanusicola | pulcher (Boulenger, 1882) | 2n = 26 9m 4sm; AN = 52 | (2) | |
| Pandanusicola | prope punctatus (Blommers-Schlösser, 1979) | 2n = 26 10m 3sm; AN = 52 | Ag-NOR [1], CB, F | (4) |
| Pandanusicola | punctatus (Blommers-Schlösser, 1979) | 2n = 26 9m 4sm; AN = 52 | (2) | |
| Mantidactylus | ||||
| Brygoomantis | alutus | 2n = 24 12m; AN = 48 | Ag-NOR [6], CB, F | (6) |
| Brygoomantis | ambohimitombi Boulenger 1918 | 2n = 24 9m 3sm; AN = 48 | (2) | |
| Brygoomantis | betsileanus (Boulenger, 1882) | 2n = 24 5m 6sm 1t; AN = 46 | (2) | |
| Brygoomantis | prope biporus (Boulenger, 1889 | 2n = 24 8m 4sm; AN = 48 | (2) | |
| Brygoomantis | sp. Ca19 | 2n = 24 7m 5sm; AN = 48 | (2) | |
| Brygoomantis | prope ulcerosus (Boettger, 1880) | 2n = 24 8m 2sm 1st 1t; AN = 46 | (2) | |
| Chonomantis | prope aerumnalis (Peracca, 1893) | 2n = 26 10m 2sm 1t; AN = 50 | (2) | |
| Chonomantis | sp. Ca11 | 2n = 26 10m 2sm 2t; AN = 50 | (6) | |
| Chonomantis | paidroa Bora, Ramilijaona, Raminosoa et Vences, 2011 | 2n = 26 6m 7sm; AN = 52 | (2) | |
| Hylobatrachus | cowanii (Boulenger, 1882) | 2n = 26 12m 1t; AN = 50 | Ag-NOR [6], CB, F | (6) |
| Hylobatrachus | lugubris (Duméril, 1853) | 2n = 26 9m 3sm 1t; AN = 50 | (2) | |
| Mantidactylus | guttulatus (Boulenger, 1881) | 2n = 26 11m 2sm; AN = 52 | (2) | |
| Ochthomantis | propefemoralis (Boulenger, 1882) | 2n = 26 9m 3sm 1t; AN = 50 | (2) | |
| Spinomantis | (2) | |||
| aglavei (Methuen et Hewitt, 1913) | 2n = 24 9m 3sm; AN = 48 | (2) | ||
| prope aglavei “North” | 2n = 24 10m 2sm; AN = 48 | Ag-NOR [7], CB, F | (6) | |
| peraccae (Boulenger, 1896) | 2n = 26 7m 6sm; AN = 48 | (2) | ||
| phantasticus | 2n = 26 13m; AN = 52 | (6) | ||
| sp. Ca3 | 2n = 26 12m 1sm: AN = 52 | Ag-NOR [6], CB, F | (6) |
Molecular taxonomic identification
DNA was extracted from cell suspensions following Sambrook (1989). A 3’ fragment of ca. 550 bp of the mitochondrial 16S rRNA gene was amplified using the primer pair 16Sa (CGCCTGTTTATCAAAAACAT) and 16Sb (CCGGTCTGAAACTCAGATCAGT) (Palumbi et al. 1991). This marker proved to be suitable for amphibian identification (Vences et al. 2005) and has been widely used for Malagasy amphibians (e.g. Vieites et al. 2009; Rosa et al. 2012; Crottini et al. 2011b, 2014; Penny et al. 2017). Amplification conditions were: initial denaturation at 94 °C for 5 min, 36 cycles at 94 °C for 30 s, 50 °C for 45s and 72 °C for 45 s, followed by a final step at 72 °C for 7 min. Amplicons were sequenced on an automated sequencer ABI 377 (Applied Biosystems, Foster City, CA, USA) using BigDye Terminator 3.1 (ABI). Chromatograms were checked and edited using Chromas Lite 2.6.6 and BioEdit 7.2.6.1 (Hall 1999). All newly determined sequences were deposited in GenBank (accession numbers: OL830846–OL830858). For taxonomic attribution we compared newly generated sequences with a curated database of reference sequences of the 3’ terminus of the 16S gene for all lineages of Malagasy mantellid frogs (Cocca 2020). Taxonomic attribution was performed using a local BLAST analysis against this reference database.
Chromosomal analysis
Cell suspensions were obtained from tissue samples as described in Mezzasalma et al. (2013). In brief, tissues were incubated for 30 min in hypotonic solution (KCl 0.075 M + sodium citrate 0.5%, 1:1) and fixed for 15 min in methanol-acetic acid, 3:1. Fixed tissues were stored at 4 °C and dissociated manually on a steel sieve. Chromosomes were obtained using the air-drying method and stained with conventional colorations (5% Giemsa solution at pH 7), Ag-NOR staining (Howell and Black 1980), C-banding according to Sumner (1972) and sequential C-banding + Fluorochromes (CMA+DAPI) following Mezzasalma et al. (2015). Ag-NOR and C-banding staining were not performed on M. sp. Ca11 and S.phantasticus, because quantity and quality of metaphase plates were not adequate for additional staining methods. Karyotype reconstruction was performed using at least five plates per sample.
Results
Molecular taxonomic identification
The selected 16S fragment was successfully amplified and sequenced from all analysed samples. All newly generated sequences showed identity scores > 97% with homologous sequences available in the mantellid frogs database generated in Cocca (2020). We followed the nomenclature used in Vieites et al. (2009), updated in Perl et al. (2014) (see Table 1).
Cytogenetic analysis
The studied specimen of Gephyromantisstriatus, Mantidactylus (Brygoomantis) alutus and Spinomantispropeaglavei “North” have a karyotype of 2n = 24 chromosomes, with the first six pairs distinctively larger than the other six pairs (Fig. 1; Table 3). In G.striatus, the pairs 1–6, 8 are biarmed while the other pairs are telocentric, with the pair 10 bearing the NOR loci (Fig. 1A; Table 3). In M. (Brygoomantis) alutus and S.propeaglavei “North” all pairs are biarmed and NOR loci were detected on the 6th and 7th pair (Fig. 1B, C), respectively.
Figure 1.

Giemsa stained karyotypes of AGephyromantisstriatus (FN 7645) BMantidactylus (Brygoomantis) alutus (FN 7945) CSpimomantispropeaglavei “North” (FN 7543) DGephyromantis sp. Ca19 (FN 7630) EMantidactylus (Chonomantis) sp. Ca11 (FN 7545) FMantidactylus (Hylobatrachus) cowanii (FAZC 11370) GSpinomantis sp. Ca3 (FN 7567) and HSpinomantisphantasticus (FG/MV 2002-970). Insets represent NOR-bearing pairs stained with Giemsa (down in the insets) and Ag-NOR method (up in the insets).
Table 3.
Chromosome morphometric parameters of the study species. LR%= % Relative Length (length of a chromosome/total chromosome length*100); CI = centromeric index (ratio between short arm/chromosome length*100). Sh = chromosome shape (m = metacentric; sm = submetacentric; t = telocentric).
| Sp. | G.striatus | M.alutus | S.propeaglavei | G. sp. Ca19 | M. sp Ca11 | M.cowanii | S. sp. Ca11 | S.phantasticus |
|---|---|---|---|---|---|---|---|---|
| Chr. | LR%-CI | LR%-CI | LR%-CI | LR%-CI | LR%-CI | LR%-CI | LR%-CI | LR%-CI |
| (sh) | (sh) | (sh) | (sh) | (sh) | (sh) | (sh) | (sh) | |
| 1 | 16.8–41.6 | 15.1–44.0 | 16.9–40.7 | 15.0–46.3 | 12.3–39.3 | 18.6–48.8 | 16.1–37.8 | 16.2–38.5 |
| (m) | (m) | (m) | (m) | (m) | (m) | (m) | (m) | |
| 2 | 12.7–36.9 | 11.8–48.5 | 14.0–32.0 | 13.7–35.6 | 12.0–34.9 | 12.9–42.3 | 14.2–42.8 | 13.8–30.9 |
| (m) | (m) | (sm) | (sm) | (sm) | (m) | (m) | (sm) | |
| 3 | 11.8–36.7 | 11.6–34.1 | 12.1–26.0 | 12.4–40.8 | 11.2–43.9 | 12.8–37.2 | 12.4–38.2 | 11.5–34.8 |
| (sm) | (sm) | (sm) | (m) | (m) | (sm) | (m) | (sm) | |
| 4 | 10.9–39.0 | 10.6–41.1 | 11.9–34.3 | 11.3–42.8 | 11.1–38.4 | 11.3–40.0 | 12.1–30.6 | 11.4–38.5 |
| (m) | (m) | (sm) | (m) | (m) | (m) | (sm) | (m) | |
| 5 | 10.2–45.2 | 10.2–44.6 | 9.7–44.7 | 10.6–36.1 | 10.0–41.7 | 19.2–44.8 | 9.1–36.0 | 10.4–35.1 |
| (m) | (m) | (m) | (sm) | (m) | (m) | (sm) | (sm) | |
| 6 | 9.7–48.7 | 10.1–48.2 | 9.7–42.6 | 6.4–31.1 | 6.2–44.7 | 5.3–47.3 | 5.5–38.2 | 6.2–33.2 |
| (m) | (m) | (m) | (sm) | (m) | (m) | (m) | (sm) | |
| 7 | 6.0–0 | 5.9–49.0 | 4.5–33.0 | 5.0–40.1 | 6.1–46.2 | 5.3–49.3 | 5.5–38.7 | 6.2–42.9 |
| (t) | (m) | (m) | (m) | (m) | (m) | (m) | (m) | |
| 8 | 5.6–39.0 | 5.9–41.4 | 4.1–47.0 | 4.8–29.3 | 6.1–41.0 | 4.8–49.6 | 5.1–39.8 | 5.9–44.5 |
| (m) | (m) | (m) | (sm) | (m) | (m) | (m) | (m) | |
| 9 | 5.4–0 | 5.8–45.8 | 3.9–47.0 | 4.4–48.8 | 5.9–43.8 | 4.4–34.4 | 4.9–43.9 | 4.4–48.8 |
| (t) | (m) | (m) | (m) | (m) | (sm) | (m) | (m) | |
| 10 | 4.6–0 | 4.9–43.0 | 3.5–39.3 | 4.3–42.9 | 5.5–0 | 4.3–41.7 | 4.2–44.1 | 3.8–48.8 |
| (t) | (m) | (m) | (m) | (t) | (m) | (m) | (m) | |
| 11 | 3.4–0 | 4.1–45.0 | 3.3–49.0 | 4.3–37.4 | 5.5–47.5 | 4.2–40.8 | 3.6–41.7 | 3.7–44.1 |
| (t) | (m) | (m) | (sm) | (m) | (m) | (m) | (m) | |
| 12 | 2.9–0 | 4.0–46.3 | 3.1–47.4 | 4.2–37.4 | 4.2–0 | 4.0–0 | 3.4–38.0 | 3.5–49.6 |
| (t) | (m) | (m) | (sm) | (t) | (t) | (m) | (m) | |
| 13 | 3.6–43.5 | 4.1–42.6 | 3.8–38.2 | 3.2–46.1 | 3.0–43.8 | |||
| (m) | (m) | (m) | (m) | (m) |
The samples of the other five species (G. sp. Ca19, M. (Chonomantis) sp. Ca11, M. (Hylobatrachus) cowanii, S.phantasticus and S. sp. Ca3) presented a karyotype of 2n = 26 chromosomes, with the first five pairs distinctively larger than the remaining eight pairs (Fig. 1D–H). In these species, all chromosome pairs resulted biarmed, with the exception of M.cowanii and of M. (Chonomantis) sp. Ca11, whose karyotype showed one (pair 12) and two pairs (10 and 12) composed of telocentric elements, respectively (Fig. 1E, F). The sixth pair is the NOR bearing one in G. sp. Ca19, M.cowanii and S. sp. Ca3 (Fig. 1D, F, G).
In G.striatus, NOR associated heterochromatin was C-banding positive (CMA + and DAPI -) and tiny centromeric C-bands were present on some chromosome pairs (Fig. 2A, A’, A”). Mantidactylusalutus and Spimomantispropeaglavei “North” showed centromeric and telomeric C-bands and NOR associated heterochromatin which were positive to CMA and DAPI negative (Fig. 2B, B’, B” and C, C’, C”). Mantidactylus (Brygoomantis) alutus also presented an additional bright centromeric band on the chromosomes of pair nine. Gephyromantis sp. Ca19 showed centromeric and telomeric C- bands, which were CMA and DAPI positive (Fig. 2D, D’, D”). Spinomantis sp. Ca3 showed solid telomeric C-bands and NOR associated heterochromatin, which resulted CMA positive and DAPI negative (Fig. 2E, E’, E”). Mantidactylus (Hylobatrachus) cowanii had centromeric C-bands on all chromosomes, which were CMA and DAPI negative (Fig. 2F, F’, F”). No heteromorphic or completely heterochromatic chromosome were found in any of the studied samples.
Figure 2.
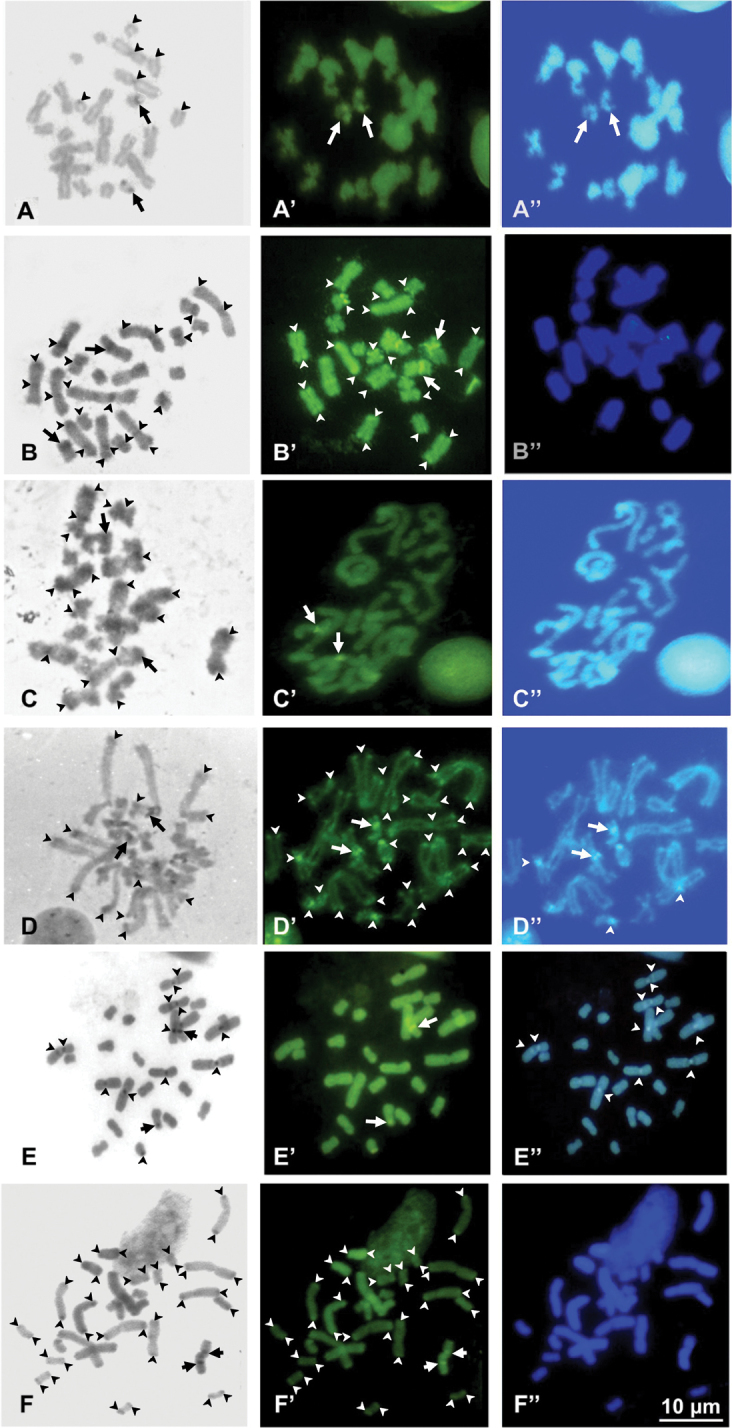
Metaphase plates of Gephyromantisstriatus (A, A’, A”), Mantidactylus (Brygoomantis) alutus (B, B’, B”), Spinomantispropeaglavei “North” (C, C’, C”), Gephyromantis sp. Ca19 (D, D’, D”), Spinomantis sp. Ca3 (E, E’, E”) and Mantidactylus (Hylobatrachus) cowanii (F, F’, F”) stained with C-banding + Giemsa (A–F), + CMA (A’–F’) + DAPI (A”–F”). Arrows point at NORs while arrowheads highlight other heterochromatin blocks.
Discussion
We here provide new karyological data on eight frog species belonging to the subfamily Mantellinae and discuss the available chromosome data on this subfamily to provide a first comprehensive assessment of its karyological diversity.
Available data on representatives of the other two Mantellidae subfamilies (Boophininae and Laliostominae) highlight the occurrence of a conserved karyotype structure in terms of chromosome number and morphology. In particular, the first karyological studies by Blommers-Schlössers (1978) on 12 species of Boophis (Boophininae) and on Aglyptodactylusmadagascariensis (Duméril, 1853) (Laliostominae) revealed a conserved karyotype of 2n = 26 with all biarmed chromosomes.
Following studies by Aprea et al. (1998, 2004) expanded the knowledge on the karyological uniformity to the position of NORs loci, invariably on the sixth chromosome pair both in Boophis and A.madagascariensis, but evidenced different patterns of heterochromatin composition and distribution. Similar karyological characters were described also in different species of the genus Mantella (belonging to the subfamily Mantellinae), all showing a karyotype of 2n = 26 with all biarmed chromosomes (Blommers-Schlössers 1978; Odierna et al. 2001). A karyotype of 2n = 26 with all biarmed elements should thus be considered the primitive condition in the whole family Mantellidae, as it is highly conserved in all subfamilies, genera and most subgenera (see Blommers-Schlösser 1978; Aprea et al. 1998, 2004; Odierna et al. 2001, see Table 2). Nevertheless, species of other genera of the subfamily Mantellinae show a wider karyological variability, both concerning chromosomes number, morphology, localizations of NORs loci and heterochromatin composition and distribution (Blommers-Schlösser 1978; Odierna et al. 2001; present study) (see also Table 2).
Concerning the variability of the chromosome number, a 2n = 26 karyotype is still the most common chromosomal configuration, but karyotypes with a reduced chromosome complement (2n = 24) have been documented in 9 species of three different genera (6 species of Mantidactylus (subgenus Brygoomantis), 2 Spinomantis and Gephyromantisstriatus) (See Fig. 1 and Table 2). Furthermore, while the 2n = 26 configuration occurs in all three subfamilies of the family Mantellidae (Mantellinae, Boophinae and Laliostominae) (e.g. Aprea et al. 1998, 2004; present study), karyotypes with 2n = 24 seem to occur in just a few phylogenetically lineages (genus Gephyromantis, Mantidactylus and Spinomantis), where the 2n = 26 configuration is also present (Blommers-Schlösser 1978; present study). In turn, the subfamily Boophinae, with all the species showing a 2n = 26 karyotype (Aprea et al. 1998, 2004), has been depicted as a basal group in the Mantellidae radiation (see e.g. Wollenberg et al. 2011). These evidences suggest that a reduction of the chromosome number from 2n = 26 to 2n = 24 occurred repeatedly and independently in different lineages of the subfamily Mantellinae, probably involving chromosome inversions and a fusion (translocation) between two elements of the smallest pairs (6–13), giving rise to an additional large (6th) chromosome pair in several species (e.g. G.striatus, M. (Brygoomantis) alutus, and S.propeaglavei “North”) (Fig. 3; Table 3). Interestingly, a similar reduction of the chromosome number driven by tandem fusions (from 2n = 26 to 24) has been documented also in the family Ranidae (Miura et al. 1995).
Figure 3.
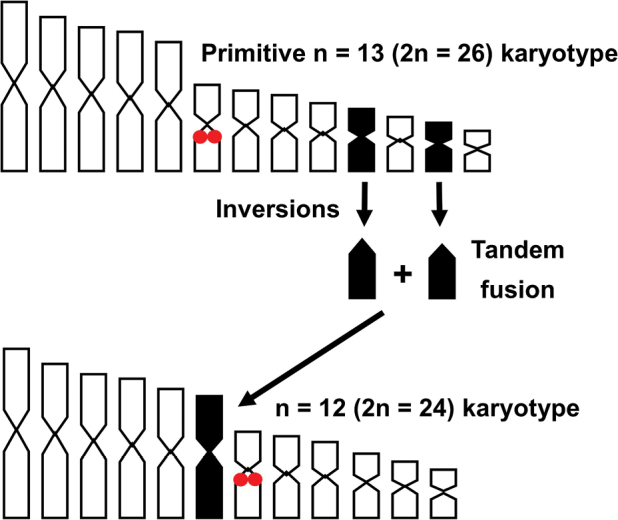
Hypothesized general model of chromosome reduction in Mantellinae from n = 13 (2n = 26) to n = 12 (2n = 24) by means of chromosome fusions. Red dots highlight the NOR bearing chromosome.
Other than tandem fusions, chromosome inversions of primitive biarmed elements also had a significant role in the morphological chromosome diversity observed in mantelline frogs. These mechanisms generated a variable number of telocentric elements in different evolutionary lineages (see Figs 1, 3 and Table 2).
Considering the position of the loci of NORs, our results and available literature data (Aprea et al. 1998, 2004; Odierna et al. 2001; Andreone et al. 2003), show that NORs occurrence on the sixth chromosome pair can be considered a primitive state, as it is described for all analysed species belonging to the genus Boophis, A.madascariensis and most Gephyromantis, Mantidactylus (subgenus Brygoomantis), and Spinomantis. On the other hand, a derivate configuration of NOR loci seems to have emerged multiple times in distinct lineages. The different positions of NOR loci in mantelline frogs suggest that these elements were also differently involved in the hypothesized chromosome fusions from 2n = 26 to 2n = 24, providing further support to multiple, independent rearrangements leading to similar karyotype configurations. In fact, while in M. (Brygoomantis) alutus the sixth large chromosome pair likely derived from a fusion involving the primitive NOR bearing pair and another smaller pair, in G.striatus and S.propeaglavei “North” the pair 6 does not include NOR loci, which are found on the 7th and 10th chromosome pair, respectively (see Fig. 1). In other species of Gephyromantis, Blommersia, Guibemantis and Mantella the relocation of NORs involved different pairs (1st, 2nd, 8th, 9th, 10th or 11th) (Odierna et al. 2001; Andreone et al. 2003; this study). It should be noted that Ag-NOR staining only evidences active NORs, and the existence of different inactive sites in the karyotypes of the studied species cannot be excluded based only on this analysis. However, we found correspondence in NOR location using both Ag-NOR and C-banding + CMA (in Figs 1, 2), which also has the power to uncover rDNA clusters (Schmid 1982; Zaleśna et al. 2017).
Various mechanisms may be responsible for NOR relocation, such as cryptic structural rearrangements, minute insertions, reintegration of rDNA genes amplified during ovogonial auxocytosis or the activation of silent sites (Nardi et al. 1977; Schmid 1978; King 1980; Mahony and Robinson 1986; Schmid and Guttenbach 1988; Mezzasalma et al. 2018). These mechanisms may be independent to other rearrangements, despite the resulting change in the configuration of NORs is a significant indicator of lineage divergence at different taxonomic level (e.g. Pardo et al. 2001; Mezzasalma et al. 2015, 2018, 2021).
Sequential C-banding did not evidence the occurrence of any sex-specific, largely heterochromatic chromosomes (generally related to differentiated heterogametic sex chromosomes, a condition not yet documented in the family Mantellidae), B chromosomes, or interchromosomal rearrangements leading to heteromorphic autosome pairs (e.g. Mezzasalma et al. 2014, 2016, 2017b; Sidhom et al. 2020). Nevertheless, C-banding showed a heterogeneous heterochromatin distribution in Mantellidae (see also Aprea et al. 1998, 2004; Odierna et al. 2001; Andreone et al. 2003), highlighting the occurrence of species-specific banding patterns. For example, G.striatus and M. (Hylobatrachus) cowanii show different amount and location of C-banding positive heterochromatin in comparison with closely related species with the same chromosome number and similar morphology (e.g. G. sp. Ca19 and M. (Brygoomantis) alutus). Interspecific variations in heterochromatin are generally due to different levels of amplification of highly repetitive DNA (Charlesworth et al. 1994). These differences mostly occurred without modifications of the chromosome morphology in Mantellidae (see also Aprea et al. 1998, 2004; Odierna et al. 2001; Andreone et al. 2003), probably by means of symmetrical addition/deletion of heterochromatin. The occurrence of distinctive species-specific banding patterns may be useful in evolutionary cytogenetic and cytotaxonomic studies in the subfamily, but comprehensive comparative analyses would benefit from more banding data on species of different genera and subgenera.
Finally, we also highlight the importance of a preliminary molecular taxonomic identification of mantellid frogs for a consistent karyotype attribution, and that future cytogenetic studies should focus on LaliostomaGlaw et al., 1998, Wakea Glaw et Vences, 2006, Boehmantis Glaw et Vences, 2006 and Tsingymantis Glaw et al., 2006, as well as on different undersampled genera and subgenera.
Conclusions
We provide new chromosomal data on eight species belonging to the subfamily Mantellinae, advancing the knowledge on their karyotype diversity, and suggesting that a reduction in the chromosome number and the relocation of NORs loci occurred repeatedly and independently in different genera of this subfamily. We hypothesize a karyotype of 2n = 26 with all biarmed elements and loci of NORs on the 6th chromosome pair as the ancestral state in the whole family Mantellidae and propose a model for the reduction of the chromosome number from 2n = 26 to 2n = 24 by means of tandem fusions.
Acknowledgements
We are grateful to Malagasy authorities for granting research and export permits. Numerous colleagues helped us in the field: Gennaro Aprea, Frank Glaw, Miguel Vences. Portuguese National Funds through FCT (Foundation for Science and Technology) support the 2020.00823.CEECIND/CP1601/CT0003 contract to AC and the ICETA 2020-37 contract to MM. This work was supported by National Funds through FCT under the PTDC/BIA-EVL/31254/2017 project.
Citation
Mezzasalma M, Andreone F, Odierna G, Guarino FM, Crottini A (2022) Comparative cytogenetics on eight Malagasy Mantellinae (Anura: Mantellidae) and a synthesis of the karyological data on the subfamily. Comparative Cytogenetics 16(1): 1–17. https://doi.org/10.3897/compcytogen.v16.i1.76260
ORCID
Marcello Mezzasalma https://orcid.org/0000-0002-7246-9831
Franco Andreone https://orcid.org/0000-0001-9809-5818
Fabio Maria Guarino https://orcid.org/0000-0002-1511-7792
Angelica Crottini https://orcid.org/0000-0002-8505-3050
References
- AmphibiaWeb (2021) University of California, Berkeley, CA, USA. https://amphibiaweb.org [Last Accessed 01 March 2021]
- Andreone F, Aprea G, Vences M, Odierna G. (2003) A new frog of the genus Mantidactylus from the rainforests of north-eastern Madagascar, and its karyological affinities. Amphibia-Reptilia 24(3): 285–303. 10.1163/156853803322440763 [DOI] [Google Scholar]
- Andreone F, Rosa GM, Noël J, Crottini A, Vences M, Raxworthy CJ. (2010) Living within dry palm leaves: the discovery of an unknown Blommersia (Mantellidae: Anura) reveals a new reproductive strategy in the amphibians of Madagascar. Naturwissenschaften 97(6): 525–543. 10.1007/s00114-010-0667-x [DOI] [PubMed] [Google Scholar]
- Aprea G, Andreone F, Capriglione T, Odierna G. (1998) Chromosome banding in several Malagasy anuran species belonging to the genera Aglyptodactylus, Boophis and Mantidactylus. Abstracts of the 9th Ordinary General Meeting of the Societas Herpetologica Europaea. Chambéry, August 25–29, 1998, Chambéry, 21–21.
- Aprea G, Andreone F, Capriglione T, Odierna G, Vences M. (2004) Evidence for a remarkable stasis of chromosome evolution in Malagasy treefrogs (Boophis, Mantellidae). Italian Journal of Zoology 71(2): 237–244. 10.1080/11250000409356641 [DOI] [Google Scholar]
- Blommers-Schlösser RMA. (1978) Cytotaxonomy of the Ranidae, Rhacophoridae, Hyperoliidae (Anura) from Madagascar with a note on the karyotype of two amphibians of the Seychelles. Genetica 48(1): 23–40. 10.1371/journal.pbio.0060118 [DOI] [Google Scholar]
- Charlesworth B, Sniegowski P, Stephan W. (1994) The evolutionary dynamics of repetitive DNA in eukaryotes. Nature 371: 215–220. 10.1038/371215a0 [DOI] [PubMed] [Google Scholar]
- Cocca W. (2020) Studying the processes of species diversification using the adaptive radiation of the mantellid frogs of Madagascar (Anura: Mantellidae) as a model system. Ph.D. Dissertation, Porto, Portugal, University of Porto. https://repositorio-aberto.up.pt/handle/10216/130002?locale=en
- Cocca W, Andreone F, Belluardo F, Rosa GM, Randrianirina JE, Glaw F, Crottini A. (2020) Resolving a taxonomic and nomenclatural puzzle in mantellid frogs: synonymization of Gephyromantisazzurrae with G.corvus, and description of Gephyromantiskintana sp. nov. from the Isalo Massif, western Madagascar. ZooKeys 951: 133–157. 10.3897/zookeys.951.51129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crottini A, Glaw F, Casiraghi M, Jenkins RKB, Mercurio V, Randrianantoandro JC, Randrianirina JE, Andreone F. (2011a) A new Gephyromantis (Phylacomantis) frog species from the pinnacle karst of Bemaraha, western Madagascar. ZooKeys 81: 51–71. 10.3897/zookeys.81.1111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crottini A, Barbuto M, Casiraghi M. (2011b) A rapid amphibian survey at Itremo-Ambatofinandrahana, central Madagascar, with confirmed absence of chytrid fungus and recommendations for future monitoring activities. North-Western Journal of Zoology 7(2): 346–351. http://biozoojournals.ro/nwjz/content/v7n2/nwjz.111214.Crottini.pdf [Google Scholar]
- Crottini A, Bollen A, Weldon C, Dalton DL, Kotze A, Noël J, Iambana B, Andreone F. (2014) Amphibian survey and current absence of Batrachochytriumdendrobatidis (Bd) in Ivoloina Park, Toamasina (eastern Madagascar). African Journal of Herpetology 63(1): 70–78. 10.1080/21564574.2013.833994 [DOI] [Google Scholar]
- Frost DR. (2021) Amphibian species of the world: an online reference. Version 6.1. Electronic Database accessible at https://amphibiansoftheworld.amnh.org/index.php. American Museum of Natural History, New York, USA. 10.5531/db.vz.0001 [DOI]
- Ganzhorn JU, Lowry II PP, Schatz GE, Somme S. (2001) The biodiversity of Madagascar: one of the world’s hottest hotspots on its way out. Oryx 35(4): 346–348. 10.1046/j.1365-3008.2001.00201.x [DOI] [Google Scholar]
- Glaw F, Vences M. (2006) Phylogeny and genus-level classification of mantellid frogs (Amphibia, Anura). Organisms, Diversity and Evolution 6(3): 236–253. 10.1016/j.ode.2005.12.001 [DOI] [Google Scholar]
- Glaw F, Vences M. (2007) A Field Guide to the Amphibians and Reptiles of Madagascar. 3rd edn. Vences M and Glaw F, Verlags GbR, Köln, 496 pp. [Google Scholar]
- Glaw F, Vences M, Böhme W. (1998) Systematic revision of the genus Aglyptodactylus Boulenger, 1919 (Amphibia: Ranidae), and analysis of its phylogenetic relationships to other Madagascan ranid genera (Tomopterna, Boophis, Mantidactylus, and Mantella). Journal of Zoological Systematics and Evolutionary Research 36(1–2): 17–37. 10.1111/j.1439-0469.1998.tb00775.x [DOI] [Google Scholar]
- Howell WM, Black DA. (1980) Controlled silver staining of nucleolus organizer regions with a protective colloidal developer: a 1 step method. Experientia 36(8): 1014–1015. 10.1007/BF01953855 [DOI] [PubMed] [Google Scholar]
- Kaffenberger N, Wollenberg KC, Köhler J, Glaw F, Vieites DR, Vences M. (2012) Molecular phylogeny and biogeography of Malagasy frogs of the genus Gephyromantis. Molecular Phylogenetics and Evolution 62(1): 555–560. 10.1016/j.ympev.2011.09.023 [DOI] [PubMed] [Google Scholar]
- King M. (1980) C-banding studies on Australian hylid frogs: Secondary constriction structure and the concept of euchromatin transformation. Chromosoma 80(2): 191–217. 10.1007/BF00286300 [DOI] [Google Scholar]
- Mahony MJ, Robinson ES. (1986) Nucleolar organiser region (NOR) location in karyotypes of Australian ground frogs (Family Myobatrachidae). Genetica 68(2): 119–127. 10.1007/BF02424409 [DOI] [Google Scholar]
- Mezzasalma M, Andreone F, Aprea G, Glaw F, Odierna G, Guarino FM. (2017a) Molecular phylogeny, biogeography and chromosome evolution of Malagasy dwarf geckos of the genus Lygodactylus (Squamata, Gekkonidae). Zoologica Scripta 46(1): 42–54. 10.1111/zsc.12188 [DOI] [Google Scholar]
- Mezzasalma M, Andreone F, Aprea G, Glaw F, Odierna G, Guarino FM. (2017b) When can chromosomes drive speciation? The peculiar case of the Malagasy tomato frogs (genus Dyscophus). Zoologischer Anzeiger 268: 41–46. 10.1016/j.jcz.2017.04.006 [DOI] [Google Scholar]
- Mezzasalma M, Andreone F, Branch WR, Glaw F, Guarino FM, Nagy ZT, Odierna G, Aprea G. (2014) Chromosome evolution in pseudoxyrhophiine snakes from Madagascar: a wide range of karyotypic variability. Biological Journal of Linnean Society London 112(3): 450–460. 10.1111/bij.12280 [DOI] [Google Scholar]
- Mezzasalma M, Andreone F, Glaw F, Petraccioli A, Odierna G, Guarino FM. (2016) A karyological study of three typhlopid species with some inferences on chromosome evolution in blindsnakes (Scolecophidia). Zoologischer Anzeiger 264: 34–40. 10.1016/j.jcz.2016.07.001 [DOI] [Google Scholar]
- Mezzasalma M, Andreone F, Glaw F, Odierna G, Petraccioli A, Guarino FM. (2018) Chromosome aneupolyploidy in an endemic Malagasy gecko (Gekkonidae: Geckolepis). Salamandra 54(1): 56–62. http://www.salamandra-journal.com [Google Scholar]
- Mezzasalma M, Andreone F, Glaw F, Guarino FM, Odierna G, Petraccioli A, Picariello O. (2019) Changes in heterochromatin content and ancient chromosome fusion in the endemic Malagasy boid snakes Sanzinia and Acrantophis (Squamata: Serpentes). Salamandra 55(2): 140–144. http://www.salamandra-journal.com [Google Scholar]
- Mezzasalma M, Glaw F, Odierna G, Petraccioli A, Guarino FM. (2015) Karyological analyses of Pseudhymenochirusmerlini and Hymenochirusboettgeri provide new insights into the chromosome evolution in the anuran family Pipidae. Zoologischer Anzeiger 258: 47–53. 10.1016/j.jcz.2015.07.001 [DOI] [Google Scholar]
- Mezzasalma M, Guarino FM, Aprea G, Petraccioli A, Crottini A, Odierna G. (2013) Karyological evidence for diversification of Italian slow worm populations (Squamata, Anguidae). Comparative Cytogenetics 7(3): 217–227. 10.3897/compcytogen.v7i3.5398 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mezzasalma M, Odierna G, Petraccioli M, Veith M, Guarino FM. (2021) Karyological diversification in the genus Lyciasalamandra (Urodela: Salamandridae). Animals 11(6): 1709. 10.3390/ani11061709 [DOI] [PMC free article] [PubMed]
- Miura I, Nishioka M, Borkin LJ, Wu Z. (1995) The origin of the brown frogs with 2n = 24 chromosomes. Experientia 51(2): 179–188. 10.1007/BF01929366 [DOI] [Google Scholar]
- Morescalchi A. (1967) Le relazioni tra il cariotipo di anuri diplasioceli: 1. I1 corredo cromosomico di alcuni Ranidae. Caryologia 20(1): 65–85. 10.1080/00087114.1967.10796248 [DOI] [Google Scholar]
- Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J. (2000) Biodiversity hot-spots for conservation priorities. Nature 403(6772): 853–858. 10.1038/35002501 [DOI] [PubMed] [Google Scholar]
- Nardi I, Barsacchi-Pilone G, Batistoni R, Andronico F. (1977) Chromosome location of the ribosomal RNA genes in Triturusvulgarismeridionalis (Amphibia, Urodela). II. Intraspecific variability in number and position of the chromosome for 18S and 28S ribosomal RNA. Chromosoma 64(1): 67–84. 10.1007/BF00292889 [DOI] [PubMed] [Google Scholar]
- Odierna G, Vences M, Aprea G, Lötters S, Andreone F. (2001) Chromosome data for Malagasy poison frogs (Amphibia: Ranidae: Mantella) and their bearing on taxonomy and phylogeny. Zoological Science 18(4): 505–514. 10.2108/zsj.18.505 [DOI] [Google Scholar]
- Palumbi SR, Martin A, Romano S, Mcmillan WO, Stice L, Grabowski G. (1991) The Simple Fool’s Guide to PCR, Version 2.0. Privately published, University of Hawaii.
- Pardo BG, Bouza C, Castro J, Martínez P, Sánchez L. (2001) Localization of ribosomal genes in Pleuronectiformes using Ag-, CMA3-banding and in situ hybridization. Heredity 86(5): 531–536. 10.1046/j.1365-2540.2001.00802.x [DOI] [PubMed] [Google Scholar]
- Penny SG, Crottini A, Andreone F, Bellati A, Rakotozafy LMS, Holderied MW, Schwitzer C, Rosa GM. (2017) Combining old and new evidence to increase the known biodiversity value of the Sahamalaza Peninsula, Northwest Madagascar. Contribution to Zoology 86(4): 273–296. 10.1163/18759866-08604002 [DOI] [Google Scholar]
- Perl RGB, Nagy ZT, Sonet G, Glaw F, Wollenberg KC, Vences M. (2014) DNA barcoding Madagascar’s amphibian fauna. Amphibia-Reptilia 35(2): 197–206. 10.1163/15685381-00002942 [DOI] [Google Scholar]
- Pintak T, Vences M, Glaw F, Böhme W. (1998) Comparative chromosome morphology of Malagasy poison frogs (Amphibia: Ranidae: Mantella). Folia Zoologica 47(3): 197–204. [Google Scholar]
- Richards CM, Nussbaum RA, Raxworthy CJ. (2000) Phylogenetic relationships within the Madagascan boophids and mantellids as elucidated by mitochondrial ribosomal genes. African Journal of Herpetology 49(1): 23–32. 10.1080/21564574.2000.9650013 [DOI] [Google Scholar]
- Rosa GM, Andreone F, Crottini A, Hauswaldt JS, Noël J, Rabibisoa NH, Randriambahiniarime MO, Rebelo R, Raxworthy CJ. (2012) The amphibians of the relict Betampona low-elevation rainforest, eastern Madagascar: an application of the integrative taxonomy approach to biodiversity assessments. Biodiversity and Conservation 21(6): 1531–1559. 10.1007/s10531-012-0262-x [DOI] [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. (1989) Molecular Cloning: A Laboratory Manual, 2nd edn. Cold Spring Harbor Lab Press, New York.
- Schmid M. (1978) Chromosome banding in Amphibia. II. Constitutive heterochromatin and nucleolar organizer regions in Ranidae, Microhylidae and Rhacophoridae. Chromosoma 77: 83–103. 10.1007/BF00287145 [DOI] [Google Scholar]
- Schmid M. (1982) Chromosome Banding in Amphibia VII. Analysis of the Structure and Variability of NORs in Anura. Chromosoma 87: 327–344. 10.1007/BF00327634 [DOI] [Google Scholar]
- Schmid M, Guttenbach M. (1988) Evolutionary diversity of reverse (R) fluorescent chromosome bands in vertebrates. Chromosoma 97(2): 101–114. 10.1007/BF00327367 [DOI] [PubMed] [Google Scholar]
- Sidhom M, Said K, Chatti N, Guarino FM, Odierna G, Petraccioli A, Picariello O, Mezzasalma M. (2020) Karyological characterization of the common chameleon (Chamaeleochamaeleon) provides insights on the evolution and diversification of sex chromosomes in Chamaeleonidae. Zoology 141: 125738. 10.1016/j.zool.2019.125738 [DOI] [PubMed]
- Sumner AT. (1972) A simple technique for demonstrating centromeric heterochromatin. Experimental Cell Research 75(1): 304–306. 10.1016/0014-4827(72)90558-7 [DOI] [PubMed] [Google Scholar]
- Vences M, Glaw F. (2005) A new species of Mantidactylus from the East Coast of Madagascar and its molecular phylogenetic relationships within the subgenus Guibemantis. Herpetological Journal 15(1): 37–44. [Google Scholar]
- Vences M, Glaw F, Mausfeld P, Böhme W. (1998) Comparative osteology of Malagasy poison frogs of the genus Mantella (Amphibia: Ranidae: Mantellinae). Bonner zoologische Beiträge 48: 205–215. https://www.biodiversitylibrary.org/page/44945242#page/221/mode/1up [Google Scholar]
- Vences M, Thomas M, van der Meijden A, Chiari Y, Vieites DR. (2005) Comparative performance of the 16S rRNA gene in DNA barcoding of amphibians. Frontiers in Zoology 2(1): 5. 10.1186/1742-9994-2-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vences M, Wahl-Boos G, Hoegg S, Glaw F, Spinelli Oliveira E, Meyer A, Perry S. (2007) Molecular systematics of mantelline frogs from Madagascar and the evolution of their femoral glands. Biological Journal of the Linnean Society 92(3): 529–539. 10.1111/j.1095-8312.2007.00859.x [DOI] [Google Scholar]
- Vences M, Wollenberg KC, Vieites DR, Lees DC. (2009) Madagascar as a model region of species diversification. Trends in Ecology and Evolution 24(8): 456–465. 10.1016/j.tree.2009.03.011 [DOI] [PubMed] [Google Scholar]
- Vieites DR, Wollenberg KC, Andreone F, Kohler J, Glaw F, Vences M. (2009) Vast underestimation of Madagascar’s biodiversity evidenced by an integrative amphibian inventory. Proceedings of the National Academy of Sciences of the United States of America 106(20): 8267–8272. 10.1073/pnas.0810821106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wollenberg KC, Vieites DR, Glaw F, Vences M. (2011) Speciation in little: the role of range and body size in the diversification of Malagasy mantellid frogs. BMC Evolutionary Biology 11(1): 217. 10.1186/1471-2148-11-217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaleśna A, Florek M, Rybacki M, Ogielska M. (2017) Variability of NOR patterns in European water frogs of different genome composition and ploidy level. Comparative Cytogenetics 11(2): 249–266. 10.3897/CompCytogen.v11i2.10804 [DOI] [PMC free article] [PubMed] [Google Scholar]