

Abstract
Arbuscular mycorrhizal (AM) fungi lack efficient exoenzymes to access organic nutrients directly. Nevertheless, the fungi often obtain and further channel to their host plants a significant share of nitrogen (N) and/or phosphorus from such resources, presumably via cooperation with other soil microorganisms. Because it is challenging to disentangle individual microbial players and processes in complex soil, we took a synthetic approach here to study 15N-labelled chitin (an organic N source) recycling via microbial loop in AM fungal hyphosphere. To this end, we employed a compartmented in vitro cultivation system and monoxenic culture of Rhizophagus irregularis associated with Cichorium intybus roots, various soil bacteria, and the protist Polysphondylium pallidum. We showed that upon presence of Paenibacillus sp. in its hyphosphere, the AM fungus (and associated plant roots) obtained several-fold larger quantities of N from the chitin than it did with any other bacteria, whether chitinolytic or not. Moreover, we demonstrated that adding P. pallidum to the hyphosphere with Paenibacillus sp. further increased by at least 65% the gain of N from the chitin by the AM fungus compared to the hyphosphere without protists. We thus directly demonstrate microbial interplay possibly involved in efficient organic N utilisation by AM fungal hyphae.
Subject terms: Stable isotope analysis, Biogeochemistry, Fungal ecology
Introduction
Arbuscular mycorrhizal (AM) fungi have accompanied terrestrial plants for approximately a half billion years [1], playing a particularly important role in their host plants’ acquisition of phosphorus (P) from soil in exchange for reduced carbon (C) provided by the plants [2–4]. The P is taken up by the AM fungal hyphae from the soil solution chiefly as orthophosphate [5, 6]. Efficient acquisition of P from organic sources such as phytic acid by the AM fungal hyphae has also been documented upon close cooperation between hyphae and such other soil microorganisms as the bacterium Rahnella aquatilis [7–10]. This is because the AM fungi lack genes coding for potent exoenzymes, thus preventing them from efficiently accessing organic nutrient sources in soil on their own [11, 12].
Earlier research has shown that AM fungal hyphae also take up nitrogen (N) from the soil as both ammonium and nitrate ions [13, 14]. Further, it has been demonstrated that the hyphae could obtain significant amounts of N also from such organic sources as plant litter or chitin, sometimes transferring part of this N to their host plants, even as they compete with those plants for limited soil N under other circumstances [15–20]. It long has been assumed that other soil microbes play important roles in mineralisation and/or further processing of organic N before it can be taken up by AM fungal hyphae, similarly as in the case of organic P [21–24]. Furthermore, bacterial grazers have been posited to facilitate release of N to the soil solution as free ammonium ions from the microbes they ingest [18, 25]. Direct experimental evidence of such inter-kingdom associations in utilisation of organic N by AM fungal hyphae and their associated mycorrhizal plants has nevertheless not been reported [26].
The aim of this research was to establish an experimental in vitro system to study interactions between AM fungal hyphae and other microbes in their hyphosphere. Using such experimental system and different chemical forms of N, we quantified rates of N transfer from a root-free zone into the roots while controlling for passive N diffusion. In so doing, we addressed three hypotheses:
The AM fungal hyphae alone cannot take up significant amounts of N from an organic N source such as chitin, in contrast to N administered as ammonium ions, even if they are capable of producing chitinases and deaminases [27].
Chitinolytic bacteria inoculated in the root-free zone will enhance hyphal access to N supplied as chitin to the same compartment.
Including protists into the root-free zone would further increase the rates of N release from the chitin and thus the amounts of N taken up by the AM fungal hyphae from that zone and eventually transferred to the roots.
We used an experimental system where Ri T-DNA transformed chicory (Cichorium intybus L.) roots were used instead of a full plant, because a green plant would require light and inevitably necessitate active cooling of the experimental system. We wanted to avoid this for logistical reasons. Moreover, some autotrophic in vitro culture systems established previously [28, 29] had required expansion of plant leaves in free air, thus imposing additional technical challenges to be overcome to prevent microbial contamination of the rhizosphere and hyphosphere zones. Such experimental systems as we employed here, which can easily be established in copious numbers due to efficient clonal propagation of roots, previously proved themselves extremely valuable for addressing such specific questions in mycorrhizal physiology and ecology as relate to trading resources between AM fungus and its plant host(s) or microbial interactions in AM fungal hyphosphere [10, 30, 31].
Materials and methods
Biological materials
For the experiments described here, we used monoxenic culture of Rhizophagus irregularis Walker & Schüßler genotype SYM5 (also known as LPA9 or BEG236) originally obtained from Asphodelus sp. rhizosphere in Greece during 1980. The culture has been maintained in vitro in association with Ri T-DNA transformed chicory roots for several years prior to the research described here [32]. To establish a non-mycorrhizal (NM) control, we used the same root culture as above without the AM fungus (i.e. NM roots). The prokaryotes used here were of various provenances (Table 1), maintained on solid lysogeny broth (1.5% agar) for at least five generations, and stored in 30% glycerol at −80 °C prior to use. Four of them exhibited chitinolytic activity when crab-shell chitin was provided as the only C and N source (Table 1 and supplementary Figs. S1–S3). Identity of the different bacterial strains was revealed by sequencing amplicons of their rRNA genes as described previously [33]. The culture of the protist Polysphondylium pallidum (Amoebozoa) was originally obtained from spruce bark compost [34] and subcultured for at least five generations using Escherichia coli co-culture on lysogeny broth agar (1.5%). Thereafter, it was inoculated separately to each of the prokaryotes (Table 1) using spores carefully recovered from sporangiophores above the surface of the agar plates in order to avoid contaminating the follow-up cultures with E. coli.
Table 1.
Identities and provenances of bacterial strains used in this study.
| Isolate ID | Bacterial culture details | Provenance | Chitinolytic activity detectedb | Sporulation of Polysphondylium observed in association with the bacteriac | Taxonomic affiliation by 16S rRNA gene fragment sequencing (sequence similarity of closest match in GenBank) |
|---|---|---|---|---|---|
| 1 | Isolate D22 | Isolated from grassland soil, Czech Republic | Yes | n.a. | Janthinobacterium sp. (100%), Oxalobacteraceae, β-proteobacteria |
| 2 | Isolate D23 | Isolated from grassland soil, Czech Republic | Yes | Yes, abundant | Janthinobacterium sp. (100%), Oxalobacteraceae, β-proteobacteria |
| 5 | Paenibacillus chitinolyticus CCM 4527 | Public culture collection, originally isolated from forest soil, Japan | Yes | Yes, abundant | Paenibacillus sp. (100%), Paenibacillaceae, Firmicutes |
| 8 | Isolate M-N | Isolated from a mycorrhizal inoculum pota | No | No | Gordonia sp. (100%), Nocardiaceae, Actinobacteria |
| 9 | Isolate M-R | Isolated from a mycorrhizal inoculum pota | No | n.a. | Rhodococcus sp. (100%), Nocardiaceae, Actinobacteria |
| 10 | Isolate NM-A | Isolated from a non-mycorrhizal inoculum pota | No | n.a. | Arthrobacter sp. (99%), Micrococcaceae, Actinobacteria |
| 12 | Isolate M-F | Isolated from a mycorrhizal inoculum pota | No | Yes | Bacillus sp. (94%), Bacillaceae 1, Firmicutes |
| 13 | Isolate M-U | Isolated from a mycorrhizal inoculum pota | No | No | Mycobacterium sp. (99%), Mycobacteriaceae, Actinobacteria |
| 15 | Isolate NM-H | Isolated from a non-mycorrhizal inoculum pota | No | Yes | Rhodococcus sp. (100%), Nocardiaceae, Actinobacteria |
| 16 | Isolate NM-O | Isolated from a non-mycorrhizal inoculum pota | No | n.a. | Nocardia sp. (100%), Nocardiaceae, Actinobacteria |
| 17 | Paenibacillus ehimensis CCM 4526 | Public culture collection, originally isolated from garden soil, Japan | Yes | Yes, abundant | Paenibacillus sp. (100%), Paenibacillaceae, Firmicutes |
15N-labelled chitin
Isotopically (15N) labelled chitin was prepared from Zygorhynchus sp. cell walls as previously [18]. Its elemental and isotopic composition as well as biochemical structure and purity were then analysed as described elsewhere [33]. Briefly, the chitin batch for Experiment 1 contained 5.8% N and 42.3% C by weight and its 15N share was 29.6 atom%. The chitin batch for Experiment 2 contained 5.3% N and 45.8% C by weight, with the N being fully isotopically labelled (>98 atom% represented by 15N).
Experimental system (microcosms)
The experimental system used here was constructed as three-compartment in vitro cultivation vessel consisting of a large, sterile Petri dish (diameter 15 cm, 2 cm height, made from polystyrene) and two smaller compartments (Fig. 1 and supplementary Fig. S4). One of the smaller compartments (the root compartment or rhizobox) was made from a lid of a small (6 cm diameter) polystyrene Petri dish with a hole for root transfer drilled into its top, and the bottom opening sealed with 42 µm nylon mesh (Silk & Progress, Brněnec, Czech Republic). The rim of the rhizobox was dipped in chloroform before pressing it against the mesh, which firmly glued the rhizobox walls to the mesh. After preparing the rhizoboxes, they were sterilised by γ-rays (>25 kGy, Bioster, Veverská Bítýška, Czech Republic). The second smaller compartment (termed here the labelling compartment) was made from the bottom of a small (6 cm diameter) Petri dish and was inserted at least 5 mm from the rhizobox. The large Petri dish was filled with 100 ml of standard modified Strullu and Romand (MSR) medium, pH 5.5 [35], containing 93 µg P and 379 µmol N, supplemented with 1% (w:v) sucrose, and solidified with 0.3% (w:v) gelling agent (Phytagel, Merck, Darmstadt, Germany). The P and N concentrations in Phytagel powder were 560 µg g−1 and 46 µmol g−1, respectively, which (together with the MSR medium) ensured luxurious P supply and rather limited N supply to the roots and AM fungi (details not shown). The sterile labelling compartment was inserted into the liquid medium freshly poured into the large Petri dish before it solidified and was held down by a sterile metal plug. The rim of the labelling compartment protruded at least 1 mm above the medium (see supplementary Fig. S4 for details). After the medium solidified, the rhizobox was placed on top of the MSR medium and the metal plug was removed from the labelling compartment. Mycorrhizal or NM roots were added to the rhizobox through the hole (supplementary Fig. S4) and incubated at 24 °C in darkness for 61 or 75 days in Experiment 1 or 2, respectively, before the roots filled the rhizobox and AM fungal hyphae (if applicable) colonised the entire volume of the MSR medium (Fig. 2). Thereafter, the labelling compartment was filled with N-free MSR medium with or without an added 15N-labelled N source and bacteria and/or protists were further added or not added as specified below. In Experiment 1, sucrose was added to both the large Petri dish volume and the labelling compartment and the N concentration in the labelling compartments was increased fourfold as compared to the rest of the microcosm, thereby establishing an N-rich patch similarly as in a previous unsterile pot experiment [18]. In Experiment 2, sucrose was omitted from the labelling compartment because we had noted possible interference between sucrose and growth of some of the bacteria. Besides, in Experiment 2, the N concentration in the labelling compartment was adjusted to equal that in the full-strength MSR medium.
Fig. 1. Experimental system (a microcosm) employed in this study.
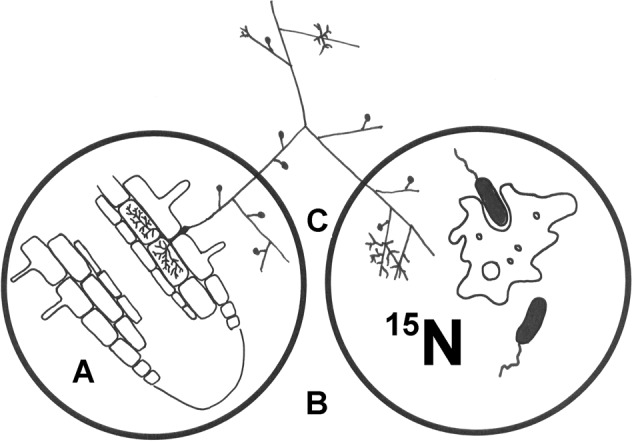
The system was prepared in a large Petri dish (not shown here). Inside that dish, a small rhizobox (A), made of a small Petri dish and delimited from the rest of the system with a 42 µm nylon mesh, contained the Ri T-DNA transformed Cichorium intybus roots, either non-mycorrhizal or mycorrhizal. Mycorrhizal fungal hyphae growing out of the mycorrhizal roots through the mesh colonised the MSR medium [35] filling the large dish volume (B) and eventually reached the labelling compartment (C). The labelling compartment was made of another small Petri dish and was filled with a modified (nitrogen-free) MSR medium, with (Experiment 1) or without (Experiment 2) sucrose and with or without an added 15N-labelled nitrogen source (either mineral or organic) and various bacteria combined or not combined with a protist grazer (more details in supplementary Fig. S4).
Fig. 2. Development of Rhizophagus irregularis hyphae and spores in Experiment 1 as affected by mineral- (ammonium chloride) or organic- (chitin) nitrogen addition to the labelling compartment (marked with asterisk) in comparison to a nitrogen-free control.

Growth of the R. irregularis was supported by Ri T-DNA transformed Cichorium intybus roots in rhizoboxes (small dishes delimited from the main dish volume by a 42 µm nylon mesh), visible in the upper row (A–C), where they are labelled with handwritten plate numbers. The AM fungal hyphae and spores developed in the main dish volume filled with the standard MSR medium [35] and supplemented with 1% sucrose after emerging from roots pre-colonised with Rhizophagus and inoculated into the rhizoboxes. After 2 months of growth, labelling compartment was filled with MSR medium supplemented with 1% sucrose and devoid of nitrogen (A, D, G, J, M), or with added 15N-labelled ammonium chloride (B, E, H, K, N) or 15N-chitin extracted from Zygorhynchus sp. cell walls (C, F, I, L, O). Fungal development was then observed and photographed at the edge of the labelling compartment under an Olympus SZX10 stereomicroscope at different time points (see left edge of panel for details).
Experiment 1
This experiment was carried out to test whether the AM fungal hyphae were able to take up and transport a significant quantity of N supplied as chitin towards the roots compared to N supplied as ammonium. Further, we tested whether any of the bacteria added to the hyphosphere would improve AM fungal access to the N bound in chitin. Because the standard cultivation media (MSR) for monoxenic AM fungal cultures contains sucrose, and also because we wanted to supply chitin as the only N source (and not the only N and C source) in the labelling compartment, we also supplied sucrose to the labelling compartment in this experiment. Altogether, 287 experimental microcosms without any visible bacterial contaminations and with AM fungal hyphae (in the mycorrhizal treatment) profusely colonising the root-free MSR medium were included into this experiment. Each labelling compartment was filled with 15 ml of N-free MSR medium (where KNO3 and Ca(NO3)2 were replaced by KCl and CaCl2, respectively) supplemented with 1% sucrose and solidified with 0.3% Phytagel. To this medium was added either 60 mg 15N-labelled chitin per compartment or an equal amount of N provided as NH4Cl (30% 15N atom%) or left without N amendment. Amendments with 15N-labelled compounds effectively elevated the N concentration in the labelling compartment fourfold as compared to the medium filling the large plate volumes. Thereafter, the plates were incubated for another 26 days to allow colonisation of the labelling compartments by AM fungal hyphae in the mycorrhizal treatment. At that time point, fresh (4 days old) bacterial liquid cultures (lysogeny broth) produced from glycerol stocks upon reciprocal shaking (110 rpm, 2 cm) at 24 °C were added or not added to the labelling compartment as 3 × 20 µl of the respective bacterial suspension or of sterile lysogeny broth. The plates were then incubated for an additional 20 days at 24 °C in darkness before harvest.
The full factorial design with 6–8 replicate plates per treatment combination included three factors: (1) AM fungus (two levels, present or absent), (2) 15N amendment of the labelling compartment (three levels, chitin, ammonium chloride, or none), and (3) bacterial inoculation of the labelling compartment (six levels). The latter included either none or one of the following bacterial strains ID 1, 5, 9, 10, or 15 (Table 1). In addition, 4–6 mycorrhizal and NM plates with chitin as the 15N-labelled N source in the labelling compartment were established per selection of other bacterial treatments (i.e. strains ID 2, 8, 16, or 17; see Table 1 for additional details).
Experiment 2
This experiment was mainly directed to testing whether including a protist into the experimental system containing plant (roots), AM fungus, and bacteria would further improve the efficiency of utilising organic N from AM fungal hyphosphere. Because we previously had noted some interferences between sucrose amendment of cultivation media and growth of several bacteria included in the research described here (details in supplementary Figs. S1 and S2), we omitted sucrose from the labelling compartment in this experiment. This effectively made the chitin the only N and the main C source for the saprotrophic microbes in the labelling compartment, considering that C inputs were comparably low from the hyphal exudates and AM fungal necromass to the labelling compartment.
This experiment included 182 experimental microcosms. All were free of bacterial contamination upon filling the labelling compartments, and all mycorrhizal plates produced extensive hyphal networks in the root-free compartment. Each labelling compartment was filled with 15 ml of N-free MSR medium devoid of sucrose and solidified with 0.3% Phytagel. Each of the labelling compartments contained 15 mg of 15N-labelled chitin, corresponding to the N concentration in the medium filling the large plates. The plates were then incubated for 26 days to allow for efficient colonisation of the labelling compartments by the AM fungal hyphae in the mycorrhizal treatment. Subsequently, fresh (4 days old) bacterial liquid cultures produced as above were added or not added to the labelling compartment as 1 × 60 µl of the bacterial suspension or of sterile lysogeny broth. The plates were incubated for a further 13 days at 24 °C in darkness. Thereafter, the spots inoculated with bacteria in the labelling compartment were either inoculated or not inoculated with spores of the protist P. pallidum produced on cultures of the respective bacteria, with the exceptions of bacteria ID 8 and 13, which had not supported protist sporulation in previous co-cultures. Protist spores recovered from aerial sporangiophores formed while co-culturing the protist with bacterium ID 2 were used instead. The plates were then incubated for an additional 9 days at 24 °C in darkness before harvest.
The full factorial design included 4–5 replicate NM plates and 6–7 replicate mycorrhizal plates per each microbial inoculation combination. Three experimental factors were considered: (1) AM fungus (two levels, present or absent); (2) bacterial inoculation of the labelling compartment (eight levels, inoculated with one of the bacterial strains ID 2, 5, 8, 12, 13, 15, 17, or left uninoculated), and protist inoculation (two levels, inoculated or uninoculated).
Harvest and analyses
Microcosms with roots escaping from the rhizobox were removed from both experiments unless the roots grew no closer than within 1 cm of the labelling compartment. Further, plates with visible bacterial contamination were excluded from the experiments unless this was strictly delimited spatially to individual and easily removable colonies in the large plate volume. Such contaminating colonies were removed prior to extracting roots and AM fungal hyphae from the plates.
Upon harvest, the plates were opened, roots from the rhizobox removed, dried at 65 °C for 3 days, weighed and then pulverised in a ball mill (MM 200, Retsch, Haan, Germany). Their C and N concentrations and isotopic composition of N were analysed using a Flash 2000 elemental analyser coupled with a Delta V Advantage isotope-ratio mass spectrometer (Thermo Fischer Scientific, Bremen, Germany). The hyphae from the root-free zone (not including the labelling compartment) were collected by filtering through Omnipore membrane filters (5 µm pore size, 47 mm diameter, Merck Millipore, Burlington, MA, USA) after the Phytagel was dissolved in 10 mM potassium citrate buffer (pH 6.0). Dry weights of the hyphae were recorded after drying at 65 °C for 3 days and their C and N concentrations and N isotopic compositions were analysed as above. Phosphorus concentrations were analysed in randomly selected root and hyphal samples from Experiment 1 by Malachite green colorimetry [36] after dry incineration of the samples (550 °C, 12 h) and extraction of the ashes with concentrated nitric acid [37]. In Experiment 2, we further quantified the development of AM fungal hyphae, bacteria, and the protist using quantitative real-time PCR (details and results in the Supplementary Information).
Calculations and statistical analyses
The N, C, and P contents of the roots and AM fungal hyphae per microcosm were calculated from the respective element concentrations and dry biomass of the samples. Excess 15N values (i.e. the amounts of N originating from the isotopically enriched inputs) in the root and the hyphal samples were calculated by considering molar N concentrations in the samples, their 15N abundance given as atom%, and using 15N abundance of samples without 15N-labeled compounds added to the labelling compartment as an isotopic baseline. Percentage of 15N transfer from the labelling compartment to the AM fungal hyphae and/or roots was then calculated using a two-source mixing model employing a mass balance equations framework as detailed elsewhere [38]. Knowing the levels of isotopic enrichment of the labelled compounds and their inputs per experimental system (i.e. microcosm), we calculated the fraction of 15N supplied per microcosm and recovered either in the roots or the AM fungal hyphae.
Data were analysed using one-, two-, or three-way analysis of variance (ANOVA), followed by post hoc Duncan’s multiple range test separating treatment means if ANOVA proved significant (p < 0.05). These analyses were carried out in Statgraphics Plus for Windows v. 3.1 (Statgraphics Technologies, Inc., The Plains, VA, USA). In case of significant heteroscedasticity of data (checked by Bartlett’s test), log (x + 0.1) transformation was employed prior to statistical analyses.
Results
Colonisation of labelling compartment by AM fungal hyphae
Rapid colonisation of the labelling compartment by finely branched AM fungal hyphae was observed when N was supplied as NH4Cl in Experiment 1. That was in contrast to a labelling compartment completely devoid of N (Fig. 2). When the labelling compartment was supplied with chitin in Experiment 1, the speed of AM fungal hyphae ingrowth was comparable to that with NH4Cl, but the hyphal morphology differed. The chitin-fed hyphae were long, spread further, and were only sparsely branched (Fig. 2). In Experiment 2, the development of AM fungus in the labelling compartment was either not affected by the different bacterial isolates or was stimulated by isolates ID 2, 5 and 12 (supplementary Fig. S5). In the same experiment, AM fungus development in the labelling compartment was not significantly affected by presence of the protist Polysphondylium pallidum (supplementary Table S1).
15N transfer to AM fungal hyphae and roots—effect of N source and microbes
In Experiment 1, transfer of isotopically labelled N from the labelling compartment to the NM roots or mycorrhizal roots with the AM fungal hyphae connected to them was affected by all three experimental factors (i.e. presence of AM fungus, identity of bacterial isolate, and form of N supplied) and their interactions (supplementary Table S2). When N was supplied as NH4Cl, transfer of N was ~10% of the added N appearing in the mycorrhizal roots and their associated AM fungal hyphae, whereas values below 1% were recorded in the NM roots (Fig. 3). Further, transfer of N added as NH4Cl from the labelling compartment to the NM roots or mycorrhizal roots with AM fungal hyphae was not affected by presence or identity of prokaryotes in the labelling compartment (Fig. 3, supplementary Table S3). When N was supplied to the labelling compartment as chitin in Experiment 1, its uptake to NM roots was low (i.e. all values below 0.17% of the supplied 15N, except a single outlier reaching 0.31%), regardless of the identity of prokaryotes added. The uptake of N from chitin by mycorrhizal roots and the AM fungal hyphae connected to them was comparable to that of the NM roots (reaching up to 0.15% of the supplied 15N) in all bacterial treatments except the bacterial isolate ID 5 (Paenibacillus chitinolyticus). That bacterium significantly increased N acquisition from chitin via AM fungal hyphae to a mean 1.43% of the supplied 15N (Fig. 3). This resulted in high statistical significance of both AM fungal presence and identity of bacteria inoculated in the labelling compartment as well as their interaction in chitin-supplemented microcosms (Fig. 3, supplementary Table S4). Further, we found that another isolate of Paenibacillus sp. (ID 17) significantly increased N transfer from chitin to the roots and the AM fungal hyphae outside the labelling compartment (reaching a mean 0.52% of the supplied 15N) as compared to the other bacteria (Fig. 4). All primary data to Experiment 1 are available as Supplementary Data 1 file accompanying this paper.
Fig. 3. Transfer rates of 15N applied into the labelling compartment to the roots together with the mycorrhizal hyphae attached to them (if applicable) in Experiment 1 according to presence or absence of mycorrhizal fungus in the experimental system (M + , mycorrhizal; NM, non-mycorrhizal) and identity of selected bacterial isolates added to the labelling compartment.
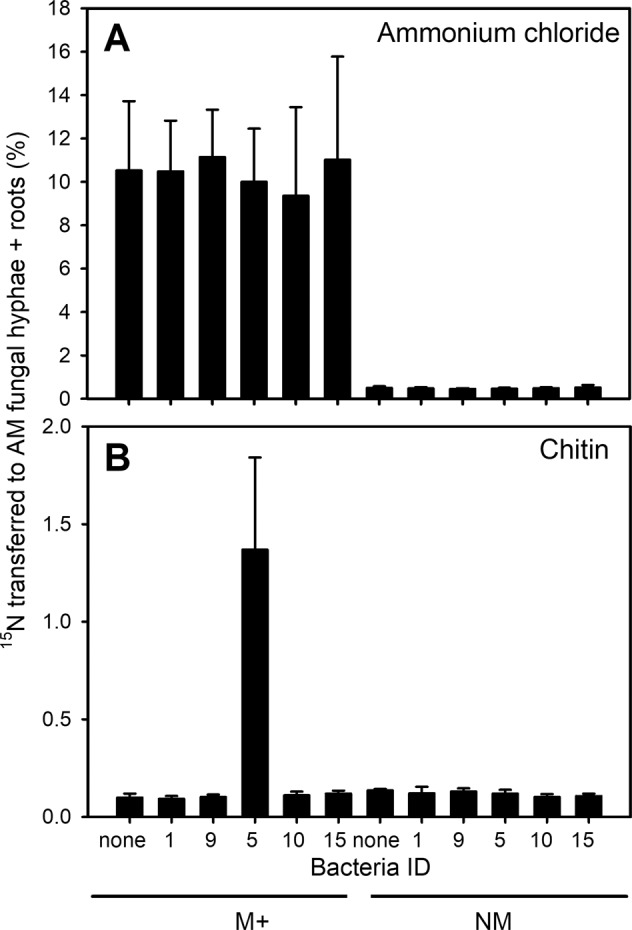
Shown are mean values (and standard deviations) of 5–8 replicates per treatment. Presented values illustrate the fraction of 15N-labelled mineral (A) or organic (B) nitrogen source applied into the labelling compartment and transferred to the non-mycorrhizal roots or mycorrhizal roots together with their attached mycorrhizal hyphae extracted from the large dish volume at the end of the experiment (%).
Fig. 4. Transfer rates of 15N-labelled nitrogen applied into the labelling compartment in the form of chitin to the roots (grey bars) or to the mycorrhizal hyphae in the root-free zone of the large Petri dish (black bars) in Experiment 1.
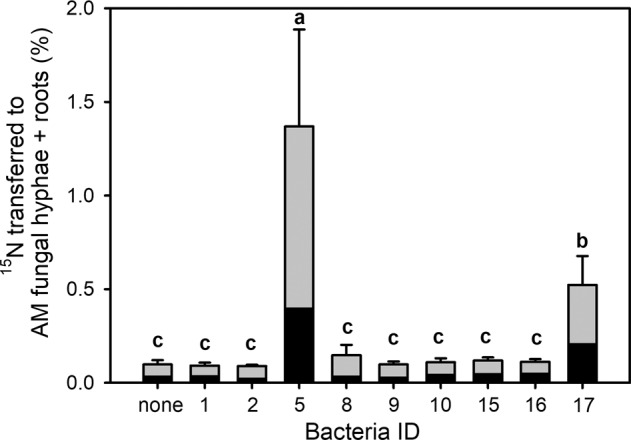
Only results from mycorrhizal microcosms are included in this analysis. Shown are mean values (and standard deviations) of 4–6 replicates per treatment of the total 15N-transfer values (i.e. excess 15N contained in the roots and their attached mycorrhizal fungal hyphae). Different letters indicate significant differences between treatment means with respect to the amount of 15N-labelled nitrogen applied as chitin into the labelling compartment and transferred to the roots and their attached mycorrhizal fungal hyphae, expressed as percentage of total 15N supplied in the labelling compartment. Means were separated by Duncan’s post-hoc test (p < 0.05), following significant ANOVA (F9,50 = 27.3, p < 0.001). Data were log(x + 0.1) transformed prior to ANOVA.
In Experiment 2, we observed generally higher rates of 15N transfer from the labelling compartment to the mycorrhizal roots with their attached AM fungal hyphae as compared to Experiment 1 (Fig. 5). Significantly higher transfer rates of N from chitin were recorded in mycorrhizal microcosms with isolate ID 5 added to the labelling compartment (mean 11.5% of the supplied 15N) as compared to other bacterial isolates or to N transfer from the labelling compartment to NM roots. In the NM microcosms, the values remained below 1.36% of the supplied 15N (Fig. 5, see supplementary Table S5 for ANOVA results). Further, presence of the protist Polysphondylium pallidum in the labelling compartment significantly increased the rate of N transfer to the roots and their associated AM fungal hyphae (supplementary Table S5). This effect obviously was driven by highly significant (p = 0.011, one-way ANOVA) increase of N transfer due to protist inoculation in mycorrhizal microcosms with added bacterium ID 5. The mean 15N transfer rates in the relevant treatments were 8.5% and 14% of the supplied 15N, which were detected in the roots and the extraradical AM fungal hyphae without and with the protists, respectively (Fig. 5). All primary data to Experiment 2 are available as Supplementary Data 2 file accompanying this paper.
Fig. 5. Transfer rates of 15N-labelled nitrogen applied to the labelling compartment in the form of chitin to the roots (grey bars) or to the mycorrhizal hyphae in the root-free zone of the large Petri dish (black bars) in Experiment 2.

Results are shown separately for non-mycorrhizal plates without protists (A), non-mycorrhizal plates with protists (B), mycorrhizal plates without protists (C), and mycorrhizal plates with protists (D). Shown are mean values (and standard deviations) of 4–7 replicates per bacterial treatment. Different lower-case letters in each graph indicate significant differences between treatment means within the individual graphs. The numbers presented are percentages of 15N-labelled nitrogen applied as chitin into the labelling compartment, and recovered outside the labelling compartment either in the roots or in the mycorrhizal hyphae. Means were separated by Duncan’s post-hoc test (p < 0.05), following significant ANOVAs (respective F values and p value ranges are shown for each individual graph). Data were log(x + 0.1) transformed prior to ANOVA. Statistical comparisons were carried out separately for each of the four scenarios (i.e. presence or absence of the mycorrhizal fungus and presence or absence of the protist). *** p < 0.001, ** 0.01 > p ≥ 0.001.
15N allocation to roots and AM fungal hyphae
Only marginal differences (ANOVA F9,50 = 1.96, p = 0.07) were observed between the bacterial treatments in terms of 15N allocation from chitin to the AM fungal hyphae and mycorrhizal roots (i.e. considering only those mycorrhizal microcosms amended with chitin) in Experiment 1 (Fig. 4). Somewhat lower 15N allocation to hyphae was noted upon inoculation with bacterium ID 2 to the labelling compartment (<28% of the total 15N transferred out of the labelling compartment), in contrast to isolates ID 16 and ID 17, which induced more than 45% of the chitin-derived 15N detected outside of the labelling compartment to be allocated to the hyphae. Allocation of 15N from ammonium chloride to the roots and the AM fungal hyphae was not significantly affected by any of the bacteria (p > 0.1). Gross mean 15N allocation to the hyphae across all bacterial treatments was 27 ± 4.5% (n = 59) of the 15N originating from the NH4Cl and detected outside of the labelling compartment in either the root or hyphal biomass.
In Experiment 2, significant differences were detected in chitin-derived 15N allocation between roots and the hyphae in mycorrhizal microcosms when the protists were present. In the absence of protists, only marginally significant differences were observed between bacterial isolates (F7,47 = 2.12, p = 0.064), ranging from <30% of the entire amount of chitin-derived N outside of the labelling compartment being detected in the hyphae (bacteria ID 8 and ID 2) to >43% (bacterium ID 15). When protists were present, allocation of the 15N to roots and the hyphae was significantly affected by bacterial identity (F7,53 = 4.26, p = 0.001), with the bacterium ID 5 showing the smallest allocation to hyphae (21%). Further, bacterium ID 8 also showed low (~33%) 15N allocation to hyphae, very similar to the bacteria-free treatment with 34% of the 15N being detected in the hyphae. All the aforementioned treatments showed significantly lower values than did the treatment with bacterium ID 17 in the labelling compartment together with the protist, in which case 46% of all chitin-derived N detected outside of the labelling compartment was allocated to the hyphae.
Discussion
Limited uptake of N from chitin by AM fungal hyphae alone
Our isotopic analyses revealed that the AM fungal hyphae could not release and take up significant amounts of N bound in chitin on their own, despite that chitinase genes have recently been uncovered in the genomes of several AM fungal species [27]. These results support our first hypothesis. On the other hand, and consistent with previous literature [20, 39, 40], the hyphae of the AM fungus could take up and transport large quantities of ammonium ions from the labelling compartment towards the roots, regardless of the identity of prokaryotes added or not added to that compartment. Passive diffusion of NH4+ ions from the labelling compartment towards the roots was obviously very effectively blocked (Fig. 3). We nevertheless noted a low but still measurable 15N transfer to roots from the labelling compartment where either NH4Cl or chitin was added in both mycorrhizal and NM microcosms. This might have been due to diffusion of NH3 gas, which has recently been proposed as an alternative and largely neglected pathway of lateral N flow from soil to AM fungal hyphae and/or roots [41]. Whereas spontaneous production of ammonia from ammonium ions in aqueous solution is long known [42], production of ammonia gas from fungal chitin would require either spontaneous degradation of chitin due to residual chitinase and/or deaminase activities or presence of free ammonium ions in the chitin preparation. Both are theoretically possible and would require further analyses to clarify, but neither of these potential issues invalidate the results of our experiments. They only would increase the background 15N.
Some chitinolytic bacteria improve utilisation of chitin-bound N by AM fungal hyphae
In our research, two isolates of chitinolytic Paenibacillus sp. increased utilisation of chitin as N source by the AM fungal hyphae, albeit to different extents or with different temporal dynamics. Differences in the latter are suggested due to differential allocation of chitin-derived 15N to the roots and the AM fungal hyphae outside of the labelling compartment between treatments added with bacteria ID 5 and ID 17 (see above).
Paenibacillus sp. has repeatedly been isolated from spores of AM fungi [43, 44], and it also has been demonstrated to bind to both vital and non-vital AM fungal hyphae under laboratory conditions [45, 46]. A number of significant interactive effects have previously been described between various AM fungi and Paenibacillus isolates [47–49]. One genotype of Paenibacillus validus has been shown to stimulate asymbiotic growth (i.e. formation of new spores from a germinating spore in the absence of a host plant) of an AM fungus Rhizophagus sp. [50]. We do not yet know how frequently members of Paenibacillus sp. actually occur in the AM fungal hyphosphere. It seems, though, that they are not restricted solely to biofilms at the AM fungal hyphae surfaces but occur both in the hyphosphere and surrounding soil [24]. This means they could be regarded as opportunistic inhabitants of the AM fungal hyphosphere and possibly involved also in recycling of AM fungal necromass [33].
Interestingly, two other chitinolytic bacteria (Janthinobacterium sp.) did not support acquisition of N from chitin by the AM fungus in our experiments. This is surprising inasmuch as we confirmed that those bacteria obviously expressed chitinolytic activity, particularly when offered chitin as the only C and N source (i.e. in absence of sucrose) and without contact to AM fungal hyphae (supplementary Figs. S1 and S2). Moreover, Janthinobacterium sp. is efficiently grazed by the protist Polysphondylium pallidum (supplementary Figs. S5 and S6). This means that, at least in the presence of protist [25], chitin-derived N should be released from the Janthinobacterium cells, allowing for detection of a significant N transfer from chitin to the AM fungal hyphae. This should be the case even if the bacterium was able to evade competition with the AM fungal hyphae for chitin’s primary degradation products. Our results thus suggest differential expression of chitin-degrading genes in the Janthinobacterium sp. depending upon the presence of AM fungal hyphae and thus indicating limited validity of our second hypothesis.
Compared to Paenibacillus sp., there is less experimental evidence for specific interactions between AM fungal hyphae and Janthinobacterium sp., although this bacterial genus (or its close relatives) has previously been reported from both AM fungal spores and hyphae [45, 51]. Our results could possibly be explained such that, in the presence of AM fungal hyphae, Janthinobacterium sp. (which was actually growing profusely in Experiment 2 in the absence of sucrose; see supplementary Fig. S5 for data) would preferentially utilise complex AM fungal exudates [7, 10, 52] as compared to exogenously provided chitin, and the chitin would possibly be utilised only in absence of the AM fungus.
Different C and N stoichiometry
It is clear that omitting sucrose from the cultivation media in the labelling compartment in Experiment 2 strongly increased chitin mineralisation by the prokaryotes, even though the AM fungal hyphae’s capacity for ammonium ions uptake obviously was not strongly hampered by sucrose (Fig. 3). Whereas the transfer of N from the labelling compartment to the AM fungal hyphae and roots in Experiment 1 (with sucrose) reached just up to 2% of the N supplied, only in the absence of sucrose (Experiment 2) did we reach N transfer rates above 10%. Only the latter results were thus comparable to those of our previous experiments conducted with unsterile potting substrates [18, 20]. This seems to be due at least in part to the fact that in Experiment 2 chitin and the living AM fungal hyphae were the only C sources for the prokaryotes in the labelling compartment. Increased need of C from the chitin by the bacteria in Experiment 2 may thus explain the order of magnitude faster mineralisation of chitin as compared to Experiment 1.
Protists speed up AM fungal uptake of N from chitin
We demonstrated that prokaryote-assisted acquisition of N by AM fungal hyphae from chitin accelerated when a eukaryotic grazer (a protist) was present together with the prokaryote in the organic N zone. These results provide strong support to our third hypothesis. However, significant variation in population size of the protists depending upon bacterial identity was noted in Experiment 2 (supplementary Fig. S5). This was not necessarily co-incident with chitin mineralisation rates and the rates of N transfer from chitin to the AM fungal hyphae. Further research is therefore justified as to the mechanisms and pathways of organic N recycling in AM fungal hyphosphere.
Conclusions and future perspectives
Resource stoichiometry and temporal dynamics of inter-kingdom interactions and population dynamics in AM fungal hyphosphere should be addressed in future research, which should also be expanded to other AM fungal species, bacteria, and protists (ideally isolated from a single ecosystem). Knowledge gained from simplified in vitro experiments should then be confirmed under unsterile soil conditions, using a full (green) plant model, stable isotope probing, co-occurrence networks, and (possibly also) by direct observation of microbial interactions and element transports, as well as utilising dense spatiotemporal sampling schemes. Other protists are likely to dominate grasslands or agricultural soils as compared to forests, and thus protists like Vermamoeba sp. may be more relevant for future studies as opposed to Polysphondylium sp. [53]. Such research will be most relevant to soils and AM fungal symbiosis-dominated ecosystems reliant on organic nutrient recycling, such as tropical rainforests, grasslands, and/or organically managed farms [54–56].
Supplementary information
Acknowledgements
Technical support is gratefully acknowledged from Věra Procházková (processing samples for mass spectrometry analyses), Hana Gryndlerová (phosphorus analyses), and David Püschel (technical photography). Two of the bacterial pure cultures were provided by CCM culture collection (Brno, Czech Republic). The original culture of the AM fungus in monoxenic culture setting was kindly provided by Symbio-M (Lanškroun, Czech Republic). Experimental work described here was funded by the Czech Science Foundation (projects 18-04892S and 21-07275S). In addition, isolation of two bacterial strains (Janthinobacterium sp.) was supported by Czech Science Foundation project 17-09946S. Further institutional support was granted by the long-term development program RVO 61388971.
Author contributions
MR and JJ conceived and developed the study. MR, PB and HH isolated, sequenced and cultured the different microbes included in the research described here. PB and MK prepared and analysed isotopically labelled chitin. MR, PB, HH, MK, MD and KG all contributed to establishment and harvest of the two main experiments and sample processing for the different (elemental, isotopic and molecular) analyses. MR conducted the bacterial cultivation assays. MR, MK, MD and KG conducted quantitative molecular analyses in Experiment 2 (including development and calibration of microbial taxa-specific qPCR markers). JJ drafted the paper. All authors contributed to its revisions and approved the final version.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
The online version contains supplementary material available at 10.1038/s41396-021-01112-8.
References
- 1.Redecker D, Kodner R, Graham LE. Glomalean fungi from the Ordovician. Science. 2000;289:1920–1. doi: 10.1126/science.289.5486.1920. [DOI] [PubMed] [Google Scholar]
- 2.Parniske M. Arbuscular mycorrhiza: the mother of plant root endosymbioses. Nat Rev Microbiol. 2008;6:763–75. doi: 10.1038/nrmicro1987. [DOI] [PubMed] [Google Scholar]
- 3.Raven JA, Lambers H, Smith SE, Westoby M. Costs of acquiring phosphorus by vascular land plants: patterns and implications for plant coexistence. N Phytol. 2018;217:1420–7. doi: 10.1111/nph.14967. [DOI] [PubMed] [Google Scholar]
- 4.Field KJ, Pressel S. Unity in diversity: structural and functional insights into the ancient partnerships between plants and fungi. N Phytol. 2018;220:996–1011. doi: 10.1111/nph.15158. [DOI] [PubMed] [Google Scholar]
- 5.Harrison MJ, Vanbuuren ML. A phosphate transporter from the mycorrhizal fungus Glomus versiforme. Nature. 1995;378:626–9. doi: 10.1038/378626a0. [DOI] [PubMed] [Google Scholar]
- 6.Smith SE, Jakobsen I, Gronlund M, Smith FA. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011;156:1050–7. doi: 10.1104/pp.111.174581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhang L, Feng G, Declerck S. Signal beyond nutrient, fructose, exuded by an arbuscular mycorrhizal fungus triggers phytate mineralization by a phosphate solubilizing bacterium. ISME J. 2018;12:23–51. doi: 10.1038/s41396-018-0171-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang L, Xu MG, Liu Y, Zhang FS, Hodge A, Feng G. Carbon and phosphorus exchange may enable cooperation between an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. N Phytol. 2016;210:1022–32. doi: 10.1111/nph.13838. [DOI] [PubMed] [Google Scholar]
- 9.Koide RT, Kabir Z. Extraradical hyphae of the mycorrhizal fungus Glomus intraradices can hydrolyse organic phosphate. N Phytol. 2000;148:511–7. doi: 10.1046/j.1469-8137.2000.00776.x. [DOI] [PubMed] [Google Scholar]
- 10.Jiang FY, Zhang L, Zhou JC, George TS, Feng G. Arbuscular mycorrhizal fungi enhance mineralisation of organic phosphorus by carrying bacteria along their extraradical hyphae. N Phytol. 2021;230:304–15. doi: 10.1111/nph.17081. [DOI] [PubMed] [Google Scholar]
- 11.Tisserant E, Malbreil M, Kuo A, Kohler A, Symeonidi A, Balestrini R, et al. Genome of an arbuscular mycorrhizal fungus provides insight into the oldest plant symbiosis. Proc Natl Acad Sci USA. 2013;110:20117–22. doi: 10.1073/pnas.1313452110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Miyauchi S, Kiss E, Kuo A, Drula E, Kohler A, Sanchez-Garcia M, et al. Large-scale genome sequencing of mycorrhizal fungi provides insights into the early evolution of symbiotic traits. Nat Commun. 2020;11:5125. doi: 10.1038/s41467-020-18795-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Johansen A, Jakobsen I, Jensen ES. Hyphal transport by a vesicular-arbuscular mycorrhizal fungus of N applied to the soil as ammonium or nitrate. Biol Fert Soils. 1993;16:66–70. [Google Scholar]
- 14.Wipf D, Krajinski F, van Tuinen D, Recorbet G, Courty PE. Trading on the arbuscular mycorrhiza market: from arbuscules to common mycorrhizal networks. N Phytol. 2019;223:1127–42. doi: 10.1111/nph.15775. [DOI] [PubMed] [Google Scholar]
- 15.Johansen A, Jensen ES. Transfer of N and P from intact or decomposing roots of pea to barley interconnected by an arbuscular mycorrhizal fungus. Soil Biol Biochem. 1996;28:73–81. [Google Scholar]
- 16.Hodge A, Campbell CD, Fitter AH. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature. 2001;413:297–9. doi: 10.1038/35095041. [DOI] [PubMed] [Google Scholar]
- 17.Hodge A, Stewart J, Robinson D, Griffiths BS, Fitter AH. Competition between roots and soil micro-organisms for nutrients from nitrogen-rich patches of varying complexity. J Ecol. 2000;88:150–64. [Google Scholar]
- 18.Bukovská P, Bonkowski M, Konvalinková T, Beskid O, Hujslová M, Püschel D, et al. Utilization of organic nitrogen by arbuscular mycorrhizal fungi–is there a specific role for protists and ammonia oxidizers? Mycorrhiza. 2018;28:465. doi: 10.1007/s00572-018-0851-y. [DOI] [PubMed] [Google Scholar]
- 19.Püschel D, Janoušková M, Hujslová M, Slavíková R, Gryndlerová H, Jansa J. Plant-fungus competition for nitrogen erases mycorrhizal growth benefits of Andropogon gerardii under limited nitrogen supply. Ecol Evol. 2016;6:4332–46. doi: 10.1002/ece3.2207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bukovská P, Rozmoš M, Kotianová M, Gančarčíková K, Dudáš M, Hršelová H, et al. Arbuscular mycorrhiza mediates efficient recycling from soil to plants of nitrogen bound in chitin. Front Microbiol. 2021;12:574060. doi: 10.3389/fmicb.2021.574060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bunn RA, Simpson DT, Bullington LS, Lekberg Y, Janos DP. Revisiting the ‘direct mineral cycling’ hypothesis: arbuscular mycorrhizal fungi colonize leaf litter, but why? ISME J. 2019;13:1891–8. doi: 10.1038/s41396-019-0403-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Nuccio EE, Hodge A, Pett-Ridge J, Herman DJ, Weber PK, Firestone MK. An arbuscular mycorrhizal fungus significantly modifies the soil bacterial community and nitrogen cycling during litter decomposition. Environ Microbiol. 2013;15:1870–81. doi: 10.1111/1462-2920.12081. [DOI] [PubMed] [Google Scholar]
- 23.Herman DJ, Firestone MK, Nuccio E, Hodge A. Interactions between an arbuscular mycorrhizal fungus and a soil microbial community mediating litter decomposition. FEMS Microbiol Ecol. 2012;80:236–47. doi: 10.1111/j.1574-6941.2011.01292.x. [DOI] [PubMed] [Google Scholar]
- 24.Emmett BD, Lévesque-Tremblay V, Harrison MJ. Conserved and reproducible bacterial communities associate with extraradical hyphae of arbuscular mycorrhizal fungi. ISME J. 2021;e-pub ahead of print 1 March 2021; 10.1038/s41396-021-00920-2. [DOI] [PMC free article] [PubMed]
- 25.Trap J, Bonkowski M, Plassard C, Villenave C, Blanchart E. Ecological importance of soil bacterivores for ecosystem functions. Plant Soil. 2016;398:1–24. [Google Scholar]
- 26.Jansa J, Hodge A. Swimming, gliding, or hyphal riding? On microbial migration along the arbuscular mycorrhizal hyphal highway and functional consequences thereof. N Phytol. 2021;230:14–6. doi: 10.1111/nph.17244. [DOI] [PubMed] [Google Scholar]
- 27.Morin E, Miyauchi S, San Clemente H, Chen ECH, Pelin A, de la Providencia I, et al. Comparative genomics of Rhizophagus irregularis, R. cerebriforme, R. diaphanus and Gigaspora rosea highlights specific genetic features in Glomeromycotina. N Phytol. 2019;222:1584–98. doi: 10.1111/nph.15687. [DOI] [PubMed] [Google Scholar]
- 28.Gil-Cardeza ML, Calonne-Salmon M, Gomez E, Declerck S. Short-term chromium (VI) exposure increases phosphorus uptake by the extraradical mycelium of the arbuscular mycorrhizal fungus Rhizophagus irregularis MUCL 41833. Chemosphere. 2017;187:27–34. doi: 10.1016/j.chemosphere.2017.08.079. [DOI] [PubMed] [Google Scholar]
- 29.Voets L, Dupre de Boulois H, Renard L, Strullu DG, Declerck S. Development of an autotrophic culture system for the in vitro mycorrhization of potato plantlets. FEMS Microbiol Lett. 2005;248:111–8. doi: 10.1016/j.femsle.2005.05.025. [DOI] [PubMed] [Google Scholar]
- 30.Kiers ET, Duhamel M, Beesetty Y, Mensah JA, Franken O, Verbruggen E, et al. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science. 2011;333:880–2. doi: 10.1126/science.1208473. [DOI] [PubMed] [Google Scholar]
- 31.van’t Padje A, Galvez LO, Klein M, Hink MA, Postma M, Shimizu T, et al. Temporal tracking of quantum-dot apatite across in vitro mycorrhizal networks shows how host demand can influence fungal nutrient transfer strategies. ISME J. 2021;15:435–49. doi: 10.1038/s41396-020-00786-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gryndler M, Šmilauer P, Püschel D, Bukovská P, Hršelová H, Hujslová M, et al. Appropriate nonmycorrhizal controls in arbuscular mycorrhiza research: a microbiome perspective. Mycorrhiza. 2018;28:435–50. doi: 10.1007/s00572-018-0844-x. [DOI] [PubMed] [Google Scholar]
- 33.Jansa J, Šmilauer P, Borovička J, Hršelová H, Forczek ST, Slámová K, et al. Dead Rhizophagus irregularis biomass mysteriously stimulates plant growth. Mycorrhiza. 2020;30:63–77. doi: 10.1007/s00572-020-00937-z. [DOI] [PubMed] [Google Scholar]
- 34.Bukovská P, Püschel D, Hršelová H, Jansa J, Gryndler M. Can inoculation with living soil standardize microbial communities in soilless potting substrates? Appl Soil Ecol. 2016;108:278–87. [Google Scholar]
- 35.Cranenbrouck S, Voets L, Bivort C, Renard L, Strullu DG, Declerck S. Methodologies for in vitro cultivation of arbuscular mycorrhizal fungi with root organs. In: Declerck S, Strullu DG, Fortin JA, editors. In vitro culture of mycorrhizas. Berlin: Springer; 2005. [Google Scholar]
- 36.Ohno T, Zibilske LM. Determination of low concentrations of phosphorus is soil extracts using malachite green. Soil Sci Soc Am J. 1991;55:892–5. [Google Scholar]
- 37.Püschel D, Janoušková M, Voříšková A, Gryndlerová H, Vosátka M, Jansa J. Arbuscular mycorrhiza stimulates biological nitrogen fixation in two Medicago spp. through improved phosphorus acquisition. Front Plant Sci. 2017;8:390. doi: 10.3389/fpls.2017.00390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Phillips DL, Gregg JW. Uncertainty in source partitioning using stable isotopes. Oecologia. 2001;127:171–9. doi: 10.1007/s004420000578. [DOI] [PubMed] [Google Scholar]
- 39.Perez-Tienda J, Valderas A, Camanes G, Garcia-Agustin P, Ferrol N. Kinetics of NH4+ uptake by the arbuscular mycorrhizal fungus Rhizophagus irregularis. Mycorrhiza. 2012;22:485–91. doi: 10.1007/s00572-012-0452-0. [DOI] [PubMed] [Google Scholar]
- 40.He XX, Chen YQ, Liu SJ, Gunina A, Wang XL, Chen W, et al. Cooperation of earthworm and arbuscular mycorrhizae enhanced plant N uptake by balancing absorption and supply of ammonia. Soil Biol Biochem. 2018;116:351–9. [Google Scholar]
- 41.Hestrin R, Weber PK, Pett-Ridge J, Lehmann J. Plants and mycorrhizal symbionts acquire substantial soil nitrogen from gaseous ammonia transport. New Phytol. 2021;e-pub ahead of print 2 June 2021; 10.1111/nph.17527 [DOI] [PubMed]
- 42.Everett DH, Wynne-Jones WFK. The dissociation of the ammonium ion and the basic strength of ammonia in water. P R Soc Lond A Mat. 1938;169:190–204. [Google Scholar]
- 43.Bidondo LF, Colombo R, Bompadre J, Benavides M, Scorza V, Silvani V, et al. Cultivable bacteria associated with infective propagules of arbuscular mycorrhizal fungi. Implications for mycorrhizal activity. Appl Soil Ecol. 2016;105:86–90. [Google Scholar]
- 44.Cruz AF, Ishii T. Arbuscular mycorrhizal fungal spores host bacteria that affect nutrient biodynamics and biocontrol of soil-borne plant pathogens. Biol Open. 2012;1:52–7. doi: 10.1242/bio.2011014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Scheublin TR, Sanders IR, Keel C, van der Meer JR. Characterisation of microbial communities colonising the hyphal surfaces of arbuscular mycorrhizal fungi. ISME J. 2010;4:752–63. doi: 10.1038/ismej.2010.5. [DOI] [PubMed] [Google Scholar]
- 46.Toljander JF, Artursson V, Paul LR, Jansson JK, Finlay RD. Attachment of different soil bacteria to arbuscular mycorrhizal fungal extraradical hyphae is determined by hyphal vitality and fungal species. FEMS Microbiol Lett. 2006;254:34–40. doi: 10.1111/j.1574-6968.2005.00003.x. [DOI] [PubMed] [Google Scholar]
- 47.Jaderlund L, Arthurson V, Granhall U, Jansson JK. Specific interactions between arbuscular mycorrhizal fungi and plant growth-promoting bacteria: as revealed by different combinations. FEMS Microbiol Lett. 2008;287:174–80. doi: 10.1111/j.1574-6968.2008.01318.x. [DOI] [PubMed] [Google Scholar]
- 48.Larsen J, Jaramillo-Lopez P, Najera-Rincon M, Gonzalez-Esquivel CE. Biotic interactions in the rhizosphere in relation to plant and soil nutrient dynamics. J Soil Sci Plant Nut. 2015;15:449–63. [Google Scholar]
- 49.Mansfeld-Giese K, Larsen J, Bodker L. Bacterial populations associated with mycelium of the arbuscular mycorrhizal fungus Glomus intraradices. FEMS Microbiol Ecol. 2002;41:133–40. doi: 10.1111/j.1574-6941.2002.tb00974.x. [DOI] [PubMed] [Google Scholar]
- 50.Hildebrandt U, Janetta K, Bothe H. Towards growth of arbuscular mycorrhizal fungi independent of a plant host. Appl Environ Microbiol. 2002;68:1919–24. doi: 10.1128/AEM.68.4.1919-1924.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cruz AF, Horii S, Ochiai S, Yasuda A, Ishii T. Isolation and analysis of bacteria associated with spores of Gigaspora margarita. J Appl Microbiol. 2008;104:1711–7. doi: 10.1111/j.1365-2672.2007.03695.x. [DOI] [PubMed] [Google Scholar]
- 52.Luthfiana N, Inamura N, Tantriani, Sato T, Saito K, Oikawa A, et al. Metabolite profiling of the hyphal exudates of Rhizophagus clarus and Rhizophagus irregularis under phosphorus deficiency. Mycorrhiza. 2021;31:403–12. doi: 10.1007/s00572-020-01016-z. [DOI] [PubMed] [Google Scholar]
- 53.Oliverio AM, Geisen S, Delgado-Baquerizo M, Maestre FT, Turner BL, Fierer N. The global-scale distributions of soil protists and their contributions to belowground systems. Sci Adv. 2020;6:eaax8787. doi: 10.1126/sciadv.aax8787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Averill C, Turner BL, Finzi AC. Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature. 2014;505:543–5. doi: 10.1038/nature12901. [DOI] [PubMed] [Google Scholar]
- 55.Mäder P, Fliessbach A, Dubois D, Gunst L, Fried P, Niggli U. Soil fertility and biodiversity in organic farming. Science. 2002;296:1694–7. doi: 10.1126/science.1071148. [DOI] [PubMed] [Google Scholar]
- 56.Cavagnaro TR. Biologically regulated nutrient supply systems: compost and arbuscular mycorrhizas—a review. Adv Agron. 2015;129:293–321. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.