

Abstract
Spiral organisms were isolated from an antral gastric mucosal biopsy specimen from a dyspeptic patient with gastritis. Only corkscrew-shaped organisms resembling “Gastrospirillum hominis” (“Helicobacter heilmannii”) but no Helicobacter pylori-like organisms were seen in histological sections. H. pylori was not cultured from specimens from this patient. On the basis of biochemical reactions, morphology, ultrastructure, and 16S DNA sequencing, the isolated “G. hominis” was shown to be a true Helicobacter sp. very similar to Helicobacter felis and the “Gastrospirillum” but was separate from H. pylori. “G. hominis” is a pleomorphic gram-negative cork-screw-shaped, motile rod with 3 to 8 coils and a wavelength of about 1 μm. In contrast to H. pylori, it has up to 14 sheathed flagellar uni- or bipolar fibrils but no periplasmic fibrils. “G. hominis” grows under microaerobic conditions at 36 and 41°C on 7% lysed, defibrinated horse blood agar plates within 3 to 7 days and can be subcultured under microaerobic but not under anaerobic conditions on media similar to those used for H. pylori and H. felis. The small translucent colonies were, in contrast to those of H. felis, indistinguishable from those of H. pylori. “G. hominis” is, like H. pylori and H. felis, motile, is oxidase, catalase, nitrite, nitrate, and urease positive, and produces alkaline phosphatase and arginine arylamidase. Like H. pylori and H. felis, it is sensitive to cephalothin (30-μg disc), resistant to nalidixic acid (30-μg disc), and sensitive to most other antibiotics. The 16S DNA sequence clusters “G. hominis” together with “Gastrospirillum,” H. felis, Helicobacter bizzozeronii, Helicobacter salmonii, Helicobacter nemestrinae, Helicobacter acinonychis, and H. pylori.
Curved and spiral organisms that cause inflammation of the gastrointestinal mucosae of humans and animals have been described regularly during the last century (2, 3, 9, 26, 33, 43). Thus, Lockard and Boler (26) described three types of spiral organisms in the gastric mucosa of dogs. Doenges (9) described two types of microorganisms in the human gastric mucosa. These were curved rods that were similar to the human microorganism Helicobacter pylori and spiral microorganisms that resembled spirochetes. Until the last decade, the majority of these curved and spiral microorganisms had not been recovered in culture. There has been great interest in these microaerobic microorganisms from the gastrointestinal mucosa since Warren and Marshall (42) first described the culture of H. pylori in 1983 and showed the relation of H. pylori to peptic ulcer disease in humans (27). Several Helicobacter species isolated from both humans and animals have been cultured and identified (6, 8, 11–13, 15–17, 18, 25, 29, 31, 34, 39, 40).
In 1987 Dent et al. (7) described corkscrew-shaped organisms in the human gastric mucosa and proposed the name “Gastrospirillum hominis” (28). “G. hominis” has been described in histological sections by several investigators (10, 14, 19, 22, 30), having a prevalence of 0.2 to 0.6% in Europe and up to 4% in China in patients with dyspepsia and occasionally in patients with peptic ulcer. “G. hominis” is usually found in foveolae of the gastric mucosa in association with inflammation and has been observed in parietal cells. They are usually less adherent to the epithelium than H. pylori. A causal relationship between gastroduodenal diseases and “G. hominis” has not been confirmed. Solnick et al. (36, 37) succeeded in determining the “G. hominis” 16S sequence from human biopsy specimens by PCR amplification of the 16S rRNA gene (rDNA) in the biopsy specimens, and the name “Helicobacter heilmannii” was proposed. A recent study indicated that mucosa-associated lymphoid tissue lymphomas are more prevalent in patients with H. heilmannii infection than in patients with H. pylori infection (41).
In this report we describe the culture, biochemical characterization, and ultrastructural histopathology of a “G. hominis” strain from the human gastric mucosa.
MATERIALS AND METHODS
Culture.
Four gastric biopsy specimens, two from the antrum and two from the corpus, from a 23-year-old male patient with persistent dyspepsia (22) were transported in serum broth (Statens Serum Institut, Copenhagen, Denmark) and were grown on nonselective, 7% defribrinated, lysed horse blood agar plates (chocolate agar plates; Statens Serum Institut) within 4 h after endoscopy. The plates (primary culture) were grown in an anaerobic chamber with 5% O2, 10% CO2, and 85% N2 and without H2 in an atmosphere with 95% humidity at 36°C for up to 7 days. The bacteria were subcultured on agar base, brucella agar, brucella agar with 1.5% NaCl, brucella agar with 1% glycine, brucella agar with trimethylamine-N-oxide hydrochloride, and lactose agar, in addition to those media mentioned in Table 1. The bacteria were grown both aerobically and anaerobically on the same media and under the same conditions, as well as under the microaerobic conditions presented in Table 1.
TABLE 1.
Growth of subcultures of “G. hominis” in a microaerobic atmosphere compared with growth of H. pylori and H. felisa
| Medium | Growth
|
Growth after the indicated incubation times
|
|||||||
|---|---|---|---|---|---|---|---|---|---|
| H. pylori | “G. hominis” | H. felis |
H. pylori
|
“G. hominis”
|
H. felis
|
||||
| 3 days | 7 days | 3 days | 7 days | 3 days | 7 days | ||||
| Chocolate agar at the following temp: | |||||||||
| 22°C | +++ | +++ | +++ | + | + | + | + | + | + |
| 36°C | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ |
| 41°C | +++ | +++ | +++ | +++ | +++ | + | +++ | 0 | 0 |
| 42°C | +++ | +++ | +++ | 0 | 0 | 0 | 0 | 0 | 0 |
| 5% Horse blood agar at the following temp: | |||||||||
| 36°C | ++ | + | +++ | + | ++ | + | ++ | + | + |
| 41°C | ++ | + | +++ | + | ++ | + | ++ | (+) | + |
| 10% Horse blood agar | +++ | ++ | +++ | ||||||
| Brucella agar + 5% horse blood | +++ | +++ | +++ | ||||||
| Mueller-Hinton agar + 5% horse blood | +++ | +++ | +++ | ||||||
| Brain heart infusion agar | +++ | + | + | ||||||
| Brain heart infusion broth | ++ | 0 | 0 | ||||||
| Selective media: | |||||||||
| Campylobacter agar (Skirrow) | +++ | + | ++ | ||||||
| Campylobacter agar (Butzler) | 0 | 0 | 0 | ||||||
| Campylobacter agar (Blaser-Wang) | + | 0 | + | ||||||
| Campylobacter agar (Preston) | + | 0 | ++ | ||||||
| Campylobacter agar (Karmali) | 0 | 0 | 0 | ||||||
| Helicobacter agar (Dent) | +++ | + | +++ | ||||||
| Blood-free agar CCDA | 0 | 0 | 0 | ||||||
0, no growth; (+), very weak growth; +, weak growth; ++, moderate growth; +++, good growth.
Bacterial strains.
The cultured “G. hominis” strain was compared with H. pylori CCUG 17874 and Helicobacter felis CCUG 28539.
Morphology.
Four gastric biopsy specimens were stained with haematoxylin-eosin and periodic acid-Schiff stains for histological examination (22).
For transmission electron microscopy, bacteria grown on agar plates for 2 to 7 days were fixed with glutaraldehyde. The colonies were detached, and after 10 min of centrifugation at 3,000 × g, the precipitates were enrobed in melted Noble agar (Difco). Small cubes with visible clusters of cells were transferred to 3% glutaraldehyde and were fixed overnight at 4°C. The specimens were postfixed in 1% (wt/vol) OsO4 and stained en bloc with 2% (wt/vol) uranyl acetate in barbiturate buffer. The procedures for the dehydration, embedding in plastics, and the further preparation of thin sections were carried out as described previously (4).
Negative staining was carried out as follows. A Formvar-coated carbon-reinforced copper grid (400 mesh) was applied, film side down, on a droplet of a culture suspension in phosphate-buffered saline (pH 7.4) and was placed on a strip of Parafilm. The grid was dried on filter paper and was stained for 30 s on droplets of 1 or 2% (wt/vol) ammonium molybdate adjusted to pH 7.4 with NH4OH. The excess liquid was then sucked off with a piece of filter paper. Electron microscopy was carried out on a Phillips 201C electron microscope at 60 kV.
Biochemical characterization.
The following biochemical test systems were used for fermentation and enzyme reactions. (i) The API Rapid ID 32A (Biomerieux) was used. (ii) Tubes for fermentation contained sterile water with 0.5% ox meat extract, 1% peptone, 0.2% disodium hydrogen phosphate, 0.3% sodium chloride, 0.0024% bromthymol blue, and 0.5% carbohydrate. The following carbohydrates were tested: glucose, lactose, saccarose, arabinose, rhamnose, maltose, mannitol, sorbitol, inositol, adonitol, dulcitol, and salicin (Statens Serum Institut). The test tubes are routinely used for testing of members of the family Enterobacteriaceae. Positive results for the tubes were read as a color change from green to yellow caused by the production of acid. (iii) Hiss medium (Statens Serum Institut) consisted of 1% peptone, 10% horse serum, 0.08% phenol red, and 2% carbohydrate. The following carbohydrates were tested: glucose, lactose, galactose, saccharose, and maltose (Statens Serum Institut). Positive results for the tubes were read as a color change from red to yellow caused by the production of acid. (iv) Brain heart infusion tubes were used for anaerobic fermentation. The tubes contained 3.7% brain heart infusion (Difco), 0.5% yeast extract, 5% lysed horse blood, 0.00005% vitamin K, 0.0005% hemin, 0.05% cystein, and 0.5% carbohydrate. The following carbohydrates were tested: glucose, lactose, saccharose, arabinose, rhamnose, maltose, mannitol, sorbitol, inositol, adonitol, salicin, fructose, cellobiose, mannose, raffinose, ribose, trehalose, xylose, erythritol, esculin, starch, and bile acid (Statens Serum Institut). The pH in the tubes was measured after incubation. (v) Hugh-Leifson medium with 1% glucose was used (Statens Serum Institut). In addition, nitrate and nitrite were tested for by using ox meat broth with 0.02% KNO3 or sodium nitrite (Statens Serum Institut), and H2S was tested for by using triple sugar iron (TSI agar) incubated anaerobically. Indoxyl acetate hydrolysis was tested for as described previously (21), and sodium hippurate hydrolysis was tested for as described by Hwang et al. (23).
Susceptibility.
The susceptibilities of “G. hominis,” H. pylori, and H. felis to nalidixic acid and cephalothin were tested under microaerobic conditions with discs (Biodisc AB, Solna, Sweden) containing 30 μg of each compound on Mueller-Hinton agar with 5% horse blood. The susceptibility of “G. hominis” to ampicillin (diffusible amount of 33 μg of antibiotic), cephalothin (66 μg), erythromycin (78 μg), tetracycline (80 μg), ciprofloxacin (10 μg), nalidixic acid (130 μg), rifampin (30 μg), amoxicillin plus clavulanate (30 and 15 μg, respectively), and metronidazole (16 μg) were tested under microaerobic conditions by the agar diffusion method with tablets (Neosensitabs; Rosco A/S, Roskilde, Denmark) on 7% defibrinated, lysed horse blood plates that were incubated for 2 days. The susceptibility of “G. hominis” to amoxicillin, tetracycline, ciprofloxacin, erythromycin, and metronidazole was tested under microaerobic conditions after 2 days by the E test (Biodisc AB). The bacteria were tested with inocula of 106 to 107 CFU/ml.
16S rDNA characterization: preparation of rDNA, performance of PCR, and cycle sequencing.
For 16S rDNA sequencing, a 72-h bacterial culture was picked from a chocolate agar plate, and the intracellular DNases were inactivated by incubation at 80°C. After centrifugation the cells were disrupted in Chelex-100 resin (Bio-Rad) (5). One microliter of the DNA-containing supernatant was used as a template in a PCR with the first and the last 16S primers (Table 2). The resulting PCR product was sequenced with the AmpliTaq FS Dye Terminator Cycle Sequencing kit (Applied Biosystems, Perkin-Elmer) and the primers indicated in Table 2. The 16S rDNA sequence of the cultured “G. hominis” strain was compared to the 16S rDNA sequences from 20 Helicobacter species listed in Table 3. For alignment and phylogenetic analysis, the MegAlign program from the DNASTAR package (Lasergene, Madison, Wis.) was used. The program uses CLUSTAL V (20) for sequence alignment and the unweighted pair group method with arithmetic means (35) combined with neighbor joining (32) for dendrogram construction.
TABLE 2.
Positions, directions, and sequences of 16S rDNA sequencing primers
| Position (direction)a | Sequenceb |
|---|---|
| 8–27 (F) | AGAGTTTGATCCTGGCTCAG |
| 516–535 (F) | GCCAGCAGCCGCGGTAAkAC |
| 784–805 (F) | GGATTAGATACCCbbGTAGTCC |
| 360–341 (R) | CCCACTGCTGCCTCCCGTAG |
| 703–684 (R) | TCTACGCATTTCACyGCTAC |
| 805–786 (R) | GGTCTACCbGGGTATCTAAT |
| 1079–1060 (R) | TCACGrCACGAGCTGACGAC |
| 1194–1174 (R) | GACGTCrTCCnCdCCTTCCTC |
| 1332–1313 (R) | TTCAyGnAGkCGrGTTGCAG |
| 1540–1521 (R) | AAGGAGGTGwTCCAvCCvCA |
E. coli 16S rDNA numbering. F, forward; R, reverse.
n = A, C, G, and T; b = C, G, and T; d = A, G, and T; v = A, C, and G; k = G and T; r = A and G; y = C and T; w = A and T.
TABLE 3.
Accession numbers and strain designations of 16S rDNA sequences used in the phylogenetic analysis
| Species | Strain | GenBank/EMBL accession no. |
|---|---|---|
| H. acinonychis | Eaton 90-119-3 | M88148 |
| H. bilis | Hb1 | U18766 |
| H. canis | MIT 95-5359 | U65102 |
| H. cinaedi | CCUG 18818 | M88150 |
| H. cholecystus | Hkb-1 | U46129 |
| H. felis | Dog-1 | U51870 |
| H. fennelliae | CCUG 18820 | M88154 |
| H. hepaticus | FRED1 | L39122 |
| H. muridarum | UNSW 1.7st | AF010140 |
| H. mustelae | 91-292-EIA Fox | M88156 |
| H. nemestrinae | ATCC 49396T | X67854 |
| H. pametentis | B8A Seymour | M88147 |
| H. pullorum | UB3659 | L36141 |
| H. pylori | 85D08 | U00679 |
| H. rodentium | MIT 95-1707 | U96296 |
| H. salomonis | CCUG 37845 | U89351 |
| H. suncus | Kaz-2 | AB006148 |
| H. trogontum | LRB 8581 | U65103 |
| “G. hominis” | G1A1 | L10079 |
| “G. hominis” | G2A2 | L10080 |
| “H. heilmannii” | RH53 | Y18028 |
Protein profile.
The protein profile of the cultured “G. hominis” strain was visualized and was compared with those of H. pylori and H. felis by sodium dodecyl sulfate-polyacrylamide gel electrophoresis as described previously (1, 24). Coomassie brilliant blue staining was performed as described by Weber and Osborn (44). Silver staining was performed with silver nitrate after fixation with 10% glutaraldehyde, and development was performed with 75 μl of 24.5% formalin in 100 ml of 3% sodium carbonate.
Nucleotide sequence accession number.
The GenBank/EMBL database accession number for the cultured “G. hominis” strain is RH53, 418028.
RESULTS
Culture.
Culture of biopsy specimens revealed uniform, small, translucent colonies after 5 days on the primary plates. The colonies were indistinguishable from those of H. pylori. By phase-contrast microscopy two forms of the organisms could be identified: 10 to 15% of the organisms were long spiral-shaped rods and the majority were short ox bow-shaped rods that resembled H. pylori. By Gram staining both forms were gram-negative rods. Subcultures of the organisms revealed H. pylori-like colonies (1 to 2 mm translucent convex colonies without swarming) after 2 to 3 days on 7% defibrinated, lysed horse blood agar. A pure culture of long spiral-shaped rods was obtained after subculture. None of the three species grew on blood-free media under aerobic or anaerobic culture conditions. The growth conditions for “G. hominis” resembled those for H. pylori except for the growth on some of the selective Campylobacter media (Table 1). “G. hominis,” like H. pylori and H. felis, did not grow on brucella agar plates with 1.5% NaCl, whether 5% horse blood was added or not. All three Helicobacter species grew on brucella agar with 1% glycine and weaker with trimethylamine-N-oxide hydrochloride when 5% horse blood was added to the plates but not when 5% horse blood was not added. No hemolysis was observed. “G. hominis” grew weakly at 22°C, it grew well at 36 and 41°C, but it did not grow at all at 42°C (Table 1). “G. hominis” did not grow under aerobic or anaerobic conditions, but it grew equally well in atmospheres with and without H2 and thus had no H2 requirement.
Biochemical properties.
The cultured “G. hominis” strain was, like H. pylori and H. felis, motile and positive for oxidase, catalase, urease, nitrate, and nitrite. All three species were negative for indoxyl acetate hydrolase, H2S (TSI agar), and indole. The organisms did not produce acid from any sugars and did not hydrolyze sodium hippurate. In the API Rapid ID 32A system the cultured “G. hominis” strain was, like H. pylori and H. felis, positive only for urease, alkaline phosphatase, and arginine arylamidase. “G. hominis” did not hydrolyze starch and grew when bile acid was present.
Morphology.
Histologic sections of gastric biopsy specimens disclosed a mildly active chronic gastritis. Long (length, 3 to 8 μm) spiral bacteria were identified in the foveolae of the antral mucosa, usually situated at a distance from the epithelium. Bacteria were inapparent in the corpus mucosa. H. pylori-like organisms were not seen. Light microscopy of subcultured “G. hominis” strains revealed corkscrew-shaped organisms that had three to eight coils, that were 3 to 10 μm in length, and that had a wavelength of about 1 μm (22).
Electron microscopy of thin sections (Fig. 1A and B) and of negatively stained specimens (Fig. 2A and B) revealed a mixture of helical organisms that ranged from organisms that were 3 to 8 μm in length and from 0.4 to 0.7 μm in width down to organisms that were 1 to 2 μm in length and rod-like. The wave lengths were measured for 70 helical profiles of “G. hominis” grown from 1 to 5 days in culture and were found to vary from 0.7 to 1.4 μm. The wavelength did not correlate with the number of coils or the time of culture. Most of the helical forms were found to be unipolar, and up to 14 sheathed flagellae were seen to originate from the polar end (Fig. 2A and B) with the characteristic electron-lucent cytoplasmic zone (Fig. 1B). Small electron-dense granules were found in several of the cultured organisms (Fig. 1A). Periplasmic fibrils were never found in the helical organisms. A small percentage of coccoid forms were found in “G. hominis” organisms cultured for 5 days or longer.
FIG. 1.

Transmission electron microscopy of subcultures of “G. hominis” grown for 1 or 2 days. (A) Thin section of a representative field of “G. hominis” grown for 1 day showing a mixture of helical and rod-like organisms. Electron-lucent areas (arrowheads) are seen beneath the flagellated pole. Small cytoplasmic inclusions (arrows) are found in several of the cells. Bar, 2 μm. (B) Thin section of “G. hominis” grown for 2 days. Two flagella (arrowheads) are seen originating from the specialized pole region. Several sheathed flagellae are found between the cells (arrow). Bar, 0.5 μm.
FIG. 2.

Negative staining with 2% ammonium molybdate of subculture of “G. hominis” grown for 2 days. (A) Three helical organisms are seen; one (arrow) is in the process of dividing. Tufts of flagellae (arrowheads) are seen protruding from only one of the poles. Bar, 2 μm. (B) Higher magnification of the flagellated pole, disclosing the sheathed flagella together with a few unsheathed flagellae (arrowheads). Bar, 0.5 μm.
Susceptibility.
Like H. pylori and H. felis, “G. hominis” is sensitive to cephalothin (30 μg/disc), but “G. hominis” is resistant to nalidixic acid, (30 μg/disc) (Biodisc), whereas H. pylori and H. felis are susceptible to nalidixic acid. By susceptibility testing with Rosco Neosensitabs, “G. hominis” had large zones (>60 mm) with ampicillin, cephalothin, erythromycin, tetracycline, rifampin, ciprofloxacin, and the combination of amoxicillin and clavulanate and small zones (<45 mm) with nalidixic acid and metronidazole. The organisms were susceptible to amoxicillin, ciprofloxacin, tetracycline, and erythromycin (MIC range, 0.016 to 0.032 μg/ml) and resistant to metronidazole (MIC, 12 μg/ml) when they were tested by the E test. The organisms were eradicated from the patient by triple therapy with amoxicillin, metronidazole, and omeprazole.
16S rDNA sequencing.
The 16S rDNA of the cultured “G. hominis” strain was aligned to 20 Helicobacter and “G. hominis” sequences found in the GenBank/EMBL database (Table 3). Due to the limited lengths of some sequences, the analysis included bases corresponding to Escherichia coli 16S rDNA positions 18 to 1488. Intervening sequences in the Helicobacter canis and Helicobacter bilis sequences have been excluded, but hypervariable regions in the 16S rDNA sequences are included so that differences between closely related species can be seen. The cultured “G. hominis” 16S rDNA sequence was most identical to the already existing sequences from “G. hominis,” Helicobacter felis, and Helicobacter salomonis, which was reflected in the dendrogram (Fig. 3) in which the cultured “G. hominis” strain clustered together with H. felis and the two “G. hominis” strains.
FIG. 3.

Phylogenetic tree of Helicobacter and “Gastrospirillum” species on the basis of 16S rDNA sequence distances. The scale bar indicates the percent difference in nucleotide sequences determined by measuring the lengths of the horizontal lines connecting any two species.
Protein profile.
Great similarities were seen in the protein profiles from the cultured “G. hominis,” H. pylori, and H. felis strains, as shown in Fig. 4. The isolated “G. hominis” strain seemed, however, to possess a major 23-kDa protein which was absent from H. pylori and H. felis. All three strains seemed to have a protein of 120 kDa, and this protein is assumed to be the cytotoxin-associated gene product CagA in H. pylori.
FIG. 4.
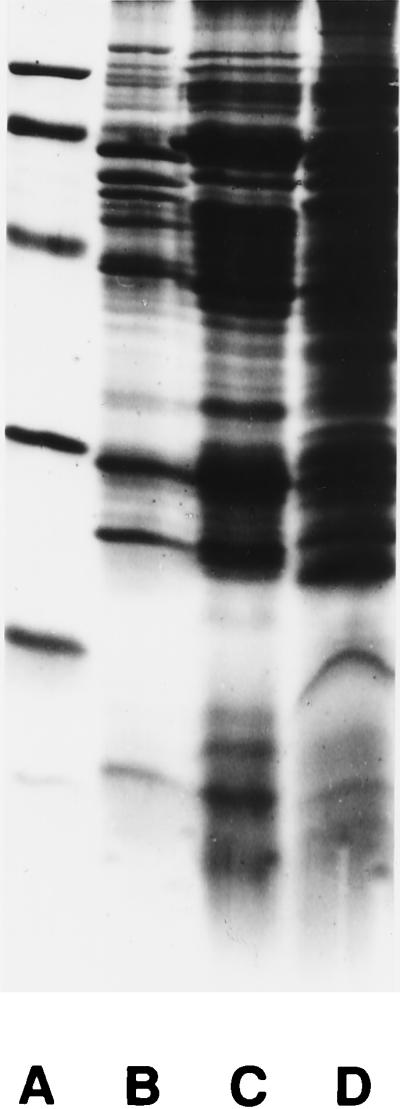
Protein profiles of H. pylori (lane B), H. felis (lane C), and “G. hominis” (lane D) visualized by Coomassie blue staining and sodium dodecyl sulfate-polyacrylamide gel electrophoresis with a 15% separation gel. The relative molecular mass standards (lane A) are 97.4, 66.2, 42.7, 31.0, 21.5, and 14.4 kDa. The most pronounced difference seen in the major bands is the 23-kDa band seen in “G. hominis” but which is not seen in H. pylori and H. felis.
DISCUSSION
On the basis of 16S rDNA sequencing, the cultured “G. hominis” strain clustered together with H. felis, Helicobacter bizzozeronii, H. salomonis, “Gastrospirillum,” Helicobacter acinonychis, Helicobacter nemestrinae, and H. pylori (Fig. 3). A close evolutionary relationship exists between H. felis, H. bizzozeronii, H. salomonis, and “G. hominis” (29, 36). The 16S rDNA sequence similarity between the members of this cluster ranges from 93.6 to 98.8% (Fig. 3). At similarity levels above 97 to 97.5%, 16S rDNA sequencing is not suited for the differentiation of species (38). The 16S rDNA sequence of the cultured “G. hominis” strain is 96.8 to 97.7% similar to the 16S rDNA sequences from the other species mentioned above. On the basis of this fact, a separate phylogenetic position of the cultured “G. hominis” isolate is possible, but no conclusion can be drawn.
The cluster of “Gastrospirillum,” H. felis, H. acinonychis, H. nemestrinae, and H. pylori had a characteristic antibiogram (nalidixic acid resistance and cephalothin sensitivity) which was different from those for other Helicobacter spp. (29). This may be a simple way of distinguishing this cluster from other clusters. No other clusters of Helicobacter spp. had a specific antibiogram.
“G. hominis” grew with distinct, small, translucent colonies that could not be distinguished from those of H. pylori, in contrast to many other spiral-shaped Helicobacter spp. such as H. felis and “Flexispira rappini,” which usually grow with a uniform translucent layer. H. felis could, however, be grown as small translucent colonies, and these colonies were usually smaller than those of H. pylori and “G. hominis.” Microscopy of the colonies revealed a pleomorphic mixture of curved and spiral-shaped organisms. The majority of the organisms could not be distinguished from H. pylori, even though the organisms seen in sections from gastric biopsy specimens were rather uniform and resembled “G. hominis,” but no H. pylori-like organisms were seen. The minority of the organisms were spiral shaped with three to eight tight coils and a wavelength of 1 μm. The proportion of spiral-shaped organisms in the colonies depended on the incubation time, with the proportion changing from more curved rods in young cultures to more spiral-shaped rods in older cultures. Thus, the microorganisms seemed to be able to change their morphologies. Recently, we cultured another “G. hominis”-like organism from a human gastric mucosa. This strain did not reveal any spiral forms during the first two subcultures. The human origin and the colony morphology exclude the possibility that other Helicobacter spp. of the cluster except H. pylori were present. The presence of H. pylori was excluded by microscopy of the colonies or histological sections. The ultrastructure revealed no periplasmic filaments in “G. hominis,” which is in contrast to the case for most other spiral-shaped Helicobacter spp. However, Eaton et al. (12) found that H. felis had the ability to loose the periplasmic filaments during subculture and that the filaments were absent from two primary isolates. In this respect “G. hominis” showed similarities to H. pylori and H. bizzozeronii, but unlike H. pylori, “G. hominis” had a bundle of up to 14 sheathed flagella in one or both ends of the bacterium. It is uncertain whether the flagella are unipolar or bipolar. In some cases they are seen only at one end of the bacterium. In some bacteria that are clearly dividing, the flagella are present at both ends, but flagella were also present at both ends of some bacteria that did not seem to be dividing. The growth conditions and biochemical reactions for “G. hominis” were very similar to those for H. pylori and H. felis except that H. felis did not grow at 41°C and did not grow as well on selective media. The discrepancies in nitrite and nitrate reactions for H. pylori in this study compared to those in other studies may be due to the test conditions because the test results are often positive after anaerobic incubation but not after microaerobic incubation. The protein profile revealed in “G. hominis” a protein of about 23 kDa which was absent from H. pylori and H. felis, but this may be a strain-specific protein and not a species-specific protein.
The cultured “G. hominis” isolate is a species of the genus Helicobacter that has not previously been cultured from specimens from humans and resembles the previously sequenced “Gastrospirillum” (30). It is probably of animal origin and cannot be distinguished from H. felis (12). The name “H. heilmannii” has been used for morphologically similar organisms, but both this species and “Gastrospirillum” spp. may consist of different related species or they may all belong to the species H. felis.
Characteristics of “G. hominis” (“H. heilmannii”).
“G. hominis” is a gram-negative, spiral-shaped, motile rod with three to eight coils and a wavelength of about 1 μm. It has up to 14 sheathed, uni- or bipolar flagella and no periplasmic filaments. “G. hominis” grows under microaerobic conditions at 36 and 41°C on 7% lysed, defibrinated horse blood agar plates within 3 to 7 days and can be subcultured under microaerobic but not anaerobic conditions on the same media used to culture H. pylori and H. felis. It grows as small translucent colonies that resemble those of H. pylori. “G. hominis” is motile and is oxidase, catalase, nitrite, nitrate, and urease positive, like H. pylori and H. felis. It produces alkaline phosphatase and arginine arylamidase; and, like H. pylori and H. felis, it is sensitive to cephalothin (30 μg/disc) and sensitive to most antibiotics, but in contrast to H. pylori and H. felis, it is resistant to nalidixic acid (30 μg/disc). The 16S rDNA sequence clusters “G. hominis” together with “Gastrospirillum,” H. felis, H. bizzozeronii, H. salmonii, H. nemestrinae, H. acinonychis, and H. pylori.
ACKNOWLEDGMENTS
We thank Bente Jensen, Jette Severinsen, and Elisabeth Brakti for excellent technical assistance.
REFERENCES
- 1.Andersen L P, Espersen F. Immunoglobulin G antibodies to Helicobacter pylori in patients with dyspeptic symptoms investigated by Western blot technique. J Clin Microbiol. 1992;30:1743–1751. doi: 10.1128/jcm.30.7.1743-1751.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Archer J R, Romero S, Ritchie A E, Hamacher M E, Steiner B M, Bryner J H, Schell R F. Characterization of an unclassified microaerophillic bacterium associated with gastroenteritis. J Clin Microbiol. 1988;26:101–105. doi: 10.1128/jcm.26.1.101-105.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bizzozero G. Sulle ghiandole tubulari del tubo gastroenterico e sui rapporti del loro coll’epitelio de rivestimento della mucosa. Atti R Accad Sci Torino. 1893;28:233–251. [Google Scholar]
- 4.Blom J, Mansa B, Wiik A. A study of Russell bodies in human monoclonal plasma cells by means of immunofluorescence and electron microscopy. Acta Pathol Microbiol Immunol Scand Sect A. 1971;84:335–349. doi: 10.1111/j.1699-0463.1976.tb00124.x. [DOI] [PubMed] [Google Scholar]
- 5.Boye, K., E. Høgdall, and M. Borre. Identification of bacteria using two degenerate 16S rDNA sequencing primers. Microbiol. Res., in press. [DOI] [PubMed]
- 6.Bronsdon M A, Goodwin C S, Sly L I, Chilvers T, Schoenknecht F D. Helicobacter nemestrinae sp. nov., a spiral bacterium found in the stomach of a pigtailed macaque (Macaca nemestrina) Int J Syst Bacteriol. 1991;41:148–153. doi: 10.1099/00207713-41-1-148. [DOI] [PubMed] [Google Scholar]
- 7.Dent J C, McNulty C A M, Uff J C, Wilkinson S P, Gear M W L. Spiral organisms in the gastric antrum. Lancet. 1987;ii:96. doi: 10.1016/s0140-6736(87)92754-1. [DOI] [PubMed] [Google Scholar]
- 8.Dewhirst F E, Seymour C, Fraser G J, Paster B J, Fox J G. Phylogeny of Helicobacter isolated from bird and swine feces and description of Helicobacter pametensis sp. nov. Int J Syst Bacteriol. 1994;44:553–560. doi: 10.1099/00207713-44-3-553. [DOI] [PubMed] [Google Scholar]
- 9.Doenges J L. Spirochetes in the gastric glands of Macacus rhesus and of man without related disease. Arch Pathol. 1939;27:469–477. [Google Scholar]
- 10.Dye K R, Marshall B J, Frierson H F, Jr, Guerrant R L, McCallum R W. Ultrastructure of another spiral organism associated with human gastritis. Dig Dis Sci. 1989;34:1787–1791. doi: 10.1007/BF01540059. [DOI] [PubMed] [Google Scholar]
- 11.Eaton K A, Dewhirst F, Radin M J, Fox J G, Paster B J, Krakowka S, Morgan D R. Helicobacter acinonyx sp. nov., isolated from cheetahs with gastritis. Int J Syst Bacteriol. 1993;43:99–106. doi: 10.1099/00207713-43-1-99. [DOI] [PubMed] [Google Scholar]
- 12.Eaton K A, Dewhirst F E, Paster B J, Tzellas N, Coleman B E, Paola J, Sherding R. Prevalence and varieties of Helicobacter species in dogs from random sources and pet dogs: animal and public health implications. J Clin Microbiol. 1996;34:3165–3170. doi: 10.1128/jcm.34.12.3165-3170.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fennell C L, Totten P A, Quinn T C, Patton D L, Holmes K K, Stamm W E. Characterization of Campylobacter-like organisms isolated from homosexual men. J Infect Dis. 1984;149:58–66. doi: 10.1093/infdis/149.1.58. [DOI] [PubMed] [Google Scholar]
- 14.Fischer R. “Gastrospirillum hominis,” another gastric spiral bacterium. Dig Dis. 1992;10:144–152. doi: 10.1159/000171352. [DOI] [PubMed] [Google Scholar]
- 15.Fox J G, Dewhirst F E, Tully J G, Paster B J, Yan L L, Taylor N S, Collins M J, Jr, Gorelick P L, Ward J M. Helicobacter hepaticus sp. nov., a microaerophillic bacterium isolated from levers and intestinal mucosal scrapings from mice. J Clin Microbiol. 1994;32:1238–1245. doi: 10.1128/jcm.32.5.1238-1245.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fox J G, Yan L L, Dewhirst F E, Paster B J, Shames B, Murphy J C, Hayward A, Belcher J C, Mendes E N. Helicobacter bilis sp. nov., a novel Helicobacter isolated from bile, liver and intestines of aged inbred mouse strains. J Clin Microbiol. 1995;33:445–454. doi: 10.1128/jcm.33.2.445-454.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Goodwin C S, Armstrong J A, Chilvers T, Peters M A, Collins D M, Sly L I, McConnell W, Harper W E S. Transfer of Campylobacter pylori and Campylobacter mustelae to Helicobacter gen. nov., as Helicobacter pylori comb. nov. and Helicobacter mustelae comb. nov., respectively. Int J Syst Bacteriol. 1989;39:397–405. [Google Scholar]
- 18.Hanninen M L, Happonen I, Saari S, Jalava K. Culture and characteristics of Helicobacter bizzozeronii, a new canine gastric Helicobacter sp. Int J Syst Bacteriol. 1996;46:160–166. doi: 10.1099/00207713-46-1-160. [DOI] [PubMed] [Google Scholar]
- 19.Heilmann K L, Borchard F. Gastritis due to spiral shaped bacteria other than Helicobacter pylori: clinical, histological, and ultrastructural findings. Gut. 1991;32:137–140. doi: 10.1136/gut.32.2.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Higgins D G, Sharp P M. Fast and sensitive multiple sequence alignment on a microcomputer. Comput Appl Biosci. 1989;5:151–153. doi: 10.1093/bioinformatics/5.2.151. [DOI] [PubMed] [Google Scholar]
- 21.Hodge D S, Borczyk A, Wat L-L. Evaluation of the indoxyl acetate hydrolysis test for the differentiation of campylobacters. J Clin Microbiol. 1990;28:1482–1483. doi: 10.1128/jcm.28.6.1482-1483.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Holck S, Ingeholm P, Blom J, Nørgaard A, Elsborg L, Adamsen S, Andersen L P. The histopathology of human gastric mucosa inhabited by Helicobacter heilmannii-like (Gastrospirillum hominis) organisms, including the first culturable case. APMIS. 1997;105:746–756. doi: 10.1111/j.1699-0463.1997.tb05080.x. [DOI] [PubMed] [Google Scholar]
- 23.Hwang M-N, Edere G M. Rapid hippurate hydrolysis method for presumptive identification of group B streptococci. J Clin Microbiol. 1975;1:114–115. doi: 10.1128/jcm.1.1.114-115.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Laemmli U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 25.Lee A, Phillips M W, O’Rourke J L, Paster B J, Dewhirst F E, Fraser G J, Fox J G, Sly L I, Romaniuk P J, Trust T J, Kouprach S. Helicobacter muridarum sp. nov., a microaerophillic helic bacterium with a novel ultrastructure isolated from the intestinal mucosa of rodents. Int J Syst Bacteriol. 1992;42:27–36. doi: 10.1099/00207713-42-1-27. [DOI] [PubMed] [Google Scholar]
- 26.Lockard V G, Boler R K. Ultrastructure of the spiralled microorganism in the gastric mucosa of dogs. Am J Vet Res. 1970;31:1453–1462. [PubMed] [Google Scholar]
- 27.Marshall B J, Warren J R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet. 1984;i:1311–1314. doi: 10.1016/s0140-6736(84)91816-6. [DOI] [PubMed] [Google Scholar]
- 28.McNulty C A M, Dent J C, Curry A, Uff J S, Ford G A, Gear M W L, Wilkinson S P. New spiral bacterium in gastric mucosa. J Clin Pathol. 1989;42:585–591. doi: 10.1136/jcp.42.6.585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mendes E N, Queiroz D M M, Dewhirst F E, Paster B J, Moura S B, Fox J G. Helicobacter trogontum sp. nov., isolated from the rat intestine. Int J Syst Bacteriol. 1996;46:916–921. doi: 10.1099/00207713-46-4-916. [DOI] [PubMed] [Google Scholar]
- 30.Morris A, Ali M R, Thomsen L, Hollis B. Tightly spiral shaped bacteria in the human stomach: another cause of active chronic gastritis? Gut. 1990;31:139–143. doi: 10.1136/gut.31.2.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Paster B J, Lee A, Fox J G, Dewhirst F E, Tordoff L A, Fraser G J, O’Rourke J L, Taylor N S, Ferrero R. Phylogeny of Helicobacter felis sp. nov. and related bacteria. Int J Syst Bacteriol. 1991;41:31–38. doi: 10.1099/00207713-41-1-31. [DOI] [PubMed] [Google Scholar]
- 32.Saitou N, Nei M. The Neighbor-joining method: a new method for reconstruction phylogenetic trees. Mol Biol Evol. 1987;4:406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 33.Salomon H. Über das Spirillum des Säugetiermagens und sein Verhalten zu den Belegzellen. Zentralbl Bakteriol Parasitenkd Infektionskr Hyg Abt 1. 1898;19:422–441. [Google Scholar]
- 34.Schauer D B, Ghori N, Falkow S. Isolation and characterization of “Flexispira rappini” from laboratory mice. J Clin Microbiol. 1993;31:2709–2714. doi: 10.1128/jcm.31.10.2709-2714.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sneath P H A, Sokal R R. Numerial taxonomy. W. H. San Francisco, Calif: Freeman & Co.; 1973. [Google Scholar]
- 36.Solnick J V, O’Rourke J, Lee A, Paster B J, Dewhirst F E, Tompkins L S. An uncultured gastric spiral organism is a newly identified Helicobacter in humans. J Infect Dis. 1993;168:379–385. doi: 10.1093/infdis/168.2.379. [DOI] [PubMed] [Google Scholar]
- 37.Solnick J V, O’Rourke J, Lee A, Tompkins L S. Molecular analysis of urease genes from newly identified uncultured species of Helicobacter. Infect Immun. 1994;62:1631–1638. doi: 10.1128/iai.62.5.1631-1638.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Stackebrandt E, Goebel B M. Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int J Syst Bacteriol. 1994;44:846–849. [Google Scholar]
- 39.Stanley J, Linton D, Burnens A P, Dewhirst F E, Owen R J, Porter A, On S L W, Costas M. Helicobacter canis sp. nov., a new species from dogs: an integrated study of phenotype and genotype. J Gen Microbiol. 1993;139:2495–2504. doi: 10.1099/00221287-139-10-2495. [DOI] [PubMed] [Google Scholar]
- 40.Stanley J, Linton D, Burnens A P, Dewhirst F E, On S L W, Porter A, Owen R J, Costas M. Helicobacter pullorum sp. nov.: polyphasic description of a new species from poultry which infects humans. Microbiology. 1994;140:3441–3449. doi: 10.1099/13500872-140-12-3441. [DOI] [PubMed] [Google Scholar]
- 41.Stolte M, Kroher G, Meining A, Morgner A, Bayerdorffer E, Bethke B. A comparison of Helicobacter pylori and Helicobacter heilmannii gastritis. A matched control study involving 404 patients. Scand J Gastroenterol. 1997;32:28–33. doi: 10.3109/00365529709025059. [DOI] [PubMed] [Google Scholar]
- 42.Warren J R, Marshall B J. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet. 1983;i:1273–1275. [PubMed] [Google Scholar]
- 43.Weber A F, Hasa O, Sautter J H. Some observations concerning the presence of spirilla in the fundic glands of dogs and cats. Am J Vet Res. 1958;19:677–680. [PubMed] [Google Scholar]
- 44.Weber K, Osborn M. The reliability of molecular weight determination by dodecyl sulfate-polyacrylamide gel electrophoresis. J Biol Chem. 1969;244:4406–4412. [PubMed] [Google Scholar]