

Summary
Biofilms are communities of bacteria, fungi or yeasts that form on diverse biotic or abiotic surfaces, and play important roles in pathogenesis and drug resistance. A generic saw palmetto oil inhibited biofilm formation by Staphylococcus aureus, Escherichia coli O157:H7 and fungal Candida albicans without affecting their planktonic cell growth. Two main components of the oil, lauric acid and myristic acid, are responsible for this antibiofilm activity. Their antibiofilm activities were observed in dual‐species biofilms as well as three‐species biofilms of S. aureus, E. coli O157:H7 and C. albicans. Transcriptomic analysis showed that lauric acid and myristic acid repressed the expressions of haemolysin genes (hla and hld) in S. aureus, several biofilm‐related genes (csgAB, fimH and flhD) in E. coli and hypha cell wall gene HWP1 in C. albicans, which supported biofilm inhibition. Also, saw palmetto oil, lauric acid and myristic acid reduced virulence of three microbes in a nematode infection model and exhibited minimal cytotoxicity. Furthermore, combinatorial treatment of fatty acids and antibiotics showed synergistic antibacterial efficacy against S. aureus and E. coli O157:H7. These results demonstrate that saw palmetto oil and its main fatty acids might be useful for controlling bacterial infections as well as multispecies biofilms.
The present study describes the antibiofilm activities of a generic saw palmetto oil and its main components, lauric acid, and myristic acid, on dual‐species biofilms as well as three‐species biofilms of Staphylococcus aureus, Escherichia coli O157:H7, and fungal Candida albicans. We demonstrate that lauric acid and myristic acid represses haemolysin genes and decreases cell surface hydrophobicity in S. aureus, represses the expression of several biofilm related genes in E. coli, and inhibits hyphal development in C. albicans. Also, lauric acid and myristic acid reduced virulence of three microbes in a nematode infection model and exhibited minimal cytotoxicity. Furthermore, combinatorial treatment of fatty acids and antibiotics showed synergistic antibacterial efficacy against S. aureus and E. coli O157:H7.
Introduction
Most antimicrobial agents aim to inhibit microbial growth, ideally without harming the host or the environment. However, the overuse of these agents has caused worldwide emergence of drug‐resistant pathogens. Bacterial and fungal biofilms play important roles in antimicrobial resistance and in a variety of device‐related infections (Donlan, 2002; Hall‐Stoodley et al., 2004; Handorf et al., 2019). Furthermore, various pathogenic microorganisms often form multispecies biofilms, which further increase tolerance to antimicrobial agents (Carlson and Johnson, 1985; Harriott and Noverr, 2011). Hence, alternative non‐toxic approaches are needed to control single and polymicrobial biofilm formation. Unlike current antimicrobial strategies, it is important to identify biofilm inhibitors that do not inhibit planktonic cell growth (Wright, 2015), which is one of an antivirulence strategy that aims to reduce pathogenesis while its consequences not affecting bacterial growth, reducing the chance of the emergence of drug resistance (Cegelski et al., 2008).
Fatty acids are widespread in all forms of life, and more than 70 natural fatty acids have been identified (Kenar et al., 2017). The antimicrobial activities of various fatty acids, mostly above hundred µg ml−1 doses, have been well reported against diverse microorganisms (Desbois and Smith, 2010; Yoon et al., 2018). However, several studies recently reported that fatty acids exhibit antibiofilm and antivirulence activities against bacteria and fungi at concentrations much lower than their MICs (Kumar et al., 2020).
Extract of the saw palmetto (Seronoa repens) berry is widely used as an alternative treatment of prostatic hyperplasia (Bent et al., 2006). The main components of saw palmetto products are fatty acids (more than 90%) and phytosterols, and the prominent fatty acids are caprylic, capric, lauric, myristic, palmitic, oleic, linolenic/linoleic and stearic acid (Penugonda and Lindshield, 2013; Booker et al., 2014). Various fatty acids, such as capric, lauric, palmitic, oleic, linoleic/linolenic and eicosadienoic acid, have been shown to possess antibiofilm activity against microbial biofilms of S. aureus and fungal C. albicans (Stenz et al., 2008; Davies and Marques, 2009; Murzyn et al., 2010; Thibane et al., 2012; Lee et al., 2017, 2020; Kim et al., 2019, 2018), and diverse fatty acid derivatives are important cell‐to‐cell signals in various plant‐associated bacteria (Ryan et al., 2015; Zhou et al., 2017). Hence, we hypothesized that fatty acid‐rich saw palmetto products and their main components would have similar antibiofilm activities against other microbes and mixed microbes.
Most microorganisms exist as multispecies consortia and often form polymicrobial biofilms that have been shown to provide several advantages for the residing species such as increased tolerance against antimicrobial agents and protozoan grazing, as well as increased virulence in infections (Roder et al., 2016). For example, fungal C. albicans often forms biofilms with other bacteria like S. aureus, P. aeruginosa and Enterococcus species on host and environmental surfaces, which formed more rapidly and have a greater antimicrobial tolerance (Harriott and Noverr, 2009). While the significance of polymicrobial biofilms in human pathology has been increasingly acknowledged, biofilm inhibition of dual‐ and three‐species has rarely been reported as of yet.
Hence, in the present study, the antibiofilm activity of saw palmetto oil was initially investigated against eleven microbes. GC‐MS analysis revealed the presence of the main fatty acid components in the oil and the antibiofilm activity of the fatty acids were further tested against single strain of S. aureus, E. coli O157:H7 and C. albicans, as well as dual‐species and three‐species. To understand how fatty acids inhibit biofilm formation, confocal laser scanning microscopy (CLSM), scanning electron microscopy (SEM) and transcriptomic analysis were used to investigate morphological changes, biofilm formation, the hyphal growth of C. albicans and gene expression. Furthermore, a Caenorhabditis elegans model was used to investigate the antimicrobial effects of fatty acids and their cytotoxic nature. In addition, synergistic effects of fatty acids with antimicrobial agents were further studied.
Results
Saw palmetto oil and main fatty acid components inhibited biofilm formation of various microorganisms
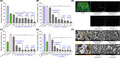
The antibiofilm activity of saw palmetto oil was initially investigated against eleven microorganisms. Saw palmetto oil at concentrations of 50 µg ml−1 significantly inhibited biofilm formation by four S. aureus strains including two methicillin‐resistant strains, a Staphylococcus epidermidis strain, two pathogenic E. coli strains and two C. albicans strains, while it could not change biofilm formation by A. baumannii and P. aeruginosa (Fig. 1A). Most notably, saw palmetto oil inhibited biofilm formation by two C. albicans strains more than 85% even at 20 µg ml−1.
Fig. 1.
Antibiofilm activities of saw palmetto oil and its main fatty acids. Biofilm formations by individual microbes were quantified in the presence of saw palmetto oil after incubation for 24 h in 96‐well plates (A). Chemical structures of main fatty acids in the saw palmetto oil (B). Biofilm inhibition by fatty acids of S. aureus MSSA 6538 (C), E. coli O157:H7 (D), C. albicans DAY185 (E) and other Staphylococcus strains (F) were quantified in the presence of lauric acid (C12:0), myristic acid (C14:0), palmitic acid (C16:0) and oleic acid (C18:1) after incubation for 24 h in 96‐well plates at 37 °C. n = 3 biologically independent samples. Error bars indicate standard deviations. *P < 0.05 versus non‐treated controls. Biofilm formations on polystyrene plates were observed in the presence of saw palmetto oil at 100 µg ml−1, C12:0 or C14:0 at 20 µg ml−1, by confocal laser microscopy (G). Scale bars represent 100 µm. At least two independent cultures were performed for microscopic experiments, and at least 10 random positions were assayed.
GC‐MS identified 25 fatty acids in saw palmetto oil (Table S3). Main components of saw palmetto oil were lauric acid (34.3%), myristic acid (14.3%), palmitic acid (9.6%) and oleic acid (29.1%) (Fig. 1B). The compositions of saw palmetto oil identified by GC‐MS analysis concur with previous studies (Penugonda and Lindshield, 2013; Booker et al., 2014).
The antibiofilm potencies of four main fatty acids, lauric acid (C12:0), myristic acid (C14:0), palmitic acid (C16:0) and oleic acid (C18:1), were further investigated against three genera such as S. aureus, E. coli and C. albicans. It appears that lauric acid and myristic acid are most active against three microorganisms. For example, lauric acid and myristic acid at 20 μg ml−1 inhibited biofilm formation by five Staphylococcus, E. coli O157:H7 and C. albicans strains more than 50% (Fig. 1C–F). For the S. aureus strain, oleic acid above 20 μg ml−1 also appreciably decreased its biofilm formation, while palmitic acid did not (Fig. 1C). However, palmitic acid and oleic acid did not change the biofilm formation by E. coli O157:H7 and C. albicans (Fig. 1D and E). Similar to the result observed with saw palmetto oil, lauric acid and myristic acid at 5 µg ml−1 mostly inhibited C. albicans biofilms (Fig. 1E), which is matched with previous reports (Prasath et al., 2019; Lee et al., 2020). The antibiofilm activity of oleic acid against S. aureus strains was previously reported (Stenz et al., 2008; Davies and Marques, 2009; Lee et al., 2017). However, it is the first observation that lauric acid and myristic acid also have the inhibitory effect on S. aureus biofilm formation. Also, we report for the first time that lauric acid and myristic acid exhibit antibiofilm activity against E. coli O157:H7.
Hence, it can be concluded that the antibiofilm activities of saw palmetto oil against S. aureus, E. coli O157:H7 and C. albicans were largely due to the presence of lauric acid and myristic acid, which were focused later on in the study. However, the MICs of saw palmetto oil and two active fatty acids against all eleven microorganisms were found to be > 500 µg ml−1 (Table S1) and three fatty acids up to 200 μg ml−1 did not much affect the planktonic cell growth (Fig. S1). These results indicate that saw palmetto oil, lauric acid and myristic acid inhibited microbial biofilm formation without affecting planktonic cell growth.
Biofilm inhibition on the bottom of 96‐well plate was analysed by CSLM. While three microorganisms formed dense biofilms (thickness > 40 µm and almost 100% surface coverage) in non‐treated controls of three microbes, saw palmetto oil at 100 µg ml−1 dramatically reduced biofilm densities and thicknesses (Fig. 1G). Also, both lauric acid and myristic acid at 20 µg ml−1 significantly inhibited biofilm formation by S. aureus, E. coli O157:H7 and C. albicans. However, saw palmetto oil and two fatty acids, myristoleic and lauric acids, at concentrations up to 100 µg ml−1 could not eradicate the pre‐existing biofilms of S. aureus, C. albicans and E. coli O157:H7 (Fig. S2).
Antibiofilm activities of saw palmetto oil and main fatty acids against polymicrobial biofilms of S. aureus, E. coli O157:H7 and C. albicans
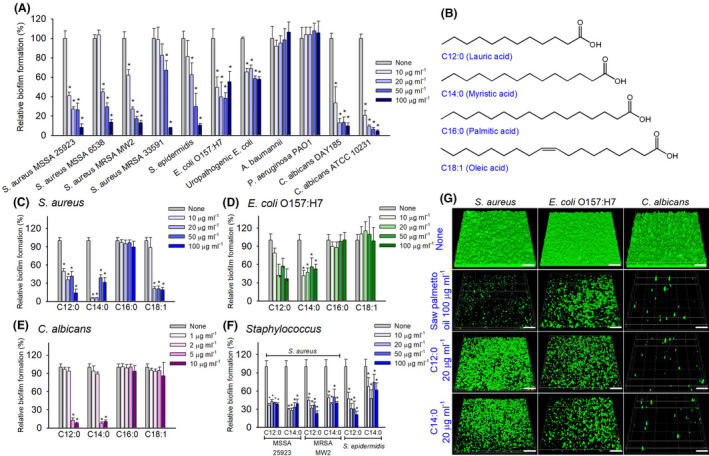
The efficacies of saw palmetto oil and fatty acids on dual and three‐species biofilms of S. aureus, E. coli O157:H7 and C. albicans were further studied. We have found proper media and inoculum to form biofilms of three sets of dual species and one set of three species, such as S. aureus and E. coli O157:H7 in LB medium, and S. aureus and C. albicans, E. coli O157:H7 and C. albicans in the 1:1 mixture of PDB and LB, and three species together in the 1:2 mixture of PDB and LB. In the non‐treated controls of two‐ or three‐species, decent biofilm formation (1 ~ 2 at OD570) was observed in all four cases (Fig. 2A–D). As expected, both lauric acid and myristic acid dose‐dependently inhibited biofilm formation of dual‐ and three‐species biofilms (Fig. 2A–D). Most notably, lauric acid and myristic acid at 10 μg ml−1 inhibited three‐species biofilm formation by 90% after culture for 24 h. To the best of our knowledge, it is the first report of three‐species biofilm inhibition of S. aureus, E. coli O157:H7 and C. albicans. Importantly, saw palmetto oil dose‐dependently inhibited biofilm formation of these four cases. Similar to single biofilms, it appears that saw palmetto oil and two fatty acids most significantly inhibited multispecies biofilms including C. albicans (Fig. 2B–D).
Fig. 2.
Biofilm inhibition by saw palmetto oil, lauric acid and myristic acid in the polymicrobial models. Inhibitory effects of saw palmetto oil, lauric acid (C12:0) and myristic acid (C14:0) against dual biofilms of S. aureus MSSA 6538 and E. coli O157:H7 in LB medium (A), dual biofilms of S. aureus 6538 and C. albicans DAY185 in a 1:1 mixture of PDB and LB media (B), dual biofilms of E. coli O157:H7 and C. albicans in a 1:1 mixture of PDB and LB media (C) and three‐species biofilms of S. aureus, E. coli O157:H7 and C. albicans biofilms in a 1:2 mixture of PDB and LB media (D) were determined after culture for 24 h at 37 °C. n = 3 biologically independent samples. *P < 0.05 versus non‐treated controls. CLSM (E) and SEM (F) observation of three‐species biofilms of S. aureus, E. coli O157:H7 and C. albicans. In the inset, the large cells are C. albicans (indicated by blue triangle), the small round cells are S. aureus (indicated by red triangle), and rod type E. coli O157:H7 cells (indicated by yellow triangle). The yellow scale bar represents 100 µm, the white scale bar represents 15 µm, and the red scale bar in inset represents 3 µm. At least two independent cultures were performed for microscopic experiments, and at least 10 random positions were assayed.
CLSM analysis confirmed that saw palmetto oil or two fatty acids at 20 μg ml−1 significantly decreased three‐species biofilm formation on the bottom of 96‐well plates (Fig. 2E). SEM observation also revealed the inhibitory effect of saw palmetto oil, lauric acid or myristic acid on biofilm formation of three‐species (Fig. 2F). We observed polymicrobial associations between S. aureus, E. coli O157:H7 and C. albicans in non‐treated control samples. It appears that C. albicans formed large hyphae and a few yeast cells that were much larger than round S. aureus cells and rod type E. coli O157:H7 cells. Notably, saw palmetto oil, lauric acid or myristic acid obviously reduced hyphal formation of C. albicans and S. aureus and E. coli O157:H7 cells were widely distributed.
Differential gene expressions induced by saw palmetto oil, lauric acid and myristic acid
To investigate the molecular basis responsible for biofilm inhibition by saw palmetto oil, lauric acid and myristic acid, qRT‐PCR was performed against three individual microorganisms with various biofilm‐related genes.
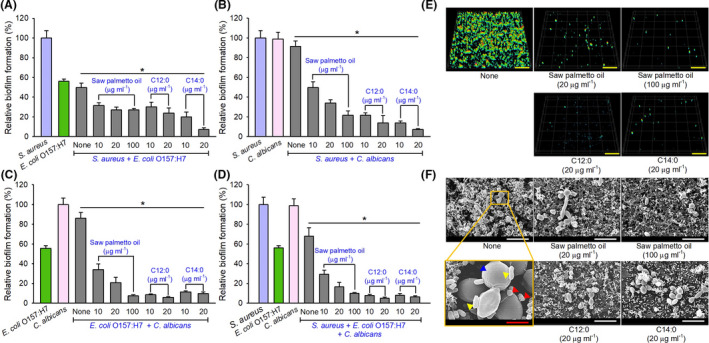
In S. aureus cells, saw palmetto oil and lauric acid, but not myristic acid, dramatically reduced the expression of alpha‐haemolysin hla gene by 20‐ and 32‐fold, respectively, and downregulated delta‐haemolysin (hld, also known as RNAIII), while other genes, such as agrA, aur, icaA and sigB, were not affected (Fig. 3A). It has been reported that hld (RNAIII) stimulates hla translation (Morfeldt et al., 1995). Hence, hla repression is partially due to the down‐regulation of hld by saw palmetto oil and lauric acid. Interestingly, saw palmetto oil and lauric acid upregulated sarA, a positive regulator of S. aureus biofilm development (Beenken et al., 2003; Valle et al., 2003). Previously, two omega fatty acids repressed the expression of hla and hld (Kim et al., 2018). Hence, it appears that antibiofilm activities of lauric acid and other omega fatty acids are partially due to down‐regulating the gene expression of the hla and hld.
Fig. 3.
Effects of fatty acids on transcriptional profiles, cell hydrophobicity and hyphal formation. S. aureus MSSA 6538 (A), E. coli O157:H7 (B) or C. albicans DAY185 (C) was incubated with or without saw palmetto oil at 100 μg ml−1, lauric acid (C12:0) at 20 μg ml−1 or myristic acid (C14:0) at 20 μg ml−1 for 6, 4 or 6 h at 37 °C with shaking at 250 rpm respectively. Transcriptional profiles were acquired by qRT‐PCR. Fold changes represent change in the transcriptions of treated vs. non‐treated controls. The experiment was conducted in duplicate (four qRT‐PCRs were tested per gene). *P < 0.05 vs. non‐treated controls (None). The cell surface hydrophobicity was determined after incubation for 24 h at 37 °C with or without saw palmetto oil, lauric acid (C12:0) and myristic acid (C14:0) in S. aureus 6538 (D), E. coli O157:H7 (E) or C. albicans DAY185 (F). Inhibition of hyphal filamentation by fatty acids in C. albicans DAY185 after incubation for 24 h at 37 °C in PDB medium (G). The scale bar represents 50 µm. At least two independent cultures were performed for microscopic experiments, and at least 10 random positions were analysed.
In E. coli O157:H7 cells, the effects of lauric acid and myristic acid on the expression of biofilm‐related genes were more significant than that of saw palmetto oil. Notably, lauric acid and myristic acid repressed the expression of two fimbriae genes (csgAB), three motility genes (fimH, flhD and motB) and two quorum‐sensing genes (luxRS) (Fig. 3B).
In C. albicans cells, it was notable that both saw palmetto oil and lauric acid repressed the expression of HWP1 gene (Fig. 3C) that is essential for hyphal development and their expressions have been shown to be correlated with cell elongation and biofilm formation (Nobile et al., 2006). Also, notably, the expression of CHT4 encoding chitinase was induced by saw palmetto oil and the two fatty acids. It was reported that CHT4 transcription was decreased upon yeast‐to‐hyphal switch (Prasath et al., 2019), which suggested that saw palmetto oil and two fatty acids could inhibit hyphal formation. Also, it was recently reported that medium‐chain fatty acids repressed the expression of HWP1 gene via mimicking of the quorum‐sensing molecule farnesol (Lee et al., 2020). Current results partially support the hypothesis of the reduction of biofilm formation (Fig. 1E) and hyphae formation (Fig. 2F).
Changes of hydrophobicity in S. aureus and hyphal inhibition in C. albicans by fatty acids
Cell surface hydrophobicity and biofilm formation are closely linked in several bacteria as hydrophobic cells adhere more to hydrophobic surfaces (Krasowska and Sigler, 2014). Hence, cell surface hydrophobicity was measured in the presence of fatty acids in the three strains (Fig. 3D–F). Interestingly, saw palmetto oil, lauric acid and myristic acid increased hydrophilicity, which means cells are less hydrophobic in S. aureus, while they did not change the hydrophilicity in either E. coli O157:H7 or C. albicans cells (Fig. 3E and F). These results partially explain the inhibition of S. aureus biofilm formation by fatty acids in hydrophobic polystyrene surfaces.
Since the yeast‐to‐hypha transition is known to be a prerequisite of biofilm development by C. albicans (Chandra et al., 2001), the effect of fatty acids on hyphal formation was observed. Saw palmetto oil, lauric acid and myristic acid substantially suppressed the hyphal transition, while untreated control cells were predominantly hyphal cells (Fig. 3G). Recently, several medium‐chain fatty acids were reported to inhibit hyphal growth in C. albicans (Lee et al., 2020). These results and transcriptomic results support that lauric acid and myristic acid potently inhibit hypha formation and reduce biofilm formation by C. albicans and even multispecies biofilm formation including that of C. albicans.
Antivirulence activities of saw palmetto oil, lauric acid and myristic acid in the nematode model with minimal cytotoxicity
Biofilm formation and cell adhesion are key virulence factors in animal hosts. Caenorhabditis elegans model is viewed as an alternative to the use of rodent models in various studies in biology, especially in the field of bacterial infection. Infections of pathogenic microorganisms, such as S. aureus (Sifri et al., 2003), E. coli O157:H7 (Chou et al., 2013) or C. albicans (Pukkila‐Worley et al., 2009) could kill the nematodes.
As expected, C. elegans was well survived with E. coli OP50, which is a common food source for the nematode, while the lifespan of C. elegans was much decreased in the presence of each pathogen. Saw palmetto oil, lauric acid and myristic acid were found to markedly prolong C. elegans survival in the presence of S. aureus (Fig. 4A), E. coli O157:H7 (Fig. 4B) or C. albicans (Fig. 4C). For example, nematode survival was almost 100 per cent without pathogen for 10 days, whereas in the presence of saw palmetto oil at 100 μg ml−1, or myristic acid at 20 μg ml−1, more than 55% of the nematodes survived. This result is in line with the observed downregulation of biofilm formation (Fig. 1) and repression of haemolysin genes (Fig. 3A). Similar antivirulence characteristics of saw palmetto oil, lauric acid and myristic acid are observed in the infection of E. coli O157:H7 (Fig. 4B) or C. albicans (Fig. 4C).
Fig. 4.
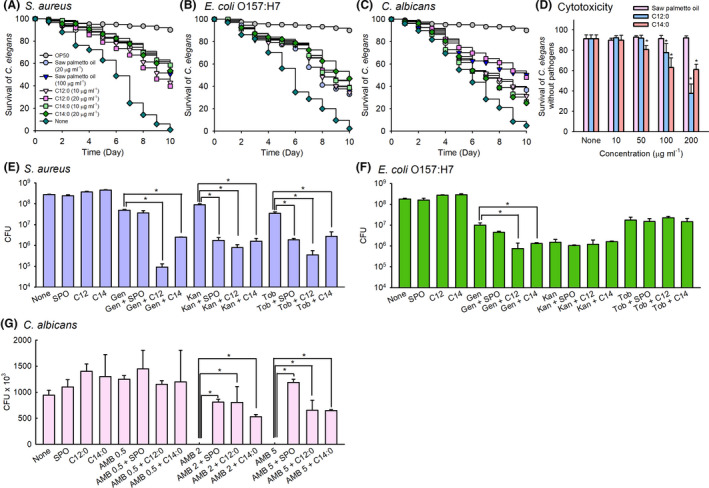
Anti‐virulence activities of fatty acids and co‐administration of fatty acids and antimicrobial agents. Nematode survivals after exposure to S. aureus MSSA 6538 (A), E. coli O157:H7 (B) or C. albicans DAY185 (C) cells in the presence of saw palmetto oil, lauric acid (C12:0) or myristic acid (C14:0). Chemical toxicities were determined by treating non‐infected C. elegans for 4 days (D). OP50 indicates E. coli OP50‐fed controls, and none indicates non‐treated controls. *P < 0.05 vs. non‐treated controls (None). OP50 represents E. coli OP50. Cell survivals were measured in the presence of antibiotic gentamicin (10 μg ml−1), kanamycin (50 μg ml−1) or tobramycin (10 μg ml−1) against S. aureus (E) or E. coli O157:H7 (F) and antifungal amphotericin B (0.5, 2 and 5 μg ml−1) against C. albicans (G). *P < 0.05 vs. non‐treated controls. None: non‐treated control. SPO: saw palmetto oil (100 μg ml−1), fatty acids C12:0 and C14:0 (20 μg ml−1), Gen: gentamicin, Kan: kanamycin, Tob: tobramycin, AMB: amphotericin B. n = 2 biologically independent samples.
In addition, to examine the cytotoxicity of saw palmetto oil, lauric acid and myristic acid, C. elegans survival was investigated with feeding only E. coli OP50, but without any pathogens. No toxic effects were observed when non‐infected nematodes were exposed to saw palmetto oil at concentrations up to 200 μg ml−1, while lauric acid and myristic acid at 100 and 200 μg ml−1 were found to be a mild titre for nematodes (Fig. 4D). These cytotoxicity results of fatty acids partially matched with a previous report (Lima et al., 2002). However, it is notable that lauric acid and myristic acid exhibit antibiofilm activity at concentrations of 10 or 20 μg ml−1, much less than the level of their cytotoxicity.
Combinatory efficacies of fatty acids and antimicrobial agents
Since combinatorial approaches have been proposed to enhance antimicrobial efficacy (Wright, 2016), combinatory efficacies of fatty acids and antimicrobial agents on microbial growth were investigated against individual microorganisms. First, antibiotic gentamicin alone at 10 μg ml−1 was tested against S. aureus or E. coli O157:H7 and antifungal amphotericin B alone at 0.5, 2 or 5 μg ml−1 was tested against C. albicans. As expected, aminoglycoside gentamicin partially killed more than 90% of the two bacteria within 1 h (Fig. 4E and F), which matched with previous reports (Weinstein et al., 1963; Yoshizawa et al., 1998). Also, amphotericin B above 2 μg ml−1 nearly abolished C. albicans growth (Fig. 4G), which also matched with a previous report (Brajtburg et al., 1990). Second, the combinations of fatty acids and antimicrobial agents were tested against three individual microorganisms. It is notable that the addition of lauric acid (C12:0) or myristic acid (C14:0) at 20 μg ml−1 caused much less survival of S. aureus (Fig. 4E) and E. coli O157:H7 (Fig. 4F), while lauric acid or myristic acid alone at 20 μg ml−1 did not affect their cell growth at all (MICs > 500 μg ml−1). It appears that the antibacterial efficacy of gentamicin was significantly enhanced in the presence of lauric acid or myristic acid, while saw palmetto oil at 100 μg ml−1 showed a marginal effect on the antibiotic efficacy. Further combinatorial assays with kanamycin and tobramycin led to different results as the efficacy of kanamycin and tobramycin in the presence of fatty acids was enhanced only in S. aureus (Fig. 4E), but not in E. coli O157:H7 (Fig. 4F).
In the case of C. albicans, unexpected results were obtained as the addition of saw palmetto oil, lauric acid or myristic acid significantly increased the survival of C. albicans (Fig. 4G). It was confirmed with two different concentrations of amphotericin B at 2 or 5 μg ml−1. Hence, it appears that fatty acids negatively affect the antifungal efficacy, which should be further investigated.
Discussion
This study presents for the first time that lauric acid and myristic acid from saw palmetto oil suppress three‐species biofilm formation by S. aureus, E. coli O157:H7 and C. albicans with minimal cytotoxic effect. The mode of action of the active fatty acids was partially revealed, and the synergistic antimicrobial efficacies of fatty acids were observed.
While broad‐spectrum antimicrobial activities of fatty acids at high doses were well documented due to its amphipathic nature (Desbois and Smith, 2010; Yoon et al., 2018), recent studies reported that fatty acids at concentrations much lower than their MICs exhibit antibiofilm and antivirulence activities against bacteria and fungi (Kumar et al., 2020). Current study demonstrates that lauric acid and myristic acid abundant in saw palmetto oil could inhibit biofilm formation of single‐, dual‐ and three‐species biofilm models of S. aureus, E. coli O157:H7 and C. albicans without affecting their planktonic cell growth. Lauric acid is mainly found in coconut oil, laurel oil and palm kernel oil, and myristic acid is found in nutmeg butter, coconut oil, laurel oil, palm kernel oil, saw palmetto oil and even breast milk (Beare‐Rogers et al., 2001). The wide‐ranging distributions of lauric acid and myristic acid indicate the importance of protective roles as not only antimicrobial components but also antibiofilm inhibitors against single or multispecies consortia. The action mechanisms of lauric acid and myristic acid are varied in three tested microbes since the three species have different systems of controlling their biofilm formation.
In S. aureus, several fatty acids such as palmitic acid (C16:0), oleic acid (C18:1) (Stenz et al., 2008), linoleic acid (C18:2), eicosadienoic acids (C20:1) (Lee et al., 2017) and omega‐3 fatty acids, such as cis‐4,7,10,13,16,19‐docosahexaenoic acid (DHA) and cis‐5,8,11,14,17‐eicosapentaenoic acid (EPA) (Kim et al., 2018; Kumar et al., 2020) at sub‐MIC levels, inhibited biofilm formation and haemolytic activity in S. aureus. Alpha‐haemolysin (Hla) is a major cytotoxic agent and is positively associated with biofilm formation in S. aureus (Caiazza and O'Toole, 2003), which is supported by the current results of the repression of hla and hld by fatty acids (Fig. 3A). Also, it has been suggested that both saturated and unsaturated fatty acids at high doses can be incorporated through a kinase‐dependent pathway, ultimately influencing the membrane fluidity in S. aureus (Liaw et al., 2004). Therefore, it appears that the decrease of haemolysin production and membrane fluidity is possible mechanism of lauric acid and myristic acid to control S. aureus biofilm formation.
In E. coli O157:H7, both lauric acid and myristic acid influenced the expression of various biofilm‐related genes, such as fimbriae genes (csgAB), motility genes (fimH, flhD and motB) and quorum‐sensing genes (luxRS) (Fig. 3B). Fimbriae are important for E. coli O157:H7 biofilm formation (Ryu and Beuchat, 2005; Rendón et al., 2007), motility plays a role in E. coli biofilm formation (Pratt and Kolter, 1998; Ren et al., 2001), and autoinducer‐2 (AI‐2) quorum sensing also positively influences the development of biofilm of commensal E. coli (González Barrios et al., 2006; Hardie and Heurlier, 2008) and of the E. coli O157:H7 strain (Bansal et al., 2008; Vikram et al., 2010). Therefore, the antibiofilm activities of lauric acid and myristic acid are due to the downregulation of the gene expression of fimbriae, motility and AI‐2 genes. Previously, palmitic acid showed antibiofilm activity against E. coli with an IC50 of 7.1 µM (Sandai et al., 2016; Kumar et al., 2020). Also, cis‐2‐decenoic acid induced biofilm dispersal of E. coli, S. aureus, C. albicans and other microbes (Davies and Marques, 2009) and cis‐2‐decenoic acid combined with antibiotics and disinfectants was found to effectively remove more than 80% of pre‐formed biofilms of E. coli (Rahmani‐Badi et al., 2014).
In C. albicans, the switching of yeast cells to hyphal cells is considered to play an important role in biofilm formation and the pathogenesis of fungal infections (Douglas, 2003). It was recently reported that lauric acid inhibited C. albicans biofilm formation by downregulation of several hyphal and biofilm‐related genes (Lee et al., 2020). Also, it was reported that myristic acid inhibited biofilm and hyphal formation by C. albicans, probably targeting several proteins involved in ergosterol synthesis, sphingolipid metabolism, multidrug resistance and the oxidative stress (Prasath et al., 2019). Transcriptomic results in the current study partially matched with the previous reports as the expression of HWP1 downregulated by fatty acids (Fig. 3C). Furthermore, the gene expression of chitinase CHT4 was induced by fatty acids (Fig. 3C) that also might play a role in the inhibition of hyphal and biofilm formation. While speculative, it appears inhibition of the hyphae formation and biofilm formation by various fatty acids is not rare in the ecosystem.
Biofilms often consist of multiple species of bacteria residing in close proximity to one another in natural environmental conditions (Elias and Banin, 2012), but current knowledge of the polymicrobial biofilms is still very limited (Roder et al., 2016). Especially, biofilm controls of three species are rarely reported, while various studies of dual‐species biofilm have been reported. For example, the efficacy of disinfectants against S. aureus, C. albicans and P. aeruginosa in multispecies biofilms was reported and multispecies biofilms were always less susceptible than single species biofilms (Kart et al., 2014). In several dual‐species biofilm studies including C. albicans, hyphal formation is a dominant factor as bacteria like S. aureus and E. coli can adhere to hyphae (Peters and Noverr, 2013; De Brucker et al., 2015; Kong et al., 2016). Recently, linoleic acid from centipede oil (Kim et al., 2020) and nepodin (Lee et al., 2019) was reported to reduce hyphal formation and biofilm formation by C. albicans and the numbers of S. aureus adhered. In this study, we developed for the first time an in vitro three‐species biofilm model including S. aureus, E. coli O157:H7 and C. albicans and further demonstrated that lauric acid and myristic acid could inhibit biofilm formation of three‐species biofilms (Fig. 2D–F). Based on SEM observation, S. aureus and E. coli O157:H7 can adhere to larger C. albicans cells in the form of yeast or hyphae cells (Fig. 2F) and saw palmetto oil and myristic acid repressed the expression of HWP1 (hyphal wall protein 1 gene) (Fig. 3C). Therefore, in polymicrobial biofilms including C. albicans, the ability of hyphal morphogenesis and its culture conditions are crucial factors for overall biofilm formation. Since several medium‐chain fatty acids also repressed the expression of HWP1 gene via mimicking of the quorum‐sensing molecule farnesol (Lee et al., 2020), it is highly possible that lauric acid and myristic acid would also interfere the quorum sensing of C. albicans. Notably, lauric acid and myristic acid repressed the expression of quorum‐sensing genes (luxRS) in E. coli (Fig. 3B), but not any quorum‐sensing gene (agrA) in S. aureus (Fig. 3A). It is intriguing that fatty acids may affect inter‐kingdom signalling systems in mono‐ and multi‐species biofilms.
Combinatorial therapy has gained importance due to the limited efficacy of antimicrobial agents against drug‐resistant pathogens and their poor penetration abilities within biofilms. Several reports showed that fatty acids can be used as an antibiotic adjuvant since combinatorial therapy of fatty acids and antimicrobial agents synergistically diminished biofilm formation and decreased bacterial pathogenesis (Ells et al., 2009; Rahmani‐Badi et al., 2014; Sepehr et al., 2014; Kumar et al., 2020), probably due to change of phospholipid composition and membrane permeability (Eder et al., 2017; Hobby et al., 2019). In the current study, synergistic antimicrobial efficacies of fatty acids were observed in two bacteria, S. aureus and in E. coli O157:H7, but not in fungal C. albicans (Fig. 4E–G). For C. albicans cells, antifungal amphotericin B binds with ergosterol, a component of fungal cell membranes, forming pores that cause rapid leakage of monovalent ions (K+, Na+, H+ and Cl−) and produced reactive oxygen species and subsequent fungal cell death (Brajtburg et al., 1990; Haraguchi et al., 1996). In this study, antifungal activity of amphotericin B was reversed by lauric acid and myristic acid. Since lauric acid and myristic acid inhibited hyphal formation (Fig. 3), while speculative, amphotericin B is likely to be more effective against hyphal cells than yeast cells. To enhance the antibiotic efficacy, combinatorial therapy and fatty acids can be simultaneously used in tandem or combination and also drug delivery systems using chitosan, natural polymers, liposomes or nanoparticles, further enhancing the bioavailability and decrease drug cytotoxicity.
The emergence of drug‐resistant microorganisms has driven the development of novel antibiotics, antifungals and even adjuvants. Control of drug‐resistant biofilm formation is important in pathogens such as S. aureus, E. coli O157:H7 and C. albicans. This study shows the antibiofilm activities of saw palmetto oil and its main fatty acids, lauric acid and myristic acid, against each pathogen and mixture of two or three species. The efficacies were confirmed in vivo in a C. elegans model with minimal cytotoxicity. Since many saw palmetto oils are commercially available as health supplements and lauric acid and myristic acid are naturally present in various plant oils, these fatty acid products could be relatively safe to be applied in multispecies infections as potent antibiofilm agents and antibiotic adjuvants.
Furthermore, fatty acid‐rich plant oils can be used to control pathogenic biofilms of single and multiple microbes by coating medical devices, synthesizing composites of polymer fatty acids, and producing emulsion with fatty acids for medical and cosmetic purposes. Also, our findings indicate that other fatty acids could be utilized for combinatorial therapy against drug‐resistant bacteria. Experiments such as bacterial adhesion and antivirulence efficacy using epithelial cells or animal model could be further investigated.
Experimental procedures
Strains, chemicals and culture materials
Eleven microorganisms used in the present study, medium, minimum inhibitory concentrations (MICs) and inoculum size for MIC measurement are listed in Table S1. All media (Luria‐Bertani (LB), tryptic soy broth (TSB), potato dextrose broth (PDB), nutrient broth and agar) were purchased from Becton Dickinson (Franklin Lakes, NJ, USA). Minimum inhibitory concentrations (MICs) were determined (Table S1) according to the Clinical Laboratory Standards Institute (CLSI) for bacteria and yeasts (CLSI, 2015; CLSI, 2017). MIC was defined as the lowest concentration that inhibited cell growth by 80% as assessed by spectrophotometry and colony counting. Experiments were performed using at least three independent cultures.
A generic saw palmetto oil was purchased from Amazon (Seattle, USA). Four fatty acids, namely dodecanoic acid (lauric acid), tetradecanoic acid (myristic acid), hexadecanoic acid (palmitic acid), and 9‐octadecenoic acid (oleic acid), and antimicrobial agents, gentamicin and amphotericin B, were purchased from Sigma‐Aldrich (St. Louis, USA) or TCI Co. (Tokyo, Japan). Dimethyl sulfoxide (DMSO) was used to dissolve all fatty acids, and 0.1% (v/v) DMSO was used as the negative control; at this concentration, it did not affect bacterial growth or biofilm formation.
Biofilm assay in 96‐well plate
Biofilm formation of various microbes was produced on 96‐well polystyrene plates, as previously described (Lee et al., 2019). Briefly, overnight cultures at an initial turbidity of OD 0.05 (~ 108 or 107 CFU ml−1) for bacteria and OD 0.1 (~ 105 CFU ml−1) for Candida at 600 nm (Table S1) were then inoculated into a proper culture media (final volume 300 μl) with or without fatty acids and incubated for 24 h without shaking at 37 °C. Biofilm cells that adhered to 96‐well plates (SPL Life Sciences, Pocheon, Korea) were stained with 0.1% crystal violet Sigma‐Aldrich (St. Louis, USA) for 20 min, washed repeatedly with sterile distilled water and resuspended in 95% ethanol. Plates were read at 570 nm, and results are presented as the means of at least six repetitions from three independent cultures. The percentage of inhibition ratio represents the relative biofilm formation (100 × biofilm formation with chemical/biofilm formation of untreated control).
Gas chromatograph/mass spectroscopy (GC‐MS)
The components of saw palmetto oil were separated and analysed by gas chromatography‐mass spectroscopy (GC‐MS) using an Agilent 6890N GC and SP‐2560 (Supelco, Sigma‐Aldrich, St. Louis, USA) with a silica capillary column (100 m × 0.25 mm i.d., film thickness 0.25 mm). Column temperature conditions and the derivatization (methylation) of fatty acids were as previously described (Kim et al., 2018). Triundecanoin (C11:0) was used as the internal standard, and quantifications were performed by integrating areas and correcting for fatty acid methylation. Supelco 37 components FAME Mix (Supelco) were used as the reference standard.
Biofilm observations by CSLM and SEM
Single, dual or triple biofilms were formed on 96‐well polystyrene plates with or without fatty acids without shaking for 24 h at 37 °C. Planktonic cells were then removed by washing with distilled water three times, and single or dual biofilms were stained with carboxyfluorescein diacetate succinimidyl ester (Invitrogen, Molecular Probes, Inc., Eugene, USA) (Lee et al., 2016). Plate bases were then visualized using a 488 nm Ar laser (emission 500 to 550 nm) under a confocal laser microscope (Nikon Eclipse Ti, Tokyo, Japan). Two independent cultures were performed under each experimental condition and at least 10 random positions were assayed.
Scanning electron microscopy was also used to observe multispecies biofilm formation on nylon membranes, as previously described (Kim et al., 2016). Briefly, a nylon membrane (Merck Millipore, Burlington, USA) was cut into 0.5 × 0.5 cm pieces and placed in 96‐well plates containing single or mixed species grown with or without fatty acids and incubated for 24 h at 37 °C. PDB medium was used for C. albicans biofilm, a 1:1 mixture of PDB and LB media was used for the dual C. albicans and S. aureus biofilms, a 1:1 mixture of PDB and LB media was used for the dual biofilms of C. albicans and E. coli O157:H7, or a 1:2 mixture of PDB and LB media was used for the triple biofilms of S. aureus, C. albicans and E. coli O157:H7. Cells that adhered to the nylon membrane were fixed with glutaraldehyde (2.5%) and formaldehyde (2%) for 24 h and then post‐fixed using osmium and dehydrated using an ethanol series (50, 70, 80, 90, 95 and 100%) and isoamyl acetate. After critical‐point drying, cells were examined and imaged using an S‐4100 scanning electron microscope (Hitachi, Tokyo, Japan) at a voltage of 15kV according to the manufacturer’s instructions.
Quantitative Real‐Time PCR (qRT‐PCR)
Three sets of transcriptomic analyses were performed with S. aureus, E. coli O157:H7 and C. albicans cells. For the S. aureus, the previous method was used (Kim et al., 2018). S. aureus MSSA 6538 cells were inoculated into 25 ml of LB broth at 37 °C in 250‐ml flasks at a starting OD600 of 0.05 and then incubated for 6 h with shaking at 250 rpm in the presence or absence of saw palmetto oil (100 µg ml−1), lauric acid (20 µg ml−1) and myristic acid (20 µg ml−1). For the E. coli O157:H7, the previous method was used (Kim et al., 2015). E. coli O157:H7 cells were inoculated into 25 ml of LB broth at 37 °C in 250‐ml flasks at a starting OD600 of 0.05 and then incubated for 4 h with shaking at 250 rpm in the presence or absence of saw palmetto oil (100 µg ml−1), lauric acid (20 µg ml−1) and myristic acid (20 µg ml−1). RNase inhibitor (RNAlater, Ambion, TX, USA) was then added, and cells were immediately chilled for 30 s in a dry ice bath having 95% ethanol to prevent RNA degradation. Cells were then harvested by centrifugation at 16 600 g for 1 min, and total RNA was isolated using a Qiagen RNeasy mini Kit (Valencia, USA) according to the manufacturer’s instructions. For the C. albicans, another previous method was used (Lee et al., 2018). 25 ml of C. albicans DAY185 at an initial turbidity of 0.1 at OD600 (~ 105 CFU ml−1) was inoculated into PDB broth in 250 ml and incubated for 6 h at 37 °C with agitation (250 rpm) in the presence or absence of saw palmetto oil (100 µg ml−1), lauric acid (20 µg ml−1) and myristic acid (20 µg ml−1). To prevent RNA degradation, RNase inhibitor (RNAlater, Ambion, TX, USA) was added to cells and cells were immediately chilled for 30 s in a dry ice bath having 95% ethanol to prevent RNA degradation. Total RNA was isolated using a hot acidic phenol method, and RNA was purified using a Qiagen RNeasy mini Kit (Valencia, USA) according to the manufacturer’s instructions.
qRT‐PCR was used to determine the expressions of various biofilm‐related genes (agrA, aur, hla, hld, icaA, sarA and sigB in S. aureus, csgA, csgB, fimH, flhD, luxR, luxS and motB in E. coli O157:H7, ALS1, ALS3, CDR1, CDR2, CHT4, CYR1, ECE1, ERG9, ERG10, HWP1, MTS1, RAS1, RBT5 and UME6, in C. albicans). The specific primers used for qRT‐PCR are listed in Table S2. Three housekeeping genes, 16s rRNA, rrsG or RDN18, were used, respectively, and the expression of housekeeping genes was not affected by fatty acids. The qRT‐PCR was done using SYBR Green master mix (Applied Biosystems, Foster City, USA) and an ABI StepOne Real‐Time PCR System (Applied Biosystems) according to the manufacturer’s instructions. At least two independent cultures were used.
Cell‐Surface hydrophilicity assay
Cell‐surface hydrophobicity was analysed as previously reported (Rosenberg et al., 1980). Briefly, cell cultures were harvested by centrifugation at 7000 g for 5 min after 20‐h incubation in LB with shaking at 250 rpm at 37 °C. Harvested cells were mixed with hexadecane (TCI chemicals, Tokyo, Japan) at a ratio of 6:1 by vortexing for 90 s. Mixtures were allowed to stand for 30 min at room temperature to enable phase separation. Aqueous phase absorbance was measured at OD600. Experiments were performed using four independent cultures per each material.
Observation of C. albicans hyphae
To observe hyphal formation, microscopic imaging system was used as previously described (Lee et al., 2020). Briefly, C. albicans DAY185 cells were re‐inoculated at 1:50 dilution in 2 ml of PDB medium with or without fatty acids (0, 10, 20 or 100 μg ml−1) and incubated for 24 h at 37 °C without shaking. After the incubation, cells were mixed and visualized using an iRiS™ Digital Cell Imaging System (Logos Bio Systems, Anyang, Korea) according to the manufacturer’s instructions. At least four independent cultures were used.
Antivirulence and cytotoxicity assays of fatty acids in the nematode model
To investigate the effects of fatty acids on the virulence of S. aureus, E. coli O157:H7 or C. albicans, we used C. elegans strain fer‐15(b26); fem‐1(hc17), as previously described (Kim et al., 2020). Briefly, approximately 30 nematodes were added to each well of 96‐well plates containing M9 for S. aures and E. coli O157:H7 or PDB:M9 (20:80) mixture for C. albicans infection. Also, the untreated control and saw palmetto oil (20 or 100 µg ml−1), lauric acid (10 or 20 µg ml−1) or myristic acid (10 or 20 µg ml−1)‐treated S. aures, E. coli O157:H7 (~ 108, 107 CFU ml−1 respectively) or C. albicans cells (~ 105 CFU ml−1) were added into the wells containing nematodes. Plates were then incubated for 10 days at 25 °C without shaking. For cytotoxicity assays, 30 non‐infected nematodes were pipetted into the single wells of a 96‐well plate containing M9 buffer and fatty acid (saw palmetto oil, lauric acid or myristic acid) at final concentrations of 0, 10, 50, 100 or 200 µg ml−1 without pathogens. Plates were then incubated for 4 days at 25 °C without shaking. Three independent experiments were performed in triplicate. Results are expressed as percentages of live nematodes (survival), determined by responses to platinum wire touching. Observations were made using an iRiS™ Digital Cell Imaging System (Logos Bio Systems, Anyang, Korea) according to the manufacturer’s instructions.
Combinatory treatment of fatty acids and antimicrobial agents
Combinatorial efficacies of fatty acids and antimicrobial agents were analysed on S. aureus, E. coli O157:H7 and C. albicans. Briefly, overnight cultures at an initial turbidity of OD 0.05 for bacteria and OD 0.1 for Candida at 600 nm were inoculated into a proper culture media (final volume 1000 μl) with antimicrobial agents, gentamicin (10 µg ml−1), kanamycin (50 μg ml−1), tobramycin (10 μg ml−1) or amphotericin B (0.5, 2, 5 µg ml−1) and/or fatty acids (saw palmetto oil at 100 µg ml−1, C12 or C14 at 20 µg ml−1). Cells were incubated for 1 h with shaking at 37 °C. Cell mixtures were diluted serially in PBS and plated on proper culture media. CFU was determined by colony counting after 24‐h incubation at 37 °C. At least four repetitions from two independent cultures were performed.
Statistical analysis
Replication numbers for assays are provided above, and results are expressed as means ± standard deviations. The statistical analysis was performed using one‐way ANOVA followed by Dunnett’s test using SPSS version 23 (SPSS Inc., Chicago, IL, USA). P values of < 0.05 were considered significant, and asterisks indicate significant differences between treated and untreated samples.
Conflict of interest
None declared.
Supporting information
Fig. S1. Planktonic cell growth in the presence of fatty acids.
Fig. S2. Dispersal of established biofilms by fatty acids.
Table S1. Strains and culture media used in this study and MICs of saw palmetto oil, lauric acid, and myristic acid.
Table S2. Primer sequences used for qRT‐PCR.
Table S3. GC‐MS analysis of saw palmetto oil.
Acknowledgements
This research was supported by grants from the Basic Science Research Program through the NRF funded by the Ministry of Education (2021R1I1A3A04037486 to J.‐H. Lee and 2019R1C1C1008329 to Y.‐G. Kim), the NRF grant funded by the Korea government (MSIT) (2021R1A2C1008368 to J. Lee) and by a grant from the Priority Research Centers Program through the NRF funded by the Ministry of Education (2014R1A6A1031189).
Microb. Biotechnol. (2022) 15(2), 590–602
Funding information
This research was supported by grants from the Basic Science Research Program through the NRF funded by the Ministry of Education (2021R1I1A3A04037486 to J.‐H. Lee, 2019R1C1C1008329 to Y.‐G. Kim), the NRF grant funded by the Korea government (MSIT) (No. 2021R1A2C1008368 to J. Lee), and by a grant from the Priority Research Centers Program through the NRF funded by the Ministry of Education (2014R1A6A1031189).
References
- Bansal, T. , Jesudhasan, P. , Pillai, S. , Wood, T.K. , and Jayaraman, A. (2008) Temporal regulation of enterohemorrhagic Escherichia coli virulence mediated by autoinducer‐2. Appl Microbiol Biotechnol 78: 811–819. [DOI] [PubMed] [Google Scholar]
- Beare‐Rogers, J. , Dieffenbacher, A. , and Holm, J.V. (2001) Lexicon of lipid nutrition. Pure Appl Chem 73: 685–744. [Google Scholar]
- Beenken, K.E. , Blevins, J.S. , and Smeltzer, M.S. (2003) Mutation of sarA in Staphylococcus aureus limits biofilm formation. Infect Immun 71: 4206–4211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bent, S. , Kane, C. , Shinohara, K. , Neuhaus, J. , Hudes, E.S. , Goldberg, H. , and Avins, A.L. (2006) Saw palmetto for benign prostatic hyperplasia. N Engl J Med 354: 557–566. [DOI] [PubMed] [Google Scholar]
- Booker, A. , Suter, A. , Krnjic, A. , Strassel, B. , Zloh, M. , Said, M. , and Heinrich, M. (2014) A phytochemical comparison of saw palmetto products using gas chromatography and H‐1 nuclear magnetic resonance spectroscopy metabolomic profiling. J Pharm Pharmacol 66: 811–822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brajtburg, J. , Powderly, W.G. , Kobayashi, G.S. , and Medoff, G. (1990) Amphotericin B: current understanding of mechanisms of action. Antimicrob Agents Chemother 34: 183–188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caiazza, N.C. , and O'Toole, G.A. (2003) Alpha‐toxin is required for biofilm formation by Staphylococcus aureus . J Bacteriol 185: 3214–3217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlson, E. , and Johnson, G. (1985) Protection by Candida albicans of Staphylococcus aureus in the establishment of dual infection in mice. Infect Immun 50: 655–659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cegelski, L. , Marshall, G.R. , Eldridge, G.R. , and Hultgren, S.J. (2008) The biology and future prospects of antivirulence therapies. Nat Rev Microbiol 6: 17–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chandra, J. , Kuhn, D.M. , Mukherjee, P.K. , Hoyer, L.L. , McCormick, T. , and Ghannoum, M.A. (2001) Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J Bacteriol 183: 5385–5394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou, T.C. , Chiu, H.C. , Kuo, C.J. , Wu, C.M. , Syu, W.J. , Chiu, W.T. , and Chen, C.S. (2013) Enterohaemorrhagic Escherichia coli O157:H7 Shiga‐like toxin 1 is required for full pathogenicity and activation of the p38 mitogen‐activated protein kinase pathway in Caenorhabditis elegans . Cell Microbiol 15: 82–97. [DOI] [PubMed] [Google Scholar]
- CLSI (2015) Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically. Approved Standard M07–A10, 10th edn. Wayne, PA, USA: Clinical and Laboratory Standards Institute. [Google Scholar]
- CLSI (2017) Reference Methods for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Standard M27, 4th edn. Wayne, PA, USA: Clinical and Laboratory Standards Institute. [Google Scholar]
- Davies, D.G. , and Marques, C.N. (2009) A fatty acid messenger is responsible for inducing dispersion in microbial biofilms. J Bacteriol 191: 1393–1403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Brucker, K. , Tan, Y. , Vints, K. , De Cremer, K. , Braem, A. , Verstraeten, N. , et al. (2015) Fungal β‐1,3‐glucan increases ofloxacin tolerance of Escherichia coli in a polymicrobial E. coli/Candida albicans biofilm. Antimicrob Agents Chemother 59: 3052–3058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desbois, A.P. , and Smith, V.J. (2010) Antibacterial free fatty acids: activities, mechanisms of action and biotechnological potential. Appl Microbiol Biotechnol 85: 1629–1642. [DOI] [PubMed] [Google Scholar]
- Donlan, R.M. (2002) Biofilms: microbial life on surfaces. Emerg Infect Dis 8: 881–890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Douglas, L.J. (2003) Candida biofilms and their role in infection. Trends Microbiol 11: 30–36. [DOI] [PubMed] [Google Scholar]
- Eder, A.E. , Munir, S.A. , Hobby, C.R. , Anderson, D.M. , Herndon, J.L. , Siv, A.W. , et al. (2017) Exogenous polyunsaturated fatty acids (PUFAs) alter phospholipid composition, membrane permeability, biofilm formation and motility in Acinetobacter baumannii . Microbiology 163: 1626–1636. [DOI] [PubMed] [Google Scholar]
- Elias, S. , and Banin, E. (2012) Multi‐species biofilms: living with friendly neighbors. FEMS Microbiol Rev 36: 990–1004. [DOI] [PubMed] [Google Scholar]
- Ells, R. , Kock, J.L.F. , Van Wyk, P.W.J. , Botes, P.J. , and Pohl, C.H. (2009) Arachidonic acid increases antifungal susceptibility of Candida albicans and Candida dubliniensis . J Antimicrob Chemother 63: 124–128. [DOI] [PubMed] [Google Scholar]
- González Barrios, A.F. , Zuo, R. , Hashimoto, Y. , Yang, L. , Bentley, W.E. , and Wood, T.K. (2006) Autoinducer 2 controls biofilm formation in Escherichia coli through a novel motility quorum‐sensing regulator (MqsR, B3022). J Bacteriol 188: 305–316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall‐Stoodley, L. , Costerton, J.W. , and Stoodley, P. (2004) Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol 2: 95–108. [DOI] [PubMed] [Google Scholar]
- Handorf, O. , Schnabel, U. , Bosel, A. , Weihe, T. , Bekeschus, S. , Graf, A.C. , et al. (2019) Antimicrobial effects of microwave‐induced plasma torch (MiniMIP) treatment on Candida albicans biofilms. Microb Biotechnol 12: 1034–1048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haraguchi, H. , Kuwata, Y. , Inada, K. , Shingu, K. , Miyahara, K. , Nagao, M. , and Yagi, A. (1996) Antifungal activity from Alpinia galanga and the competition for incorporation of unsaturated fatty acids in cell growth. Planta Med 62: 308–313. [DOI] [PubMed] [Google Scholar]
- Hardie, K.R. , and Heurlier, K. (2008) Establishing bacterial communities by 'word of mouth': LuxS and autoinducer 2 in biofilm development. Nat Rev Microbiol 6: 635–643. [DOI] [PubMed] [Google Scholar]
- Harriott, M.M. , and Noverr, M.C. (2009) Candida albicans and Staphylococcus aureus form polymicrobial biofilms: Effects on antimicrobial resistance. Antimicrob Agents Chemother 53: 3914–3922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harriott, M.M. , and Noverr, M.C. (2011) Importance of Candida‐bacterial polymicrobial biofilms in disease. Trends Microbiol 19: 557–563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hobby, C.R. , Herndon, J.L. , Morrow, C.A. , Peters, R.E. , Symes, S.J.K. , and Giles, D.K. (2019) Exogenous fatty acids alter phospholipid composition, membrane permeability, capacity for biofilm formation, and antimicrobial peptide susceptibility in Klebsiella pneumoniae . MicrobiologyOpen 8: e00635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kart, D. , Tavernier, S. , Van Acker, H. , Nelis, H.J. , and Coenye, T. (2014) Activity of disinfectants against multispecies biofilms formed by Staphylococcus aureus, Candida albicans and Pseudomonas aeruginosa . Biofouling 30: 377–383. [DOI] [PubMed] [Google Scholar]
- Kenar, J.A. , Moser, B.R. , and List, G.R. (2017) Chapter 2 – Naturally occurring fatty acids: source, chemistry, and uses. In Fatty Acids. Ahmad, M.U. (ed). Urbana, IL, USA: AOCS Press, pp. 23–82. [Google Scholar]
- Kim, H.‐S. , Ham, S.‐Y. , Jang, Y. , Sun, P.‐F. , Park, J.‐H. , Lee, J.H. , and Park, H.D. (2019) Linoleic acid, a plant fatty acid, controls membrane biofouling via inhibition of biofilm formation. Fuel 253: 754–761. [Google Scholar]
- Kim, Y.‐G. , Lee, J.‐H. , Kim, S.‐I. , Baek, K.‐H. , and Lee, J. (2015) Cinnamon bark oil and its components inhibit biofilm formation and toxin production. Int J Food Microbiol 195: 30–39. [DOI] [PubMed] [Google Scholar]
- Kim, Y.‐G. , Lee, J.‐H. , Gwon, G. , Kim, S.‐I. , Park, J.G. , and Lee, J. (2016) Essential oils and eugenols inhibit biofilm formation and the virulence of Escherichia coli O157:H7. Sci Rep 6: 36377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, Y.‐G. , Lee, J.‐H. , Raorane, C.J. , Oh, S.T. , Park, J.G. , and Lee, J. (2018) Herring oil and omega fatty acids inhibit Staphylococcus aureus biofilm formation and virulence. Front Microbiol 9: 1241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, Y.‐G. , Lee, J.‐H. , Park, J.G. , and Lee, J. (2020) Inhibition of Candida albicans and Staphylococcus aureus biofilms by centipede oil and linoleic acid. Biofouling 36: 126–137. [DOI] [PubMed] [Google Scholar]
- Kong, E.F. , Tsui, C. , Kucharikova, S. , Andes, D. , Van Dijck, P. , and Jabra‐Rizk, M.A. (2016) Commensal protection of Staphylococcus aureus against antimicrobials by Candida albicans biofilm matrix. MBio 7: e01365‐01316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krasowska, A. , and Sigler, K. (2014) How microorganisms use hydrophobicity and what does this mean for human needs? Front Cell Infect Microbiol 4: 112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar, P. , Lee, J.‐H. , Beyenal, H. , and Lee, J. (2020) Fatty acids as antibiofilm and antivirulence agents. Trends Microbiol 28: 753–768. [DOI] [PubMed] [Google Scholar]
- Lee, J.‐H. , Kim, Y.‐G. , Ryu, S.Y. , and Lee, J. (2016) Calcium‐chelating alizarin and other anthraquinones inhibit biofilm formation and the hemolytic activity of Staphylococcus aureus . Sci Rep 6: 19267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, J.‐H. , Kim, Y.‐G. , Park, J.G. , and Lee, J. (2017) Supercritical fluid extracts of Moringa oleifera and their unsaturated fatty acid components inhibit biofilm formation by Staphylococcus aureus . Food Control 80: 74–82. [Google Scholar]
- Lee, J.‐H. , Kim, Y.‐G. , Choi, P. , Ham, J. , Park, J.G. , and Lee, J. (2018) Antibiofilm and antivirulence activities of 6‐gingerol and 6‐shogaol against Candida albicans due to hyphal inhibition. Front Cell Infect Microbiol 8: 299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, J.‐H. , Kim, Y.‐G. , Khadke, S.K. , Yamano, A. , Watanabe, A. , and Lee, J. (2019) Inhibition of biofilm formation by Candida albicans and polymicrobial microorganisms by nepodin via hyphal‐growth suppression. ACS Infect Dis 5: 1177–1187. [DOI] [PubMed] [Google Scholar]
- Lee, J.‐H. , Kim, Y.‐G. , Khadke, S.K. , and Lee, J. (2020) Antibiofilm and antifungal activities of medium‐chain fatty acids against Candida albicans via mimicking of the quorum‐sensing molecule farnesol. Microb Biotechnol 1–14. 10.1111/1751-7915.13710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liaw, S.J. , Lai, H.C. , and Wang, W.B. (2004) Modulation of swarming and virulence by fatty acids through the RsbA protein in Proteus mirabilis . Infect Immun 72: 6836–6845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lima, T.M. , Kanunfre, C.C. , Pompeia, C. , Verlengia, R. , and Curi, R. (2002) Ranking the toxicity of fatty acids on Jurkat and Raji cells by flow cytometric analysis. Toxicol in Vitro 16: 741–747. [DOI] [PubMed] [Google Scholar]
- Morfeldt, E. , Taylor, D. , von Gabain, A. , and Arvidson, S. (1995) Activation of alpha‐toxin translation in Staphylococcus aureus by the trans‐encoded antisense RNA, RNAIII. EMBO J 14: 4569–4577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murzyn, A. , Krasowska, A. , Stefanowicz, P. , Dziadkowiec, D. , and Lukaszewicz, M. (2010) Capric acid secreted by S. boulardii inhibits C. albicans filamentous growth, adhesion and biofilm formation. PLoS One 5: e12050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobile, C.J. , Nett, J.E. , Andes, D.R. , and Mitchell, A.P. (2006) Function of Candida albicans adhesin Hwp1 in biofilm formation. Eukaryot Cell 5: 1604–1610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penugonda, K. , and Lindshield, B.L. (2013) Fatty acid and phytosterol content of commercial saw palmetto supplements. Nutrients 5: 3617–3633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters, B.M. , and Noverr, M.C. (2013) Candida albicans‐Staphylococcus aureus polymicrobial peritonitis modulates host innate immunity. Infect Immun 81: 2178–2189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasath, K.G. , Sethupathy, S. , and Pandian, S.K. (2019) Proteomic analysis uncovers the modulation of ergosterol, sphingolipid and oxidative stress pathway by myristic acid impeding biofilm and virulence in Candida albicans . J Proteomics 208: 103503. [DOI] [PubMed] [Google Scholar]
- Pratt, L.A. , and Kolter, R. (1998) Genetic analysis of Escherichia coli biofilm formation: roles of flagella, motility, chemotaxis and type I pili. Mol Microbiol 30: 285–293. [DOI] [PubMed] [Google Scholar]
- Pukkila‐Worley, R. , Peleg, A.Y. , Tampakakis, E. , and Mylonakis, E. (2009) Candida albicans hyphal formation and virulence assessed using a Caenorhabditis elegans infection model. Eukaryot Cell 8: 1750–1758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahmani‐Badi, A. , Sepehr, S. , Mohammadi, P. , Soudi, M.R. , Babaie‐Naiej, H. , and Fallahi, H. (2014) A combination of cis‐2‐decenoic acid and antibiotics eradicates pre‐established catheter‐associated biofilms. J Med Microbiol 63: 1509–1516. [DOI] [PubMed] [Google Scholar]
- Ren, D. , Sims, J.J. , and Wood, T.K. (2001) Inhibition of biofilm formation and swarming of Escherichia coli by (5Z)‐4‐bromo‐5‐(bromomethylene)‐3‐butyl‐2(5H)‐furanone. Environ Microbiol 3: 731–736. [DOI] [PubMed] [Google Scholar]
- Rendon, M.A. , Saldana, Z. , Erdem, A.L. , Monteiro‐Neto, V. , Vazquez, A. , Kaper, J.B. , et al. (2007) Commensal and pathogenic Escherichia coli use a common pilus adherence factor for epithelial cell colonization. Proc Natl Acad Sci USA 104: 10637–10642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roder, H.L. , Sorensen, S.J. , and Burmolle, M. (2016) Studying bacterial multispecies biofilms: where to start? Trends Microbiol 24: 503–513. [DOI] [PubMed] [Google Scholar]
- Rosenberg, M. , Gutnick, D. , and Rosenberg, E. (1980) Adherence of bacteria to hydrocarbons: a simple method for measuring cell‐surface hydrophobicity. FEMS Microbiol Lett 9: 29–33. [Google Scholar]
- Ryan, R.P. , An, S.Q. , Allan, J.H. , McCarthy, Y. , and Dow, J.M. (2015) The DSF family of cell‐cell signals: an expanding class of bacterial virulence regulators. PLoS Pathog 11: e1004986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryu, J.H. , and Beuchat, L.R. (2005) Biofilm formation by Escherichia coli O157:H7 on stainless steel: effect of exopolysaccharide and Curli production on its resistance to chlorine. Appl Environ Microbiol 71: 247–254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandai, D. , Tabana, Y.M. , Ouweini, A.E. , and Ayodeji, I.O. (2016) Resistance of Candida albicans biofilms to drugs and the host immune system. Jundishapur J Microbiol 9: e37385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sepehr, S. , Rahmani‐Badi, A. , Babaie‐Naiej, H. , and Soudi, M.R. (2014) Unsaturated fatty acid, cis‐2‐decenoic acid, in combination with disinfectants or antibiotics removes pre‐established biofilms formed by food‐related bacteria. PLoS One 9: e101677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sifri, C.D. , Begun, J. , Ausubel, F.M. , and Calderwood, S.B. (2003) Caenorhabditis elegans as a model host for Staphylococcus aureus pathogenesis. Infect Immun 71: 2208–2217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stenz, L. , Francois, P. , Fischer, A. , Huyghe, A. , Tangomo, M. , Hernandez, D. , et al. (2008) Impact of oleic acid (cis‐9‐octadecenoic acid) on bacterial viability and biofilm production in Staphylococcus aureus . FEMS Microbiol Lett 287: 149–155. [DOI] [PubMed] [Google Scholar]
- Thibane, V.S. , Ells, R. , Hugo, A. , Albertyn, J. , van Rensburg, W.J.J. , Van Wyk, P.W.J. , et al. (2012) Polyunsaturated fatty acids cause apoptosis in C. albicans and C. dubliniensis biofilms. Biochim Biophys Acta 1820: 1463–1468. [DOI] [PubMed] [Google Scholar]
- Valle, J. , Toledo‐Arana, A. , Berasain, C. , Ghigo, J.M. , Amorena, B. , Penades, J.R. , and Lasa, I. (2003) SarA and not σB is essential for biofilm development by Staphylococcus aureus . Mol Microbiol 48: 1075–1087. [DOI] [PubMed] [Google Scholar]
- Vikram, A. , Jesudhasan, P.R. , Jayaprakasha, G.K. , Pillai, S.D. , and Patil, B.S. (2010) Citrus limonoids interfere with Vibrio harveyi cell‐cell signalling and biofilm formation by modulating the response regulator LuxO. Microbiology 157: 99–110. [DOI] [PubMed] [Google Scholar]
- Weinstein, M.J. , Luedemann, G.M. , Oden, E.M. , Wagman, G.H. , Rosselet, J.P. , Marquez, J.A. , et al. (1963) Gentamicin, a new antibiotic complex from micromonospora. J Med Chem 6: 463–464. [DOI] [PubMed] [Google Scholar]
- Wright, G.D. (2015) Solving the antibiotic crisis. ACS Infect Dis 1: 80–84. [DOI] [PubMed] [Google Scholar]
- Wright, G.D. (2016) Antibiotic adjuvants: rescuing antibiotics from resistance. Trends Microbiol 24: 862–871. [DOI] [PubMed] [Google Scholar]
- Yoon, B.K. , Jackman, J.A. , Valle‐Gonzalez, E.R. , and Cho, N.J. (2018) Antibacterial free fatty acids and monoglycerides: Biological activities, experimental testing, and therapeutic applications. Int J Mol Sci 19: 1114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshizawa, S. , Fourmy, D. , and Puglisi, J.D. (1998) Structural origins of gentamicin antibiotic action. EMBO J 17: 6437–6448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou, L. , Zhang, L.H. , Camara, M. , and He, Y.W. (2017) The DSF family of quorum sensing signals: diversity, biosynthesis, and turnover. Trends Microbiol 25: 293–303. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. Planktonic cell growth in the presence of fatty acids.
Fig. S2. Dispersal of established biofilms by fatty acids.
Table S1. Strains and culture media used in this study and MICs of saw palmetto oil, lauric acid, and myristic acid.
Table S2. Primer sequences used for qRT‐PCR.
Table S3. GC‐MS analysis of saw palmetto oil.