

Abstract
Fifteen newborn chickens were isolated in separate cages after 1 month of living together, divided into three groups, and challenged for 5 weeks with seed food which either was supplemented with avoparcin (10 mg/kg of animal food) or tylosin (40 mg/kg) or was nonsupplemented. At 9 weeks of age and after the 5-week challenge, all chickens received nonsupplemented feed for 4 additional weeks. At 4, 9, and 13 weeks of life, feces were collected and inoculated on M-Enterococcus agar plates with and without vancomycin (4 μg/ml). vanA-containing Enterococcus hirae was isolated from 11 of 15 chickens before antibiotic challenge, without detection of vancomycin-resistant Enterococcus faecium. At 9 weeks of age and after the 5-week avoparcin challenge, vanA E. hirae strains were no longer detected, but five of five chickens now had vanA E. faecium. At a lower frequency, vanA E. faecium had also displaced vanA E. hirae in both the tylosin group (one of four chickens) and the control group (two of five chickens). One month after avoparcin discontinuation, the number of chickens colonized with vanA E. faecium decreased from five to one. All vanA-containing E. hirae strains detected in the first month of life and most of the vanA-containing E. faecium strains detected in the second month of life showed identical ApaI and SmaI restriction patterns, respectively, when analyzed by pulsed-field gel electrophoresis. All vanA E. hirae isolates transferred glycopeptide and macrolide resistance to Enterococcus faecalis JH2-2 in vitro; the level of glycopeptide resistance was higher in the transconjugants than in the donor E. hirae strains. These data suggest that E. hirae may be a significant source of vanA determinants with the potential of transfer to other enterococcal species from humans or animals.
Modern antimicrobial therapy has classified enterococci among the more important nosocomial pathogens. Due to the intrinsic resistance of this genus to several antibiotics, and the increasing prevalence of strains with high-level penicillin (mainly Enterococcus faecium) and aminoglycoside resistance, glycopeptides could be considered as drugs of choice in some severe enterococcal infections. Unfortunately, the emergence of vancomycin resistance (Vanr) over the past few years has become an increasing problem in medical centers throughout the United States and Europe (35). In the United States, the nosocomial prevalence of vancomycin-resistant enterococci (VRE) increased from 0.3 in 1989 to 11 to 13% in non-intensive care unit patients by 1996 (6, 18, 42), and clonal spread of resistant strains has been found (7, 37). The origin of such strains remains controversial. In one study in the United States, VRE (vanA or vanB) were found in stools from 16% of high-risk hospitalized patients, but not from community-based volunteers without hospital exposure, nor from the environment or animals (10). In Europe, VRE are infrequently found among clinical isolates (5, 46, 52), and there is often great heterogeneity among typed isolates (4, 23). Recent studies have consistently found VRE in the environment, including sewage (24, 44), animal samples, and food of animal origin (1, 3, 13, 25, 26, 50). The reported rates of VRE fecal carriage in nonhospitalized Europeans range from 2 to 28% (23, 26, 48, 49). Information about the degree of VRE colonization in carriers and whether this colonization is transient or persistent remains scarce (10, 49). In any case, colonization precedes most infections (15).
The glycopeptide avoparcin has been used as a growth promoter in animal husbandry in Europe (especially in poultry and pigs) since the middle 1970s. An association between the rates of vancomycin resistance in humans and avoparcin usage in animals has been suggested (1, 26, 34, 48). Therefore, the European Union has proposed banning the use of avoparcin as a growth promoter in animals since April 1997. Tylosin is a 16C-macrolide that has also been used for growth promotion. vanA containing Enterococcus strains are frequently resistant to the 14C-macrolide erythromycin. The present study was designed to evaluate the influence of avoparcin and tylosin antibiotics on the selection and evolution of intestinal vanA-containing enterococci in chicken. We also examined the pulsed-field gel electrophoresis (PFGE) patterns of enterococcal isolates collected during the study in order to determine the relatedness between the isolates.
(This study was presented in part at the 37th Interscience Conference on Antimicrobial Agents and Chemotherapy, Toronto, Canada, 28 September to 1 October 1997. [36a].)
MATERIALS AND METHODS
Samples and strain identification.
Fifteen newborn chickens were analyzed. One-day-old animals were obtained from a commercial broiler company in Northern Spain and were maintained during the period of the study in a private family-run farm in the same region, which had not used antibiotics as feed supplements. The chicks were maintained in the same cage for 1 month (required for survival), and received non-antibiotic-supplemented seed food. After this month, chickens were separated into newly decontaminated individual cages and individual fecal samples were collected (sample I). Measures were taken to decrease the risk of cross-contamination among cages during the period of the experiment (such as having the cages separated to avoid contact among chickens). Three groups of animals (five animals each) were formed. Group 1 (cages 1C to 5C) animals were maintained with nonsupplemented food, group 2 (cages 1A to 5A) animals were fed with avoparcin-supplemented food (10 mg/kg of animal feed), and group 3 (cages 1T to 5T) animals were fed with tylosin-supplemented food (40 mg/kg). After a time of exposure to the antibiotic-supplemented food of 5 weeks, individual fecal samples were obtained from each chicken (sample II). Antibiotics were then discontinued from the food of all groups. 1 month later, another fecal sample from each chicken was analyzed (sample III). Sample processing was performed as follows. Approximately 1 g of fecal sample was suspended in 3 ml of sterile saline solution and serially 10-fold diluted. A 50-μl aliquot and a 10-μl aliquot of the dilutions were used to inoculate M-Enterococcus agar medium (bioMérieux, Marcy-l’Etoile, France), with and without vancomycin (4 μg/ml), respectively. Plates were incubated at 37°C and examined at 24 and 48 h. Bacterial counts were performed from both the vancomycin-supplemented and nonsupplemented agar plates, and all colonies with the appearance of enterococci that grew on the vancomycin-containing plates were identified by the API 20 Strep system (bioMèrieux), supplemented with biochemical tests as previously recommended (17). To corroborate the identification to the species level, the isolates were tested for the presence of genes coding for E. faecalis antigen A (EfaA) (41), chromosomal E. faecium aminoglycoside acetyltransferase-6′ [AAC(6′)-Ii] (11), and E. hirae muramidase-2 (8), by colony lysis hybridization. Intragenic probes for efaA (730 bp) and aac(6)′-li (323 bp) were generated by PCR from E. faecalis TX4002 and E. faecium TX0016, respectively. The muramidase gene of E. hirae cloned into Escherichia coli pUC19 (9) (kindly provided by Lolita Daneo-Moore) was used as a gene probe for E. hirae. Plasmid DNA was prepared with the Wizard Plus Minipreps kit (Promega, Madison, Wis.) and was digested with EcoRI and EcoRV. These probes were cleaned and labelled with 32P for hybridization (41). Preparation of colony lysates containing denatured enterococcal genomic DNA and hybridizations under high-stringency conditions were carried out by using modified standard protocols (41).
Susceptibility testing.
Susceptibility testing was performed with all of the colonies obtained from each vancomycin-containing plate. Only one isolate of a given species per specimen was selected after susceptibility testing, unless a different antibiotic resistance phenotype was observed. MICs of vancomycin (Eli Lilly & Co., Indianapolis, Ind.); avoparcin (Roche, Basel, Switzerland), teicoplanin, and erythromycin (Roussel Uclaf, Paris, France); tylosin, streptomycin, kanamycin, gentamicin, ampicillin, rifampin, and fusidic acid (Sigma Chemical Co., St. Louis, Mo.); and quinupristin-dalfopristin (Rhone-Poulene Rorer, Vitry sur Seine, France) were determined by the agar dilution method according to the National Committee for Clinical Laboratory Standards (36). An Enterococcus isolate for which the MIC of streptomycin or kanamycin was ≥2,000 μg/ml and that of gentamicin was >500 μg/ml was considered as having high-level resistance to these aminoglycosides; borderline results for streptomycin (MIC, 1,000 μg/ml) were tested by using highly charged (300-μg) aminoglycoside disks (38). β-Lactamase was determined by using nitrocefin disks (Becton Dickinson Microbiology Systems, Cockeysville, Md.).
Characterization of resistance genes.
PCR was performed, as previously described, to amplify the vanA (53), vanB (9), vanC1 (32), and vanC2 genes (14). As positive controls for vanA, vanB, vanC1, and vanC2 reactions, the strains E. faecium AR1 (44), E. faecium SF299 (19), Enterococcus gallinarum 970, and E. casseliflavus 969 (Spanish Culture Type Collection), respectively, were used. Genomic DNA of vancomycin-resistant strains was used for dot-blot hybridization with a vanA probe obtained by PCR (from E. faecium AR1) and labeled with digoxigenin (Boehringer Mannheim DNA labeling and detection kit, Mannheim, Germany). PCR was also performed with high-level gentamicin- and kanamycin-resistant Enterococcus strains to amplify genes coding for the bifunctional AAC(6′)-APH(2") enzyme and the phosphotransferase APH(3′) enzyme, using primers and conditions previously described (47).
Conjugation experiments and plasmid extraction.
Transfer of aminoglycosides and vancomycin resistance to the recipient strains E. faecalis JH2-2 (plasmid free, susceptible to vancomycin and erythromycin, and without high-level resistance to streptomycin and kanamycin, but resistant to rifampin and fusidic acid) (22) and E. faecium GE-1 (susceptible to vancomycin, tylosin, with intrinsic high-level resistance to kanamycin, but not to streptomycin or gentamicin, and resistant to rifampin and fusidic acid) (16) was performed by the filter-mating method. Donor and recipient strains were mixed in a 1:10 ratio. The selective agar plates for transconjugant cells contained vancomycin (4 μg/ml), rifampin (100 μg/ml), and fusidic acid (25 μg/ml). Plasmid DNAs of vanA-containing Enterococcus isolates and their transconjugants were obtained as previously described, including lysozyme treatment (39).
Bacteriocin activity.
Screening for bacteriocin activity was performed by the agar spot test method (45).
PFGE of genomic DNA.
All vanA-containing E. hirae and E. faecium strains were analyzed by PFGE. Genomic DNA was prepared in Incert agarose plugs as previously described (43), except for the lysis step, where bacteria in plugs were lysed in EC lysis solution (6 mM Tris, 1 M NaCl, 100 mM EDTA [pH 7.6], 0.5% Brij 58, 0.5% sarcosyl, 0.2% deoxycolic acid, 20 μg of RNase/ml, and 1 mg of lysozyme/ml) for 4 to 6 h only before being treated overnight in ESP solution (0.5 M EDTA [pH 9], 50 μg of proteinase K/ml, and 1% sarcosyl) at 50°C in a shaking water bath. Restriction enzyme ApaI (Gibco BRL, Life Technologies, Gaithersburg, Md.) was used to digest E. hirae DNA in plugs, while SmaI (Gibco BRL) was used to digest E. faecium genomic DNA. One percent I.D.NA agarose (FMC BioProducts Rockland, Maine), prepared in 0.25× TBE (1× TBE is 0.089 M Tris, 0.002 M EDTA, and 0.089 M boric acid) buffer and DNA samples were electrophoresed by using a clamped homogeneous electric field with a CHEF-DR II (Bio-Rad Laboratories, Richmond, Calif.) system with a pulse ramping time from 2 s to 22 s for 16 h. Gels were stained in ethidium bromide and photographed against UV light. Once the isolates having identical patterns were analyzed, a representative isolate of the group was used to compare its restriction pattern with those of the other isolates. Isolates were classified as indistinguishable, closely related, possibly related, or different according to previously published criteria for bacterial strain typing (33).
Statistical analysis.
Statistical analyses were performed by geometric means and t test (paired) for comparison of the avoparcin group with the control and tylosin groups by using the statistical program StatView (Abacus Concepts, Berkeley, Calif.).
RESULTS
Identification and PCR results.
VRE from chicken fecal samples were identified as E. hirae and E. faecium by the API 20 system supplemented with biochemical tests (17). The DNA probe for aac(6′)-li hybridized under high-stringency conditions only to E. faecium, and the DNA probe for muramidase-2 hybridized only to E. hirae. Negative results were obtained with the E. faecalis probe, efaA. Positive PCR amplification of vanA and negative results for vanB, vanC1, and vanC2 were found in all vancomycin-resistant Enterococcus strains isolated from chicken fecal samples. The DNA probe for vanA hybridized to all VRE isolated from fecal samples.
vanA-containing enterococci in chickens receiving avoparcin-supplemented feed.
At the end of the first month of life and immediately before antibiotic challenge, most chickens (11 of 15) harbored vanA-positive E. hirae strains, at concentration of 3.8 × 103 CFU/g of feces (Table 1). During the period of intervention, the proportion of chickens in the control group with vanA E. hirae decreased (from three to one), but vanA E. faecium emerged in two chickens, one of which had vanA E. hirae simultaneously with a vanA E. faecium strain. One month after the period of intervention, two of five chickens from the control group harbored vanA E. faecium strains.
TABLE 1.
vanA-containing Enterococcus strains isolated from 15 chickens during three observation periods
| Chicken | Results for observation perioda:
|
|||||
|---|---|---|---|---|---|---|
| I
|
II
|
III
|
||||
| Species | CFU/g of feces | Species | CFU/g of feces | Species | CFU/g of feces | |
| Control | ||||||
| 1C | E. hirae | 6.0 × 102 | E. faecium | 4.5 × 103 | NR | <6.0 × 102c |
| 2C | E. hirae | 3.0 × 104 | E. faecium/E. hirae | 1.5 × 103 | E. faecium | 6.0 × 102 |
| 3C | NRb | <6.0 × 102c | NR | <6.0 × 102c | NR | <6.0 × 102c |
| 4C | NR | <6.0 × 102c | NR | <6.0 × 102c | E. faecium | 6.0 × 102 |
| 5C | E. hirae | 6.0 × 103 | NR | <6.0 × 102c | NR | <6.0 × 102c |
| Geometric mean | 1.9 × 103 | 9.7 × 102d | 5.4 × 103 | |||
| Avoparcin | ||||||
| 1A | NR | <6.0 × 102c | E. faecium | 1.8 × 104 | NR | <6.0 × 102c |
| 2A | E. hirae | 1.2 × 103 | E. faecium | 1.2 × 105 | NR | <6.0 × 102c |
| 3A | E. hirae | 7.2 × 104 | E. faecium | 3.0 × 104 | NR | <6.0 × 102c |
| 4A | NR | <6.0 × 102c | E. faecium | 6.0 × 102 | E. faecium | 1.8 × 103 |
| 5A | E. hirae | 2.7 × 104 | E. faecium | 3.5 × 103 | NR | <6.0 × 102c |
| Geometric mean | 3.6 × 103 | 1.1 × 104d | 6.5 × 102 | |||
| Tylosin | ||||||
| 1T | E. hirae | 7.3 × 103 | Chicken died | Chicken died | ||
| 2T | E. hirae | 1.8 × 104 | NR | <6.0 × 102c | NR | <6.0 × 102c |
| 3T | E. hirae | 4.2 × 103 | E. faecium | 1.0 × 104 | NR | <6.0 × 102c |
| 4T | E. hirae | 3.0 × 103 | NR | <6.0 × 102c | E. faecium | 6.0 × 102 |
| 5T | E. hirae | 1.8 × 104 | E. hirae | 3.0 × 103 | E. faecium | 1.2 × 104 |
| Geometric mean | 7.8 × 103 | 1.6 × 103d | 1.2 × 103 | |||
I, baseline (before antibiotic challenge); II, 4 weeks after antibiotic challenge; III, after 5 weeks of antibiotic discontinuation.
NR, no vanA Enterococcus strains detected.
The value 5 × 102 was used instead of <6.0 × 102 to calculate geometric means.
Difference not statistically significant.
In the avoparcin-challenged group, three of five chickens originally harbored vanA-containing E. hirae; these were replaced after challenge by vanA E. faecium. In the other two chickens, vanA E. faecium was also found after challenge, but was the first vanA isolate. VanA-type E. faecium strains were found at a mean concentration of 1.1 × 104 CFU/g of feces. Interestingly, vanA E. faecium isolates were not recovered from three of the five animals harboring this resistant organism in the avoparcin group after antibiotic discontinuation. The proportion of vanA-containing enterococci among the total enterococcal strains was quite variable among animals, but this proportion in the avoparcin-treated group showed a broader range in different animals (range, 0.8 to 45%; mean, 17.9%) than in the control group (range, 0 to 5.5%; mean, 1.6%).
vanA-containing enterococci in chickens receiving tylosin-supplemented feed.
All chickens included in the group receiving tylosin-supplemented feed originally harbored vanA E. hirae (five of five). After tylosin challenge, vanA E. hirae disappeared in three chickens, but a vanA E. faecium strain emerged in a single chicken (one of four). One chicken died during this period. Four weeks after tylosin discontinuation, vanA E. faecium was detected in two of four animals (Table 1). The proportion of vanA-containing enterococci among the total enterococcal strains in the tylosin group had a range of 0 to 5% between different animals (mean, 1.2%), similar to that of the control group (mean, 1.6%).
Characteristics of vanA-containing enterococci.
In all cases, vanA-containing E. faecium strains showed the following antibiotic susceptibility profile: resistance to vancomycin (MIC, 512 to 1,024 μg/ml), avoparcin (MIC, 64 to 512 μg/ml), teicoplanin (MIC, 128 to 256 μg/ml), erythromycin (MIC, >512 μg/ml), and tylosin (MIC, >256 μg/ml) and absence of high-level resistance to aminoglycosides (streptomycin, gentamicin, and kanamycin) and quinupristin-dalfopristin (Table 2). All E. faecium strains were susceptible to ampicillin (MIC, ≤0.5 to 8 μg/ml). Glycopeptide resistance was always transferred by conjugation, and resistance to macrolides was acquired by the recipient strain in 9 of 11 isolates (Table 2).
TABLE 2.
PFGE patterns and MICs of different antibiotics for VanA Enterococcus strains isolated from chicken samples and transconjugants with E. faecalis JH2-2 as the recipient strain
| Donor (no. of isolates) | PFGE pattern(s) | MIC (μg/ml) for donora
|
Phenotype of Vanr transconjugants | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Avo | Van | Tei | Ery | Tyl | Str | Gen | Kan | Amp | Q/D | |||
| E. hirae | ||||||||||||
| 6 | Ehi-A | 128 | 64–128 | 16–32 | >512 | >256 | 1,000–4,000 | ≤16 | >4,000 | ≤0.5–4 | 1 | Ery Tyl Kanr |
| 4 | Ehi-A and A1 | 64–128 | 64–128 | 16–32 | >512 | >256 | 1,000 | ≤16 | >4,000 | 2–16 | 1 | Ery Tyl Kans |
| 3 | Ehi-A | 128 | 64–128 | 8–16 | >512 | >256 | 1,000 | ≤16 | >4,000 | 1–2 | 1 | Ery Tyl Kans and Ery Tyl Kanr |
| 1 | Ehi-A | 64 | 64 | 16 | >512 | >256 | 1,000 | 2,000 | >4,000 | 2 | 1 | Ery Tyl Kans and Ery Tyl Kanr |
| E. faecium | ||||||||||||
| 4 | Efm-A | 256–512 | 512–1,024 | 128 | >512 | >256 | 32 | ≤16 | ≤128–256 | ≤0.5–2 | 1 | Ery Tyl Kanr |
| 1 | Efm-A | 256 | 1,024 | 128 | >512 | >256 | 32 | ≤16 | 256 | 1 | 1 | Ery Tyl Kans |
| 2 | Efm-A | 256 | 1,024 | 256 | >512 | >256 | 128 | ≤16 | ≤128 | 8 | 1 | None obtained |
| 1 | Efm-B | 64 | 1,024 | 128 | >512 | >256 | ≤16 | ≤16 | ≤128 | 8 | 1 | Ery Tyl Kanrb |
| 2 | Efm-C | 64–128 | 512 | 128 | >512 | >256 | ≤16–32 | ≤16 | ≤128–256 | ≤0.5–4 | 1 | Ery Tyl Kans |
| 1 | Efm-C | 64 | 256 | 128 | >512 | >256 | 32 | ≤16 | 256 | 1 | 1 | Ery Tyl Kanr |
| 1 | Efm-C1 | 256 | 512 | 128 | >512 | >256 | 128 | ≤16 | ≤128 | 8 | 0.5 | Ery Tyl Kans and Ery Tyl Kanr |
| 1 | Efm-D | 64 | 1,024 | 128 | >512 | >256 | ≤16 | ≤16 | ≤128 | 4 | 0.5 | Ery Tyl Kanrb |
Avo, avoparcin; Van, vancomycin; Tei, teicoplanin; Ery, erythromycin; Tyl, tylosin; Str, streptomycin; Gen, gentamicin; Kan, kanamycin; Amp, ampicillin; Q/D, quinupristin-dalfopristin.
Conjugation with E. faecium GE-1 as recipient.
All vanA-containing E. hirae isolates were resistant to vancomycin (MIC, 64 to 128 μg/ml), avoparcin (MIC, 64 to 128 μg/ml), erythromycin (MIC, >512 μg/ml), and tylosin (MIC, >256 μg/ml), but showed lower-level resistance to teicoplanin (MIC, 8 to 32 μg/ml) (Table 2). All of these isolates were highly resistant to streptomycin and kanamycin, and one isolate was also highly resistant to gentamicin (E. hirae B-71). The gene coding for APH(3′) was detected by PCR in these E. hirae isolates, and a positive amplification product (220 bp) was obtained from E. hirae B-71 by PCR with the primers for the aph2"-aac6′ gene. All of these isolates were susceptible to quinupristin-dalfopristin. One of 14 E. hirae isolates showed low-level ampicillin resistance (MIC, 16 μg/ml), and no β-lactamase production was detected. Glycopeptide resistance was transferred by conjugation to E. faecalis JH2-2 from all isolates, and transconjugants expressed high-level resistance to both vancomycin (MIC, >256) and teicoplanin (MIC, ≥256), even higher than that in the donor strains. Transfer of vancomycin resistance was always associated with transfer of streptomycin resistance. Resistance to erythromycin and tylosin was also cotransferred with resistance to glycopeptides from 10 of 14 E. hirae isolates; transfer of macrolide resistance was always associated with transfer of kanamycin resistance.
In 11 vanA-containing Enterococcus strains, the plasmid-content was investigated. A high-molecular-weight plasmid was detected in five of six E. hirae isolates and in three of five E. faecium isolates. Transfer of vancomycin resistance was associated with the acquisition of this high-molecular-weight plasmid in three of the E. faecium isolates; in only one case was erythromycin resistance cotransferred. Plasmids were not detectable in vancomycin-resistant transconjugants from E. hirae isolates.
No bacteriocin production by the vanA E. faecium strains could be demonstrated when assayed against the vanA E. hirae strains of this study.
PFGE.
Thirteen E. hirae isolates collected from chicken fecal samples demonstrated identical ApaI restriction patterns when analyzed by PFGE and were assigned the pattern designation Ehi-A (Fig. 1). One isolate differed by three bands from the other 13 isolates and was characterized as a variant of the Ehi-A pattern (Fig. 1). Analysis of 13 E. faecium isolates by SmaI digestion revealed four different PFGE patterns (Fig. 2). Eight isolates of E. faecium showed identical SmaI restriction patterns and were designated as the Efm-A pattern. The pattern designation Efm-C was assigned to two E. faecium strains with identical PFGE patterns. A third E. faecium strain showed a closely related (single-band difference) PFGE pattern, Efm-C1. Two other E. faecium strains showed PFGE patterns completely different from each other and from the other isolates and were designated as Efm-D and Efm-B, respectively (Fig. 2).
FIG. 1.
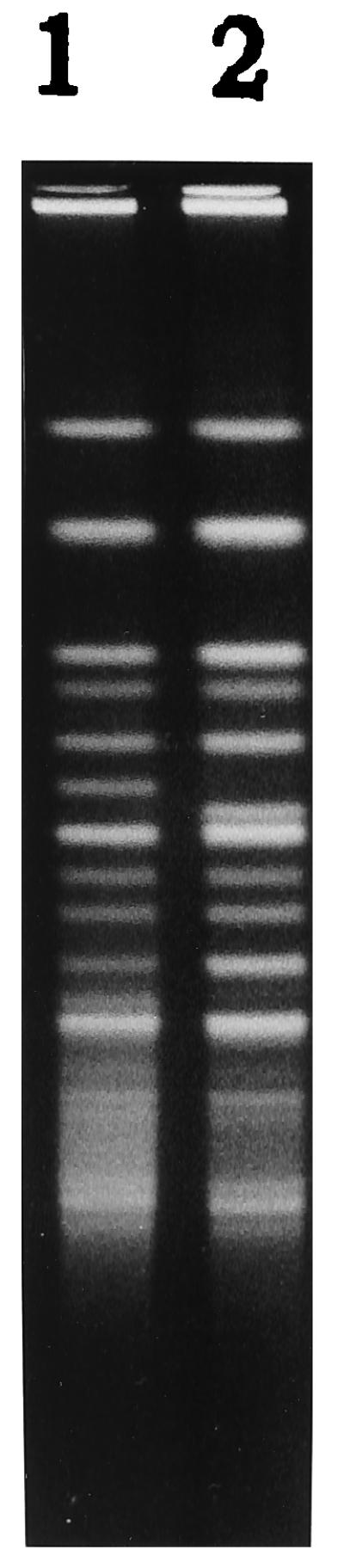
PFGE of ApaI-digested genomic DNA from vanA-containing E. hirae strains. Lanes: 1, Efm-A1 pattern; 2, Efm-A pattern.
FIG. 2.

PFGE of SmaI-digested genomic DNA from vanA-containing E. faecium strains. Lanes: 1, Efm-A pattern; 2, Efm-C pattern; 3, Efm-C1 pattern; 4, Efm-D pattern; 5, Efm-B pattern.
DISCUSSION
E. hirae isolates containing vanA were consistently detected in the feces of newborn chickens fed with nonsupplemented seeds. The origin of the resistant strains may be the broiler company. The seed may be contaminated with Enterococcus (20), but we were unable to detect vanA-containing enterococci when seed preparations were inoculated on selective culture media containing vancomycin (data not shown). The presence of a common strain of vanA-containing E. hirae in most animals under observation likely relates to the common occupancy of a single cage by all chickens during the initial period of the experiment. Isolation is impossible at very early stages of chicken growth, because it is followed by high spontaneous mortality. The proportion of vanA-containing E. hirae strains in the total number of Enterococcus strains was variable among animals, suggesting that some were more likely to serve as a source for cross-contamination. Similar spread would be expected to occur under normal circumstances in chicken farms. In both antibiotic-supplemented and nonsupplemented animals, vanA E. hirae tended to disappear during the second month of life, confirming its role as a member of the early bacterial community in the chicken gut, as has been previously shown with vancomycin-susceptible E. hirae strains (12). Nevertheless, at this stage, the vanA determinant was now detectable in E. faecium. It is possible that the original population of E. hirae strains may have been naturally replaced by a formerly minority vanA E. faecium subpopulation undetected in previous samples. On the other hand, the possibility of in vivo transfer of vanA from E. hirae to E. faecium can also be considered. Such transfer occurs under in vitro conditions; moreover, in at least one animal, both vanA E. hirae and vanA E. faecium strains were simultaneously present in the intestine. Specific replacement due to bacteriocin production was not suggested by the results of in vitro competition assays.
The addition of avoparcin to the food was associated with the appearance of vanA-containing E. faecium strains in all challenged animals; discontinuation of the antibiotic for 1 month was followed by the failure to detect these isolates in four of the five chickens. The differences in numbers of chickens colonized and the number of VRE were not statistically significantly different, perhaps because of the small number of chickens studied and the low concentration of avoparcin feed. The average cell density of VRE in feces from avoparcin-treated chickens was about 11 times higher than that in nontreated animals. These data suggest that avoparcin had some selective effect and that, in the absence of selection, vancomycin-susceptible enterococci present in the intestinal content may overgrow the resistant organisms. In some animals, colonization by resistant enterococci may last much longer; Bager et al. (2) detected vancomycin (vanA)-resistant strains 6 months after discontinuation of avoparcin feeding in pigs and poultry.
Tylosin supplementation did not appear to select for vanA-containing strains, despite the fact that all vancomycin-resistant strains were tylosin resistant. The use of other growth promoters in animals, tylosin in this case and virginiamycin in the report of Welton et al. (51), did not appear to select for vanA-containing enterococci.
All vanA E. hirae strains detected in the first month of the chickens’ lives in the control, avoparcin, and tylosin groups showed identical ApaI restriction patterns when analyzed by PFGE (pattern Ehi-A). In the second month, when addition of avoparcin and tylosin started, vanA E. hirae strains tended to disappear and to be replaced by vanA E. faecium. All vanA E. faecium isolates recovered in the second month showed identical SmaI patterns (pattern Efm-A), except for two strains detected in the avoparcin group that showed a completely different pattern (Efm-C and its closely related Efm-C1 pattern). However, after discontinuation of the antibiotic for 1 month, three different vanA E. faecium patterns were found in the avoparcin and tylosin groups, while in the control group, the vanA E. faecium population, pattern Efm-A was the only one detected. The variability of the patterns detected in the avoparcin and tylosin groups and the continuity in control group suggest that avoparcin and tylosin may have contributed to expansion of the vanA E. faecium population.
The vanA-containing E. hirae strains expressed a lower level of glycopeptide resistance than E. faecium. This phenomenon occurred both with vancomycin (mode MIC, 64 to 128 versus 512 to 1,024 μg/ml, respectively) and teicoplanin (mode MIC, 8 to 32 versus 128 to 256 μg/ml). The glycopeptide MICs of the recipient E. faecalis strains after conjugation with E. hirae were in the E. faecium range, suggesting that the same vanA gene cluster may have a lower level of expression in E. hirae than in E. faecalis or E. faecium. The vancomycin resistance gene cluster may have different levels of expression in different species (21, 29).
Different patterns of cotransfer of macrolide resistance and glycopeptide resistance were found by using E. hirae strains as donors, despite the fact that all of the E. hirae isolates showed a common PFGE pattern. In 10 of 14 instances, E. faecalis transconjugants with both macrolide and glycopeptide resistance were recovered, but in 4 of these 10, there were also transconjugants in which only glycopeptide resistance was transferred. E. faecalis transconjugants were obtained with both macrolide and glycopeptide resistance from 9 of 11 E. faecium donors, but in 1 of 9 of these strains, transconjugants also were derived that showed only vancomycin resistance; 2 of 11 had only macrolide resistance. These results suggest that both resistances are in separate replicons that are frequently cotransferred.
One of the vanA E. hirae strains showed high-level gentamicin resistance, and the gene coding for the bifunctional AAC(6′)-APH(2") enzyme was detected by PCR. This strain showed the same ApaI restriction pattern as the others that were gentamicin susceptible, which suggests that the gene encoding the bifunctional AAC(6′)-APH(2") may have been located in a plasmid and transferred in vivo. McNamara et al. (31) found high-level gentamicin resistance in 34% of E. hirae clinical isolates, and the genetic determinant for the bifunctional enzyme in this species was shown to be homologous to that characterized in other species, such as E. faecalis and E. faecium (29).
vanA E. hirae strains have been previously isolated from water (27) and humans (40). The incidence of this species in the human flora has probably been underestimated, but may account for 3% of enterococcal clinical isolates (31). The rate of acquisition of vancomycin-resistant E. hirae strains of chicken origin by human populations and its role as a source of vancomycin and macrolide resistance determinants remain to be evaluated.
These results suggest that vancomycin resistance may be common in newborn chickens in Northern Spain, and the epidemiology of vanA E. hirae strains may be of interest, because this species may well be a reservoir for glycopeptide resistance. Despite the low number of chickens studied, our data indicate that avoparcin supplementation may increase the selection of glycopeptide-resistant enterococci, but this effect may be transient. Tylosin was not associated with glycopeptide selection, suggesting that the selective measure was mainly exerted on the non-vanA-containing Enterococcus populations. Because the acquisition of tylosin resistance implies cross-resistance with all macrolide antibiotics, and avoparcin resistance correlates with vancomycin and teicoplanin resistance, a closer surveillance of the chicken reservoir of antibiotic-resistant Enterococcus should be urgently considered.
ACKNOWLEDGMENTS
This work was supported by a grant from the Ministerio de Salud y Consumo of Spain (FIS 98/0282).
We thank Carmen Robledo for helping us care for the chickens, Emilia Cercenado for providing E. faecium SF299, and Teresa Coque for critical review of the manuscript.
REFERENCES
- 1.Aarestrup F M. Occurrence of glycopeptide resistance among Enterococcus faecium isolates from conventional and ecological poultry farms. Microb Drug Res. 1995;1:255–257. doi: 10.1089/mdr.1995.1.255. [DOI] [PubMed] [Google Scholar]
- 2.Bager F, Madsen M, Christensen J, Aarestrup F M. Avoparcin used as a growth promoter is associated with the occurrence of vancomycin-resistant Enterococcus faecium on Danish poultry and pig farms. Prev Vet Med. 1997;31:95–112. doi: 10.1016/s0167-5877(96)01119-1. [DOI] [PubMed] [Google Scholar]
- 3.Bates E M, Jordens J Z, Griffiths D T. Farm animals as a putative reservoir for vancomycin resistant enterococcal infections in man. J Antimicrob Chemother. 1994;34:507–516. doi: 10.1093/jac/34.4.507. [DOI] [PubMed] [Google Scholar]
- 4.Bingen E H, Denamur E, Lambert-Zechovsky N Y, Elion J. Evidence for the genetic unrelatedness of nosocomial vancomycin-resistant Enterococcus faecium strains in a pediatric hospital. J Clin Microbiol. 1991;29:1888–1892. doi: 10.1128/jcm.29.9.1888-1892.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brown D F J, Courvalin P the European Glycopeptide Resistance Group. Program and abstracts of the 36th Interscience Conference on Antimicrobial Agents and Chemotherapy. Washington, D.C: American Society for Microbiology; 1996. European glycopeptide susceptibility survey: susceptibility of Enterococcus spp to teicoplanin and vancomycin, abstr. E26; p. 85. [Google Scholar]
- 6.Centers for Disease Control and Prevention. Nosocomial enterococci resistant to vancomycin—United States, 1989–1993. Morbid Mortal Weekly Rep. 1993;42:597–599. [PubMed] [Google Scholar]
- 7.Chow J W, Kuritza A, Shlaes D M, Green M, Sahm D F, Zervos M J. Clonal spread of vancomycin-resistant Enterococcus faecium between patients in three hospitals in two states. J Clin Microbiol. 1993;31:1609–1611. doi: 10.1128/jcm.31.6.1609-1611.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chu C-P, Kariyama R, Daneo-Moore L, Shockman G D. Cloning and sequence analysis of the muramidase-2 gene from Enterococcus hirae. J Bacteriol. 1992;174:1619–1625. doi: 10.1128/jb.174.5.1619-1625.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Clark N C, Cooksey R C, Hill B C, Swenson J M S, Tenover F C. Characterization of glycopeptide-resistant enterococci from U.S. hospitals. Antimicrob Agents Chemother. 1993;37:2311–2317. doi: 10.1128/aac.37.11.2311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Coque T M, Tomayko J F, Ricke S C, Okhyusen P C, Murray B E. Vancomycin-resistant enterococci from nosocomial, community, and animal sources in the United States. Antimicrob Agents Chemother. 1996;40:2605–2609. doi: 10.1128/aac.40.11.2605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Costa Y, Galimand M, Leclercq R, Duval J, Courvalin P. Characterization of the chromosomal aac(6′)-Ii gene specific for Enterococcus faecium. Antimicrob Agents Chemother. 1993;37:1896–1903. doi: 10.1128/aac.37.9.1896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Devriese L A, Hommez J, Wijfels R, Haesebrouck F. Composition of the enterococcal and streptococcal intestinal flora of poultry. J Appl Bacteriol. 1991;71:46–50. [PubMed] [Google Scholar]
- 13.Devriese L A, Ieven M, Goossens H, Vandamme P, Pot B, Hommez J, Haesebrouck F. Presence of vancomycin-resistant enterococci in farm and pet animals. Antimicrob Agents Chemother. 1996;40:2285–2287. doi: 10.1128/aac.40.10.2285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dutka-Malen S, Evers S, Courvalin P. Detection of glycopeptide resistance genotypes and identification to the species level of clinically relevant enterococci by PCR. J Clin Microbiol. 1995;33:24–27. doi: 10.1128/jcm.33.1.24-27.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Edmond M B, Ober J F, Weinbaum D L, Pfaller M A, Hwang T, Sanford M D, Wenzel R Y. Vancomycin-resistant Enterococcus faecium bacteremia: risk factors for infection. Clin Infect Dis. 1995;20:1126–1133. doi: 10.1093/clinids/20.5.1126. [DOI] [PubMed] [Google Scholar]
- 16.Eliopoulos G M, Wennersten C, Zighelboim-Daum S, Goldmann D, Moellering R C., Jr High-level resistance to gentamicin in clinical isolates of Streptococcus (Enterococcus) faecium. Antimicrob Agents Chemother. 1988;32:1528–1532. doi: 10.1128/aac.32.10.1528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Facklam R R, Collins M D. Identification of Enterococcus species isolated from human infections by a conventional test scheme. J Clin Microbiol. 1989;27:731–734. doi: 10.1128/jcm.27.4.731-734.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gaynes R, Edwards J. National Nosocomial Infection Surveillance (NNIS) System. Nosocomial vancomycin resistant enterococci (VRE) in the United States, 1989–1995: the first 1000 isolates. Infect Control Hosp Epidemiol. 1996;17(Suppl.):P18. doi: 10.1086/646686. [DOI] [PubMed] [Google Scholar]
- 19.Gold H S, Cercenado E, Thauvin-Eliopoulos C, Eliopoulos G M, Wennerstern C B, Moellering R C., Jr A gene conferring resistance to vancomycin but not teicoplanin in isolates of Enterococcus faecalis and Enterococcus faecium demonstrates homology with vanB, vanA, and vanC genes of enterococci. Antimicrob Agents Chemother. 1993;37:1604–1609. doi: 10.1128/aac.37.8.1604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Guillot J F, Chaslus-Dancla E, Lafont J P. Spontaneous implantation of antibiotic-resistant Enterobacteriaceae in the digestive tract of chickens in the absence of selective pressure. Antimicrob Agents Chemother. 1977;12:697–702. doi: 10.1128/aac.12.6.697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hayden M K, Picken R N, Sahm D F. Heterogeneous expression of glycopeptide resistance in enterococci associated with transfer of vanB. Antimicrob Agents Chemother. 1997;41:872–874. doi: 10.1128/aac.41.4.872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jacob A E, Hobbs S J. Conjugal transfer of plasmid-borne multiple antibiotic resistance in Streptococcus faecalis var. zymogenes. J Bacteriol. 1974;117:360–372. doi: 10.1128/jb.117.2.360-372.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jordens J Z, Bates J, Griffiths D T. Faecal carriage and nosocomial spread of vancomycin-resistant Enterococcus faecium. J Antimicrob Chemother. 1994;34:515–528. doi: 10.1093/jac/34.4.515. [DOI] [PubMed] [Google Scholar]
- 24.Klare I, Heier H, Claus H, Witte W. Environmental strains of Enterococcus faecium with inducible high-level resistance to glycopeptides. FEMS Microbiol Lett. 1993;106:23–30. doi: 10.1111/j.1574-6968.1993.tb05930.x. [DOI] [PubMed] [Google Scholar]
- 25.Klare I, Heier H, Claus H, Reissbrodt R, Witte W. vanA-mediated high-level glycopeptide resistance in Enterococcus faecium from animal husbandry. FEMS Microbiol Lett. 1995;125:165–172. doi: 10.1111/j.1574-6968.1995.tb07353.x. [DOI] [PubMed] [Google Scholar]
- 26.Klare I, Heier H, Claus H, Reissbrodt R, Witte W. Enterococcus faecium strains with vanA-mediated high-level glycopeptide resistance isolated from animal foodstuffs and fecal samples of humans in the community. Microb Drug Res. 1995;1:265–272. doi: 10.1089/mdr.1995.1.265. [DOI] [PubMed] [Google Scholar]
- 27.Knudtson J M, Hartman P A. Antibiotic resistance among enterococcal isolates from environmental and clinical sources. J Food Prot. 1993;56:489–492. doi: 10.4315/0362-028X-56.6.489. [DOI] [PubMed] [Google Scholar]
- 28.Leclercq R, Derlot E, Duval J, Courvalin P. Plasmid-mediated resistance to vancomycin and teicoplanin in Enterococcus faecium. N Engl J Med. 1988;319:157–161. doi: 10.1056/NEJM198807213190307. [DOI] [PubMed] [Google Scholar]
- 29.Leclercq R, Derlot E, Weber M, Duval J, Courvalin P. Transferable vancomycin and teicoplanin resistance in Enterococcus faecium. Antimicrob Agents Chemother. 1989;33:10–15. doi: 10.1128/aac.33.1.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Mangan M W, McNamara E B, Smyth E G, Storrs M J. Molecular genetic analysis of high-level gentamicin resistance in Enterococcus hirae. J Antimicrob Chemother. 1997;40:377–382. doi: 10.1093/jac/40.3.377. [DOI] [PubMed] [Google Scholar]
- 31.McNamara E B, King E M, Smyth E G. A survey of antimicrobial susceptibility of clinical isolates of Enterococcus spp. from Irish hospitals. J Antimicrob Chemother. 1995;35:185–189. doi: 10.1093/jac/35.1.185. [DOI] [PubMed] [Google Scholar]
- 32.Miele A, Bandera M, Goldstein B P. Use of primers selective for vancomycin resistance genes to determine van genotype in enterococci and to study gene organization in vanA isolates. Antimicrob Agents Chemother. 1995;39:1772–1778. doi: 10.1128/aac.39.8.1772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Murray B E, Singh K V, Markowitz S M, Lopardo H A, Patterson J E, Zervos M J, Rubeglio E, Eliopoulos G M, Rice L B, Goldstain F W, Jenkins S G, Caputo G M, Nasnas R, Moore L S, Wong E S, Weinstock G. Evidence for clonal spread of a single strain of β-lactamase-producing Enterococcus (Streptococcus) faecalis to six hospitals in five states. J Infect Dis. 1991;163:780–785. doi: 10.1093/infdis/163.4.780. [DOI] [PubMed] [Google Scholar]
- 34.Murray B E. What can we do about vancomycin resistant enterococci? Clin Infect Dis. 1995;20:1134–1136. doi: 10.1093/clinids/20.5.1134. [DOI] [PubMed] [Google Scholar]
- 35.Murray B E. Vancomycin-resistant enterococci. Am J Med. 1997;102:284–293. doi: 10.1016/S0002-9343(99)80270-8. [DOI] [PubMed] [Google Scholar]
- 36.National Committee for Clinical Laboratory Standards. 4th ed. Approved standard M7-A4. Wayne, Pa: National Committee for Clinical Laboratory Standards; 1997. Methods for dilution antimicrobial susceptibility testing on bacteria that grow aerobically. [Google Scholar]
- 36a.Robredo B, Torres C, Ruiz F, Baquero F. Program and abstracts of the 33rd Interscience Conference on Antimicrobial Agents and Chemotherapy. Washington, D.C: American Society for Microbiology; 1997. Evolution of vanA Enterococcus populations in faeces of newborn chickens receiving avoparcin and tylosin as food additives, abstr. C-132; p. 69. [Google Scholar]
- 37.Sader H S, Pfaller M A, Tenover F C, Hollis R J, Jones R N. Evaluation and characterization of multiresistant Enterococcus faecium from 12 U.S. medical centers. J Clin Microbiol. 1994;32:2840–2842. doi: 10.1128/jcm.32.11.2840-2842.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sahm D F, Torres C. High-content aminoglycoside disks for determining aminoglycoside-penicillin synergy against Enterococcus faecalis. J Clin Microbiol. 1988;26:257–260. doi: 10.1128/jcm.26.2.257-260.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sambrook J, Fritsch E F, Maniatis T. Molecular cloning: a laboratory manual. 2nd ed. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1989. pp. 1.25–1.27. [Google Scholar]
- 40.Simonsen G S, Haaheim H, Kruse H, Dahl K H, Olsvik Ø, Sundsfjord A. Program and abstracts of the 37th Interscience Conference on Antimicrobial Agents and Chemotherapy. Washington, D.C: American Society for Microbiology; 1997. The possible horizontal transmission of the vanA gene cluster between chicken and human GRE-strains at avoparcin-exposed farms, abstr. C-131; p. 69. [Google Scholar]
- 41.Singh, K. V., T. M. Coque, G. M. Weinstock, and B. E. Murray. In vivo testing of an Enterococcus faecalis efaA mutant and use of efaA homologs for species identification. FEMS Immunol. Med. Microbiol. 21:323–331. [DOI] [PubMed]
- 42.Tenover F C, Gaynes R. Dissemination of vancomycin-resistant enterococci in the United States. In: Brun-Buisson C, Eliopoulos G M, Leclercq R, editors. Bacterial resistance to glycopeptides. Paris, France: Medicine Sciences Flammarion; 1998. pp. 102–110. [Google Scholar]
- 43.Tenover F C, Arbeit R D, Goering R V, Mickelsen P A, Murray B E, Persing D H, Swaminathan B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33:2233–2239. doi: 10.1128/jcm.33.9.2233-2239.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Torres C, Reguera J A, SanMartin M J, Pérez-Díaz J C, Baquero F. vanA-mediated vancomycin-resistant Enterococcus spp. isolated from sewage. J Antimicrob Chemother. 1994;33:553–561. doi: 10.1093/jac/33.3.553. [DOI] [PubMed] [Google Scholar]
- 45.Uhlman L, Schillinger U, Rupnow J R, Holzapfel W H. Identification and characterization of two bacteriocin-producing strains of Lactococcus lactis isolated from vegetables. Int J Food Microbiol. 1992;16:141–151. doi: 10.1016/0168-1605(92)90007-p. [DOI] [PubMed] [Google Scholar]
- 46.Vandamme P, Vercauteren E, Lammens C, Pensart N, Ieven M, Pot B, Leclercq R, Goossens H. Survey of enterococcal susceptibility patterns in Belgium. J Clin Microbiol. 1996;34:2572–2576. doi: 10.1128/jcm.34.10.2572-2576.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.van de Klundert J A M, Vliegenthart J S. PCR detection of genes coding for aminoglycoside-modifying enzymes. In: Persing D H, Smith T F, Tenover F C, White T J, editors. Diagnostic molecular microbiology. Principles and applications. Washington, D.C: American Society for Microbiology; 1993. pp. 547–552. [Google Scholar]
- 48.Van den Bogaard A E, Stobbergingh E E. Time to ban all antibiotics as animal growth-promoting agents? Lancet. 1996;348:619. doi: 10.1016/s0140-6736(05)64838-6. [DOI] [PubMed] [Google Scholar]
- 49.Van der Auwera P, Pensart N, Korten V, Murray B E, Leclercq R. Incidence of oral glycopeptides on the fecal flora of human volunteers: selection of highly glycopeptide resistant enterococci. J Infect Dis. 1996;173:1129–1136. doi: 10.1093/infdis/173.5.1129. [DOI] [PubMed] [Google Scholar]
- 50.Wegener H C, Madsen M, Nielsen N, Aarestrup F M. Isolation of vancomycin resistant Enterococcus faecium from food. Int J Food Microbiol. 1997;35:57–66. doi: 10.1016/s0168-1605(96)01221-4. [DOI] [PubMed] [Google Scholar]
- 51.Welton L A, Thal L A, Perry M B, Donabedian S, McMahon J, Chow J W, Zervos M J. Antimicrobial resistance in enterococci isolated from turkey flocks fed virginiamycin. Antimicrob Agents Chemother. 1998;42:705–708. doi: 10.1128/aac.42.3.705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Witte W, Klare I. Glycopeptide resistant Enterococcus faecium outside hospitals: a commentary. Microb Drugs Res. 1995;3:259–263. doi: 10.1089/mdr.1995.1.259. [DOI] [PubMed] [Google Scholar]
- 53.Woodford N, Morrison D, Johnson A P, Briant V, George R C, Cookson B. Application of DNA probes for rRNA and vanA genes to investigation of a nosocomial cluster of vancomycin-resistant enterococci. J Clin Microbiol. 1993;31:653–658. doi: 10.1128/jcm.31.3.653-658.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]