

Abstract
Background:
Recent evidence suggesting that polysubstance use is the norm rather than the exception highlights the need for a better understanding of interactions amongst the abuse-related effects of commonly co-abused drugs. Synthetic cathinones remain one of the most popular families of novel psychoactive substances and are typically used in preparations containing multiple stimulants. Evaluating the reinforcing effects of drugs under both single-operant procedures and procedures in which alternatives are available can provide a more complete characterization of their reinforcing effects and economic interactions.
Methods:
These studies utilized a drug-versus-drug choice procedure in 18 male Sprague-Dawley rats to evaluate economic interactions between the synthetic cathinone, MDPV, and cocaine in addition to how a history of concurrent access impacts reinstatement behavior.
Results:
When equi-effective doses of MDPV and cocaine were made concurrently available, approximately half of the subjects responded exclusively on the MDPV-reinforced lever whereas the other half responded exclusively on the cocaine-reinforced lever. Allocation of responding was reversed when the cost of the preferred drug increased, or the cost of the non-preferred drug decreased. Drug-paired cues and MDPV, cocaine, and methamphetamine pretreatments reinstated responding on both drug levers, regardless of preference.
Conclusion:
These data demonstrate that MDPV and cocaine act as economic substitutes and suggest that measures of reinforcing effectiveness determined under a progressive ratio schedule of reinforcement can predict drug choice. These data also suggest that environmental stimuli associated with a particular drug might stimulate class-specific drug-seeking, however, further studies are needed to test the generality of this claim.
Keywords: Concurrent drug access, MDPV, cocaine, self-administration, reinstatement
1. Introduction
Synthetic cathinones first emerged in the United States in 2009 in the form of “bath salts” preparations and quickly became popular as “legal” alternatives to illicit stimulants (e.g., cocaine, methamphetamine, and MDMA) (Jerry et al., 2012; Ross et al., 2012; Spiller et al.,2011). During this time, they garnered much attention in the popular press primarily due to frequent reports of bizarre behaviors, and adverse psychiatric (paranoia, hallucinations, aggression) and physiological (tachycardia, hyperthermia) events leading to large numbers of emergency room visits (Centers for Disease Control and Prevention (CDC), 2011; Johnson and Johnson, 2014; Kyle et al., 2011; Murray et al., 2012; Spiller et al., 2011). Despite the United States Drug Enforcement Administration placing 14 synthetic cathinones under Schedule I regulations, they remain one of the largest and most popular families of novel psychoactive substances (NPS) as evidenced by recent reports from the United Nations Office on Drugs and Crime (UNODC, 2020) as well as estimates suggesting that high school age individuals in the some parts of the U.S. use synthetic cathinones at rates comparable to either heroin or methamphetamine (Johnston et al., 2018; Palamar et al., 2019). One synthetic cathinone commonly found in “bath salts” preparations is 3,4-methylenedioxypyrovalerone, or MDPV. MDPV is a monoamine uptake inhibitor that is ~700 times more selective for inhibiting uptake at the dopamine transporter over the serotonin transporter (Baumann et al., 2013; Gannon et al., 2018; Simmler et al., 2013). Consistent with survey reports (Johnson and Johnson, 2014; Palamar et al., 2016), preclinical models have consistently demonstrated that MDPV has a high potential for abuse. Drug discrimination studies have shown that MDPV has discriminative stimulus effects that overlap with other commonly abused stimulants (e.g., cocaine, methamphetamine) (Berquist and Baker, 2017; Collins et al., 2016; Fantegrossi et al., 2013; Gannon et al., 2016; Gatch et al., 2013; Risca and Baker, 2019; Seaman Jr et al., 2021) and is readily self-administered by both rodents (Baumann et al., 2013; Gannon et al., 2018, 2017; Watterson et al., 2014) and non-human primates (Collins et al., 2019). Furthermore, when quantified by progressive ratio schedules of reinforcement as well as behavioral economic assessments of demand, MDPV has been shown to function as a more effective reinforcer than either cocaine or methamphetamine (Aarde et al., 2013; Collins et al., 2019; Gannon et al., 2018, 2017; Watterson et al., 2014; but note de Moura et al., 2021).
A complementary method to investigate the relative reinforcing effects of a drug is to provide subjects the opportunity to choose between multiple drugs (Katz, 1990). An index of choice can thereby be calculated as the number of times one drug is chosen relative to the number of total infusions earned. Compared to single-operant self-administration procedures, choice procedures provide not only information about the rate of responding and drug intake, but also information about the behavioral allocation towards each alternative. Choice procedures are based on the principle that, assuming all other factors are equal, subjects will allocate more behavior towards the larger or more effective alternative. For instance, monkeys provided access to two operanda reinforced by identical doses of the same drug allocated responding equally to each operandum. However, when monkeys were allowed to choose between two different doses of the same drug, they allocated behavior towards the operandum reinforced by the larger dose of drug (Anderson and Woolverton, 2003; Koffarnus and Woods, 2008). These demonstrations aid in interpreting allocation of behavior as an expression of preference that is determined by the magnitude of the reinforcing effects of each alternative.
Evaluating the reinforcing effects of drugs under both single-operant procedures (e.g., progressive ratio, economic demand) as well as procedures in which alternatives are available (e.g., choice) can provide a more complete characterization of their reinforcing effects. For instance, the reinforcing properties of different drugs shown to maintain similar breakpoints under a progressive ratio schedule of reinforcement do not necessarily produce equal levels of responding under conditions in which these same drugs are concurrently available. Ward and colleagues demonstrated that when cocaine, or mixtures of cocaine and heroin were available under a progressive ratio schedule of reinforcement, at no point did mixtures of cocaine and heroin maintain breakpoint greater than those maintained by cocaine alone. However, when these same doses of cocaine were made concurrently available with the same pairs of doses of cocaine and heroin mixtures, rats allocated greater levels of responding towards the lever reinforced by certain mixtures of cocaine and heroin than the lever reinforced by cocaine alone (Ward et al., 2005). Providing subjects concurrent access to two drugs also allows for investigation of the economic interactions between them by manipulating the price of one drug (e.g., increase of the ratio requirement or decrease of the available unit dose) while keeping the price of the alternative constant. If intake of the alternative drug increases, decreases, or stays the same, the drugs are deemed substitutes, complements, or independents of one another, respectively (Bickel et al., 1995; Hursh, 2000). Overall, the use of drug-versus-drug choice assays can provide important information about the reinforcing effects of a drug as well as how those effects are altered in the presence of an alternative that cannot be easily predicted from single-operant self-administration assays.
To this end, the current studies investigated drug-versus-drug choice behavior in rats that had concurrent access to MDPV and cocaine in order to test the hypotheses that: (1) when functionally equivalent doses are made concurrently available, rats will be indifferent, and allocate equal amounts of responding towards both MDPV and cocaine; (2) when the available dose of one drug is increased (or decreased), rats will allocate responding towards the lever reinforced by the functionally larger dose (i.e., MDPV and cocaine will act as economic substitutes); and (3) following extinction of responding for both drug alternatives, reinstatement of responding by re-exposure to the drug-paired stimuli will occur at comparable levels on levers previously reinforced by MDPV and cocaine whereas priming injections of MDPV or cocaine will reinstate greater levels of responding on the levers previously reinforced by MDPV or cocaine, respectively.
2. Methods
2.1. Subjects:
Eighteen male Sprague-Dawley rats (275–300 g upon arrival) were purchased from Envigo (Indianapolis, IN, USA) and maintained in a temperature- and humidity-controlled vivarium. Rats were individually housed and maintained on a 14/10-hour light/dark cycle (lights on at 6:00 AM). All experiments were conducted during the light cycle with sessions conducted at approximately the same time each morning. Rats were provided ad libitum access to Purina rat chow and water except during experimental sessions. All procedures were conducted in accordance with Institutional Animal Care and Use Committee at the University of Texas Health Science Center at San Antonio and the Guide for Care and Use of Laboratory Animals (National Research Council, 2011).
2.2. Surgery:
Rats were anesthetized with 2-3% isoflurane and prepared with chronic indwelling catheters in the left and right femoral veins using procedures similar to those previously described (Gannon et al., 2017; Seaman Jr and Collins, 2021). Catheters were tunneled under the skin and attached to a vascular access button placed in the mid-scapular region. Immediately following surgery, rats were administered Penicillin G (60,000 U/rat) subcutaneously to prevent infection. Rats were allowed 5–7 days to recover during which time both catheters were flushed daily with 0.5 ml of heparinized saline (100 U/ml). Thereafter, catheters were flushed daily with 0.2 ml of saline prior to, and 0.5 ml of heparinized saline after the completion of self-administration sessions. Catheter patency was assessed using 5 mg/kg methohexital as needed (e.g., an increase in pressure when flushing). If a rat was unresponsive to an intravenous injection of methohexital, a replacement catheter was implanted into the right jugular vein.
2.3. Drugs:
Racemic MDPV HCI was synthesized by Kenner Rice (Bethesda, MD). Cocaine HCI and heroin were provided by the National Institute on Drug Abuse Drug Supply Program (Bethesda, MD). D-methamphetamine was purchased from Sigma-Aldrich (St. Louis, MO, USA). All drugs were dissolved in sterile 0.9% saline and administered intravenously in a volume of 0.1 ml/kg (for self-administration) or 1 ml/kg (for reinstatement tests) body weight.
2.4. Apparatus:
All experiments were conducted in standard operant conditioning chambers located within ventilated, sound-attenuating enclosures (Med Associates, Inc., St. Albans, VT). Each chamber was equipped with two response levers located 6.8 cm above the grid floor and 1.3 cm from the right or left wall. Visual stimuli were provided by two sets of green, yellow, and red LEDs, one set located above each of the two levers, and a white house light located at the top center of the opposite wall. Drug solutions were delivered by a variable speed syringe pump through Tygon tubing connected to a stainless-steel fluid swivel and spring tether, which was held in place by a counterbalanced arm. Experimental events were controlled, and data were collected using MED-PC IV software and a PC-compatible interface (Med Associates, Inc.).
2.5. Self-administration
2.5.1. Acquisition:
Rats were initially allowed to respond for either 0.032 mg/kg/infusion of MDPV or 0.71 mg/kg/infusion of cocaine under a fixed ratio (FR) 1: timeout (TO) 5-second schedule of reinforcement during daily 90-minute sessions. Doses were chosen based on historical data from our laboratory in which full dose-response curves were generated for both MDPV and cocaine under a progressive ratio schedule of reinforcement in 61 rats (Gannon et al., 2018, 2017). Using the linear regressions that defined these functions, the dose of cocaine required to maintain the same number of infusions as 0.032 mg/kg/infusion of MDPV was estimated to be 0.71 mg/kg/infusion; thus these two doses were considered to be functionally equivalent in terms of reinforcing effectiveness (infusions earned). Two sets of conditioned stimuli (discriminative and infusion-paired) were used in these studies. One discriminative stimulus consisted of the illumination of a yellow LED above the active lever (left or right; counterbalanced across rats) that signaled drug availability (cocaine or MDPV; counterbalanced across rats). Completion of the response requirement on this lever resulted in a drug infusion (0.1 ml/kg over ~1 second) that was paired with the illumination of the yellow, green, and red LEDs above that lever as well as the houselight; these stimuli remained illuminated for the duration of the 5-second post-infusion timeout period during which drug was made unavailable. The other set of discriminative stimuli consisted of the illumination of green and red LEDs above the active lever (left or right; counterbalanced across rats) that signaled drug availability (cocaine or MDPV; counterbalanced across rats). Completion of the response requirement on this lever resulted in a drug infusion (0.1 ml/kg over ~1 second) that was paired with the flashing of the yellow, green, and red LEDs as well as the houselight, at 1 hz; this occurred throughout the 5-second post-infusion timeout period during which drug was made unavailable. Responses on the inactive lever, as well as responses on either lever during timeouts, were recorded but had no scheduled consequences. Acquisition criteria were as follows: ≥15 infusions for two consecutive days with ≥80% responding occurring on the active relative to inactive lever. Response requirements were subsequently increased to an FR 5 where they remained for the duration of the study. Once behavior met stability criteria for the initial drug (±25% of the mean of three consecutive sessions and no increasing or decreasing trend), rats were allowed to respond on the alternate lever for the alternate drug (and alternate set of conditioned stimuli) under an FR 5 schedule. The initially active lever now became inactive (i.e., the discriminative stimuli were omitted and responding had no programmed consequences). Throughout the entire acquisition period (i.e., acquisition of responding for both MDPV and cocaine; counterbalanced), the catheter through which drug infusions were delivered alternated daily in order to ensure that both catheters functioned equivalently.
2.5.2. Concurrent access:
After reaching stability under an FR 5 schedule for the second drug, rats were provided access to both drugs(or saline) and their associated stimuli under a concurrent FR5:FR5 schedule of reinforcement during daily 90-minute sessions. For all rats, the following conditions were evaluated in quasi-random order: 1) concurrent access to 0.032 mg/kg/infusion of MDPV and saline; 2) concurrent access to 0.71 mg/kg/infusion of cocaine and saline; and 3) concurrent access to 0.032 mg/kg/infusion of MDPV and 0.71 mg/kg/infusion of cocaine. Conditions remained in place for at least 7 days and until stability was reached, defined as ±25% of the mean of three consecutive sessions before moving to the next condition. Each session began with a 1-minute blackout followed by two forced choice trials, one on each lever, for the associated drug (or saline) and stimulus conditions. These forced choice trials were separated by a 1-minute blackout, and following completion, were followed another 1-minute blackout. The order of forced trails (i.e., drug and stimuli) was counter-balanced across rats but remained constant for a given rat throughout the course of the study. The session counter did not begin until both forced trials were completed. Throughout the remainder of the 90-minute session, rats had concurrent access to both drugs (or one drug and saline) and associated stimuli.
2.5.3. Dose-substitution:
Since responding was most often binary for one infusion or the other, a subset of rats (n=8) were used to evaluate the economic interactions between MDPV and cocaine by increasing the cost of the preferred drug or decreasing the cost of the non-preferred drug by decreasing or increasing the available unit dose, respectively. For instance, if a rat preferentially self-administered 0.032 mg/kg/infusion of MDPV when 0.71 mg/kg/infusion of cocaine was concurrently available, the unit dose of MDPV was decreased (0.0032 mg/kg/infusion MDPV versus 0.71 mg/kg/infusion cocaine) or the unit dose of cocaine was increased (0.032 mg/kg/infusion MDPV versus 1.78 mg/kg/infusion cocaine). Likewise, if a rat preferentially self-administered 0.71 mg/kg/infusion of cocaine when 0.032 mg/kg/infusion of MDPV was concurrently available, the unit dose of cocaine was decreased (0.032 mg/kg/infusion MDPV versus 0.071 mg/kg/infusion cocaine) or the unit dose of MDPV was increased (0.32 mg/kg/infusion MDPV versus 0.71 mg/kg/infusion cocaine). The order of these dose manipulations was quasi-random, with each condition maintained for at least 7 days and until stability was reached, defined as ±25% of the mean of three consecutive sessions.
2.6. Extinction and reinstatement:
The remaining subset of rats (n=10) were used to determine whether initial preferences for MDPV (0.032 mg/kg/infusion) or cocaine (0.71 mg/kg/infusion) would differentially impact the reinstatement of responding under conditions in which: 1) both the MDPV- and cocaine-associated stimuli were reintroduced; and 2) pretreatments with MDPV, cocaine, methamphetamine, or heroin were administered prior to sessions in which MDPV- and cocaine-associated stimuli were reintroduced. Briefly, under extinction conditions, completion of response requirements on the cocaine or MDPV levers had no programmed consequences (i.e., no infusions or infusion-paired stimuli were delivered). Extinction conditions remained in place for at least 7 sessions, and until the total number of lever responses on both levers was ≤15% of baseline responding. Once extinction criteria were met, reinstatement testing was initiated. A series of 5 reinstatement tests were performed as described previously (Doyle et al., 2021). Briefly, reinstatement tests were identical to self-administration conditions with the exceptions that: 1) intravenous pretreatments of saline (cue-induced reinstatement) or drug (cue + drug-primed reinstatement) were administered 5 minutes before the session; 2) forced trials were omitted from the session; and 3) saline rather than drug was delivered upon completion of response requirements. Cue-induced reinstatement always occurred first followed by four additional cue+drug-primed reinstatement tests. Intravenous pretreatments with cocaine (3.2 mg/kg), MDPV (0.32 mg/kg), methamphetamine (1 mg/kg), or heroin (0.1 mg/kg) occurred in a quasi-random order, with each reinstatement test separated by at least two extinction sessions; additional extinction sessions were conducted until the extinction criterion was met.
2.7. Data analysis
All data are presented as the mean ±S.E.M. For dose-substitution studies, the percent choice of 0.032 mg/kg/infusion of MDPV is shown as a function of cocaine dose (or saline) whereas the percent choice of 0.71 mg/kg/infusion of cocaine is shown as a function of MDPV dose (or saline). Data represent the final day of each dose-substitution period and were analyzed via a mixed-effects repeated measure one-way analysis of variance (ANOVA) and post-hoc Dunnett’s test. Extinction data were analyzed both on the group level and by preference via a two-way repeated measure ANOVA and post-hoc Dunnett’s test. Similarly, reinstatement data were analyzed via a two-way repeated measure ANOVA and post-hoc Dunnet’s test when comparing total responding to extinction, and Bonferonni’s test when comparing allocation of responding on each lever produced by each pretreatment.
3. Results
3.1. Single-drug access
All rats provided access to MDPV (0.032 mg/kg/infusion) met acquisition criteria by day ten, with no apparent difference of which catheter drug was infused through (Fig. 1; upper left). MDPV intake was maintained upon increasing the fixed ratio to five (Fig. 1; upper right). When cocaine (0.71 mg/kg/infusion) was initially substituted for MDPV and reinforced by the previously inactive lever, there were comparable responses made on the newly active and inactive levers. However, by the end of seven days, rats responded nearly exclusively on the cocaine-reinforced lever (Fig. 1; upper right). Similarly, by day ten, acquisition criteria were met in all rats provided access to cocaine (0.71 mg/kg/infusion), with no apparent difference of which catheter drug was infused through (Fig. 1; lower left), and intake was maintained upon increasing the fixed ratio to five (Fig. 1; lower right). When MDPV (0.032 mg/kg/infusion) was substituted for cocaine and reinforced by the previously inactive lever, responding was allocated to the newly active lever over the course of the seven days (Fig. 1; lower right).
Figure 1:
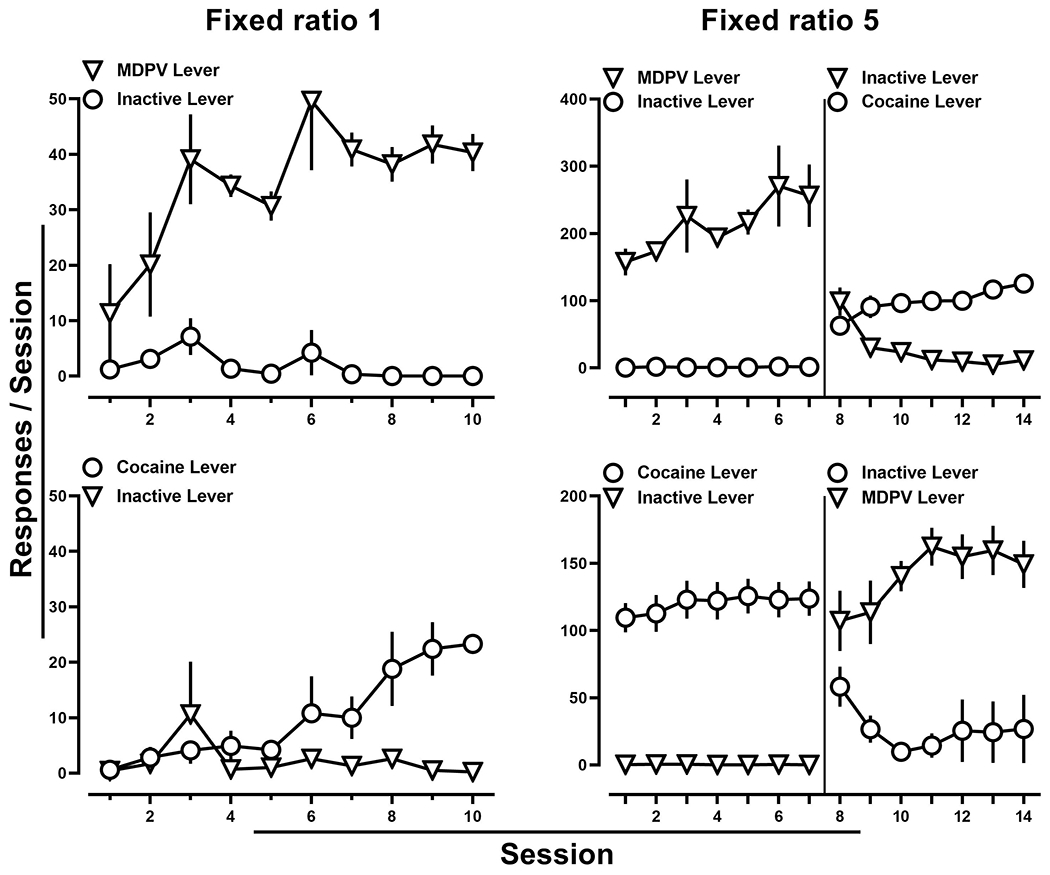
Number of active and inactive lever responses throughout a 10-day acquisition period for MDPV (0.032 mg/kg/infusion) (upper left) or cocaine (0.71 mg/kg/infusion) (lower left). Subsequent responding under a fixed ratio five schedule of reinforcement for MDPV and when cocaine was substituted on the previously inactive lever (upper right). Similarly, responding under a fixed ratio five schedule of reinforcement for cocaine followed by MDPV substitution on the previously inactive lever (lower right). The solid line represents when the alternate drug and drug-paired stimuli were made available on the alternate lever. Data represent the mean ±S.E.M., and each point represents 8-10 rats.
3.2. Concurrent access
Subsequently, rats were provided access to either 0.032 mg/kg/inf of MDPV and saline, 0.71 mg/kg of cocaine and saline, or 0.032 mg/kg/inf of MDPV and 0.71 mg/kg of cocaine, in a pseudorandom order. When MDPV and saline (Fig. 2; left) or cocaine and saline (Fig 2. right) were concurrently available, responding was always primarily allocated towards the lever that was reinforced by drug. In contrast, when these doses of MDPV and cocaine were made concurrently available, responding was comparable for both MDPV and cocaine, on the group level (Fig. 3; left). However, examination of individual subject data revealed that one population of rats (n=10) responded exclusively on the MDPV-reinforced lever (Fig. 3; middle), whereas the remaining rats (n=8) responded exclusively on the cocaine-reinforced lever (Fig. 3; right). These populations were deemed, “MDPV-preferring” and “cocaine-preferring” rats, respectively.
Figure 2:
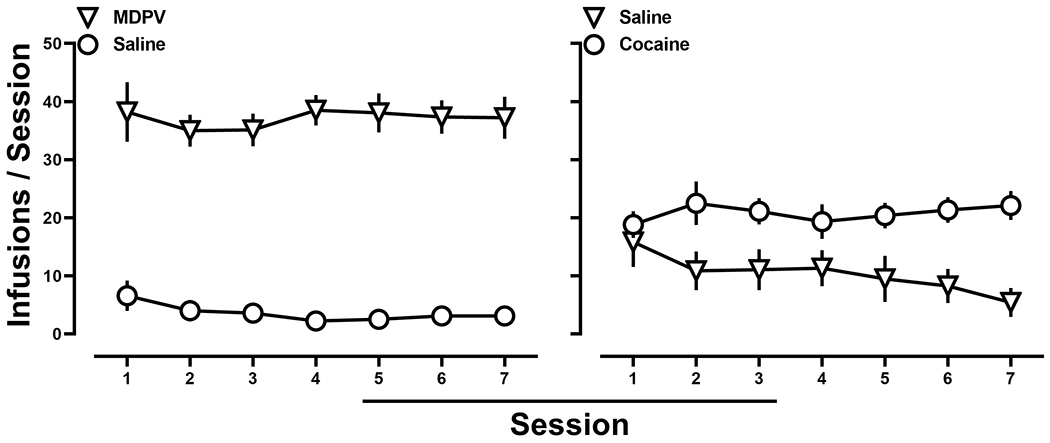
Left; Average number of infusions of 0.032 mg/kg/infusion MDPV (inverted triangles) and saline (filled circles) when MDPV and saline were concurrently available. Right; Average number of infusions of 0.71 mg/kg/infusion cocaine (circles) and saline (filled inverted triangles) when cocaine and saline were concurrently available. Data represent the mean ±S.E.M., and each point represents 18 rats.
Figure 3:
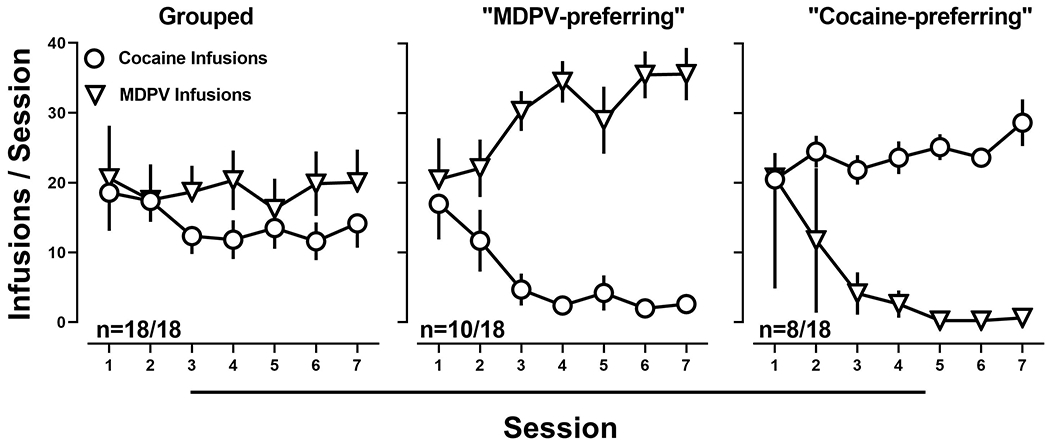
Average number of infusions of 0.032 mg/kg/infusion of MDPV (inverted triangles) and 0.71 mg/kg/infusion of cocaine (circles) when MDPV and cocaine were concurrently available on the group level (left; n=18), and in rats categorized as MDPV-preferring (center; n=10) and cocaine-preferring (right; n=8). Data represent the mean ±S.E.M.
3.3. Dose substitution
To evaluate whether MDPV and cocaine may act as economic substitutes, in a subset of rats the cost of one drug was either increased or decreased whereas the cost of the alternative drug remained fixed. When the price of MDPV was fixed (0.032 mg/kg/infusion), choice of MDPV increased as the cost of cocaine increased (i.e., choice of MDPV increased as the dose of cocaine decreased) (Fig. 4; left). A significant effect of dose (F [3,20] = 23.7; p < 0.0001) was revealed by a one-way repeated measure ANOVA. Post-hoc tests revealed a significant decrease in percent MDPV choice when either 0.71 mg/kg/infusion (47.5 ±18%) or 1.78 mg/kg/infusion (1 ±1%) of cocaine was made concurrently available, compared to saline (98.1 ±1.2%). Similarly, when the price of cocaine remained constant (0.71 mg/kg/infusion), choice of cocaine increased as the cost of MDPV increased (i.e., choice of cocaine increased as the dose of MDPV decreased) (Fig. 4; right). A significant effect of dose (F [3,20] = 20; p < 0.0001) was revealed by a one-way repeated measure ANOVA. Post-hoc tests revealed a significant decrease in percent cocaine choice when either 0.032 mg/kg/infusion (52.5 ±18%) or 0.32 mg/kg/infusion (7.1 ±7.1%) of MDPV was made concurrently available, compared to saline (95.6 ±2.2%). Data from individual subjects are presented in Supplemental Fig. 1.
Figure 4:
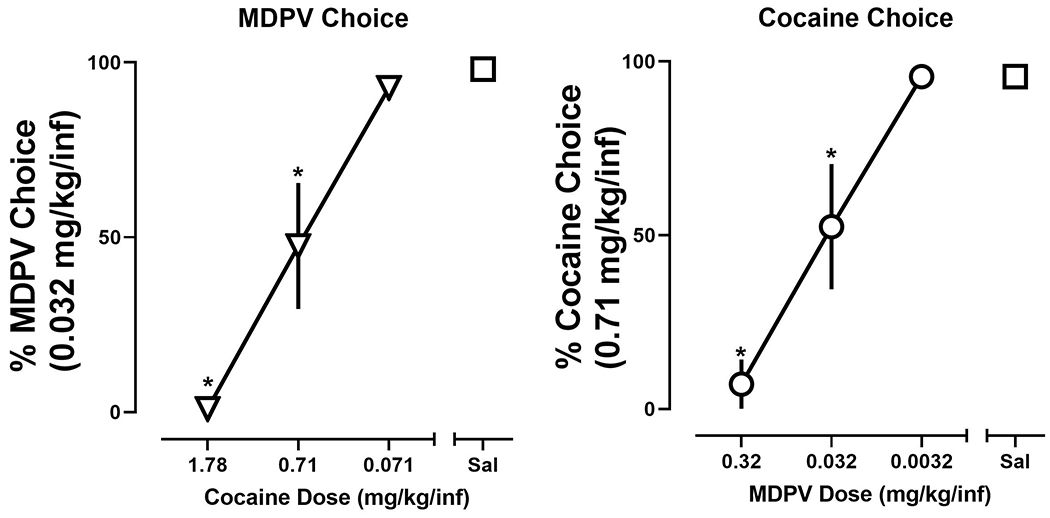
Percent choice of 0.032 mg/kg/infusion of MDPV as a function of concurrently available cocaine dose (or saline) (left). Percent choice of 0.71 mg/kg/infusion of cocaine as a function of concurrently available MDPV dose (or saline) (right). Data represent the mean ±S.E.M. Asterisks represent a significant decrease from saline (p < 0.05).
3.4. Extinction and reinstatement
The remaining rats were used to investigate the impact of concurrent access to two drugsas well as drug preference on extinction and reinstatement behavior. Again, on the group level,responding for MDPV and cocaine were comparable when made concurrently available, the result of a subset of rats self-administering MDPV nearly exclusively whereas the remaining rats self-administered cocaine nearly exclusively. Upon instituting extinction conditions, responding on the MDPV and cocaine levers decreased across days with extinction criteria being met on day 4 ±0.7. A two-way repeated-measure ANOVA revealed that there was no interaction between time and lever (F [6,54] = 0.44; p > 0.05) and no difference in extinction of responding on the MDPV and cocaine levers (F [1,9] =0.03; p > 0.05), but there was a main effect of time (F [6,54] = 5; p < 0.001) (Fig. 5; left). When evaluating extinction behavior based on preference, MDPV-preferring rats decreased responding across days with extinction criteria being met on day 4 ±1. A two-way repeated-measure ANOVA revealed that there was no interaction between time and lever (F [6,30] = 1.7; p > 0.05) and no difference in extinction of responding on the MDPV and cocaine levers (F [1,5] =0.2; p > 0.05), but there was a main effect of time (F [6,30] = 7.8; p < 0.0001) (Fig. 5; middle). Similarly, in cocaine-preferring rats, responding decreased across days with extinction criteria being met on day 4 ±.08. A two-way repeated-measure ANOVA revealed that there was no interaction between time and lever (F [6,18] = 1.5, p > 0.05), no difference in extinction of responding on the MDPV and cocaine levers (F [1,3] =3.6; p > 0.05), and no main effect of time (F [6,18] = 0.6; p > 0.05) (Fig. 5; right).
Figure 5:
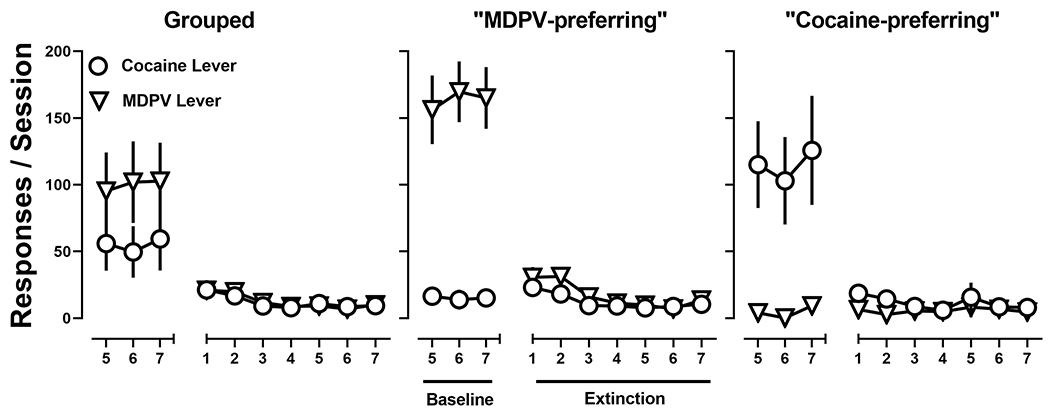
Responses made on the MDPV lever (inverted triangles) and cocaine lever (circles) on the group level (left), in MDPV-preferring rats (middle; n=6) and cocaine-preferring rats (right; n=4) rats the final three days of concurrent access to MDPV and cocaine self-administration, and extinction conditions.
Subsequently, a series of reinstatement tests were conducted. A two-way repeated measures ANOVA revealed no significant interaction between pretreatment and lever (F [1.1,9.9] = 3.1; p > 0.05), but significant main effects of both pretreatment (F [1.5,13.3] = 18; p < 0.001) and lever (F [1,9] = 6; p < 0.05). Post-hoc analyses revealed that reinstatement of responding on both the MDPV and cocaine levers was significantly increased (p < 0.05) when drug-paired cues were reintroduced, as well as drug-paired cues in conjunction with pretreatments of MDPV, cocaine, heroin, and methamphetamine, relative to the final day of extinction (Fig. 6). Furthermore, post-hoc analyses revealed that cocaine-primed reinstatement led to significantly more responding on the cocaine relative to MDPV lever (p < 0.05), but no other statistically significant differences between MDPV and cocaine lever responding were observed throughout reinstatement tests (Fig. 6). In addition, comparable allocation of responding was observed regardless of MDPV (open symbols) or cocaine (filled symbols) preference.
Figure 6:
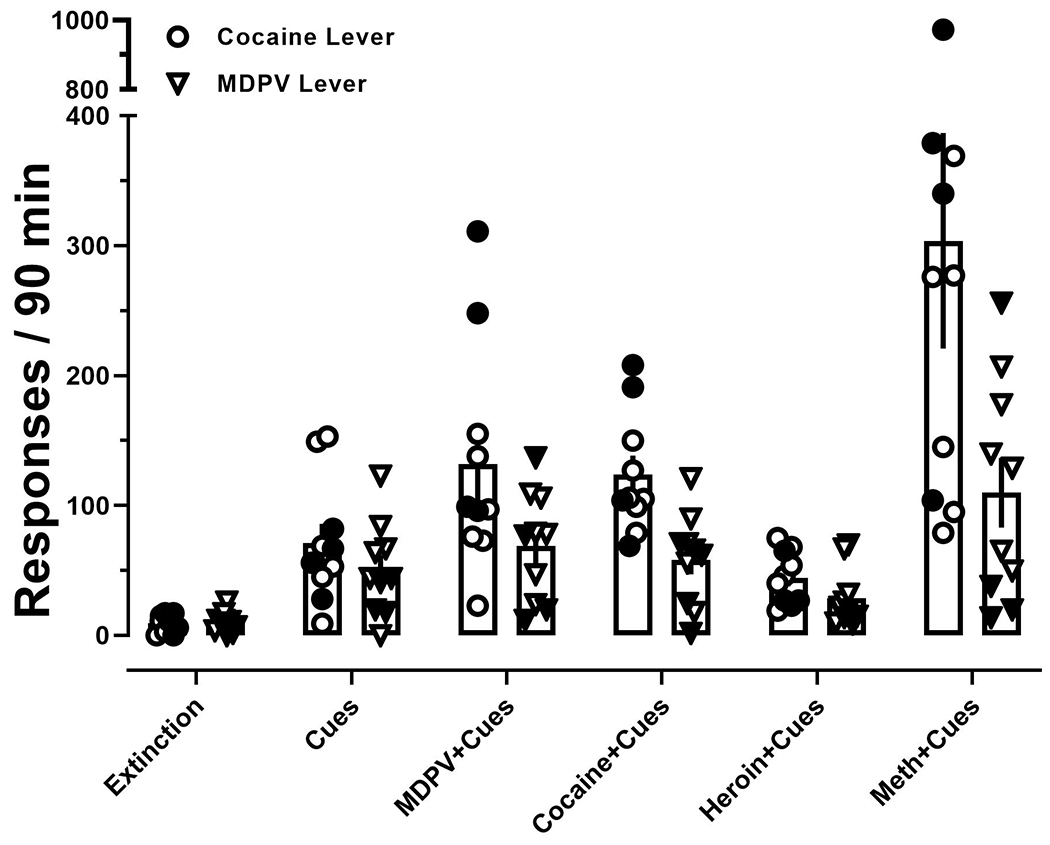
Responses made on the MDPV lever (inverted triangles) and cocaine lever (circles) in MDPV-preferring rats (open symbols) and cocaine-preferring rats (filled symbols) on the final day of extinction and during reinstatement tests. Data represent the mean ±S.E.M.
4. Discussion
Despite similar mechanisms of action, MDPV has been shown to be more reinforcing than cocaine when assessed via progressive ratio schedules of reinforcement or demand curve analyses (Collins et al., 2019; Gannon et al., 2018, 2017). The current studies extend these findings by evaluating choice behavior under conditions in which MDPV and cocaine were concurrently available. When provided concurrent access to functionally equivalent doses of MDPV and cocaine (i.e., doses that maintain similar breakpoints under a progressive ratio schedule of reinforcement), MDPV and cocaine maintained comparable levels of responding on the group level. This group-level indifference resulted from half of the rats responding exclusively on the MDPV-reinforced lever (MDPV-preferring rats) and the remaining half responding exclusively on the cocaine-reinforced lever (cocaine-preferring rats). Substituting a smaller dose of the preferred drug resulted in a reallocation of responding toward the initially non-preferred drug. Similarly, substituting a larger dose of the initially non-preferred drug also resulted in reallocation of behavior away from the preferred drug and toward the initially non-preferred alternative. Lastly, reinstatement of extinguished responding by reintroduction of drug-paired cues, or drug-paired cues in addition to a priming injection of MDPV, cocaine, or methamphetamine, produced responding on both the MDPV and cocaine levers, with no differences observed as a function of preference. Taken together, these data demonstrate that MDPV and cocaine act as economic substitutes and that reinforcing effectiveness determined under a progressive ratio schedule of reinforcement can, at least in the case of economic substitutes, predict drug choice.
When rats were provided concurrent access to doses of MDPV (0.032 mg/kg/inf) and cocaine (0.71 mg/kg/inf) that maintain comparable breakpoints under a progressive ratio schedule of reinforcement (Gannon et al., 2018, 2017), responding was equally distributed between the MDPV- and cocaine-reinforced levers, suggesting that rats were indifferent between these doses of MDPV and cocaine, at least at the group level. Examination of individual subject data illustrated that approximately half of the subjects responded nearly exclusively on the MDPV-reinforced lever, whereas the remaining rats responded nearly exclusively on the cocaine-reinforced lever. Importantly, these preferences were not simply a result of the drug upon which a rat acquired responding, the order of forced trials for a given rat, or perseverative responding. One likely explanation for the apparent preferences observed for either MDPV or cocaine in the current studies is that slight individual differences exist with regard to the relative reinforcing effects of the training doses of MDPV and cocaine. For MDPV-preferring rats, 0.032 mg/kg/inf MDPV was likely more reinforcing than 0.71 mg/kg/inf cocaine, whereas the opposite was likely true in cocaine-preferring rats. Comparing MDPV and cocaine through a concurrent access procedure aids in provided a more complete characterization of the reinforcing effects of these drugs. Indeed, when compared using a single-drug access, economic demand curve procedure, two doses of MDPV on the descending limb of the MDPV fixed ratio dose-response curve (0.032 and 0.1 mg/kg/inf) resulted in a smaller alpha value than two doses of cocaine at comparable positions on the descending limb of the cocaine fixed ratio dose response curve (0.32 and 1 mg/kg/inf), suggesting that the reinforcing effects of MDPV greater than those of cocaine (Gannon et al., 2018). However, in the current studies, when large (i.e., located on the descending limbs of a fixed ratio dose-response curve) doses of MDPV and cocaine were made concurrently available, choice was not exclusively allocated towards MDPV, as might be predicted, but rather was dictated by the relative cost of each alternative. An important future study is to directly compare the doses of MDPV and cocaine that maintain the greatest amount of responding under a progressive ratio schedule of reinforcement (i.e., 1.78 mg/kg/inf cocaine vs. 0.178 mg/kg/inf MDPV; Gannon et al., 2018) in order to determine progressive ratio breakpoints can be used to predict choice behavior.
It is also worth noting that the training dose of MDPV (0.032 mg/kg/inf) maintained higher rates of responding than an “equi-effective” dose of cocaine (0.71 mg/kg/inf), an effect that was observed both during acquisition (i.e., when rats only had access to one drug) as well as in the concurrent access portion of the study. Although these differences could be explained by an imperfect selection of “equi-effective” doses, it is also likely influenced by other pharmacokinetic and pharmacodynamic properties of the drugs that can impact both rates and total amounts (e.g., breakpoints) of responding under both fixed and progressive ratio schedules of reinforcement (Griffiths et al., 1979; Katz, 1990; Ko et al., 2002; Richardson and Roberts, 1996; Roberts, 1993). For instance, GBR 12909, a relatively selective inhibitor of reuptake at the dopamine transporter with a longer duration of action than cocaine, maintains less responding than cocaine under a fixed ratio schedule of reinforcement, but similar maximal breakpoints under a progressive ratio schedule of reinforcement (Roberts, 1993).
In order to probe whether these initial preferences were determined by relative reinforcing effectiveness (or price), a series of dose manipulations were conducted. Indeed, when the dose of the preferred drug was reduced (price of preferred drug was increased) allocation of responding was reversed. These findings are consistent with what has been observed in rhesus monkeys provided concurrent access to different doses of cocaine available for responding on two operanda. As has been outlined previously, increasing the price for one drug (i.e., decreasing the dose made available or increasing the response requirement) while the price of a concurrently available drug is held constant produces different types of economic interactions (Bickel et al., 1995; Hursh, 2000). The current data suggest that MDPV and cocaine act as substitutes, in that when the cost of one drug was increased, intake of the alternative drug increased, and vice-versa. These studies aid in laying the groundwork to investigate economic interactions between commonly used drugs from different classes (e.g., opioids and stimulants) in rats. Although only male subjects were used in the current studies due to the historical progressive ratio data that were analyzed to determine equi-effective doses of MDPV and cocaine being collected in all male subjects, future studies should incorporate females to determine whether this same economic relationship exists between MDPV and cocaine.
Finally, the current studies found that cue-induced reinstatement produced similar levels of responding on the MDPV- and cocaine-paired levers, regardless of drug preference. This could be, in part, due to both sets of cues acquiring conditioned properties as a function of their being paired with either MDPV- or cocaine-reinforcement, given that rats had a history of responding exclusively on each lever. An additional consideration is that all subjects had a history of concurrent access to MDPV and saline, and cocaine and saline, prior to extinction of responding on both levers, which could contribute to rats responding on both levers during reinstatement tests. Similar to what was observed when cues alone were reintroduced, pretreatment with MDPV, cocaine, or methamphetamine produced responding on both the MDPV and cocaine levers, albeit more responding generally occurred on the cocaine lever, regardless of MDPV or cocaine preference. A priming injection of heroin paired with conditioned stimuli did not result in responding on either lever that was greater than those produced by the conditioned stimuli (i.e., cues) alone. The underlying driver of greater levels of cocaine-lever responding in response to pretreatments of MDPV, cocaine, and methamphetamine is unclear but could be a result of the single doses of each priming drug being evaluated. The cumulative records from these reinstatement tests show that the initial ten ratios completed tended to be on the cocaine lever, regardless of initial drug preference, further supporting that the doses evaluated could have produced more cocaine-like discriminative stimulus effects. Evaluation of additional doses would provide greater resolution on the relationship between drug preference and possible drug seeking-related behaviors. Given the overlapping discriminative stimulus effects of MDPV, cocaine, and methamphetamine (Berquist and Baker, 2017; Collins et al., 2016; Fantegrossi et al., 2013; Gannon et al., 2016; Gatch et al., 2013; Risca and Baker, 2019; Seaman Jr et al., 2021), and the role of these effects in mediating drug-primed reinstatement, it is perhaps unsurprising that responding was allocated to both levers and produced more responding than the conditioned stimuli alone. If the effects observed in the current studies are due to an overlap in the discriminative stimulus effects of the two drugs, then one would expect rats with a history of concurrent access to drugs from different classes (e.g., a stimulant and an opioid) to reinstate only on the lever previously reinforced by the priming drug (or another drug from that class). However, if the reinstatement of responding on both levers is due to the fact that rats had a history of concurrent drug-reinforcement on the two levers then one would expect rats with a history of concurrent access to drugs from different classes to also reinstate on both levers regardless of the priming drug. Lastly, MDPV being more selective than cocaine for inhibiting the norepinephrine transporter relative to the serotonin transporter (Baumann et al., 2013; Gannon et al., 2018; Simmler et al., 2013) could result in the discriminative stimulus effects of stressors (e.g., footshock, yohimibine) that are known to reinstate responding being more similar to those of MDPV than cocaine. If true, and assuming discriminative stimulus effects play an important role in directing the reinstatement of responding under a concurrent access procedure, then one would predict that stress-primed reinstatement might produce a selective, or at least preferential, reinstatement of responding on the MDPV relative to cocaine lever.
5. Conclusions
Despite the growing awareness that polysubstance use is the norm rather than the exception, the vast majority of preclinical substance use research continues to focus on the effects of individual drugs, studied in isolation. This dichotomy highlights the need to characterize interactions amongst commonly co-abused drugs in order to better understand the determinants of polysubstance use. Indeed, the current studies demonstrate that MDPV and cocaine act as economic substitutes, suggesting that preferences in the drug-taking behavior of individuals that use multiple drugs from the same class (e.g., cocaine and MDPV) is likely determined in large part by availability and cost. If an individual typically uses cocaine, they may use MDPV when cocaine is too expensive or difficult to find. It could also be the case that these preferences may switch altogether when less expensive alternatives become readily available (e.g., cheap “bath salts” products available for purchase online or in shops). Furthermore, reinstatement of responding by drug-paired stimuli and priming injections of MDPV, cocaine, and methamphetamine produced similar allocation of responding on both MDPV- and cocaine-paired levers. This suggests that environmental stimuli associated with a particular drug (e.g., cocaine) might stimulate drug-seeking that is class-specific, however, further studies with concurrent access to drugs from different classes are needed to better understand these complex relationships. Future studies allowing rats concurrent access to opioids and stimulants could provide important information regarding economic interactions between these drugs and conditions that contribute to drug choice behavior, with the ultimate goal of developing better therapeutic outcomes for individuals engaging in polysubstance use.
Supplementary Material
Highlights.
Polysubstance use is far more prevalent than use of a single substance.
Drug-vs-drug choice can be used to characterize interactions between different drugs.
At the group level rats were indifferent to equi-effective doses of MDPV and cocaine.
Increasing MDPV cost resulted in increased cocaine intake and vice-versa.
Level of reinstatement was not determined by initial preference for MDPV or cocaine.
Acknowledgements
The authors would like the thank Rachel DeSantis, Victoria Campos, Karen Jimenez, and Juan Morales for their technical assistance in the completion of these studies.
Role of funding source
These studies were funded in part by research grants from the National Institutes of Health and National Institute on Drug Abuse (R01DA039146 [GTC]), and the Intramural Research Programs of the National Institute on Drug Abuse and the National Institute of Alcohol Abuse and Alcoholism (KCR). The sponsors did not play a role in the study design, the collection, analysis and interpretation of data, the writing of the manuscript, or in the decision to submit the article for publication.
Funding:
This research was supported by National Institutes of Health and National Institute on Drug Abuse (R01DA039146 [GTC], and the Intramural Research Programs of the National Institute on Drug Abuse and the National Institute of Alcohol Abuse and Alcoholism (KCR).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Aarde SM, Huang PK, Creehan KM, Dickerson TJ, Taffe MA, 2013. The novel recreational drug 3,4-methylenedioxypyrovalerone (MDPV) is a potent psychomotor stimulant: self-administration and locomotor activity in rats. Neuropharmacology 71, 130–140. 10.1016/j.neuropharm.2013.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson KG, Woolverton WL, 2003. Effects of dose and infusion delay on cocaine self-administration choice in rhesus monkeys. Psychopharmacology (Berl.) 167, 424–430. 10.1007/s00213-003-1435-9 [DOI] [PubMed] [Google Scholar]
- Baumann MH, Partilla JS, Lehner KR, Thorndike EB, Hoffman AF, Holy M, Rothman RB, Goldberg SR, Lupica CR, Sitte HH, Brandt SD, Tella SR, Cozzi NV, Schindler CW, 2013. Powerful cocaine-like actions of 3,4-methylenedioxypyrovalerone (MDPV), a principal constituent of psychoactive “bath salts” products. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol 38, 552–562. 10.1038/npp.2012.204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berquist MD, Baker LE, 2017. Characterization of the discriminative stimulus effects of 3,4-methylenedioxypyrovalerone in male Sprague-Dawley rats. Behav. Pharmacol 28, 394–400. 10.1097/FBP.0000000000000310 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bickel WK, DeGrandpre RJ, Higgins ST, 1995. The behavioral economics of concurrent drug reinforcers: a review and reanalysis of drug self-administration research. Psychopharmacology (Berl.) 118, 250–259. 10.1007/BF02245952 [DOI] [PubMed] [Google Scholar]
- Centers for Disease Control and Prevention (CDC), 2011. Emergency department visits after use of a drug sold as “bath salts”--Michigan, November 13, 2010-March 31, 2011. MMWR Morb. Mortal. Wkly. Rep 60, 624–627. [PubMed] [Google Scholar]
- Collins GT, Abbott M, Galindo K, Rush EL, Rice KC, France CP, 2016. Discriminative Stimulus Effects of Binary Drug Mixtures: Studies with Cocaine, MDPV, and Caffeine. J. Pharmacol. Exp. Ther 359, 1–10. 10.1124/jpet.116.234252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins GT, Sulima A, Rice KC, France CP, 2019. Self-administration of the synthetic cathinones 3,4-methylenedioxypyrovalerone (MDPV) and α-pyrrolidinopentiophenone (α-PVP) in rhesus monkeys. Psychopharmacology (Berl.) 236, 3677–3685. 10.1007/s00213-019-05339-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Moura FB, Sherwood A, Prisinzano TE, Paronis CA, Bergman J, Kohut SJ, 2021. Reinforcing effects of synthetic cathinones in rhesus monkeys: Dose-response and behavioral economic analyses. Pharmacol. Biochem. Behav 202, 173112. 10.1016/j.pbb.2021.173112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doyle MR, Sulima A, Rice KC, Collins GT, 2021. Interactions between reinforcement history and drug-primed reinstatement: Studies with MDPV and mixtures of MDPV and caffeine. Addict. Biol 26, e12904. 10.1111/adb.12904 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fantegrossi WE, Gannon BM, Zimmerman SM, Rice KC, 2013. In vivo effects of abused “bath salt” constituent 3,4-methylenedioxypyrovalerone (MDPV) in mice: drug discrimination, thermoregulation, and locomotor activity. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol 38, 563–573. 10.1038/npp.2012.233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gannon BM, Baumann MH, Walther D, Jimenez-Morigosa C, Sulima A, Rice KC,Collins GT, 2018. The abuse-related effects of pyrrolidine-containing cathinones are related to their potency and selectivity to inhibit the dopamine transporter. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol 43, 2399–2407. 10.1038/S41386-018-0209-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gannon BM, Galindo KI, Rice KC, Collins GT, 2017. Individual Differences in the Relative Reinforcing Effects of 3,4-Methylenedioxypyrovalerone under Fixed and Progressive Ratio Schedules of Reinforcement in Rats. J. Pharmacol. Exp. Ther 361, 181–189. 10.1124/jpet.116.239376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gannon BM, Williamson A, Suzuki M, Rice KC, Fantegrossi WE, 2016. Stereoselective Effects of Abused “Bath Salt” Constituent 3,4-Methylenedioxypyrovalerone in Mice: Drug Discrimination, Locomotor Activity, and Thermoregulation. J. Pharmacol. Exp. Ther 356, 615–623. 10.1124/jpet.115.229500 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gatch MB, Taylor CM, Forster MJ, 2013. Locomotor stimulant and discriminative stimulus effects of “bath salt” cathinones. Behav. Pharmacol 24, 437–447. 10.1097/FBP.0b013e328364166d [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffiths RR, Bradford LD, Brady JV, 1979. Progressive ratio and fixed ratio schedules of cocaine-maintained responding in baboons. Psychopharmacology (Berl.) 65, 125–136. 10.1007/BF00433038 [DOI] [PubMed] [Google Scholar]
- Hursh SR, 2000. Behavioral economic concepts and methods for studying health behavior. Reframing Health Behav. Change Behav. Econ 27–60. [Google Scholar]
- Jerry J, Collins G, Streem D, 2012. Synthetic legal intoxicating drugs: the emerging “incense” and “bath salt” phenomenon. Cleve. Clin. J. Med 79, 258–264. 10.3949/ccjm.79a.11147 [DOI] [PubMed] [Google Scholar]
- Johnson PS, Johnson MW, 2014. Investigation of “bath salts” use patterns within an online sample of users in the United States. J. Psychoactive Drugs 46, 369–378. 10.1080/02791072.2014.962717 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnston LD, Miech RA, O’Malley PM, Bachman JG, Schulenberg JE, Patrick ME, n.d. 2018. Overview Key Findings on Adolescent Drug Use 126. [Google Scholar]
- Katz JL, 1990. Models of relative reinforcing efficacy of drugs and their predictive utility. Behav. Pharmacol 1, 283–301. 10.1097/00008877-199000140-00003 [DOI] [PubMed] [Google Scholar]
- Ko MC, Terner J, Hursh S, Woods JH, Winger G, 2002. Relative reinforcing effects of three opioids with different durations of action. J. Pharmacol. Exp. Ther. 301, 698–704. 10.1124/jpet.301.2.698 [DOI] [PubMed] [Google Scholar]
- Koffarnus MN, Woods JH, 2008. Quantification of drug choice with the generalized matching law in rhesus monkeys. J. Exp. Anal. Behav 89, 209–224. 10.1901/jeab.2008.89-209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyle PB, Iverson RB, Gajagowni RG, Spencer L, 2011. Illicit bath salts: not for bathing. J. Miss. State Med. Assoc 52, 375–377. [PubMed] [Google Scholar]
- Murray BL, Murphy CM, Beuhler MC, 2012. Death following recreational use of designer drug “bath salts” containing 3,4-Methylenedioxypyrovalerone (MDPV). J. Med. Toxicol. Off. J. Am. Coll. Med. Toxicol 8, 69–75. 10.1007/s13181-011-0196-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palamar JJ, Barratt MJ, Ferris JA, Winstock AR, 2016. Correlates of new psychoactive substance use among a self-selected sample of nightclub attendees in the United States. Am. J. Addict. 25, 400–407. 10.1111/ajad.12403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palamar JJ, Rutherford C, Keyes KM, 2019. “Flakka” use among high school seniors in the United States. Drug Alcohol Depend. 196, 86–90. 10.1016/j.drugalcdep.2018.12.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson NR, Roberts DC, 1996. Progressive ratio schedules in drug self-administration studies in rats: a method to evaluate reinforcing efficacy. J. Neurosci. Methods 66, 1–11. 10.1016/0165-0270(95)00153-0 [DOI] [PubMed] [Google Scholar]
- Risca HI, Baker LE, 2019. Contribution of monoaminergic mechanisms to the discriminative stimulus effects of 3,4-methylenedioxypyrovalerone (MDPV) in Sprague-Dawley rats. Psychopharmacology (Berl.) 236, 963–971. 10.1007/s00213-018-5145-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts DC, 1993. Self-administration of GBR 12909 on a fixed ratio and progressive ratio schedule in rats. Psychopharmacology (Berl.) 111, 202–206. 10.1007/BF02245524 [DOI] [PubMed] [Google Scholar]
- Ross EA, Reisfield GM, Watson MC,Chronister CW, Goldberger BA, 2012. Psychoactive “bath salts” intoxication with methylenedioxypyrovalerone. Am. J. Med 125, 854–858. 10.1016/j.amjmed.2012.02.019 [DOI] [PubMed] [Google Scholar]
- Seaman RW Jr, Collins GT, 2021. Impact of Morphine Dependence and Withdrawal on the Reinforcing Effectiveness of Fentanyl, Cocaine, and Methamphetamine in Rats. Front. Pharmacol 12, 1266. 10.3389/fphar.2021.691700 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seaman RW Jr Doyle MR, Sulima A, Rice KC, Collins GT, 2021. Discriminative stimulus effects of 3,4-methylenedioxypyrovalerone (MDPV) and structurally related synthetic cathinones. Behav. Pharmacol 10.1097/FBP.0000000000000624 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simmler LD, Buser TA, Donzelli M, Schramm Y, Dieu L-H, Huwyler J, Chaboz S, Hoener MC, Liechti ME, 2013. Pharmacological characterization of designer cathinones in vitro. Br. J. Pharmacol 168, 458–470. 10.1111/j.1476-5381.2012.02145.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spiller HA, Ryan ML, Weston RG, Jansen J, 2011. Clinical experience with and analytical confirmation of “bath salts” and “legal highs” (synthetic cathinones) in the United States. Clin. Toxicol. Phila. Pa 49, 499–505. 10.3109/15563650.2011.590812 [DOI] [PubMed] [Google Scholar]
- United Nations Office on Drugs and Crime, 2020. Current NPS Threats, Vol III, October 2020. [Google Scholar]
- Ward SJ, Morgan D, Roberts DCS, 2005. Comparison of the reinforcing effects of cocaine and cocaine/heroin combinations under progressive ratio and choice schedules in rats. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol 30, 286–295. 10.1038/sj.npp.1300560 [DOI] [PubMed] [Google Scholar]
- Watterson LR, Kufahl PR, Nemirovsky NE, Sewalia K, Grabenauer M, Thomas BF, Marusich JA, Wegner S, Olive MF, 2014. Potent rewarding and reinforcing effects of the synthetic cathinone 3,4-methylenedioxypyrovalerone (MDPV). Addict. Biol 19, 165–174. 10.1111/j.1369-1600.2012.00474.x [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.