

Abstract
Objective
Many women are affected by vulvodynia, but medical therapies to date have proven ineffective. We performed a pilot study using gel-based proteomics to develop a map of proteins present in vaginal/vestibular secretions and identify proteins that could be considered for future evaluation as potential therapeutic targets.
Materials and Methods
We collected vestibular fluid from 4 controls and 4 patients with vulvodynia by placing a cotton swab in the vestibule and extracting the absorbed proteins. The proteins underwent 2-dimensional difference gel electrophoresis and mass spectrometry to develop a protein map. Immunohistochemistry was used to validate proteomic findings.
Results
A map was constructed of 32 of the more abundant proteins in vestibular fluid and their levels compared in control subjects and vulvodynia patients. Among these were annexin A1, interleukin 1 receptor antagonist, protein S100 A9, and a number of antiproteases and proteases. Many of these proteins differed by at least 50% between groups, but only annexin A1, one of the protease inhibitors, and immunoglobulin G κ chain were significantly different. The results with annexin A1 were validated by similar findings with immunohistochemistry.
Conclusions
The findings of this pilot study demonstrate a set of vestibule mucosa proteins that differ significantly—either increasing or decreasing—in vulvodynia patients compared with controls, and several others that exhibited greater than 1.5-fold change but did not reach statistical significance. This study constitutes a proof-of-principle that an open, unbiased proteomic approach can identify molecular participants in vulvodynia, some of which had not been identified to date by hypothesis-driven studies.
Key Words: protease, antiprotease, vulvodynia, inflammation, pain, allodynia, vestibulitis, gel-based proteomics
Allodynia, or pain out of proportion to a stimulus, in the presence of a normal appearing vulvar epithelium, is an important characteristic of vulvodynia.1 Several studies of this condition have demonstrated an increase in neurite outgrowth in the vulvar vestibule.2–4 Neuronal alterations demonstrated in patients with vulvodynia have been associated with the infiltration of mast cells and B lymphocytes and several other molecules. A wide range of etiologies have been proposed for the allodynia, including neurite outgrowth and associated cellular and molecular changes, as well as hormonal, psychosocial, and structural changes, but these proposed etiologies have not led to effective therapies that reduce allodynia.5–8
Our hypothesis for this pilot study was that vulvodynia was caused or perpetuated by specific protein molecules, so we used a discovery proteomics approach to explore this possibility. We focused on proteins present in the fluid covering the vestibule that might participate in the neuronal alterations of vulvodynia or be involved in other mechanisms. These proteins could be of epithelial origin, from the underlying tissue, derived from the microbiome, etc. We used an unbiased gel-based discovery proteomics approach as a hypothesis-generating tool to obtain novel insights into the pathophysiologic mechanism of allodynia at the most common site of pain, the vulvar vestibule. This approach targets the most abundant proteins in those samples and allows the direct comparison of multiple samples. The goals of this proteomics study are 3-fold: first, to develop a protein map of vestibular fluid from healthy controls, determining normal individual protein levels; second, to use a discovery proteomic method to identify proteins whose expression is altered in vulvodynia patients and potential pathways in which these proteins could be involved; and third, to validate these differences with immunohistochemistry. We analyzed proteins in the fluid overlying the vestibule using 2-dimensional difference gel electrophoresis (2D-DIGE) coupled with mass spectrometry (MS) to determine the identity of proteins in vulvodynia and control subjects.9–11
MATERIAL AND METHODS
Study Subjects
The characteristics of the subjects in the control and vulvodynia groups are listed in Table 1. Each group consisted of 4 subjects with similar ages. All subjects were having regular menstrual cycles, and samples were obtained at the midluteal phase. All had negative Gram stains and had not had fungal infections for at least 6 months. The vulvodynia subjects were patients at the Penn State Vulvodynia/Vulvovaginitis Clinic and exhibited both spontaneous pain and provoked pain from a cotton swab test. Control subjects did not report spontaneous pain and cotton swab tests were negative. The exclusion criteria for both groups in the study included current treatment for vulvovaginal pain, a prior vestibulectomy, or having dermatologic or other medical conditions that could potentially contribute to vulvovaginal pain.
TABLE 1.
Study Subject Characteristics/Inclusion Criteria
| Characteristic/group | Control | Vulvodynia |
|---|---|---|
| n | 4 | 4 |
| Age mean (range), y | 32.5 (21–48) | 31.5 (25–42) |
| Menstrual status | Regular cycles | Regular cycles |
| Cycle phase at sampling | Midluteal | Midluteal |
| Fungal infection (last 6 mo) | Negative | Negative |
| Gram stain | Negative | Negative |
| Spontaneous pain | no | yes |
| Provoked pain (cotton swab test) | no | yes |
The characteristics of the study population are listed, along with inclusion and exclusion criteria.
Exclusion criteria: current treatment for vulvar/vaginal pain, prior vestibulectomy, and potential contributing dermatologic/medical conditions.
Vulvodynia patients for immunohistochemical studies were identified by a record search of the archives of the pathology department for those who previously underwent vestibulectomy and did not have other dermatologic or inflammatory disorders. They were subject to the same inclusion criteria described previously. Control subjects were recruited from a group of healthy reproductive age women consenting to participate as controls in a related study.12 Tissue from control subjects were obtained by biopsy after 1% lidocaine analgesia using a 3-mm Keys punch biopsy tool from the posterior half of the vulvar vestibule. Samples were placed in 10% buffered formalin and embedded in paraffin.
This study and the procedures for obtaining archival tissue and control biopsy tissue were approved by the Penn State College of Medicine Institutional Review Board.
Sample Collection for Proteomic Studies
All cotton swab samples for proteomic studies were collected during the luteal phase of the menstrual cycle. Vestibular mucosal fluid was collected by placing a cotton swab in the posterior vestibule at the 6 o’clock position immediately external to the hymenal ring for 1 minute. Swabs were stored at −80°C until ready for extraction.
Two-Dimensional Difference Gel Electrophoresis and Densitometry
The procedure to perform 2D-DIGE and the analysis of resulting gels has been described in detail previously.11 Adaptations of these procedures for the present study are described subsequently.
Fluid samples from the vestibule were extracted from the swabs with PBS, 1-mM EDTA, and collected by centrifugation in a recovery vessel. A series of 3 extractions were done for each swab and proteins were precipitated from the extracts using trichloroacetic acid/acetone. The precipitated protein was resuspended in cell lysis buffer (30-mM Tris-HCl [pH 8.5], 2-M thiourea, 7-M urea, 4% [wt/vol] CHAPS). The protein concentration of each extract was determined using the Bio-Rad Protein Assay (Bio-Rad Laboratories, Hercules, CA) and adjusted to 1 mg/mL with cell lysis buffer for labeling. Two-dimensional DIGE gels were prepared and run as described previously11 and imaged using a Typhoon 9400 fluorescent imager. Images were imported into Progenesis SameSpots v2.0 for analysis. Additional information about the acquisition and processing of 2D-DIGE data is provided in our published work.11
Gel images were analyzed by densitometry to determine whether there are differences in the amounts of specific proteins between groups. All gels included a reference sample that allowed us to normalize the values of all spots in the gels of each sample so that the samples could be compared with one another. The resulting quantitation of each spot is referred to as its normalized volume and is a quantitative measure of that protein spot. There were many protein spots that have been confirmed by MS (see hereinafter) to represent the same protein because of differences in isoform or proteoform patterns and could reflect both changes in protein sequence or posttranslational modification or processing.13 The normalized volumes for these spots were added together and the resulting sums used to compare values obtained in vulvodynia samples to those found in controls. Preliminary studies showed that vestibular fluid contains only small amounts of the most abundant plasma proteins, so depletion of these proteins was not needed, as was the case with plasma samples.
Mass Spectrometry
Mass spectrometry was performed to identify the protein making up each spot on the gel as described in our previous publication.11 All 658 gel spots were picked by manual excision and processed for analysis by MALDI-ToF/ToF MS (5800 Proteomic Analyzer Applied Biosystems, Foster City, CA) in the MS Core Facility at the Penn State University College of Medicine. The MS and MS/MS data were submitted to the MASCOT search engine using the National Center for Biotechnology Information nonredundant database and human taxonomy for identification.
Immunohistochemistry
Immunohistochemistry was performed on multiple samples from vulvodynia and control subjects to determine tissue localization within the vestibule of selected proteins that distinguished vulvodynia patients from controls and as a validation for proteomic results. Paraffin-embedded tissues from vulvodynia patients who had undergone vestibulectomy and control subjects from a previous study were sectioned (4 μm), deparaffinized, rehydrated, and processed by low-temperature antigen retrieval as previously described.14 Sections were blocked with normal horse serum and immunoreacted overnight at room temperature with the primary antibody for annexin A1 (Abcam), S100 A9 (Abcam), and glial fibrillary acidic protein (Abcam) at dilutions from 1:500 to 1:2000. The signals were amplified with Vector Immpress-AP horse antirabbit IgG Polymer Detection Kit and Vector Red used as the chromogen. Control sections were incubated with only the secondary antibody. Images were recorded using a Nikon DXM 1200 digital camera under Nomarski optics and using Nikon ACT1, version 2, software.
Statistical Analysis
Multivariate analysis of data from 2D-DIGE protein spots was performed using Progenesis Stats (Nonlinear) for the t tests on individual spots and for principal components analysis of the entire vulvodynia versus control data set. As shown in Figure 1, many of the identified proteins consisted of multiple gel spots. In these cases, the normalized volumes were added together for “whole protein” analysis.
FIGURE 1.
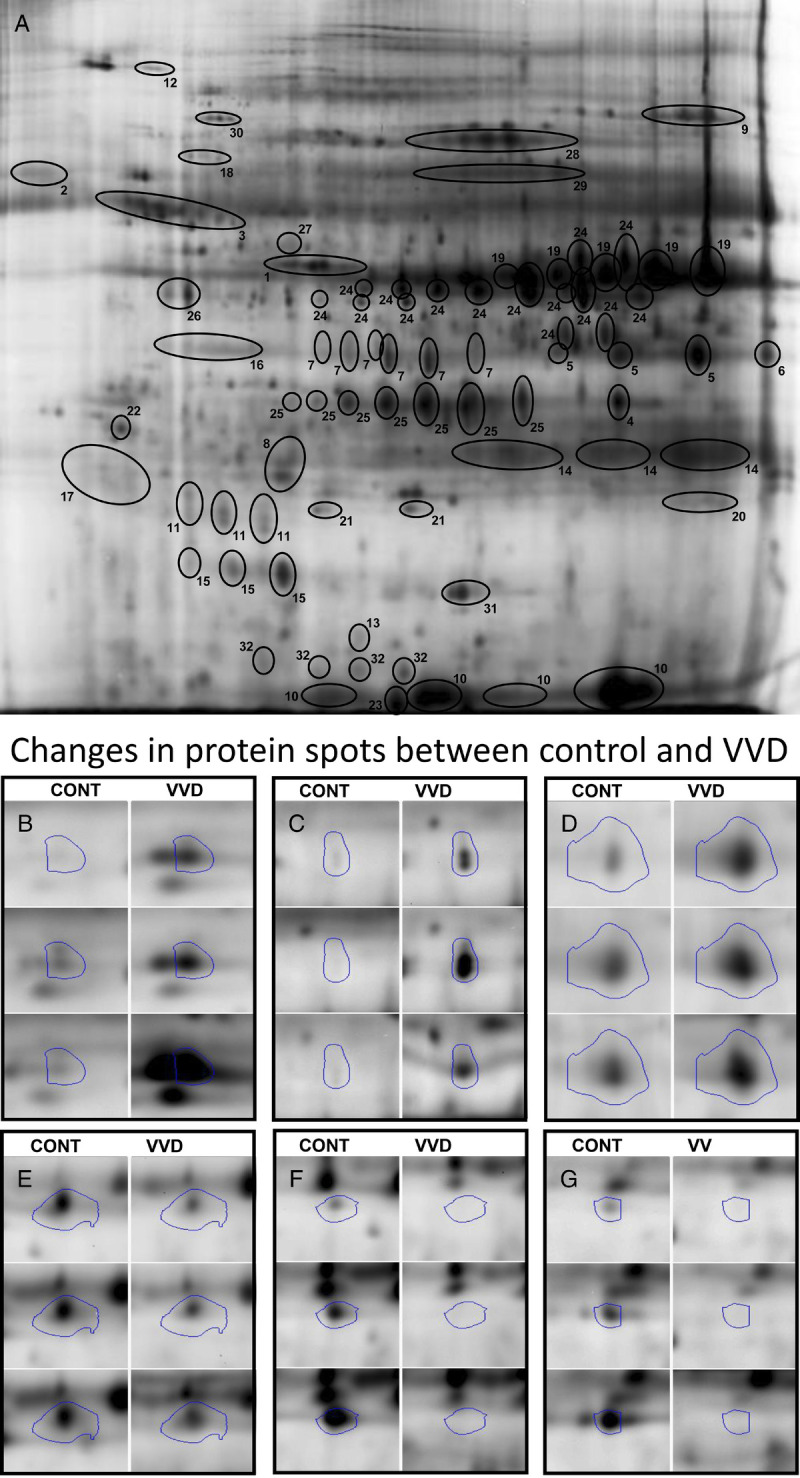
A, A spot map of a reference gel image of vestibular swab extract proteins identified by 2D-DIGE. Panel A depicts the identified proteins. Some of the proteins may include multiple spots reflecting different proteoforms/isoforms, fragments, or multimers. 1, Actin, cytoplasmic 2; 2, α-1-antichymotrypsin precursor; 3, α-1-antitrypsin precursor; 4, α-2-macroglobulin–like protein 1 precursor; 5, annexin A1; 6, annexin A2 isoform 2; 7, annexin A3; 8, cathepsin B preproprotein; 9, complement C3 precursor; 10, fatty acid–binding protein, epidermal; 11, γ-glutamylcyclotransferase; 12, glial fibrillary acidic protein; 13, hyaluronan mediated motility receptor isoform a; 14, IgG κ chain; 15, interleukin 1 receptor antagonist protein; 16, keratin 10; 17, keratin, type I cytoskeletal 13; 18, keratin, type I cytoskeletal 9; 19, leukocyte elastase inhibitor (serpin B1); 20, neutrophil gelatinase–associated lipocalin precursor; 21, peroxiredoxin 2; 22, proteasome subunit α type-5 isoform 1; 23, protein S100-A9; 24, serpin B3 (squamous cell carcinoma antigen 1); 25, serpin B4 (squamous cell carcinoma antigen 2); 26, serpin B6; 27, serpin peptidase inhibitor, clade A (α-1 antiproteinase, antitrypsin), member 1; 28, serum albumin; 29, serum albumin preproprotein; 30, Sp|P0A6M8|EFG_ECOLI elongation factor G (EF-G, Escherichia coli); 31, superoxide dismutase (Cu-Zn); 32, transthyretin precursor. Panels B to D depict examples of protein spots that are increased in vulvodynia (VVD) patients compared with control subjects (CONT). Panels E and F depict spots that are decreased in VVD patients.
Role of the Funding Sources
This research was supported by a grant from the National Vulvodynia Association (J.F.) and a kind gift from the Kenneth and Susan Stoudt (C.M.). The funding sources did not have a role in the study design, collection, analysis, interpretation, writing of the manuscript, or in the decision to submit the manuscript for publication.
RESULTS
Study Subject Characteristics
All 4 vulvodynia patients in proteomic studies reported unprovoked pain in the vulvovaginal area for 3 to 5 years and demonstrated, by cotton swab test over at least 3 months of evaluation by author CM, pain in response to touch in the vulvar vestibule. The appearance of the vulva was normal. No patients were receiving medications for vulvodynia, and none had undergone vestibulectomy in the past.
The 4 control subjects were free of vulvar and vaginal pain based on review of symptoms and cotton swab testing for pain.
All vulvodynia patients for tissue sampling had undergone vestibulectomy as a treatment for pain and that archival tissue was used for the immunohistochemical analysis. Pathology findings reported for vulvodynia subjects did not reveal other dermatologic or inflammatory disorders. Control subjects for immunohistochemical studies were free of vulvar and vaginal pain, and cotton swab testing for pain was negative.
Protein Differences Between Vulvodynia Patients and Control Samples
Gel electrophoresis and densitometry
Two-dimensional DIGE gels revealed a total of 658 protein spots that were quantified by densitometry. Of these, 55 protein spots (8.4% of total) differed significantly between vulvodynia (VVD) patients and controls (CONT). Of the 55 protein spots, 23 (3.5%) were increased in vulvodynia patients and 32 (4.9%) were decreased in vulvodynia patients, indicating that the changes between groups were not due to a generalized increase or decrease. Figure 1 depicts a representative normalization gel in panel A. Protein spots that were identified are circled and numbered with the protein names listed in the legend for Figure 1. Panels B, C, and D show representative comparisons of three of the spots that increased significantly in vulvodynia patients versus controls, and panels E, F, and G show three of the spots that were significantly decreased in vulvodynia patients. A principal component analysis of all spots on the gels is shown in Figure 2. Note that the cluster of vulvodynia patients is distinct from the control subjects.
FIGURE 2.
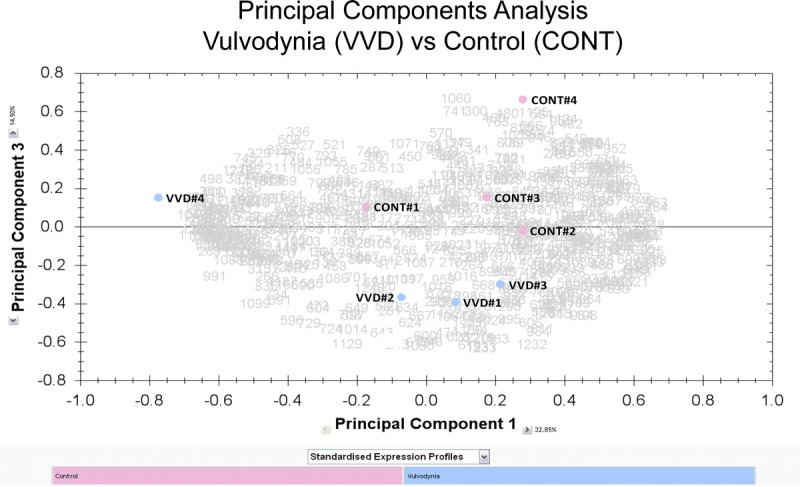
Principal component analysis. Values for the normalized volumes for all of the 658 spots in the 2D-DIGE gels were used to perform a principal component analysis of the gel data. The samples are designated CONT #1 to #4 and VVD #1 to #4.
Mass spectrometry
The protein spots were excised from a preparative gel for MS analysis. Of these, MS MALDI-ToF/ToF identified, with MASCOT CI scores greater than 95%, a peptide from 140 excised spots, or 21% of the total excised spots. These spots constituted more than 67% of the densitometrically detected protein on the gels. A number of spots were found to identify the same peptide, indicating multiple proteoforms/isoforms of the whole protein.13 When these spots were considered together, 32 whole proteins were identified. We combined the normalized spot volumes from spots containing the same whole protein and performed statistical analysis using the total normalized spot volume of each whole protein, reported in Table 2, listing the protein names, accession numbers, means, and SDs for each group of 4. Thirteen of the proteins that differed by more than 50% between control and vulvodynia subjects are highlighted in the fifth column of Table 2 and represent a variety of functions. Proteins that were 50% greater in vulvodynia vs control (>1.5) are described as increased and shown in bold, underlined blue type (i.e., annexin A1). Those that were 50% lower in vulvodynia vs control (<0.67) are described as decreased and shown in bold, underlined red type (i.e., α-1-antitrypsin precursor). Levels of all proteins were compared by t test and the resulting p values are listed. Three proteins were found to differ significantly between groups and are shown in bold type and marked with an asterisk. Annexin A1 was significantly increased and immunoglobulin G (IgG) κ chain and serpin peptidase inhibitor, clade A (α-1 antiproteinase), member 1, were significantly decreased in vulvodynia patients.
TABLE 2.
Vestibular Swab Extract Proteins Identified by 2D-DIGE With Values and Note of Significant Changes
| Protein name | Accession number | Control | Vulvodynia | Fold difference VVD/con | p |
|---|---|---|---|---|---|
| Actin, cytoplasmic 2 | gi|4501887| | 4.06 (1.552) | 9.7 (11.521) | 2.11 | .37 |
| α-1-Antichymotrypsin precursor | gi|50659080| | 3.88 (2.021) | 3.36 (0.906) | 0.87 | .655 |
| α-1-Antitrypsin precursor | gi|50363217| | 13.66 (2.354) | 8.34 (4.31) | 0.61 | .074 |
| α-2-Macroglobulin–like protein 1 precursor | gi|74271845| | 1.07 (0.884) | 1.07 (0.706) | 1 | 1 |
| Annexin A1 | gi|4502101| | 1.93 (0.523)* | 3.64 (1.22)* | 1.89 | .042* |
| Annexin A2 isoform 2 | gi|4757756| | 0.95 (0.164) | 1.12 (0.264) | 1.18 | .299 |
| Annexin A3 | gi|4826643| | 5.37 (0.978) | 6.75 (2.162) | 1.26 | .288 |
| Cathepsin B preproprotein | gi|4503139| | 2.44 (0.473) | 2.4 (1.172) | 0.98 | .949 |
| Complement C3 precursor | gi|115298678| | 3.11 (2.203) | 4.5 (2.223) | 1.45 | .409 |
| Fatty acid–-binding protein, epidermal | gi|4557581| | 7.89 (3.054) | 6.11 (1.48) | 0.77 | .334 |
| γ-Glutamylcyclotransferase | gi|13129018| | 6.97 (2.956) | 5.39 (0.856) | 0.77 | .345 |
| Glial fibrillary acidic protein | gi|4503979| | 0.79 (0.173) | 3.87 (5.498) | 4.9 | .305 |
| Hyaluronan-mediated motility receptor isoform a | gi|217272802| | 0.96 (0.18) | 1.12 (0.686) | 1.17 | .669 |
| IgG κ chain | gi|4176418 | 10.1 (1.857)* | 6.71 (1.697)* | 0.66 | .036* |
| Interleukin 1 receptor antagonist protein | gi|10835147| | 2.94 (1.06) | 3.42 (1.757) | 1.16 | .659 |
| Keratin 10 | gi|269849769| | 3.2(0.563) | 4.19 (1.042) | 1.31 | .147 |
| Keratin, type I cytoskeletal 13 | gi|131412228| | 13.91 (7.745) | 6.93 (2.543) | 0.5 | .138 |
| Keratin, type I cytoskeletal 9 | gi|55956899| | 4.2 (1.195) | 3.81 (3.134) | 0.91 | .823 |
| Leukocyte elastase inhibitor (serpin B1) | gi|13489087| | 5.19 (1.086) | 5.13 (0.688) | 0.99 | .921 |
| Neutrophil gelatinase–associated lipocalin precursor | gi|38455402| | 3.26 (1.044) | 2.76 (0.731) | 0.85 | .458 |
| Peroxiredoxin 2 | gi|32189392| | 1.44 (0.399) | 2.48 (1.078) | 1.72 | .119 |
| Proteasome subunit α type-5 isoform 1 | gi|23110942| | 0.42 (0.185) | 1.59 (2.052) | 3.79 | .299 |
| Protein S100-A9 | gi|4506773| | 0.65 (0.237) | 1.4 (0.84) | 2.15 | .139 |
| Serpin B3 (squamous cell carcinoma antigen 1) | gi|5902072| | 20.82 (8.024) | 11.47 (5.219) | 0.55 | .099 |
| Serpin B4 (squamous cell carcinoma antigen 2) | gi|28076869| | 7.31 (3.385) | 7.69 (3.308) | 1.05 | .879 |
| Serpin B6 | gi|41152086| | 2.39 (0.972) | 2.03 (0.868) | 0.85 | .605 |
| Serpin peptidase inhibitor, clade A (α-1 antiproteinase), member 1 | gi|15990507 | 1.68 (0.498)* | 0.94 (0.296)* | 0.56 | .044* |
| Serum albumin | gi|28592 | 6.9 (5.38) | 9.66 (6.132) | 1.4 | .525 |
| Serum albumin preproprotein | gi|4502027| | 7.19 (4.859) | 8.7 (5.802) | 1.21 | .705 |
| ECOLI elongation factor G (EF-G, E. coli) | gi|62288080| | 1.34 (0.687) | 5.17 (5.624) | 3.86 | .225 |
| Superoxide dismutase (Cu-Zn) | gi|4507149| | 2.39 (0.798) | 3.71 (1.307) | 1.55 | .134 |
| Transthyretin precursor | gi|4507725| | 4.91 (0.856) | 3.41 (1.139) | 0.69 | .08 |
List of all proteins identified by 2D-DIGE and mass spec and their accession numbers. Values are means (SD) of the normalized volumes of control (Con) or vulvodynia (VVD) subjects. For proteins with multiple isoforms/proteoforms, the normalized volumes for all spots were added together. The fold difference (VVD/Con) is given in the fifth column. Proteins with a 50% increase in vulvodynia vs control (>1.5) are in bold blue underlined type. Values with a 50% decrease in vulvodynia (<0.67) are in bold red underlined type. The right-hand column gives the p values resulting from a t test of control versus vulvodynia values (n = 4/group) for each protein. Significant differences (p < .05) are in bold type and marked with an asterisk.
Immunohistochemistry
Having identified a set of proteins in the fluid layer coating the vestibule mucosa that differed between vulvodynia patients and controls, we used immunohistochemistry to attempt to validate the proteomics findings for several of the proteins. Multiple samples from vulvodynia patients and control subjects were subjected to immunostaining. In representative samples, we show that annexin A1 is located within the vestibular mucosa and that its expression pattern differs between VVD and control. Annexin A1 immunoreactivity is present in control subjects in a uniform distribution in the superficial layer of the vestibular mucosa (see Figure 3A; arrowheads; ▼). In vulvodynia subjects, annexin immunoreactivity is more intense and the staining pattern is variable (see Figure 3B). The stratum basalis (arrows; ↑) displays an equally intense level of immunostaining to that of the superficial layer. Staining is also apparent in the lamina propria of the vulvodynia samples.
FIGURE 3.
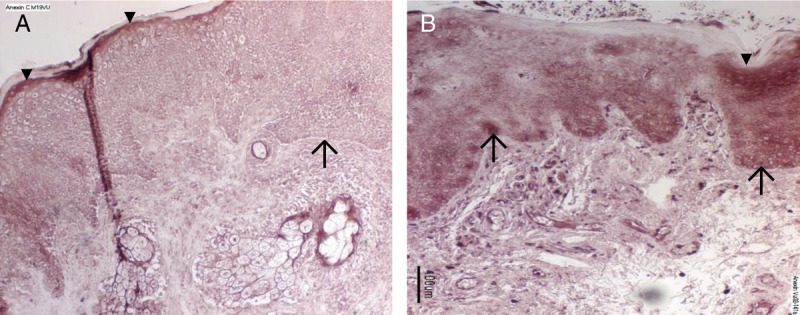
Immunostaining for annexin A1. Tissue samples from a representative control (panel A) and vulvodynia subject (panel B) were immunoreacted with anti-annexin A1 and detected with Vector Red. There is absent immunostaining in slides that were reacted with only the secondary antibody as controls (data not shown). Annexin A1 immunoreactivity (dark staining) is present in control subjects in a uniform distribution in the superficial layer of the epithelium (arrowheads; ▼) of the vestibular mucosa (panel A). In vulvodynia subjects compared with controls, annexin immunoreactivity in the superficial layer is more intense, but the staining pattern is more variable (panel 3B). In tissue from vulvodynia patients (panel 3B), the stratum basalis (arrows; ↑) displays an equally intense level of immunostaining to that of the superficial layer. However, staining is barely detectable in the stratum basalis of the control samples (panel 3A). In the vulvodynia samples, below the epithelium, there were also intensely stained lymphocytes present within blood vessels and scattered throughout the lamina propria.
We also stained tissue samples for S100 A9 and glial fibrillary acidic protein, but despite the differences seen with proteomics, we could not distinguish any differences between control and vulvodynia samples (data not shown).
DISCUSSION
Using an unbiased discovery proteomics approach, we conducted a pilot study to investigate changes in proteins from the surface of the vulvar vestibule of vulvodynia patients and control subjects as a way of gaining insight into molecules potentially involved in the pathogenesis of vulvodynia. We recovered enough of the protein-containing fluid samples for a gel-based proteomic analysis revealing 658 protein spots. Using MS, we were able to identify 32 proteins consisting of one to several spots. It is noteworthy that 9 of these proteins were either proteases or antiproteases. The findings presented offer new clues to vulvodynia's pathophysiology, provide a basis for testable hypotheses, and attest to the possibility of obtaining material for such studies from a swab applied to the surface of the vestibule.
We describe a set of vestibular mucosal proteins that are significantly different in vulvodynia patients compared with control subjects, and a number of others that differed by at least 50% but did not reach statistical significance. Several of the identified proteins have previously been suspected to have a role in vulvodynia. These include annexin A1,15 interleukin 1 receptor antagonist,16 protein S100-A9,17 and a number of protease inhibitors.18 Findings of increased nerve endings in vulvodynia4 could be reflected by increases in glial fibrillary acidic protein observed in the present study.
Annexin A1 and S100-A9
The findings of significantly increased annexin A1, a phospholipid binding protein active in membrane trafficking and considered to have anti-inflammatory activity, and elevated levels of S100-A9, a calcium binding protein with chemotactic activity, are interesting in light of recent reports implicating these proteins in other chronic mucosal conditions, particularly recurrent vulvovaginal candidiasis. Fidel et al15,17 have postulated that annexin A1 and S100-A9, produced by epithelial cells, are causally implicated in worsening vulvovaginal symptoms in the face of relatively low numbers of candida. Annexin A1 has been identified in mast cell granules, an important observation because mast cell degranulation has been implicated in vulvodynia.
Antiproteases
The finding that serpin peptidase inhibitor, Clade A (α-1 antiproteinase), member 1, was significantly lower in vulvodynia versus control indicates that a shift in vulvodynia in serine protease/antiprotease balance in favor of proteolysis may occur. This interpretation is supported by the finding by Jayaram et al18 of decreases in enzymatic activity in 2 additional serpins, secretory leukocyte protease inhibitor, and human epididymis protein 4, in fluid collected from a swab applied briefly to the posterior vaginal surface. This is an appealing interpretation because of its translational potential. Our findings add credence to this idea in that most of the 7 protease inhibitors we detected in this study were at lower levels, albeit not significantly, in vulvodynia patients. There is compelling evidence linking proteases to pain, a hallmark of vulvodynia, as well as to chronic inflammatory responses that are implicated in vulvodynia's pathophysiology.5,19
Notable among the links between physiological as well as pathological pain and inflammation are the 4 protease activated receptors (PARs). These are G-protein coupled transmembrane receptors expressed ubiquitously in a wide variety of cells, including nociceptive neurons, immune, and glial cells. They are activated by extracellular proteases and in their activated forms PARs serve as sensors and transducers of information to cells about proteases in the extracellular environment. For example, activation of PAR2 has been shown to potentiate pain mediated by the transient receptor potential vanilloid receptor 1, a member of the transient receptor potential cation channel subfamily,20 shown to be upregulated in vulvodynia neurites.21 Besides linking pain and inflammatory response, activation of PARs is linked to other homeostatic/allostatic functions relevant to vulvodynia (i.e., vascular permeability22–24 and synaptic plasticity).23 There is also evidence that activation of PAR2 by mast cell tryptase in vitro can compromise the barrier function of vaginal epithelial cells.25,26 Together, these observations have led to recognition of the critical contribution of the neuroimmune system via PARs to neuroinflammation-associated diseases and the impetus to develop agents to treat such diseases by blocking PARS.27 However, developing effective inhibitors specific to any of the many known functions is proving to be challenging because of the complexity of the PARS regulation.27–29 One major recent discovery has been the potential of PARs for biased signaling, namely, the ability of ligands acting at the same G protein–coupled receptor to stabilize multiple, distinct, receptor conformations linked to different functional outcomes. The prominent representation of proteases and antiproteases in vestibular fluid in the present study raises the possibility that alterations in protease balance could affect the regulation of PARS. The data indicate that there may be alterations in vestibular protease/antiprotease balance in vulvodynia. These findings point to a potential for mechanism-based therapeutic options that can be linked to allodynia pain.
Many of the protein comparisons failed to achieve statistical significance because of an extreme value in 1 of the 4 patients tested. However, the outlying value was not always in the same patient. This leads us to speculate that there may be subgroups of vulvodynia patients. Although we did stratify our patients into the subcategory of generalized unprovoked vulvodynia and collected all samples in the luteal phase, a more complex categorization (i.e., atopic history vs. infection history, oral contraceptives, etc.) will be necessary to determine whether there are additional subtypes and pathogenic processes within each subtype. This is a possibility that could be addressed by expanding the number of patients in future studies.
Strengths and Limitations
The strengths of our study lie in its approach. The open approach provided by 2D-DIGE, a hypothesis-generating method, allows the detection of proteins that were not suspected of having a role in vulvodynia pathogenesis, could be derived from different cellular sources, and therefore could not have been predicted in a typical hypothesis-based approach. In addition, 2D-DIGE, coupled with MS, allows the detection of proteins for which there is not an available antibody.9,10,30
One limitation of our study is that 2D-DIGE is most useful in the detection of more abundant proteins, so rarer protein species may not be identified. Another inherent limitation is that we are unable to determine contributions made by processes occurring in the underlying tissue. To assemble a more complete picture of processes involved in vulvodynia pathogenesis, one could study changes in tissue protein expression in parallel to mucosal fluid expression by performing parallel proteomic assays on vestibule tissue biopsies and surface fluid from the same vulvodynia patients. Finally, although we can detect changes in protein, we cannot determine whether the changes are a cause or a consequence of vulvodynia.
CONCLUSIONS
In conclusion, our proteomic study of vestibule fluid defined a set of proteins that undergo changes in expression in vulvodynia and based on the available literature these proteins may be involved, as discussed in several pathways. The fact that these proteins were obtained reproducibly with a simple cotton swab could facilitate sample procurement and enable some of the future studies noted previously. The present findings lay a foundation toward a better understanding of vulvodynia pathophysiologic processes and the development of mechanism-based therapies.
Footnotes
The authors have declared they have no conflicts of interest.
This research was supported by a grant from the National Vulvodynia Association (J.F.) and a kind gift from Kenneth and Susan Stoudt (C.M.).
This study was reviewed and approved by the Institutional Review Board of the Penn State University College of Medicine, Hershey, PA.
J.W. is deceased.
A preliminary report of this study was presented at the XXI World Congress of the International Society for the Study of Vulvovaginal Disease (ISSVD), Paris, France, October 13, 2011.
Contributor Information
Todd Umstead, Email: tmu1@psu.edu.
Debra Shearer, Email: dshearer@pennstatehealth.psu.edu.
Judith Weisz, Email: jxw7@psu.edu.
David S. Phelps, Email: dsp4@psu.edu.
Joanna Floros, Email: jfloros@psu.edu.
REFERENCES
- 1.Bachmann GA Rosen R Pinn VW, et al. Vulvodynia: a state-of-the-art consensus on definitions, diagnosis and management. J Reprod Med 2006;51:447–56. [PubMed] [Google Scholar]
- 2.Halperin R Zehavi S Vaknin Z, et al. The major histopathologic characteristics in the vulvar vestibulitis syndrome. Gynecol Obstet Invest 2005;59:75–9. [DOI] [PubMed] [Google Scholar]
- 3.Tommola P Unkila-Kallio L Paetau A, et al. Immune activation enhances epithelial nerve growth in provoked vestibulodynia. Am J Obstet Gynecol 2016;215:768.e1–8. [DOI] [PubMed] [Google Scholar]
- 4.Bornstein J, Goldschmid N, Sabo E. Hyperinnervation and mast cell activation may be used as histopathologic diagnostic criteria for vulvar vestibulitis. Gynecol Obstet Invest 2004;58:171–8. [DOI] [PubMed] [Google Scholar]
- 5.Falsetta ML Foster DC Bonham AD, et al. A review of the available clinical therapies for vulvodynia management and new data implicating proinflammatory mediators in pain elicitation. BJOG 2017;124:210–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lynch PJ. Vulvodynia as a somatoform disorder. J Reprod Med 2008;53:390–6. [PubMed] [Google Scholar]
- 7.Khandker M Brady SS Stewart EG, et al. Is chronic stress during childhood associated with adult-onset vulvodynia? J Womens Health (Larchmt) 2014;23:649–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Reissing ED Brown C Lord MJ, et al. Pelvic floor muscle functioning in women with vulvar vestibulitis syndrome. J Psychosom Obstet Gynaecol 2005;26:107–13. [DOI] [PubMed] [Google Scholar]
- 9.Phelps DS Umstead TM Silveyra P, et al. Differences in the alveolar macrophage proteome in transgenic mice expressing human SP-A1 and SP-A2. J Proteom Genom Res 2013;1:2–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Phelps DS, Umstead TM, Floros J. Sex differences in the response of the alveolar macrophage proteome to treatment with exogenous surfactant protein-A. Proteome Sci 2012;10:44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Phelps DS Umstead TM Quintero OA, et al. In vivo rescue of alveolar macrophages from SP-A knockout mice with exogenous SP-A nearly restores a wild type intracellular proteome; actin involvement. Proteome Sci 2011;9:67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.MacNeill C de Guzman G Sousa GE, et al. Cyclic changes in the level of the innate immune molecule, surfactant protein-a, and cytokines in vaginal fluid. Am J Reprod Immunol 2012;68:244–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Smith LM, Kelleher NL. Proteoform: a single term describing protein complexity. Nat Methods 2013;10:186–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.MacNeill C Umstead TM Phelps DS, et al. Surfactant protein A, an innate immune factor, is expressed in the vaginal mucosa and is present in vaginal lavage fluid. Immunology 2004;111:91–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lilly EA, Yano J, Fidel PL, Jr. Annexin-A1 identified as the oral epithelial cell anti-Candida effector moiety. Mol Oral Microbiol 2010;25:293–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Foster DC, Sazenski TM, Stodgell CJ. Impact of genetic variation in interleukin-1 receptor antagonist and melanocortin-1 receptor genes on vulvar vestibulitis syndrome. J Reprod Med 2004;49:503–9. [PubMed] [Google Scholar]
- 17.Yano J Palmer GE Eberle KE, et al. Vaginal epithelial cell-derived S100 alarmins induced by Candida albicans via pattern recognition receptor interactions are sufficient but not necessary for the acute neutrophil response during experimental vaginal candidiasis. Infect Immun 2014;82:783–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jayaram A Esbrand F Dulaveris G, et al. Decreased concentration of protease inhibitors: possible contributors to allodynia and hyperalgesia in women with vestibulodynia. Am J Obstet Gynecol 2015;212:184.e1–4. [DOI] [PubMed] [Google Scholar]
- 19.Tommola P Butzow R Unkila-Kallio L, et al. Activation of vestibule-associated lymphoid tissue in localized provoked vulvodynia. Am J Obstet Gynecol 2015;212:476.e1–8. [DOI] [PubMed] [Google Scholar]
- 20.Vellani V Kinsey AM Prandini M, et al. Protease activated receptors 1 and 4 sensitize TRPV1 in nociceptive neurones. Mol Pain 2010;6:61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tympanidis P Casula MA Yiangou Y, et al. Increased vanilloid receptor VR1 innervation in vulvodynia. Eur J Pain 2004;8:129–33. [DOI] [PubMed] [Google Scholar]
- 22.De Campo BA, Henry PJ. Protease-activated receptors (PARs) are partly pro-inflammatory and partly anti-inflammatory: will PAR agonists or antagonists participate in future drug therapies? Curr Drug Targets 2006;7:629–37. [DOI] [PubMed] [Google Scholar]
- 23.Almonte AG, Sweatt JD. Serine proteases, serine protease inhibitors, and protease-activated receptors: roles in synaptic function and behavior. Brain Res 2011;1407:107–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ramachandran R Noorbakhsh F Defea K, et al. Targeting proteinase-activated receptors: therapeutic potential and challenges. Nat Rev Drug Discov 2012;11:69–86. [DOI] [PubMed] [Google Scholar]
- 25.Li Y Feng C Wei X, et al. Activation of protease-activated receptor-2 disrupts vaginal epithelial barrier function. Cell Biol Int 2014;38:1247–51. [DOI] [PubMed] [Google Scholar]
- 26.Zhang R, Ge J. Proteinase-activated receptor-2 modulates Ve-cadherin expression to affect human vascular endothelial barrier function. J Cell Biochem 2017;118:4587–93. [DOI] [PubMed] [Google Scholar]
- 27.Soualmia F, El Amri C. Serine protease inhibitors to treat inflammation: a patent review (2011–2016). Expert Opin Ther Pat 2018;28:93–110. [DOI] [PubMed] [Google Scholar]
- 28.Ramachandran R Altier C Oikonomopoulou K, et al. Proteinases, their extracellular targets, and inflammatory signaling. Pharmacol Rev 2016;68:1110–42. [DOI] [PubMed] [Google Scholar]
- 29.Hamilton JR, Trejo J. Challenges and opportunities in protease-activated receptor drug development. Annu Rev Pharmacol Toxicol 2017;57:349–73. [DOI] [PubMed] [Google Scholar]
- 30.Zegels G Van Raemdonck GA Tjalma WA, et al. Use of cervicovaginal fluid for the identification of biomarkers for pathologies of the female genital tract. Proteome Sci 2010;8:63. [DOI] [PMC free article] [PubMed] [Google Scholar]