

Abstract
Jiedu-Yizhi formula (JDYZF) is prescribed for the treatment of Alzheimer's disease (AD) and was created by Jixue Ren, a master of traditional Chinese medicine, based on the “marrow deficiency and toxin damage” theory. In our clinic, this formula has been used for the treatment of AD for many years and has achieved good results. However, the mechanism by which JDYZF improves cognitive impairment has not been determined. In this study, we confirmed that orally administered JDYZF reversed the cognitive deficits in an Aβ25–35-induced rat model, increased the number of neurons in the hippocampal CA1 area, improved their structure, decreased the deposition of β-amyloid (Aβ), reduced the expression of proteins related to the NLRP3/Caspase-1/GSDMD and LPS/Caspase-11/GSDMD pyroptosis pathways, and reduced the levels of interleukin 1β (IL-1β) and IL-18, thereby inhibiting the inflammatory response. In addition, JDYZF exerted no hepatotoxicity in rats. In short, these results provide scientific support for the clinical use of JDYZF to improve the cognitive function of patients with AD.
1. Introduction
Alzheimer's disease (AD) is characterized by the degeneration of the central nervous system. AD is the most common cause of dementia, and its symptoms include memory loss, personality changes, and social and emotional problems [1]. At present, approximately 50 million patients are suffering from AD worldwide. This number continues to rapidly increase and is expected to reach 152 million by 2050 [2]. Owing to its hidden onset, difficult diagnosis, and lack of a cure, AD imposes substantial economic and medical burdens on individuals and society [3, 4].
Many hypotheses have been proposed regarding the aetiological and pathological mechanisms of AD, but no exact theory has been confirmed. At present, drugs such as donepezil and memantine improve the symptoms of AD for only a short time. However, these agents cannot reverse or delay the progression of AD and have some side effects and toxicity [5]. New drugs that target the typical pathological characteristics of AD—senile plaques (SPs) formed by β-amyloid (Aβ) and neurofibrillary tangles (NFTs) caused by hyperphosphorylation of the tau protein—have achieved favourable results in only animals and not in clinical experiments [6, 7]. Therefore, under the guidance of alternative and integrated medicine, it is particularly important to select known effective methods for the treatment of AD, such as acupuncture, electroacupuncture, music therapy, aromatherapy, pulsed electromagnetic fields, and traditional Chinese medicines, to improve the clinical symptoms to the greatest extent and explore the mechanisms of these treatment methods [8–13]. Among them, traditional Chinese medicine formulas with multitarget therapeutic effects have been widely reported to improve the cognitive function of AD patients and have gradually attracted the attention of researchers [14].
Jiedu-Yizhi formula (JDYZF) is a traditional Chinese medicine compound that tonifies the kidney and marrow, resolves phlegm, and activates blood circulation and detoxification processes that were developed by Ren Jixue, a master of Chinese medicine. Our clinical department has used this formula for many years, and this treatment has a significant effect on AD. In a previous study on the mechanism of JDYZF, we found that the extract of Coptis chinensis polysaccharide, the main drug component of JDYZF, improved the lifespan of the transgenic Caenorhabditis elegans CL4176 model of AD, reduced the paralytic rate, inhibited nematode head Aβ deposition, increased heat shock protein hsp16.2 and hsp16.41 gene expression levels [15], reduced the Aβ25–35-induced expression of cleaved-caspase-3 and Bax in PC12 cells, increased the expression of Bcl-2 and inhibited apoptosis [16]. However, the mechanism of JDYZF in the treatment of AD remains unclear.
A strong inflammatory reaction occurs in the AD brain, and Aβ can activate microglia to trigger neuroinflammation; thus, neuroinflammation may play an important role in the pathogenesis of AD [17]. In recent years, pyroptosis has been shown to potentially promote neuroinflammation. Pyroptosis is a highly inflammatory programmed cell death mechanism mediated by cysteinyl aspartate-specific proteinases (caspases). Its characteristic manifestations are swelling of the cell body, rupture of the cell membrane, and release of proinflammatory factors to trigger an inflammatory cascade reaction [18, 19]. Pyroptosis is achieved through the assembly of inflammasomes, which are pattern recognition receptors such as NOD-like receptors (NLRs, including NLRP1 and NLRP3) that recognize pathogen-associated molecular patterns (PAMPs) and endogenous damage-associated molecular patterns (DAMPs) that recruit the adaptor protein apoptosis-associated speck-like protein containing a CARD (ASC). This adaptor is further connected to the downstream receptor caspase-1. The aggregation of caspase-1 leads to its autocleavage and to the production of its active form, which can induce the maturation of proinflammatory cytokines such as interleukin 1β (IL-1β) and IL-18. Concomitantly, gasdermin D (GSDMD) is cleaved to produce an N-terminal fragment (GSDMD-NT) that can initiate pyroptosis. GSDMD-NT can perforate the cell membrane, leading to cell death and to the outflow of proinflammatory factors [20–23]. Among the NLR family members, NLRP3 is the most well studied. NLRP3 is expressed by neuronal cells and microglia and can be activated by Aβ- and Aβ-induced oxidative stress products such as reactive oxygen species (ROS) and other substances to initiate pyroptosis [24, 25]. In addition, lipopolysaccharides (LPSs) in the cytosol can directly induce the activation of caspase-4/caspase-5 in humans or caspase-11 in rodents and the cleavage of GSDMD to cause nerve cell pyroptosis [26]. Therefore, altering pyroptosis and inhibiting neuritis could be breakthroughs in the treatment of AD. At the same time, inflammation is closely related to blood stasis [27]. Whether JDYZF, which ameliorates blood stasis, exerts a therapeutic effect by altering pyroptosis and inhibiting inflammatory reactions remains unknown.
In this study, we evaluated the effects of JDYZF on the cognitive and memory function of AD model rats. We also assessed the effects of JDYZF on the expression of Aβ and the NLRP3/caspase-1/GSDMD and LPS/caspase-11/GSDMD pyroptosis pathways to provide scientific support for its clinical application.
2. Materials and Methods
2.1. Animals
Adult male Sprague–Dawley rats (weight: 200–220 g) were purchased from Changchun Yisi Experimental Animal Technology Co., Ltd. (Changchun, China). According to the regulations of the experimental animal centre of the Changchun University of Chinese Medicine, the rats were allowed to drink freely and were housed in a feeding room at a temperature of 25 ± 3°C and a relative humidity of 55 ± 5% on a 12-hour light/dark cycle. The animal experimental procedure was approved by the Experimental Animal Ethics Committee of the Changchun University of Chinese Medicine (no. 2021207).
2.2. Preparation of the Aβ25-35 Oligomer
One milligram of Aβ25-35 dry powder (A4559, Sigma) was dissolved in 500 μL of 0.9% normal saline to make a 2 μg/μL solution; the solution was sonicated for 5 minutes using a bath sonicator and then incubated at 37°C for 7 days. The incubated Aβ25–35 became flocculent and was stored in a refrigerator at 4°C for later use [28, 29].
2.3. Preparation of the JDYZF Decoction
JDYZ is composed of seven Chinese medicines, Coptis, wine-treated rhubarb, Ligusticum wallichii, Pheretima, tortoise shell glue, Cornus officinalis, and Alpiniae oxyphyllae fructus. The traditional Chinese medicines were purchased from Hongjian Pharmacy (Changchun, China). These herbs were mixed at a ratio of 1:1:1:1:1:1:2, soaked in distilled water (in a volume 5 times the volume of the medicine) for 1 hour, boiled for 1 hour, and then boiled twice. The extraction solutions were combined and concentrated to 1.0 g/mL, placed in sterile containers, sealed, and stored at −20°C.
2.4. Animal Modelling and Treatment
SD rats were anaesthetized with sodium pentobarbital and fixed on a brain stereotactic instrument. Holes were drilled in the rat skull 3 mm below the anterior fontanelle and 2 mm on both sides of the midline. After detecting the dura mater with a microsyringe, the needle was lowered 2.6 mm into the hippocampal CA1 area. Five microlitres of the Aβ25-35 solution was injected within 5 minutes. After 15 minutes, the needle was slowly withdrawn over a period of 5 minutes. The wound was quickly closed with paraffin after the ejection of the needle [30]. Seven days after the injection, the rats were randomly divided into five groups. The rats were administered an equivalent dose by gavage based on the body surface areas of humans and rats. Eleven rats each in the low-dose group (JDYZ.L), middle-dose group (JDYZ.M), and high-dose group (JDYZ.H) received 3.6 g/kg, 7.2 g/kg, and 14.4 g/kg JDYZF decoction, respectively, by gavage. Meanwhile, 10 rats in the positive drug group (PG) were administered donepezil hydrochloride (0.9 mg/kg) by gavage. Furthermore, 10 rats in the model group (MG) and 9 rats in the control group (CG) were administered normal saline (1 mL/100 g) by gavage. All animals were treated once per day for 8 weeks.
2.5. Morris Water Maze Test
The Morris water maze test was performed after 8 weeks of drug intervention. The water maze (diameter × height: 150 cm × 50 cm) was equally divided into four quadrants. Different patterns were placed on the wall of each quadrant to serve as clues. The platform (diameter: 10 cm) was placed 1 cm below the water surface of the fixed quadrant. The water was dyed black with melanin to ensure that the rats could not see the platform. A total of four water entry points were placed in each quadrant. The water temperature was maintained between 23°C and 27°C. Each rat was placed at the entry point facing the pool wall and was allowed 60 s to find and climb the platform. The time it took to find and climb the platform was recorded as the escape latency. If the rat did not climb the platform within the specified time, the time was recorded as 60 s. Subsequently, the rats were allowed to stay on the platform for 15 s, with an interval of 5 minutes between each entry. The experiment was performed for 4 days, and the order of entry points differed each day. Four days later, the platform was removed, and the rats were placed in the quadrant opposite the platform for the probe trial. Overall, the following information was recorded: the time it took for them to swim in the platform quadrant within 60 s, and the time it took for them to cross the platform [31]. The EthoVision XT, Version 11.0 system (Noldus, Netherlands) was used for recording and analysis.
2.6. Haematoxylin and Eosin (HE) Staining
The anaesthetized rats were injected intraperitoneally with 2% pentobarbital sodium (45 mg/kg) 24 hours after the water maze test. The required organs were collected quickly and stored according to the regulations. The hippocampus and liver were embedded after dehydration, cut into 5-µm-thick sections, and stained with HE [32]. The pathological changes in the hippocampus and liver were observed under an optical microscope.
2.7. Western Blotting
The rat hippocampi were homogenized and placed in a radioimmunoprecipitation assay lysis buffer containing protease and phosphatase inhibitors and phenylmethylsulfonyl fluoride. The samples were lysed on ice for 30 minutes and centrifuged at 13,000 r/min for 10 minutes at 4°C, and the supernatant was aspirated. The bicinchoninic acid assay was performed to determine the protein concentration. The samples were then stored at −80°C until further use. Western blotting was performed using standard protocols [33]. The membranes were incubated overnight with primary antibodies, including anti-NLRP3 (1:500, Novus, USA), anti-GSDMD (1:1000, Abcam, UK), anti-caspase-1 (1:1000, Novus, USA), and anti-caspase-11 (1:200, Novus, USA), and then with the appropriate secondary antibodies. ImageJ software was used to measure the gray values of the target bands.
2.8. Immunohistochemistry
The hippocampal slices were prepared at a thickness of 4 μm and then incubated with 0.3% hydrogen peroxide (which was added dropwise) at room temperature for 10 minutes to block endogenous peroxidase activity. The slices were washed 3 times with phosphate-buffered saline (PBS) and blocked with 5% bovine serum albumin for 30 minutes. The slices were incubated overnight with Aβ (MOAB-2, recognizes unaggregated, oligomeric and fibrillar forms of beta amyloid 42 and unaggregated beta amyloid 40, 1:500, Novus, USA), NLRP3 (1:50, Novus, USA), and GSDMD (1:1000, Abcam, UK) antibodies. Subsequently, the slices were washed 3 times with PBS and incubated with biotin secondary antibodies for 20 minutes and then treated with 3,3′-diaminobenzidine for 3 to 5 minutes. After washing, they were counterstained with haematoxylin and mounted after dehydration [34]. The slices were observed using a Cytation 5 (BioTek, USA) image reader. ImageJ software was used to analyse the average optical density (AOD) values and positive cell counts.
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
Aβ1-42 (Elabscience, China), IL-1β (mlbio, China), and IL-18 (mlbio, China) ELISA kits were used to determine the levels of Aβ1-42 and the inflammatory factors IL-1β and IL-18 in the hippocampus, cortex, colon, and serum. A microplate reader was used to detect the absorbance of the sample at 450 nm.
2.10. Statistical Analysis
All data are presented as the mean ± standard deviation. SPSS 20 software was used for data analysis. One-way analysis of variance (one-way ANOVA) was used to compare the differences among multiple groups. If the variances were uniform, the least significant difference test was used. If the variances were not uniform, Dunnett's T3 test was used. A p value of <0.05 was considered statistically significant, and a p value of <0.01 was considered highly statistically significant.
3. Results
3.1. JDYZF Can Improve Cognitive Impairment in AD Rats Induced with Aβ25–35
The spatial learning and memory of rats were evaluated using the water maze test by calculating the escape latency of each group during the first 4 days and the average escape latency. The results showed no significant difference between the MG and other groups (except JDYZ.M) on the first day. On days 2 to 4, the escape latencies of the MG and other groups differed significantly (Figure 1(a)). The average escape latency was significantly higher in the MG group than in the other groups (p < 0.01) (Figure 1(b)). In addition, after the removal of the platform, the rats in the MG group spent less time swimming in the quadrant in which the platform was located (p < 0.05 except PG) (Figure 1(c)) and crossed the platform fewer times than those in the other groups (p < 0.05 except PG and JDYZ.H) (Figure 1(d)). Moreover, after removing the platform, the swimming trajectories of the MG and PG groups were sparse and irregular, while that of the CG group was dense and tended to be in the quadrant with the platform; the trajectories of the JDYZF groups were similar to those of the CG group to varying degrees (Figure 1(e)). This finding indicates that the injection of Aβ25–35 into the hippocampal CA1 area impaired the spatial learning and memory abilities of the rats. JDYZF and donepezil attenuated the learning and memory impairments to varying degrees. Notably, the effect of JDYZF was more prominent, especially in the JDYZ.L group.
Figure 1.
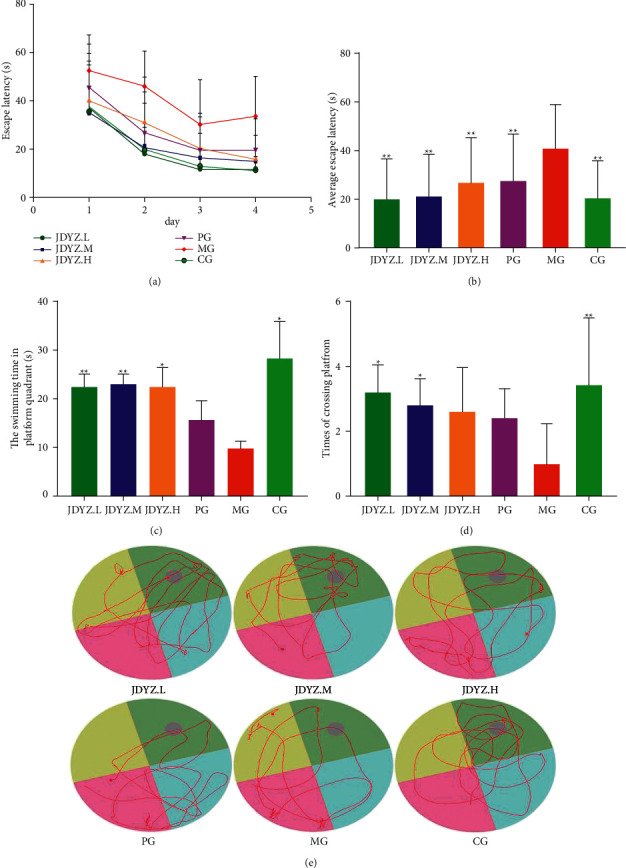
Oral administration of JDYZF rescues cognitive deficits in Aβ25–35-induced rats: (a) the rats in each group were trained in the water maze for 4 days, and the escape latency (expressed as means ± SEM) was gradually decreasing. The average escape latency (b), the swimming time in platform quadrant (c), and the number of crosses in the quadrant (d) were measured and recorded. (e) Representative swimming trajectories of different groups rats after removing the platform. Use one-way ANOVA to compare differences between multiple groups. ∗p < 0.05 and ∗∗p < 0.01, other groups compared with the model group.
3.2. JDYZF Attenuates Hippocampal Neuron Damage in AD Rats Induced with Aβ25–35
The deposition of Aβ can lead to neuronal cell loss and degeneration, which are the basis for the impairment of cognitive function in AD [35]. H&E staining showed that the pyramidal cells in the hippocampal CA1 area in the CG had the following features: tightly arranged, distinct layers, clear and complete morphology, rich cytoplasm, round nuclei, obvious nucleoli, and normal staining. In addition, the MG had the following features: disordered pyramidal cell arrangement, reduced number of layers, changed morphology, unclear boundary between the cytoplasm and nucleus, pyknotic nuclei, and deepened staining. The number and morphological structure of pyramidal cells in the CA1 area of the hippocampus were greatly recovered in the JDYZ.M, JDYZ.H, and PG groups compared with the MG. However, the effect was not as good as that in the JDYZ.L group (Figure 2). This finding indicates that JDYZF has some protective effect on hippocampal neurons.
Figure 2.
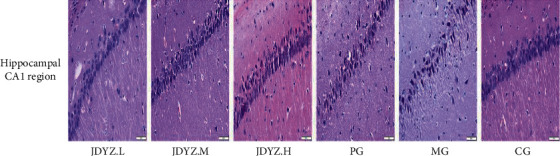
Representative histopathological photos of hippocampal CA1 area tissue sections from each group. JDYZF reduces Aβ1–42 levels in multiple tissues in AD model rats.
The Aβ23–35 fragment is neurotoxic to Aβ1–42; after brain injection, the Aβ23–35 fragment causes a large amount of expression and deposition of Aβ oligomers, thereby triggering AD [36]. Owing to the presence of the gut-brain axis, AD cause changes in the intestinal ecology. Intestinal bacteria may increase the secretion of Aβ homologous proteins in the intestine, spread through the gut-brain axis to the brain, and trigger a vicious cycle of Aβ deposition [1]. We used an ELISA kit to measure the expression levels of Aβ1–42 in the hippocampus, cortex, colon, and serum samples of rats in each group. Compared with the vehicle injection, the Aβ23–35 hippocampal injection increased the expression level of Aβ1–42 in the rat hippocampus, cortex, colon, and serum. Orally administered donepezil slightly attenuated the increases in the Aβ1–42 levels; however, the effect was not as good as that of the JDYZF concoctions, especially that of JDYZ.L (Figures 3(a)–3(d)). This finding indicates that JDYZF can reduce the expression level of Aβ in AD model rats.
Figure 3.
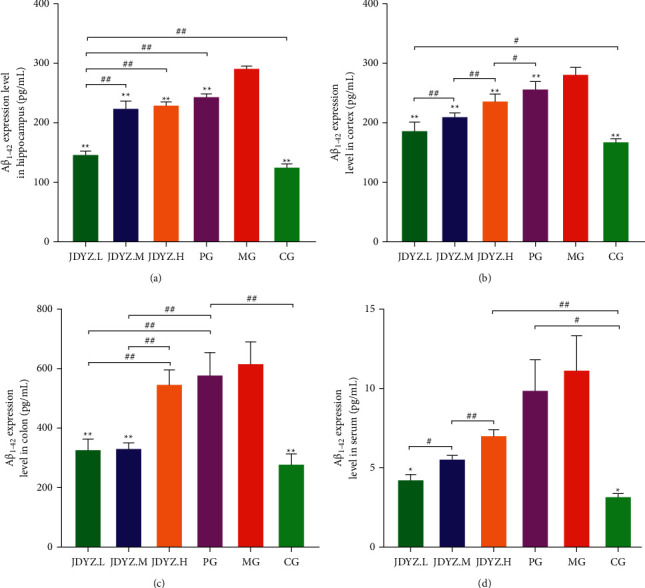
JDYZF oral administration can reduce the content of Aβ1–42 in the hippocampus (a), cerebral cortex (b), colon (c), and serum (d) of AD rats induced by Aβ25–35. ∗p < 0.05 and ∗∗p < 0.01, other groups compared with the model group. In addition to the model group, #p < 0.05 and ##p < 0.01, and the other groups are compared in pairs.
3.3. JDYZF Inhibits the Expression of Proteins Related to the Pyroptosis Pathway
Pyroptosis-related proteins are highly expressed in the brains of APP/PS1 transgenic mice [37]. Aβ triggers neuroinflammation by activating the pyroptosis pathway, causing neuronal damage, which may be an important mechanism of AD pathogenesis. Therefore, we assessed the expression of proteins related to the classical pyroptosis pathway in the rat hippocampus using Western blotting. Quantitative analysis revealed that Aβ23–35 hippocampal injection led to significant increases in NLRP3, GSDMD, pro-caspase-1, and caspase-1 P20 expression levels. Compared with donepezil, which slightly downregulated the expression of these proteins, JDYZF appeared to be more effective (Figure 4(a)). Notably, in the nonclassical pathway of pyroptosis, caspase-11 is directly activated by LPS to induce pyroptosis. Accordingly, we assessed the expression of the caspase-11 precursor and its cleaved products. Surprisingly, the results were consistent with those of the levels of proteins in the classical pathway (Figure 4(b)). This finding was further confirmed by the immunohistochemical staining of hippocampal samples from each group (Figure 5). These data suggest that JDYZF reduces nerve injury and improves cognitive function by downregulating the expression of proteins related to the pyroptosis pathway.
Figure 4.
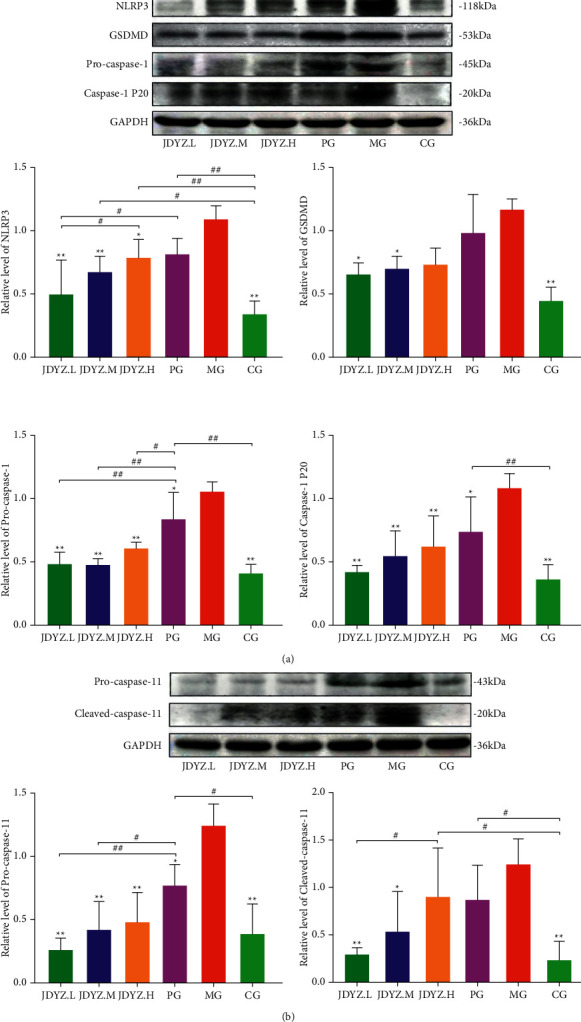
Compared to MG, oral administration of JDYZF and donepezil can reduce the expression levels of NLRP3, GSDMD, pro-caspase-1, and caspase-1 p20 in AD rats hippocampi (a). In addition, by reducing the expression of caspase-11 precursor and spliceosome, the activity of the nonclassical pyrolysis pathway is inhibited and the cognitive impairment of AD rats is improved (b). The relative level is equal to the gray value of the relevant protein divided by the gray value of GAPDH. ∗p < 0.05 and ∗∗p < 0.01, other groups compared with the model group. #p < 0.05 and ##p < 0.01, the other groups are compared in pairs except the model group.
Figure 5.
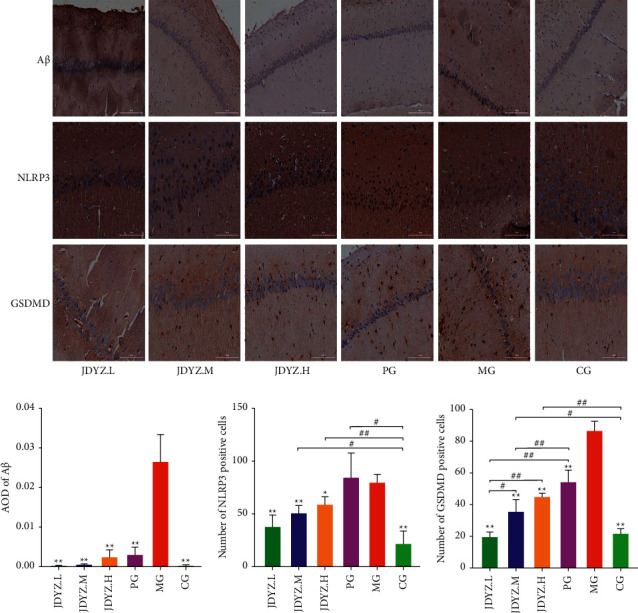
Oral administration of JDYZF can reduce the deposition of Aβ and the expression of NLRP3 and GSDMD in the hippocampi of AD rats, which is consistent with the results of ELISA and Western blot. Use ImageJ software to measure the average optical density of Aβ in the hippocampal slices, and calculate the number of positive cells of NLRP3 and GSDMD in each field of view. The scale bar of Aβ is 200 um, the scale bar of NLRP3, and GSDMD is 100 um. Statistics and results display methods are the same as before.
3.4. JDYZF Reduces the Levels of IL-1β and IL-18 in Multiple Tissues of AD Model Rats
Activated caspase-1 and caspase-11 cleave pro-IL-1β and pro-IL-18 and produce mature/activated IL-1β and IL-18, which aggravate local inflammation. We measured the levels of IL-1β and IL-18 in the hippocampus, cortex, colon, and serum samples of rats in each group. The levels of IL-1β and IL-18 were higher in the MG than in the CG. Donepezil and JDYZF decreased the levels of IL-1β and IL-18; however, the effect of JDYZF, especially JDYZ.L, was more apparent (Figures 6(a)–6(h)). This finding indicates that JDYZF may inhibit the inflammatory reaction in AD model rats.
Figure 6.
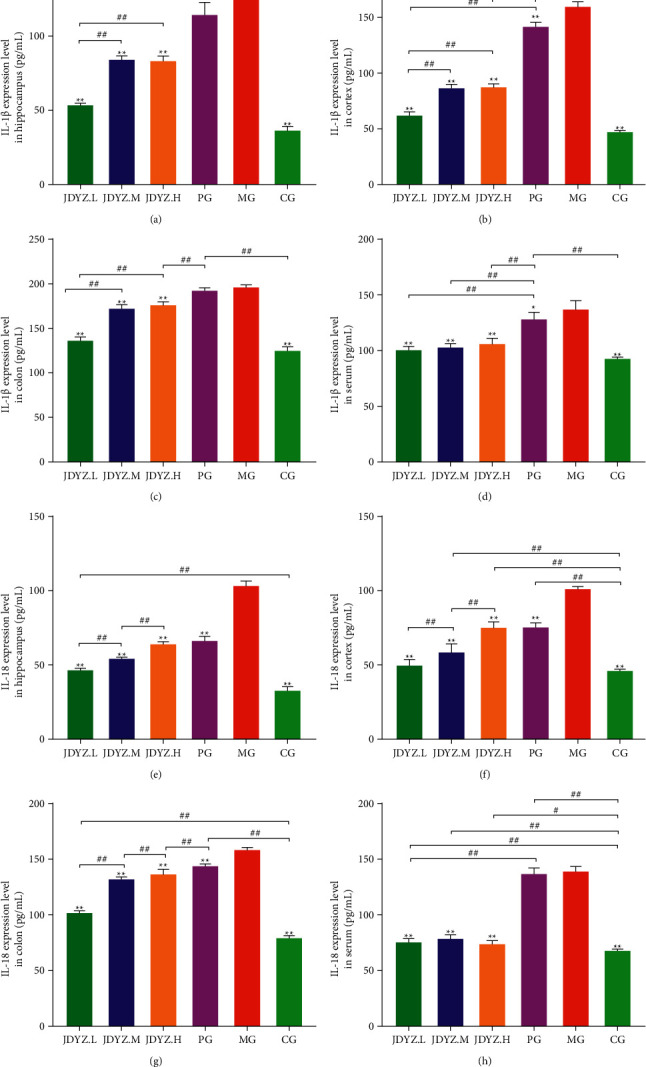
JDYZF can reduce the inflammatory response in AD rats by inhibiting the expression of IL-1β in the hippocampus (a), cerebral cortex (b), colon (c), and serum (d) and the expression of IL-18 in the hippocampus (e), cerebral cortex (f), colon (g), and serum (h).
3.5. JDYZF Has No Hepatotoxicity
AD, as a chronic neurological disease, requires long-term oral drug treatment. Therefore, the safety of the drug is particularly important. We found that the administration of JDYZF did not cause significant changes in the liver structure based on the histological analyses of the livers of rats in each group (Figure 7). This finding indicates that oral JDYZF is safe and reliable for the treatment of AD.
Figure 7.
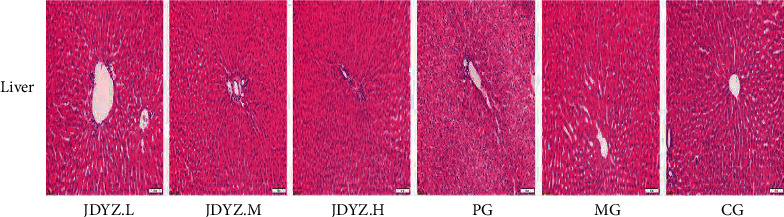
Representative images of HE staining of tissue sections from the rat liver. Aβ hippocampal injection and oral administration of JDYZF did not cause significant liver structural damage, indicating that JDYZF has no hepatotoxicity, while oral donepezil caused slight hepatic edema.
4. Discussion
Currently, when the development of specific drugs to treat AD is blocked, various alternative methods are used. Clinical studies have shown that acupuncture at acupoints that replenish qi, resolve phlegm, and promote blood circulation can reduce Alzheimer's Disease Assessment Scale-cognitive subscale (ADAS-cog) and Clinician's Interview-Based Impression of Change-Plus (CIBIC-Plus) scores of AD patients [38]. In an animal experiment with similar acupoints, acupuncture reduced the protein expression levels of NLRP3, Caspase-1, and IL-1β in the hippocampi of AD model mice, inhibited the activation of microglia, and improved cognitive function [39]. Electroacupuncture at acupoints on the head has also been reported to improve the Montreal Cognitive Assessment (MoCA) scores of AD patients [40]. Animal experiments have also confirmed that electroacupuncture at acupoints on the head can enhance hippocampal and prefrontal cortex neuroprotection and regulate synaptic plasticity [41]. In addition, music therapy can alter the levels of neurotransmitters, autonomic nerve function, and neuronal connections to improve the memory and language ability of AD patients and reduce their mental symptoms [42, 43]. Aromatherapy can enhance neurogenesis in the limbic system of the brain through the projection of odour stimulation to improve cognitive impairment [11]. Evidence also supports the biological effects of pulsed electromagnetic fields in the treatment of AD, and their ability to improve cognitive impairment may be achieved by the modulation of insulin growth factors (IGFs) [12]. The emergence of these alternative therapies has enriched the treatment methods of AD and can be used in clinical treatment according to the specific conditions of the patient to maximize improvements in their clinical symptoms. In contrast, traditional Chinese medicine formulas are unique in the treatment of AD because of their multicomponent, multitargeted, and multipathway regulatory effects [14]. For example, the classic formula Shen-Zhi-Ling Oral Liquid, which is used for the treatment of AD, can reduce APP mRNA expression in the hippocampus, reduce amyloid deposition, reduce Caspase-3 expression, reduce neuronal apoptosis [44, 45], and increase hippocampal haemoglobin. The expression of oxygenase-1 (HO-1) and biliverdin reductase (BVR) can resist oxidative damage [46], regulate the insulin signal transduction pathway INR/PI3K/Akt, and improve glucose uptake, transport, and glycolysis in the brain [47]. The methods for improving AD are diversified and conform to the various pathological mechanisms of AD, thus achieving good clinical results.
A clinical study of JDYZF showed that it improved the Mini-Mental State Examination (MMSE) and MoCA scores in patients with cognitive impairment and had a positive effect on patients with AD [48]. In a previous mechanistic study, we found that the effect of Coptis polysaccharide, the extract of Coptis chinensis and the main component of JDYZF, on improving AD was multifaceted, which led us to hypothesize that JDYZF is more effective than other treatments.
The production and excessive accumulation of the toxin Aβ is an important factor driving the occurrence and progression of AD, and SPs are a pathological hallmark of AD [49]. SPs are formed by the accumulation of excess Aβ, which can induce the formation of NFTs, damage blood vessels, and induce neuronal loss [50]. Studies have shown that before numerous Aβ aggregates produce SPs, soluble Aβ oligomers cause nerve damage in many ways [51]. These oligomers can inhibit long-term potentiation (LTP) by enhancing the response of the N-methyl-D-aspartate receptor (NMDA) [52]. Oligomers can also directly interact with cell membranes to disrupt their permeability and bind with the cellular prion protein (PrPc) to disrupt the synaptic function of hippocampal neurons [53, 54]. Oligomers can bind with other disease-causing proteins associated with AD, such as TAR DNA-binding protein 43 (TDP-43), to form mixed oligomers and cause neuronal death [55]. Before the formation of SPs, neuroinflammation occurs in the AD brain [56], which may be related to the inflammatory response mediated by Aβ oligomers through receptors for advanced glycation end products (RAGEs), Toll-like receptors, and NLRs [57]. In this study, ELISA and immunohistochemical analyses confirmed that JDYZF reduced the expression level of soluble Aβ1–42 in the hippocampi of AD rats and reduced the Aβ deposition in plaques. This outcome may account for the mechanism by which JDYZF induces neuroprotective effects and reduces the neuroinflammatory response.
The response of NLRP3 to Aβ oligomers leads to the assembly of ASC and caspase-1 to form inflammasomes with NLRP3, which in turn activates GSDMD. Cell death and exogenous inflammatory factors are thereby induced via a process termed pyroptosis. During this process, ASC can bind to Aβ to further induce the production of Aβ oligomers [58], and inflammasomes can weaken the phagocytic function of microglia to take up Aβ, disrupting Aβ clearance [59]. The local inflammatory response caused by exogenous inflammatory factors can also promote the production and aggregation of Aβ [60], forming a vicious cycle between the occurrence of Aβ and pyroptosis and promoting the progression of AD. AD worsening can be slowed only by breaking this cycle. Studies have shown that knocking out the NLRP3 or Caspase-1 gene in APP/PS1 mice reduces the Aβ deposition in the brain and returns the LTP to nearly the baseline level [61]. Some researchers believe that the functional state of microglia in APP/PS1/NLRP3-/- or APP/PS1/Caspase-1-/- mice is shifted towards the M2 phenotype, which has anti-inflammatory effects and an enhanced ability to clear Aβ, which in turn has a neuroprotective effect [62]. Therefore, intervening in the NLRP3 inflammasome has become a new strategy for the treatment of AD. We herein showed that JDYZF reduced the protein expression levels of NLRP3, pro-caspase-1, and caspase-1 P20 in the hippocampi of AD rats and inhibited the excessive activation of NLRP3 inflammasomes, thereby inhibiting the maturation of GSDMD, breaking the vicious cycle between Aβ and pyroptosis and improving cognitive impairment in AD rats. In addition, studies have confirmed that activation of the NLRP3/caspase-1/GSDMD pathway damages the blood-brain barrier (BBB) [63]. When an imbalance in the intestinal flora in AD patients leads to an increase in intestinal LPS, BBB damage may lead to an increase in the LPS level in the brain [1, 64, 65]. Caspase-11 can directly recognize LPS and cleave GSDMD to initiate pyroptosis. Caspase-11 can also activate the NLRP3/caspase-1/GSDMD pathway, resulting in the expansion of pyroptosis [66], representing another vicious cycle that leads to the progression of AD. This study showed that JDYZF also reduced the expression of the caspase-11 precursor and its splicing body and interfered with the nonclassical pyroptosis pathway. Therefore, we hypothesize that JDYZF improves the intestinal flora imbalance in AD rats and reduces the levels of LPS in the intestine and brain. However, this hypothesis needs to be further explored in future studies.
IL-1β, located upstream of the inflammatory response in the brain, recruits peripheral immune cells to infiltrate the CNS and produce proinflammatory factors, induces APP expression to increase and promote the production and aggregation of Aβ [67–69], and plays an important role in the formation of a vicious cycle of Aβ and pyroptosis. In addition, the overexpression of IL-1β and IL-18 can damage the BBB, promote the expansion of central pyroptosis, and aggravate the inflammatory response [69]. Herein, JDYZF reduced the expression levels of IL-1β and IL-18 in multiple AD rat tissues, reduced the inflammatory response, and weakened the effect of inflammatory factors on the vicious cycle of Aβ production and pyroptosis.
5. Conclusion
In summary, the pathogenesis of AD is complicated and unclear. Clinicians should choose comprehensive treatment methods that are suitable for patients to improve their condition to the greatest extent. JDYZF has achieved good results as a clinical treatment for AD. It may reduce the expression and deposition of Aβ, inhibit the expression levels of pyroptosis-related proteins, reduce the neuroinflammatory response, and block the vicious cycles between Aβ and pyroptosis, pyroptosis and LPS, and inflammation, Aβ and pyroptosis, thereby improving cognitive impairment in AD rats. By exploring AD treatment mechanisms, we hope to provide an alternative treatment.
Acknowledgments
The authors thank Professor Linbo Zhang of Jilin Agricultural University and researcher Chao Xu of the Institute of Specialty of Chinese Academy of Agricultural Sciences for their technical support. This work was supported by the project of Jilin Provincial Health Commission (grant no. 2019J058).
Data Availability
The data used to support the results of this study are included in the article.
Conflicts of Interest
The authors declare no conflicts of interest.
Authors' Contributions
M. L. and X. Z. conceived the project, designed the experiments, and revised the manuscript. J. W. designed and performed most of the experiments, analysed the data, and wrote the manuscript. Y. L., P. Z., and T. W. assisted with some experiments.
References
- 1.Sochocka M., Donskow-Łysoniewska K., Satler Diniz B. The gut microbiome alterations and inflammation-driven pathogenesis of Alzheimer’s disease—a critical review. Molecular Neurobiology . 2018;56(3):1841–1851. doi: 10.1007/s12035-018-1188-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Breijyeh Z., Karaman R. Comprehensive review on Alzheimer’s disease: causes and treatment. Molecules . 2020;25(24):p. 5789. doi: 10.3390/molecules25245789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hodson R. Alzheimer’s disease. Nature . 2018;559 doi: 10.1038/d41586-018-05717-6.7715 [DOI] [PubMed] [Google Scholar]
- 4.Selkoe D., Mandelkow E., Holtzman D. Deciphering alzheimer disease. Cold Spring Harbour Protocols in Medicine . 2012;2(1) doi: 10.1101/cshperspect.a011460.a011460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Liu S., Li X., Gao J., Liu Y., Shi J., Gong Q. Icariside II, a phosphodiesterase-5 inhibitor, attenuates beta-amyloid-induced cognitive deficits via BDNF/TrkB/CREB signaling. Cellular Physiology and Biochemistry . 2018;49(3):1010–1025. doi: 10.1159/000493232. [DOI] [PubMed] [Google Scholar]
- 6.Jia Y., Yang H., Ren J. CRMP2-derived peptide ST2-104 (R9-CBD3) protects SH-SY5Y neuroblastoma cells against Aβ25-35-induced neurotoxicity by inhibiting the pCRMP2/NMDAR2B signaling pathway. Chemico-Biological Interactions . 2019;3(5):28–39. doi: 10.1016/j.cbi.2019.03.005. [DOI] [PubMed] [Google Scholar]
- 7.Costandi M. Ways to stop the spread of Alzheimer’s disease. Nature . 2018;559 doi: 10.1038/d41586-018-05723-8.7715 [DOI] [PubMed] [Google Scholar]
- 8.Zeng B.-Y., Salvage S., Jenner P. Effect and mechanism of acupuncture on Alzheimer’s disease. International Review of Neurobiology . 2013;111:181–195. doi: 10.1016/b978-0-12-411545-3.00009-2. [DOI] [PubMed] [Google Scholar]
- 9.Yang W.-D., Dong W.-G. Mechanisms of electroacupuncture for improving Alzheimer’s disease from reducing β amyloid protein level. Zhen Ci Yan Jiu . 2020;45(5):426–431. doi: 10.13702/j.1000-0607.180885. [DOI] [PubMed] [Google Scholar]
- 10.Garcia-Casares N., Moreno-Leiva R. M., Garcia-Arnes J. A. Music therapy as a non-pharmacological treatment in Alzheimer’s disease. A systematic review. Revue Neurologique . 2017;65(12):529–538. [PubMed] [Google Scholar]
- 11.Jimbo D., Kimura Y., Taniguchi M., Inoue M., Urakami K. Effect of aromatherapy on patients with Alzheimer’s disease. Psychogeriatrics . 2009;9(4):173–179. doi: 10.1111/j.1479-8301.2009.00299.x. [DOI] [PubMed] [Google Scholar]
- 12.Li Y., Zhang Y., Wang W., et al. Effects of pulsed electromagnetic fields on learning and memory abilities of STZ-induced dementia rats. Electromagnetic Biology and Medicine . 2019;38(2):123–130. doi: 10.1080/15368378.2019.1591437. [DOI] [PubMed] [Google Scholar]
- 13.Howes M.-J. R., Fang R., Houghton P., Houghton J. Effect of Chinese herbal medicine on Alzheimer’s disease. International Review of Neurobiology . 2017;135:29–56. doi: 10.1016/bs.irn.2017.02.003. [DOI] [PubMed] [Google Scholar]
- 14.Su Y., Wang Q., Wang C. The treatment of Alzheimer’s disease using Chinese medicinal plants: from disease models to potential clinical applications. Journal of Ethnopharmacology . 2014;152(3):403–423. doi: 10.1016/j.jep.2013.12.053. [DOI] [PubMed] [Google Scholar]
- 15.Li Y., Guan S., Liu C., et al. Neuroprotective effects of Coptis chinensis Franch polysaccharide on amyloid-beta (Aβ)-induced toxicity in a transgenic Caenorhabditis elegans model of Alzheimer’s disease (AD) International Journal of Biological Macromolecules . 2018;113(113):991–995. doi: 10.1016/j.ijbiomac.2018.03.035. [DOI] [PubMed] [Google Scholar]
- 16.Li Y., Liu C., Zhu X. Inhibiting c-Jun N-terminal kinase(JNK)-mediated apoptotic signaling pathway inPC12 cells by a polysaccharide(CCP)from coptis chinensis against amyloid-β(Aβ)-induced neurotoxicity. International Journal of Biological Macromolecules . 2019;134:5651–6574. doi: 10.1016/j.ijbiomac.2019.05.041. [DOI] [PubMed] [Google Scholar]
- 17.Han C., Yang Yi, Guan Q. New mechanism of nerve injury in Alzheimer’s disease: β-amyloid-induced neuronal pyroptosis. Journal of Cellular and Molecular Medicine . 2020;24(14):8078–8090. doi: 10.1111/jcmm.15439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Feng J., Li M., Qian W. Unconjugated bilirubin induces pyroptosis in cultured rat cortical astrocytes. Journal of Neuroinflammation . 2018;15(1) doi: 10.1186/s12974-018-1064-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bergsbaken T., Fink S. L., Cookson B. T. Pyroptosis: host cell death and inflammation. Nature Reviews Microbiology . 2009;7(2):99–109. doi: 10.1038/nrmicro2070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liao Z., Su J. Progresses on three pattern recognition receptor families (TLRs, RLRs and NLRs) in teleost. Developmental and Comparative Immunology . 2021;122 doi: 10.1016/j.dci.2021.104131.104131 [DOI] [PubMed] [Google Scholar]
- 21.Srinivasula S. M., Poyet J.-L., Razmara M. The PYRIN-CARD protein ASC is an activating adaptor for caspase-1. Journal of Biological Chemistry . 2002;277(24):21119–21122. doi: 10.1074/jbc.C200179200. [DOI] [PubMed] [Google Scholar]
- 22.Zhao Y., Shao F. Diverse mechanisms for inflammasome sensing of cytosolic bacteria and bacterial virulence. Current Opinion in Microbiology . 2016;29:37–42. doi: 10.1016/j.mib.2015.10.003. [DOI] [PubMed] [Google Scholar]
- 23.Wang Li, Arthur V. The NLRP3 inflammasome: mechanism of action, role in disease and therapies. Molecular Aspects of Medicine . 2020;76 doi: 10.1016/j.mam.2020.100889.100889 [DOI] [PubMed] [Google Scholar]
- 24.Patel M. N., Carroll R. G. Inflammasome priming in sterile inflammatory Disease. Trends in Molecular Medicine . 2017;23(2):165–180. doi: 10.1016/j.molmed.2016.12.007. [DOI] [PubMed] [Google Scholar]
- 25.Heneka M. T., Kummer M. P., Stutz A., et al. NLRP3 is activated in Alzheimer’s disease and contributes to pathology in APP/PS1 mice. Nature . 2013;493(7434):674–678. doi: 10.1038/nature11729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Rathinam V. A. K., Zhao Y., Shao F. Innate immunity to intracellular LPS. Nature Immunology . 2019;20(5):527–533. doi: 10.1038/s41590-019-0368-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ma C.-Y., Liu J.-H., Liu J.-X., et al. Relationship between two blood stasis syndromes and inflammatory factors in patients with acute coronary syndrome. Chinese Journal of Integrative Medicine . 2017;23(11):845–849. doi: 10.1007/s11655-016-2746-0. [DOI] [PubMed] [Google Scholar]
- 28.Wei Y. L.. M., Yue K. Study on urine metabolic profile of Aβ25-35-Induced Alzheimer’s disease by using UHPLC-Q-TOF-MS. Neuroscience . 2018;10(1):30–43. doi: 10.1016/j.neuroscience.2018.10.001. [DOI] [PubMed] [Google Scholar]
- 29.Zuo H., Liu X., Wang D., et al. RKIP-Mediated NF-κB Signaling is involved in ELF-MF-mediated improvement in AD rat. International Journal of Medical Sciences . 2018;15(14):1658–1666. doi: 10.7150/ijms.28411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wu W., He X., Xie S. Protective effects of Huang-Lian-Jie-Du-Tang against Ab25–35-induced memory deficits and oxidative stress in rats. Journal of International Medical Research . 2020;48(3) doi: 10.1177/0300060519893859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Deng-Bryant Y., Leung L. Y., Caudle K., Tortella F., Shear D. Cognitive evaluation using Morris water maze in neurotrauma. Methods in Molecular Biology . 2016;1462:539–551. doi: 10.1007/978-1-4939-3816-2_29. [DOI] [PubMed] [Google Scholar]
- 32.Cardiff R. D., Miller H., Munn R. J. Manual hematoxylin and eosin staining of mous tissue sections. Cold Spring Harbour Protocols . 2014;2014(6):655–658. doi: 10.1101/pdb.prot073411. [DOI] [PubMed] [Google Scholar]
- 33.Ni D., Xu P., Gallagher S. Immunoblotting and Immunodetection. Current Protocols in Cell Biology . 2017;74:1–37. doi: 10.1002/cpcb.18. [DOI] [PubMed] [Google Scholar]
- 34.Ramos-Vara J. A. Principles and methods of immunohistochemistry. Methods in Molecular Biology . 2017;1641:115–128. doi: 10.1007/978-1-4939-7172-5_5. [DOI] [PubMed] [Google Scholar]
- 35.Viola K. L., Klein W. L. Amyloid β oligomers in Alzheimer’s disease pathogenesis, treatment, and diagnosis. Acta Neuropathologica . 2015;129(2):183–206. doi: 10.1007/s00401-015-1386-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Li W.-Z., Li W.-P., Huang D.-K. Dexamethasone and Aβ25–35 accelerate learning and memory impairments due to elevate amyloid precursor protein expression and neuronal apoptosis in 12-month male rats. Behavioural Brain Research . 2012;227(1):142–149. doi: 10.1016/j.bbr.2011.10.038. [DOI] [PubMed] [Google Scholar]
- 37.Song X., Cui Z., He J. κ-opioid receptor agonist, U50488H,inhibits pyroptosis through NLRP3 via the Ca2+/CaMKII/CREB signaling pathway and improves synaptic plasticity in APP/PS1 mice. Molecular Medicine Reports . 2021;24(1):529–538. doi: 10.3892/mmr.2021.12168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jia Y., Zhang X., Yu J. Acupuncture for patients with mild to moderate Alzheimer’s disease, 2017 disease: a randomized controlled trial. Molecular Neurobiology . 2017;17(1):556–564. doi: 10.1186/s12906-017-2064-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.He C., Huang Z.-S., Chen H.-Ru. Effect of pretreatment of acupuncture on learning-memory ability and related protein of NLRP3 inflammasome in hippocampus in Alzheimer’s disease like rats. Zhongguo Zhen Jiu . 2020;40(12):1323–1327. doi: 10.13703/j.0255-2930.20191012-k0004. [DOI] [PubMed] [Google Scholar]
- 40.Xia K.-P., Pang J., Li S.-L. Effect of electroacupuncture at governor vessel on learning-memory ability and serum level of APP, Aβ1-42 in patients with Alzheimer’s disease. Zhongguo Zhen Jiu . 2020;40(4):375–378. doi: 10.13703/j.0255-2930.20190728-0003. [DOI] [PubMed] [Google Scholar]
- 41.Zheng Yu, Qin Z., Tsoi B. Electroacupuncture on trigeminal nerve-innervated acupoints ameliorates poststroke cognitive impairment in rats with middle cerebral artery occlusion: involvement of neuroprotection and SynapticPlasticity. Neural Plasticity . 2020;2020 doi: 10.1155/2020/8818328.8818328 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Peck K. J., Girard T. A., Russo F. A., Fiocco A. J. Music and memory in Alzheimer’s disease and the potential underlying mechanisms. Journal of Alzheimer’s Disease . 2016;51(4):949–959. doi: 10.3233/jad-150998. [DOI] [PubMed] [Google Scholar]
- 43.Lyu J., Zhang J., Mu H., et al. The effects of music therapy on cognition, psychiatric symptoms, and activities of daily living in patients with Alzheimer’s disease. Journal of Alzheimer’s Disease . 2018;64(4):1347–1358. doi: 10.3233/jad-180183. [DOI] [PubMed] [Google Scholar]
- 44.Pan W., Wang Q., Kwak S. Shen-zhi-ling oral liquid improves behavioral and psychological symptoms of dementia in Alzheimer’s disease. Evidence-Based Complementary and Alternative Medicine . 2014;2014:6. doi: 10.1155/2014/913687.913687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Chi H., Liu T., Pan W., et al. Shen-Zhi-Ling oral solution improves learning and memory ability in Alzheimer’s disease mouse model. Journal of traditional Chinese medicine . 2019;39(5):667–677. [PubMed] [Google Scholar]
- 46.Xing S., Shen D., Chen C., Wu B., Chi H. Effect of the herbal formulation Shen-Zhi-Ling on an APP/PS1 mouse model of Alzheimer’s disease by modulating the biliverdin reductase/heme oxygenase 1 system. Experimental and Therapeutic Medicine . 2017;14(3):1961–1966. doi: 10.3892/etm.2017.4732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Qin G., Dong Y., Liu Z. Shen-Zhi-Ling oral liquid ameliorates cerebral glucose metabolism disorder in early AD via insulin signal transduction pathway in vivo and in vitro. Chinese Medicine . 2021;16(1):p. 128. doi: 10.1186/s13020-021-00540-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Li M., Wang Q., Liu C. Jiedu Yizhi decoction in the treatment of non-dementia vascular cognitive impairment. Jilin Journal of Chinese Medicine . 2018;38(9):1057–1060. [Google Scholar]
- 49.Gouras G., Gouras K., Olsson T. T., Hansson O. β-Amyloid peptides and amyloid plaques in Alzheimer’s disease. Neurotherapeutics . 2015;12(1):3–11. doi: 10.1007/s13311-014-0313-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hardy J. A., Higgins G. A. Alzheimer’s disease: the amyloid cascade hypothesis. Science . 1992;256(5054):184–185. doi: 10.1126/science.1566067. [DOI] [PubMed] [Google Scholar]
- 51.Gandy S., Simon A. J., Steele J. W., et al. Days to criterion as an indicator of toxicity associated with human Alzheimer amyloid-beta oligomers. Annals of Neurology . 2010;68(2):220–230. doi: 10.1002/ana.22052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Li S., Jin M., Koeglsperger T., Shepardson N. E., Shankar G. M., Selkoe D. J. Soluble A oligomers inhibit long-term potentiation through a mechanism involving excessive activation of extrasynaptic NR2B-containing NMDA receptors. Journal of Neuroscience . 2011;31(18):6627–6638. doi: 10.1523/jneurosci.0203-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kayed R., Lasagna-Reeves C. A. Molecular mechanisms of amyloid oligomers toxicity. Journal of Alzheimers Disease . 2013;33:S67–S78. doi: 10.3233/JAD-2012-129001. [DOI] [PubMed] [Google Scholar]
- 54.Freir D., Freir B., Nicoll A. J., et al. Interaction between prion protein and toxic amyloid β assemblies can be therapeutically targeted at multiple sites. Nature Communications . 2011;2(1):336–345. doi: 10.1038/ncomms1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Fang Y.-S., Tsai K.-J., Chang Y.-J., et al. Full-length TDP-43 forms toxic amyloid oligomers that are present in frontotemporal lobar dementia-TDP patients. Nature Communications . 2014;5(1):4824–4837. doi: 10.1038/ncomms5824. [DOI] [PubMed] [Google Scholar]
- 56.Sengupta U., Nilson A. N., Kayed R. The role of amyloid-β oligomers in toxicity, propagation, and immunotherapy. EBioMedicine . 2016;6:42–49. doi: 10.1016/j.ebiom.2016.03.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Minter M. R., Taylor J. M., Crack P. J. The contribution of neuroinflammation to amyloid toxicity in Alzheimer’s disease. Journal of Neurochemistry . 2016;136(3):457–474. doi: 10.1111/jnc.13411. [DOI] [PubMed] [Google Scholar]
- 58.Venegas C., Kumar S., Franklin B. S., et al. Microglia-derived ASC specks cross-seed amyloid-β in Alzheimer’s disease. Nature . 2017;552(7685):355–361. doi: 10.1038/nature25158. [DOI] [PubMed] [Google Scholar]
- 59.Zhang Y., Dong Z., Song W. NLRP3 inflammasome as a novel therapeutic target for Alzheimer’s disease. Signal Transduction and Targeted Therapy . 2020;5(1):37–361. doi: 10.1038/s41392-020-0145-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tan M.-S., Yu J.-T., Jiang T., Zhu X.-C., Tan L. The NLRP3 inflammasome in Alzheimer’s disease. Molecular Neurobiology . 2013;48(3):875–882. doi: 10.1007/s12035-013-8475-x. [DOI] [PubMed] [Google Scholar]
- 61.Heneka M. T. Inflammasome activation and innate immunity in Alzheimer’s disease. Brain Pathology . 2017;27(2):220–222. doi: 10.1111/bpa.12483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Dempsey C., Rubio Araiz A., Bryson K. J., et al. Inhibiting the NLRP3 inflammasome with MCC950 promotes non-phlogistic clearance of amyloid-β and cognitive function in APP/PS1 mice. Brain, Behavior, and Immunity . 2017;61:306–316. doi: 10.1016/j.bbi.2016.12.014. [DOI] [PubMed] [Google Scholar]
- 63.Bai R., Lang Y., Shao J. The role of NLRP3 inflammasome in cerebrovascular diseases pathology and possible therapeutic targets. ASN Neuro . 2021;13:1–38. doi: 10.1177/17590914211018100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hill J. M., Lukiw W. J. Microbial-generated amyloids and Alzheimer’s disease (AD) Frontiers in Aging Neuroscience . 2015;7:9–14. doi: 10.3389/fnagi.2015.00009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhao Y., Walter J. Microbiome-generated amyloid and potential impact on amyloidogenesis in Alzheimer’s disease (AD) Journal of Natural Sciences . 2015;1(7):138–150. [PMC free article] [PubMed] [Google Scholar]
- 66.Young-Su Y. Caspase-11 non-canonical inflammasome: a critical sensor of intracellular lipopolysaccharide in macrophage-mediated inflammatory responses. Immunology . 2017;152(2):207–217. doi: 10.1111/imm.12787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Díaz A., Rojas K., Espinosa B. Aminoguanidine treatment ameliorates inflammatory responses and memory impairment induced by amyloid-beta 25-35 injection in rats. Neuropeptides . 2014;48(3):153–159. doi: 10.1016/j.npep.2014.03.002. [DOI] [PubMed] [Google Scholar]
- 68.Lynch M. A. Neuroinflammatory changes negatively impact on LTP: a focus on IL-1β. Brain Research . 2015;1621:197–204. doi: 10.1016/j.brainres.2014.08.040. [DOI] [PubMed] [Google Scholar]
- 69.Song L., Pei L., Yao S., Wu Y., Shang Y. NLRP3 inflammasome in neurological diseases, from functions to therapies. Frontiers in Cellular Neuroscience . 2017;11:63–80. doi: 10.3389/fncel.2017.00063. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data used to support the results of this study are included in the article.