

Abstract
A variety of 1β-methylcarbapenem derivatives were screened to identify inhibitors of IMP-1 metallo-β-lactamase, a class B β-lactamase, in an automated microassay system using nitrocefin as a substrate. The structure–inhibitory-activity relationship study revealed that three types of 1β-methylcarbapenems having benzothienylthio, dithiocarbamate, or pyrrolidinylthio moieties at the C-2 position showed good inhibitory activity. Among the compounds screened, J-110,441, having a benzothienylthio moiety at the C-2 position of 1β-methylcarbapenem, was the most potent inhibitor of class B metallo-β-lactamases with Ki values of 0.0037, 0.23, 1.00, and 0.83 μM for IMP-1 encoded by the blaIMP gene, CcrA from Bacteroides fragilis, L1 from Stenotrophomonas maltophilia, and type II from Bacillus cereus, respectively. In a further characterization study, J-110,441 also showed inhibitory activity against TEM-type class A serine β-lactamase and chromosomal class C serine β-lactamase from Enterobacter cloacae with Ki values of 2.54 and 0.037 μM, respectively. Combining imipenem or ceftazidime with J-110,441 had a synergistic effect on the antimicrobial activity against β-lactamase-producing bacteria. Against the isolates of IMP-1-producing Serratia marcescens, the MICs of imipenem decreased to levels ranging from 1/64 to 1/4 in the presence of one-fourth of the MIC of J-110,441. Against E. cloacae producing high levels of class C β-lactamase, the MIC of ceftazidime decreased from 64 to 4 μg/ml in the presence of 4 μg of J-110,441 per ml. This is the first report to describe a new class of inhibitor of class B and class C β-lactamases including transferable IMP-1 metallo-β-lactamases.
One of the most important mechanisms of microbial resistance to β-lactam antibiotics is hydrolysis by β-lactamases. Since carbapenems have a broader antimicrobial spectrum than do other β-lactam antibiotics and are not hydrolyzed by many clinically relevant serine β-lactamases, the medical use of carbapenems would be expected to increase. However, there are several carbapenem-hydrolyzing β-lactamases that preferentially hydrolyze carbapenems in addition to penicillins and cephalosporins (28). The class B metallo-β-lactamases, which have zinc atoms at the active site, are a group of such carbapenem-hydrolyzing enzymes (1, 5) and are minimally inhibited by β-lactamase inhibitors such as tazobactam (4, 23, 28). Besides, widely used serine β-lactamase inhibitors behave as substrates of class B β-lactamases (27).
The first metallo-β-lactamase-producing Pseudomonas aeruginosa strain was isolated in Japan in 1991 (38), and the outbreak of carbapenem-resistant organisms such as members of the family Enterobacteriaceae, especially Serratia marcescens, and pseudomonads in various districts in Japan was reported afterwards (2, 15, 31, 32). The mechanism of resistance was ascribed to the production of IMP-1 metallo-β-lactamase encoded on the blaIMP gene (2). The systematic survey of distribution by Senda et al. indicated the horizontal spread of blaIMP in Japan (31), and recently, blaIMP-carrying clinical isolates have been reported in Korea (19) and Europe (8). Thus, the spread of metallo-β-lactamase is a serious concern for antimicrobial chemotherapy with β-lactam antibiotics including carbapenems (20, 28), because the blaIMP gene is transferable among gram-negative bacteria (2, 31) and an effective inhibitor has not been developed yet.
In the screening for IMP-1 metallo-β-lactamase inhibitors, we found a potent β-lactamase inhibitor, J-110,441, having the benzothienylthio moiety at the C-2 position of the 1β-methylcarbapenem nucleus (21). In this paper, we describe the relationship between the structure and the β-lactamase-inhibitory activity of the carbapenem derivatives tested and discuss the properties of J-110,441 as a potential inhibitor of various β-lactamases.
MATERIALS AND METHODS
Compounds.
All carbapenems screened in this study (see Tables 1 to 3) were synthesized at the Tsukuba Research Institute, Banyu Pharmaceutical Co., Ltd., Tsukuba, Japan. Typical compounds shown in Table 4 were synthesized as described previously (13, 14, 24, 36). Imipenem, cilastatin sodium, and ampicillin were the products of Banyu Pharmaceutical Co., Ltd., Tokyo, Japan. The following drugs were commercially available: ceftazidime (Tanabe Pharmaceutical Co., Ltd., Osaka, Japan), aztreonam (Eisai Co., Ltd., Tokyo, Japan), cephaloridine and flomoxef (Shionogi Pharmaceutical Co., Osaka, Japan), and nitrocefin (Oxoid, Basingstoke, England).
TABLE 1.
Structure and IMP-1 metallo-β-lactamase-inhibitory activity of J-110,441 analogs

| Compound | R1 | R2 | IC50 (μM)a |
|---|---|---|---|
| J-110,441 |  |
H | <0.1 |
| 1a | H | 0.70 | |
| 1b | Me | >10 | |
| 1c | H | >10 | |
| 2a | H | 2.4 | |
| 2b | H | >10 | |
Nitrocefin was used as a substrate.
TABLE 3.
Structure and IMP-1 metallo-β-lactamase-inhibitory activity of pyrrolydinylthio carbapenem analogs
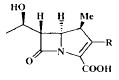
| Compound | R | IC50 (μM)a | Compound | R | IC50 (μM)a | |
|---|---|---|---|---|---|---|
| 4a |  |
<0.1 | 6a |  |
<0.1 | |
| 4b | 0.40 | 6b | 0.40 | |||
| 4c | 5.0 | 6c | 5.0 | |||
| 6d | 6.2 | |||||
| 5a (J-111,225) | 0.70 | 6e | 9.0 | |||
| 5b | >10 | 6f | 9.0 | |||
| 6g | >10 | |||||
| Side chain of 5a | >10 | |||||
| 7a | <0.1 | |||||
| 7b | <0.1 | |||||
| 7c | 0.80 | |||||
Nitrocefin was used as a substrate.
TABLE 4.
Stability and affinity for various types of β-lactamases
| β-Lactamase (original host) | J-110,441
|
Hydrolysis (relative %)a
|
|||
|---|---|---|---|---|---|
| Hydrolysis (relative %)a | Kib(μM) | Compound 3a | Compound 7a | Imipenem | |
| Metallo-β-lactamases | |||||
| IMP-1 (P. aeruginosa) | 1 | 0.0037 | 2 | 2 | 100 |
| CcrA (B. fragilis) | 3 | 0.23 | 22 | 18 | 100 |
| L1 (S. maltophilia) | 6 | 1.00 | 61 | 40 | 100 |
| Type II (B. cereus) | 8 | 0.83 | 7 | 63 | 100 |
| Serine β-lactamases | |||||
| Group 2b,d class A (E. coli) | <1 | 2.54 | 2 | NTc | <1 |
| Group 1,d class C (E. cloacae) | <1 | 0.037 | <1 | NT | <1 |
Taking the hydrolysis rates of imipenem and cephaloridine as 100% for metallo- and serine β-lactamases, respectively.
Imipenem and cephaloridine were used as the substrates for metallo- and serine β-lactamases, respectively.
NT, not tested.
Bush’s classification (3).
Bacterial strains.
Clinical isolates that produce IMP-1 metallo-β-lactamase were collected from various districts in Japan starting in 1994, and the blaIMP gene was detected by the PCR method (32). P. aeruginosa GN17203, which harbors blaIMP-carrying plasmid pMS350, and Bacteroides fragilis GAI30079 were generous gifts from M. Inoue, School of Medicine, Kitasato University, Kanagawa, Japan, and K. Watanabe, Institute of Anaerobic Bacteriology, School of Medicine, Gifu University, Gifu, Japan, respectively.
Susceptibility test.
MICs were determined by the twofold serial broth microdilution method with Mueller-Hinton broth (Difco Laboratories, Detroit, Mich.) for aerobes and GAM broth (Nissui Seiyaku Co., Ltd., Tokyo, Japan) for B. fragilis. A culture of aerobes grown at 37°C for 6 h in Mueller-Hinton broth was diluted to 107 CFU/ml, and a culture of B. fragilis grown at 37°C for 18 h under anaerobic conditions in GAM broth was diluted to 108 CFU/ml. Each dilution was inoculated into the drug-containing broth with an inoculum apparatus (MIC-2000; Dynatech Laboratories, Inc., Chantilly, Va.). The final inoculum sizes of aerobes and B. fragilis were 105 and 106 CFU/ml, respectively. The MIC was defined as the lowest antibiotic concentration that completely prevented visible growth after incubation at 37°C for 20 h.
The combined effect of J-110,441 with imipenem or ceftazidime was determined by the checkerboard method (29) under the same conditions as those for the MIC determination described above. To estimate synergism, the fractional inhibitory concentration (FIC) index was calculated according to the method of Elison et al. (9).
Preparation of β-lactamase.
IMP-1 metallo-β-lactamase was purified from P. aeruginosa GN17203 harboring the blaIMP gene. Cells were incubated at 4°C for 1 h in 10 mM 3-(N-morpholino)propanesulfonic acid (MOPS) buffer (pH 7.0), containing 27% sucrose and 2 mg of lysozyme (Sigma Chemical Co., St. Louis, Mo.) per ml, and then disrupted by sonication. The cellular debris was removed by centrifugation (16,000 × g, 15 min, 4°C), and supernatant was precipitated by an 80% saturated ammonium sulfate. This fraction was dialyzed against 10 mM MOPS buffer (pH 7.0) and applied to a CM-Sephadex C-50 column (Pharmacia Biotech AB, Uppsala, Sweden) equilibrated with 10 mM MOPS buffer (pH 7.0), and the enzyme was eluted with a linear NaCl gradient. The active fractions were pooled, dialyzed against 10 mM MOPS buffer (pH 7.0) containing 1 μM ZnCl2, and concentrated by ultrafiltration with a UK-10 ultrafilter (Advantec Toyo Co., Tokyo, Japan). The purity of the preparation was checked by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and the purified enzyme solution was stored at −80°C.
CcrA metallo-β-lactamase was prepared from B. fragilis GAI30079. Cells were suspended in 50 mM sodium phosphate buffer (pH 7.0) and disrupted by sonication. The cellular debris was removed by centrifugation (13,500 × g, 15 min, 4°C), and the supernatant was dialyzed against 50 mM sodium phosphate buffer (pH 7.8). This fraction was applied to a DEAE-Toyopearl 650M column (Toso Co., Tokyo, Japan) equilibrated with 50 mM sodium phosphate buffer (pH 7.8), and the enzyme was eluted with a linear NaCl gradient. The pooled active fractions were dialyzed against 50 mM sodium phosphate buffer (pH 7.0), concentrated by ultrafiltration with a UK-10 ultrafilter, and rechromatographed with a Sephadex G-100 column (Pharmacia Biotech AB) equilibrated with 50 mM sodium phosphate buffer (pH 7.0). The concentrated crude enzyme preparation was dialyzed against 50 mM sodium phosphate buffer (pH 7.0) and stored at −80°C.
L1 metallo-β-lactamase was prepared from Stenotrophomonas maltophilia GN12873 as described previously (30).
Type II metallo-β-lactamase from Bacillus cereus, TEM-1 penicillinase from Escherichia coli, and cephalosporinase from Enterobacter cloacae were obtained from Sigma Chemical Co. TEM-1 penicillinase and E. cloacae cephalosporinase correspond to group 2b and group 1 of Bush’s classification (3), respectively.
Determination of β-lactamase activity.
The activity of the metallo-β-lactamase preparation was determined at each step by monitoring the hydrolysis of 100 μM imipenem (Δɛ = 9.04 mM−1 · cm−1 at 299 nm) at 30°C in 10 mM MOPS buffer (pH 7.0) containing 100 μM ZnCl2. One unit of β-lactamase activity was defined as the amount of enzyme that hydrolyzed 1 μmol of imipenem per min at 30°C.
Determination of IC50.
The 50% inhibitory concentration (IC50) for IMP-1 metallo-β-lactamase was determined by measuring the enzymatic hydrolysis of a chromogenic cephalosporin, nitrocefin, in the presence of inhibitors. This automated assay system was a modification of a previously reported method (26). To avoid identifying metal chelators, 10 mM MOPS buffer (pH 7.0) containing 100 μM ZnCl2 was used in this microassay. Inhibitors were dissolved in 10 mM MOPS buffer (pH 7.0) or dimethyl sulfoxide at final concentrations of 0.1, 1.0, and 10 μM. After 1 μl of each inhibitor and 25 μl of IMP-1 metallo-β-lactamase (3 to 6 mU/ml) were mixed in a 98-well microplate, the assay was initiated within 1 min by the rapid addition of 75 μl of nitrocefin to create a final concentration of 72.7 μM. The reaction mixtures in the absence of inhibitor, enzyme, or both were prepared as the controls. Assay plates were incubated with slow shaking in an M-36 microincubator (Taitec Co., Tokyo, Japan) at 30°C, and the hydrolysis of nitrocefin was measured after incubation for 15 min by detecting the increase of absorbance at 492 nm in an MTP-120 plate reader (Corona Electric Co., Ibaraki, Japan). Under these conditions, substrate consumption in control experiments was <10% of initial concentration. The mean initial rates of hydrolysis at each inhibitor concentration were calculated from the triplicate measurements for each inhibitor concentration. The IC50s (micromolar concentrations) were determined by plotting percentages of inhibition against inhibitor concentrations.
β-Lactamase assays.
Kinetic studies were performed at 30°C in 10 mM MOPS buffer (pH 7.0), and the hydrolysis of the substrate was monitored in a UV-2200 temperature-controlled spectrophotometer (Shimadzu, Tokyo, Japan). The initial velocity of test compounds at a 100 μM concentration was determined, and velocities relative to those of imipenem and cephaloridine for metallo- and serine β-lactamases, respectively, were calculated. The molar extinction coefficient was as follows: cephaloridine, Δɛ = 10.2 mM−1 · cm−1 at 260 nm; J-110,441, Δɛ = 18.4 mM−1 · cm−1 at 306 nm; compound 3a, Δɛ = 6.37 mM−1 · cm−1 at 300 nm; and compound 7a, Δɛ = 10.5 mM−1 · cm−1 at 300 nm. Kinetic parameters were derived from at least two independent experiments by Hanes-Woolf plots of the initial velocity of substrate hydrolysis. To calculate the Ki values by Dixon plotting, rates of substrate hydrolysis at concentrations ranging from 10 to 100 μM were determined in the presence of various concentrations of inhibitors. Imipenem and cephaloridine were used for metallo- and serine β-lactamases, respectively. The assay was initiated by the addition of enzyme to the mixture of substrate and inhibitor. The total reaction volume was 1.0 ml in all cases.
RESULTS
Structure–inhibitory-activity relationship.
Several compounds have been identified as IMP-1 metallo-β-lactamase inhibitors in an automated microassay system using nitrocefin as a substrate. In this assay, IC50s of marketed β-lactam antibiotics such as panipenem, ampicillin, ceftazidime, cephaloridine, and aztreonam were over 100 μM. Meropenem and flomoxef showed weak inhibitory activity with IC50s of 25 and 60 μM, respectively. The IC50s of metal chelators such as dipicolinic acid (39) and EDTA were within the range of 80 to 200 μM. Cilastatin sodium, an inhibitor of renal membrane dipeptidase (dehydropeptidase I) (16) and a weak inhibitor of CphA metallo-β-lactamase from Aeromonas acidophila (17), did not show inhibitory activity against IMP-1 metallo-β-lactamase at the concentration of 10 μM.
The structure–inhibitory-activity relationship of J-110,441 having benzothienylthio moiety as a side chain and related analogs is summarized in Table 1. The most remarkable finding in this study was that the introduction of a 2-substituted benzothiophene at the C-2 position of 1β-methylcarbapenem, which resulted in J-110,441, led to strong inhibitory activity with an IC50 of <0.1 μM. Replacement of the 2-substituted benzothiophene with a 3-substituted benzothiophene (compound 1a), a benzothiazole (compound 2a), or a benzimidazole (compound 2b) resulted in decreased inhibitory activity. Introduction of the methyl ester to the carboxylic acid of the carbapenem nucleus (compound 1b) or insertion of a pyrrolidine ring as a spacer between the carbapenem nucleus and the benzothienylthio moiety (compound 1c) produced a marked reduction in inhibitory activity.
Dithiocarbamate carbapenems showed relatively good inhibitory activity; the IC50s of the compounds tested ranged from <0.1 to 1.8 μM, as shown in Table 2. Among the dithiocarbamate compounds tested, those of the 1,2,5,6-tetrahydropyridin-1-ylthiocarbonylthio series (compounds 3a to 3e) had better inhibitory activity than did the others (compounds 3f and 3g). The introduction of a cationic side chain (compare compounds 3a to 3d with compound 3e) produced an increase in inhibitory activity, especially in the case of dicationic 1,4-diazabicyclo[2,2,2]octane (DABCO) having a carbamoylmethyl or hydroxyethyl substituent in the side chain (compounds 3a and 3b), which had an IC50 below 0.1 μM. The carbamoylmethyl- or hydroxyethyl-substituted DABCO itself did not inhibit IMP-1 metallo-β-lactamase at the concentration of 10 μM.
TABLE 2.
Structure and IMP-1 metallo-β-lactamase-inhibitory activity of dithiocarbamate carbapenem analogs

| Compound | R | IC50 (μM)a |
|---|---|---|
| 3a |  |
<0.1 |
| 3b | <0.1 | |
| 3c | 0.22 | |
| 3d | 0.22 | |
| 3e |  |
0.50 |
| 3f | 1.4 | |
| 3g | 1.8 | |
Nitrocefin was used as a substrate.
The structure–inhibitory-activity relationship of pyrrolidinylthio carbapenem derivatives is shown in Table 3. The introduction of an aminomethyl group to the benzene ring (compare compounds 4a and 4b with 4c) and alkylation of the nitrogen of pyrrolidine with hydroxyethyl (compare compound 6a with 6b) brought about stronger inhibitory activity. An additional aminomethyl (compare compound 4a with 4b) increased inhibitory activity. A decrease in inhibitory activity was caused by (i) replacement of the aminomethyl group attached to the benzene ring with hydroxymethyl (compare compound 6b with 6d), (ii) substitution on the benzylic methylene position (compare compound 6b with 6c), (iii) introduction of a methyl group on the pyrrolidine ring (compare compound 5a with 5b), and (iv) use of cyclohexane instead of the benzene ring (compare compound 6b with 6e). The thiol side chain of compound 5a did not inhibit the enzyme at the concentration of 10 μM, although the IC50 of compound 5a was 0.7 μM (21). The important observation was that direct attachment of a benzene ring to pyrrolidine was necessary for inhibitory activity because the introduction of spacers between pyrrolidine and the benzene ring (compound 6f) or cleavage of the pyrrolidine ring (compound 6g) caused a decrease in inhibitory activity.
With respect to the stereochemistry of the pyrrolidine ring of compound 5a, the Ki values of the compounds with trans (compound 5a) and cis configurations were 0.18 and 0.12 μM, respectively. The results suggested that the C-5 stereochemistry of the pyrrolidine ring was not a crucial factor for the inhibitory potential for IMP-1 enzyme, while the inhibitory activity was markedly reduced by inversion of the C-3 chiral center of the pyrrolidine ring (IC50, >10 μM) regardless of C-5 stereochemistry.
It was noteworthy that replacement of the benzene ring with thiophene (compare compound 7a with 4b) showed increased inhibitory activity. Alkylation of the amino group did not affect the inhibitory activity (compound 7b); however, a 3-thienyl analog (compound 7c) was over 10 times less potent than the 2-thienyl compound (compound 7b).
Among the pyrrolidinylthio carbapenems tested, compound 7a was more stable under the hydrolysis by and more potent against various metallo-β-lactamases than was compound 4a or 6a.
Inhibitory activity against various β-lactamases.
Some compounds with an IC50 below 0.1 μM were selected and were characterized by studying their affinity for class B metallo-β-lactamases and class A and class C serine β-lactamases and their stability under hydrolysis by these enzymes (Table 4).
Among all the compounds tested, J-110,441, having a benzothienylthio moiety at the C-2 position of 1β-methylcarbapenem, was found to be the most stable under hydrolysis by all of the metallo- and serine β-lactamases tested in terms of relative hydrolysis rate, which ranged from <1 to 8% for IMP-1, CcrA, L1, and B. cereus type II metallo-β-lactamases and group 2b and group 1 serine β-lactamases. In particular, it is noted that J-110,441 exhibited better inhibitory activity than did the other inhibitors against metallo-β-lactamases, since the Ki values were 0.0037, 0.23, 1.00, and 0.83 μM for IMP-1 encoded on the transferable blaIMP gene, CcrA from B. fragilis, L1 from S. maltophilia, and type II from B. cereus, respectively. J-110,441 inhibited IMP-1 in a competitive manner (Fig. 1) and also showed inhibitory activity against class A and class C serine β-lactamases with Ki values of 2.54 and 0.037 μM, respectively. The inhibition by J-110,441 was reversible. Thus, J-110,441 was suggested to have potential as a new class of β-lactamase inhibitor with a broad spectrum.
FIG. 1.

Dixon plot consistent with competitive inhibition of IMP-1 by J-110,441. Imipenem was used as the substrate at the indicated concentration (conc.).
As shown in Table 4, compounds 3a and 7a also showed appreciable relative stability under hydrolysis by IMP-1 metallo-β-lactamase, which was similar to that of J-110,441, and the inhibitory activity against these enzymes was observed as Ki values of 0.09 and 0.01 μM, respectively. However, the compounds were more susceptible to hydrolysis by the CcrA and L1 enzymes than to that by the IMP-1 enzyme. Against class A and class C serine β-lactamases, the inhibitory activity of these compounds was inferior to that of J-110,441.
Combined effect on antimicrobial activity.
Table 5 presents the antimicrobial activities of J-110,441, imipenem, ceftazidime, and their combination against β-lactamase producers. The organisms that produce class B enzymes showed resistance to imipenem and ceftazidime, while E. cloacae, which is a derepressed producer of class C enzyme, remained susceptible to imipenem but showed high resistance to ceftazidime. The activity of imipenem or ceftazidime was potentiated in the presence of J-110,441. Especially against clinical isolates of S. marcescens producing IMP-1, the MICs of imipenem, ranging from 64 to 256 μg/ml, decreased to levels ranging from 4 to 64 μg/ml in the presence of one-fourth of the MIC of J-110,441. Against class C β-lactamase-producing E. cloacae, the MIC of ceftazidime decreased from 64 to 4 μg/ml in the presence of 4 μg of J-110,441 per ml. Synergy (FIC index, ≤0.5) of imipenem or ceftazidime with J-110,441 was observed against 9 of 11 strains tested that were resistant mainly due to the production of class B and/or class C β-lactamases. However, the results indicated that some strains are still clinically resistant to imipenem even in combination with J-110,441.
TABLE 5.
Potentiation of antimicrobial activity against β-lactamase producers
| Sp. and strain | Type of class B β-lactamase | MICa (μg/ml) of drug:
|
FIC indexb | ||||
|---|---|---|---|---|---|---|---|
| Imipenem alone | Ceftazidime alone | J-110,441 alone | Imipenem + 1/4 MIC of J-110,441 | Ceftazidime + 1/8 MIC of J-110,441 | |||
| S. marcescens | |||||||
| BB6231 | IMP-1 | 64 | >128 | 128 | 4 | 0.31 | |
| BB6232 | IMP-1 | 64 | 128 | 256 | 4 | 0.31 | |
| BB6233 | IMP-1 | 256 | >128 | 256 | 4 | 0.27 | |
| BB6234 | IMP-1 | 256 | >128 | 128 | 16 | 0.31 | |
| BB5886 | IMP-1 | 256 | >128 | 64 | 64 | 0.50 | |
| P. aeruginosa | |||||||
| BB6298 | IMP-1 | 16 | >128 | 128 | 4 | 0.50 | |
| GN17203 | IMP-1 | 128 | >128 | 128 | 64 | 0.75 | |
| S. maltophilia | |||||||
| BB6299 | L1 | 128 | >128 | 32 | 128 | 1.25 | |
| GN12873 | L1 | 128 | >128 | 64 | 32 | 0.50 | |
| B. fragilis GAI30079 | CcrA | 64 | >128 | 32 | 16 | 0.50 | |
| E. cloacae GN7471 | —c | 0.5 | 64 | 32 | 4 | 0.19 | |
Determined by broth microdilution method.
FIC index: ≤0.5, synergism; 1.0, additivity; >2.0, indifference or antagonism.
—, not produced.
DISCUSSION
The results described above show that a series of 1β-methylcarbapenems exhibited a variety of potencies against IMP-1 metallo-β-lactamase. The carbapenem nucleus probably played an important role in the affinity for IMP-1 enzyme, as suggested previously for B. fragilis metallo-β-lactamase (7). Introduction of a thiophene or DABCO moiety to the side chain brought about stronger inhibitory activity. Conversely, introduction of a spacer between the carbapenem nucleus and the side chain or into the side chain caused a marked decrease of inhibitory activity. In the case of 3,5-disubstituted pyrrolidinylthio-1β-methylcarbapenem, C-5 stereochemistry of the pyrrolidine ring was not a crucial factor for the inhibitory potential against IMP-1 enzyme, though it affected the antibacterial activity (33).
The most remarkable result was that J-110,441 was found to be an inhibitor of various β-lactamases, especially against transferable IMP-1 metallo-β-lactamase and class C serine β-lactamase with Ki values of 0.0037 and 0.037 μM, respectively. J-110,441 showed strong inhibitory activity against IMP-1 enzyme, but the activity decreased against other metalloenzymes; in particular, it was over 100-fold less active against the L1 and type II enzymes. These results may be ascribed to the diversity in metallo-β-lactamases (28).
A synergistic effect was observed between J-110,441 and imipenem or ceftazidime in the bacterial susceptibility study using clinical isolates of β-lactamase producers. Against class C serine β-lactamase-producing E. cloacae, the antimicrobial activity of ceftazidime was potentiated in the presence of J-110,441, although the possibility of compensatory inhibition of penicillin binding proteins by cephem and carbapenem antibiotics should not be excluded. The combined effects of imipenem and J-110,441 showed much variation among the blaIMP gene-carrying organisms tested. It might be explained by varied mechanisms of resistance in clinical isolates such as production of different enzymes including IMP-1 and chromosomal AmpC enzyme, low outer membrane permeability (porin deficiency), or efflux (22).
It has been reported elsewhere that the commercially available β-lactamase inhibitors are clinically ineffective against IMP-1 metallo-β-lactamase and many class C serine β-lactamases (3, 23). Also, the inhibitory activity of synthetic metallo-β-lactamase inhibitors is reportedly insufficient for development for clinical use (12, 17, 25, 34, 37). Needless to say, the spread of metallo-β-lactamase is a future threat to antimicrobial chemotherapy with β-lactams including carbapenems (28, 31, 38). The X-ray crystallography studies have revealed three-dimensional structures of B. cereus type II (6, 10), CcrA (7, 11), and L1 (35) to have two zinc atoms at the active sites of these enzymes, and the models were based on not only native enzyme crystal structures but those with substrate (7, 10, 35) or inhibitor (34) at the active site. Recently, two zinc atoms in IMP-1 enzyme were determined enzymatically (18). Because crystallographic information is still limited, further study is necessary for the computer modeling for designing effective inhibitors. Our present data yielded various pieces of information on the structure–inhibitory-activity relationship and could assist in the development of specific inhibitors of bacterial β-lactamases and/or an antibiotic stable against various β-lactamases.
REFERENCES
- 1.Ambler R P. The structure of β-lactamases. Philos Trans R Soc Lond B Biol Sci. 1980;289:321–331. doi: 10.1098/rstb.1980.0049. [DOI] [PubMed] [Google Scholar]
- 2.Arakawa Y, Murakami M, Suzuki K, Ito H, Wacharotayankun R, Ohsuka S, Kato N, Ohta M. A novel integron-like element carrying the metallo-β-lactamase gene blaIMP. Antimicrob Agents Chemother. 1995;39:1612–1615. doi: 10.1128/aac.39.7.1612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bush K. Characterization of β-lactamases. Antimicrob Agents Chemother. 1989;33:259–263. doi: 10.1128/aac.33.3.259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bush K, Macalintal C, Ramussen B A, Lee V J, Yang Y. Kinetic interactions of tazobactam with β-lactamases from all major structural classes. Antimicrob Agents Chemother. 1993;37:851–858. doi: 10.1128/aac.37.4.851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bush K, Jacoby G A, Medeiros A A. A functional classification scheme for β-lactamases and its correlation with molecular structure. Antimicrob Agents Chemother. 1995;39:1211–1233. doi: 10.1128/aac.39.6.1211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Carfi A, Dunee E, Galleni M, Frere J M, Dideberg O. Metallo-beta-lactamase from Bacillus cereus 569h9. 1997. Brookhaven Protein Data Bank entry 1BME. [Google Scholar]
- 7.Concha N O, Ramussen B A, Bush K, Herzberg O. Crystal structure of the wide-spectrum binuclear zinc β-lactamase from Bacteroides fragilis. Structure. 1996;4:823–836. doi: 10.1016/s0969-2126(96)00089-5. [DOI] [PubMed] [Google Scholar]
- 8.Cornagalia G, Riccio M L, Mazzariol A, Lauretti L, Fontana R, Rossolini G M. Appearance of IMP-1 metallo-β-lactamase in Europe. Lancet. 1999;353:899–900. doi: 10.1016/s0140-6736(98)05954-6. [DOI] [PubMed] [Google Scholar]
- 9.Elison G B, Singer S, Hitchings C H. Antagonists of nucleic acid derivatives. VIII. Synergism in combination of biochemically related antimetabolites. J Biol Chem. 1954;208:477–488. [PubMed] [Google Scholar]
- 10.Fabiane S M, Sofi M K, Wan T, Payne D J, Bateson J H, Mitchell T, Sutton B J. Crystal structure of zinc-dependent β-lactamase from Bacillus cereus at 1.9A resolution: binuclear active site with features of a mononuclear enzyme. Biochemistry. 1998;37:12404–12411. doi: 10.1021/bi980506i. [DOI] [PubMed] [Google Scholar]
- 11.Fitzgerald P M D, Wu J K, Toney J H. Unanticipated inhibition of the metallo-β-lactamase from Bacteroides fragilis by 4-morpholineethanesulfonic acid (MES): a crystallographic study at 1.85-A resolution. Biochemistry. 1998;37:6791–6800. doi: 10.1021/bi9730339. [DOI] [PubMed] [Google Scholar]
- 12.Gilpin M L, Fulston M, Payne D, Cramp R, Hood I. Isolation and structure determination of two novel phenazines from a Streptomyces with inhibitory activity against metallo-enzymes, including metallo-β-lactamase. J Antibiot. 1995;48:1081–1085. doi: 10.7164/antibiotics.48.1081. [DOI] [PubMed] [Google Scholar]
- 13.Hashizume, T., and R. Nagano. August 1998. Metallo-β-lactamase inhibitors. Japan patent 10-256065.
- 14.Imamura, H., N. Ohtake, A. Shimizu, H. Sato, Y. Sugimoto, S. Sakuraba, H. Kiyonaga, C. Suzuki-Sato, M. Nakano, R. Nagano, K. Yamada, T. Hashizume, and H. Morishima. Novel trans-3,5-disubstituted pyrrolidinylthio-1β-methylcarbapenems. Antibacterial activities of J-111,225, J-114,870, J-114,871 and their related compounds. Submitted for publication. [DOI] [PubMed]
- 15.Ito H, Arakawa Y, Ohsuka S, Wacharotayankun R, Kato N, Ohta M. Plasmid-mediated dissemination of the metallo-β-lactamase gene blaIMP among clinically isolated strains of Serratia marcescens. Antimicrob Agents Chemother. 1995;39:824–829. doi: 10.1128/aac.39.4.824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kahan F M, Kropp H, Sundelof J G, Birnbaum J. Thienamycin: development of imipenem-cilastatin. J Antimicrob Chemother. 1983;12(Suppl. D):1–35. doi: 10.1093/jac/12.suppl_d.1. [DOI] [PubMed] [Google Scholar]
- 17.Keynan S, Hooper N M, Felici A, Amicosante G, Turner A J. The renal membrane dipeptidase (dehydropeptidase I) inhibitor, cilastatin, inhibits the bacterial metallo-β-lactamase enzyme CphA. Antimicrob Agents Chemother. 1995;39:1629–1631. doi: 10.1128/aac.39.7.1629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Laraki N, Franceschini N, Rossolini G M, Santucci P, Meunier C, Pauw E D, Amicosante G, Frere J M, Galleni M. Biochemical characterization of the Pseudomonas aeruginosa 101/1477 metallo-β-lactamase IMP-1 produced by Escherichia coli. Antimicrob Agents Chemother. 1999;43:902–906. doi: 10.1128/aac.43.4.902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lee K, Chong Y, Shin H B, Yong D. Abstracts of the 38th Interscience Conference on Antimicrobial Agents and Chemotherapy. Washington, D.C: American Society for Microbiology; 1998. Rapid increase of imipenem-hydrolyzing Pseudomonas aeruginosa in a Korean hospital, abstr. E-85; p. 193. [Google Scholar]
- 20.Livermore D M. Acquired carbapenemases. J Antimicrob Chemother. 1997;39:673–676. doi: 10.1093/jac/39.6.673. [DOI] [PubMed] [Google Scholar]
- 21.Nagano R, Adachi Y, Hashizume T, Morishima H. Abstracts of the 38th Interscience Conference on Antimicrobial Agents and Chemotherapy. Washington, D.C: American Society for Microbiology; 1998. J-111,225—a novel trans-3,5-disubstituted pyrrolidinylthio-1β-methyl-carbapenem: inhibitor of IMP-1 metallo-β-lactamase, abstr. F-55; p. 247. [Google Scholar]
- 22.Nikaido H. Prevention of drug access to bacterial targets: permeability barriers and active efflux. Science. 1994;264:382–388. doi: 10.1126/science.8153625. [DOI] [PubMed] [Google Scholar]
- 23.Ohsuka S, Arakawa Y, Norii T, Ito H, Ohta M. Effect of pH on activities of novel β-lactamases and β-lactamase inhibitors against these β-lactamases. Antimicrob Agents Chemother. 1995;39:1856–1858. doi: 10.1128/aac.39.8.1856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ohtake N, Imamura H, Jona H, Kiyonaga H, Shimizu A, Moriya M, Sato S, Nakano M, Ushijima R, Nakagawa S. Novel dithiocarbamate carbapenems with anti-MRSA activity. Bioorg Med Chem. 1998;6:1089–1101. doi: 10.1016/s0968-0896(98)00069-8. [DOI] [PubMed] [Google Scholar]
- 25.Payne D J, Bateson J H, Gasson B C, Proctor D, Khushi T, Farmer T H, Tolson D A, Bell D, Skett P W, Marshall A C, Reid R, Ghosez L, Combret Y, Brynaert J-M. Inhibition of metallo-β-lactamases by a series of mercaptoacetic acid thiol ester derivatives. Antimicrob Agents Chemother. 1997;41:135–140. doi: 10.1128/aac.41.1.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Payne D J, Coleman K, Cramp R. The automated in vitro assessment of β-lactamase inhibitors. J Antimicrob Chemother. 1991;28:775–776. doi: 10.1093/jac/28.5.775. [DOI] [PubMed] [Google Scholar]
- 27.Prosperi-Meys C, Llabres G, de Seny D, Soto R P, Valladares M H, Laraki N, Frere J-M, Galleni M. Interaction between class B β-lactamases and suicide substrates of active-site serine β-lactamases. FEBS Lett. 1999;443:109–111. doi: 10.1016/s0014-5793(98)01689-5. [DOI] [PubMed] [Google Scholar]
- 28.Rasmussen B A, Bush K. Carbapenem-hydrolyzing β-lactamases. Antimicrob Agents Chemother. 1997;41:223–232. doi: 10.1128/aac.41.2.223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sabath L D. Synergy of antimicrobial substances by apparently known mechanisms. 1968. pp. 210–217. . Antimicrob. Agents Chemother. 1967. [PubMed] [Google Scholar]
- 30.Saino Y, Kobayashi F, Inoue M, Mitsuhashi S. Purification and properties of inducible penicillin β-lactamase isolated from Pseudomonas maltophilia. Antimicrob Agents Chemother. 1982;22:564–570. doi: 10.1128/aac.22.4.564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Senda K, Arakawa Y, Nakashima K, Ito H, Ichikawa S, Shimokawa K, Kato N, Ohta M. Multifocal outbreaks of metallo-β-lactamase-producing Pseudomonas aeruginosa resistant to broad-spectrum β-lactams, including carbapenems. Antimicrob Agents Chemother. 1996;40:349–353. doi: 10.1128/aac.40.2.349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Senda K, Arakawa Y, Ichiyama S, Nakashima K, Ito H, Ohsuka S, Shimokata K, Kato N, Ohta M. PCR detection of metallo-β-lactamase gene (blaIMP) in gram-negative rods resistant to broad-spectrum β-lactams. J Clin Microbiol. 1996;34:2909–2913. doi: 10.1128/jcm.34.12.2909-2913.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shimizu A, Sugimoto H, Sakuraba S, Imamura H, Sato H, Ohtake N, Ushijima R, Nakagawa S, Suzuki C, Hashizume T, Morishima H. Abstracts of the 38th Interscience Conference on Antimicrobial Agents and Chemotherapy. Washington, D.C: American Society for Microbiology; 1998. Novel trans-3,5-disubstituted pyrrolidinylthio-1-methylcarbapenems with potent activity against MRSA and Pseudomonas aeruginosa, abstr. F-52; p. 246. [Google Scholar]
- 34.Toney J H, Fitzgerald P M, Grover-Sharma N, Olson S H, May W J, Sundelof J G, Vanderwall D E, Cleary K A, Grant S K, Wu J K, Kozarich J W, Pompliano D L, Hammond G G. Antibiotic sensitization using biphenyl tetrazoles as potent inhibitors of Bacteroides fragilis metallo-β-lactamase. Chem Biol. 1998;5:185–196. doi: 10.1016/s1074-5521(98)90632-9. [DOI] [PubMed] [Google Scholar]
- 35.Ullah J, Walsh T R, Tailor I A, Emery D C, Gamblin S J, Spencer J. The crystal structure of the L1 metallo-β-lactamase from Stenotrophomonas maltophilia at 1.7Å resolution. J Mol Biol. 1998;284:125–136. doi: 10.1006/jmbi.1998.2148. [DOI] [PubMed] [Google Scholar]
- 36.Waddell S T, Ratcliffe R W, Szumiloski S P, Wildonger K J, Wilkening R R, Blizzard T A, Huber J, Kohler J, Dorso K, St. Rose E, Sundelof J G, Hammond G G. Benzothiazolylthio carbapenems: potent anti-MRSA agents. Bioorg Med Chem Lett. 1995;5:1427–1432. [Google Scholar]
- 37.Walter M W, Felici A, Galleni M, Soto R P, Adlington R M, Baldwin J E, Frere J-M, Gololobov M, Shofield C J. Trifluoromethyl alcohol and ketone inhibitors of metallo-β-lactamases. Bioorg Med Chem. 1996;6:2455–2458. [Google Scholar]
- 38.Watanabe S, Iyobe D, Inoue M, Mitsuhashi S. Transferable imipenem resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 1991;35:147–151. doi: 10.1128/aac.35.1.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yang Y, Roll D M, Wildey M J, Lee M, Greenstein M, Maiese W M, Bush K. Abstracts of the 34th Interscience Conference on Antimicrobial Agents and Chemotherapy. Washington, D.C: American Society for Microbiology; 1994. Inhibition of metallo-β-lactamases by LL-10G568α and LL-10G568β, abstr. C56; p. 88. [Google Scholar]