

Abstract
Mutations of the rpoB gene conferring resistance to rifampin were analyzed in 40 methicillin-resistant Staphylococcus aureus isolates obtained from six countries. Interestingly, the majority of clinical isolates showed multiple mutations within rpoB. The amino acid substitution 481His→Asn was the most prevalent one, capable of conferring low-level resistance on its own. Cross-resistance to rifampin, rifabutin, and rifapentine was demonstrated for all mutants identified. The level of resistance to rifamycins correlated with both the mutation position and type of amino acid substitution.
Multiresistance as a common feature in methicillin-resistant Staphylococcus aureus (MRSA) is a growing problem not only in hospital settings (5, 10, 22). Glycopeptides are the antibiotics of choice in the treatment of infections caused by MRSA (15). A combination therapy, however, with an antibiotic such as rifampin that reveals strong activity and good tissue penetration often is required to reach deep-seated infections effectively (6, 9, 16). Rifampin acts by inhibiting bacterial RNA polymerase (23). Previous studies on other bacteria provide evidence that mutations in rpoB, the gene which encodes the β subunit of RNA polymerase, are responsible for rifampin resistance (Rifr) (2, 8, 12, 17, 18, 20). By using pairs of isogenic S. aureus isolates gathered before and after acquisition of resistance under rifampin therapy, Aubry-Damon et al. recently demonstrated the occurrence of amino acid substitutions within a short conserved region of the β subunit and correlated the level of rifampin resistance with the mutations involved (2).
The present study was aimed at determining the distribution of mutations in the rpoB gene in clinical isolates of rifampin-resistant MRSA as well as correlating the MICs of rifampin, rifabutin, and rifapentine with the locations and nature of the amino acid substitutions.
A total of 35 Rifr MRSA clinical isolates and five in vitro mutants generated from two rifampin-susceptible (Rifs) epidemic MRSA strains were analyzed in this study. Fifteen of the Rifr MRSA isolates were furnished by laboratories in the United States, France, Italy, Poland, and Slovenia, whereas 20 of the Rifr initial isolates were obtained between 1993 and 1998 from German hospitals. All MRSA isolates were analyzed by pulsed-field gel electrophoresis as described previously (24). Resistance to rifampin (Sigma, Deisenhofen, Germany), rifabutin (Pharmacia-Upjohn, Milan, Italy), and rifapentine (Hoechst Marion Roussel, Kansas, Mo.) was determined by the agar dilution method with Mueller-Hinton agar (Oxoid, Basingstoke, England) with an inoculum of 104 CFU per spot. Two oligonucleotide primers (Life Technologies, Eggenstein, Germany), rpoB1 (5′-ACC GTC GTT TAC GTT CTG TA) and rpoB2 (5′-TCA GTG ATA GCA TGT GTA TC), were designed to amplify and sequence a 460-bp PCR fragment encompassing clusters I and II of the rifampin resistance mutation sites of the S. aureus rpoB gene (1). Amplification was performed on a Gene Amp PCR system 2400 (Perkin-Elmer, Weiterstadt, Germany) under standard conditions. Direct sequencing of purified PCR products was carried out by using a dye reaction terminator cycle sequencing kit (PE Applied Biosystems, Weiterstadt, Germany) according to the protocol described by the manufacturer. Amplified DNA was sequenced in both directions by using the 310 genetic analyzer (Perkin-Elmer).
All MRSA isolates were characterized with regard to their genotypes and the MICs of rifampin, rifabutin, and rifapentine as shown in Table 1. Comparable MICs of rifampin and rifapentine could be demonstrated for all MRSA isolates, whereas the MICs of rifabutin generally were two to eight times lower. The degree of resistance allowed classification of the strains in the categories of low-level resistance (MICs, 1 to 4 μg/ml) and high-level resistance (MICs, ≥8 μg/ml). Restriction analysis of chromosomal DNA demonstrated the unrelatedness of the majority of isolates and revealed 19 different genotypes, with MRSA T38 and T23 representing the southern and northern German epidemic strains, respectively (Fig. 1). Sequence analysis of 40 MRSA isolates from six countries revealed missense mutations in a short region of the rpoB gene equivalent to clusters I and II of Escherichia coli (18) (Fig. 2). Twelve mutational changes at 10 positions were identified, with 473Ala→Thr representing a new mutation site. New amino acid substitutions, 465Gln→Arg, 466Leu→Ser, 468Gln→Lys, and 477Ala→Thr in cluster I and 527Ile→Met and 529Ser→Leu in cluster II, were described, thereby emphasizing the high variability of these amino acid positions (2, 4, 7, 8, 18–21). Sequence findings allowed the categorization of all of the rifampin-resistant MRSA isolates into 12 different genotypes with respect to the mutations involved (Table 2). Despite the different geographical origins of the isolates (Table 1), codon 481 was mutated on 32 separate occasions, which indicates a central role of this amino acid. All in vivo isolates that demonstrated two or three amino acid changes exhibited high-level resistance. Interestingly enough, all of these isolates showed the mutational change 481His→Asn, which is capable of conferring low-level resistance on its own, thereby indicating a two-step resistance mechanism in vivo to high-level resistance within these isolates. High-level resistance in vivo, however, was not demonstrated to occur through multiple mutations alone. The single amino acid substitution 468Gln→Lys also causes high-level resistance.
TABLE 1.
Strain characterization
| Strain | Origin | Genotype | MIC (μg/ml) of:
|
||
|---|---|---|---|---|---|
| Rifampin | Rifabutin | Rifapentine | |||
| T236 | United States | K | 2 | 1 | 2 |
| T248 | France | C | 256 | 32 | 256 |
| T249 | France | B | 256 | 32 | 256 |
| T250 | France | B | 256 | 32 | 256 |
| T262 | Slovenia | I | 512 | 64 | 512 |
| T264 | Poland | J | 512 | 64 | 256 |
| T295 | Poland | J | 256 | 64 | 256 |
| T296 | Poland | J | 256 | 64 | 256 |
| T297 | Poland | J | 256 | 64 | 256 |
| T397 | Italy | S | 512 | 512 | 256 |
| T398 | Italy | S | 128 | 64 | 128 |
| T399 | Italy | Q | 256 | 64 | 256 |
| T400 | Italy | R | 256 | 128 | 256 |
| T401 | Italy | B | 512 | 64 | 512 |
| T402 | Italy | B | 128 | 32 | 128 |
| T4 | Germany | A | 2 | 1 | 2 |
| T20 | Germany | B | 512 | 64 | 512 |
| T25 | Germany | B | 1 | 1 | 2 |
| T36 | Germany | L | 512 | 64 | 512 |
| T46 | Germany | E | 1,024 | 128 | 512 |
| T59 | Germany | M | 512 | 128 | 1,024 |
| T66 | Germany | H | 512 | 128 | 512 |
| T109 | Germany | D | >1,024 | 512 | 512 |
| T112 | Germany | B | 256 | 64 | 256 |
| T113 | Germany | D | 512 | 512 | 256 |
| T115 | Germany | N | 2 | 1 | 2 |
| T118 | Germany | G | 256 | 64 | 256 |
| T124 | Germany | F | 512 | 256 | 256 |
| T161 | Germany | O | 256 | 32 | 256 |
| T166 | Germany | H | 512 | 128 | 512 |
| T211 | Germany | P | 2 | 1 | 2 |
| T212 | Germany | B | 256 | 32 | 256 |
| T382 | Germany | D | 2 | 1 | 2 |
| T403 | Germany | D | 2 | 1 | 2 |
| T404 | Germany | D | 2 | 1 | 2 |
| T38 | Germany | D | ≤0.008 | ≤0.008 | ≤0.008 |
| T38aa | This study | D | 512 | 256 | 512 |
| T38bb | This study | D | 512 | 512 | 512 |
| T23 | Germany | B | ≤0.008 | ≤0.008 | ≤0.008 |
| T23aa | This study | B | 4 | 2 | 4 |
| T23aac | This study | B | >1,024 | 512 | 512 |
| T23bb | This study | B | 1,024 | 256 | 512 |
Rifampin concentration of 0.0625 μg/ml used for selection of mutants from Rifs MRSA.
Rifampin concentration of 64 μg/ml used for selection of mutants from Rifs MRSA.
Rifampin concentration of 64 μg/ml used for selection of mutants from T23a.
FIG. 1.

Pulsed-field gel electrophoresis patterns of SmaI digests of total DNA from representatives of each pulsotype (indicated by capital letters above lanes). Lanes m, size markers. Molecular sizes (in kilobases) are indicated on the right.
FIG. 2.

Alignment of E. coli, Mycobacterium tuberculosis, and S. aureus rpoB sequences representing clusters I and II (1–3, 11, 14, 25, 26). The amino acid alignment is presented in a single-letter code. Asterisks symbolize identity to E. coli sequence. Positions involved in rifampin resistance are marked; mutations are indicated by downward-pointing arrows; insertions are indicated by upside-down triangles; and deletions are underlined. The new Rifr mutation site found in this study is indicated by a double downward-pointing arrow. Amino acid substitutions involved in Rifr found in this study are presented in italics. Amino acid substitutions not previously described are indicated in boldface.
TABLE 2.
Correlation of mutations in the rpoB gene and the level of resistance to rifampin
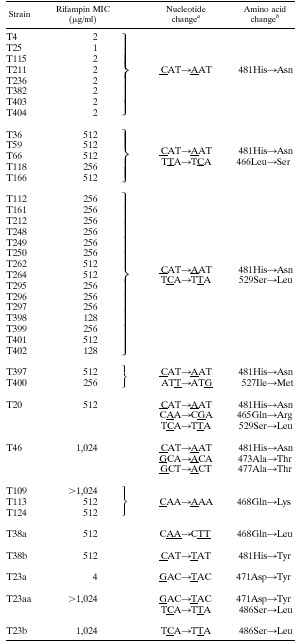 |
The mutations listed are for all strains within a grouping. Base changes are underlined.
The amino acid changes listed are for all strains within a grouping.
Generation of Rifr mutants in vitro resulted in both of the phenotypes observed. Low-level resistance followed by a second mutation leading to high-level resistance also was demonstrated in vitro (Table 2). Rifs strain T23 cultured on 0.0625 μg of rifampin-supplemented agar per ml revealed a new low-level resistant mutant with the amino acid substitution 471Asp→Tyr (T23a). When, in turn, T23a was plated on 64 μg of rifampin per ml, a new mutant readily acquired a second mutation, 486Ser→Leu, resulting in high-level resistance (T23aa). Rifs strain T38 was cultured on rifampin-supplemented agar at concentrations of 0.0625 and 64 μg/ml, and two single high-level resistance mutations were found with 468Gln→Lys (T38a) and 481His→Tyr (T38b), respectively. A substitution at amino acid position 481 within isolates T38b and T382 conferred low- or high-level resistance depending on the nature of the new amino acid (Table 2).
With respect to the pharmacokinetics of rifampin (27) and the findings presented in this study, we agree with the suggestion made by Aubry-Damon et al. (2) to revise the breakpoints for rifampin set by the National Committee for Clinical Laboratory Standards (13). Accordingly, new breakpoints of ≤0.5 and ≥8 μg/ml for the categorization of S. aureus relative to rifampin seem reasonable.
The goal of the present study was to elucidate the distribution of mutations within the rpoB gene in rifampin-resistant MRSA and to assess the in vitro effectiveness of different rifamycins against these isolates. By presenting seven new mutations, the study confirms that rpoB mutations are responsible for the common Rifr phenotype in MRSA and that the level of resistance to any rifamycin is dependent on both the type and the location of the mutation within the rpoB gene. The occurrence of multiple mutations within rpoB for S. aureus is first described in this study. Their presence might be explained by the epidemic nature of many MRSA strains and, consequently, the frequent exposure of these strains to rifampin chemotherapy. Finally, we were able to demonstrate that the mutations involved confer cross-resistance to rifampin, rifabutin, and rifapentine, thereby indicating that these antibiotics are likely to exhibit comparable effectiveness in the treatment of S. aureus infection.
Acknowledgments
We thank Denia Franck and Michael Stappenbeck for their technical assistance as well as Sebastian Walpen for sequencing support.
REFERENCES
- 1.Aboshkiwa M, Rowland G, Coleman G. Nucleotide sequence of the Staphylococcus aureus RNA polymerase rpoB gene and comparison of its predicted amino acid sequence with those of other bacteria. Biochim Biophys Acta. 1995;1262:73–78. doi: 10.1016/0167-4781(95)00054-k. [DOI] [PubMed] [Google Scholar]
- 2.Aubry-Damon H, Soussy C J, Courvalin P. Characterization of mutants in the rpoB gene that confer rifampin resistance in Staphylococcus aureus. Antimicrob Agents Chemother. 1998;42:2590–2594. doi: 10.1128/aac.42.10.2590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bodmer T, Zürcher G, Imboden P, Telenti A. Mutation position and type of substitution in the β-subunit of the RNA polymerase influence in-vitro activity of rifamycins in rifampicin-resistant Mycobacterium tuberculosis. J Antimicrob Chemother. 1995;35:345–348. doi: 10.1093/jac/35.2.345. [DOI] [PubMed] [Google Scholar]
- 4.Carter P E, Abadi F J R, Yakubu D E, Pennington T H. Molecular characterization of rifampicin-resistant Neisseria meningitidis. Antimicrob Agents Chemother. 1994;38:1256–1261. doi: 10.1128/aac.38.6.1256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chambers H F. Methicillin resistance in staphylococci: molecular and biochemical basis and clinical implications. Clin Microbiol Rev. 1997;10:781–791. doi: 10.1128/cmr.10.4.781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fujii K, Tsuji A, Miyazaki S, Yamaguchi K, Goto S. In vitro and in vivo antibacterial activities of KRM-1648 and KRM-1657, new rifamycin derivatives. Antimicrob Agents Chemother. 1994;38:1118–1122. doi: 10.1128/aac.38.5.1118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Honore N, Cole S T. Molecular basis of rifampin resistance in Mycobacterium leprae. Antimicrob Agents Chemother. 1993;37:414–418. doi: 10.1128/aac.37.3.414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jin D J, Gross C A. Mapping and sequencing of mutations in the Escherichia coli rpoB gene that lead to rifampicin resistance. J Mol Biol. 1988;202:45–58. doi: 10.1016/0022-2836(88)90517-7. [DOI] [PubMed] [Google Scholar]
- 9.Kunin C M. Antimicrobial activity of rifabutin. Clin Infect Dis. 1996;22:S3–S14. doi: 10.1093/clinids/22.supplement_1.s3. [DOI] [PubMed] [Google Scholar]
- 10.Michel M, Gutmann L. Methicillin-resistant Staphylococcus aureus and vancomycin-resistant enterococci: therapeutic realities and possibilities. Lancet. 1997;349:1901–1906. doi: 10.1016/s0140-6736(96)11192-2. [DOI] [PubMed] [Google Scholar]
- 11.Miller L P, Crawford J T, Shinnick T M. The rpoB gene of Mycobacterium tuberculosis. Antimicrob Agents Chemother. 1994;38:805–811. doi: 10.1128/aac.38.4.805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Morrow T O, Harmon S A. Genetic analysis of Staphylococcus aureus RNA polymerase mutants. J Bacteriol. 1979;137:374–383. doi: 10.1128/jb.137.1.374-383.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.National Committee for Clinical Laboratory Standards. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically. 4th ed. 1997. Approved standard M7-A4. National Committee for Clinical Laboratory Standards, Wayne, Pa. [Google Scholar]
- 14.Ovchinnikov Y A, Monastyrskaya G S, Gubanov V V, Guryev S O, Chertov O Y, Modyanov N N, Grinkevich V A, Makarova I A, Marchenko T V, Polovnikova I N, Lipkin V M, Sverdlov E D. The primary structure of Escherichia coli RNA polymerase. Eur J Biochem. 1981;116:621–629. doi: 10.1111/j.1432-1033.1981.tb05381.x. [DOI] [PubMed] [Google Scholar]
- 15.Rahman M. Alternatives to vancomycin in treating methicillin-resistant Staphylococcus aureus infections. J Antimicrob Chemother. 1998;41:325–328. doi: 10.1093/jac/41.3.325. [DOI] [PubMed] [Google Scholar]
- 16.Schmitz F J, Jones M E. Antibiotics for treatment of infections caused by MRSA and elimination of MRSA carriage. What are the choices? Int J Antimicrob Agents. 1997;9:1–19. doi: 10.1016/s0924-8579(97)00027-7. [DOI] [PubMed] [Google Scholar]
- 17.Severinov K, Mustaev A, Severinova E, Kozlov M, Darst S A, Goldfarb A. The β-subunit rif-cluster I is only angstroms away from the active center of Escherichia coli RNA polymerase. J Biol Chem. 1995;270:29428–29432. doi: 10.1074/jbc.270.49.29428. [DOI] [PubMed] [Google Scholar]
- 18.Severinov K, Soushko M, Goldfarb A, Nikiforov V. Rifampicin region revisited. J Biol Chem. 1993;268:14820–14825. [PubMed] [Google Scholar]
- 19.Severinov K, Soushko M, Goldfarb A, Nikiforov V. RifR mutations in the beginning of the Escherichia coli rpoB gene. Mol Gen Genet. 1994;244:120–126. doi: 10.1007/BF00283512. [DOI] [PubMed] [Google Scholar]
- 20.Taniguchi H, Aramaki H, Nikaido Y, Mizuguchi Y, Nakamura M, Koga T, Yoshida S. Rifampicin resistance and mutation of the rpoB gene in Mycobacterium tuberculosis. FEMS Microbiol Lett. 1996;144:103–108. doi: 10.1111/j.1574-6968.1996.tb08515.x. [DOI] [PubMed] [Google Scholar]
- 21.Telenti A, Imboden P, Marchesi F, Lowrie D, Cole S, Colston M J, Matter L, Schopfer K, Bodmer T. Detection of rifampicin-resistance mutations in Mycobacterium tuberculosis. Lancet. 1993;341:647–650. doi: 10.1016/0140-6736(93)90417-f. [DOI] [PubMed] [Google Scholar]
- 22.Voss A, Milatovic D, Wallrauch-Schwarz C, Rosdahl V T, Braveny I. Methicillin-resistant Staphylococcus aureus in Europe. Eur J Clin Microbiol Infect Dis. 1994;13:50–55. doi: 10.1007/BF02026127. [DOI] [PubMed] [Google Scholar]
- 23.Wehrli W. Rifampin: mechanisms of action and resistance. Rev Infect Dis. 1983;5:S407–S411. doi: 10.1093/clinids/5.supplement_3.s407. [DOI] [PubMed] [Google Scholar]
- 24.Wichelhaus T A, Schulze J, Hunfeld K P, Schäfer V, Brade V. Clonal heterogeneity, distribution, and pathogenicity of methicillin-resistant Staphylococcus aureus. Eur J Clin Microbiol Infect Dis. 1997;16:893–897. doi: 10.1007/BF01700555. [DOI] [PubMed] [Google Scholar]
- 25.Williams D L, Spring L, Collins L, Miller L P, Heifets L B, Gangadharam P R J, Gillis T P. Contribution of rpoB mutations to development of rifamycin cross-resistance in Mycobacterium tuberculosis. Antimicrob Agents Chemother. 1998;42:1853–1857. doi: 10.1128/aac.42.7.1853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yang B, Koga H, Ohno H, Ogawa K, Fukuda M, Hirakata Y, Maesaki S, Tomono K, Tashiro T, Kohno S. Relationship between antimycobacterial activities of rifampicin, rifabutin and KRM-1648 and rpoB mutations of Mycobacterium tuberculosis. J Antimicrob Chemother. 1998;42:621–628. doi: 10.1093/jac/42.5.621. [DOI] [PubMed] [Google Scholar]
- 27.Yao J D C, Moellering R C. Antibacterial agents. In: Murray P R, Baron E J, Pfaller M A, Tenover F C, Yolken R H, editors. Manual of clinical microbiology. 6th ed. Washington, D.C.: American Society for Microbiology; 1995. pp. 1281–1307. [Google Scholar]