

Executive summary
The multi-functional, hypothalamic hypocretin/orexin (HCRT) producing neurons regulate an array of physiological and behavioral states including arousal, sleep, feeding, emotions, stress, and reward. How a presumably uniform HCRT neuron population regulates such a diverse set of functions is not clear. The role of the HCRT neuropeptides may vary depending on the timing and localization of secretion and neuronal activity. Moreover, HCRT neuropeptides may not mediate all functions ascribed to HCRT neurons. Some could be orchestrated by additional neurotransmitters and neuropeptides that are expressed in HCRT neurons. We hypothesize that HCRT neurons are segregated into genetically-, anatomically- and functionally-distinct subpopulations. We discuss accumulating data that suggest the existence of such HCRT neuron subpopulations that may effectuate the diverse functions of these neurons in mammals and fish.
Keywords: hypocretin, orexin, zebrafish, heterogeneity, sleep
Introduction
The hypothalamus region in the brain regulates diverse physiological and behavioral processes such as arousal, sleep, feeding, reproduction and autonomic control, by integrating external and internal stimuli [1]. Among the millions of neurons in the hypothalamus, a group of only a few thousand hypocretin/orexin (HCRT) producing neurons stands out as a key regulator of sleep and wakefulness. HCRT neurons secrete the HCRT1 and HCRT2 neuropeptides and are located in the lateral hypothalamus area (LHA) and the adjacent perifornical area (PeFLH) [2,3]. In mice, approximately 5,000 HCRT neurons project to wide areas in the brain, including the tuberomammillary nucleus (TMN), paraventricular thalamic nucleus, arcuate nucleus, and monoaminergic nuclei, which express the two G-protein coupled receptors HCRTR1 and HCRTR2 [4–6]. In accordance with the broad anatomical projections, the function of HCRT neurons includes primarily the regulation of the sleep/wake cycle, but also that of feeding, energy homeostasis, emotion, stress response, motivation, and reward.
Now, how could one type of neuron defined through the expression of HCRT peptides serve as a multi-functional hypothalamic system supporting such diverse physiological processes? The secretion of HCRT neuropeptides at different effector sites in the brain could, for instance, be differentially regulated. The differential and spatially restricted release of HCRT1 and HCRT2, and the different signaling through differentially expressed HCRTR1 and HCRTR2 could be another determining factor. In addition, HCRT neurons also contain other neurotransmitters and proteins that may be involved in regulating their development, plasticity and diverse functions.
Soon after the discovery of the two HCRT neuropeptides [2,3], it was found that loss of HCRT signaling, either by loss of the ligand or receptor, results in the sleep disorder narcolepsy in humans, and causes narcolepsy-like symptoms in animals [7–9]. Narcolepsy is characterized by fragmented sleep and wakefulness during day and night, and a signature symptom; cataplexy – a sudden skeletal muscle atonia during wakefulness, which is triggered by emotional stimuli [10]. Several discoveries have linked the HCRT system to narcolepsy, including the isolation of mutated HCRTR2 in narcoleptic dogs [7], the narcolepsy-like phenotype of HCRT knockout mice [2,8], the detection of low HCRT protein levels in narcoleptic patients [9], and the severe loss of HCRT neurons in the brains of narcoleptic patients [11–13]. This tight association between the HCRT system, narcolepsy, and sleep disturbances established the HCRT system as a key regulator of arousal and sleep/wake transitions.
The HCRT system is unique to vertebrates, although recent work has characterized a functional HCRT neuropeptide in the basal chordate, amphioxus [14]. Since 2004, the HCRT system has been studied in the simple vertebrate, zebrafish [15,16]. This diurnal animal emerged as a popular model to study neurodevelopment and behavior, including the hypothalamic regulation of sleep [17]. The zebrafish has a simple, but conserved, brain anatomy and function with mammals [18]. In addition, it is amenable for genetic manipulation, using transposase- [19] and clustered regularly interspaced short palindromic repeats (CRISPR)-associated protein 9 (Cas9)- based genome editing [20]. Unlike most other vertebrates, the brain of the larva zebrafish is transparent, making it suitable for time-lapse imaging of single cells and organelles [21,22], optogenetics, and brain-wide imaging of genetically encoded calcium markers [23,24]. Furthermore, the zebrafish is well suited for large-scale genetic-, pharmacological-, and behavioral-screens [25,26]. Zebrafish sleep was defined using behavioral criteria, specifically through reduction of voluntary movement during the night, clock-controlled rhythmic activity that peaks during the day, reversibility, specific posture, increased arousal thresholds, and sleep rebound post-sleep deprivation [27–32]. Two major sleep signatures have been characterized in zebrafish; slow bursting sleep and propagating wave sleep, which share commonalities with mammalian slow-wave sleep and paradoxical or rapid-eye-movement (REM) sleep, respectively [33].
The zebrafish HCRT neuronal network is simple: comprised of about 20 neurons in larvae and about 60 neurons in adults (Table 1) [16,27,31,34]. The zebrafish HCRT system includes a single hcrt gene, which encodes both HCRT1 and HCRT2 peptides [34], and only one HCRT receptor, which is structurally more similar to the mammalian HCRTR2. Similar to mammals, zebrafish HCRT neurons are located in the LHA and project to the telencephalon, diencephalon, mesencephalon, rhombencephalon, and the pineal gland, toward the noradrenergic, dopaminergic, serotonergic, cholinergic, histaminergic and melatonergic nuclei [15,16,21,27,31,34–36]. Overexpression of HCRT in the entire body of larval zebrafish induces wakefulness [27], and both the HCRT neuron-ablated larvae and the HCRT-receptor mutant (Hcrtr−/−) adult zebrafish exhibit sleep fragmentation, as is observed in narcoleptic patients [29,31]. Overall, the relative simplicity of the HCRT system in zebrafish makes this vertebrate an ideal model to study the diverse functions of HCRT neuronal networks.
Table 1.
The HCRT system in zebrafish, rodents, and humans
| Zebrafish | Rodents | Humans | |
|---|---|---|---|
| Number of HCRT neurons | ~20–60 | ~5,000 | ~70,000 [12] |
| Type of peptides | HCRT1 and HCRT2 | HCRT1 and HCRT2 | HCRT1 and HCRT2 [2] |
| Type of receptors | HCRTR2 | HCRTR1 and HCRTR2 | HCRTR1 and HCRTR2 [2] |
| Brain regions | LHA | LHA, DMH and PeFLH | LHA and PHA [93] |
| Main functions studied | Sleep and wakefulness, feeding | Sleep and wakefulness, feeding, energy homeostasis, emotions, stress response, and reward | Sleep and wakefulness, feeding, emotions, social interaction, and reward [94,95] |
DMH, dorsomedial hypothalamus; HCRT, hypocretin; HCRTR, hypocretin receptor; LHA, lateral hypothalamic area; PeFLH, perifornical part of lateral hypothalamus area; PHA, posterior hypothalamic area.
It is generally assumed that HCRT neurons are a relatively homogeneous population; however, finer granularity may exist. In this chapter, we discuss the hypothesis that HCRT neurons can control multiple functions due to their heterogeneous genetics, anatomy, and functionality. We raise the possibility of functional HCRT subpopulations with distinct gene expression, activity, and connectivity profiles.
The genetic profile of HCRT neurons
The molecular marker that defines the HCRT neurons is the HCRT neuropeptide. However, these neurons contain other distinct proteins, which are expressed in small to large subsets of HCRT neurons, in addition to other regions of the brain. In order to study the comprehensive molecular identity of HCRT neurons, several studies have been conducted in human, rodents, and fish. They have revealed an array of genes that are expressed in subpopulations of HCRT neurons or in almost all of the cells, suggesting molecular heterogeneity (Table 2).
Table 2.
Specific gene and protein expression signatures that define HCRT neuron populations in rodents
| Proteins and mRNAs expressed in most HCRT neurons | Proteins and mRNAs expressed in a subset of HCRT neurons | ||
|---|---|---|---|
| PDYN | Chou et al., 2001 [37] | IGFBP3 | Honda et al., 2009 [47] |
| NARP/NPTX2* | Reti et al., 2002 [38] | TRIB2 | Cvetkovic-Lopes et al., 2010 [48] |
| CCKAR | Tsujino et al., 2005 [42] | PCSK1, NGB, DLK1, GPX3, LGALS3, LHX9* | Dalal et al., 2013 [50] |
| ENTPD3/NTPDase3* | Belcher et al., 2006 [46] | CART, PENK | Mickelsen et al., 2017 [40] |
| NNAT, CELF6, STAT5B, RFX4* | Dalal et al., 2013 [50] | PEG3, TLR2, ARHGAP36, IRS4 | Seifinejad et al., 2019 [55] |
| NUCB2 | Mickelsen et al., 2017 [40] | QRFP | Takahashi et al., 2020 [53] |
Only mRNAs/proteins that were anatomically verified are listed. Proteins/mRNAs that are expressed in HCRT neurons of both zebrafish and rodents are marked with *.
ARHGAP36, Rho GTPase-activating protein 36; CART, cocaine and amphetamine regulatory transcript; CCKAR, cholecystokinin A receptor; CELF6, CUGBP elav-like family member 6; DLK1, delta like non-canonical notch ligand 1; ENTPD3/NTPDase3, ectonucleoside triphosphate diphosphohydrolase 3; GPX3, glutathione peroxidase 3; IGFBP3, insulin-like growth factor-binding protein 3; IRS4, insulin receptor substrate 4; LGALS3, galectin 3; LHX9, LIM homeobox 9; NARP/NPTX2, neuronal pentraxin 2; NGB, neuroglobin; NNAT, neuronatin; NUCB2, nucleobindin 2; PCSK1, proprotein convertase subtilisin/kexin type 1; PDYN, prodynorphin; PEG3, paternally-expressed gene 3; PENK, proenkephalin; QRFP, pyroglutamylated RFamide peptide; RFX4, regulatory factor X4; STAT5B, signal transducer and activator of transcription 5B; TLR2, toll-like receptor 2; TRIB2, tribbles homolog 2.
Among the mRNA and/or proteins expressed in almost all HCRT neurons are the secreted proteins, prodynorphin (PDYN) and neuronal activity-regulated pentraxin II (NARP/NPTX2). These two proteins have been used to show that the cells, and not only the HCRT ligand, are absent in narcoleptic patients [11,37–39]. In human brains, at least 80% of the HCRT-producing neurons also contain Pdyn mRNA and NARP protein [11]. In the rat hypothalamus, all (>99%) HCRT-positive neurons are NARP-positive [38] and nearly all (94 ± 2%) are PDYN-positive [37]. These two proteins are also expressed widely in additional brain areas, and not restricted to the LHA. Confirming these results, studies in mice have also shown co-expression of Hcrt and Pdyn mRNA in 98.4% of the HCRT neurons using in situ hybridization (ISH) [40]. NARP can enhance postsynaptic responses to glutamate, while PDYN inhibits neuron activity via the κ-opioid receptor [37,41]. However, the role of NARP and PDYN in the HCRT system is undetermined.
Another protein found in almost all HCRT neurons in mice is the cholecystokinin receptor (CCKAR) [42]. CCK suppresses food intake [43] and locomotor activity [44]. Moreover, studies in rats have reported on co-localization of ectonucleoside triphosphate diphosphohydrolase 3 (ENTPD3/NTPDase3) and the ATP ligand-gated cation channel receptor P2X2R in almost all HCRT neurons [45,46]. The co-localization of P2X2R and ENTPD3 in mammalian HCRT neurons suggests that purinergic signaling might modulate the functions of the HCRT system, such as the regulation of sleep and energy metabolism.
The expression of these genetic markers in essentially the whole HCRT neuronal population suggests, at first sight, that these neurons would be homogenous. However, other genes are only expressed in a portion of HCRT neurons. For example, comparison of the transcriptome of narcoleptic versus control postmortem human brains, and transgenic mice lacking HCRT neurons versus wildtype mice, revealed that insulin-like growth factor-binding protein 3 (IGFBP3) colocalizes in ~80% of HCRT neurons in mice but only in 10–20% of HCRT neurons in humans [47]. It was suggested that elevated IGFBP3 levels in HCRT cells could decrease HCRT production and wakefulness under physiological conditions. Another protein expressed only in a subset of HCRT neurons is tribbles homolog 2 (TRIB2), an intracellular protein produced by HCRT neurons and other cell types that is targeted by antibodies in a fraction of patients with narcolepsy [48,49]. TRIB2-specific antibody-containing serum of a narcoleptic patient showed that 87% of HCRT neurons in the mouse hypothalamus are co-stained by the serum [48].
Using the translating ribosome affinity purification assay, 188 transcripts were identified as robustly enriched in HCRT neurons [50], including the known HCRT cell markers Pdyn, Narp, and Igfbp3. Moreover, few novel genes and gene products had substantial overlap with HCRT neurons, including CUGBP elav-like family member 6 (CELF6), neuronatin (NNAT), signal transducer and activator of transcription 5b (STAT5b) and regulatory factor x4 (RFX4), with the latter expressed only within these neurons. In contrast, several gene products were expressed only in a subset of HCRT neurons, revealing molecular heterogeneity in these neurons: proprotein convertase subtilisin/kexin type 1 (PCSK1, 42 ± 4.6 % co-localization), neuroglobin (NGB, 54 ± 3.6 %), glutathione peroxidase 3 (GPX3, 29 ± 9.3 %), galectin 3 (LGALS3, 35 ± 5.8 %), delta like non-canonical notch ligand 1 (DLK1, 44 ± 10 %) and LIM homeobox 9 mRNA (Lhx9, 76 ± 3.6 % co-localization) [50]. Ablation of Lhx9 resulted in more than 30% loss of HCRT neurons, without a general disruption of hypothalamic development. It appears that Lhx9 can regulate the development and specification of a subset, but not of all, HCRT neurons [50]. It remains to be determined whether these genetic variations reflect the transient expression state of the neurons at a given state or rather represent functionally-distinct subsets of HCRT populations.
Single-cell RNA-sequencing (scRNA-seq) of the mouse hypothalamus found partial co-localization of pyroglutamylated RFamide peptide (Qrfp) and Hcrt mRNA [51]. QRFP is a hypothalamic peptide that has been implicated in food intake, activity, anxiety, cardiovascular functions, arousal and hibernation-like state [52–54]. QRFP is lost together with HCRT neurons in Hcrt-ablated mice [55]. However, IHC and transgenesis showed that QRFP and HCRT do not colocalize in the same hypothalamic neurons in zebrafish and mice [54,56]. This can be explained by the differences in sensitivity of the sequencing and histological techniques. Surprisingly, recent IHC assay in mice showed that few QRFP-positive neurons also express HCRT [53]. Altogether, these experiments suggest that a low number of HCRT neurons co-express QRFP, which may help mediate the function of a small subpopulation of HCRT neurons.
RNA-sequencing of purified HCRT neurons and comparison to other hypothalamic cells revealed an array of differentially expressed genes in mice. This work used IHC to show that paternally-expressed gene 3 (PEG3), toll-like receptor 2 (TLR2), rho GTPase-activating protein 36 (ARHGAP36) and insulin receptor substrate 4 (IRS4) partially colocalize with HCRT [55]. Moreover, loss of Peg3 in mice significantly reduced HCRT cell numbers, while knock-down of a Peg3 in zebrafish completely abolished Hcrt expression, resulting in a two-fold increase in the amount of sleep [55].
The development of single-cell gene expression analysis enables through-profiling of the genetic identity of distinct neurons. This method was used to show in mice that the most extensively co-expressed transcripts in HCRT neurons are Pdyn (100% co-localization), Lhx9 (100%), Igfbp3 (100%), Nptx2/Narp (100%), Celf6 (100%), and Nnat (98%), which were discussed above. This method also revealed new candidate genes, such as nucleobindin 2 (Nucb2, 100%), proenkephalin (Penk, 76.8%), and cocaine and amphetamine regulatory prepropeptide (Cartpt, 42.0%) [40]. Validation of the results using ISH and IHC showed that although Nucb2 mRNA (84.4%) and NUCB2 protein (99.2%) colocalize with the majority of HCRT neurons, Penk (31.4%) and Cartpt (8.4%) positive cells had limited overlap with HCRT neurons, and immunoreactive CART protein was barely detected in HCRT neurons (0.8%) [40]. This last observation suggests that few HCRT neurons contain Cartpt mRNA but only a small amount of detectable protein.
These extensive studies have resulted in a list of genes that are expressed in many or in a few of HCRT neurons (Table 2). However, the complexity of the mammalian brain, which includes thousands of dense HCRT neurons that are intermingled with other hypothalamic neurons, have raised experimental challenges to isolate and study an uncontaminated HCRT neuronal population. In contrast, the relatively simple brain of the zebrafish enables the identification of specific HCRT neurons. The Hcrt promoter and transgenic tg(Hcrt:EGFP) zebrafish [35] provides an optimal model to profile the genetic identity of HCRT neurons. In zebrafish, similar to the findings in mammals, the Entpd3 and the P2X receptor P2rx8, which mediate purinergic signaling, are expressed in a small subset of HCRT neurons in larvae. These findings suggest that HCRT neurons mediate purinergic signaling by cell surface P2X receptors and Entpd3, which control the extracellular concentration and accessibility of ATP [57]. As adenosine signaling has been linked to sleep regulation in zebrafish [58], these results suggest that some HCRT neurons regulate metabolism and the sleep/wake cycle by sensing purine nucleotides. An additional gene found to be expressed in a subset of HCRT neurons in the zebrafish lateral hypothalamus (LH) is Nptx2b [21]. Moreover, a study of the leptin-neurotensin (Nts)-HCRT neuronal circuit demonstrated that a portion of both larvae and adult HCRT neurons express the Nts receptor [59], suggesting that Nts neurons regulate the function of HCRT neurons in zebrafish, as in mammals [60].
We isolated and sequenced the entire HCRT neuronal population using fluorescence-activated cell sorting (FACS) and whole transcriptome RNA-sequencing in zebrafish larvae. Hundreds of novel HCRT-neuron-specific transcripts were identified [61]. The functional roles of the genes were diverse and included regulation of metabolism, sleep, synaptogenesis, and synaptic plasticity. A comparison between the HCRT-neuron-specific candidate genes isolated in zebrafish [61] and mice [50] showed that at least eight genes (Rfx4, Lhx9, Scg2, Vgll2, Ptprn, Creb3l1, Sgsm1, and Fam46a) are found in HCRT neurons in both vertebrates. Verification of the results using double-staining showed that while several genes, including Kcnh4a, Hmx3, Lhx9 and Dennd1b, were expressed in most HCRT neurons, the majority, including Star, Fam46a, and Zgc171844 were expressed only in a subset of HCRT neurons. Notably, the portion of co-localization with HCRT in larvae was similar to that in adults. Intriguingly, similar to the finding in mice, Lhx9 can induce the specification of HCRT neurons in zebrafish [50,61,62].
Altogether, the studies in mammals and zebrafish suggest that the functional diversity of HCRT neurons can be regulated by their diverse molecular actors. Considering that the HCRT neuropeptides are only two of the secreted players, and additional proteins and neurotransmitters are likely to play a role in the function of the neurons, we suggest that the HCRT neurons do not necessarily act as one homogenous unit, but rather are divided into functionally-distinct subpopulations. Understanding the function of the genes that are differentially expressed in HCRT subpopulations will provide the basis to elucidate the mechanism that regulates the multi-functions of HCRT neurons.
The various HCRT interactions with other neuronal populations
Another approach to classify the heterogeneity of HRCT neurons is by connectivity of their upstream and downstream projections. Previous studies have examined HCRT neuronal afferent and efferent projections using traditional and genetically targeted neuroanatomical tracing methods in rodents. In general, HCRT neurons integrate a wide variety of neural signals from multiple sources distributed all over the brain, including the TMN, paraventricular thalamic nucleus, arcuate nucleus, cholinergic and monoaminergic nuclei [4–6,63–65]. Additionally, HCRT-expressing neurons are excited by a wide variety of neurotransmitters, including glutamate [66], ATP [67], corticotropin releasing factor [68], thyrotropin releasing hormone [69], noradrenaline [70], and acetylcholine [66,70], and they are inhibited by GABA [66] and adenosine [71,72]. In turn, HCRT neurons project diffusely throughout the brain, with especially dense excitatory projections to brain areas regulating arousal (e.g. locus coeruleus [LC] and TMN), reward (e.g. ventral tegmental area [VTA] and nucleus accumbens [NAc]) and autonomic function (e.g. brainstem sympathetic and respiratory control centers). An open question remains as to whether there are distinct HCRT cell clusters that exhibit different afferent and efferent projection maps?
To systematically determine the afferents to the HCRT neurons, researchers used retrograde and anterograde tracing to identify direct inputs to HCRT neurons in rat. They found that anterograde tracers injected into the brainstem and reward-related nuclei preferentially mark the lateral part of the HCRT field, while injections into the ventromedial hypothalamus (VMH) and other hypothalamic regions preferentially stain HCRT neurons in the medial and PeFLH field [65].
Output projections from HCRT neurons also demonstrate a preference between brain regions. A number of studies have shown that HCRT neurons can be anatomically subdivided into at least two separate subpopulations: one subpopulation is thought to reside in the medial HCRT field in the PeFLH and dorsomedial hypothalamus (DMH), sending projections to the LC and TMN, while the other subpopulation is thought to reside in the lateral HCRT field in the LHA, sending projections to the VTA and NAc (Figure 1) [73–77]. Moreover, only a portion of HCRT neurons project to the sublaterodorsal tegmental nucleus (SLD) and regulate the activation of REM sleep [78]. In contrast, a study in mice suggests that although HCRT neurons can be classified based on their downstream projections, these classifications do not show a lateral/medial topographic location within the hypothalamus. Based on retrograde tracing of target neurons, HCRT neurons that project to both the LC and TMN or to both the VTA and NAc were identified, however, these subpopulations were intermingled [79].
Fig. 1. Possible functional heterogeneity of HCRT neurons.
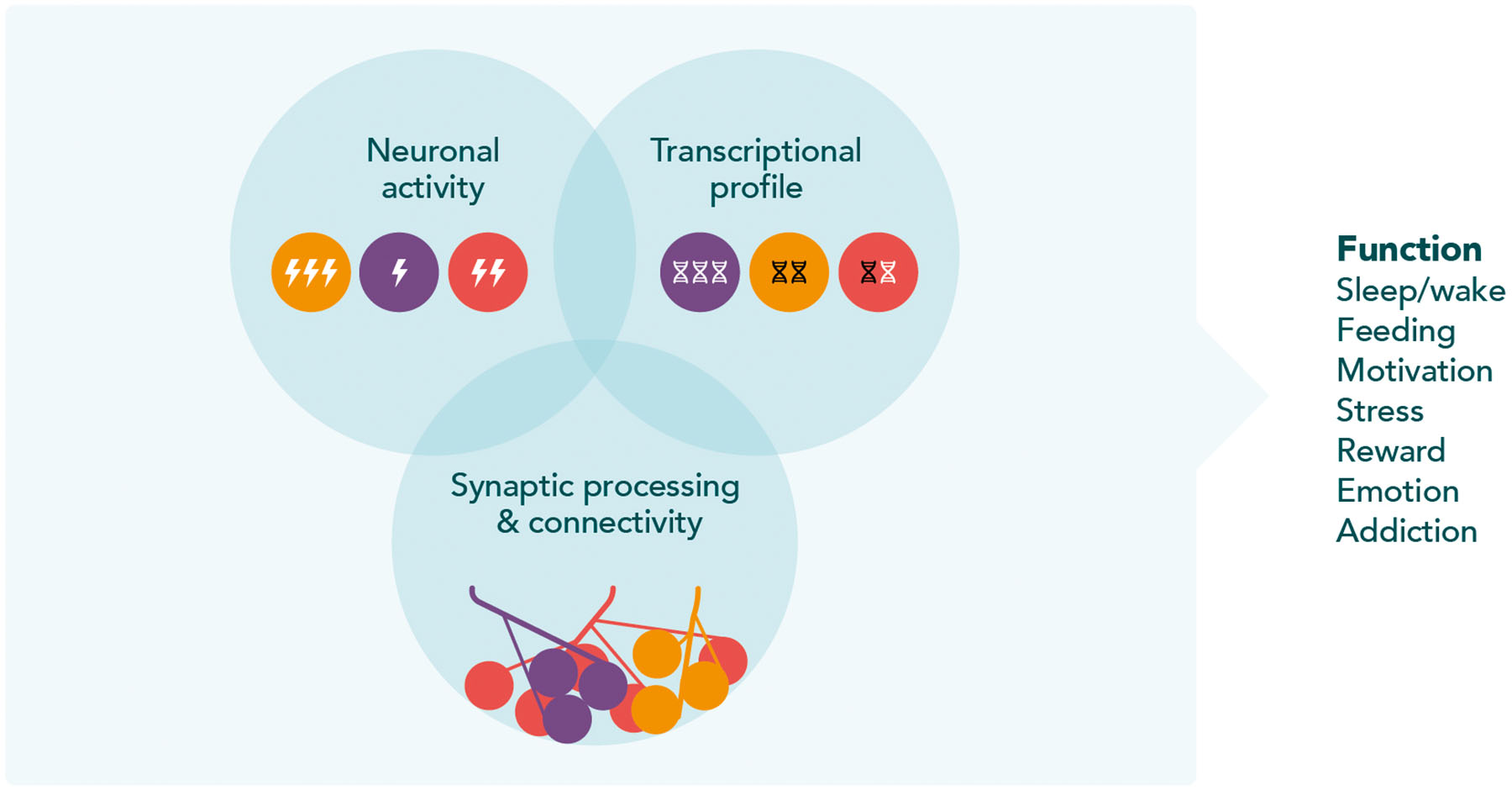
Schematic illustration of proposed functional heterogeneity of HCRT neurons. The unique transcriptional profile, neuronal activity, and synaptic processing and connectivity of HCRT neurons suggest that the multi-functional HCRT neurons can be divided into distinct functional subpopulations in all vertebrates.
In agreement with findings in rodents, in zebrafish, HCRT neurons have been shown to innervate and functionally interact with noradrenergic neurons [80] and the histaminergic system [81] in the LC and TMN, respectively. Further study in mammals and comparison to simpler vertebrates can shed light on the projection variabilities within the HCRT neuronal populations. Segregated HCRT neuronal projections may interact with their targets selectively. Whether these anatomically distinct HCRT populations display autonomous activity patterns, physiological and behavioral characteristics, require further investigation.
The diverse functions of HCRT neurons
Considering the heterogeneity of gene expression and neuronal connectivity, the diverse functions of HCRT neurons in regulating various behaviors such as feeding, sleep, stress, motivation and reward, might indeed be regulated by different subpopulations. Accumulating data suggest that HCRT neurons are divided into distinct functional subpopulations because of variability in intrinsic electrical and synaptic properties [82], or because of differential regulation by drugs and behavioral paradigms [74]. In this section, we discuss data that suggest that HCRT neurons are functionally diverse.
At the cellular level, HCRT neurons can be functionally subdivided into two populations based on their electrophysiological properties. Unbiased statistical analysis of electrical properties of HCRT neurons in combination with 3-D analysis of their shape, pointed to an existence of two subgroups of HCRT neurons in the mouse brain that have unique ‘electrical fingerprints’ and distinct ways of receiving information from other neurons [82]. Moreover, electrophysiological and calcium imaging data have demonstrated a wide array of neuronal activity patterns in HCRT neurons, although retrograde tracers injected into either the LC or VTA do not seem to preferentially label specific active classes of HCRT neurons [83].
It has been proposed that HCRT neurons in the LH regulate motivation and reward, whereas HCRT neurons located in the PeFLH and DMH regulate arousal and response to stress (Figure 1) [73]. For example, during wakefulness, the expression of the neuronal activity marker Fos is increased in PeFLH HCRT neurons, but not in the lateral HCRT neurons [84]. In a rodent model of depression, HCRT neurons located in the PeFLH and DMH regions are found to be more sensitive to unpredictable chronic mild stress than HCRT neurons located in the LH [85]. In contrast, the reward of food or drug induces activity in the lateral HCRT neurons [74]. Studies have shown that morphine administration increase Fos expression in the lateral HCRT neurons [75] and intermittent access to cocaine activates the lateral HCRT subpopulation, while selective knock-down of lateral HCRT neurons reduces motivation for cocaine [86].
Similar to mammals, the HCRT system in zebrafish consolidates sleep and wakefulness [15], induces food intake [87], and modulates neural circuitry of stress [88]. Neural activity of freely behaving larvae, monitored using Ca2+-dependent bioluminescence, was associated with periods of increased locomotor activity [89]. However, functional differences between subclasses of HCRT neurons in zebrafish is unknown and require further research.
Future directions
The HCRT system has been extensively studied in humans and multiple animal models, ranging from zebrafish and cavefish to rodents and primates [15,90–92]. Since their discovery, HCRT neurons have been defined based on their cellular marker – the HCRT neuropeptides – and largely treated as one homogenous population. However, accumulating data suggest functional, anatomical, and genetic heterogeneity of HCRT neurons because of the diverse expression profiles of molecular actors and synaptic connections, as well as cell-specific activity. These findings offer new interpretations for previous studies in the field and may help to understand the role of the HCRT system in health and disease.
Further studies are required in order to decipher and characterize the anatomy and function of subpopulations of HCRT neurons. For example, single-cell RNA-seq experiments can identify and profile genetically defined HCRT neuronal subpopulations. These genetically distinct subpopulations can be functionally characterized using imaging of neural activity markers during the sleep/wake cycle, feeding, stress and reward states. These experiments can be followed by manipulation of HCRT neuronal subpopulations using optogenetics, synaptic silencing, and genetically-induced neuron ablation, in order to understand the unique role of each HCRT neuronal networks.
Key Take-Home Points.
The hypothalamic hypocretin/orexin (HCRT) system regulates diverse functions, ranging from arousal and feeding to stress and reward.
Based on their unique gene expression profile, connections, and activity, we propose that the HCRT neurons are divided into distinct functional subpopulations in all vertebrates.
The relatively simple zebrafish is an attractive model to study the structure and function of subpopulation of HCRT neurons in single neuron resolution.
Funding sources
This work was supported by the United States—Israel Binational Science Foundation (BSF, grant no. 2017105) and the National Institutes of Health (NIH, 1RO1MH116470-01A1).
Footnotes
Conflicts of interest
The authors have no conflicts of interest to declare.
References
- 1.Simerly RB. Chapter 13 - Organization of the Hypothalamus. In: Paxinos G, editor. The Rat Nervous System (Fourth Edition) [Internet]. San Diego: Academic Press; 2015. p. 267–94. [Google Scholar]
- 2.Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, Tanaka H, et al. Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell. 1998;92(4):573–85. [DOI] [PubMed] [Google Scholar]
- 3.Lecea L de, Kilduff TS, Peyron C, Gao X-B, Foye PE, Danielson PE, et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc Natl Acad Sci U S A. 1998;95(1):322–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sakurai T The neural circuit of orexin (hypocretin): maintaining sleep and wakefulness. Nat Rev Neurosci. 2007;8(3):171–81. [DOI] [PubMed] [Google Scholar]
- 5.de Lecea L Optogenetic control of hypocretin (orexin) neurons and arousal circuits. Curr Top Behav Neurosci. 2015;25:367–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Peyron C, Kilduff TS. Mapping the hypocretin/orexin neuronal system: An unexpectedly productive journey. J Neurosci. 2017;37(9):2268–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lin L, Faraco J, Li R, Kadotani H, Rogers W, Lin X, et al. The sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor 2 gene. Cell. 1999;98(3):365–76. [DOI] [PubMed] [Google Scholar]
- 8.Chemelli RM, Willie JT, Sinton CM, Elmquist JK, Scammell T, Lee C, et al. Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell. 1999;98(4):437–51. [DOI] [PubMed] [Google Scholar]
- 9.Nishino S, Ripley B, Overeem S, Lammers GJ, Mignot E. Hypocretin (orexin) deficiency in human narcolepsy. Lancet. 2000;355(9197):39–40. [DOI] [PubMed] [Google Scholar]
- 10.Krahn LE, Lymp JF, Moore WR, Slocumb N, Silber MH. Characterizing the emotions that trigger cataplexy. J Neuropsychiatry Clin Neurosci. 2005;17(1):45–50. [DOI] [PubMed] [Google Scholar]
- 11.Crocker A, España RA, Papadopoulou M, Saper CB, Faraco J, Sakurai T, et al. Concomitant loss of dynorphin, NARP, and orexin in narcolepsy. Neurology. 2005;65(8):1184–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Thannickal TC, Moore RY, Nienhuis R, Ramanathan L, Gulyani S, Aldrich M, et al. Reduced number of hypocretinneurons in human narcolepsy. Neuron. 2000;27(3):469–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Peyron C, Faraco J, Rogers W, Ripley B, Overeem S, Charnay Y, et al. A mutation in a case of early onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains. Nat Med. 2000;6(9):991–7. [DOI] [PubMed] [Google Scholar]
- 14.Wang P, Wang M, Zhang L, Zhong S, Jiang W, Wang Z, et al. Functional characterization of an orexin neuropeptide in amphioxus reveals an ancient origin of orexin/orexin receptor system in chordate. Sci China Life Sci. 2019;62(12):1655–69. [DOI] [PubMed] [Google Scholar]
- 15.Elbaz I, Levitas-Djerbi T, Appelbaum L. The hypocretin/orexin neuronal networks in zebrafish. Curr Top Behav Neurosci. 2017;33:75–92. [DOI] [PubMed] [Google Scholar]
- 16.Kaslin J The orexin/hypocretin system in zebrafish is connected to the aminergic and cholinergic systems. J Neurosci. 2004;24(11):2678–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Levitas-Djerbi T, Appelbaum L. Modeling sleep and neuropsychiatric disorders in zebrafish. Curr Opin Neurobiol. 2017;44:89–93. [DOI] [PubMed] [Google Scholar]
- 18.Evans DH, editor. The physiology of fishes. 2nd ed. Boca Raton: CRC Press; 1998. 519 p. (Marine science series). [Google Scholar]
- 19.Kawakami K Tol2: A versatile gene transfer vector in vertebrates. Genome Biol. 2007;8(1):S7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hwang WY, Fu Y, Reyon D, Maeder ML, Tsai SQ, Sander JD, et al. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat Biotechnol. 2013;31(3):227–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Appelbaum L, Wang G, Yokogawa T, Skariah GM, Smith SJ, Mourrain P, et al. Circadian and homeostatic regulation of structural synaptic plasticity in hypocretin neurons. Neuron. 2010;68(1):87–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Elbaz I, Zada D, Tovin A, Braun T, Lerer-Goldshtein T, Wang G, et al. Sleep-dependent structural synaptic plasticity of inhibitory synapses in the dendrites of hypocretin/orexin neurons. Mol Neurobiol. 2017;54(8):6581–97. [DOI] [PubMed] [Google Scholar]
- 23.Naumann EA, Fitzgerald JE, Dunn TW, Rihel J, Sompolinsky H, Engert F. From whole-brain data to functional circuit models: The zebrafish optomotor response. Cell. 2016;167(4):947–960.e20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Thiele TR, Donovan JC, Baier H. Descending control of swim posture by a midbrain nucleus in zebrafish. Neuron. 2014;83(3):679–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rihel J, Prober DA, Arvanites A, Lam K, Zimmerman S, Jang S, et al. Zebrafish behavioral profiling links drugs to biological targets and rest/wake regulation. Science. 2010;327(5963):348–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Granato M, van Eeden FJ, Schach U, Trowe T, Brand M, Furutani-Seiki M, et al. Genes controlling and mediating locomotion behavior of the zebrafish embryo and larva. Dev Camb Engl. 1996;123:399–413. [DOI] [PubMed] [Google Scholar]
- 27.Prober DA, Rihel J, Onah AA, Sung R-J, Schier AF. Hypocretin/Orexin overexpression induces an insomnia-like phenotype in zebrafish. J Neurosci. 2006;26(51):13400–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zada D, Appelbaum L. Chapter 9 - Behavioral criteria and techniques to define sleep in zebrafish. In: Gerlai RT, editor. Behavioral and Neural Genetics of Zebrafish [Internet]. Academic Press; 2020. p. 141–53. [Google Scholar]
- 29.Elbaz I, Yelin-Bekerman L, Nicenboim J, Vatine G, Appelbaum L. Genetic ablation of hypocretin neurons alters behavioral state transitions in zebrafish. J Neurosci. 2012;32(37):12961–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zhdanova IV, Wang SY, Leclair OU, Danilova NP. Melatonin promotes sleep-like state in zebrafish. Brain Res. 2001;903(1):263–8. [DOI] [PubMed] [Google Scholar]
- 31.Yokogawa T, Marin W, Faraco J, Pézeron G, Appelbaum L, Zhang J, et al. Characterization of sleep in zebrafish and insomnia in hypocretin receptor mutants. PLoS Biol. 2007;5(10):e277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Elbaz I, Foulkes NS, Gothilf Y, Appelbaum L. Circadian clocks, rhythmic synaptic plasticity and the sleep-wake cycle in zebrafish. Front Neural Circuits. 2013;7:9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Leung LC, Wang GX, Madelaine R, Skariah G, Kawakami K, Deisseroth K, et al. Neural signatures of sleep in zebrafish. Nature. 2019;571(7764):198–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Faraco JH, Appelbaum L, Marin W, Gaus SE, Mourrain P, Mignot E. Regulation of hypocretin (orexin) expression in embryonic zebrafish. J Biol Chem. 2006;281(40):29753–61. [DOI] [PubMed] [Google Scholar]
- 35.Appelbaum L, Wang GX, Maro GS, Mori R, Tovin A, Marin W, et al. Sleep-wake regulation and hypocretin-melatonin interaction in zebrafish. Proc Natl Acad Sci U S A. 2009;106(51):21942–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Panula P Hypocretin/orexin in fish physiology with emphasis on zebrafish. Acta Physiol Oxf Engl. 2010;198(3):381–6. [DOI] [PubMed] [Google Scholar]
- 37.Chou TC, Lee CE, Lu J, Elmquist JK, Hara J, Willie JT, et al. Orexin (hypocretin) neurons contain dynorphin. J Neurosci Off J Soc Neurosci. 2001;21(19):RC168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Reti IM, Reddy R, Worley PF, Baraban JM. Selective expression of Narp, a secreted neuronal pentraxin, in orexin neurons. J Neurochem. 2002;82(6):1561–5. [DOI] [PubMed] [Google Scholar]
- 39.Blouin AM, Thannickal TC, Worley PF, Baraban JM, Reti IM, Siegel JM. Narp immunostaining of human hypocretin (orexin) neurons: loss in narcolepsy. Neurology. 2005;65(8):1189–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mickelsen LE, Kolling FW, Chimileski BR, Fujita A, Norris C, Chen K, et al. Neurochemical heterogeneity among lateral hypothalamic hypocretin/orexin and melanin-concentrating hormone neurons identified through single-cell gene expression analysis. eNeuro. 2017;4(5): ENEURO.001317.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Muschamp JW, Hollander JA, Thompson JL, Voren G, Hassinger LC, Onvani S, et al. Hypocretin (orexin) facilitates reward by attenuating the antireward effects of its cotransmitter dynorphin in ventral tegmental area. Proc Natl Acad Sci U S A. 2014;111(16):E1648–1655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Tsujino N, Yamanaka A, Ichiki K, Muraki Y, Kilduff TS, Yagami K, et al. Cholecystokinin activates orexin/hypocretin neurons through the cholecystokinin A receptor. J Neurosci Off J Soc Neurosci. 2005;25(32):7459–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kopin AS, Mathes WF, McBride EW, Nguyen M, Al-Haider W, Schmitz F, et al. The cholecystokinin-A receptor mediates inhibition of food intake yet is not essential for the maintenance of body weight. J Clin Invest. 1999;103(3):383–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hirosue Y, Inui A, Miura M, Nakajima M, Okita M, Himori N, et al. Effects of CCK antagonists on CCK-induced suppression of locomotor activity in mice. Peptides. 1992;13(1):155–7. [DOI] [PubMed] [Google Scholar]
- 45.Florenzano F, Viscomi MT, Mercaldo V, Longone P, Bernardi G, Bagni C, et al. P2X2R purinergic receptor subunit mRNA and protein are expressed by all hypothalamic hypocretin/orexin neurons. J Comp Neurol. 2006;498(1):58–67. [DOI] [PubMed] [Google Scholar]
- 46.Belcher SM, Zsarnovszky A, Crawford PA, Hemani H, Spurling L, Kirley TL. Immunolocalization of ecto-nucleoside triphosphate diphosphohydrolase 3 in rat brain: Implications for modulation of multiple homeostatic systems including feeding and sleep–wake behaviors. Neuroscience. 2006;137(4):1331–46. [DOI] [PubMed] [Google Scholar]
- 47.Honda M, Eriksson KS, Zhang S, Tanaka S, Lin L, Salehi A, et al. IGFBP3 colocalizes with and regulates hypocretin (orexin). PloS One. 2009;4(1):e4254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cvetkovic-Lopes V, Bayer L, Dorsaz S, Maret S, Pradervand S, Dauvilliers Y, et al. Elevated Tribbles homolog 2-specific antibody levels in narcolepsy patients. J Clin Invest. 2010;120(3):713–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kawashima M, Lin L, Tanaka S, Jennum P, Knudsen S, Nevsimalova S, et al. Anti-tribbles homolog 2 (TRIB2) autoantibodies in narcolepsy are associated with recent onset of cataplexy. Sleep. 2010;33(7):869–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Dalal J, Roh JH, Maloney SE, Akuffo A, Shah S, Yuan H, et al. Translational profiling of hypocretin neurons identifies candidate molecules for sleep regulation. Genes Dev. 2013;27(5):565–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Romanov RA, Zeisel A, Bakker J, Girach F, Hellysaz A, Tomer R, et al. Molecular interrogation of hypothalamic organization reveals distinct dopamine neuronal subtypes. Nat Neurosci. 2017;20(2):176–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Takayasu S, Sakurai T, Iwasaki S, Teranishi H, Yamanaka A, Williams SC, et al. A neuropeptide ligand of the G protein-coupled receptor GPR103 regulates feeding, behavioral arousal, and blood pressure in mice. Proc Natl Acad Sci U S A. 2006;103(19):7438–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Takahashi TM, Sunagawa GA, Soya S, Abe M, Sakurai K, Ishikawa K, et al. A discrete neuronal circuit induces a hibernation-like state in rodents. Nature. 2020;583(7814):109–14. [DOI] [PubMed] [Google Scholar]
- 54.Okamoto K, Yamasaki M, Takao K, Soya S, Iwasaki M, Sasaki K, et al. QRFP-deficient mice are hypophagic, lean, hypoactive and exhibit increased anxiety-like behavior. PLoS ONE. 2016;11(11):e0164716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Seifinejad A, Li S, Mikhail C, Vassalli A, Pradervand S, Arribat Y, et al. Molecular codes and in vitro generation of hypocretin and melanin concentrating hormone neurons. Proc Natl Acad Sci U S A. 2019;116(34):17061–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chen A, Chiu CN, Mosser EA, Khan S, Spence R, Prober DA. QRFP and its receptors regulate locomotor activity and sleep in zebrafish. J Neurosci. 2016;36(6):1823–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Appelbaum L, Skariah G, Mourrain P, Mignot E. Comparative expression of p2x receptors and ecto-nucleoside triphosphate diphosphohydrolase 3 in hypocretin and sensory neurons in zebrafish. Brain Res. 2007;1174:66–75. [DOI] [PubMed] [Google Scholar]
- 58.Gandhi AV, Mosser E, Oikonomou G, Prober DA. Melatonin is required for the circadian regulation of sleep. Neuron. 2015;85(6):1193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Levitas-Djerbi T, Yelin-Bekerman L, Lerer-Goldshtein T, Appelbaum L. Hypothalamic leptin-neurotensin-hypocretin neuronal networks in zebrafish. J Comp Neurol. 2015;523(5):831–48. [DOI] [PubMed] [Google Scholar]
- 60.Leinninger GM, Opland DM, Jo Y-H, Faouzi M, Christensen L, Cappellucci LA, et al. Leptin action via neurotensin neurons controls orexin, the mesolimbic dopamine system and energy balance. Cell Metab. 2011;14(3):313–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yelin-Bekerman L, Elbaz I, Diber A, Dahary D, Gibbs-Bar L, Alon S, et al. Hypocretin neuron-specific transcriptome profiling identifies the sleep modulator Kcnh4a. Elmquist JK, editor. eLife. 2015;4:e08638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Liu J, Merkle FT, Gandhi AV, Gagnon JA, Woods IG, Chiu CN, et al. Evolutionarily conserved regulation of hypocretin neuron specification by Lhx9. Dev Camb Engl. 2015;142(6):1113–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.González JA, Iordanidou P, Strom M, Adamantidis A, Burdakov D. Awake dynamics and brain-wide direct inputs of hypothalamic MCH and orexin networks. Nat Commun. 2016;7(1):11395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sakurai T, Nagata R, Yamanaka A, Kawamura H, Tsujino N, Muraki Y, et al. Input of orexin/hypocretin neurons revealed by a genetically encoded tracer in mice. Neuron. 2005;46(2):297–308. [DOI] [PubMed] [Google Scholar]
- 65.Yoshida K, McCormack S, España RA, Crocker A, Scammell TE. Afferents to the orexin neurons of the rat brain. J Comp Neurol. 2006;494(5):845–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Henny P, Jones BE. Innervation of orexin/hypocretin neurons by GABAergic, glutamatergic or cholinergic basal forebrain terminals evidenced by immunostaining for presynaptic vesicular transporter and postsynaptic scaffolding proteins. J Comp Neurol. 2006;499(4):645–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Wollmann G, Acuna-Goycolea C, van den Pol AN. Direct excitation of hypocretin/orexin cells by extracellular ATP at P2X receptors. J Neurophysiol. 2005;94(3):2195–206. [DOI] [PubMed] [Google Scholar]
- 68.Winsky-Sommerer R, Yamanaka A, Diano S, Borok E, Roberts AJ, Sakurai T, et al. Interaction between the corticotropin-releasing factor system and hypocretins (orexins): A novel circuit mediating stress response. J Neurosci. 2004;24(50):11439–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Hara J, Gerashchenko D, Wisor JP, Sakurai T, Xie X (Simon), Kilduff TS. Thyrotropinreleasing hormone increases behavioral arousal through modulation of hypocretin/orexin neurons. J Neurosci. 2009;29(12):3705–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bayer L, Eggermann E, Serafin M, Grivel J, Machard D, Muhlethaler M, et al. Opposite effects of noradrenaline and acetylcholine upon hypocretin/orexin versus melanin concentrating hormone neurons in rat hypothalamic slices. Neuroscience. 2005;130(4):807–11. [DOI] [PubMed] [Google Scholar]
- 71.Thakkar MM, Winston S, McCarley RW. Orexin neurons of the hypothalamus express adenosine A1 receptors. Brain Res. 2002;944(1–2):190–4. [DOI] [PubMed] [Google Scholar]
- 72.Liu Z-W, Gao X-B. Adenosine inhibits activity of hypocretin/orexin neurons via A1 receptor in the lateral hypothalamus: a possible sleep-promoting effect. J Neurophysiol. 2007;97(1):837–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Harris GC, Aston-Jones G. Arousal and reward: a dichotomy in orexin function. Trends Neurosci. 2006;29(10):571–7. [DOI] [PubMed] [Google Scholar]
- 74.Harris GC, Wimmer M, Aston-Jones G. A role for lateral hypothalamic orexin neurons in reward seeking. Nature. 2005;437(7058):556–9. [DOI] [PubMed] [Google Scholar]
- 75.Richardson KA, Aston-Jones G. Lateral hypothalamic orexin/hypocretin neurons that project to ventral tegmental area are differentially activated with morphine preference. J Neurosci Off J Soc Neurosci. 2012;32(11):3809–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hasegawa E, Yanagisawa M, Sakurai T, Mieda M. Orexin neurons suppress narcolepsy via 2 distinct efferent pathways. J Clin Invest. 2014;124(2):604–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Mochizuki T, Arrigoni E, Marcus JN, Clark EL, Yamamoto M, Honer M, et al. Orexin receptor 2 expression in the posterior hypothalamus rescues sleepiness in narcoleptic mice. Proc Natl Acad Sci U S A. 2011;108(11):4471–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Feng H, Wen S-Y, Qiao Q-C, Pang Y-J, Wang S-Y, Li H-Y, et al. Orexin signaling modulates synchronized excitation in the sublaterodorsal tegmental nucleus to stabilize REM sleep. Nat Commun. 2020;11(1):3661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Iyer M, Essner RA, Klingenberg B, Carter ME. Identification of discrete, intermingled hypocretin neuronal populations. J Comp Neurol. 2018;526(18):2937–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Singh C, Oikonomou G, Prober DA. Norepinephrine is required to promote wakefulness and for hypocretin-induced arousal in zebrafish. eLife. 2015;4:e07000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Sundvik M, Panula P. Interactions of the orexin/hypocretin neurones and the histaminergic system. Acta Physiol Oxf Engl. 2015;213(2):321–33. [DOI] [PubMed] [Google Scholar]
- 82.Schöne C, Venner A, Knowles D, Karnani MM, Burdakov D. Dichotomous cellular properties of mouse orexin/hypocretin neurons. J Physiol. 2011;589(Pt 11):2767–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.González JA, Jensen LT, Fugger L, Burdakov D. Convergent inputs from electrically and topographically distinct orexin cells to locus coeruleus and ventral tegmental area. Eur J Neurosci. 2012;35(9):1426–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Estabrooke IV, McCarthy MT, Ko E, Chou TC, Chemelli RM, Yanagisawa M, et al. Fos expression in orexin neurons varies with behavioral state. J Neurosci. 2001;21(5):1656–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Nollet M, Gaillard P, Minier F, Tanti A, Belzung C, Leman S. Activation of orexin neurons in dorsomedial/perifornical hypothalamus and antidepressant reversal in a rodent model of depression. Neuropharmacology. 2011;61(1):336–46. [DOI] [PubMed] [Google Scholar]
- 86.James MH, Stopper CM, Zimmer BA, Koll NE, Bowrey HE, Aston-Jones G. Increased number and activity of a lateral subpopulation of hypothalamic orexin/hypocretin neurons underlies the expression of an addicted state in rats. Biol Psychiatry. 2019;85(11):925–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Yokobori E, Kojima K, Azuma M, Kang KS, Maejima S, Uchiyama M, et al. Stimulatory effect of intracerebroventricular administration of orexin A on food intake in the zebrafish, Danio rerio. Peptides 2011;32(7):1357–62. [DOI] [PubMed] [Google Scholar]
- 88.Pavlidis M, Theodoridi A, Tsalafouta A. Neuroendocrine regulation of the stress response in adult zebrafish, Danio rerio. Prog Neuropsychopharmacol Biol Psychiatry. 2015;60:121–31. [DOI] [PubMed] [Google Scholar]
- 89.Naumann EA, Kampff AR, Prober DA, Schier AF, Engert F. Monitoring neural activity with bioluminescence during natural behavior. Nat Neurosci. 2010;13(4):513–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Jaggard JB, Stahl BA, Lloyd E, Prober DA, Duboue ER, Keene AC. Hypocretin underlies the evolution of sleep loss in the Mexican cavefish. Elife. 2018;7:e32637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Adamantidis AR, Zhang F, Aravanis AM, Deisseroth K, de Lecea L. Neural substrates of awakening probed with optogenetic control of hypocretin neurons. Nature. 2007;450(7168):420–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Diano S, Horvath B, Urbanski HF, Sotonyi P, Horvath TL. Fasting activates the nonhuman primate hypocretin (orexin) system and its postsynaptic targets. Endocrinology. 2003;144(9):3774–8. [DOI] [PubMed] [Google Scholar]
- 93.Moore RY, Abrahamson EA, Van Den Pol A. The hypocretin neuron system: an arousal system in the human brain. Arch Ital Biol. 2001;139(3):195–205. [PubMed] [Google Scholar]
- 94.Sutcliffe JG, Lecea L de. The hypocretins: Excitatory neuromodulatory peptides for multiple homeostatic systems, including sleep and feeding. J Neurosci Res. 2000;62(2):161–8. [DOI] [PubMed] [Google Scholar]
- 95.Blouin AM, Fried I, Wilson CL, Staba RJ, Behnke EJ, Lam HA, et al. Human hypocretin and melanin concentrating hormone levels are linked to emotion and social interaction. Nat Commun. 2013;4:1547. [DOI] [PMC free article] [PubMed] [Google Scholar]